
LE PARADOXE DE LA GESTATION CHEZ
LES MAMMIFÈRES EUTHÉRIENS
Conflit parental, immunité et grossesse (Lyon, 9/3/2018)

PLAN DU COURS
Les gènes soumis à empreinte – pourquoi? Démonstration de leur présence chez les mammifères (Solter et
Surani)
Apparition dans l’évolution des vertébrés
Les gènes soumis à empreinte dans la fonction placentaire
Mécanismes de l’empreinte parentale: un cas particulier de régulation épigénétique Définitions
Mécanismes
Liens avec l’évolution biologique
Le paradoxe immunologique de la gestation Bases théoriques
Fausses couches à répétition et prééclampsie

UNE ILLUSTRATION DE LA NON-ÉQUIVALENCE DES
GÉNOMES PATERNELS ET MATERNELS
3

LES EXPÉRIENCES DE SURANI: NON
ÉQUIVALENCE DES GÉNOMES
4

IDEM CHEZ L’HUMAIN
(Wake et al., 1978)
5

APPARITION DE L’EMPREINTE CHEZ LES
MAMMIFÈRES PLACENTAIRES
6 (Renfree et al, Philos Trans R Soc Lond B Biol Sci. 2013)

PAS DE GÈNES SOUMIS À EMPREINTE CHEZ LES
OVIPARES
(Proudhon et Bourc’his, 2010)

(Proudhon et Bourc’his, 2010)
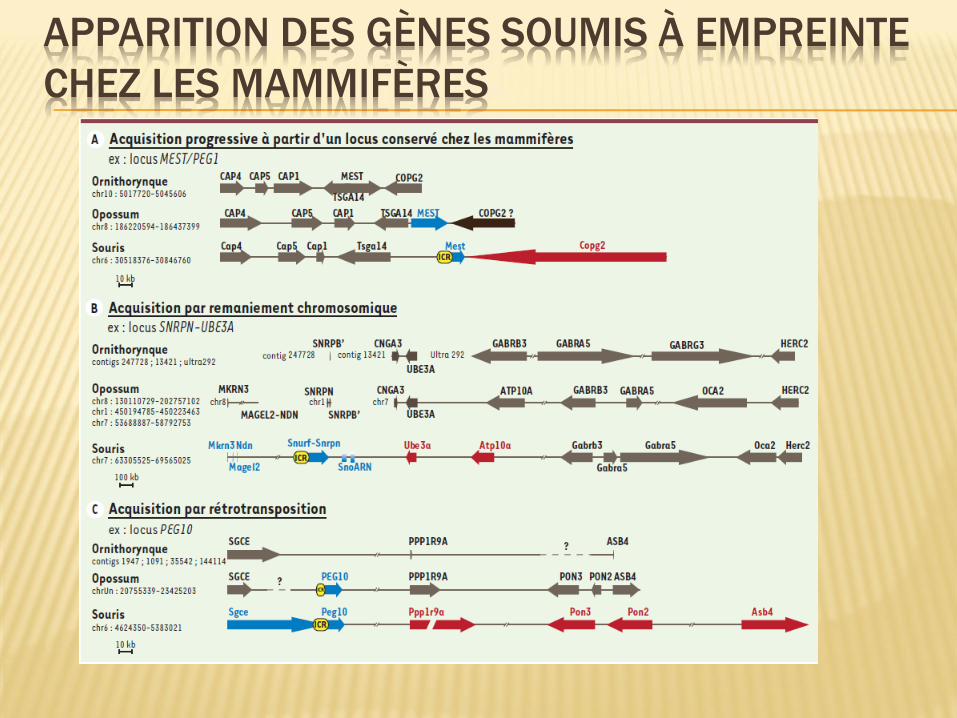
APPARITION DES GÈNES SOUMIS À EMPREINTE
CHEZ LES MAMMIFÈRES

LES GENES SOUMIS A EMPREINTE DANS
LE PLACENTA

LE PARADIGME H19/IGF2

LE RÔLE DES GÈNES SOUMIS À EMPREINTE DANS
LE CONFLIT MATERNO-PARENTAL
Constancia, Nature 2004 12

Constancia, Nature 2004
LE RÔLE DES GÈNES SOUMIS À EMPREINTE
DANS LE CONFLIT MATERNO-PARENTAL
13

MÉCANISMES DE L’EMPREINTE: LA
MACHINERIE ÉPIGÉNÉTIQUE

UNE DÉFINITION DE JBS HALDANE
« Dear Professor, I
cannot believe that in
only a few million years,
we have passed from
one-cell organisms to
complex beings as we
are. »
A lady (name not
remembered by history)
« Dear Madam, you did it
yourself in nine months! »

UN PEU DE TEXTE…
Epigenetics : heritable changes in gene expression or cellular phenotype caused by mechanisms other than changes in the underlying DNA sequence
These changes may remain through cell divisions for the remainder of the cell's life and may also last for multiple generations. However, there is no change in the underlying DNA sequence of the organism; instead, non-genetic factors cause the organism's genes to behave (or "express themselves") differently.
One example of epigenetic changes is the process of cellular differentiation. Totipotent stem cells become the various pluripotent cell lines which in turn become fully differentiated cells.
Trans-mitotic or Trans-meiotic?
The case of imprinted genes

DU CODE AU MESSAGE
Information Message Action
DNA ARNm Protéine
ARN de structure

LES TYPES CELLULAIRES ET L’ÉPIGÉNÉTIQUE
Hépatocyte Neurone

WADDINGTON (1905-1975) SUGGÈRE QUE LES
PHÉNOTYPES S’ÉTABLISSENT AU COURS DU
DÉVELOPPEMENT PAR UN POSITIONNEMENT DES
CELLULES DANS UN PAYSAGE ÉPIGÉNÉTIQUE.
S Yamanaka Nature 460, 49-52 (2009)

LE CONTEXTE DE L’ÉPIGÉNÉTIQUE
Gènes
Codant pour des protéines (20-30000)
Acteurs de l’épigénétique Méthylation de l’ADN
Modifications des histones
ARNs non-codant
•Phosphorylations
•Acétylations
•Méthylations
•Sumoylations
•Glycosylations
•Ubiquitinations
Modifications post-
traductionnelles
Transcription
ARNs non-codant (~30000?)
Au cours du développement, plus de 200
types cellulaires apparaissent

DES MARQUES ÉPIGÉNÉTIQUES QUI
TRAVERSENT PLUSIEURS GÉNÉRATIONS

EFFETS DE CONTAMINANTS
ALIMENTAIRES SUR LA GAMÉTOGENÈSE
Anway, 2005, Science

HISTOLOGIE TESTICULAIRE

EFFETS TRANSGÉNÉRATIONNELS
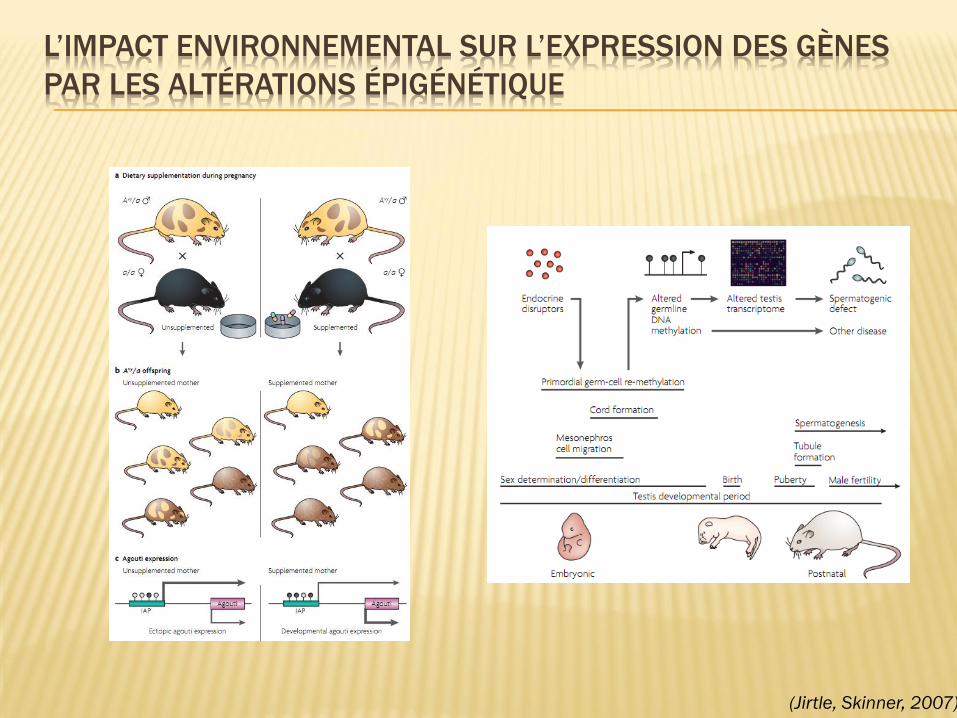
L’IMPACT ENVIRONNEMENTAL SUR L’EXPRESSION DES GÈNES
PAR LES ALTÉRATIONS ÉPIGÉNÉTIQUE
(Jirtle, Skinner, 2007)

L’ÉPIGÉNÉTIQUE EST ELLE UN MODE
D’ÉVOLUTION LAMARCKIEN?

LE « VRAI » APPORT DE L’ÉPIGÉNÉTIQUE À
L’ÉVOLUTION DU VIVANT
Adaptation à la température 25-35 °C Adaptation à la température 25-35 °C
Cyprinodon salinus salinus
A1 A2 A3 A4 A5 AA
Adaptation à la température 25-35 °C Adaptation à la température 25-35 °C
A2 A3 AA
Adaptation à la température 25-35 °C Adaptation à la température 25-35 °C
A3 AA
Adaptation à la température 25-35 °C
AA
Mort de l’espèce
An
né
e f
roid
e
An
né
e t
ièd
e
An
né
e c
ha
ud
e
An
né
e f
roid
e

DES ESPÈCES ÉPIGÉNÉTIQUES?

DEUX PHÉNOTYPES: UNE SÉQUENCE
GÉNÉTIQUE DANS LE GÈNE LCYC

LES MACHINERIES MOLÉCULAIRES À LA
BASE DE L’ÉPIGÉNÉTIQUE

DNA methylation, Histone
methylation/acetylation and other post-
translational modifications, miRNA
expression control gene expression

LES ACTEURS MOLÉCULAIRES DE
L’ÉPIGÉNÉTIQUE DNA methylation
DNA methyltransferases
Maintenance: DNMT1
De novo methylation: DNMT3A,
DNMT3B
Regulatory: DNMT3L
Acting on RNA targets: DNMT2
Demethylases:
TET1, TET2, TET3
Gadd45
Targets: CpG islands
Stability
Correlation with gene expression ?
Histone code
>70 post-translational
modifications on N-ter
amino-acids of H3 and
H4
A complex code
H3K4Me3 open
H3K27Me3 closed
H3/H4 Acet open
Numerous enzymes
involved
HAT
HDAC
Swi-Snf
P300/CEBP
LSD1
Documented
modifications by cellular
processes/stresses
MiRNA synthesis and
regulation
Targeting hundred of
genes
Powerful identification
tools now available
Quantification eaasy by
qRT-PCR
Biological targets can be
identified
Molecular mechanisms at
least partially elucidated
Drosha
Dicer
RISC complex
Argonaute proteins

LA MÉTHYLATION DE L’ADN SUR LES CYTOSINES

MÉTHYLATION DE L’ADN ET DÉVELOPPEMENT

MÉTHYLATION DE L’ADN ET TRANSCRIPTION ?

UN NOUVEAU PARADIGME
P Exons
P Exons P Exons
P Exons P Exons

TOUJOURS TOUT FAUX
Gene name N of corr N of significant
corr
Best Pearson corr
(p-value) Promoter Gene body
APAF1 2 0 --- --- ---
BCL2 1 1 0.451 (0.002) --- 1 (+)
CASP3 2 0 --- --- ---
CD247 12 12 -0.595 (<0.0001) --- 12 (-)
CD28 2 2 -0.495 (0.0005) 2 (+) ---
CD3D 4 1 -0.500 (0.0004) --- 1 (-)
CD3E 2 2 -0.546 (<0.0001) --- 2 (-)
CD3G 3 1 -0.335 (0.02) 1 (-) ---
FCER1G 3 0 --- --- ---
HLA-A 3 0 --- --- ---
HLA-B 2 0 --- --- ---
HLA-C 2 0 --- --- ---
HLA-DMA 3 1 0.382 (0.009) --- 1 (+)
HLA-DMB 9 7 0.464 (0.001) --- 7 (+)
HLA-DOB 2 1 0.307 (0.04) 1 (-) ---
HLA-DQB1 12 7 0.656 (<0.0001) 1 (+) 4 (+), 2 (-)
HLA-DRB1 17 11 0.898 (<0.0001) 1 (-) 6 (+), 4 (-)
HLA-E 7 0 --- --- ---
IFNGR2 3 0 --- --- ---
IKBKB 1 0 --- --- ---
IL1B 1 1 -0.291 (0.05) --- 1 (-)
IL1RAP 1 0 --- --- ---
INS 1 0 --- --- ---
IRF1 1 0 --- --- ---
LTA 13 10 -0.377 (0.01) 1 (+) 9 (-)
MAP2K3 2 0 --- --- ---
MAP3K14 1 0 --- --- ---
MAP3K5 3 0 --- --- ---
MAPK14 1 0 --- --- ---
MAPK9 1 0 --- --- ---
NFKBIA 1 0 --- --- ---
NGFR 2 1 -0.360 (0.01) 1 (-)
SOCS2 1 1 0.503 (0.0004) 1 (+)
STAT1 4 2 -0.418 (0.004) 1 (-)
TNFRSF1B 1 1 -0.305 (0.04) 1 (-)

(Fraga, 2005)
MISE EN ÉVIDENCE DES MODIFICATIONS
ÉPIGÉNÉTIQUES AU COURS DE LA VIE ET DES
EXPOSITIONS DE L’INDIVIDU

ET LE CODE DES HISTONES?

CODE DES HISTONES ET TRANSCRIPTION

ALIMENTATION ET ÉPIGÉNÉTIQUE: ORIGINE DES
RÉSIDUS MÉTHYLÉS
• Plantes feuillues
(épinards, asperges,
laitue)
• Pois, lentilles, haricots
• Jaune d’œuf
• Levure de boulanger
• Céréales de petit
déjeuner
• Foie
• Rognons

EPIGÉNÉTIQUE ET MICROARN
(Udali et al, 2012)

3
Long Noncoding RNAs in Cardiac Development and Pathophysiology. Schonrock, Nicole; Harvey, Richard; Mattick, John Circulation Research. 111(10):1349-1362, October 26, 2012. DOI : 10.1161/CIRCRESAHA.112.268953
Figure 2 . Overview of the current landscape of non coding RNAs (ncRNAs). A, ncRNAs can be superficially divided into 2 classes based on their size, long (>200 nt) and short (B, Characteristics of 4 long ncRNA classes. Enhancer RNAs (eRNAs) derived from bidirectional transcription of enhancer domains marked by histone 3 lysine 4 monomethylation (H3K4me1) are 5' capped but generally are not polyadenylated. Long intergenic ncRNAs (lincRNAs) are generated from independent promoters, characterized by the presence of histone 3 lysine 4 trimethylation (H3K4me3), distal to and not overlapping any protein-coding gene. In contrast, natural antisense transcripts (NATs) arise from independent transcription of the opposite DNA strand and may be intronic, intragenic, or bidirectional to protein-coding genes, and may share some sequence complementarity with their sense transcript. Despite having a 5' cap, 3' untranslated region (UTR)-derived RNAs (uaRNAs) are unlikely to be independently transcribed and may be a cleavage product of a longer messenger RNA (mRNA) precursor.
LA COMPLEXITÉ DES ARN NON CODANTS VA AU-
DELÀ DES MICRORNA

EN RÉSUMÉ…
Méthylation de l’ADN
A quel endroit?
Qui le fait?
Le code des histones
Ce sont des modifications post-traductionnelles
A des positions particulières (plutôt près de la queue N-terminale
Souvent des méthylations et des acétylations
Les ARN non codants

Comment l’organisme maternel peut-il, chez les
mammifères vivipares, accepter pendant des
semaines, des mois, voire plus d’une année, un
organisme pour moitié d’origine paternelle, comme
une véritable greffe ?
P Medawar 1953

IMMUNITÉ ET PLACENTATION

LES LIMITES DE LA TOLERANCE IMMUNITAIRE

QUELLES SONT LES CAUSES DE
L’ACCOUCHEMENT?
(Marcellin et al J Immunol, 2017)

QUELLES SONT LES CAUSES DE
L’ACCOUCHEMENT?
(Marcellin et al J Immunol, 2017)

UN ÉQUILIBRE FRAGILE
Défauts immunologiques = placentation
déficientes
Flux utéro-placentaire anormal
RCIU
Prééclampsie

LA PRÉÉCLAMPSIE: UNE MALADIE
GESTATIONNELLE HUMAINE MAJEURE
• Preeclampsia:
3% to 8% of pregnancies
A major cause of materno-foetal morbidity and mortality
Only cure: the placenta and foetus delivery
Risk factors
Primiparity
Chronic hypertension
Diabetes
Obesity
preeclampsia is a multifactorial disease
Specifically human

CE QUE L’ON SAIT
Clinical definition (from the 20th week of amenorrhea):
hypertension (>140:90; >160:110 for severe cases)
proteinuria (>300 mg/day)
Physiopathology :
Early defect : abnormal placentation
Shallow invasion hypoperfusion
• How does the placental defect lead to a systemic maternal syndrome ?
Normal Preeclampsia
(Redman, Sargent, 2005)

LES CAUSES ET LES OUTILS

DU DÉFAUT IMUNITAIRE À L’ANOMALIE DE
PLACENTATION, VERS LA DYSFONCTION ENDOTHÉLIALE
GÉNÉRALISÉE

POURQUOI (COMMENT) LA PRÉECLAMPSIE
EXISTE-T-ELLE?
Un paradoxe biologique
Une maladie grave: pays en voie de développement, Europe avant la médecine moderne
Une maladie pouvant tuer les femmes quand elles vont donner la vie
Une maladie fréquente
Un avantage sélectif?
Modélisation:
Une pathologie de la première gestation
La gravité diminue avec les grossesses consécutives s’il n’y a pas de changement de partenaire
Hypothèses: Une probabilité d’avoir des allèles à risque
Une probabilité de décéder si la pathologie se déclare
Ce qui change les fréquences alléliques à la génération suivante

Education par
les mères Education
par les
pères
Education par les mères
Education par les
pères
Encéphalisation
Corrélation conceptualisée entre
accouplement et gestation
notion de filiation paternelle qui
apparait dans l’évolution
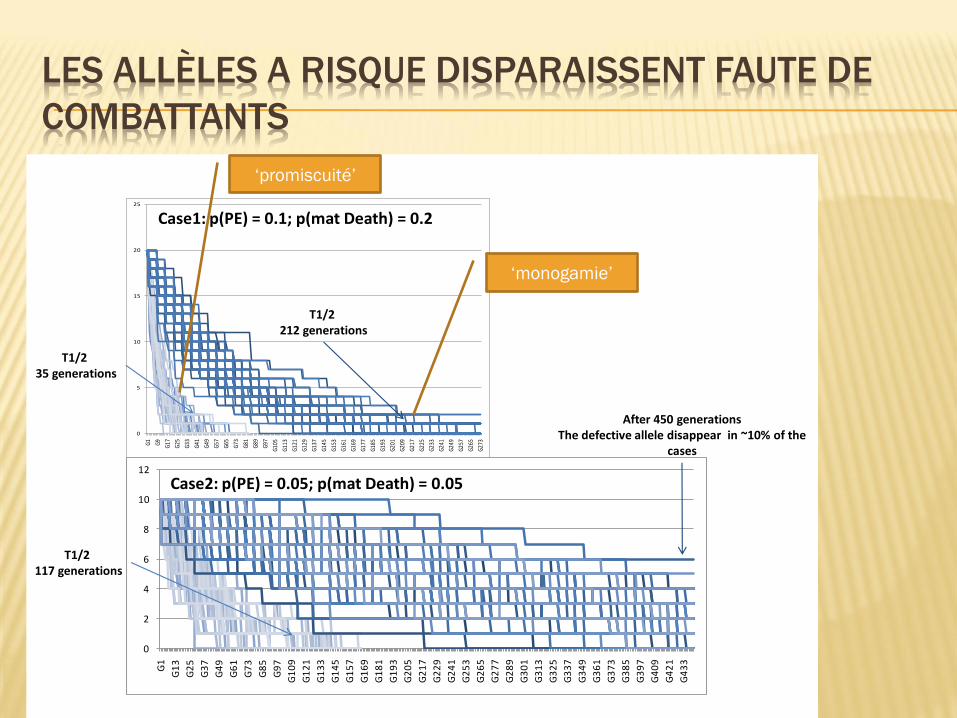
LES ALLÈLES A RISQUE DISPARAISSENT FAUTE DE
COMBATTANTS
0
5
10
15
20
25
G1
G9
G17
G25
G33
G41
G49
G57
G65
G73
G81
G89
G97
G10
5
G11
3
G12
1
G12
9
G13
7
G14
5
G15
3
G16
1
G16
9
G17
7
G18
5
G19
3
G20
1
G20
9
G21
7
G22
5
G23
3
G24
1
G24
9
G25
7
G26
5
G27
3
0
2
4
6
8
10
12
G1
G1
3
G2
5
G3
7
G4
9
G6
1
G7
3
G8
5
G9
7
G1
09
G1
21
G1
33
G1
45
G1
57
G1
69
G1
81
G1
93
G2
05
G2
17
G2
29
G2
41
G2
53
G2
65
G2
77
G2
89
G3
01
G3
13
G3
25
G3
37
G3
49
G3
61
G3
73
G3
85
G3
97
G4
09
G4
21
G4
33
Case1: p(PE) = 0.1; p(mat Death) = 0.2
Case2: p(PE) = 0.05; p(mat Death) = 0.05
T1/2 35 generations
T1/2 212 generations
T1/2 117 generations
After 450 generationsThe defective allele disappear in ~10% of the
cases
Figure 1 (Chelbi et al.)
‘monogamie’
‘promiscuité’

CONCLUSION ET QUESTIONS
Les allèles ‘à risque’ entraînent une disparition
de leurs porteurs
Leur fréquence diminue puis disparaît
Quid de nos proches cousins?

UN INVESTISSEMENT PATERNEL DIFFÉRENT
?

UN ÉQUILIBRE À TROUVER
Fitness
Complete endogamyIncreased consanguinity
Decreases Fitness
Complete exogamyIncreased risk of PEDecreases Fitness
Figure 2 (Chelbi et al.)Des gènes répondent-ils à ce modèle?

CONCLUSIONS ET QUESTIONS
Estimation du rapport coût/bénéfice
Evolution à venir de la viviparité?
Les mêmes mécanismes sont ils à l’œuvre
dans les autres groupes vivipares du monde
vivant
Les plantes supérieures
Des vertébrés dispersés dans l’arbre évolutif
empreinte parentale?
Recommended

















