
Roux' Archiv ffir En~wicklungsmech~nik 154, 405-416 (1963)
Aus dem Zoologischen Institut der Universiti~t Bonn
DIE E N T S T E H U N G DES FARBMUSTERS BEIM L A K E N F E L D E R H U H N
Von ROLF DANNEEL und HELGA SCHUl~ANN
Mit 5 Textabbfldungen (12 Einzelbilder)
(Eingegangen am 14. Miirz 1963)
Die unumstrittene Tatsaehe, dal~ alle Pigmentzellen der Wirbeltiere aus der embryonalen Neuralleiste stammen und yon da aus aktiv an ihren Bestimmnngsort wandern, erSffnet neue Wege zur Efforschung der Musterbildung, die sich aus logisehen ~Jberlegungen ergeben.
Zun~ehst besteht natfir]ieh die MSgliehkeit, dal~ die Neuralleiste iiberhaupt keine Melanoblasten produziert. Dieser Fall ist beispiels- weise beim wei•en Wiener Kaninehen verwirklieht, das zwar blaue Augen hat, weft das Retinapigment an Or~ und Stelle, d. h. unabhi~ngig yon der Neuralleiste entsteht, das aber in der Epidermis und in den Haar- wurzeln zu keiner Zeit der Entwicklung Melanoeyten besitzt.
Zweitens kann dutch genetische Einfifisse die Anzahl der in der Neuralleiste gebildeten Pigmentzellen reduziert sein. In diesem Falle ist zu erwarten, dal~ bei einer gleiehmi~l~igen Verteilung der wenigen verffigbaren Pigmentzellen ein helleres Farbkleid entsteht, bei un- gleieher, vom Zufall abh~ngiger Verteilung dagegen ein unregelm~l~iges Farbmuster wie bei den hier zu besprechenden Lakenfelder Hiihnern.
Eine dritte Variante ist dann zu erwarten, wenn zwar Melanocyten in ausreichender Anzahl entstehen, aus irgendwelehen Grfinden aber fiberhaupt nieht oder nur unter bestimmten Bedingungen zur Pigment- synthese befahigt sind. Beispiele hierfiir liefern die Albinos und die l~ussenkaninchen. Bei beiden Mutanten enthalten die Haarwurzeln eine normale Anzahl yon Melanocyten, die jedoeh bei Albinos fiberhaupt kein Pigment bilden kSnnen, bei den Russenkaninchen nur bei relativ geringer Hauttemperatur. In beiden Fifllen reicht, wie man schon lange weiB, die vorhandene Tyrosinase nieht zur Pigmentbildung aus (DAN~EL 1941).
Eine weitere MSgliehkeit der Musterbildung besteht darin, da~ die yon der Neuralleiste gelieferten Pigmentzellen versp~tet gebildet werden oder langsamer wandern. Auf diese Weise entsteht z. B. die weil~e Zone der gescheckten Haubenrat te und die weil3e Blesse der Maus (WAGwN]~ 1959~ SCHUMAN~ 1960). In beiden F~llen wirkt sich die Versp~tung
Wilhelm Roux' Arch. Entwickl.-Mech. Org., Bd. 154 28

406 ROLF DANNEEL und I-[ELGA SCHUMANN:
der P igmentze l len dah in aus, dab die Melanocy ten n icht mehr alle KSrper reg ionen rechtzei t ig , d . h . vor Beginn der Hautd i f fe renz ie rung und -verd ich tung erreichen. Diese Stel len ble iben dann weilL
B e k a n n t is t aueh die W i r k u n g des Gens d (dilute), das nach MAR- X]~T und SILVerS (1959) bei M~usen eine R e d u k t i o n der P igment - ze l ldendr i ten und so ve rmut l i eh eine StSrung der U b e r t r a g u n g des P igmen t s auf die Haarze l l en bewirk t .
Zu e rwghnen is t sehlieBlich noch, dal~ schon vorhandene tg t ige Melanocy ten zum Teil nacht r~gl ich wieder ver loren gehen kSnnen. Dies is t z . B . be im Si lberkan inchen und be im Al te r se rgrauen der Fa l l (I~EINTJS, S ]957, WEISSE~FV~LS 1954).
D a m i t s ind die mSgl iehen Wege zur Mus te rb i ldung aber sieher noch n ich t ersehSpft , da es bei S~uget ieren und VSgeln eine Reihe yon Streifen- u n d F l e e k e n m u s t e r n gibt , yon deren E n t s t e h u n g m a n sich vore rs t noch keine begrf indete Vors te l lung m a c h e n kann.
Das Lakenfe lder Huhn , von dem hier die Rede sein soll, in teress ier te uns deswegen, weft das F a r b m u s t e r dieser Tiere demjen igen der t L u b e n - r a t t e sehr ~hnlich sieht, t robzdem aber auf ganz andere A r t zus tande kommt .
Da m a n bei Un te r suchungen fiber die Musterbf ldung ein Bezugs- sys tem brauch t , haben wi t zum Vergleieh eine einfarbige schwarze Hfihnerrasse herangezogen, n~mhch sehwarze l~heinl~nder.
Material und Methoden
Die ve rwende ten Lakenfe lder Hf ihner s t ammten , ebenso wie die schwarzen Rheinl / inder , yon Spezia lzf ichtern u n d wurden bei uns wei ter ve rmehr t .
Zur Untersuchung der Hautdifferenzierung und der Melanocytenwanderung wurden die Embryonen naeh verschieden langer Bebriitung dem Ei entnommen und je nach GrSl~e unzer]egt fixiert (BouI~) oder zuvor in Stiicke zerschnitten. Den schon geschliipften Kiicken wurden nur Hautstiicke entnommen und mit der Unterseite auf Filtrierpapier geklebt, damit sich die Explantate bei der Fixierung nieht einrollten. Die 7--10 # dicken Serienschnitte wurden in allen F~]len mit Azan oder Kerneehtrot ge~rbt bzw. nach ~/[ASSON versflbert. Auf eine Nach- f~rbung der versilberten Sehnitte haben wir verzichtet, weil das Pigment junger Melanocyten in gef~rbten Pr~paraten leieht fibersehen werden kann.
Fiir die Gewebekultttren verwendeten wir 4 Tage alte Embryonen, die in drei Absehnitte zerteilt wurden, n~mlich den Kopf, den vorderen Rumpf und das hintere Drit~el mit dem Sehwanz. Die drei Teile wurden dann jeweils in Stfickehen geeigneter Gr6~e gesehnitten. So konnten wir das Verhalten der Me]anocyten in den drei KSrperregionen gesondert und much vergleichend untersuchen.
Zum Ansetzen der Kulturen verwendeten wir eine vereinfachte Methode, well je Embryo etwa 30 Explantate anfielen, deren Zfichtung in Maximow-Kammern oder Rollertubes sehr umst~ndlieh gewesen wgre. Wir fibertrugen die Kulturen auf Deekgl~ser, legten diese in Petrischalen ein und iibersehichteten sie gemeinsam mit dem fiblichen Nahrungsgemiseh aus Embryonalextrakt, Hiihnerplasma,

Entstehung des Farbmusters beim Lakenfelder l~uhn 407
menschlichem Nabelschnurserum und PC-LSsung. Die Kulturen wurden dann 4 T~ge lang bei 370 gehalten, anschlie~end n~ch BOUIN fixiert, nach MAssoN versilbert und ohne I~achf~trbung in Eukitt eingedeckt.
1. Das Federkleid des erwachsenen Tieres
a) Die verschiedenen Federn und ihre Fiirbung I m Gefieder eines I-Iuhnes kann man drei Federtypen unterscheiden,
die Konturfedern, die Dunenfedern und die H~arfedern.
Die Kontur]edern tragen am Federsehaft zweireihig ~ngeordnete Seiteniiste, die , ,Rami", deren sekundi~re Verzweigungen, die ,,Radii", kleine H~kchen besitzen, die ,,lgadioli", die ihrerseits dazu dienen, die Seiteni~ste lest miteinander zu verbinden. So entsteht die fiir die Konturfeder char~kteristisehe Fahne. Die Dunen/edern, denen die lgadioli fehlen, bilden keine Fahne aus, wi~hrend die Haar/edern aus einem sehr dfinnen Sehaft bestehen, der an seinem distalen Ende in der Regel eine kleine, rudiment~re Fahne tr~gt.
Die Konturfeder besitzt auger der eigentliehen F~hne an ihrer Basis stets einen Dunenteil. Das Verh~ltnis der Fahne zum Dunenabschnitt ist jedoeh bei den einzelnen Konturfedern verschieden. So steht dem grof$en Fahnenteil der Steuer- und Schwungfedern ein sehr kleiner Dunenabsehnitt gegenfiber, w~hrend es bei den Deckfedern des l~umpfes umgekehrt ist. Bei t tfihnern finder sich ferner bekanntlieh ein Ge- sehleehtsdimorphismus. So sind beim Hahn vor Mlem die Konturfedern des Sehwanzes, des Sattels und des tIalsbehanges reicher differenziert Ms bei der Itenne.
Das Federpigment der Lakenfelder ttiihner geh6rt zu den Melaninen und ~5rd yon Melanoeyten synthetisiert, die wie gesagt, aus der NeurM- leiste s tammen und durch die Subcutis in die Epidermis und yon da aus ~n die Federwurzeln einwandern, wo sie alsbald mit der Pigment- bildung beginnen. Die F~rbung der Federn hi~ngt dann yon der Anzahl, der Lage und der Aktivits der vorhandenen Melanocyten ab.
Das Farbmuster des erwachsenen La/cen]elder Huhnes (Abb. 1) er- innert, wie gesagt, an die Zeiehnung der t taubenrat te . Kopf, Hals und Sehwanz der Tiere sind schw~rz, w~ihrend der Rumpf mehr oder weniger weil3 aussieht. Das Lakenfelder Huhn ist aber nicht rein sehwarzwei~ gemustert, sondern besitzt sowoh] in den sehw~rzen Ms vor Mlem in der weil~en Gefiederregion stets eine mehr oder weniger grol~e Anzahl yon gemusterten Federn (PADB~G 1957). In diesen gemusterten Federn liegen weil~e und pigmentierte Areale in unregelm~i~iger Anordnung nebeneinander. Die mikroskopische Beobachtung hat darfiber hinaus ergeben, dal~ sich der Ubergang yon den gef~rbten zu den weiBen Feder~bsehnitten stets a]lm~hlieh vollzieht.
28*

408 t~OLP D A ~ E L und HELGA SC/tlJMANN:
Ein Zusammenhang zwisehen der Waehstumsgesehwindigkeit der Federn und ihrem Zeiehnungsmuster besteht nicht. Dies lgl~t sieh be- sonders gut an den Sehwungfedern der Fliigel beobaehten, wo die Form und die GrSl3e der Federn keinerlei Beziehung zum Zeichnungsmuster aufweist.
Aueh eine physiologiseh bedingte rhythmisehe Pigmentproduktion, wie sie den Melanoeyten maneher gebgnderter Federn eigen ist, liegt hier nieht vor. Alle unsere Beobaehtungen (s. Kapitel 2) spreehen viel- mehr dafiir, dag die Zeiehnung der Feder bei den Lakenfelder H/ihnern
Abb . I a u. b. H a h n u n d I-Ienne der L a k e n f e l d e r H i i h n e r r a s s e
eine Folge der ersten zuf/~liigen Besiedlung mit Melanoeyten ist und dutch deren Anzahl und Anordnung im Federfollikel bestimmt wird. Eine Ausnahme hiervon machen lediglich die langen sehmalen Sattel- federn des Hahnes, die alle das gleiehe Zeichnungsmuster besitzen. Ihr Dunenteil bleibt stets weil3, w/~hrend die Fahne etwa zur Hs aus- gef~rbt ist.
Die Gesamtf/~rbung des Gefieders ist im iibrigen bei den einzelnen Individuen reeht versehieden. Insbesondere variiert das zahlenmgBige Verhgltnis der sehwarzen bzw. wei6en Federn zu den gemusterten Federn von Tier zu Tier, wobei allerdings die Anzahl der einfarbigen Federn in den entspreehenden Bezirken stets weir tiberwiegt. Nut die Sehwungfedern der Flfigel sind bei allen Individuen ausnahmslos ge- mustert, doch gibt es aueh hier alle Abstufungen von fast sehwarz bis zu fast weiB.
b) Die Fiirb~ng au]einander/olgender Fede~yenerationen Wie WILLI~ und I~AWL~S (1940) nachweisen konnten, besthnmen
die in den Federkeim eingedrungenen Melanoeyten nieht nut die Farbe, sondern aueh das Zeiehnungsmuster der Feder. Die Federn eines Ilaut- transplantates fgrben sieh deshalb nieht wirtsgemgl3, sondern spender-
Entstehung des Farbmusters beim Lakenfelder Huhn 409
gem/iI3 aus. Vorher hatte schon K u ~ x (1932)gezeigt, dal3 die ge- musterten Federn verschiedener Tauben- und Hiihnerrassen bei Rupf- versuchen fiber mehrere Federgenerationen ihre jeweilige Zeichnung beibehielten. In diesem Fall handelte es sich aber um genetisch fest- gelegte Muster, die bei allen Individuen der betreffenden Rasse fiberein- stimmten. Uns interessierte daher die Frage, ob anch die so uneinheitlieh gemusterten Federn des Lakenfelder Huhnes ihre Zeiehnung yon einer Federgeneration zur anderen beibehalten, oder ob beim Federweehsel eine Umordnung der zuriiekgebliebenen Melanoeyten stattfinden kann.
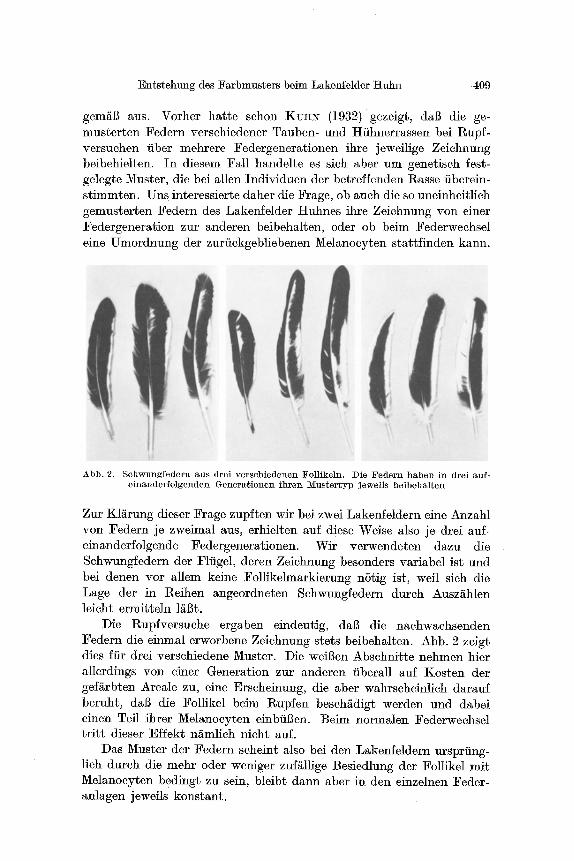
Abb. 2. Schwnngfedera aus drei verschiedenen Follikeln. Die Federn haben in drei anf- e inanderfolgenden Genera,tionen ihren 1Vlustertyp jeweils be ibehal ten
Zur K1/irung dieser Frage zup~ten wir bei zwei Lakenfeldern eine Anzahl von Federn je zweimal aus, erhielten auf diese Weise also je drei auf- einanderfolgende Federgenerationen. Wir verwendeten dazu die Schwungfedern der Flfigel, deren Zeichnung besonders variabel ist und bei denen vor allem keine Follikelmarkierung nStig ist, weft sieh die Lage der in Reihen angeordneten Sehwungfedern dutch Ausz/ihlen leieht ermitteln ]/~Bt.
Die Rupfversuche ergaben eindeutig, dal~ die nachwachsenden Federn die einmal erworbene Zeichnung stets beibehalten. Abb. 2 zeigt dies ffir drei verschiedene Muster. Die weil~en Abschnitte nehmen hier allerdings yon einer Generation zur anderen iibera]l auf Kosten der gef~rbten Areale zu, eine Erseheinung, die aber wahrscheinlieh darauf beruht, daI3 die Follikel beim l~upfen beseh~digt werden und dabei einen Tell ihrer Melanocyten einbfil3en. Beim normalen Federweehsel t r i t t dieser Effekt ni~mlich nieht auf.
Das Muster der Federn seheint also bei den Lakenfeldern ursprting- lieh dureh die mehr oder weniger zuf~llige Besiedlung der Follikel mit Melanocyten beding~ zu sein, bleibt dann aber in den einzelnen Feder- anlagen jewefls konstant.
410 ROLF DANNEEL und HELGA SCHUMANN:
2. Das Dunenkleid des Kiikens
Wie Untersuchungen yon WAGENX~ (1959) an der Haubenra t te und von S c ~ u ~ A ~ (1960) an Blessem~usen gezeigt haben, wird d~s Schek-
kungsmuster bei den Iq~ge- tieren w~hrend der Embryo- nalentwicklung festgelegt und bleibt d~nn unver~ndert. Auch in diesem Punkt ver- halten sieh die Lakenfelder Hiihner anders. Hier wer- den n~mlich zwei verschie- dene F~rbkleider ausgebildet, das Kiikenmuster und das Gefiedermuster des erwach- senen Tieres.
Das Federkleid des friseh geschlfipften Kfikens be- steht aus Erstlingsdunen, die schon in der zweiten H~lfte der Embryon~lzeit ange]egt werden. Auch bei den Embryonaldunen k~nn man drei verschiedene F~rb- typen unterseheiden, n~m- Heh hellgelbe, gemusterte und schw~rze Dunen. Die gelben Dunen gleichen den Embryonaldunen anderer Hiihnerrassen, wie z. B. der Leghorns, nnd sollen bier nicht weiter besprochen werden, zumal dus gelbe Kfikenpigment nieht yon Melanocyten produziert wird.
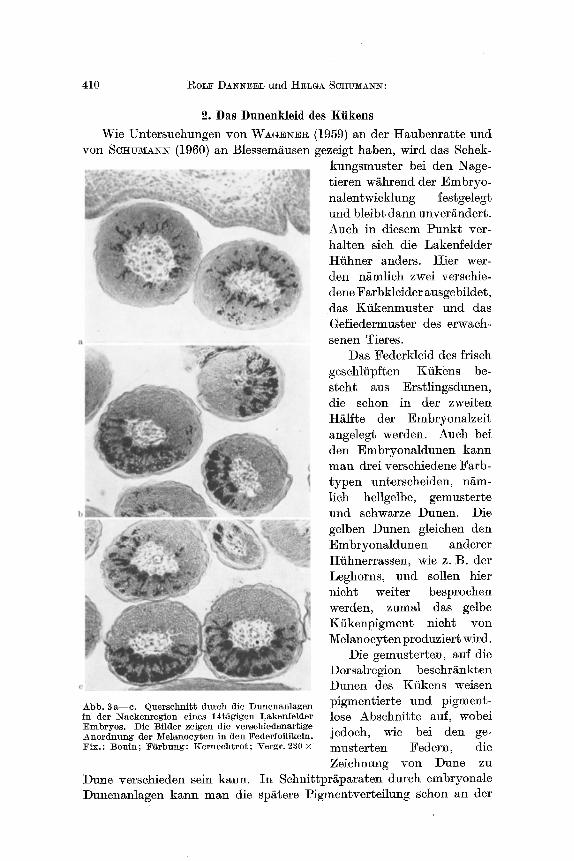
Die gemusterten, ~uf die Dorsalregion beschrs Dunen des Ktikens weisen
Abb. 3 a--c. Querschnitt durch die Dunenanlagen pigmentierte und pigment- in der l~ackenregion eines 14tfigigen Lakenfelder lose Abschnit te auf, wobei Embryos. Die Bilder zeigen die verschiedenartige jedoch, wie bei den ge- Anordnung tier l~{el~nocyten in den Federfollikeln. Fix.: Bonin; F&rbung: Kernechtrot; Vergr. 230 x musterten Federn, die
Zeichnung yon Dune zu Dune verschieden sein kann. In Schnit tpr~paraten dureh embryonale Dunenanlagen karm man die sp~tere Pigmentverteilung sehon an der

Entstehung des Farbmusters beim Lakenfelder Huhn 411
Melanoeytenanordnung im Federkeim erkennen. Sie ist auBerordentlieh variabel, wie die Abb. 3a - -c fiir die Dunenanlagen im Naeken eines 14t/tgigen Lakenfelder Embryos zeigen.
Diese ungleiche Besiedlung benachbarter Follikel h/~ng~ ganz often- bar mit den nieht genauer bekannten und daher zuf/~llig erseheinenden Wanderwegen der einzelnen Melanoeyten zusammen, die sich ja auger- dem in dem jungen Federkeim noch eine Zeitlang dutch Teilung ver- mehren k6nnen.
Gemusterte und schwarze Dunen gibt es, wie gesag~, nut in der dorsalen Region des Lakenfelder Kiikens, doch ist das Ausbreitungs-
A b b . 4 a u . b . D i e g l e i c h e n T i e r e w i e a u f _A_bb. 1 a l s 5 T a g e a l t e K i i k e n
gebiet der dunkel pigmentierten Zone bei den einzelnen Tieren sehr verschieden gro$. Abb. 4a und b zeigen zwei 5 Tage alte Lakenfelder Kiiken. W/~hrend das eine Tier (Abb. 4a) nur am ttinterkopf einen kleinen Bezirk mit dunkel gef/~rbten Dunen aufweist, besitzt das andere (Abb. 4b) auf der l~iiekenmitte einen langen, pigmentierten Streifen, der am Hinterkopf beginnt und sich bis zum Sehwanz erstreekt. An den Fliigeln beider Kfiken sind schon die ersten gemusterten Federn zu sehen.
Wie bei vielen VSge]n kann man auch beim Lakenfelder Huhn nicht yon dem Zeichnungsmuster des Dunenkleides auf dasj enige des sp/~teren Federk]eides sehlieBen. Die beiden Bilder Abb. 1 zeigen dieselben Tiere, wie die Photos der Abb. 4a und b, diesmal aber kurz vor der Ge- schlechtsreife. Hahn nnd Henne sehen jetzt fast gleich aus, obwohl das Dunenkleid des Hahnenkiiekens fast einfarbig gelb war, w/ihrend die Henne als Kfiken einen breiten pigmentierten Dorsalstreifen besal~.
3. Die embryonalen Melanocyten der Lakenfelder und der einfarbig sehwarzen Rheinl~inder
Die ersten Pigmentzellen des einfarbig schwarzen Rheinliinders lassen sich beim 5--6 Tage alten Embryo nachweisen. Am 5. Embryonaltage sind die Melanocyten zwar noch auf die Epidermis der Hinterkopfregion

412 ROL~ DA~NE~L undHELGA SeHv~A~:
beschr/inkt, breiten sieh aber schon am 6. Entwicklungstage fiber den ganzen Rficken aus. Sie erscheinen yon vorneherein in recht groBer Anzahl und enthalten viel Pigment.
Die weitere Ausbreitung der Pigmentzellen verl~uft von dorsal naeh ventral und geht sehr schnell vor sich, so dab am 8. Embryonaltage bereits alle KSrperregionen Melanoeyten enthalten. Die stark ver- zweigten, ganz mit Pigment angefiillten Wanderzellen sammeln sieh in den nun folgenden Tagen fiberalI in den gerade entstehenden Dunen- anlagen, so dab die Epidermis der RheinlKnder sehon am 12. Embryonal- rage keine Pigmentzellen mehr enthi~lt. Das Rheinl~nder~Kfiken ist also einfarbig sehwarz wie das erwaehsene Tier.
Ganz anders liegen die Verhi~ltnisse beim Laken/elder Embryo. Die ersten Pigmentzellen lassen sich bier frfihestens am 7.--8. Embryonal- rage nachweisen, vergliehen mit den Rheinliindern also mit zweiti~giger Versp~tung. Die Melanoeyten, die von vorneherein in auffallend ge- ringer Anzahl auftauchen, enthalten nur ganz vereinzelte Pigment- kSrnehen und besitzen aueh noch keine Ausl~iufer. Am 7. Embryonal- tage findet man nur in der Epidermis des I-Iinterkopfes Melanocyten, am 8. Tage aueh in der Lumbalregion, we naeh LcB~ow (1957) ein zweites Entstehungszentrum ]iegt.
Erst am 10 . - - l l . Entwicklungstage erscheinen weitere Pigment- zellen, die mehr Pigment produzieren und deutliche Ausliiufer bflden. Aueh diese Melanoeyten sind zun/ichst noch auf die Hinterkopf- und Lumbalregion besehr~nkt, dringen aber schon in grSBerer Anzahl aus der Epidermis in die Dunenanlagen ein. An diesen beiden KSrperstellen entstehen also sparer sehwarze Dunen. Die wenigen, noeh fibrigen Melanoeyten breiten sieh nun allm~ihlich nach den Flanken und dem Baueh zu aus, we die ersten schwaeh pigmentierten Pigmentzellen am 16. Embryonaltage eintreffen. Ffir die Ausf~irbung der Dunen, die in- zwischen sehon fast ausgewachsen sind, kommen diese Melanoeyten jedoeh zu sp~i4. Die Kfieken bleiben daher hier gelb.
Im Zusammenhang mit den Versuehen an Haubenrat ten und Blesse- m~usen interessierte uns schliel~lieh noeh die Frage, wann die Differen- zierung der Haut bei den }tfihnerembryonen beginnt und ob aueh hier Unterschiede zwisehen den Lakenfeldern und den einfarbigen Hfihnern bestehen. Die Differenzierung der Haut verl~uft jedoch bei den Laken- feldern und den Rheinl~ndern vSllig gleich. Sie geht bei beiden tIfihner- rassen, wie bei den Ratten- und M~useembryonen, yon den Flanken aus und beginnt am 12. Entwieklungstage damit, da]~ die Epidermis zweischichtig wird und dab das Bindegewebe anf~ngt, sieh in das Cerium und die Subeutis zu gliedern. Die Hautdifferenzierung greift dann auf die benaehbarten Bezirke fiber und erreieht 3--4 Tage spi~ter die dorsale und ventrale Mittellinie des KSrpers.

Entstehung des Farbmusters beim Lakenfelder tIuhn 413
Die Scheckung beruht beim Lakenfelder Huhn also nicht, wie bei den erw/~hnten Nagetieren auf einer Ko]lision zwisehen der Melanocyten- ausbreitung und der ttantdifferenzierung.
4. Beobachtungen an Gewebekulturen
Die Melanocyten der Lakenfelder und der Rheinl/~nder verhalten sich nicht nur in vivo0 sondern auch in vitro ganz verschieden. In
Abb. 5 a u . b . Melanocyten in Gewebekul tur . a Von E m b r y o n e n tier schwarzen l%hein- l~nder, b Von Lakenfe lder E m b r y o n e n . Fix. : Bo~in; Versi lberung nach I~ASSON,
Vergr . 640 •
Explantaten aus viert/igigen Embryonen schwarzer Rheinl~nder taueh- ten schon naeh 4 Kulturtagen zahlreiche Pigmentzellen auf, n/~mlieh im Durchschnitt 98 Melanocyten pro Kultur. Explantate aus Laken- felder Embryonen gleichen Alters enthielten unter denselben Kultur- bedingungen dagegen insgesamt nur 24 Melanocy~en in 20 Kulturen.
Die pigmentbildenden Zellen der beiden Rassen nnterseheiden sich ferner auch in Gewebekulturen hinsichtlieh ihrer Pigmentproduktion. Wie die Abb. 5 a und b zeigen, enthielten die Melanoeyten der sehwarzen Rheinl/~nder (Abb. 5a) stets sehr viel mehr Pigment als diejenigen gleichaltriger Kulturen yon Lakenfeldern (Abb. 5b).
Da fiir diese Versuche jeweils arteigener Embryonalextrakt ver- wendet worden war, haben wir in Kontrollexperimenten die Extrakte ausgetauscht, d.h. Transplantate aus Lakenfelder Embryonen in Ex- trakten aus Rheinl~ndern gezfiehtet und umgekehrt. Am Ergebnis /~nderte sich dadurch jedoch niehts. Das Verhalten der Melanocyten ist also genetiseh bedingt und wird dureh das Milieu nicht beeinflu$t.

414 ROLF DANN]~ET, und tt]~LGA SC~IU~ANN:
Diskussion
Das Farbmuster des Lakenfelder Huhnes hat /~ul]erlieh groSe Ahn- liehkeit mit der Scheckung der Haubenratte, kommt aber auf ganz andere Art zustande. Insbesondere hat die Hautdifferenzierung, die bei der l~attenscheckung eine ausschlaggebende 1%olle spielt, fiir die Entstehung des Lakenfelder Musters keine Bedeutung, weft sie bei H~ihnern so sp~t einsetzt, da$ selbst sehr langsam wandernde Pigment- zellen noeh alle KSrperregionen rechtzeitig, d. h. vor der Itautdifferen- zierung besiedeln kSnnen.
Behn Vergleich der Lakenfelder mit einfarbig schwarzen Rhein- l~ndern fallen drei Unterschiede im Verhalten tier Pigmentzellen auf, die eine Erkl/~rung tier Musterbildung zulassen. Die Melanoeyten der Lakenfelder Embryonen sind yon Anfang an in relativ geringer Anzahl vorhanden, sie besiedeln ferner die verschiedenen KSrperregionen mit erheblicher Versp/itung und produzieren erst sehr sp~t die zur Aus- f/~rbung der Federn notwendige Pigmentmenge.
Die F/irbung des Dunenkleides l~ftt sich hiernaeh zwanglos deuten. Die Dunen werden n~mlich sehon am 7. Embryonaltage angelegt, wachsen sehr schnell und erreichen sehon vor dem Schlfipfen ihre volle L/~nge. Bis zum 14. Entwieklungstage sind abet beim Lakenfelder Embryo die P~gmentzellen noch auf die Hinterkopfregion und die Sehwanzwurzel beschr/~nkt. Erst nach diesem Zeitpunkt breiten sich die wenigen noch vorhandenen Melanocyten in ventraler Richtung weRer aus. Da die Dunen jetzt aber schon so gut wie ausgewachsen sind, hat eine Melanoeytenbes~edlung der Follikel nach dem 14. Em- bryonaltage keinen Einflu$ mehr auf ihre Ausf/~rbung. Schwarze Dunen gibt es beim Kiicken daher nur in den Regionen, die yon den Pigment- zellen zuerst erreicht werden, d. h. am Kopf und am Itinterende. Alle iibrigen Dunen bleiben gelb.
Ffir das Federkleid, das auf die Dunen folgt, haben diese frfih- embryonalen Vorg/~nge zum Tell keine Bedeutung mehr. Schon kurz vor dem Schlfipfen hat sich n/imlich sowohl die verzSgerte Melanoeyten- wanderung als aueh die versp/~tete Pigmentsynthese ausgegliehen. Jetzt wirkt sieh aber die ger~nge Anzahl tier verffigbaren Melanoeyten aus, die bei weitem nicht ausreicht, um das gesamte Gefieder auszuf/~rben. Nut die Folhkel des Kopfes und tier Schwanzregion, die zuerst bes~edelt werden, erhalten yon vornherein so viele Pigmentzellen, da$ dort vorwiegend schwarze Federn entstehen kSnnen. Die meisten Feder- folhkel des Rumpfes bleiben dagegen frei yon Melanoeyten, w~hrend andere so sp/~rlieh versorgt werden, da$ nut noch mehr oder weniger stark gemusterte Federn zustande kommen.
Die Entstehung des Lakenfelder Musters ist also entwicklungs- physiologisch durchaus verstKndlich. Weniger leicht zu beantworten ist die Frage nach dem Angriffspunkt des hierffir verantwortlichen

Ents~ehung des Farbmusters beim Lakenfelder Huhn 415
Erbfaktors. Eine vergleiehbare Situation seheint aber bei den Weigen Leghorns vorzuliegen, deren rein weiBe Farbe im Laufe der Jahre dutch Eliminierung der st6renden Modifikationsgene herausgezfichtet worden ist, bei denen aber immer wieder Tiere auftreten, die in ihrem weiBen Gefieder einzelne gefgrbte Federn enthalten (NAcETSlt~IM 1957). Die Leghorns haben, wie die Lakenfelder pigmentierte Augen, sind also keine Albinos. Sie besitzen vielmehr einen Pigment-Inhibitor I, der aber offensichtlieh nieht inlmer absolut dominant ist.
Bei M/~usen gibt es eine dominante Mutation ,,Splotch", die eben- fulls zu einem Seheekungsmuster fiihrt und nach AV~BAC~ (1954) prim/~r Ver/inderungen in der NeurMleiste bewirkt, wonaeh sekund/ir auch die Melanoblastenentstehung gest6rt ist. Wit halten es ffir m6g- lich, dub ffir die Lakenfelder und Leghorns /~hnliehes gilt, doeh kann hieriiber natiirlieh erst das Experiment entseheiden.
Zusammenlassung
Das Gefieder des erwachsenen Lakenfelder tIuhnes ist im groBen ganzen schwarzweiB gescheckt, doch enthalten sowohl die schwarzen als auch die weiBen Gefiederregionen stets eine mehr oder weniger groBe Anzahl yon gemusterten Federu.
Obwohl die Zeichnung dieser gemusterten Federn sehr variabel ist, behalten die Federn aus ein und demselben Follikel in aufeinander- folgenden Federgenerationen ihr Muster jeweils bei.
Das Kiicken der Lakenfelder besitzt ein anderes Muster als das erwachsene I-Iuhn. Wie ein Vergleich zwischen den Embryonen der einfarbig schwarzen Rheinlgnder und denjenigen der Lakenfelder zeigt, entstehen die Melanocyten bei der letztgenannten I-Ifihnerrasse in viel geringerer Anzahl, besiedeln die verschiedenen K6rperregionen ver- sp/itet und bilden auch weniger Pigment.
Die langsamere Wanderung und die sp/~tere Pigmentsynthese fiihren zur Ausbildung des Kfickenmusters, w/ihrend das Muster des erwach- senen Huhnes vor allem auf der verringerten Melanocytenanzahl beruht. Nur an denjenigen K6rperstellen, die in unmittelbarer N~he der beiden Entstehungszentren der Melanocyten, d .h . am Kopf und am Hinter- ende liegen, erhalten die Federanlagen so viele Pigmentzellen, dub hier schwarze Federn entstehen k6nnen. Die wenigen, weiterwandernden Melanocyten dringen nut noch hier und dort in einzelne Federkeime ein und ffihren so zu der Entstehung der in das weiBe t~umpfgefieder eingestreuten mehr oder weniger stark gemusterten Federn.
Aueh in vitro bildet Embryonalgewebe yon Lakenfeldern sehr viel weniger Melanocyten als gleichMtriges Gewebe yon sehwarzen t~hein- 1/~ndern.

416 I~OLF DA~NEEn eL al. : Entstehung des Farbmusters beim Lakenfelder Huhn
L i t e r a t u r
AU~RBACH, R. : Analysis of the developmental effects of a lethal mutation in the house mouse. J. exp. Zool. 122, 305--324 (1954).
DANNEEL, R.: Ph~nogenetik der Kaninchenfgrbung. Ergebn. Biol. 18, 55--87 (1941).
Kul:~, 0 . : Entwicldungsphysiologische Untersuchungen an der Vogelfeder. Wilhelm Roux' Arch. Entwickl.-Meeh. Org. 127, 456--541 (1932).
LUBNOW, E.: Die Pigmentierung des japanischen Seidenhuhns. Biol. Zbl. 76, 316--342 (1957).
M~KER% C. L., and W. K. SILVERS: Pigment cell biology. New York: Academic Press 1959.
NACHTS~EIN, H. : Pigmentierte Federn beim weigen Leghorn. Zfichtungskunde 29, 285--288 (1957).
PADBm~G, M. : Untersuchungen fiber die Pigmentierung des Lakenfelder IIuhnes. Diss. Bonn 1957.
t{EI~TJES, M. : Untersuchungen fiber die Silberung des ttauskaninchens. Biol. Zbl. 76, 414--428 (1957).
SCHVMAgN, H. : Die Entstehung der Scheckung bei M~usen mit weiBer Blesse. Develop. Biol. 2, 501--515 (1960).
WAGERER, G. : Die Entstehung der Scheckung bei der Haubenratte. Biol. Zbl. 78, 4 5 1 4 6 0 (1959).
WEISSENFELS, N. : Das natiirliche Ergrauen und die Depigmentierung der Haare. Biol. Zbl. 73, 3 9 9 4 1 4 (1954).
WILLIER, B. H., and M. RAWLES : The control of feather eolour pattern by melano- phores grafted from one embryo to another of different breed of fowl. Physiol. Zool. 111, 177--191 (1940).
Prof. Dr. 1~. DANNEEL , 53 Bonn, Zoologisches Insti tut der Universit~t, Poppelsdoffer Schlog
Recommended

















