Embed Size (px)
Citation preview

1 Acts Biotechnol. 7 (1987) 2, 147-155 I
Zur Wechselwirkung zwischen einer Penicillium-Cellulase und wasserloslichen Cellulosederivaten
KASULKE, U.l, PHILIPP, B.1, POLTER, E.B
Akademie der Wissenschaften der DDR Institut fiir Polymerenchemie KantstraBe 55, Teltow-Seehof, 1530 DDR Akademie der Wissenschaften der DDR Institut fiir Biotechnologie PermoserstraBe 15, Leipzig, 7050 DDR
Summary
Employing anionic and non-ionic cellulose ethers, differing in type of substituent and degree of substitution, as substrates, the pH-profile of enzyme activity and the parameter K , and V,,, of the MICHAELIS-MENTEN kinetics have been determined with Penicillium citrioviride cellulase in an homogeneous system. Within rather wide limits, a linear correlation was found between the DS of the substrate and K, or Vm,,, respectively. Also the pH-profile was found to depend on DS main- ly, being bimodal at higher DS. On the other hand, no significant difference was observed between anionic and non-ionic cellulose ethers of about the same DS with regard to the parameters deter- mined. It is assumed, that substrate-enzyme interaction is governed mainly by the length of non- derivatized chain sequences mainly and not by Coulomb interactions.
In fruheren Arbeiten hatten wir uber den Einflufi von Substitutionsgrad (DS) und Ar t des Substituenten wasserloslicher Cellulosederivste auf deren homogene enzymatische Hydrolyse durch Kulturfiltrate von Trichodem &ride und Gliocludium sp. berichtet [l--31: Unsere in [4] zusammengefafiten Versuche zum EinfluB der Ar t des Substituen- ten, insbesondere seines Lsdungszustandes, auf die Enzym-Su bstrat-Wechselwirkung durch Sorptionsversuche mit PeniciUium citrioviride-Cellulase an oberflachenmodifi- zierten Cellulosepartikeln unter Heterogenbedingungen, hatten wegen uberlagerung solcher Substituenteneffekte mit dem dominierenden EinfluB einer veranderten physi- kalischen Struktur der Substratoberflache nur,zu recht begrenzten Aussagen gefuhrt. In weiteren Untersuchungen mit Gliocladium sp.-Cellulase hatten wir gepriift, wie sich bei verschiedenen wasserloslichen Cellulosederivaten die Art des Substituenten und der Substitutionsgrad auf die Aktivitat des Enzymkomplexes auswirken [5]. Die Lage des pH-Optimums wurde bei diesem Cellulasesystem durch den Ladungszustand des Sub- strates beeinflufit. Im folgenden Beitrag werden Ergebnisse zur Bestimmung der Konstanten der IMICHA- Em-MENTEN-Kinetik (K, und v,,,) fiir eine grofiere Anzahl verschiedenartiger Cellu- losederivate dargestellt und hinsichtlich des Einflusses von DS und Art des Substituen- ten diskutiert, wobei wir als Enzymsystem die Cellulase von Penicillium cit~ioviride ein- setzten. Zur Festlegung optimaler Versuchsbedingungen fur die Ermittlung von K , und V,,, bestimmten wir das pH-Profil der Enzymmirkung gegeniiber den verwende- ten Cellulosederivaten.
14.8 Acts Biotechnol. 7 (1987) 2
Versuchsmethodik Als Substrate dienten die in Tabelle 1 aufgefuhrten wasserfoslichen Cellulosederivate im DS-Be- reich 0,2 bis 1,8, wobei es sich zumeist um im Laboratorium hergestellte Praparate handelte. Als EnzymlBsung stand ein Kulturfiltrat von Penicillium citrioviride [6] zur Verfiigung. Zur Bestim- mung des pH-Profils der Enzymwirkung wurde dieses Filtrat in einer Verdiinnung von 1 : 50 ein- gesetzt, wobei der pH-Wert des Reaktionssystems mit 0,l M Citronensaure-Phosphat-Puffern im pH-Bereich von 3- 10 variiert wurde. Die Versuche wurden mit einer Substratkonzentration von 1% bei 40 "C 0,l "C durchgefiihrt. Nach Vortemperieren der entsprechenden Losungen wurde die Hydrolyse durch Zugsbe der Enzymlosung gestartet und nach 30 min Reaktionszeit mit Fem- cyanidreagens [7] bzw. - im Falle einer sehr schnellen Bildung reduzierender Zucker bei den nied- rig substituierten Proben - mit 0,5 N Natronlauge abgebrochen.
Tabelle 1. pH-Optima der Cellulase von Penicillium citrioviride bei verschiedenen Cellulosederivaten
Probe DS pH-Optimum
Carboxymethylcellulose (CNC) 0,63 3 3 Carbocymethylcellulose (CMC) 0.70 3.8 Carboxymethylcellulose (CMC) 1,oo 4,2 und 7,65 Carboxymethylcellulose (CNC) 1,36 4,2 und 7,50 Carboxyethylcellulose (CEC) 0,30 3.8 Carboxyethylcellulose (CEC) 0,52 3 3 Cellulosesulfat (CS) 0,20 323 Cellulosesulfat (CS) 0,50 3,8 Hydroxyethylcellulose (HEC) - 0,3 3.8 Hydroxyethylcellulose (HEC) - 0,9 4,3 und 7,65 kationisierte Hydroxyethyl- - 0,3 3.8 cellulose (K-HEC) Methylcellulose (MC) - 1,4 4,O und 7,65 Hydroxypropylcellulose (HPC) - 1,8 4,s und 8.00
Die Parameter der bf.ICHAELrs-MEnTEx-Kinetik (K, und v,,,) errnittelten wir fur jedes Substrat sowohl beim wirkungsoptimalen pH-Wert als auch bei einem konstanten Wert von pH = 5. Ziir Festlegung geeigneter Reaktionsbedingungen (,,linearer Hydrolyseverlauf") wurde jeweils die Zeitabhangigkeit der RS-Bildung bei einer Verdunnung des Kulturfiltrates auf 1 : 50, 1 : 100 und 1 : 250 verfolgt. Zumeist reichte eine Verdiinnung von 1 : 100 aus, um bei einer Reaktionszeit von
-0,s 0 0,s 1 S - l l . g - ' l
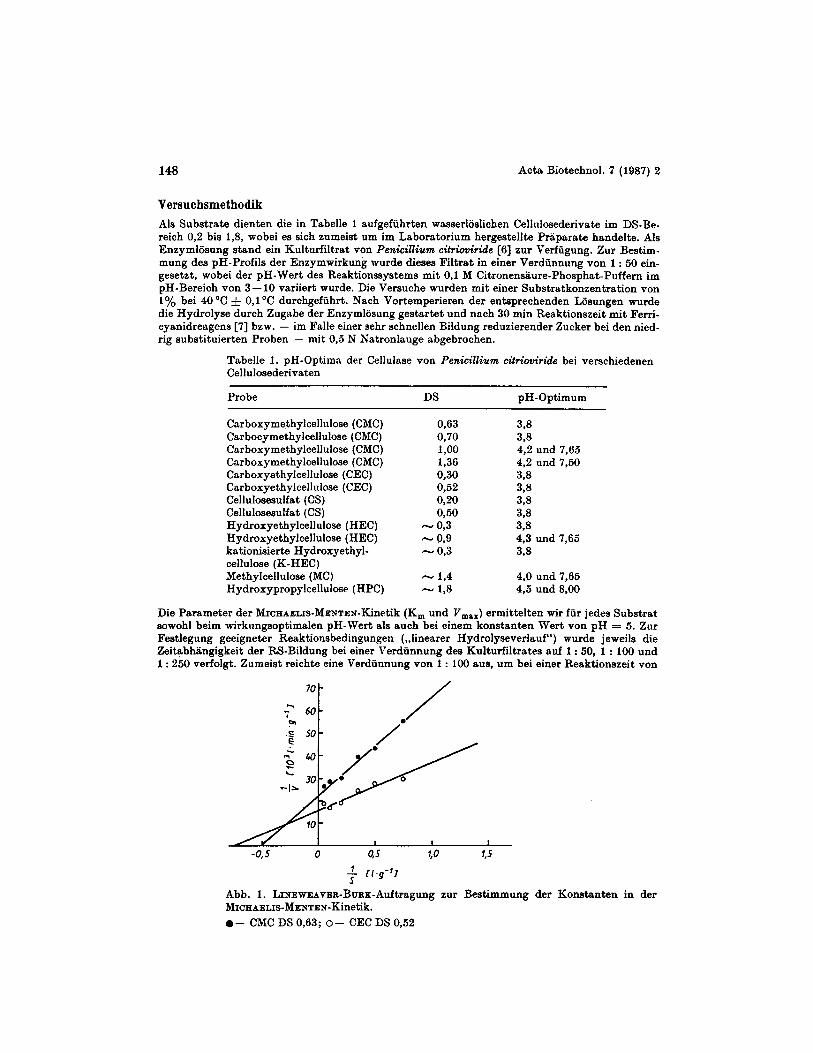
Abb. I. LINEWEAVER-BnRK-Auftragung zur Bestimmung der Konstanten in der MICHAELIS-MENTEN-Kinetik. 0 - CMC DS 0,63; O - CEC DS 0.52
KASIJLKE, U., PHLLDPP, B. u. a., Penicillium-Cellulase und wasserliisliche Cellulosederivate 149
30 min 10% des maximal erreichbaren RS-Wertes nicht zu uberschreiten. Die Bestimmung von K , und V,,, erfolgte zumeist unter Verwendung von 5 verschiedenen Substratkonzentrationen. Die Auswertung nahmen wir nach LINEWEAVER-BURK [S] vor (Abb. 1).
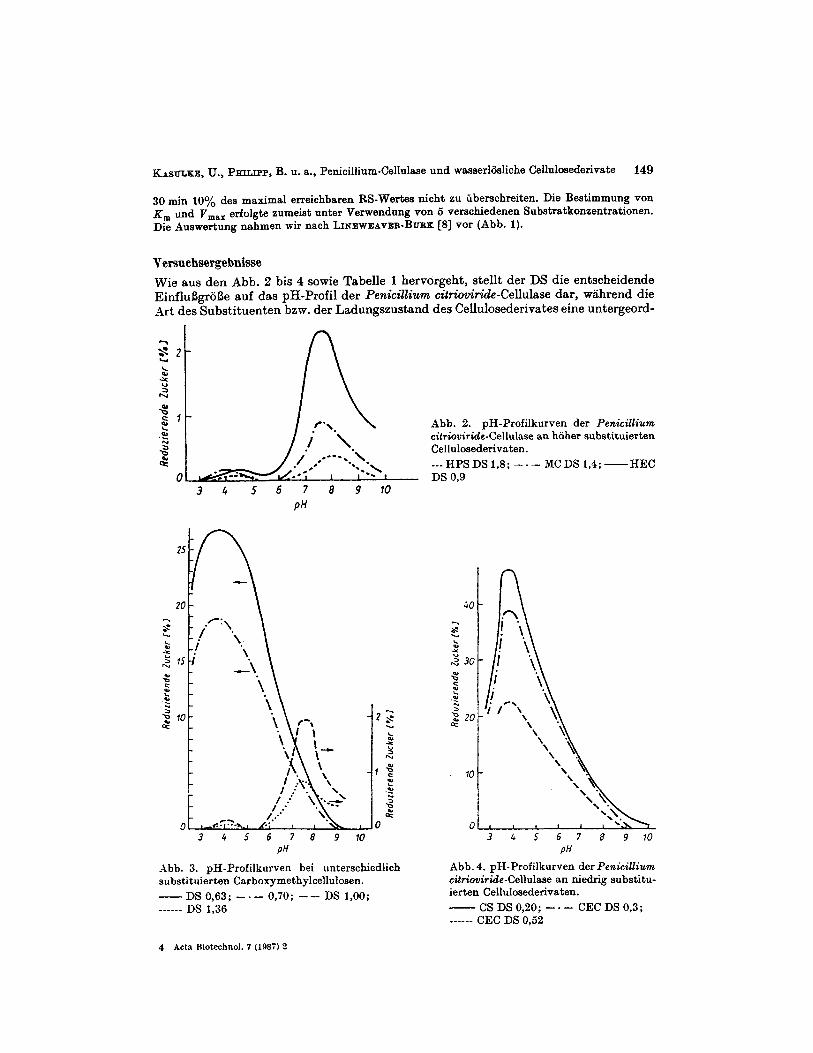
Versuehsergebnisse Wie aus den Abb. 2 bis 4 somie Tabelle 1 hervorgeht, stellt der DS die entscheidende EinfluSgroSe auf das pH-Profil der Penicillium citn'oviride-Cellulase dar, wahrend die Art des Substituenten bzm. der Ladungszustand des Cellulosederivates eine untergeord-
c al
Abb. 2. pH-Profilkurven der Penicillium citriouiride-Cellulase an hoher substituierten Cellulosederivaten.
DS 0,9
al '- w 5
---HPSDSl,S; -.- MCDS 1,4;-HEC
3 4 5 6 7 8 9 1 0 PH
3 4 5 6 7 8 9 10 PH
Abb. 3. pH-Profilkurven bei unterschiedlich substituierten Carboxymethylcellulosen.
_ - _ _ - _ DS 1,36 - DS 0,63; -. - 0,70; - - DS 1,OO;
3 4 5 6 7 8 9 1 0
Abb. 4. pH-Profilkorven der Penicillium cit7iouiride-Cellulase an niedrig substitu- ierten Cellulosederivaten.
___--- CEC DS 032
PH
- CS DS 0,20; - - CEC DS 0,3;
4 Acta Riotechnol. 7 (1987) 2

150 Acts Biotechnol. 7 (1987) 2
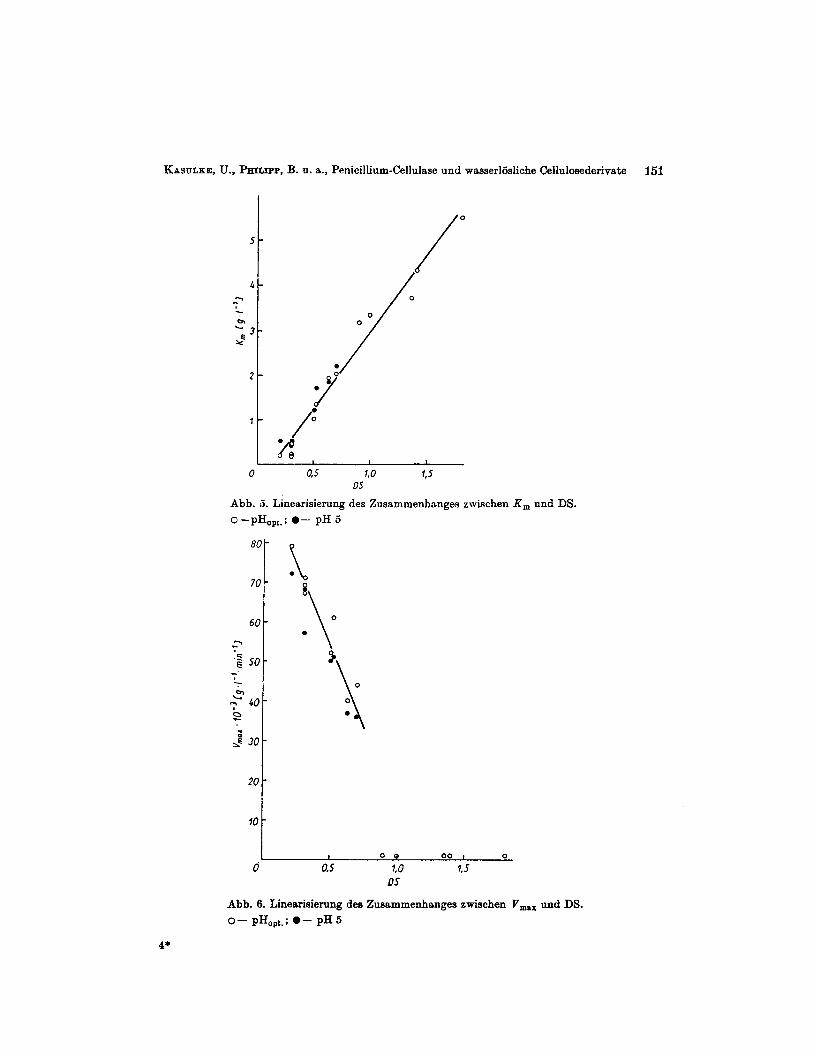
nete Rolle spielen. Bei DS-Werten oberhalb von 0,9 wird bei nichtionischen ebenso wie bei anionischen Substraten, eine bimodale Kurve fur die Abhangigkeit der Enzymwir- kung vom pH-Wert des Mediums erhalten, wobei das en te Wirkungsmaximum bei p H 4,O bis 4,5, das zweite zwischen pH 7,5 und pH 8,O lag (siehe Abb. 2 und 3). Eine gewisse Abhangigkeit der Lsge der beiden Wirkungsmaxima von der Art des Substituenten ist nicht auszuschlieBen, lieB sich jedoch unter Beriicksichtigung der fur hoher substituierte Proben groBeren Fehlerbreite der Messungen nicht sicher nachweisen. Diese Feststellung gilt zugleich hinsichtlich eines moglichen Einflusses des Ladungszustandes des Substra- tes auf das pH-Profil der Enzymwirkung, da die geringfugige pH-Verschiebung in Richtung des alkalischen Bereichs bei den beiden Wirkungsmaxima der Hydroxypropyl- cellulose im Vergleich zu den anderen verwendeten Cellulosederivaten mit DS- Werten 2 0,9 noch nicht als signifikant anzusehen ist. Bei niedrig substituierten Substratproben (DS 5 0,7) findet man hingegen nur ein Wirkungsmaximum (Abb. 3 und 4). Dieses liegt unabhangig von der Art des Substituenten, des Mungszustandes dks Derivates und des DS der Proben bei pH 3,s f 0,l. Die pH-Profile der Kurven weisen annahernd eine symmetrische Form auf. Bei den hoher substituierten Derivaten mird ein ausgepragtes Wirkungsminimum des hier untenuchten Cellulasesystenis im pH-Bereich vori 5 bis 6 beobachtet (Abb. 2 und 3). Die nach 30 min Reaktionszeit gebildete Menge an redu- zierenden Zuckern im pH-Optimum nimmt verstiindlicherweise niit steigendem DS ab. Dies stimmt mit friiher publizierten Ergebnissen uberein, bei denen nach 100 h Hydro- lyse mit Cellulase von Trichoderma viride [l] und Gliocladium sp. [2, 31 eine starke Ab- hangigkeit der maximal gebildeten RS- bzw. Glucosemenge vom DS festgestellt wurde. Die Parameter der MICHAELIs-IMENTE~-Kinetik ( K , und v,,,) wurden im pH-Optimum und bei dem in der Literatur oft angewandten pH-Wert von 5,O ermittelt. Fur die hoch- substituierten Cellulosederivate (DS 2 0,9), die bimodale pH-Kurven aufwiesen, be- stimmten wir die Konstanten der iMICHIELTs-PUIENTEN-Kinetik bei den iin sauren Bereich liegenden Maxima. Auf Grund des ausgepragten Wirkungsminiiiiunis bei pH 5 konnten wir keine Konstanten erhalten. Wie aus Tabelle 2 und Abb. 5 hervorgeht, zeigen die K,"- Werte eine lineare Zunahlne mit steigendem DS (Korrelationskoeffizient r > 0,98). Ein EinfluB der Art des Substituenten bzw. des Ladungszustandes des Derivats war nicht nachweisbar. Auch fur die maximale Abbaugeschwindigkeit V,,, ist als entscheidende EinfluBgroSe die Hohe des DS und nicht die Art des Substituenten anzusehen (vgl.
Tabelle 2. Konstnnten der Michaelis-Menten-Kinetik der Celliilase von Penicillium citriociride bei verschiedenen Cellulosederivaten
Probe DS Km v * a x [g/ll. . g/1 . min]
PH,,,. PH 5 PHOP'. PH 5
CMC CMC CMC CMC CEC CEC cs cs HEC HEC
MC HPC
K-HEC
0,63 1,92 0,70 2,03 1 9 0 0 3,35 1,36 3,72 0,30 0,46 0,52 1.34 0,20 0,20 0,50 1,03 - 0,3 0,19 - 0.9 3,18 - 0.3 0,21 - 1.4 4,32 - 1.8 5,50
1.84 2,20 - - 0.43 1,71 0,52 1,21 0,42
0,48 -
40 43 1 1
67 61 79 52 69 1
71 1 1
37 36
- 57 51 72 50 67
68 -
-
KASULKE, U., PHILIPP, B. u. a., Penicillium-Cellulase und wasserlijsliche Cellulosederivate 151
5 -
c - - h c -
c. 3 - 2
2 -
1 -
/ O
// 0
yo 1
2 , I t
I 0 0 00 I 0
0.5 t 0 1.5 DS
R, und DS.
Abb. 6. Linearisierung des Zusammenhanges zwischen V,,, und DS. 0- pHopt.; 0- pH 5
4+

152 Acte Biotechnol. 7 (1987) 2
Tabelle2, Abb. 6). Wahrend der lineare Zusammenhang zwischen K , und DS im Wir- kungsoptimum des Enzyms fur den gesamten hier betrachteten DS-Bereich beobachtet wurde, war eine lineare Korrelation zum DS fur V,,, nur bis zu DS-Werten von etwa 1,0 feststellbar (Korrelationskoeffizient r > 0,96). Bei weiterer Erhohung des DS trat keine signifikante Anderung von V,,, auf, wobei allerdings auch hier die mit steigendem DS wachsende Fehlerbreite der Bestimmung in Betracht zu ziehen ist. Soweit bei pH 5 entsprechende Aussagen moglich waren, wurde hier ebenfalls ein linearer Zusammenhang zwischen K , bzw. V,,, und dem DS gefunden. Die K,-Werte lagen fur die einzelnen Substratproben generell etwas hoher als im Wirkungsoptimum ; fur V,,, war eine unterschiedlich starke Abnahme beim Wechsel vom optimalen pH zu pH 5 festzustellen (siehe Tabelle 2).
Zusammenfassende Diskussiori und SchluUfolgerungen
Die hier dargestellten Ergebnisse sprechen recht eindeutig dafiir, da13 fur das Cellulase- system von Penicillium citrioviride sowohl daspH-Profil der Enzymwirkung als auch die Konstanten der MICHAELIS-MENTEX-Kinetik ( K , und V,,,) entscheidend vom DS des wasserloslichen Cellulosederivats bestimmt werden. Die chemische Struktur des Sub- stituenten bzw. der Ledungszustand des Derivats spielen allenfalls eine recht unterge- ordnete Rolle. Wahrend unsere fruheren Untersuchungen zum pH-Profil der Erizymwirkung des Cellu- lasesystems von GlioclcGdium sp. gegenuber verschiedenen nichtionischen und anionischen Cellulosederivaten eine deutliche Ahhiingigkeit des wirkungsoptimalen pH- Wertes vom DS und Ladungszustand bei Carboxymethyl- und Carboxyethylgruppen tragenden l)eri- vaten ergebeii hatten [5], fanderi wir bei den hier durchgefuhrten Untersuchungen mit Penicillium citrioviride-Cellulase fur eine grol3ere Zehl loslicher Cellulosederivate eine Konstanz des wirkungsoptimalen pH- Wertes im vergleichbareii DS-Bereich. Eine bi- modale pH-Wirkungskurve, wie wir sie fiir PeniciUiurn citrioviride im Falle hoher sub- stituierter Substratproben fanden, wurde friiher von REESE [9] fur Penicillium pusillurn beobachtet, wobei ein DS-Wert fiir die von ihm verwendete CMC-Probe nicht angegeben wurde. Auch andere Autoren fanden hei den Cellulasen verschiedener Mikroorganismen und Molusken bi- z. T. auch trirnodale pH-Profile. Zum Beispiel weist die Cellulase von iklyu arenariu bei der Bestimmung der CMC-sse Aktivitat em trimodales pH-Spektrum auf [lo]. Gegenuber Filterpapier konnte bei Poronia oedipus eine bimodale Aktivitats- kurve festgestellt werden, die C,-Aktivitat zeigte diese Erscheinung nicht [ 11 1. Ahnlich unterschiedliches Verhalten zeigte die Cellobiohydrolase-Komporiente von Penicillium funiculosum-Cellulase. Gegeniiber diirch Phosphorsaure gequollener Cellulose wurde eine bimodale, bei Avicel eirie unimodale pH-Aktivitgtskurve erhalten [ 121. Bei unseren fruheren Untenuchungen mit Gliocladium sp.-Cellulase deutet sich im pH-Bereich von 2 bis 7 in keinem Falle ein bimodales A4ktivitatsprofil an, wobei allerdings die dort betrachteten Proben - abgesehen von einer CMC mit DS = 1,0 - nur niedrige DS- Werte 5 0,7 aufwiesen, die auch mit Penicillium citrioviride-Cellulase zu unimodalen pH-Wirkungs-Kurven f uhrten. Zur Deutung der bimodalen pH-Aktivitatskurve bei Penicillium-Cellulase nahmen wir in Ubereinstimmung mit Vontellungen von REESE [9] an, da13 mindestens 2 verschie- dene Enzymkomponenten mit unterschiedlichen optimalen pH-Werten wirksam werden, von denen sehr wahncheinlich die eine mit dem Wirkungsoptimum im sauren Bereich bevorzugt Bindungen zwischen unsubstituierten Anhydroglucoseeinheiten spaltet, wah- rend die andere mit einem Wirkungsoptimum im schwach alkalischen Bereich moglicher- weise auch Bindungen zwischen einer unsubstituierten und einer substituierten Einheit angreift .

KASULKE, U., PHILIPP, B. u. a., Penicillium-Cellulase und wasserlosliche Cellulosederivate 153
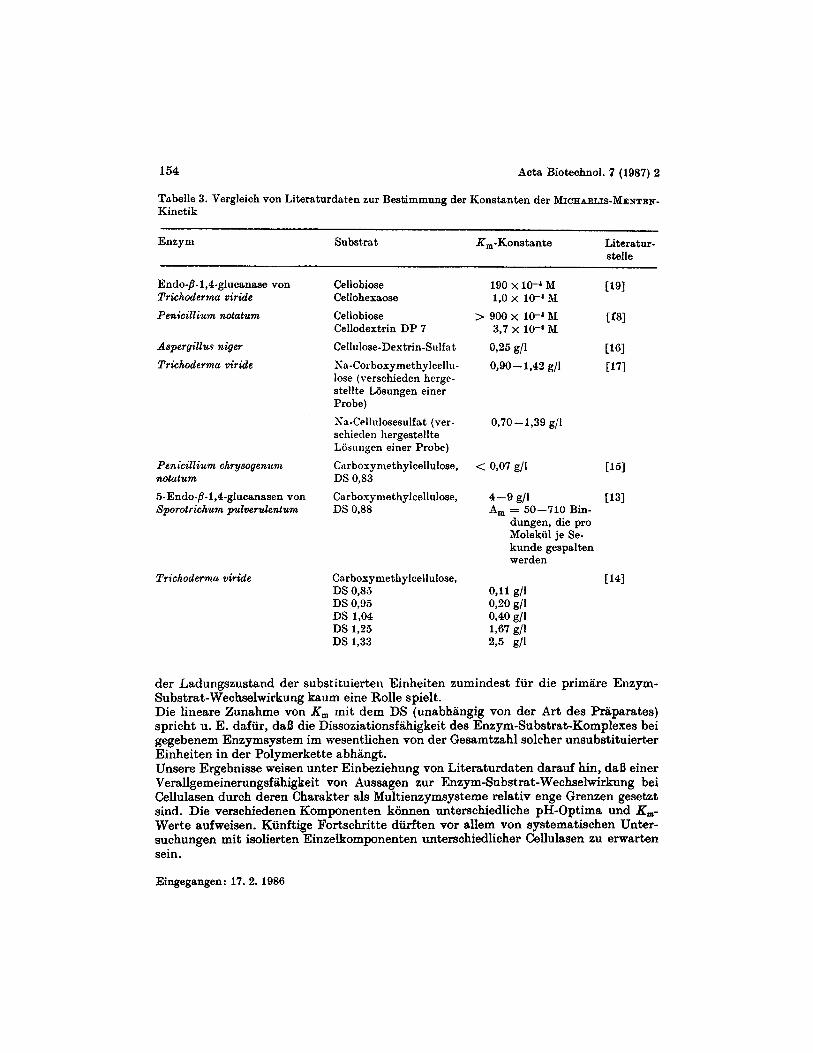
Bei einer Probe mit einem DS von 0,63 waren bei gleichmafliger Verteilung der Substi- tuenten analog SYURLIN [20] theoretisch 46,3y0 Glucose moglich, von denen man 34,4% in Sequenzen von mindestens Diadenlange und 22,7% in Sequenzen von mindestens Triadenlange erwarten konnte. Bei einem DS von 1,OO reduziert sich die Glucosemenge auf 29,3yo, von denen nur noch 14,8y0 in Sequenzen von mindestens Diadenlange und 6,2% von mindestens Triadenlange vorlagen, wenn man annimmt, daB die unsubsti- tuierte Glucose nach einer BERNommI-Statistik entlang der Cellulosekette verteilt ist [21]. Mit ansteigendem DS der zu untersuchenden Proben verringert sich also standig der pro2entuale Anteil unsubstituierter Glucose und damit auch die Menge der zu erwar- tenden unsubstituierten Glucosesequenzen unterschiedlicher Lange. Im Falle niedrig substituierter Substratproben mit DS-Werten 5 0,7, bei denen ein unimodales pH-Profil mit einem Maximum bei pH = 3,8 beobachtet wird, fallt offenbar nur der sich durch Spaltung langerer unsubstituierter Sequenzen ergebende Anteil fiir den RS-Wert ins Gewicht, das Wirkungsoptimum wird durch die*diesen ProzeI3 totali- sierende Enzymkomponente festgelegt. Die Annahme eines Wirksamwerdens unterschiedlicher Enzymkomponenten mit unt er- schiedlichen wirkungsoptimalen pH-Werten 1aBt auch ein unterschiedliches pH-Yrofil der Enzymwirkung fur die Cellulasen verschiedener Mikroorganismen - z. B. von Gliocladium sp. einerseits, Penicillium citrioviride andererseits - im Prinzip verstandlich werden. Zur Klarung der Verallgemeinerungsfahigkeit und Systematisierbarkeit solcher Aussa- gen erscheinen allerdings weitere experimentelle Untersuchungen an venchiedenart igen Cellulasesystemen bzw. deren isolierten Einzelkomponenten unter Verwendung eines vorher festgelegten Substratspektrums notwendig. Einer Diskussion der von uns ermittelten MrCHAELIs-MENTEN-Parameter sol1 eine kurze Betrachtung der aus der Literatur zuganglichen Angaben fur K , und V,,, anhand von Tabelle 3 vorangestellt werden, in die auch einige Werte fur die sogenannte ,,molekulare Aktivitat" A,, d. h. die Anzahl der pro Enzymmolekul je Sekunde gespaltenen Bindun- gen, mit aufgenommen wurden. Nach Auffassung von ERIKSSON u. a. [13] ermoglicht AJI oft eine bessere Differenzierung zwischen verschiedenen Enzymsystemeri bzw. Enzymkomponenten als die alleinigen Angaben von K,.Vergleichsweise wurden in die Tabelle auch einige K,-Werte fiir unsubstituierte, wasserlosliche Oligomere der Cellulose aufgenommen. Aus den Zahlenwerten der Tabelle 3 ist u. a. zu entnehmen, daB
- im Oligomerbereich K , mit steigendem D P abnimmt, d. h. die Bindungsstarke irn Enzym-Substrat-Komplex ansteigt ;
- von ERIKSSON u. a. auf viskosimetrischem Wege bereitsfriiher [ 141 eine Zunahme von K, rnit steigendem DS der als Substrate eingesetzten Carboxymethylcelluloseii fiir Trichoderma vide-Cellulase festgestellt wurde ;
- verschiedene aus einem Enzymkomplex von Sporotrichum pulverulentum isolierte Endoglucanasen sich in ihren K,-Werten erheblich unterscheiden konnen.
Die von uns uber eine RS-Bestimmung fur Penicillium citrioviride gefundene lineare Abhangigkeit zwischen K,,, und dem DS einer grol3eren Anzahl verschiedener Cellulose- derivate bestatigt als Trendaussage die von EBIESSON mit Trichodermu vide-Cellulase gefundene Zunahme von K, mit steigendem DS der als Substrat eingesetzten CMC-Pro- hen. Eine quantitative ubereinstimmung war aufgrund der unterschiedlichen Enzym- systeme nicht zu erwarten und wurde auch nicht gefunden. Insgesamt fuhren unsere Ergebnisse am Cellulasesystem von Penicillium citrioviride zu dem SchluB, daS fur die Wechselwirkung im wesentlichen die Anzahl und Lange un- substituierter Kettensequenzen mal3gebend sind, wahrend die chemische Struktur u n d
154 Acta Biotechnoi. 7 (1987) 2
Tilbelle 3. Vergleich von Literaturdaten zur Bestimmung der Konstanten der MICHAELIS-MENTEN- Kinetik
Enzyni Substrat R,-Konstante Literntur- stelle
Endo-,9-1,4-glucanarre von Cellobiose Trichoderma viride Celloheraose
190 x lo-' &I ~ 9 1 l,o x 10-4 31
Penicillium notatum Cellobiose > 900 x lo-' DI [f81
Aspergillm niger Cellulose-Destrin-Sulfa t 0,25 g/1 C1GI
Triehoderma viride ~a-CorbosymethylceIlii- 0,90 - 1,42 g/1 r171
Cellodextrin DP 7 3,7 x 10-4 31
lose (verschieden hergr- stellte Losungen einer Probe)
schieden liergestellte Losungen einer Probe)
Sa-Cellnlosesulfat (ver- 0,70-1,39 g/1
Pen icillium chr ysogenuni Carbovymethylcellulose, < 0,07 g/l ~ 5 1
5-Endo-,9-1,4-glucanasen von Carboxyniethylcellulose, 4-9 g/l 1131
notutum DS O,S3
Sporotrichum pulverulentum DS 0,88 A, = 50-710 Bin- dungen, die pro Molekiil je Se- kunde gespnlten werden
Trichodernia viridz Csrboryme thy lcellulose, [I41 DS 0,85 O J 1 g/l DS 0,95 0,20 g/l DS 1,04 0,40 g/l DS 1,25 1,67 g/l DS 1,33 2 3 E/I
der Ladungszustand der substituierteri Einheiten zumiridest fur die primare Enzym- Substrat-Wechselwirkung kaum eine Rolle spielt. Die lirieare Zunahme von K, mit dem DS (unabhiingig von der Art des Mparates) spricht u. E. dafur, daB die Dissoziationsfahigkeit des Enzym-Substrat-Komplexes bei gegebenem Enzymsystem im wesentlichen von der Gesamtzahl solcher unsubstituierter Einheiten in der Polymerkette abhiingt. Unsere Ergebnisse weisen unter Einbeziehung von Literaturdaten darauf hin, daB einer Verallgemeinerungsfiihigkeit von Aussagen zur Enzym-Substrat-Wechselwirkung bei Cellulasen durch deren Charakter als Multienzymsysteme relativ enge Grenzen gesetzt sind. Die verschiedenen Komponenten konnen unterschiedliche pH-Optima und Km- Werte aufweisen. Kiinftige Fortschritte diirften vor allem von systematischen Unter- suchungen mit isolierten Einzelkomponenten unterschiedlicher Cellulasen zu erwarten sein.
Eingegangen: 17.2.1986

KASULKE, U., PHILIPP, B. u. a., Penicillium-Cellulase und wasserlosliche Cellulosederivate 155
Li teratur
[l] PHILIPP, B., KASULKE. U., LCKANOFF, B., JACOPIAN, V., POLTER, E.: Acta Polymerica 33
[2] PHILIPP, B., KASULKE, U., DAUTZENBERG, H., POLTER, E., H ~ ~ B E R T , S.: $cta Polymerica 34
[3] KASULKE, U.. DAUTZENBERG, H., POLTER, E., PHILIPP, B.: Cell. Chem. Technol. 15 (1983),
[4] PHILIPP, B., KASULKE, U., SCHCLZ, W., HIRTE, W.: Actit Biotechnol. (im Druck). [a] POLTEH, E., KASULKE, U.. PHILIPP, B.: Acta Biotechnol. 4 (1984), 347. [6] HIRTE, W. F., GLATHE, I.: WPC 12 D/189403. [7] GHOSE, T.: Measurement of celltilase activity (Biotechnology Commission, International
[8] HOFMANN, E.: Dynamische Biochemie, Teil 2, Berlin: Skademie-Verlng, 1979. [9] REESE, E. T., GILLIOAX. W., NORKRANS, B.: Physiol. Plant. 5 (1952)’, 379.
(1982). 714.
(1983), 651.
423.
Union of Pure st Applied Chemistry, July 1980).
[lo] MIRZA. M., SERBAN, M.: Rev. roum. Biochim. 18 (1981), 33. [ll] DENISON, D. 8.: Mycologia 69 (1977), 592. [12] WOOD, T. &I., MCCRAE, S. I., MACFARLANE, C. C.: Biochem. J. 1S9 (1980), 51. [13] XLNIN, K. E., ERIKSSOS, L E . , PETTERSON, B.: Eur. J. Biochem. 51 (1975), 207. [14] ERIKSSON, K.-E., HOLLMARK, B. H.: Biochem. Biophys. 133 (1969), 233. [la] ALMIN, K. E., ERIKSSON, K.-E., JANSSON, C.: Biochim. Biophys. Acta 139 (1967), 248. [16] CLARKE, A., STONE, €3.: Biochem. J. 96 (1965), 802. [17] MANDELS, M., REESE. E. T.: Enzym. hydrol. cellul.. Ed.: E. T. REESE, Pergamon Press,
[IS] PETTERSSON, G.: Arch. Biochem. Biophys. 130 (1969), 286. [19] KING, K. W., VESSAL, 31. J.: Adv. Chem. Ser. 95 (1969), 7. [20] SPURLIN, I(. M.: J. Am. Chem. SOC. 61 (1939), 2222. [21] I(ASULKE, U.: Dissertation A, Akademie der Wissenschaften der DDR, 1986.
1963, p. 115.





























