Embed Size (px)
Citation preview

Zvyšování konkurenceschopnosti studentů oboru botanika a učitelství biologie
CZ.1.07/2.2.00/15.0316

Využití houbových organismů v genovém inženýrství
MIKROORGANISMY - bakterie, kvasinky a houby využíval člověk po staletí. Lidé je VYUŽÍVAJÍ K PRODUKCI POTRAVIN, KRMIV A LÉČIV či ke kontrole životního prostředí. Na druhou stranu jsou některé mikroorganismy a viry patogenní. V minulém století PO OBJEVU DNA a možnosti genového transferu zahájili vědci VÝZKUM GENETICKY MODIFIKOVANÝCH ORGANISMŮ. Mikroorganismy se využívají jako DONORY ČI RECIPIENTY GENŮ - nosiče nějakých vlastností. Účelem této snahy je VYLEPŠENÍ CHARAKTERISTIK EXISTUJÍCÍCH ORGANISMŮ přidáním vhodných genů, nebo vystřižení a eliminace genů nesoucích nežádoucí vlastnosti. V České republice Ministerstvo životního prostředí je autoritou zodpovědnou za biologickou bezpečnost. V souladu se zákonem 78/2004 Sb. je povolen POUZE UZAVŘENÝ CHOV RŮZNÝCH GENETICKY MODIFIKOVANÝCH MIKROORGANISMŮ. Hlavním polem využití genetického inženýrství mikroorganismů jsou ZDRAVOTNÍ PÉČE, KULTIVACE ROSTLIN, PRODUKCE BIOCHEMIKÁLIÍ, FARMACEUTIK A POTRAVINÁŘSKÝCH ČI KRMIVÁŘSKÝCH PRODUKTŮ. Na poli ochrany životního prostředí je výzkum geneticky modifikovaných organismů zaměřen na BIOREMEDIACI.
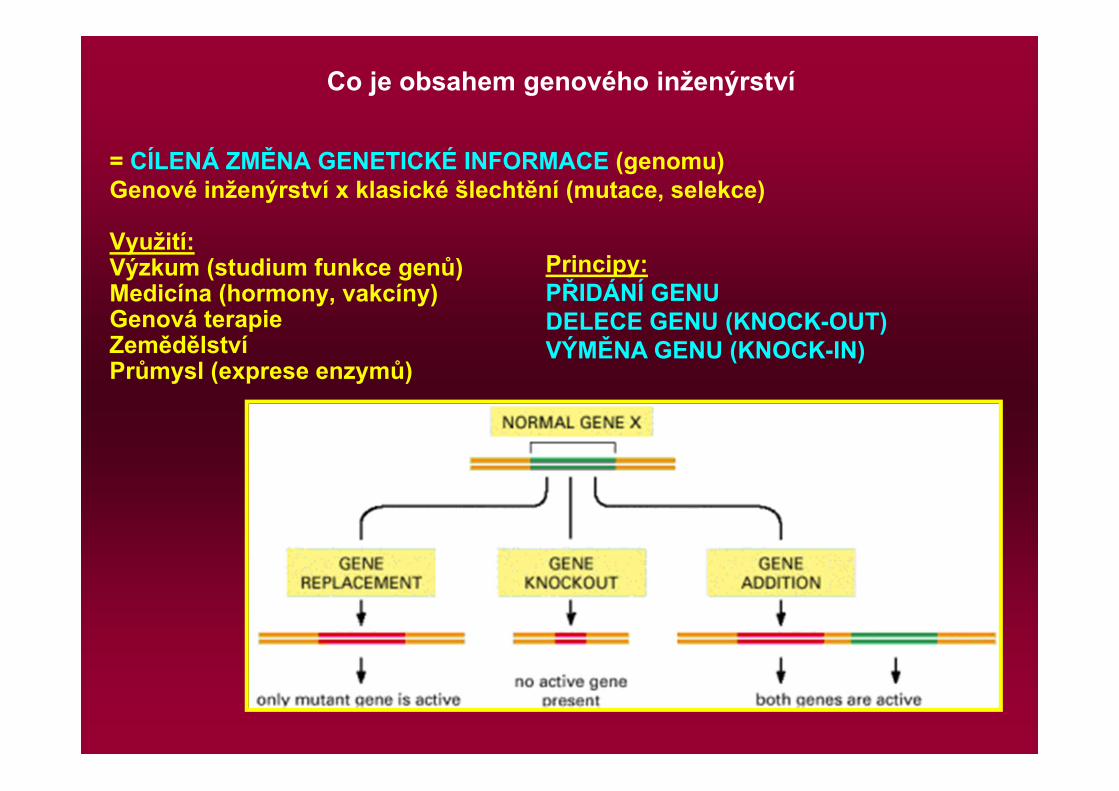
Co je obsahem genového inženýrství
= CÍLENÁ ZMĚNA GENETICKÉ INFORMACE (genomu)Genové inženýrství x klasické šlechtění (mutace, selekce)
Využití:Výzkum (studium funkce genů)Medicína (hormony, vakcíny)Genová terapieZemědělstvíPrůmysl (exprese enzymů)
Principy:PŘIDÁNÍ GENUDELECE GENU (KNOCK-OUT)VÝMĚNA GENU (KNOCK-IN)

Genetická manipulace u průmyslových hub
Z kvasinek je nejlépe prozkoumaní Saccharomyces cerevisiae, která má genetický systém více pozměnitelný k modifikaci.
Nejčastějšími objekty klasických genetických studií jsou druhy Neurospora crassa a Aspergillus nidulans, ale naneštěstí zase mají malé nebo žádné průmyslové využití.
Do roku 1974 byly nejpoužívanějšími technikami zlepšování kmenů hub MUTACE, SCREENING A SELEKCE. Tyto postupy byly velmi účinné na poli produkce penicilínu – nově vyšlechtěné kmeny Penicillium chrysogenum produkují 50 x více penicilínu než původní flemingovo P. notatum.
Aspergillus nidulans Neurospora crassa

MUTACE A MUTAGENEZE Mutace jsou DĚDIČNÉ ZMĚNY, ke kterým DOCHÁZÍ V GENOMU, může zahrnovat jednotlivé nukleotidy, sekvence nukleotidů, nebo celé oblasti na chromozómech. Frekvence přirozených mutací je nízká a tak v PRŮMYSLOVÉM VYUŽITÍ – MUTAGENNÍ ČINIDLA (X-paprsky, UV zaření, chemické mutageny (demethylsulphonate, ethylmethanesulphonat).
PARASEXUÁLNÍ SYSTÉM Mnoho průmyslově důležitých hub – Aspergillus, Penicillium POSTRÁDÁ POHLAVNÍ STADIUM, jeho náhradou zajišťující variabilitu je PARASEXUÁLNÍ PROCES
Může k němu dojít v laboratoři, když se KULTIVUJÍ SPOLEČNĚ různé KMENY téhož druhu.
V současné době tento proces probíhá ŘÍZENĚ POMOCÍ FŮZE PROTOPLASTŮ. Zahrnuje odstranění buněčné stěny enzymy, následuje ELEKTROFÚZE, kultivace PROTOPLASTOVÉHO AGREGÁTU, nutná selekce. Např. šlechtění Aspergillus niger pro vyšší produkci glukosamylázy. Dále pokusy s fůzí protoplastů u Penicillium, Claviceps, Giberella, Trichoderma. Beauveria.
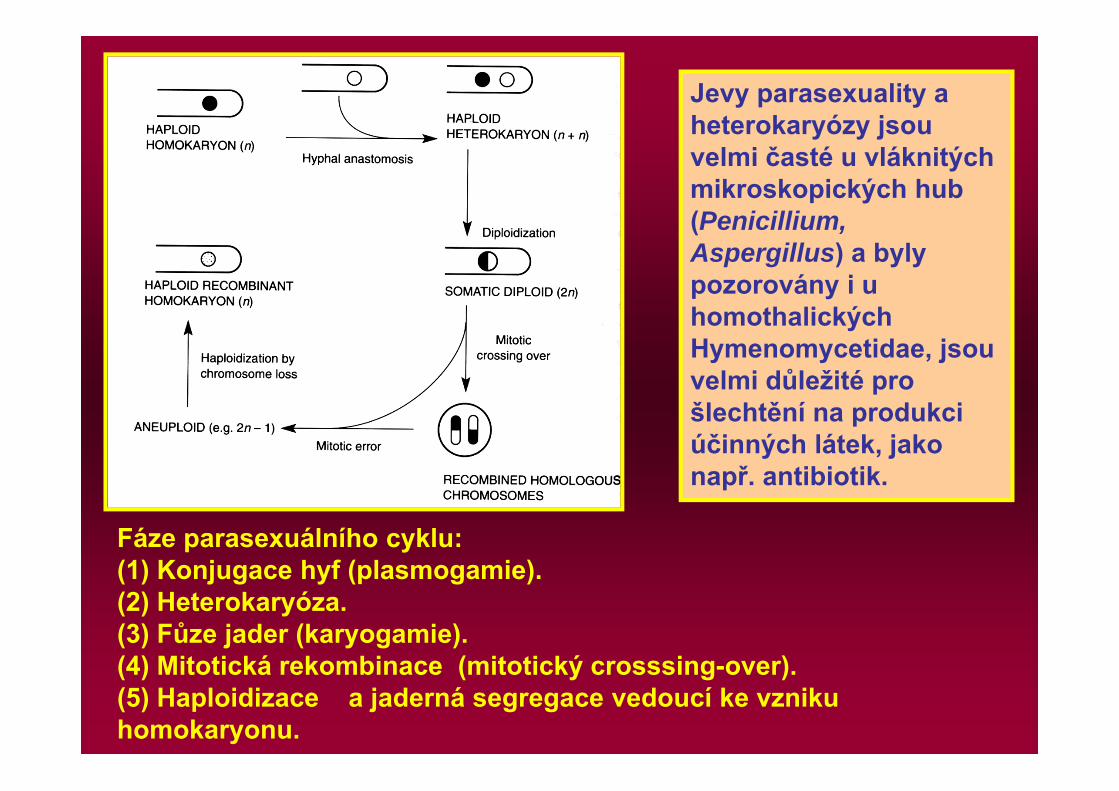
Fáze parasexuálního cyklu: (1) Konjugace hyf (plasmogamie). (2) Heterokaryóza. (3) Fůze jader (karyogamie). (4) Mitotická rekombinace (mitotický crosssing-over). (5) Haploidizace a jaderná segregace vedoucí ke vzniku homokaryonu.
Jevy parasexuality a heterokaryózy jsou velmi časté u vláknitých mikroskopických hub (Penicillium, Aspergillus) a byly pozorovány i u homothalických Hymenomycetidae, jsou velmi důležité pro šlechtění na produkci účinných látek, jako např. antibiotik.

Pohlavní rozmnožování ParasexualitaVysoce organizovaná a často taky přísně načasovaná událost, která je geneticky programovaná
Zahrnuje sekvenci ne zcela častých událostí, které se zdají být spíše příležitostné než v určitém plánu
Jaderná fůze je často zprostředkovaná genetickými faktory, vyjádřenými jako párovací typy ('mating types‚) dochází k ní ve vysoce specializovaných strukturách a často zahrnuje mnoho párů kompatibilních jader.
Jaderná fůze je izolovaná událost, není zprostředkovaná pomocí párovacích typů, nebyla nalezena ve specializovaných strukturách a zahrnuje pouze jednotlivá jádra.
Během meiózy, ke crossing-overu pravděpodobně dochází v každém homologním páru chromozómů a mnohonásobný crossing-over je běžný.
Během somatické rekombinace, ke crossing-overu běžně dochází pouze u jednoho nebo několika chromozómů, a nikdy to není tak často jako u meiózy.
V meiózi, k segregaci dochází vysoce organizovaným způsobem během 2 specializovaných jaderných dělení.
Somatická haploidizace se pravděpodobně vyskytuje jako výsledek postupné ztráty chromozómů z aneuploidního jádra (2n-1) během několika mitotických dělení než je ustanoven stabilní haploid
SROVNÁNÍ POHLAVNÍHO CYKLU A PARASEXUÁLNÍHO CYKLU

TRANSFORMACE
V 70-tých letech bylo objeveno, že kvasinka Saccharomyces cerevisiaeOBSAHUJE PLASMID. Saccharomyces cerevisiae má poměrně velké rozměry, snadnou kultivaci a není to nebezpečný organismus. Jsou schopny produkovat PROTEINY I Z ŽIVOČIŠNÝCH GENŮ. Plasmidy jsou do kruhu stočené molekuly DNA, které NOSÍ DĚDIČNOST NEZÁVISLOU NA CHROMOZÓMECH. U kvasinek plasmidy se nacházejí volně v cytoplasmě nebo nukleoplasmě. Jeden plasmid může být z BUŇKY DO BUŇKY předán procesem TRANSFORMACE Plasmid může být použit jako VEKTOR, kterým se PŘENÁŠEJÍ POTŘEBNÉ ČÁSTI buněčné DNA z jednoho kmene do druhého. Pro tento účel je obvyklé KLONOVAT plasmid do VHODNÉ BAKTERIE – obvykle Escherichia coli Při klonování, hybridní nebo chimerický plasmid je vytvořen použitím ČÁSTI PLASMIDU KVASINKY a části nebo celým BAKTERIÁLNÍM PLASMIDEM. Část bakteriálního nebo kvasinkového plazmidu musí obsahovat specifický gen –např. pro rezistenci k antibiotikům, nebo syntézu určité aminokyseliny. Plasmid může být kultivován a nebo KLONOVÁN V BAKTERII. Po kultivaci plazmidu v bakterii se požadovaný úsek DNA zpět INKORPORUJE DO BUŇKY KVASINKY V současnosti bylo TRANSFORMACÍ DOSAŽENO i u vláknitých hub Aspergillus nidulans a Neurospora crassa.

KVASINKOVÝ VEKTOR (A)
Vektor obsahuje BAKTERIÁLNÍ POČÁTEK REPLIKACE (ori) (umožňuje být rozmnožován jako plazmid E.coli) a GEN PRO REZISTENCI VŮČI ANTIBIOTIKUampicillin (Ampr). Vektor obsahuje KVASINKOVÝ POČÁTEK REPLIKACE a MARKEROVÝ GEN (LEU2), dovolující selekci transformovaných kvasinek. LEU2gen kóduje enzym nutný pro SYNTÉZU AMINOKYSELINY LEUCINU, aby mohly být transformované kvasinky, které nemají tento enzymy mohly být selektovány na médiu kde chybí leucin.
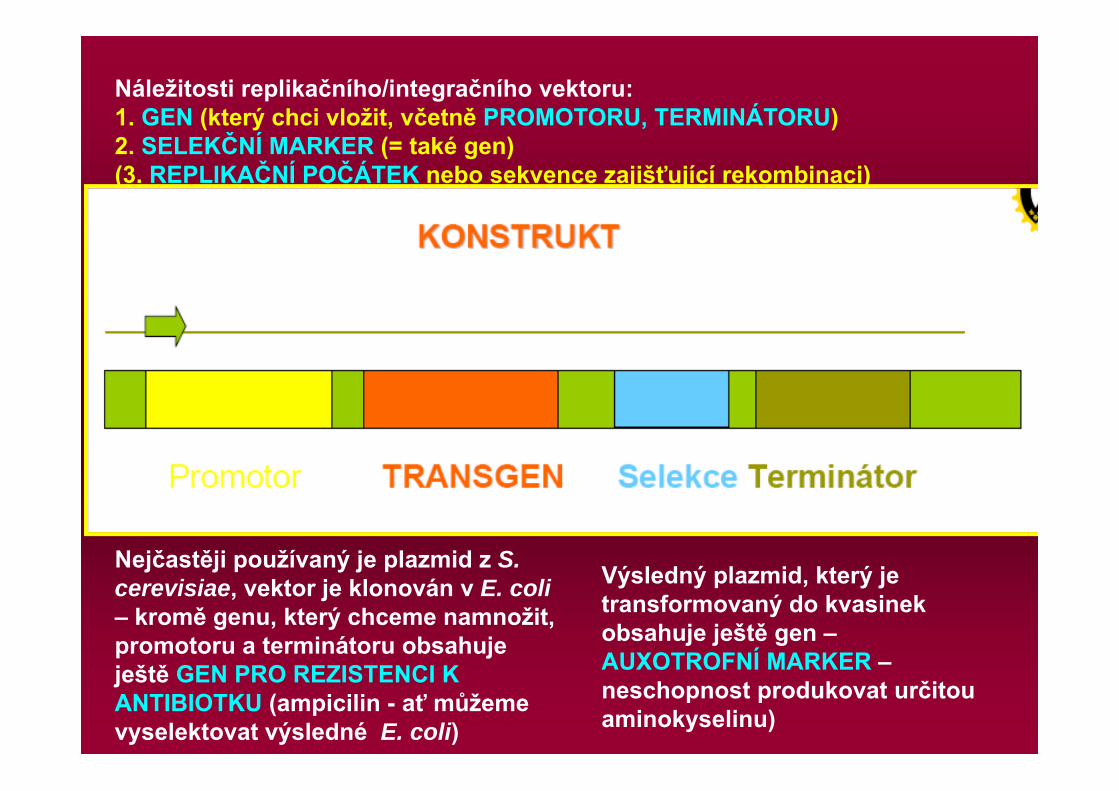
Náležitosti replikačního/integračního vektoru:1. GEN (který chci vložit, včetně PROMOTORU, TERMINÁTORU)2. SELEKČNÍ MARKER (= také gen)(3. REPLIKAČNÍ POČÁTEK nebo sekvence zajišťující rekombinaci)
Nejčastěji používaný je plazmid z S. cerevisiae, vektor je klonován v E. coli– kromě genu, který chceme namnožit, promotoru a terminátoru obsahuje ještě GEN PRO REZISTENCI K ANTIBIOTKU (ampicilin - ať můžeme vyselektovat výsledné E. coli)
Výsledný plazmid, který je transformovaný do kvasinek obsahuje ještě gen –AUXOTROFNÍ MARKER –neschopnost produkovat určitou aminokyselinu)
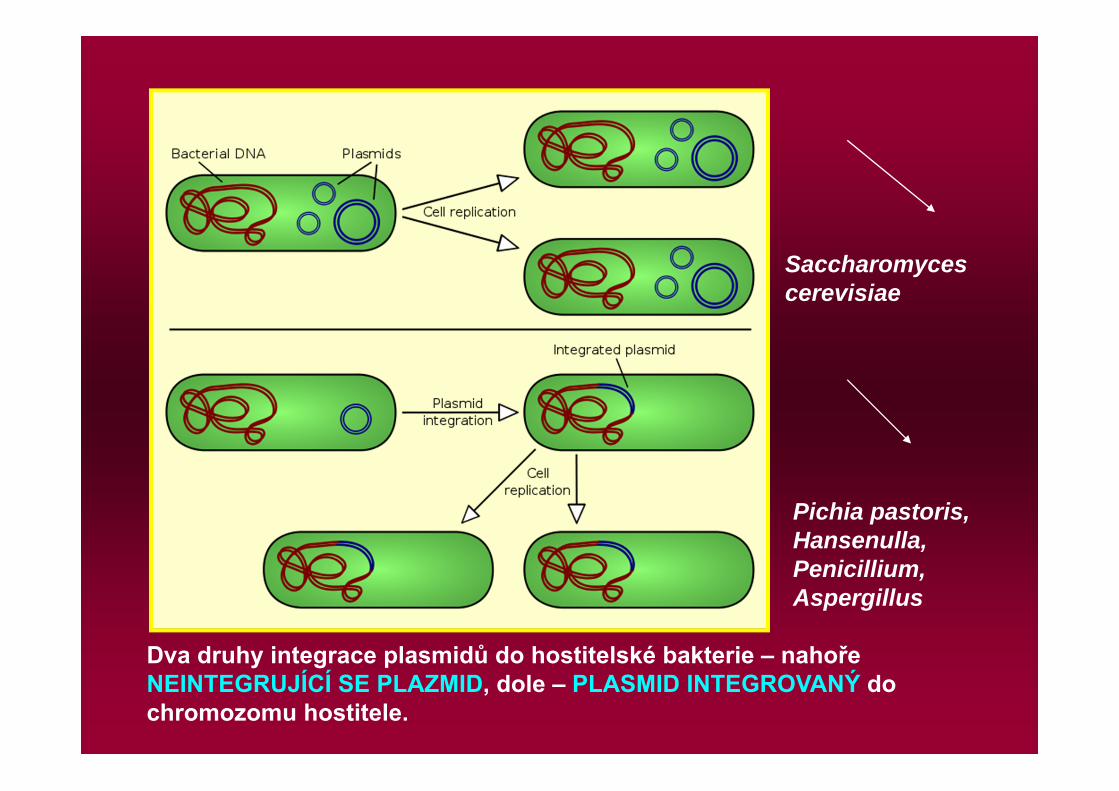
Dva druhy integrace plasmidů do hostitelské bakterie – nahoře NEINTEGRUJÍCÍ SE PLAZMID, dole – PLASMID INTEGROVANÝ do chromozomu hostitele.
Pichia pastoris, Hansenulla, Penicillium, Aspergillus
Saccharomyces cerevisiae
SACCHAROMYCES CEREVISIAE – MODELOVÝ OBJEKT
Velký objem poznatků BUNĚČNÉ BIOLOGIE, GENETIKY, možnost velké variability kultivačních podmínek, zdroj silných promotorů, přirozená PŘÍTOMNOST 2цm PLASMIDU a schopnost relativně široké škály POSTTRANSLAČNÍCH MODIFIKACÍ BÍLKOVIN, včetně obecně akceptované NERIZIKOVOSTI TOHOTO TAXONU určuje současný rozsah aplikací tohoto kmene v KOMERČNÍ PRODUKCI (PŘEDEVŠÍM) ŽIVOČIŠNÝCH PROTEINŮ (asi 20, bio-terapeutika, vakciny, diagnostika - antigeny; asi 50 % světové PRODUKCE INSULINU).
Použitelné vektory. Typ: epizomální (YEps), plasmidyYIps, umělé chromozomy (YACs).

PICHIA PASTORIS
Významná ALTERNATIVA S. cerevisiae (S. cerevisiae - nízká produkce některých typů bílkovin, zadržení bílkoviny v periplazmatickém prostoru, produkce cytotoxického etanolu a další).
Pichia pastoris – metylotrof (schopné UTILIZOVAT METHANOL jako zdroj uhlíku; jsou extrémně citlivé na jeho přítomnost nebo nedostatek v médiu). Nepřítomnost etanolu umožňuje realizovat kultivace dosahující vysokých buněčných denzit. Velká schopnost tohoto taxonu ZACHOVAT AUTENTICKÝ CHARAKTER HETEROLOGNÍHO PROTEINU (prověřeno asi 100 bílkovin). Systém genového vyjádření na bázi YAC. Produkce např . Hirudinu proti srážení krve

KVASINKOVÝ GENOM
Obecná charakteristika eukaryotické DNA
JÁDRO kvasinek je zřetelně ohraničeno JADERNOU MEMBRÁNOU a tvoří ho CHROMATIN, tedy komplex dsDNA, histonů a proteinů nehistonové povahy. Organely, především MITOCHONDRIE, jsou odděleny MEMBRÁNOU od ostatních kompartmentů buňky a je v nich přítomna alespoň jedna MOLEKULA KRUŽNICOVÉ dsDNA. Většina genetické informace kvasinkové buňky je uložena v chromozómech v jádře. Kromě nich může jádro obsahovat také KRUHOVOU MOLEKULU DNA tzv. 2 µm PLAZMID nebo 3 µm DNA.
Jaderný genom
Většina genů je soustředěna v JÁDŘE v CHROMOZÓMECH. Počet chromozómů byl odvozován z výsledků genetických studií. U nejlépe prostudované kvasinky, tj. Saccharomyces cerevisiae, bylo zjištěno 17 CHROMOZÓMŮ v haploidním jádře. Kvasinkový chromozóm obsahuje jednu dlouhou lineární molekulu dsDNA, jejíž velikost se pohybuje kolem 1.107 bp.

Mitochondriální genom
Mitochondriální dvoušroubovicová DNA se od chromozomální liší NIŽŠÍ DENZITOU A MENŠÍ MOLEKULOVOU HMOTNOSTÍ a je označována jako mtDNA. Molekuly mtDNA se výrazně liší svou velikostí (17 až 108 kb), většinou jsou kružnicové, ale u některých kvasinek jsou přítomny i lineární mtDNA.
Plazmidová DNA
Plazmidová DNA zpravidla NENÍ PRO BUŇKU ESENCIÁLNÍ, protože neobsahuje genetickou informaci kódující nezbytné životní funkce. Zatím jen u malého počtu eukaryotických buněk byla zjištěna její přítomnost. Ve většině laboratorních kmenů kvasinky Saccharomyces cerevisiae lze však v jádře nalézt 2 µM PLAZMID, který představuje kovalentně uzavřenou kružnicovou dsDNA nazvanou podle své obvodové délky. Buňka pak obsahuje plazmidovou DNA asi v 60 kopiích v haploidním a ve 120 kopiích v diploidním stádiu. Tento 2 µm plazmid se stal ZÁKLADEM PRO KONSTRUKCI MNOHA VEKTORŮ používaných v genových manipulacích u kvasinek

EXPRESE HETEROLOGNÍCH PROTEINŮ
HETEROLOGNÍ PROTEIN – rekombinantní protein – znamená produkce určitého GENOVÉHO PRODUKTU v organismu pro který to NENÍ NORMÁLNÍ.
Recentní aplikace technik s REKOMBINANTNÍ DNA NA KVASINKY A VLÁKNITÉ HOUBY otvírá nové možnosti ve vztahu ke konstrukci vysoce produktivních kmenů.
Heterologní genové expresní systémy se studují u mnoha houbových organismů - NEJINTENZIVNĚJŠÍ využití je KVASINEK Saccharomyces cerevisiae, VLÁKNITÉ HOUBY jsou nyní zkoumány jako POTENCIÁLNÍ HOSTITELSKÉ ORGANISMY k produkci heterologních proteinů. Neuropsora, Aspergillus, Penicillium. V mnoha případech pro základní výzkum, v dalších pro komerční aplikace.

EXPRESE HETEROLOGNÍCH PROTEINŮ
Příklady heterologních proteinů produkovaných houbovými organismy
Saccharomyces cerevisiae :
glukoamyláza, endoglukanáza, lidský a-interferon, Insulin, Interleukin, myší a-amyláza, myší imunoglobin
Pichia pastoris:
pšeničná a-amyláza, kvasinková invertáza, HIV antigeny, lidský serový albumin, hovězí lysozym, tumor nekrosis faktor,
Aspergillus nidulans:
lidský a-interferon, růstové faktory, růstové hormony
Trichoderma reesei:
produkce celulázy

Kvasinka (Saccharomyces cerevisiae)
Rok 1996 – KOMPLETNĚ ZMAPOVANÝ GENOM jako první eukaryotický mikroorganismus.
Tento organismus se stal klíčem pro POST-GENOMICKÝ VÝZKUM – u každého genu bylo zjišťováno jako se PROJEVUJE VE FENOTYPU, jaké jsou interakce jednotlivých proteinů.
Pro klonování se využívá umělých kvasinkových chromozómů YAC (Yeasts Artifical Chromosomes)
MAPOVÁNÍ GENOMU HOUBOVÝCH ORGANISMŮ
Další mikroorganismy, u kterých bylo dokončeno mapování genomu:
Schizosaccharomyces pombe, Aspergillus fumigatus, Candida albicans, Cryptococcus neoformans, Ustillago maydis, Magnaporthe grisea.





























