Embed Size (px)
Citation preview

Helgol~inder wlss. Meeresunters. 22, 377-400 (1971)
Zur Trematodenfauna der Herzmuscheln Cardium edule und Cardium lamarcki
G. LAUCKNER
Biologische Anstalt Helgoland (Zentrale); Hamburg 50, Bundesrepublik Deutschland
ABSTRACT: On the trematode fauna of the cockles Cardium edule and Cardium lamarcki. Thirteen species of larval Digenea were recovered from the soflc parts of their intermediate hosts, the common cockle Cardlum edule L. and LAMARCK'S cockle Cardium lamarcki R ~ w , collected from North Sea tidal fiats and from shallow reaches of the German Baltic Sea coast. These comprise the encysted metacercariae of Himasthla elongata, H. continua, H. interrupta (all with 29 collar spines), one undetermined, rare Himasthla species (with 31 collar spines), Renicola roscovita, Psilostomum brevicolle, Asyrnphylodora demeli, and one undetermined metacercaria resembling LEBOUR'S (1911.) Cercaria mytiIL Cockles are favourite intermediate hosts for gymnophallids. C. edule harbours the sporocyst stage of Gymnophallus choledochus. Sporocysts of another, yet unidentified gymnophallid, occur in C. lamarcki. Unencysted meta- cercariae of Gymnophallus gibberosus are common to both Cardiurn species, while Meiogyrnno- phallus minutus and MeiogymnophaIlus sp. occur in C, edule only, thus exhibiting marked host-specificity. Morphologically similar eehinostomatid metacercarial cysts could be distin- guished by their diameters, which were found to have a species-specific constant size regardless of the salinity and the host species in which they are formed. Application of biometricaI methods revealed that the cysts of Renicola roscovita, Asymphylodora demeli and Metacer- carla sp. grow within their hosts. These observations are confirmed by experimental infections of different mollusc species. Gymnophallid metacercariae showed marked preference for certain microhabitats within the host, and are readily distinguished by their situs: MeiogyrnnophaIlus minutus occurs in "chalky spots" just under the umbo, while MeiogymnophalIus sp. inhabits "pits" between mantle and shell of C. edule. G. gibberosus is restricted to the tissues of the anterior adductor muscle. Mierohabkat selection occurs also in encysted metacercariae, but not to such a pronounced degree as in the family Gymnophallidae. No interspecific exclusion of larval trematodes was encountered. Where incidence and intensity of infection are highest, up to 11 of the reported ~3 species may parasitize the same host specimen. Infections with gymnophallids were always higher in C. edule than in C. lamarcki.
E I N L E I T U N G
Die beiden litoralen Herzmuschelarten Cardiurn edule L. und Cardiurn larnarcki REEVE slnd Zwischenwirte einer ganzen Reihe digener Trematoden, deren Adult i in SeevSgeln oder Fischen parasitieren. Die ersten Untersuchungen/fiber Helminthenbefall yon C. edule gehen auf VILLOT (1878), HU~T (1888a, b), JAMESON (1902), JAMESON & NICOLL (1913), JOHNSTON~ (1905), L~BO~R (1905, 1907, 1908a, b, 1911, 1914), NICOLL (1906) und PELSENEErl (1906) zuriick.
Die Zahl der seither erschienenen, iiberwiegend parasitologisch-systematisch orien-

378 G. LAucKNZR
tierten Untersuchungen, in denen Trematoden-Entwi&lungsstadien aus Cardium edule beschrieben werden oder in denen die Herzmuschet als deren Zwis&enwirt genannt wird, ist beachtli&; bier seien nur die wi&tigsten Arbeiten angeftihrt" ]3ARTOLI (1965), BOWERS (1969), Bow~I~S & JAMES (1967), COLa (1935, 1938), HOTTON (1952), Loos- FI~AI,~K (1967, 1968, 1969, 1971a, b), MARKOWSKI (1936), PALOMBI (1934), PREVOT (1965), RE~CQ & PREVOT (1962), R~Es (1939), REIMZR (1964, 1970), STUlVKAI~D & UZMANN (1958), ~VERDING (I969).
Trotz dleser Vielzahl yon VeriSffentlic~ungen fehlt bis heute eine zusammenfas- sende r0bersi&t tiber die Trematodenfauna der Herzmuscheln. Die einzige, yon COL~ (1956) zusammengestellte Liste der Parasiten und Kommensalen yon C. eduIe ist un- vollst~ndig und in bezug auf die Taxonomie fehlerhatt. Das gleiche gilt ftir viele vor allem der ~Iteren Arbeiten.
H~iufig wurden die bei der Untersu&ung yon Herzmus&eln gefundenen larvalen Trematoden allein auf Grund morphologischer Khnli&keiten einem bekannten Adultus zugeordnet, ohne dat~ der Nachweis der Identit~it dur& experimentelle Endwirtinfek- tion erbra&t worden w~ire. Dadurch kam es vor allem bei den artenreichen und in Car- dium h~iufig vorkommenden Genera Himasthla und Gymnophallus zu erheblichen Konfusionen. Mehrfachbeschreibungen und -benennungen sind eher die Regel als die Ausnahme (STtJNKARD 1939, LOos-FI~ANK 1967, 1969, 1971a, 1971b).
Heute hat sich generell die Ansicht durchgesetzt, dafg die sichere Bestimmung yon Larvenstadien digener Trematoden nur auf dem Wege tiber die experimentelle Voll- endung des Entwicktungszyklus und an Hand des Adultus mSglich ist. Obgleich die er- wa&senen Wtirmer durch Angaben tiber ihre KSrperdimensionen und -proportionen und unter Hinzuziehung ~Skologischer Daten (Wirtsspezifit~it, Situs im Endwirt usw.) charakterisiert werden, mangelt es an entsprechenden exakten Untersuchungen zur Bio- metrie und Ukologie der Larvalstadien. Das tiber sie vorliegende sp~irliche Daten- material und vor allem die widersprtichlichen Befunde verschiedener Autoren unter- streichen die Notwendigkeit experimenteller Entwicklungsstudien.
Andererseits schlieflen Routine- und Freilanduntersuchungen an Mollusken diesen zeitaufwendigen Weg in den meisten Fiilten aus. Fiir den tJkologen kann es jedoch yon Bedeutung sein, die Artzugeh~Srigkeit der vorge£undenen Trematodentarven zu kennen, denn die verschiedenen Parasitenarten kSnnen ihren Wirt in unterschiedlichem Mage physiologisch bceinflussen (LAuc~N~R, in Vorbereitung).
Bei der Interpretation parasitologisch-meristischer Daten wirkt sich die Tatsache erschwerend aus, daf~ in der Literatur Mat~angaben generell in statistisch inad~iquater Form dargeboten werden. Dadurch sind die Mefgdaten verschiedener Autoren kaum ur~d nur unter grogen Vorbehalten untereinander vergldchbar. Exakte Untersuchun- gen tiber die Variabili6/t einzelner KSrperdimensionen der zur Diskussion stehenden Parasitenarten fehlen vSlIig. Damit bleibt unklar, welchen Aussagewert - das heigt, wel&e statistis&e Si&erheit - die in der parasitologischen Literatur zur Artunters&ei- dung herangezogenen Metgdaten als taxonomis&e Kriterien besitzen.
Die vorliegende Arbeit gibt einen 13berbli& tiber die Trematodenfauna yon Car- dium edule und C. lamarcki aus dem Bereich der deutschen Nord- und Ostseektiste. Es wird der Versu& unrernommen, die in Herzmuscheln parasitierenden Larvenstadien nahe verwandter Arten an Hand meristischer Merkmale zu unterscheiden.

Zur Trematodenfauna der Herzmuscheln 379
MATERIAL UND METHODEN
Der iiberwiegende Teil der auf Trematodenbefall untersuchten Herzmuscheln stammt aus dem Nordsylter Watt (,-, 27 bis 30 0/0o S), aus der Flensburger F~Srde (I-Iabernis/Neukir&en, ~-, 15 bis 19 °/0o S) und aus dem Burger See (Fehmarn, ~ 10 bis 12 °/0o S). An einigen Standorten des Nordsylter Watts und in der Flensburger FSrde kommeri beide Herzmus&elarten in ausgedehnten Mis&populationen vor; der Burger See wird auss&lief~lich yon C. lamarcki besiedelt (LAucKNm~, im Dru&).
Encystierte Metacercarien wurden zur Untersuchung aus dem Wirtsgewebe her- ausgel&t und unter Vermeiden yon De&glasdru& mit Hilfe yon Mikroskop und Okularmikrometer lebend, Cercarien sowie freibewegliche Metacercarien nach der Me- rhode yon LOos-FI~ANK (1967, 1970) in fixiertem Zustand vermessen. Bei ovalen Cysten wurde der grSgere Durchmesser bestimmt.
Aus den Megwerten wurden die statistischen Matgzahten - arithmetis&es Mittel (7), Standardabweichung (s) und Variabilit~itskoeffizient (V) - errechnet und die er- haltenen H~iufigkeitsverteilungen mit Hilfe des x2-Goodness-of-fit-Test auf I~berein- stimmung :nit der theoretisch zu erwartenden Normalverteilung untersucht. Mittel- wertunterschiede wurden mittels t-Test auf statistische Signifikanz geprii~ und Einzel- daten zweier oder mehrerer Proben unter Anwendung der Varianzanalyse miteinan- der verglichen.
Fiir die Auswertung der Megergebnisse stand ein ,,WANG 700 B"-Elektronen- rechner (DFG-Leihgabe) zur Verfiigung. Rechenprogramme und Methodik sind an anderer Stelle ausfiihrlich beschrieben (LAucKN~R, im Druck).
Die unretuschierten Abbildungen 2 bis 5 sind Lebendaufnahmen (LEITZ-OR- THOLUX mit Aufsatz-LEICA und MICROBLITZ 300). Da die Objekte zur Verbes- serung der Detailwiedergabe unter wechselndem De>asdru& aufgenommen worden
Abb. I: Cardium edule L. (links) und Cardium lamarcki RE~w (rechts)

380 G. LAucKNeR
sind, wurde auf die Eintragung yon Magstgben in die Abbildungen verzichtet. Ver- bindli&e Magangaben enth~.lt Tabelle 2; die Met~methodik entspri&t der yon Loos- FlVaNK (1970) angewendeten.
ERGEBNISSE
U n t e r s c h e i d u n g d e r C a r d i u m - A r t e n
In keiner der bisher ver~Sffentlichten Arbeiten tiber die Trematodenfauna der Herzmuscheln wird zwischen Cardium edule und C. lamarcki unterschieden, sondern stets nur ,,C. edule" als Wirtsart genannt. Im Verlauf dieser Untersu&ung zeigte sich aber, dat~ das Befallsbild beider Mus&elarten sowohI qualitative als auch quantitative Unterschiede aufweist. Literaturunstimmigkeiten finden dadur& zumindest eine teiI- weise Erkl~irung.
C. edule (Abb. 1, links) hat gerundete, mit quergestellten Schuppen besetzte Rip- pen, die auch im posterioren Bereich voll ausgebildet sind. Der hintere Tell des Schalen- spaltes schlieflt infolge der Verzahnung beider Klappen in einer wellenf6rmigen Linie. Das Periostrakum ist bei dieser Art nur schwa& ausgebildet, das Ligament fang und kr~i~ig.
C. Iamarcki (Abb. 1, rechts) hat spitz-dachkantartige Rippen ohne Schuppen, jedoch mit schuppen~ihnlichen Auffaltungen des bei dieser Art sehr kr~iitig ausgebil- deten Periostrakums. Im posterioren Bereich sind die Rippen stark abgeflacht oder feh- Ien fast ganz; der Schalenspalt schlieBt daher in einer geraden Linie, Das Ligament ist kurz und schmal.
Die mittlere Anzahl der Radi~irrippen ist bei C. edule h~Sher als bei C. Iamarcki. Weitere Angaben zur Schalenmorphologie und Biometrie beider Arten finden sich in einer gesonderten Darstellung (LAucKNEIq im Druck).
Eine Unters&eidung ist au& an Hand der Weichk6rper m6glich: Die Tubuli der Mitteldarmdriise sind bei C. edule hellgrtin bis dunkelgrtin, bei C. lamarctei hellbraun bis dunkelbraun gef~irbt. Die Aut~enseite der Siphonen ist bei C. eduIe hellgetb, bei C. lamarcki dunkler und augerdem braun gesprenkelt.
T r e m a t o d e n - E n t w i c k l u n g s s t a d i e n a u s C a r d i u m e d u l e u n d C a r d i u m I a m a r c k i
Die im Untersuchungsgebiet in Herzmus&eln angetroffenen larvalen Trematoden sind in Tabelle 1 zusammengestellt.
Abb. 2: A Himasthla etongata, Cyste. B H. elongata, aus der Cyste herauspr~iparierte Metacer- carie. C H. elongata, Kopfstacheln. D H, elongata beim Eindringen in die Fuf~muskulatur yon C. edule. Die Cercarie hat den Schwanz bereits abgeworfen. E Fut~ eines jungen C. edule mit starkem H. elongata-Befall aus experimenteller Infektion. F Nach 400t~igiger Inkubation bei 4 ° C vom Wirtstier gebildete, eine H. elongata-Cyste umgebende Kalkkapsel. Die unter Deck-
glasdruck befreite Metacercarie war noch aktiv
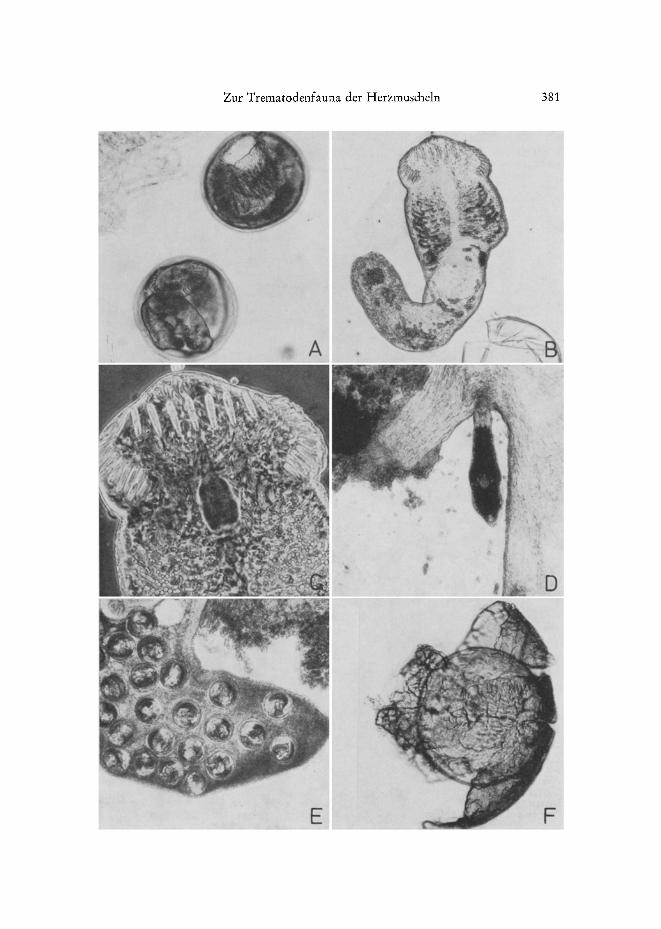
Zur Trematodenfauna der Herzmuscheln 381

382 G. Lauc~NER
Himasthla elongata (Abb. 2 A-F): Der Durchmesser der runden bis leicht ovalen Metacercariencysten betr~igt zwischen 225 und 270 ]~m. Sie finden sich ganz iiber- wiegend in der Fut~muskulatur und nur sehr selten in anderen KSrperabschnitten. Der Entwi&Iungszyklus yon H. elongata wurde yon WERDING (1969) aufgekl~rt. Erster Zwischenwirt ist dana& Littorina littorea, Endwirte sind Grof~m~Swen (Loos-Fr, AN~ 1967). Hinsichtlich der Wahl des zweiten Zwischenwirtes zeigt H. elongata keine Wirtsspezifit~it. Die Cercarien encystieren sich in zahlreichen Muschel- und Schnecken- arten. Auch der erste Zwischenwirt, L. Iittorea, wird befallen.
Himasthla continua (Abb. 3A), mit einem Cystendurchmesser yon 185 bis 225 pro, bef~illt ebenfalls den Futg der Muscheln und encystiert sich vorwiegend in des- sen oberem Tell in der N~ihe des Eingeweidesackes. Da sich die Cystendurchmesser- Spannweiten beider genannten Arten im Bereich um 225/~m beriihren und einander gelegentlich sogar ieicht iiberlappen, kann es in dieser Gr/Sigenklasse zu Verwechslungen kommen. Grot~e H. continua-Cysten kSnnen fiir kleine H. elongata gehalten werden - und umgekehrt.
Himasthla interrupta (Abb. 3 B): Die Cysten messen zwischen 150 und 185 #m. In adulten Cardien besetzen sie vor atlem den Mantelrand; nur ausnahmsweise finder man sie neben den beiden anderen Arten in der Fut~muskulatur und noch seltener in anderen Organen. Bei unter 6 bis 7 mm langen Muscheln der Nullgruppe werden dem- gegeniiber auch Hepatopancreas und Ful~ starker befallen. Offenbar ist der sehr schmale Mantelrand dieser kleinen Cardien fiir die Cercarien no& wenig attraktiv.
H. continua und H. interrupta wurden yon Loos-FgAN~ (1967) als neue Arten be- schrieben und ihr Entwicklungszyklus aufgekl~rt. Erster Zwischenwirt ist in beiden Fiillen Hydrobia ulvae, als Endwirte fungieren verschiedene MSwenarten. Die Metav cercarien atler drei Himasthla-Species zeigen aui~er ihrer unterschiedlichen GrSt~e keine die Artentrennung erm~Sglichenden morphologischen Besonderheiten. Die Zahl der Kopfstacheln betr~gt iibereinstimmend 29.
Im Eingeweidesa& von C. edute aus dem Kampener Watt wurden in drei F~illen echinostomatide Metacercarien mit einem Durchmesser um 220 # m u n d 31 Kopf- stacheln gefunden. An den iibrigen Standorten trat diese Himasthla-Art niche auf. Die Vermutung, dat~ sie sehr selten ist, wird durch die Angaben yon Loos-FRAN~ (1967) untermauert, nach denen alle sechs im deutschen Nordseeraum vorkommenden Ver- treter dieses Genus 29 Kopfstacheln haben. Demnach diirfie der zu dieser Metacercarie geh6rende Adultus im Gebiet nut ausnahmsweise auftreten. Als erster Zwischenwirt kSnnte die im Sylter Vogelkojenwatt in einem sehr isolierten Bestand vorkommende Netzreusenschnecke, Nassarius reticulatus, in Betracbt gezogen werden. Nach ZI~GEL- ~EIeR (pers6nliche Mitteilung) schied eine im Labor gehakene Netzreusenschne&e grot~e Himasthla-Cercarien aus. Kopfstachdz,ihtungen wurden allerdings nicht durch- gefiihrt. STUNI~AI~D (1938) beschreibt aus Nassarius obsoletus Himasthla quissetensis, eine Art mit ebenfatls 31 Kopfstacheln.
Abb. 3" A Himasthla continua. Aus der Cyste herausgepret~te Metacercarie. B H. interrupta, Metacercarie. C Renicola roscovita, junge Metacercarie aus dem Hepatopancreas eines 6 mm langen C. edule. D Palpus yon C. edule (Ausschnitt) mit starkem natiirlichen Befall yon Reni- cola roscovita. E Psilostomum brevicolle, Metacercarie aus dem Eingeweidesa& yon C. la-
rnarckL F P. brevicolle, aus der Cyste herauspr~iparierte Metacercarie
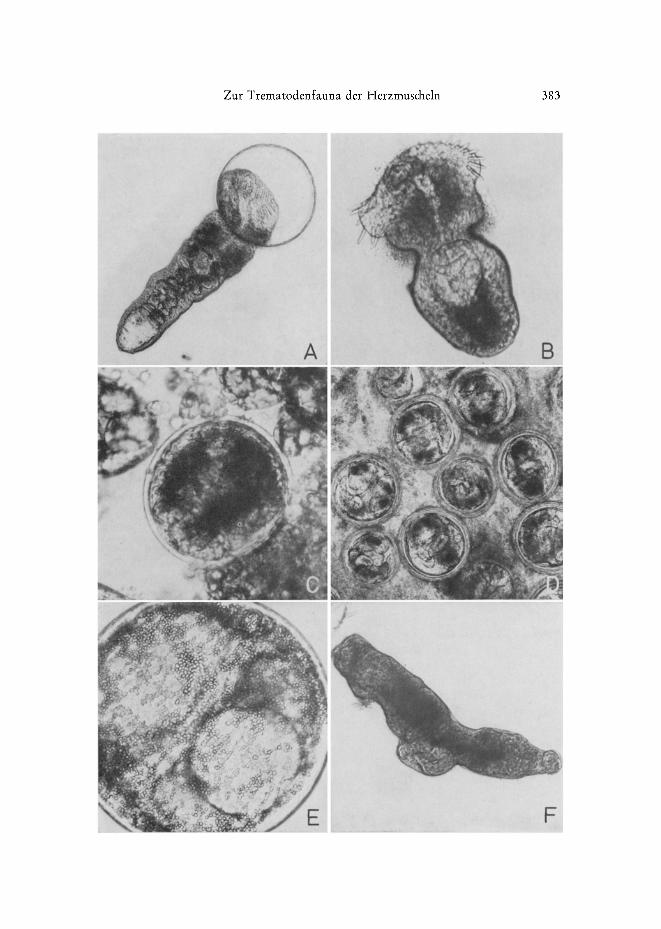
N
N

384 G. LAuCKNER
Die Himasthla-Metacercarien zeigen im Wirtsk/Srper kein Wachstum. Ihnen steht eine Gruppe yon Trematoden unters&iedticher systematis&er Stellung gegentiber, de- ren Cysten im Wirtsgewebe eine mit einer GrSt~enzunahme verbundene Entwi&lung durchmachen. Diese Gruppe wird im vorliegenden Material durch Renicola roscovita, Asymphyloclora demeli und Metacercaria spec., eine ohne experimentelle Endwirt- infektion ni&t einzuordnende Form, repr~isentiert. Wahrs&einlich geh6rt ihr auch Psilostomum brevicoIIe an. Die wenigen Funde lassen aber keine konkreten Aussagen iiber ein eventuelles Wachsturn dieser Metacercarie zu.
RenicoIa roscovita (Abb. 3 C-D): Der Dur&messer der kugelrunden Cysten be- tr~igt zwischen 110 und 180 #m und s&wankt damit in einem sehr weiten Bereich. Aus- gewachsene Metacercarien sind zwischen 160 und 180 #m grog. Befallen werden bei adulten Herzmus&eln ganz tiberwiegend die Palpen. Erst wenn sie di&t an dicht mit Cysten besetzt sind (Abb. 3 D), wei&en die Cercarien auf den Mantelrand (hiiufig), die Mitteldarmdriise (seltener) und die Kiemenbasis (sehr selten) aus. Bei Nullgruppen- Cardien, deren Mantelr~,inder noch s&wa& entwi&elt sind, wird dagegen zunii&st die Mitteldarmdrtise befallen. Der Entwi&lungsgang dieser Art wurde yon WERDING (I969) bes&rieben. Erster Zwis&enwirt ist Littorina littorea.
Metacercaria spec. (Abb. 5 E): Die Cyste hat grotge _Khnli&keit mit der yon R. roscovita, ist aber im Mittel gr6ger und di&wandiger und nimmt im Laufe ihrer Entwi&lung im zweiten Zwis&enwirt ebenfatls noch an Gr6t~e zu. Im September 1970 betrug der Durchmesser yon Metacercarien dieser Art aus C. lamarcki des Burger Sees 160 bis 190 #m; in Nullgruppen-edule aus dem Sylter Watt wurden bereits im August Cysten yon 180 bis 195 #m angetroffen. Dem Kurvenverlauf der H~iufigkeitsvertei- lung nach zu urteilen, diir&e der mittlere Durchmesser ausgewa&sener Metacercarien bei etwa 190 his 200 #m liegen.
Da sich die Durchmesser-Spannweite unausgewachsener Metacercaria spec. mit der yon R. roscovita stark iiberschneidet, MSnnen Verwechslungen vorkommen. Metacer- caria spec. bef~illt jedo& nie die Palpen, sondern auss&Iiet~lich Mitteldarmdriise und Eingeweidesa& mit einer deutli&en Konzentration in der N~ihe der Mund~iffnung.
Metacercaria spec. stimmt gut mit der Abbildung yon LEBOtJr~s (1911) ,,Cercaria mytili" und au& mit deren Situsangabe (Mitteldarmdriise) iiberein. Die sehr knappe Beschreibung der Autorin - ,,The cyst is spherical and thi&-walled but quite trans- parent, 0.20 mm across" (p. 450) - rei&t trotz der vollkommenen Ubereinstimmung far eine si&ere Identifikation nicht aus. Da ohnehin der zu ,,Cercaria mytili" ge- h~irende Adultus unbekannt ist, t~igt si& au& die Zugeh~irigkeit yon Metacercaria spec. nur auf experimentellem Wege kl~iren.
Psilostomum brevicolle (Abb. 3 E-F): Die 200 bis 230 #m grogen Metacercarien sind, was bereits LOos-FRANK (1968) bei der Aufkl~irung des Entwi&lungszyklus auf- rid, auf die Mitteldarmdriise (nach eigenen Beoba&tungen daneben auf den Ein- geweidesa&) spezialisiert. Dur& ihr diffuses Exkretionsnetz und den bei geeigneter Drehung der Larve auch durch die Cystenwandung hindurch erkennbaren Bauchsaug- napf-Sphinkter unters&eiden sie sich auff~illig und unverwe&selbar yon den iibrigen, im Hepatopancreas und Eingeweidesa& der Herzmuscheln parasitierenden Trema- toden. Erster Zwis&enwirt dieser Art ist Hydrobia utvae, Endwirte sind Seev/Sget.
Asymphylodora demeli (Abb. 5 F): Die zwis&en 160 und 380 #m grogen, diinn-

Zur Trematodenfauna der Herzmuscheln 385
wandigen Metacercarien dieses Fis&trematoden wurden ausschliei~lich in C. lamarcki aus dem Burger See gefunden. Sie sind auf das Nierengewebe spezialisiert und wachsen im Laufe ihrer Entwicklung auf mehr als das Zehnfache ihrer AnfangsgrSge heran, wo- durch sie das Wirtstier physiologisch ungewShnlich stark belasten. Der Entwicklungs- gang von A. demeli wird yon MA~KOWSKI (1936) und REIMER (1970) beschrieben. Erste Zwischenwirte sind Hydrobia ulvae und H. ventrosa, Endwirte vor allem Cypri- niden und Gobiiden.
Im Gegensatz zu den bisher aufgefiihrten Formen sind die Metacercarien der in Herzmuscheln parasitierenden Gymnophalliden nicht encystiert, wenngMch in einigen F~llen yon Wirtsgewebe eng umschlossen. Diese bindegewebigen H~illen wurden zum Teil als Sporocysten beschrieben, was in der Literatur zu einiger Verwirrung fahrte. Freigesetzt fiihren die Metacercarien nahezu ununterbrochen Kontraktions- und Streckbewegungen aus. Infolge der sich dabei s6indig stark veriindernden K~Srperform sind Messungen am lebenden Tier wenig sinnvoll.
Die Vertreter dieser Familie find ausnahmslos Vogelparasiten (S'ruNKARD & UZ- MANN 1958). Fiir drei von fiinf, in der vorliegenden Arbeit beschriebenen, in Herz- muscheln des Untersuchungsgebietes vorkommenden Gymnophalliden stellt Cardium den zweiten und fiir die beiden weiteren Arten den ersten Zwischenwirt dar (Tab. 1).
Weitgehende morphologische 13bereinstimmung insbesondere der Larvenstadien macht die Gymnophatlidae zu einer augerordentlich problematlschen Gruppe. Mehr- fachbeschreibungen und -benennungen kommen bei ihnen h~iufiger vor aIs in anderen Gruppen. Loos-FI~ANIf (1971b) fiihrt eine Liste yon 67 Artnamen auf. Davon sind nut acht Entwicklungszyklen roll aufgekl~irt; die iibrigen sind nur als Larval- oder als Aduttenstadium bekannt. STUNKAI~D & UZMA~ (1958) umreigen das Problem mit den treffenden Worten (p. 285): ,,The situation is chaotic and one of utter confusion."
Meiogymnophallus minutus (Abb. 4 A-B): Die Metacercarien liegen, yon Binde- gewebe lest umschlossen, in der N~ihe des Schalenwirbels unmittelbar unter dem Liga- ment. Freipr~iparierte Metacercarien bewegen sich sehr tr~ige. Die Schenkel der Exkre- tionsblase find m~ichtig aufgetrieben und prall mit stark lichtbrechenden Granula ge- fiillt, wodurch die Larven im Durchlicht fast schwarz erscheinen. Habitus und Situs dieser Art schliegen Verwechslungen mit anderen Gymnophalliden des Gebiets aus. BowE~s & JaMes (1967) sowie Loos-FRa~qK (1971b) geben Beschreibungen yon Meta- cercarie und Adultus. Der ersre Zwischenwirt dieser in den Wattengebieten der Nor& see wahrscheinlich hgufigsten Art ist noch immer unbekannt. BAR'rOLI (1965) beschreibt eine ~ihnliche, aber mit M. minutus nicht identische Art aus C. edule siidfranz/Ssischer Gew~isser als Gymnophallus fossarum. Der Angabe des Locus typicus nach diirRe es sich nicht um C. edule, sondern urn C. lamarcki (syn. C. glaucum sensu MARS 1951, 1966; vgl. LAOCi~NrI~, im Dru&) handeln.
Meiogymnophallus spec. (Abb. 4 B): Die Metacercarien, die etwas kleiner als die yon M. minutus und im Gegensatz zu diesen sehr beweglich sind, liegen, enggepa&t, zwischen Mantel und Schale in Gruppen wechselnder Gr5t~e zusammen. Bei guter Be- leuchtung kann man die in der Masse lei&t r6tlich erscheinenden Ansammlungen durch das dtinne MantelepitheI der Mus&el hindurchschimmern sehen. Sie liegen nicht frei, sondern in Heine Gruben in der Schalenwandung eingesenkt und find yon einer auf~er- ordentlich dtinnen Hiille umgeben. Diese steht offenbar mit dem Mantelepithel in Ver-

386 G. LAucKNER
bindung, denn beim Freipr~iparieren der Parasiten zerreit~t sie fast immer, wobei die Larven aus ihren Gruben herausgespiilt werden. Unverletzt bMbt die Hiille, wenn ab- gestorbene, bereits in Verwesang iibergehende Cardien mit grN~ter Vorsicht pr~ipariert werden. Die Parasiten iiberleben den Tod ihres Wirtes geraume Zeit.
LOos-FRANK (1971b) h~lt die bier als Meiogymnophallus spec. gefiihrte Form fiir junge Stadien yon M. minutus. F ii r diese Aufnahme spricht, dag beide die gleiche Wimperflammen-Formel - 2 [(2 + 2) + (2 + 2)] = 16 Protonephridien - und beide abgerundete Tegumentstachetn besitzen, w~ihrend nach LOos-FRANK (1971b) fiir atle anderen im Gebiet vorkommenden Gymnophalliden sp#ze Sta&eln charakteristisch sind. Ein weiteres Indiz ftir die IdentitY, it beider Formen ist der geringe FiilIungsgrad der Exkretionsblase yon MeiogyrnnophalIus spec.; er iiigt den Schlug zu, dab es sich um junge, noch nicht infekti~se Stadien handelt. D a g e g e n spricht der ,,station~ire Zustand", die Ansammlung in dichten, yon einer HiilIe umgebenen Gruppen, in dem diese Metacercarie auftritt. Eine weitere Stiitze ffir die Giiltigkeit der Alternativhypo- these liefert der VergMch der K6rperdimensionen beider Formen mit t-liFe der Va- rianzanalyse. Sie ergab beim Vergleich der Liingen-Breiten-ReIation yon je 10 fixierten Metacercarien yon Meiogyrnnophallus minutus und Meiogymnophallus spec. mit F = 19,41 und p = 0,34- 10 -~ extrem hohe Signifikanz. Eine endgiiltige Kl~irung kann jedoch erst das Infektionsexperiment erbringen.
Gymnophallus gibberosus (Abb. 4 C-F): Metacercarie (aus C. edule und Macoma bahica) und Adultus (aus Entenv/Sgeln) wurden yon LOos-FI~ANK (1971a) beschrieben. Der erste Zwischenwirt ist noch unbekannt. Die Situsangabe der Originalbeschreibung - ,,zwischen Mantel und Schale" - l~/f~t sich an Hand der eigenen Befnnde noch pr~izisie- ren: G. gibberosus parasitiert ausschliel~lich im Gewebe des k 1 e i n e n (vorderen) Schalenadduktors und des benachbarten vorderen Fugretraktors, und zwar iiberwie- gend an den Grenzfl~ichen zwischen Schale und Muskel. Wird der gesamte Weichk/Srper einschliefllich der Schliegmuskeln yon der Schale freipr~ipariert und in Wasser aus- geschiittelt, so fallen die G. gibberosus-Metacercarien heraus. Andererseits kann der Parasit teicht iibersehen werden, wenn bei der Pr~iparation nur der Mantel enffernt wird, der Schliel~muskel aber an der Schale verbleibt.
Die Metacercarien dieser Art ,,ffessen" sich aktiv - wahrscheinlich mit Hilfe pro- teolytischer Enzyme - durch das Schliet~muskelgewebe hindurch und veranlassen das Wirtstier zur Bildung yon Kalkkonkrementen, die entweder der Schale im Bereich der Muskelansatzsteilen mehr oder weniger lest aufsitzen oder abet als ,,Perlen" lose im Adduktorgewebe tiegen.
G. gibberosus w~chst im zweiten Zwischenwirt bedeutend st~irker als die beiden vorgenannten Meiogymnophaltus-Arten. Junge Metacercarien bewegen sich frei, aus- gewachsene Exemplare sind ot~ - wie die yon Meiogymnophallus spec., aber im Gegen- satz zu diesen einzeln - yon einer zarten Halle umgeben. Intakte Hiillen werden wie-
Abb. 4: A Meiogyrnnophaltus rninutus. Von Wirtsgewebe umgebene Metacercarien (Durch- licht). B Meiogymnophatlus spec. (links oben) zum Vergleich mit M. minutus (re&ts unten im Bild). C GymnophaIlus gibberosus. Metacercarie unter starkem Deckglasdruck. D G. gibberosus, ~iltere Metacercarie (ohne Deckglasdruck). E, F Kalkkonkremente ira Gewebe des kleinen Ad- duktormuskels yon Cardium tamarcki (schwarz); Pfeil in E: Stelle, an der sich die G. gibbero-
sus-Metacercarie befand
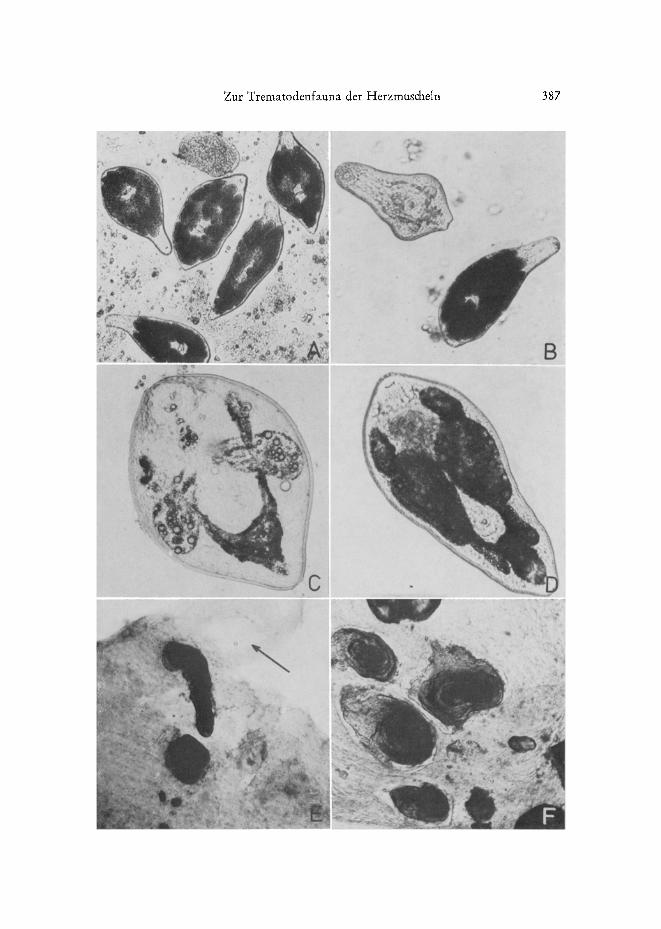
N
0 f~
L~
c~

388 G. LAUCKNER
derum nur bei der Pr~iparation stark verwester Herzmuscheln erhalten, deren Adduk- tormusket sich bereits auf leichten Zug hin yon der Schale lgst.
Bei s&wa&em Befall wird dieser Parasit h~ufig iibersehen. Auff~illiger als die Tiere setbst sind die auf ihrer Spur zurii&bleibenden Kalkkonkremente; sie sind ein si&eres Indiz fiir deren Anwesenheit. Keiner der iibrigen, in Cardium parasitierenden Gymnophalliden induziert derartige Bildungen (Abb. 4 E--F).
GymnophalIus choledochus (Abb. 5 A-B): Far diese Art stellt C. edule den ersten Zwischenwirt dar. Die furcocerce Cercarien oder - je nacb Jahreszeit - auch schwanz- lose Metacercarien enthaltenden Sporocysten liegen in grotger Zahl in allen Teilen des Eingeweidesackes. G. choledochus zeichnet sich durch Alternativzyklen aus (Loos- FRANK 1969): Die die Muschel verlassenden Cercarien befallen versch~edene Polychae- ten-Arten und gelangen haupts~chlich auf diesem Wege in die eine Gruppe yon End- wirten (Lariden). Ungeschliip~e Cercarien werfen im Herbst die Schwiinze ab und wachsen, ohne ihren ersten Zwischenwirt verlassen zu haben, in den Sporocysten zu infekti/Ssen Metacercarien heran. Samt der sie beherbergenden Muschel gelangen sie in die zweite Gruppe yon Endwirten (Limicolen), die ausgesprochene Muschelfresser sin&
Gymnophallus spec. (Abb. 5 C-D): Die Cercarien dieser Art, die sich ebenfalls in Sporocysten entwickeln, gleichen denen yon G. choledochus fast vollkommen, nur sind sie bedeutend kleiner. Da diese Art aber ausschlieglich in kleinen C. lamarcki aus dem Burger See gefunden wurde und beispielsweise auch G. gibberosus in der kieineren Macoma baltica (Loos-FI~ANK 1971a) und ebenso in C. lamarcki (eigeneBeobachtungen) eine geringere EndgrSt~e erreicht als in C. edule, wurden die Stadien aus dem Burger See zuniichst fiir G. choledochus gehalten. Erst die Weiterverfolgung des Entwi&lungs- ganges lieg Zweifel an der Identit~it aufkommen:
Die Metacercarien yon G. choledochus finden sich ,invariablement dans les para- podes" (Pl~vox 1965, p. 451). Die Untersuchungen yon LOos-FRANII (1969) sowie eigene Infektionsexperimente mit vom Trocbophora-Stadium parasitenfrei aufgezoge- hen Nerds diversicolor best~itigen die strenge Spezialisierung auf diesen Situs. Nach- dem N. diversicolor im Experiment mit einer grogen Zahl ausgeschwiirmter Cercarien von GymnophalIus spec. zusammengebracht worden waren, wurde an der Basis der Parapodialborsten in keinem Fall auf choledochus hindeutender Metacercarienbefall festgestellt. Demgegeniiber fanden sich zahlreiche Gymnophallidenlarven in der Mus- kulatur des Kieferapparates, was als sicheres Zeichen far das Vorliegen einer neuen Art anzusehen ist.
W i r t s s p e z i f i t ~ i t u n d B e f a l l s b i l d
In den Fiillen, in denen die Herzmuscheln als Zwischenwirte ftir encystierte Meta- cercarien - also ffir ausgespro&ene Ruhestadien - fungieren, ist yon vornherein keine Wirtsspezifit~t auf dem Gattungs- oder gar Artniveau zu erwarten. Die bier behandel- ten Formen lassen sich auch in anderen litoralen Mollusken- Lamellibranchiern wie
Abb. 5: A Cercarie von Gymnophallus choledochus. B Sporocyste yon G. choledochus. C Cer- carie yon Gymnophallus spec. aus C. tamarcki 7 Burger See. D Sporocyste yon Gymnophallus
spec. E Metacercaria spec. F Metacercarie yon Asymphylodora derneli
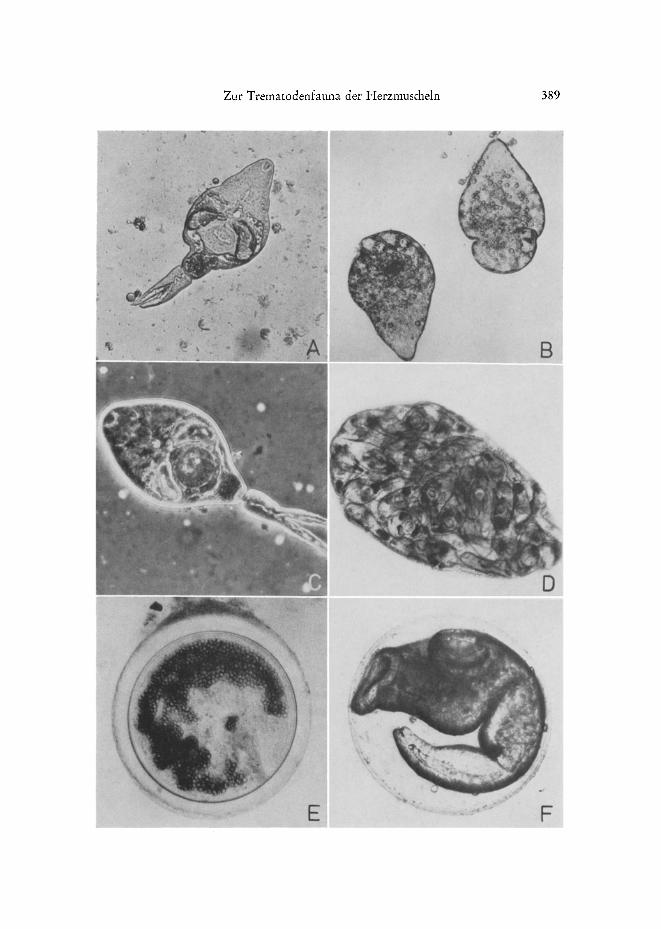
N
~D
N
c~o

390 G. LAUCKNER
Gastropoden - nachweisen. Das Befallsbild wird im wesentlichen durch das gleichzeitige Auftreten yon End- und Zwischenwirten bestimmt. So wird das Fehlen yon HimasthIa elongata, eines ausgesprochenen Grotgm6wentrematoden (Loos-F•ANK 1967) an den Standorten Flensburger F6rde und Burger See (Tab. 1) mit der geringen Abundanz bzw. dem Fehlen der Endwirte in diesem Gebiet zusammenh~ingen (HoFF 1941), denn die Zwischenwirte sind in allen FHlen vorhanden und die hydrographischen Voraus- setzungen ffir die Infektion des ersten Zwischenwirtes giinstig.
Bei Asymphylodora demeIi scheinen andere Faktoren eine verbreitungslimitie- rende Rolle zu spielen, denn diese Art fehlt mit Ausnahme des Burger Sees im Gebiet auch dort, wo Zwis&en- und Endwirte vorhanden sind. Zwar bes&reibt REIM~R (1970) das Auftreten dieser Art in C. edule aus FlachgewS.ssern um Riigen. Da abet nicht zwischen C. edule und C. lamarcki unterschieden wurde (REIMER, persSnliche Mitteilung), bleibt ungekliirt, ob dieser Parasit tats~i&li& in beiden Herzmus&elarten vorkommt oder auf C. Iamarcki beschr~inkt ist. Letzteres ist wenig wahrscheinli&, da A. demeli-Metacercarien yon ANKEL (1962) auch in Hydrobia ulvae und H. ventrosa angetroffen, wenn au& fiir E&inostomatiden-Entwi&Iungsstadien gehalten wurden. Na& WIKGREN (1956) kommt Asymphylodora nut in Bu&ten mit reicher Unterwas- servegetation vor. Diese Bedingungen sind im Burger See erfiillt.
V611ig entgegengesetzte VerhS.ltnisse werden bei den Gymnophalliden angetroffen. Bei den Arten, far die Cardium der erste Zwis&enwirt ist - G. choledochus und Gym- nophallus spec. - ist mit einer mehr oder weniger ausgepriigten Wirtsspezifit~it zu rech- nen. Far Gymnophallus spec. ist die Frage, ob beide Herzmus&elarten glei&ermagen befallen werden, an Hand des vorliegenden Materials ni&t zu beantworten, denn die- ser Parasit wurde bisher nut im Burger See gefunden - und dort kommt auss&lietg- li& C. lamarcki vor. Gymnophallus-Sporocysten wurden au& in C. lamarcki aus den Mischpopulationen des Sylter Watts gefunden. Da bier G. choledochus in C. edule vor- kommt, wurde ohne Vergleich der Cercarienmage angenommen, dag es sich bei den Sporocysten aus C. lamarcki ebenfalls um G. choledochus handelt. Erst die - sp~itere - Entde&ung der nenen Art, Gymnophallus spec., stelh die Ri&tigkeit dieser Annahme in Frage; sie bedarf daher weirerer Untersuchung.
Wie aus Tabelle 1 hervorgeht, kommt die Metacercarie yon G. gibberosus in bei- den Herzmuschelarten und an allen drei Standorten vor. Da sie augerdem in Macoma baltica parasitiert (Loos-FRANK 1971a, b), bietet sie ein Beispiel fiir geringe Wirts- spezififiit. Das genaue Gegenteil gilt fiir die beiden Meiogymnophallus-Arten. Rund 3600 C. edule yon verschiedenen Standorten des Sylter Watts wurden in den Jahren 1969 bis 1971 auf Trematodenbefall untersucht; alIe waren ausnahmslos mit M. minu- tus und Meiogymnophallus spec. infiziert. Von 2700 C. Iamarcki aus demselben Gebiet wies keines diese Parasiten auf. Auch in C. edule aus der Flensburger NSrde wurde Meiogymnophallus gefunden, wenngMch Befallsrate und Befallsintensit{it hier erheb- lich niedriger waren. C. lamarcki desselben Standortes erwiesen si& wiederum als un- befallen. Angesi&ts dieser Befunde kann angenommen werden, datg M. minutus und Meiogymnophallus spec. streng auf C. edule als zweiten Zwischenwirt spezialisiert sin&
Interspezifische Konkufrenz zwischen den in Herzmuscheln parasitierenden lar- valen Trematoden wurde in keinem Fall beoba&tet. So kommen in adulten C. edule

Zur Trematodenfauna der Herzmuscheln 391
aus dem Sylter Vogetkojen- und K6nigshafenwatt, wo der st~irkste Befall festgestellt wurde, Mischinfektionen mit in der Regel neun (und im Extremfall elf) der in Tabelle I aufgefiihrten 13 Arten im s e 1 b e n Wirtstier vor; in C. Iamarcki sind es gew6hnlich sieben und maximal neun.
Wirtsspezifische q u a n t i t a t i v e Unterschiede wurden beim Befall mit en- cystierten Metacercarien nicht gefunden, zeigren sich aber deutlich bei der Infektion mit gymnophalliden Metacercarien. Auch an Standorten mit Cardiurn-Mischpopula- tionen und Gymnophalliden-Massenbefall bei C. edule (rund 300 M. rninutus, 200 Meiogymnophallus spec. und 60 G. gibberosus pro Muschel von durchschnittlich 30 mm SchalenlSinge) enthielten C. larnarcki immer nut wenige G. gibberosus (4 bis 20 pro Muschel), die dariiber hinaus kleiner waren als die Exemplare aus C. edule. Im ganzen entstand der Eindru&, dag C. lamarcki kein idealer Zwischenwirr far GymnophaI- liden-Metacercarien ist. In diesem Zusammenhang sollte erw~ihnt werden, dag die eher marine Art C. edule eine geringere Gefrierresistenz aufweist als C. larnarcki. Bei den Parasiten liegen die Verh~iltnisse iihnlich: Die frostresistenten encystierten Metacer- carien kommen in beiden Wirrsspecies vor, die kiilteempfindlichen Gymnophalliden- Metacercarien bevorzugen dagegen die ebenfalls empfindlichere Muschelart (LAucKN~R, unver6ffentlichte Ergebnisse).
B i o m e t r i e
Die biometrische Analyse der Verteilungskurven von Metacercarien-Durchmessern ergab zu Beginn dieser Untersuchung ein verworrenes Bild und schien die in der Litera- tur allgemein vertretene Ansicht zu best~itigen, nach der eine Bestimmung yon Meta- cercarien an Hand ihres Cystendurchmessers nicht m~Sglich ist.
So zeigten die H~iufigkeitspolygone der Metacercariendurchmesser aus natiir- lichen Infektionen mehr oder weniger ausgepr~igte Abweichungen yon der bei homo- genera Material theoretisch zu erwartenden Normalverteilung. Auf~erdem schwankte der mittlere Durchmesser innerhalb weiter Grenzen. Ein ganz extremes Bild boten die Cysten yon Renicola roscovita. Daraufhin wurde der Versuch unternommen, das Aus- maB der Variabilit~it der Cystendurchmesser experimenteI1 zu bestimmen.
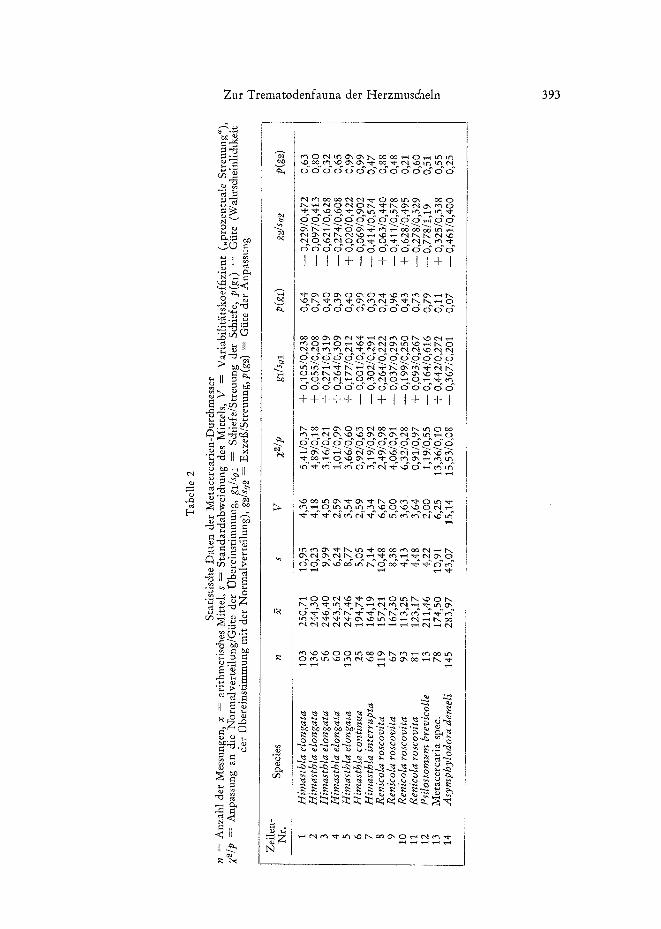
Himasthla elongata: Aus der Ostsee stammende Exemplare mehrerer Mollusken- arten (Littorina, Cardiurn, Mya, Mytilus, Macorna), die keinen natiirlichen Befall auf- wiesen, wurden mit Cercarien yon H. elongata - nach WERDING (1969) der einzigen, im Gebiet in Littorina Iittorea parasitierenden Hirnasthla-Species - bei 30 °/00 und 15 %0 S infiziert. Einem Teil der Cercarien wurde kein Wirtstier geboten; sie encystier- ten sich frei an den Glasw~inden der Versuchsgef~it~e. Der mittlere Cystendurchmesser wurde in verschiedenen Zeitabst~inden nach der Infektion bestimmt. Einige Ergebnisse sind in Tabelle 2 (Zeile 1 bis 5) zusammengestellt. Sie beziehen sich auf folgende Wirts- tier-Salzgehalts-Kombinationen:
Zeile 1: C. edule (Null-Gruppe) bei 30 O/o0 S, Zeile 2: in 30 °/oo S frei encystiert, Zeile 3: L. littorea bei 30'0/00 S, Zeile 4: C. lamarcki (Null-Gruppe) bei 15 °/o0 S, Zeile 5: in 15 °/o0 S frei encystiert.

Tab
elle
1
Ent
wic
klun
gsst
adie
u di
gene
r T
rem
atod
en a
us C
ardi
um
edul
e un
d C
. la
mar
cki
aus
der
Nor
d- u
nd O
stse
e (X
=
vo
rkom
men
d, -
- =
nich
t ge
- fu
nden
, -[
- =
mlt
zie
mli
cher
Sic
herh
eit
nich
t vo
rkom
men
d, ?
=
Ric
htig
keit
pos
itiv
en B
efun
des
frag
lich
)
Par
asit
E
ntw
ickl
ungs
- I
Syl
t/N
ords
ee
stad
ium
[
C.
edul
e C
. la
mar
cki
1 E
chin
osto
mat
idae
H
imas
thla
elo
ngat
a (M
EH
LIS
, 183
1)
Him
asth
la
cont
inua
LO
os-F
RA
NK
, 19
67
Him
asth
la i
nter
rupt
a LO
OS-
Fi~A
NK
, 196
7 H
imas
thla
spe
c.
Ren
icol
idae
Ren
icol
a ro
scov
ita
(STU
NK
ARD
, 1932
)
Mon
orch
iida
e
Asy
mph
ylod
ora
dem
eli
MA
RK
OW
SKI, 1
935
Psi
lost
omat
idae
Psi
lost
omum
bre
vico
lle
(cR
EPL
IN,
1829
)
ency
stle
rte
Met
acer
cari
en
Met
acer
cari
e un
sich
erer
sy
stem
atis
cher
Ste
llun
g:
Met
acer
cari
a sp
ec.
Gym
noph
alli
dae
Mei
ogym
noph
alIu
s m
inut
us (
Co~
B O
LD, 1
859)
! M
eiog
ymno
phal
lus
spec
. I
Gyr
nnop
hall
us g
ibbe
rosu
s LO
os-F
RA
NK
, 197
1 I' I
Gym
noph
allu
s ch
oled
ochu
s OnH
NE
R,
1900
I I
Gym
noph
allu
s sp
ec.
frei
e M
etac
erca
rien
Spo
rocy
sten
m
it C
erca
rien
un
d M
etac
erca
rien
[
X
X
X
X
X
X
X
Fle
nsbu
rger
Ft/
rde
C.
edul
e C
. la
mar
cki
--
X
X
--
X
--
..
..
X
X
X
X
X
Bur
ger
See
C.
lam
arck
i
-I- x X
X
X
_m
X
X
I X
X
X
X
x 12
X X
X
X
?
X
X
X
-I-
X
X
X
X
-i
X
X
b~ t-*
> z t-a
Tab
elle
2
Sta
tist
isch
e D
aten
der
Met
acer
cari
en-D
urch
mes
ser
n =
Anz
ahl
der
Mes
sung
en,
~ =
arit
hmet
isch
es M
itte
l, s
~
Sta
ndar
dabw
eich
ung
des
Mit
tels
, V
=
Var
iabi
lit~
itsk
oeff
izie
nt (
,,pr
ozen
tual
e S
treu
ung"
),
X~/
p ~
Anp
assu
ng a
n di
e N
orm
alve
rtei
lung
/Gii
te d
er l
~be
rein
stim
mun
g, g
l/Sg
l =
Sch
iefe
/Str
euun
g de
r Sc
hief
e, P
(gl)
=
Gii
te
(Wah
rsch
einl
ichk
eit
der
~be
rein
stim
mun
g m
it d
er N
orm
alve
rtei
lung
), g
~/s
~ ~
E
xzef
l/S
treu
ung,
p(g
2) ~
G
iite
der
Anp
assu
ng
N
Zei
len-
N
r.
Spe
cies
n
~ s
V
Z2/p
gl
ls~l
P
(gl)
g~
/s~2
P
(g2)
1 H
imas
thla
eto
ngat
a 10
3 25
0,7t
10
,95
4,36
5,
41J0
,37
+ 0,
105/
0,23
8 0,
64
-- 0
,229
/0,4
72
0,63
2
Him
asth
la e
long
ata
136
244,
30
10,2
3 4,
18
4,89
/0,1
8 +
0,05
5/0,
208
0,79
--
0,0
97/0
,413
0,
80
3 H
imas
thla
elo
ngat
a 56
24
6,40
9,
99
4,05
3,
16/0
,21
+ 0,
271/
0,31
9 0,
40
-- 0
,621
/0,6
28
0,32
4
Him
asth
la e
long
ata
60
243,
52
6,24
2,
59
1,01
/0,9
9 4-
0,2
64/0
,309
0,
39
-- 0
,274
/0,6
08
0,65
5
Him
asth
la e
long
ata
130
247,
46
8,77
3,
54
3,66
/0,6
0 4-
0,1
77/0
,212
0,
40
4- 0
,020
/0,4
22
0,99
6
Him
asth
la c
onti
nua
25
194,
74
5,05
2,
59
0,92
/0,6
3 --
0,0
01/0
,464
0,
99
-- 0
,069
/0,9
02
0,99
7
Him
asth
la i
nter
rupt
a 68
16
4,19
7,
14
4,34
3,
19/0
,92
-- 0
,302
/0,2
91
0,30
--
0,4
14/0
,574
0,
47
8 R
enic
ola
rosc
ovit
a 11
9 15
7,21
10
,48
6,67
2,
49/0
,98
+ 0,
264/
0,22
2 0,
24
+ 0,
063/
0,44
0 0,
88
9 R
enic
ola
rosc
ovit
a 67
16
7,30
8,
38
5,00
4,
06/0
,91
-- 0
,037
/0,2
93
0,96
--
0,4
11/0
,578
0,
48
10
Ren
icol
a ro
scov
ita
93
113,
25
4,13
3,
63
6,32
/0,2
8 --
0,1
99/0
,250
0,
43
4- 0
,628
/0,4
95
0,21
11
R
enic
ola
rosc
ovit
a 81
12
3,17
4,
48
3,64
0,
91/0
,97
4- 0
,093
/0,2
67
0,73
--
0,2
78/0
,529
0,
60
12
Psi
tost
omum
bre
vico
lle
13
211,
46
4,22
2,
00
1,19
/0,5
5 --
0,1
64/0
,616
0,
79
-- 0
,778
/1,1
9 0,
51
13
Met
acer
cari
a sp
ec.
78
174,
50
10,9
1 6,
25
13,3
6/0,
10
4- 0
,442
/0,2
72
0,11
+
0,32
5/0,
538
0,55
14
A
syrn
phyl
odor
a de
mel
i 14
5 28
3,97
43
,07
15,1
4 15
,53/
0,08
--
0,3
67/0
,201
0,
07
-- 0
,461
/0,4
0.0
0,25
N
kao
k~

394 G. LAucKNER
Die Messungen yon Metacercarien aus Mya, Mytilus und Macoma erbrachten die gleichen Resultate; auf ihre Mitteilung kann hier verzichtet werden. Das Ergebnis der statistischen Auswertung der H~iufigkeitsverteilungen I~iflt sich folgendermai~en zusam- menfassen (Tab. 2, Zeite 1 bis 5).
Der Durchmesser der bei experimentellen Infektionen erhaltenen Metacercarien yon Himasthla elongata ist unabh~ingig davon, ob sich die Cystenbildung bei 30 °/00 oder bei 15 °/0o S und im KSrper eines Wirtstieres oder frei im Wasser vollzieht. Es er- folgt kein weiteres Wachstum. Die Ubereinstimmung der empirischen mit der theo- retischen Verteilung ist im ganzen gut (Tab. 2, Spalte Z2/p); auch Schiefe und Exzei~ zeigen keine signifikanten Abweichungen. Die zwischen etwa 2,5 und 4,4 % schwan- kende relative Streuung der Cystendurchmesser (Spalte V) deutet auf eine geringe Variabilit~it hin. Parallelversuche mit Cercarien yon Himasthla continua und H. inter- rupta erbrachten gleiche Ergebnisse. Damit ist der Cystendurchmesser der Metacer- carien ein geeignetes Merkmal zur Unterscheidung der Arten.
Aus den in Tabelle 2 (Zeile I ~';~s 5) wiedergegebenen Einzeldaten ergeben sich fiir H. elongata die gewogenen Werte-~c = 246,70, s = 9,65, V = 3,91 (n = 485). Diese Werte sowie die in Zeile 6 und 7 flit H. continua und H. interrupta gegebenen Daten kSnnen als Referenzstandards verwendet werden.
Der Vergleich der Mittelwerte mittels t-Test ergibt ffir die Kombination H. inter- rupta - H. continua t = 19,63/p = 0,18 • 10 -~ und fiir H. continua - H. elongata t = 26,72/p = 0,72 • 10 -98 und damit extreme Signifikanz. Wegen der gro~en Mittel- wertsunterschiede eriibrigt sich der Test; die Verschiedenheit der Verteilungen ist often- sichtlich (strenggenommen darf der t-Test im vorliegenden Fail gar nicht angewendet werden, denn beim ersten Vergleichspaar liefert der F-Test F = 2,00/p = 0,03 und beim zweiten Paar F = 3,65/p = 0,0002).
Renicola roscovita: W~I~DmG (1969) gibt den Durchmesser yon R. roscovita- Cysten mit 160 bis 180 #m an. Im eigenen Untersuchungsmateriai schwankte er zwi- schen 110 und 180 #m. Die Verteilungskurven yon Cysten aus natiirlichen Infektionen zeigten - je nach Jahreszeit - entweder extreme positive oder negative Schiefe oder aber Zwei- bzw. Mehrgipfligkeit. In allen F~illen lag die prozentuaIe Streuung zwi- schen 6 und 8 (Beispiet: Tabelle 2, Zeile 8).
Bei experimentellen Infektionen wurde am 1. Tag post infectionem ein mittlerer Metacercariendurchmesser yon 113,25#m und am 7. Tag p.i. ein solcher yon 123,27 ~m erhalten (Tab. 2, Zeile 10 und 11). Damit zeigte sich, dab die Metacercarien dieser Art im Wirtstier wachsen und die bei Cysten aus natiirlichen Infektionen erhal- tenen schiefen und mehrgipfligen Verteilungsmuster als das Ergebnis yon Befallssuk- zessionen zu werten sind. Die beste Ubereinstimmung mit der Normalverteilung - und gleichzeitig den hSchsten Mittelwert - weisen die R. roseovita-Metacercarien am Ende einer Befallsperiode auf. Die Daten in Zeile 9 (Tab. 2), die sich auf Messungen vom Winter 1970/71 beziehen, kSnnen als Referenzstandard fiir ausgewachsene Metacer- carien dieser Art betrachtet werden.
Psilostomum brevicolIe: Die Zahl der Beobachtungen (Tab. 2, Zeile 12) ist zu ge- ring, um exakte Aussagen tiber ein eventuelles Wachstum dieser Metacercarien zu er- mSglichen.
Metacercaria spec.: Die Date n in Zeile 13 beziehen sich auf eine Probe aus dem

Zur Trematodenfauna der Herzmuscheln 395
Burger See vom September 1970. Die hohe prozentuale Streuung und die deutliche Ab- weichung yon der Normalverteitung (na& SACHS t969 auf dem 10-°/0-Niveau be- urteilt) sowie die dicht an der Signifikanzschwelle liegende Schiefe sind sichere Indizien dafiir, dag sich die Mehrzahl der vermessenen Metacercarien no& in der Wachstums- phase befand. Metacercaria spec. unterscheidet sich auger durch den etwas grSgeren Durchmesser und eine di&ere Wandung morphologisch nicht yon Renicola roscovita. Es ist an Hand des vorliegenden Datenmaterials nicht auszuschliegen, dag es sich bei Metacercaria spec. um atypische Cysten yon R. roscovita handelt.
Asymphylodora demeti: Die Daten der Zeile 14 beziehen sich ebenfatls auf die Probe aus dem Burger See yore September 1970. Variabititiit, Abweichung yon der Normalverteilung und Schiefe erreichen hier extreme Werte. Auf Grund der voran- gegangenen Betrachtungen ist die Deutung dieser Befunde einfach: Asymphylodora demeli w~ichst im zweiten Zwischenwirt augerordentlich stark. Die hier vorliegende Probe enth~ilt Cysten sehr unterschiedlichen Alters.
Im Fall der drei letztgenannten Arten sind die in Tabelle 2 angegebenen mittleren Cystendurchmesser nur mit Vorbehalt als Referenzstandard verwendbar.
DISKUSSION
In der vorliegenden Arbeit wird versucht, die in Herzmuscheln parasitierenden larvalen Trematoden (a) auf Grund ihres Situs im WirtskSrper, (b) unter Berii&sichti- gung morphologischer Besonderheiten und (c) an Hand meristis&er Merkmale zu un- terscheiden und zu bestimmen. Die Zuverl~issigkeit dieser drei Kriterien und die Frage, ob sie zur Unterscheidung der Arten fiir sich allein oder in Kombination anwendbar und ausreichend sind, ist zu diskutieren.
Zur Frage der Spezialisierung adulter Trematoden auf einen bestimmten Situs in ihrem (Vertebraten-)Endwirt liegen zahlreiche Untersuchungen vor. HOLMES (1971) gibt ein quantitatives, statistisch abgesichertes Beispiel und resiimiert die einschl~igige Literatur. Uber Situspr~iferenzen in den Zwischenwirten existieren verglei&sweise sp~irliche, oft ungenaue und teils widerspriichliche Angaben. Redien und Sporocysten mariner Trematoden parasitieren bis auf wenige Ausnahmen in Mitteldarmdriise und Gonaden des ersten (Evertebraten-)Zwischenwirtes. Metacercarien zeigen im zweiten Zwis&enwirt bereits eine deutlichere Mikrohabitatdifferenzierung. Die strengste Spe- zialisierung findet man bei den Gymnophalliden. Durch zuverl~issige Literaturangaben und eigene Beobachtungen wird betegt, dag Meiogymnophatlus-minutus-Be£all auf einen engbegrenzten Bereich unterhalb des Ligaments yon C. edule bes&r~inkt ist. Auf Grund eigener Untersuchungen an mehr als 8000 I-Ierzmuscheln beider Arten liegen sich fiir die iibrigen, in Cardium des Gebietes vorkommenden Gymnophalliden die Literaturangaben - ,,(frei) zwis&en Mantel und Schale" - pr~izisieren (G. gibberosus: vgl. p. 386, Meiogymnophallus spec.: vgl. p. 385). Zur systematischen Einordnung die- set drei Trematodenlarven ist somit die Situsangabe ausreichend. Es sei betont, dab eine solche Vereinfachung selbstverst~indli& nur ftir das bier bearbeitete Gebiet gilt und Fehlbestimmungen durch das Auftreten weiterer (seltener) Arten in den normalerweise

396 G. LAUCKNER
von den obengenannten Species besetzten /Skologischen Nischen nicht auszuschliel~en sind. Die grot~e Zahl der bier untersuchten Cardien (> 8000) setzt aber die Wahr- scheinlichkeit solcher Fehlbestimmungen sehr stark herab.
Unter den encystierten, im zweiten Zwischenwirt wachsenden Metacercarien sind nut dsymphylodora demeli (Niere) und Psilostomum brevicolle (MitteldarmdriJse, Eingeweidesack) streng lokalisiert. Sie sind auf~erdem ihrem Habitus nach unverwech- selbar. Die beiden anderen Arten dieser Gruppe zeigen eine weniger ausgepr~igte Mikrohabitatpr~ferenz, aber auch hier gibt der Situs die M6gli&keit zur Anwendung einer ,,Differentialdiagnose" : Funde yon Cysten in den Palpen der Muscheln (= Reni- cola roscovita) schlief~en Metacercaria spec. und Psilostornum brevicolle aus, denn beide befallen nie die Palpen. Andererseits kann Renicola roscovita bei starkem Pal- penbefall oder bei Jungmuscheln mit no& zu kleinen Palpen auch in anderen Organen auftreten. Im Bereich des Eingeweidesackes (und nur hier) besteht Verwechslungsgefahr zwischen R. roscovita und Metacercaria spec., wobei die letztere abet im Mittel gr~5t~er und dickwandiger ist. In diesem Fall kann nut die genaue statistische Analyse der Durchmesser-Verteilungskurve Aufschlug tiber eine m/Sgliche Heterogenitiit des Unter- suchungsmaterials geben und als Grundlage zur weiteren Differenzierung dienen. Eine Verwechslung yon im Mantelrand encystierten R. roscovita-Metacercarien mit den un- gef~ihr gteichgrogen Cysten yon Himasthla interrupta ist auszuschliei~en, da sich die letzteren durch ihre Kopfkragenstacheln eindeutig als echinostomatide Larven aus- weisen.
Als problematisch gilt die Gruppe der Himasthla-Arten. Zahlreiche Ver~Sffent- lichungen haben die Verh~ilmisse hier eher verschMert als erhellt, und so ist . . . . . the solution of the problem.., not to be obtained by the reexamination of old material. It is rather to be sought in the application of the experimental method for the measure- ment of specific variation" (STUNKARD 1939, p. 721).
Dieser For&rung ist in der vorliegenden Untersuchung entsprochen worden. Als Ausgangspunkt der experimentellen Arbeiten dienten jeweils die aus den ersten Zwi- schenwirten freigesetzten, an Hand der Angaben yon LOos-FRANK (1967) und WEI~DING (I969) einwandfrei identifizierten Cercarien yon Himasthla interrupta, H. continua und H. elongata. Die Vermessung der nach Infektion der als zweite Zwis&enwirte fungierenden Muscheln und S&necken gewonnenen Metacercarien und die eingehende statistische Analyse der Daten erbrachte den angesichts der gegenteiligen Literatur- angaben tiberraschenden Befund, dai~ die Cystendurchmesser aller drei Arten bei guter f3bereinstimmung mit der theoretisch zu erwartenden Normatverteilung eine nut ge- ringe Variabilit~it aufweisen und daf~ sich die drei Hiiufigkeitsverteilungen praktisch nicht /Jberschneiden, sondern in ihren Randbereichen allenfalls ber~ihren. Weiterhin zeigte sich, dag die Mittelwerte - unabh~ingig yon Satzgehalt und Wirtsart - konstant und die zwischen ihnen bestehenden Unterschiede statistisch hochsignifikant sin&
Damit ist eine sichere Unterscheidung der drei hier zur Diskussion stehenden Himasthla-Arten allein an Hand ihrer Cystendurchmesser m/Sglich. Die Trennung nach diesem Verfahren setzt selbstversfiindlich voraus, daf~ keine weiteren Arten mit 29 Kopfstacheln auftreten, deren Verteilungskurven mit denen der zu trennenden identisch sind oder sich mit ihnen st~irker tiberlappen. Diese Grundvoraussetzung ist im Gebiet erfiJllt: Von den im deutschen Nordseeraum vorkommenden sechs Himasthla-Arten

Zur Trematodenfauna der Herzmuscheln 397
benutzen nur die drei bier aufgefiihrten Species Motlusken als zweiten Zwischenwirt (Loos-FI~ANK 1967).
Das Ergebnis der experimentellen Untersuchungen gestattete auch eine Deutung der bei Messungen an Cysten aus natiirlichen Infektionen anfiinglich auftretenden Un- stimmigkeiten. W~ihrend im Versuch alle drei Arten in guter N~iherung normalverteilte Durchmesser-H~iufigkeitskurven lieferten, wurde in natiirlichen Infekten Normalver- teilung nur bei den Mantelrand-Metacercarien gefunden, deren Durchmesser im Mittel bei 165 #m lag. Die Cysten aus dem Fut~ der Muschelfi ergaben regelm~igig mehr oder weniger deutlich ausgepr~igt negativ schiefe Verteilungeti, deren Mittel bei 240/~m lag. Die Interpretation dieser Befunde ist an Hand des Vorangegangenen einfach" Im Man- telrand encystierten sich nahezu auss&liegli& H. interrup~a~ yon der Hiiufigkeitskurve dieses homogenen Materials war daher auch Normalverteilung zu erwarten. Im Fug encystierte sich neben H. elongata in geringerer Zahl auch H. continua. Die Hetero- genit~it des Materials spiegelt si& in schiefen, manchmal sogar angedeutet bimodalen Verteilungen hoher Variabilit~it wider. Erst nach Vorliegen der experimentellen Daten wurde eine Trennung in zwei Verteilungen m6gli&.
Interessant ist in diesem Zusammenhang, dat~ die beiden im Fug yon Cardiurn nebeneinander vorkommenden HirnasthIa-Arten auch im Endwirt in enger Nachbar- s&att leben. Sowohl H. elongata als au& H. continua parasitieren im Duodenum yon Grotgm~Swen, H. interrupta dagegen im Ileum.
Datg den bisherigen Bearbeitern, die den Cystendurchmesser yon H. eIongata mit 210 his 250 ~m (NmoLL 1906) bzw. 210 bis 270 #m (\V~RDING 1969) angeben, die Heterogenit~it ihres Untersuchungsmaterials entgangen ist, diirtte vor allem an der in der Parasitologie iiblichen Verfahrensweise liegen. Generell werden nicht Mittelwert und Standardabwei&ung aus den Einzelwerten exakt berechnet, sondern lediglich deren Spannweite (range) angegeben - und fast ausnahmslos ohne glei&zeitige Mit- teilung der Anzahl der Beobachtungswerte. Nun hat abet ,,the observed range. . , many drawba&s and is not a good measure of dispersion.., and it may be assumed to be an adequate representation of a distribution and a significant measure of variability, which it is n o t . . . Unless the total frequency is also given, an observed range is thus meaningless" (SIMPSON et al. 1960, p. 80). Abet selbst wenn die Anzahl der Messungen angegeben und damit eine Rii&berechnung der (wahrscheinlichen) Streuung der Daten erm6glicht wird, entgeht die in den Originaldaten enthaltene Information beziigli& Verteilungsmodus, Schiefe usw. ihrer Entde&ung. Die in dieser Arbeit gegebenen Bei- spiele verdeutli&en, dag der Informationsgehalt parasitologis&-meristis&er Daten er- hebli& h~Sher zu bewerten ist, als dies gemeinhin ges&ieht und bei statistisch ad~iquater Bearbeitung zur L6sung taxonomischer Probleme entscheidend beitragen kann.
ZUSAMMENFASSUNG
1. 13 larvale Trematoden werden aus Herzmuscheln yon 3 Stationen der deutschen Nord- und Ostseekiiste beschrieben. Von 9 Arten sind die Entwi&lungszyklen ganz oder teilweise bekannt (in jedem Fall der Adultus). Von 4 Arten, die nicht identifi- ziert wurden, ist mindestens eine unbeschrieben.

398 G. LAUCKNER
2. Die gefundenen encystierten Metacercarien zeigten keine Wirtsspezifit~tt. Die un- encystierten Metacercarien yon Meiogyrnnophallus minutus und Meiogyrnnophallus spec. sind dagegen hochgradig wirtsspezifis& und befallen nur Cardium edule L., ni&t aber C. Iarnarcki REEW.
3. Als Kriterien fiir die Unterscheidung der Arten haben sich als brauchbar erwiesen: (a) der Sitlls im Zwis&enwirt, (b) raorphologische MerkmaIe und (c) meristische Merkmale. Ihre RangfoIge wechselt von Gruppe zu Gruppe: Die Gymnophalliden- Metacercarien kSnnen allein an Hand ihres Situs unterschieden llnd bestimmt werden. Auf die Gruppe Asymphylodora demeli - Renicola roscovita - Psilo- stomum brevicolle - Metacercaria spec. sind im Sinne einer Differentialdiagnose alle drei Kriterien anzuwenden. Die morphologisch nicht unterscheidbaren Cysten der Himasthla-Arten mit 29 Kopfstacheln weisen statistisch hochsignifikante Durch- messer-Unterschiede auf.
4. Es werden die statistischen Maf~zahlen der Metacercarien-Durchmesser mitgeteilt. Sie kSnnen bei Felduntersuchungen als Referenzstandard verwendet werden.
Danksagungen. Frau Dr. B. FRANK (Universit~it Hohenheim) gew~hrte mir freundlicher- weise Einsicht in ihre zur Zeit meiner eigenen experimentellen Untersuchungen noch unver- 5ffentlichten Manuskripte (1970, 1971a, b). I& verdanke ihr weiterhin anregende Diskussionen und die Durchsicht dieses Manuskripts. Die Herren Prof. Dr. F. K~/~c~R (BAH) und D. TIto- M~TSC~t~K (Fa. WANG) gaben mir dankenswerterweise Gelegenheit zur Benutzung ihrer WANG-Elektronenrechner. Die Herren Dr. MICI-tAt~LIS (Forschungsstelle Norderney), Dr. E. ZIEG~LMeIER, E. KOCH und M. S6HL (BAH) versorgten reich mit Material. Herr G. RIECttEY (Campingplatz Wulfener Hals) untersttitzte mich bei meinen Untersuchungen im Burger See. Frau Dipl.-Bibl. I. SCHRITT und Herr Dipl.-Bibl. W. MeIss beschafiten die tells schwer zuggng- liche Literatur. Ihnen allen sei an dieser Stelle herzlichst fiir die Unterstiitzung gedankt.
Z ITIERTE LITERATUR
ANXEL, F., I962. Hydrobia ulvae PENNANT und Hydrobia ventrosa MONTAGU als Wirte lar- valet Trematoden. Vidensk. Meddr dansk naturh. Foren. 124, 1-100.
BARTOLI, P., 1965. D&eloppement exp&imental d'une m&acercaire nouvelle de la famille des Gymnophallidae MoRozov 1955 (Trematoda: Digenea), parasite de lamellibranches matins. Bull. Soc. zool. Ft. 90, 571-580.
BowERs, E. A., 1965. A description of Meiogymnophatlus jarnesoni sp. nov. (Trematoda: Gym- nophallidae) from the intestine of the common scorer, Melanitta nigra L. Ann. Mag. nat. Hist. (Ser. 13) 8, 277-283.
- - & JAM~S, B. L., 1967. Studies on the morphology, ecology and life-cycle of Meiogymno- phallus rninutus (CoB~OLD 1859), comb. nov. (Trematoda: Gymnophallidae). Parasitology ,57, 281-300.
COL~, H. A., 1935. On the larval trematode parasites of the mussel (Mytilus edulis) and the cockle (Cardiurn eduIe), Parasitology 27, 276-280.
- - 1938. On some larval trematode parasites of the mussel (Mytilus edulis) and the cockle (Cardium edule). P. 2. Parasitology 30, 40-43.
COLE, H. E., 1956. Benthos and the shellfish of commerce. In: Sea Fisheries. Ed. by M. GRA- HAM. Arnold, London, 139-206.
HoFv, C. C., 1941. A case of correlation between infection of snail hosts with Cryptocotyle lingua and the habits of the gulls. J. Parasit. 27, 539.

Zur Trematodenfauna der Herzmuscheln 399
HOLMES, J. C., 1971. Habitat segregation in sanguinicolid blood flukes (Digenea) of scorpaenid rockfishes (Perciformes) on the Pacific coast of North America. J. Fish. Res. Bd Can. 28, 903-909.
HUET, L., 1888a. Note sur un parasite nouveau du Cardium edule. Bull. Soe. linn. Normandie (Set. 4) 2, 149-152.
- - 1888b. Nouvelle note sur le Bucephalus hairneanus. Bull. Soc, linn. Normandie 7, 40-41. HUTTON, R. F., 1952. Studies on the parasites of Cardium edule L.: Cercaria fulbrighti n. sp.,
a Gymnophallus larva with a forked tail. J. mar. biol. Ass. U.K. 31,317-326. JAMESON, H. L., 1902. On the origin of pearls. Proc. zool. Soc. Lond. 1, 140-166. - - & NICOLL, W., 1913. On some parasites of the scorer duck (Oederaia nigra) and their
relation to the pearl-inducing trematode in the edible mussel (Mytilus edulis). Proc. zool. Soe. Lond. 12, 53-63.
JOHnSTOnE, J., 1905. Internal parasites and diseased conditions of fishes. Proc. Trans. Lpool biol. Soc. 19, 278-300.
LEBOUR, M. V., 1905. A preliminary note on a trematode parasite in Cardium edule. North- umb. Sea Fish. Rep. for 1904, 82-85.
- - 1907. Larval trematodes of the Northumberland coast. Trans. nat. Hist. Soc. Northumb. 1, 437-454.
- - 1908a. Trematodes of the Northumberland coast. II. Trans. nat. Hist. Soc. Northumb. 3 , 28-45.
- - 1908b. A contribution to the life-history of Echinostoma secundum NICOLL. Parasitology 1, 352-358.
- - 1911. A review of the British marine cercariae. Parasitology 4, 416-456. - - 1914. Some larval trematodes from Millport. Parasitology 7, 1-11. LEsves, M. CH., 1857. Observations sur quelques cercaires parasites de mollusques marins.
Annls Sci. nat. (Ser. 4) 7, 113-117. LOos-FRANK, B., 1967. Experimentelle Untersuchungen fiber Bau, Entwicklung und Systematik
der Himasthlinae (Trematoda, Echinostomatidae) des Nordseeraumes. Z. ParasitKde 28, 299-351.
- - 1968. Der Entwicklungszyklus yon Psilostomum brevicolle (CREVLIN 1829) (Syn.: P. platy- urum [M/~HLING 1896]) (Trematoda, Psilostomatidae). Z. ParasitKde 31, 122-131.
- - 1969. Zur Kennmis der gymnophalliden Trematoden des Nordseeraumes. I. Die Alternativ- Zyklen yon Gymnophallus choledochus ODI~NER 1900. Z. ParasitKde 32, 135-156.
- - 1970. Zur Kenntnis der gymnophalliden Trematoden des Nordseeraumes. IL Lacunoverrnis macomae (LEI3OUR i908) n. comb. (Syn.: Gymnophallus macroporm JAMESON & NICOLL 1913) und seine Metacercarie. Z. ParasitKde 35, 130-139.
- - t971a. Zur Kenntnis der gymnophalliden Trematoden des Nordseeraumes. III. Gymno- phallus gibberosus n. sp. und seine Metacercarie. Z. ParasitKde 35, 270-281.
- - 1971b. Zur Kenntnis der gymnophalliden Trematoden des Nordseeraumes. IV. Dbersicht fiber die gymnophalliden Larven aus Mollusken der Gezeitenzone. Z. ParasitKde 36, 206- 232.
MAI~KOXCSKI, S., 1936. Uber die Trematodenfauna der bahischen Mollusken aus der Umgebung der Halbinsel Hel. Bull. int. Acad. pol. Sci. Lett. (C1. Sci. math. nat., Set. B 2) 2, 285-317.
MARS, P., 1951. Essai d'interpr&ation des formes g6n6ralement group6es sous le nora de Car- dium edule LINNE. Bull. Mus. Hist. nat. Marseille 11, 1-31.
- - 1966. Recherches sur quelques &angs du littoral tvl6diterran6en fran~ais et sur Ieurs faunes malacologiques. Vie Milieu (Suppl. Bd) 20, 1-359.
NICOLL, W., 1906. Notes on trematode parasites of the cockle (Cardium edule) and mussel (Mytilus eduIis). Ann. mag. nat. Hist. (Set. 7) 17, 147-155.
PALOMUI, A., 1934. Gli stadi larvali dei trematodi del Golfo di Napoli. I. Contributo allo studio della morfologia, biologia e sistematica delle cercarie marine. Pubbl. Staz. zool. Napoli 14, 51-94.
PELSrN~R, P., 1906. Tr6matodes parasites de mollusques marins. Bull. Sci. Fr. Belg. 40, 161-186.

400 G. LAUCKNER
PREVOT, G., 1965. Ddvdoppement exp&imental d'une m&acercaire de GymnophaUus (Trema- toda: Digenea). Bull. Soc. zool. Fr. 90, 45t-456.
REBECQ, J. & PREVOT, G., 1962. Ddveloppement exp&imental d'un Gymnophallus (Trematoda: Digenea). C. r. hebd. S~anc. Aead. Sci., Paris 225, 3272-3274.
REEs, F. G., 1939. Cercaria strigata L~BOUR from Cardiurn edule and TelIina tenuis. Parasito- logy 81,458-463.
REIMm~, L. W., 1970. Digene Trematoden und Cestoden der Ostseefische ais natiirliche Fisch- marken. Parasit. SchrReihe 20, 1-144.
SACHS, L., 1969. Statistische Auswertungsmethoden. Springer, Berlin, 671 pp. SIMPSON, G. G., ROE, A. & LEWONTIN, R. C., 1960. Quantitative Zoology. Harcourt, Brace
& Co., New York, 440 pp. STUNKAI~D, H. W., 1939. Determination of species in the trematode genus Hirnasthla. Z. Para-
sitKde 10, 719-721. - - & UZMANN, J. R., 1958. Studies in digenetic trematodes of the genus Gymnophallus and
Parvatrerna. Biol. Bull. mar. biol. Lab., Woods Hole 115, 276-302. VILLOT, F. C. A., 1878. Organisation et d~veloppement de quelques esp~ces de trematodes
endoparasites marins. Annls Sci. nat. (Ser. 6) 49, 1-40. WERmN% B., 1969. Morphologie, Entwi&lung und iDkologie digener Trematoden-Larven der
Strandschnecke Littorina Iittorea. Mar. Biol. 3, 306-333. WI~Gt~N, B. J., 1956. Studies on Finnish larval flukes with a list of known Finnish adult
flukes. Acta zooI. fenn. 91, 1-t06.
Ans&rift des Autors: G. LAUCKNER Biologische Anstalt Helgoland (Zentrale) 2 Hamburg 50 Patmaille 9 Bundesrepublik Deutschland


























![[Canada Research] Cardium Junior Oil Amp Gas Producers](https://img.pdfslide.net/doc/110x75/55cf9945550346d0339c84de/canada-research-cardium-junior-oil-amp-gas-producers.jpg)

