
Biochemistry 201Biological Regulatory MechanismsJanuary 26, 2015C. Gross, Lecture 3
Eukaryotic transcriptional regulation
References
A few of the many insights from RNA polymerase structures
Cramer, P. (2002) Multisubunit RNA polymerases. Curr Opin Struct Biol 12:89-97.
Murakami KS, Darst SA. (2003) Bacterial RNA polymerases: the holo story. Curr Opin Struct Biol 13:31-9.
*Cramer, P. (2004) RNA polymerase II structure: from core to functional complexes. Curr Opin Genet Dev 14:218-26. Review.
Wang, D. Bushnell DA, Westover KD, Kaplan, CD, Kornberg RD. Structural basis of transcription: role of the trigger loop in substrate specificity and catalysis. Cell. 2006 Dec 1;127(5):941-54.
Kostrewa D, Zeller ME, Armache KJ, Seizl M, Leike K, Thomm M, Cramer P.(2009) RNA polymerase II-TFIIB structure and mechanism of transcription initiation. Nature. 462:323-30.
Chromosome conformation capture (CCC)de Wit, E. and de Laat, W. (2012) A decade of 3C technologies: insights into nuclear organization. Genes Dev. 26: 11-24.
Mediator and Other Components*Kornberg, R.D. (2005) Mediator and the mechanism of transcriptional activation. Trends in Biochemical Sciences 30:235-239.Fan, X, Chou, DM, & Struhl, K. (2006). Activator-specific recruitment of Mediator in vivo. Nature Structural & Molecular Biology, 13(2), 117-20.Sikorski TW and Buratowski. (2009). The basal initiation machinery: Beyond the general transcription factors. Current Opinion in Cell Biology. 21 344-351.

What do activators do?Cosma, MP, Tanaka, T, & Nasmyth, K. (1999). Ordered recruitment of transcription and chromatin remodeling factors to a cell cycle- and developmentally regulated promoter. Cell, 97(3), 299-311.Bryant, GO, & Ptashne, M. (2003). Independent recruitment in vivo by Gal4 of two complexes required for transcription. Molecular Cell, 11(5), 1301-9.Bhaumik, S.R., Raha, T. Aiello, D.P., and Green, M.R. (2004) In vivo target of a transcriptional activator revealed by fluorescence resonance energy transfer. Genes Dev 18: 333-343.Vakoc, CR, Letting, DL, Gheldof, ... Blobel, GA (2005) Proximity among Distant Regulatory Elements at the B–Globin Locus Requires GATA-1 and FOG-1. Molecular Cell 17:453-462Fishburn, J., Mohibullah, N. and Hahn, S. (2005) Function of a eukaryotic transcription activator during the transcription cycle. Molecular Cell 18:369-378.Bulger M and Groudine M. Functional and Mechanistic Diversity of Distal Transcription Enhancers (2011). Cell 144:327-39
Role of the RNA Pol II CTD
*McCracken, S, Fong, N, Yankulov, K, et al. (1997). The C-terminal domain of RNA polymerase II couples mRNA processing to transcription. Nature, 385(6614), 357-61.
Tietjen,J. ……Ansari, A. Chemical-genomic dissection of the CTD code (2010) NMSB: 17: 1154-1162
Mayer, A. ….Cramer, P. Uniform transitions of the general Pol II transcription apparatus (2010) NMSB 17:1272-79
Buratowski, S (2009) progression through the RNA polymerase II CTD cycle ( Review). Mol Cell 36: 541-546
Chapman, R… Eick, D. Molecular evolution of the RNA polymerase CTD. Trends in Genetics (2008): Jun;24(6):289-96. Epub 2008 May 9. Review.PMID: 18472177
Genome wide elongation technologiesChurchman LS, Weissman JS. (2011) Nascent transcript sequencing visualizes transcription at nucleotide resolution. Nature 469: 368-73. doi: 10.1038/nature09652. (Net-seq)
Kwak, H… and Lis, JT (2013) Precise maps of RNAP reveal how promoters direct initiation and pausing. Science 339: 950 (PRO-seq)

Elongation Control
BBA2013-- Issue 1874 devoted to reviews of transcription elongationYamaguchi, Shibata and Handa (2013):Transcriptionelongation factors DSIF and NELF: Promoter proximal Pausing and Beyond. BBA 1829: 98-104Note: this issue of BBA has many elongation reviews; most not too good.
Kwak and Lis (2013) Control of transcriptional Elongation Annual review Genetics 47: 483-508
Jonkers, Kwak and Lis (2014) Genome-wide dynamics of PolII elongation and its interplay with promoter proximal pausing, chromatin and exons. E-life: 2014;3:e02407. DOI: 10.7554/eLife.02407
Veloso, Kirkconnell, magnuson, Biewen, Paulsen, Wilson and Mats Ljungman. (2014) Rate of elongation by RNA polymerase II is associated with specific gene features and epigenetic modifications. Genome research 24:896–905
Core, Waterfall and Lis (2008) Nascent RNA Sequencing Reveals widespread pausing and divergent initiation at Human promoters. Science 322: 1845-1848
Zhou Q, Li T, Price DH (2012) RNA polymerase II elongation control .Annu Rev Biochem. 2012;81:119-43. Rougvie A and Lis JT (1988) The RNA Polymerase II Molecule at the 5’ end of the uninduced hsp70 gene of D. melangaster is transcriptionally engaged. Cell 54: 795-804
Zobeck, KL….Lis Jt (2010) Recruitment timing and dynamics of transcription factors at the Hsp70 Loci in Living Cells Mol Cell 40 965-75
Nechaev S…..Adelman K. (2010) Global Analysis of short RNAs reveals widespread Promoter Proximal Stalling and Arrest of Pol II in Drosophila Science 327: 335-38
Gilchrist, DA,……Adelman, K. (2010) Pausing of RNA Polymerase II Disrupts DNA specified Nucleosome Organization to enable precise gene regulation. Cell 143: 540-51Chen, Y.,….Handa, H.(2009) DSIF, the Paf1 complex and Tat-SF1 have nonredundant, cooperative roles in RNA polymerase II elongation. Genees Dev 23: 2765 -77
Liu, Y……Hahn, S. (2009) Phosphorylation of the transcription elongation factor Spt5 by yeast Bur1 kinase stimulates recruitment of the PAF complex. MCB 29: 4852-63Wu, C-H,….Gilmour, D. ( 2003)NELF and DSIF cause promoter proximal pausing on the Hsp70 promoter in drosophila. Genes Dev 17: 1402-14

RNA polymerase elongation rate and splicingBraberg, Jin, Moehle, Chan, Wang, Shales……Guthrie, Kaplan Krogan ( 2013) From Structure to Systems: High resolution Quantitative genetic Analysis of RNA polymerase II. Cell 154, 775-788
Fong, Kim, Zhou, Jim Qiu, Saldi Diesner, Jones, Fu and Bentley (2014) Ore0mRNA splicing is facilitated by an optimal RNA polymerase II elongation rate Genes and Dev 28: 2663-2676

Important Points
1. DNA looping allows proteins bound to distant sites on DNA to interact.
2. Transcription initiation at Pol II promoters on naked DNA templates in vitro requires the general transcription factors in addition to RNA polymerase II.
3.. In vivo, transcription initiation also requires activators – proteins that bind directly to enhancers – as well as Mediator and enzymes that modify chromatin structure.
4. At a typical eukaryotic promoter, activators guide the assembly of Mediator, the general transcription factors, RNA polymerase and chromatin-modifying enzymes, often through weak, relatively non-specific interactions. There appears to be no set order of assembly from one promoter to the next. Moreover, different promoters have different requirements for these components.
5. The RNA polymerase CTD is a long series of 7-amino acid repeats. When transcription is initiated, serine 5 of the repeat is phosphorylated by TFIIH. As elongation proceeds, serine 5 is gradually dephosphorylated and serine 2 is gradually phosphorylated by enzymes carried along with the RNA polymerase. This dynamic pattern of modification couples transcription to processing of the newly-synthesized RNA.
6. Promoter proximal pauses ( 20-50 nucleotides) are extensively used for regulation in eukaryotes.
7. Transcription takes place in a chromatin world and they exhibit a 2-way interaction

New challenges for transcription in eukaryotic cells
1. Three polymerases
2. Much more complex pattern of gene expression
3. Transcription takes place in a chromatin world
4. Complex processing of mRNA
Proliferation of trx factors Regulation at a distance ( enhancers)Combinatorial control
Constant 2-way interplay between trx and chromatin
PolII CTD serves as a platform for coordinating processes

PolII initiation factors and core promoter

Purification scheme for partially purified general transcription factors. Fractionation of HeLa nuclear extract (Panel A) and nuclear pellet (Panel B) by column chromatography and the molar concentrations of KCl used for elutions are indicated in the flow chart, except for the Phenyl Superose column where the molar concentrations of ammonium sulfate are shown. A thick horizontal (Panel A) or vertical (Panel B) line indicates that step elutions are used for protein fractionation, whereas a slant line represents a linear gradient used for fractionation. The purification scheme for pol II, starting from sonication of the nuclear pellet, followed by ammonium sulfate (AS) precipitation is shown in Panel B. (Figures are adapted from Flores et al., 1992 and from Ge et al., 1996)
NAME # OF SUBUNITS FUNCTION
TFIIA 3 Antirepressor; stabilizes TBP-TATA complex; coactivator
TFIIB 1 Recognizes BRE;Start site selection; stabilize TBP-TATA; pol II/TFIIF recruitment
TFIID TBP 1 Binds TATA box; higher eukaryotes have multiple TBPs TAFs ~10 Recognizes additional DNA sequences; Regulates TBP binding; Coactivator;
Ubiquitin-activating/conjugating activity; Histone acetyltransferase; multiple TAFs
TFIIF 2 Binds pol II; facilitates pol II promoter recruitment and escape; Recruits TFIIE and TFIIH; enhances efficiency of pol II elongation
TFIIE 2 Recruits TFIIH; Facilitates forming initiation-competent pol II; promoter clearance TFIIH 9 ATPase/kinase activity. Helicase: unwinds DNA at transcription startsite; kinase
phosphorylates ser5 of RNA polymerase CTD; helps release RNAP from promoter

The Pol II promoter has many recognition regions
Positions of various DNA elements relative to the transcription start site (indicated by the arrow above the DNA). These elements are:
BRE (TFIIB recognition element); there is also a second BRE site downstream of TATA
TATA (TATA Box);
Inr (initiator element);
DPE (downstream promoter element);
DCE (downstream core element).
MTE (motif ten element; not shown) is located just upstream of the DPE.

The first two steps of Eukaryotic transcription
Many archae have a proliferation of TBPs and TFBs, suggesting that they provide choice in promoters, akin to alternative s.
On a supercoiled naked DNA template and in archae, TBP and TFB are sufficient for formation of the pre-initiation complex (PIC), suggesting that they are key to the mechanism of transcription initiation in eukaryotes
Promoter
TFBTBP

Transcription Initiation by RNA Pol II
The stepwise assembly of the Pol II preinitiation complex is shown here. Once assembled at the promoter, Pol II leaves the preinitiation complex upon addition of the nucleotide precursors required for RNA synthesis and after phosphorylation of serine resides within the enzyme’s “tail”.
PIC = preinitiation complex

The GTF’s are not enough—what else is needed?
The concept of a co-activator

The GTFs are not sufficient to mediate activation: Discovery and isolation of Mediator from Yeast
GTFs and RNA Pol II
Tx
1 unit
1 unit
10 units
crude lysate
4 years
50 units
mediator
VP 16
GAL4

Is Mediator Required for Transcription of all Pol II -transcribed genes?
mRNAs decrease over time according to their half-lives.A few mRNAs remain at some level (stable mRNAs).
Control: ts subunit of Pol II
Compare levels of all mRNAs using microarrays
Rpb1
Rpb1ts (37˚C for 45 min)
high T
Subunit of Mediator
Same basic pattern
Same basic pattern
Subunit of TF II H
Srb4ts
Kin28ts

The Head region interacts with PolII-TFIIF complex; the mutants with general effects on Trx are located in this region; the tail region interacts with activators; mutants have more specific effects on transcription
Mediator is very large and has diverse roles
A) PIC model from EM-study of polII (brown)-TFIIF (light blue) and X-ray structure of TBP (white) -IIB (yellow)--DNA; Arrow indicates direction of trx
B) Model for PIC-mediator was produced by superimposing an EM structure of Mediator-PolII on the PIC in A; head, middle and tail regions shown

SAGA is another important complex with multiple roles in transcription, including being a coactivator
The core of SAGA, containing the Taf substructure (Yellow), is surrounded by three domains responsible for distinct functions: activator binding (Tra-1), histone acetylation Gcn5), and TBP regulation (Spt3). This structural organization illustrates an underlying principle of modularity that may be extended to our understanding of other multifunctional transcription complexes.

The TAFs in TFIID also serve as coactivators

Assembly of PIC in presence of mediator, activators and chromatin remodelers

The factors and assembly pathways used to form transcriptionally competent preinitiation complexes can be promoter dependent. (1) TBP assembling onto promoter regions via TFIID leads to recruitment of the other basal initiation factors, as outlined in the stepwise assembly pathway. In S. cerevisiae, this pathway is most often utilized at TATA-less genes. At some mammalian promoters, histone H3K4 trimethylation helps to recruit the TFIID complex. (2) Mediator bridges interactions between activators and the basal initiation machinery, and can stimulate basal transcription as well. At some promoters Mediator can recruit TFIIH and TFIIE independently of RNApoII. (3) TBP can also be brought to promoters by the SAGA complex. In S. cerevisiae, this pathway is most utilized at TATA containing promoters. The Mot1 and NC2 complexes can repress this pathway by actively removing TBP from the TATA element. (4) Mot1 and NC2 can also have a positive role in transcription by removing non-productive TBP complexes from DNA, thereby allowing functional PICs to form.
Many Paths to the PIC
Buratowski, 2009

Genomic Level Snapshots of transcription factor binding sites
ChIP
ChIP-ChIP
ChIP-seq
ChIP-exo
Bioinformatic approaches
Phylogenetic footprinting
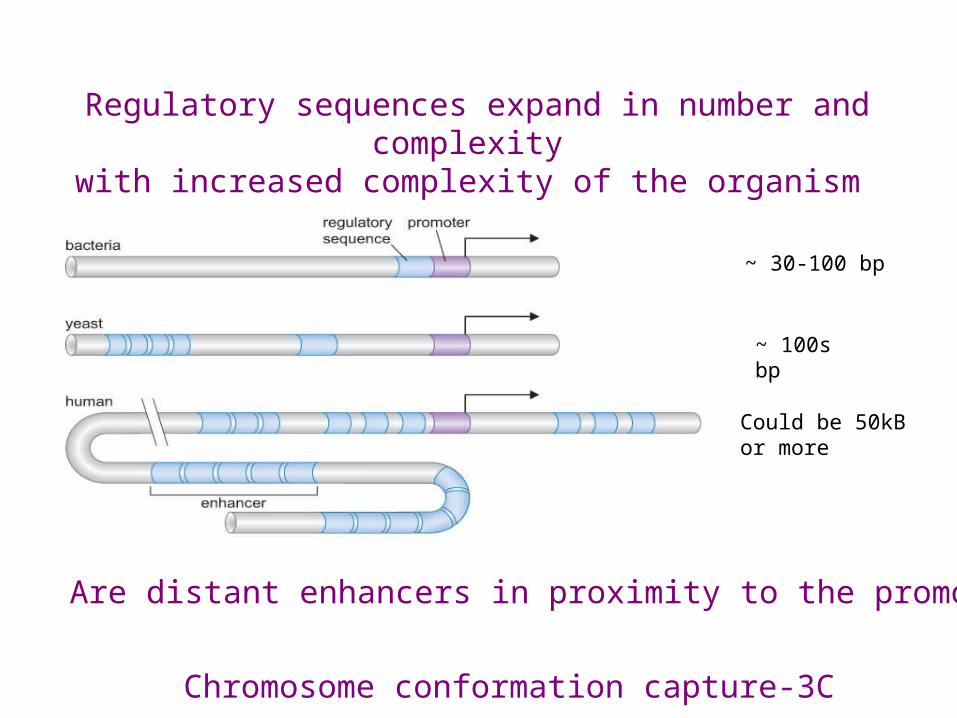
Regulatory sequences expand in number and complexity with increased complexity of the organism
~ 30-100 bp
~ 100s bp
Could be 50kB or more
Are distant enhancers in proximity to the promoter?
Chromosome conformation capture-3C

5C3C
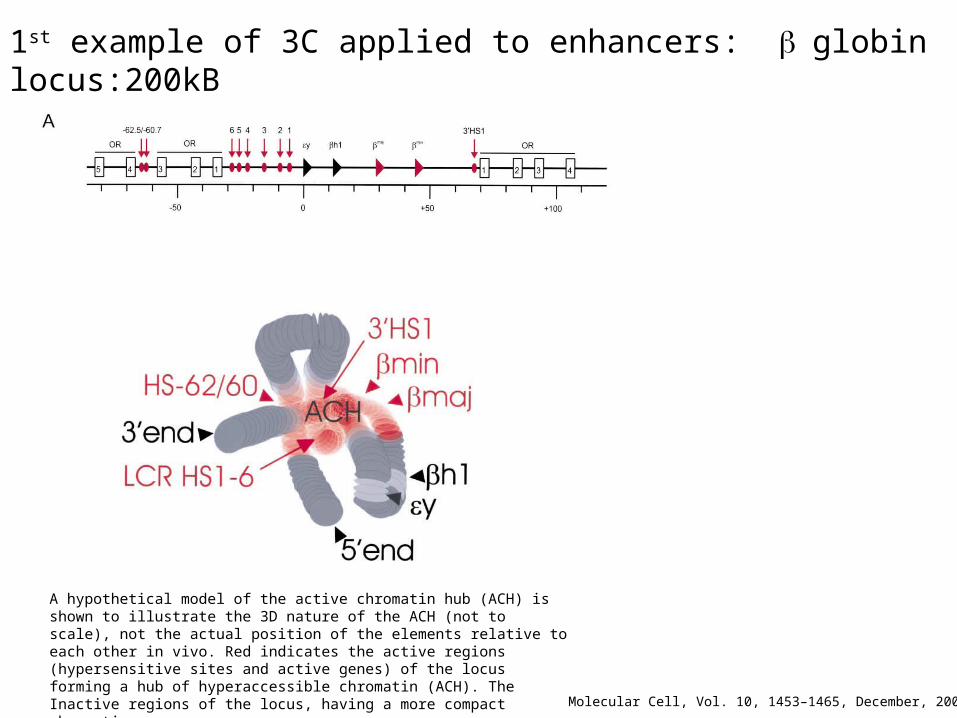
A hypothetical model of the active chromatin hub (ACH) is shown to illustrate the 3D nature of the ACH (not to scale), not the actual position of the elements relative to each other in vivo. Red indicates the active regions (hypersensitive sites and active genes) of the locus forming a hub of hyperaccessible chromatin (ACH). The Inactive regions of the locus, having a more compact chromatin structure are indicated in gray, with the inactive h1 and y genes in lighter gray.
1st example of 3C applied to enhancers: globin locus:200kB
Molecular Cell, Vol. 10, 1453–1465, December, 2002

Elongation control in eukaryotes

RNA polymerase II CTD
YSPTSPSP P P
Plasmodium: 5 Yeast: 26Mammals: 52
Heptad repeat unit
2 5 7
Palangat M et al. PNAS 2005;102:15036-15041©2005 by National Academy of Sciences
CTD is located adjacent to RNA exit channel

Phosphorylation state of PolI CTD during transcription
YSPTSPS2 5 7
TF II H, Mediator
pTEFb/Cdk9In S. cerevisiae:Cdk1 and Bur 1
Phosphatases (Rtr1(2?)
Phosphatases (Fcp1, ssu72)
Stage of transcription Kinase/phosphatase
Initiation YSPTSPS(Unphosphorylated)
YSPTSPS
P
Transition to elongation(Ser5)
YSPTSPS
P P
Elongation(Ser 2,5)
YSPTSPS
P
Further elongation(Ser2)
YSPTSPSTermination(Unphosphorylated)

A new step in transcription in higher eukaryotes
Initiation transition Elongation/termination
KB KfAbortive Initiation
ElongatingComplex RPoRPcR+P
NTPsProkaryotes
Higher Eukaryotes: same through abortive initiation, but new step before elongation
PIC ( does abortive initiation)
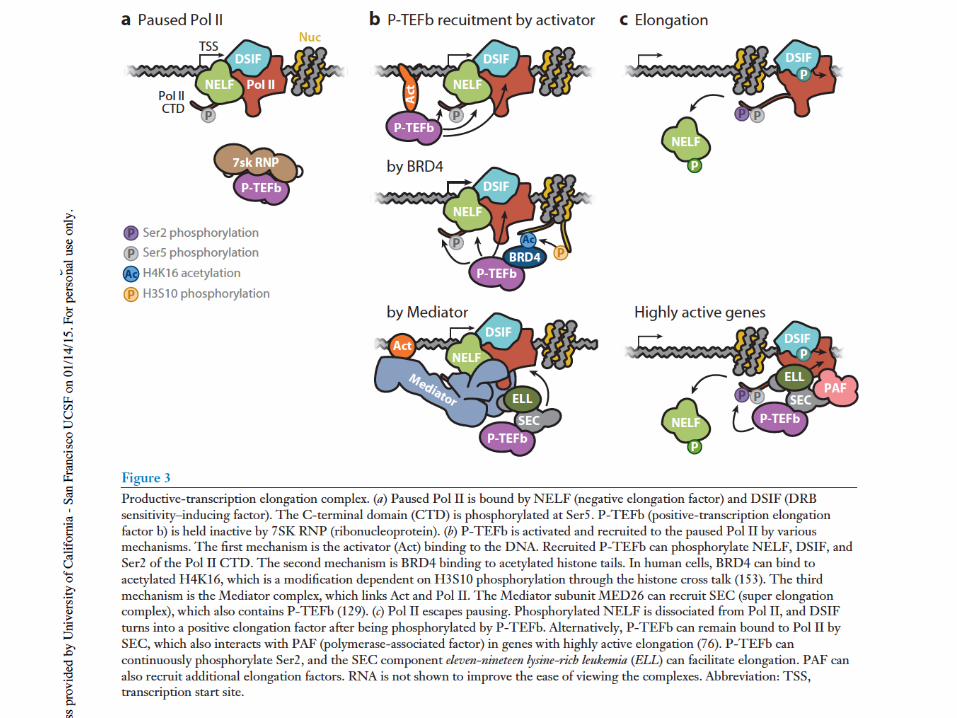
NELF: Negative elongation factor; DSIF: DRB (5,6-dichloro-1-beta-D-ribofuranosyl-benzimidazole) sensitivity-inducing factor-Spt4+5
Promoter-proximal pause: +20 - +50
Potential roles for paused polymerase: antagonize nucleosome formation to keep promoter open; synchronous activation; checkpoint

5 - amaR CTD
Mouse RNA Pol II
wt
52
What is the major role of the Pol II CTD?
examine RNAs
50 hrs.
HeLacells Introduce
CTD construct - amanitin
Splicing, processing of 3’ end, termination were all affected
Nature 385: 357 (1997)

Specific Processes are connected to each Phosphorylated Form of the CTD
CTD Status Transcription RNA-Processing Chromatin
Unphosphorylated Activation (mediator)
Serine 5P early termination mRNA capping H3K4 modification (ScN4E1 complex) (capping enzyme) Set1 complex
progression to elongation Nucleosome mobility (Cdk9 kinase via capping Cdk9/bur1 for Spt5
enzyme); Bur1kinase)
Serine 2P/5P H3K36 methylation (Set 2 )
Serine 2P late termination polyadenylation histone chaperone (Rtt103) (Pcf11) Spt6

Histone marks: transcriptional regulators or by-products?
Approach: track residue specific modifications in living cells in a strain with aGFP-glucocorticoid receptor and an inducible tandem array
Results:1.Preacetylated prior to addition of glucocorticoid2.High acetylation correlated with rapid gene activation and rapid transition to elongation
Kimura doi:10.1038/nature13714

TECs are community organizers
The major steps in mRNA processing (trx, 5’ capping, polyA addition, splicing) all occur together on a transcript extruded from the exit channel of RNAP although they can be reconstituted independently in vivo Principles of “cotranscriptionality” to integrate nuclear metabolism1.Permits coupling between different biogenesis steps; eg crosstalk; suspected when decreasing one step has effects on 2 nd; could always be indirect
a. Landing pad—increase concentration of reactants—proteins involved in capping etcb. Allosteary: guanosyl transferase of capping enzyme activated by interaction with phosphorylated CTDc. Kinetic coupling—optimize timing
2. Impose order or control
a. Juxtaposition of proteins permits assembly, competitive interactions handoffs; often mutually exclusive PPisb. directions emanating from phopshorylation state of CTD
3. A locator for nuclear machines –DNA repair, modification etc
Bentley: Cotranscriptionality Mol cell rev 2009

Kinetic coupling model for splicing
Approach: Measure splicing efficiency in a series of slow and fast RNAP; Importantly double slow + fast returned splicing to normal
Caveats: not true for all splice sites; some reverse, some unaffected
Recommended








![[VI]. Post-Transcriptional Processing and Post- Transcriptional Control of Gene Expression Processing of eukaryotic pre-mRNA About 60% of human genes](https://img.pdfslide.net/doc/110x75/56649ddb5503460f94ad2a9e/vi-post-transcriptional-processing-and-post-transcriptional-control-of.jpg)








