
Ergebnisse 86
Obwohl nach Untersuchungen von Emig (1999) einige V-ATPase-Untereinheiten (UE) in
fast allen Saccharosedichtefraktionen nachzuweisen waren, war das Ziel der vorliegenden
Untersuchungen, die für die V-ATPase-Aktivität bedeutenden Untereinheiten zu
identifizieren, um zu prüfen, ob in den AtNHX1- und VM23-markierten Dichtebereichen
der Saccharosegradienten ein funktionsfähiges Enzym der V-ATPase vorlag.
Das Antiserum ATP88b, das gegen das Holoenzym der V-ATPase aus K. daigremontiana
gerichtet war (Haschke et al., 1989), erkannte in den HL-Präparaten aus M. crystallinum
sieben Polypeptide apparenter Molmassen von 100, 69, 55, 41, 34, 28, und 16 kDa (Abb.
23 25 31 28 33 34 36 4438 40 4241 46 47 48
17 23 31 27 34 35 37 44 49 38 40 42 41 45 46 48
A
B
34 & 32
100 kDa
5541
69
28
16
26
41
100 kDa6955
34
28
16
Saccharose [ % (w/w)]
Abbildung 3.24: ATP88b-Immunomarkierung der Saccharosedichtefraktionen:
A) Membranpräparate HL-gestresster Pflanzen, eingesetzte Proteinmenge 10 µg;
B) Membranpräparate salzgestresster Pflanzen, eingesetzte Proteinmenge 10 µg;
Zahlen mit den Pfeilen geben Molmassen von markierten Polypeptiden an.
Saccharose [ % (w/w)]
48

Ergebnisse 87
3.24 A). Dabei waren die Banden von 100 und 41 kDa sehr schwach markiert und konnten
nur in den Fraktionen von 28 % bis 34 % (w/w) Saccharose und in den Fraktionen von 41
% (w/w) Saccharose und höher erkannt werden. Das Polypeptid von 28 kDa wurde nur in
den dichten Fraktionen von 44 % (w/w) Saccharose und höher immunodekoriert. Die
anderen Polypeptide (69, 55, 34 und 16 kDa) wurden in allen Membranfraktionen
gefunden. Ein Molmassenvergleich mit den V-ATPase-Untereinheiten aus anderen
Pflanzenarten deutete darauf hin, dass diese Polypeptide V-ATPase-Untereinheiten
darstellen: a (100 kDa), A (69 kDa), B (55 kDa), C (41 kDa), D (34 kDa), Ei (28 kDa) und
c (16 kDa) (Lüttge und Ratajczak, 1997; Ratajczak, 2000). In den Präparaten aus
salzbehandelten Pflanzen wurden alle oben erwähnten Polypeptide über den gesamten
Gradienten markiert (Abb. 3.24 B). Zusätzlich konnten Markierungen von zwei Proteinen
mit einer Molmasse von 32 und 26 kDa beobachtet werden. Das Polypeptid von 32 kDa
stellt die Di-UE der V-ATPase dar (Bremberger und Lüttge, 1992: Krisch, 1999). Das
andere Protein von 26 kDa wurde auch in den früheren Untersuchungen mit den
polyklonalen V-ATPase-Antisera in M. crystallinum gefunden, aber bisher noch nicht
identifiziert (Bremberger und Lüttge, 1992; Krisch, 1999). In der vorliegenden Arbeit
konnten die Polypeptide von 32 und 26 kDa interessanterweise sehr deutlich in Fraktionen
von 44 % (w/w) Saccharose und höher bei salzgestressten Pflanzen beobachtet werden. In
früheren Untersuchungen waren diese Polypeptide im Gegensatz dazu in der
Tonoplastenfraktion von 25 % (w/w) Saccharose aus salzgestressten Pflanzen und aus
Pflanzen mit altersbedingtem CAM zu detektieren (Bremberger und Lüttge, 1992;
Ratajczak et al., 1994 b; Zhigang et al., 1996; Krisch, 1999). In den leichten
Membranfraktionen salzgestresster Pflanzen und in allen Fraktionen der HL-gestressten
Pflanzen waren diese beiden Polypeptide in den vorliegenden immunologischen Versuchen
nicht zu sehen. Dies könnte aber unter anderem auf Unterschiede in der Gesamtmenge der
V-ATPase-Komplexe in diesen Fraktionen zurückzuführen sein.
Studien an V-ATPasen aus unterschiedlichen Spezies haben gezeigt, dass die
Untereinheiten A, B und c für die Funktion dieses Enzyms essentiell sind (Forgac, 1989;
Lüttge und Ratajczak, 1997). Alle erwähnten Untereinheiten waren sowohl in leichten, als
auch in dichten von AtNHX1 und VM23 markierten Fraktionen sowohl bei HL-, als auch
bei salzgestressten Pflanzen vorhanden. Die Untereinheiten aus dem Stielbereich der V-
ATPase (C, Di, Ei) sowie die a-UE, die wie die c-UE ein membranintegrales Protein
darstellt (Lüttge und Ratajczak, 1997; Kluge, 2002), waren auch in den Fraktionen von
43 % (w/w) Saccharose und höher in Membranpräparaten salzgestresster Pflanzen

Ergebnisse 88
angereichert. Daraus konnte man ableiten, dass in der schweren Membranpopulation alle
für M. crystallinum beschriebenen Untereinheiten des V-ATPase-Enzyms vorhanden
waren (Ratajczak et al., 1994 b; Ratajczak, 2000).
3.9.2.3 Untersuchungen zur Aktivität der V-ATPase Die vorliegenden immunologischen Studien haben gezeigt, dass alle essentiellen
V-ATPase-UE bei HL- und salzgestressten Pflanzen in den Dichtefraktionen von 44 / 49 %
(w/w) Saccharose vorhanden waren. Jedoch waren einige der erwähnten V-ATPase-UE
über den gesamten Gradienten markiert (z.B A-, B-, D, c-UE), insbesondere bei
salzgestressten Pflanzen (siehe Kapitel 3.9.2.2). Das Vorhandensein der V-ATPase-UE in
nicht-tonoplastidären Endomembranen (z.B. ER oder Golgi) wird oft durch
Proteinsynthese- und Sortierungsvorgänge erklärt (Hurley und Taiz, 1989; Herman et al.,
1994; Oberbeck et al., 1994; Matsuoka et al., 1997). Dennoch zeigt dieses Enzym in den
meisten Fällen seine höchste Aktivität und Sensitivität gegen spezifische Inhibitoren am
Ort der endgültigen Lokalisierung, d.h. im Tonoplasten (Lüttge und Ratajczak, 1997). Zur
Tonoplastenidentifizierung sollten daher neben den immunologischen Untersuchungen
auch Aktivitätsmessungen der V-ATPase herangezogen werden.
Die spezifische V-ATPase-Aktivität in Membranfraktionsproben wurde anhand einer
Hemmung der ATP-Hydrolyse bei pH 8,0 (Struve und Lüttge, 1987 / 1988) durch ein zur
V-ATPase hoch affines Makrolid Concanamycin (Dröse et al., 1993; Dröse und Altendorf,
1997) bestimmt. Die Aktivitätskurven der Gradientenfraktionen beider Versuchsgruppen
zeigten einen komplexen Verlauf (Abb. 3.25 A und B). Es fiel auf, dass in
Membranpräparaten von 27 / 34 % (w/w) Saccharosekonzentration sowohl bei HL-, als
auch bei salzgestressten Pflanzen ein deutliches Hydrolyse-Maximum zu beobachten war.
Der Höchstwert lag dabei in den Präparaten salzgestresster Pflanzen im Vergleich zu HL-
gestressten Pflanzen um den Faktor vier höher. Der Unterschied zwischen den
Membranpräparaten aus HL- und salzgestressten Pflanzen war in den Fraktionen von ca.
47 % (w/w) Saccharose wesentlich größer (um den Faktor 20), wo bei salzgestressten
Pflanzen noch ein Maximum der spezifischen ATP-Hydrolyse zu sehen war.
Der Verlauf der Aktivitätsprofile der V-ATPase deutete an, dass in Membranpräparaten
aus HL- und salzgestressten Pflanzen zwei Aktivitätsmaxima der V-ATPase vorlagen,
eines bei 33 % (w/w) Saccharose und eines bei den höheren Dichten von 47 % (w/w)
Saccharose. Das erste Hydrolyse-Maximum repräsentierte die in der Literatur und in der
vorliegenden Arbeit angesprochene Tonoplastenpopulation von 25 / 30 % (w/w)

Ergebnisse 89
Saccharose, die leichte Verschiebung in den dichteren Bereich hin war vermutlich
präparationsbedingt. Der zweite und größte Peak der ATP-Hydrolyse, der in den
Präparaten aus salzgestressten Pflanzen auftrat, korrespondierte mit der
Membranpopulationen, in denen ebenfalls Markierungsmaxima einiger V-ATPase-UE (a,
D, Di, Ei und C) zu beobachten waren (siehe Kapitel 3.9.2.2).
3.9.2.4 V-PPase-Immunmarkierung Neben der Immunmarkierung der V-ATPase wurde in weiteren Experimenten die
Fraktionierung der zweiten tonoplastidären Protonenpumpe, der vakuolären
15 20 25 30 35 40 45 500
2
4
6
8
10
µmol
ATP
* h
-1 *
mg
Prot
ein-1
Saccharose [% (w/w)] 15 20 25 30 35 40 45 50
0
20
40
60
80
100
120
Abbildung 3.25: Concanamycin-sensitive ATP-Hydrolyse:
A) Membranpräparate HL-gestresster Pflanzen (MW mit SD; n = 3);
B) Membranpräparate salzgestresster Pflanzen (MW mit SD; n = 3).
A
B
µmol
ATP
* h
-1 *
mg
Prot
ein-1

Ergebnisse 90
Pyrophosphatase (V-PPase) untersucht. Ein besonderes Interesse zu diesem Ansatz konnte
dadurch begründet werden, dass eine Markierung der V-PPase in einer Kombination mit
TIPs und der V-ATPase oft zum Nachweis der Tonoplastenmembran eingesetzt wird (Hoh
et al., 1995; Kirch et al., 2000; Jiang et al., 2001).
Die Immunmarkierung durch einen gegen das V-PPase-Polypeptid gerichteten Antikörper
anti-V-PPase (Maeshima und Yoshida, 1989) konnte in den Membranpräparaten aus
salzgestressten Pflanzen in zwei Dichtebereichen beobachtet werden (Abb. 3.26). Eine
markierte Membranpopulation befand sich in den Fraktionen von 17 / 31 % (w/w)
Saccharose. Die Dichteposition dieser Fraktionen stimmte mit den
Sedimentationseigenschaften der oben angesprochenen Tonoplastenpopulation von 25 %
(w/w) Saccharose überein. Die zweite von anti-V-PPase markierte Membranpopulation lag
im Gradientenfraktionen von 40 % (w/w) Saccharose und höher. Im Vergleich zu den
leichten Fraktionen war die Kreuzreaktion mit anti-V-PPase in den schweren Fraktionen
wesentlich stärker, was mit Ergebnissen von vergleichbaren immunologischen Studien an
M. crystallinum von Emig (1999) übereinstimmte. Während in den hier untersuchten 17 /
31 % (w/w)-Fraktionen nur ein Polypeptid von der passenden Molmasse von 72 kDa leicht
markiert wurde, konnte in den Fraktionen hoher Dichte eine Doppelbande von 70 kDa und
72 kDa beobachtet werden. Die Doppelbande in Membranfraktionen von M. crystallinum
im Salzstress-CAM war ein auffälliges Ergebnis. Ein vergleichbares Markierungsmuster
wurde für die V-PPase aus K. daigremontiana und Lycopersicum esculentum beschrieben
17 26 3331 35 36 38 4439 40 4341 45 48 49
Abbildung 3.26: V-PPase-Immunmarkierung der Saccharosedichtefraktionen
salzgestresster Pflanzen, eingesetzte Proteinmenge 10 µg; Zahlen mit den Pfeilen
geben Molmassen der markierte Polypeptide an.
Saccharose [ % (w/w)]
72 kDa70

Ergebnisse 91
und im Zusammenhang mit dem Vorhandensein einer CAM-spezifischen Isoform der V-
PPase oder eines präparationsbedingten bzw. physiologischen Proteolyse-Abbaus des 72
kDa-Polypeptids diskutiert (Becker et al., 1995; Jiang et al., 2001).
Die Studien von Oberbeck et al. (1994), Robinson et al. (1996) und Ratajczak et al. (1999)
haben gezeigt, dass die V-PPase auch am Plasmalemma und in Membranen des ER, Golgi-
Apparats sowie multivesikulärer Bodies und Clathrin-coated-Vesikeln vorkommt, aber im
Vergleich zu Tonoplasten in einer nicht-aktiven Form, oder ihre Aktivität im Vergleich zu
Tonoplasten geringer ist. Folglich sollte die durchgeführte Immunmarkierung der V-PPase
durch Enzymaktivitätsmessungen ergänzt werden, um den Schluss über die
Tonoplastenidentität der hier angesprochenen Membran schwerer Dichte zu untermauern.
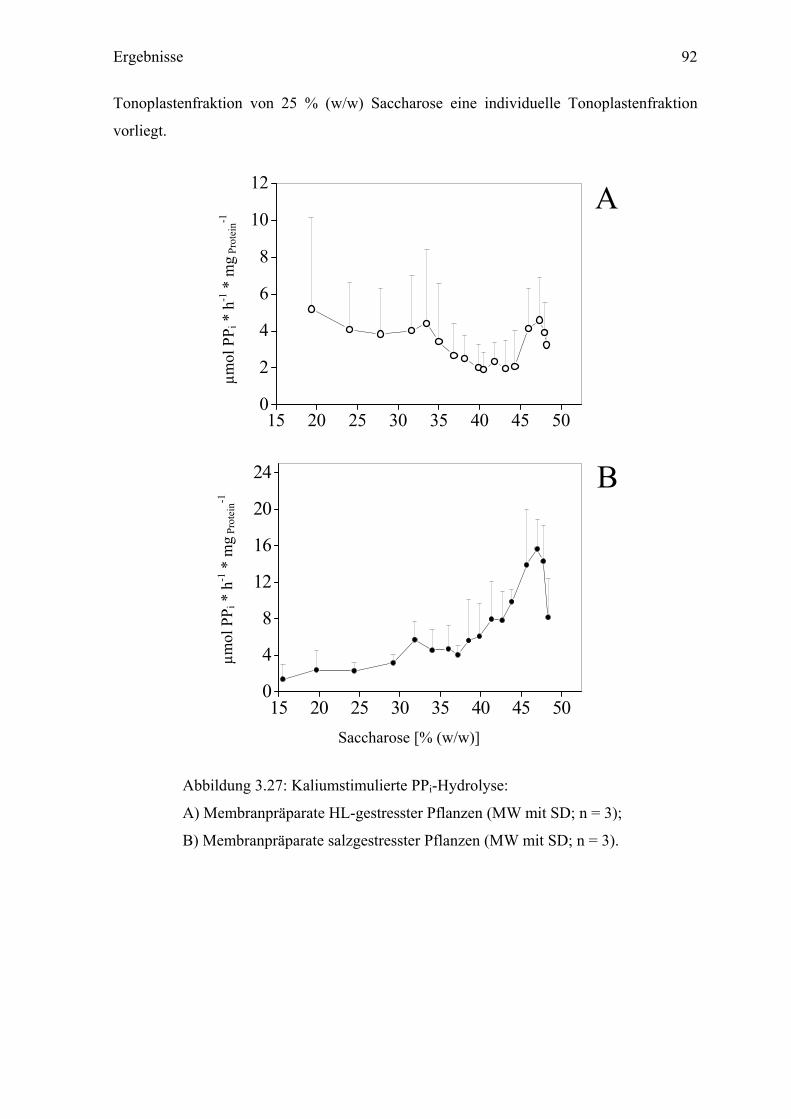
3.9.2.5 Untersuchungen zur Aktivität der V-PPase Zur Bestimmung der V-PPase wurde die kaliumstimulierte PPi-Hydrolyse bei pH 8,0
gemessen (Maeshima, 2000). In den Fraktionen bis zu 38 % (w/w) Saccharose zeigte sich
in den Membranpräparaten sowohl aus HL-, als auch salzgestressten Pflanzen eine relativ
niedrige Aktivität der V-PPase (Abb.3.27 A und B). Bei HL-Pflanzen war die V-PPase-
Aktivität in den dichteren Fraktionen ebenso niedrig mit < 6 µmol PPi * h-1 * mgprotein-1.
Hingegen stieg die PPi-Hydrolyse in den Präparaten salzgestresster Pflanzen zu den
schwereren Fraktionen hin an und zeigte ein deutliches Maximum bei 47 % (w/w)
Saccharose mit einem Hydrolyse-Wert von über 15 µmol PPi * h-1 * mg protein-1. Das in den
Membranpräparaten aus salzgestressten Pflanzen gefundene PPi-Hydrolyse-Maximum
korrelierte mit dem Maximum der V-PPase-Immunmarkierung (siehe Kapitel 3.9.2.4). In
vergleichbaren Membranpräparaten in Studien von Emig (1999) war die PPi-Hydrolyse-
Aktivität trotzt einer deutlichen V-PPase-Markierung niedriger. Die Ursache für eine hohe
PPi-Hydrolyserate in den vorliegenden Messungen könnte die Benutzung zusätzlicher
Proteaseninhibitoren, Benzamidin und Leupeptin, während der Membranisolation sein, die
möglicherweise zum Erhalt der Enzymaktivität beigetragen haben.
Fazit Zusammenfassend wies die im Rahmen dieser Arbeit durchgeführte Kofraktionierung vier
tonoplastspezifischer Proteine (Na+-H+-Antiporter, VM23, V-ATPase und V-PPase),
darauf hin, dass zumindest bei salzgestressten Pflanzen von M. crystallinum in den dichten
Gradientenfraktionen von 44 / 49 % (w/w) Saccharose zusätzlich zur bekannten
Ergebnisse 92
Tonoplastenfraktion von 25 % (w/w) Saccharose eine individuelle Tonoplastenfraktion
vorliegt.
µmol
PP i
* h
-1 *
mg
Prot
ein-1
Saccharose [% (w/w)]
µmol
PP i
* h
-1 *
mg
Prot
ein-1
Abbildung 3.27: Kaliumstimulierte PPi-Hydrolyse:
A) Membranpräparate HL-gestresster Pflanzen (MW mit SD; n = 3);
B) Membranpräparate salzgestresster Pflanzen (MW mit SD; n = 3).
15 20 25 30 35 40 45 500
2
4
6
8
10
12
15 20 25 30 35 40 45 500
4
8
12
16
20
24
B
A

Diskussion 93
4 Diskussion 4.1 Zwei Vakuolentypen im Blattgewebe von M. crystallinum Die Betrachtung der Vakuole als ein einheitliches saures Kompartiment mit multiplen
Funktionen, darunter lytischen, speichernden, homeostatischen und turgoraufbauenden ist
allgemein verbreitet (Taiz, 1992; Wink, 1993). Jedoch wurden in spezialisierten
pflanzlichen Geweben, wie Samen-Endosperm oder Kotyledonen vakuoläre
Kompartimente mit differentiellen Speicherfunktionen gefunden: proteinspeichernde
Vakuolen (protein storage vacuoles (PSVs)) mit neutralem pH und lytische Vakuolen mit
sauren pH-Werten (Holwerda et al., 1990; Holwerda et al., 1992; Nakamura und Matsuola,
1993; Hoh et al., 1995; Jiang et al., 2001). In PSVs werden gespeicherte Proteine (z.B.
Lektin und Sporamin in Gerste oder Phaseolin und Phytohemaglutinin in Bohnen) von den
in lytischen Vakuolen lokalisierten Proteasen (z.B. Aleurain) geschützt. Laut einiger
Studien ist die Koexsistenz der PSVs und lytischen Vakuolen in einer Zelle transient und
tritt z. B. bei der Entwicklung von Kotyledonen auf, wenn lytische Vakuolen degradieren
und von PSVs ersetzt werden (Hoh et al., 1995). Andererseits koexistieren diese zwei
Vakuolentypen für einen längeren Zeitraum in Aleuronzellen von Gerste und in
Tabakprotoplasten (Swanson und Jones, 1996; Di Sansebastiano et al., 1998; Swanson et
al., 1998). Ein anderes Beispiel unterschiedlicher Vakuolenspezialisierung in einer Zelle
stellen Tannin- und Wasservakuolen in motorischen Pulvinizellen von Mimosa pudica dar
(Fleurat-Lessard et al., 1997). Außer in spezialisierten Gewebetypen wurden auch in
vegetativen Geweben aus verschiedenen C3-Pflanzen Vakuolen unterschiedlicher Azidität
beschrieben, z.B. in Blättern, Wurzelspitzen und Zellkulturen von Tabak, sowie Tomaten
(Di Sansebastiano et al., 1998; Paris et al., 1996; Jiang und Rogers, 1998; Jiang et al.,
2001, Di Sansebastiano et al., 2001). In diesen Fällen sind die Funktionen der
unterschiedlichen Vakuolenkompartimente jedoch noch nicht ganz verstanden.
Eine besondere Bedeutung hat das vakuoläre Kompartiment beim CAM, indem Vakuolen
die Funktion einer transienten Speicherung der Äpfelsäure übernehmen und daher
beträchtliche Oszillationen des luminalen pH-Wertes im Tag-Nacht-Rhythmus aufweisen.
In der vorliegenden Arbeit wurde die Existenz von Vakuolen mit unterschiedlichen pH-
Werten, saure und neutrale Vakuolen, im Mesophyll von M. crystallinum im CAM-
Zustand zum Zeitpunkt der maximalen Ansäuerung (am Ende der Dunkelphase)
nachgewiesen (siehe Kapitel 3.2.1). Einen besonderen Befund stellt die Identifizierung
saurer und neutraler Vakuolen mit einem variablen Größenverhältnis innerhalb einzelner
Mesophyllzellen dar (siehe Kapitel 3.2.2). Vor diesem Hintergrund sollte betont werden,

Diskussion 94
dass neutrale Vakuolen von M. crystallinum im Vergleich zu PSVs aus C3-Pflanzen eine
andere Speicherfunktion erfüllen sollten, da bisher keine massive Proteinspeicherung in
Blättern von M. crystallinum, weder im C3- noch im CAM-Zustand, nachgewiesen wurde
(Adams et al., 1998).
4.1.1 Separate Sequestrierung von NaCl in neutralen Vakuolen und von organischen
Säuren in sauren Vakuolen beim Salzstress-CAM
Ohne Salzstress wiesen die meisten Vakuolen von M. crystallinum im CAM-Zustand, z.B.
in HL-gestressten CAM-Pflanzen, eine homogene nächtliche Ansäuerung auf. Der Anteil
von neutralen Vakuolen stieg hingegen in CAM-Pflanzen unter Salzstress bis zu Hälfte an
(siehe Tab. 4.1). Der Inhalt neutraler Vakuolen hatte offensichtlich eine niedrige
Pufferkapazität, da er den sauren pH-Wert des Gesamtblattpresssaftes aus salzgestressten
Pflanzen nicht oder nur wenig beeinflusste. Dies bedeutet, dass in neutralen Vakuolen
offensichtlich starke Salze akkumuliert werden. Eine Kristallanalyse, die eindeutig eine
massive NaCl-Speicherung in der Population neutraler Vakuolen salzgestresster Pflanzen
zeigte, bestätigte diese Annahme. Keine Kristalle von löslichen organischen Säuren oder
ihrer Salze wurden in neutralen Vakuolen gefunden. Kristalle, die denen von Äpfel- oder
Citronensäure ähnlich waren, wurden ausschließlich in der Population saurer Vakuolen
detektiert. Dies bedeutet, dass bei der Ausbildung von CAM unter Salzstress eine
differentielle Substanzspeicherung im Mesophyllvakuolen von M. crystallinum stattfand:
organische Säuren waren in sauren Vakuolen und NaCl in neutralen Vakuolen getrennt
gespeichert.
4.1.2 Speicherfunktion saurer und neutraler Vakuolen und Aktivität der
tonoplastidären Transportproteine
Natriumtransport Der Transport von Natrium in Vakuolen von M. crystallinum wird durch einen
elektroneutralen sekundär aktiven Na+-H+-Antiporter vermittelt (Barkla et al., 1995 / 2002;
Blumwald et al., 2000). Die Immunmarkierung des Na+-H+-Antiporter-Proteins in partiell
getrennten sauren und neutralen Vakuolen salzgestresster Pflanzen zeigte, dass es in einer
höheren Menge in Membranen neutraler, salzakkumulierender Vakuolen im Vergleich zu
säurehaltigen Vakuolen vorlag (siehe Tab. 4.1). Demnach konnte ein direkter

Diskussion 95
Zusammenhang zwischen der Proteinausrüstung der Tonoplasten neutraler Vakuolen und
ihrer Speicherfunktion nachgewiesen werden.
Malattransport In früheren Untersuchungen wurde in unterschiedlichen Pflanzenspezies ein einwärts
rektifizierender malatselektiver Anionenkanal nachgewiesen, der an der
Malatakkumulation in der Vakuole, u. a. von CAM-Pflanzen, beteiligt ist (Iwasaki et al.,
1992; Cerana et al., 1995; Hafke et al., 2003). In den vorliegenden Patch-clamp-Studien
wurde gezeigt, dass auch in Vakuolen von M. crystallinum ein malatselektiver Kanal
vorliegt, der hinsichtlich seiner Charakteristika, wie z.B. der Aktivierung nur bei negativen
Membranspannungen, der langsamen Aktivierungskinetik und der Diskriminierung von
Chlorid den bekannten vakuolären Malatkanälen ähnelt. Die Bedeutung dieses Malatkanals
für die vakuoläre CAM-bedingte Malatakkumulation in M. crystallinum ließ sich aus der
Tatsache erkennen, dass in Vakuolen, in denen eine Speicherung von organischen Säuren
nachgewiesen wurde, im Vergleich zu neutralen Vakuolen eine wesentlich höhere
Aktivität des Malatkanals zu registrieren war (siehe Tab. 4.1).
Das Vorhandensein des aktiven Malatkanals in Vakuolen mit der Äpfelsäure-Speicherung
ist ein entscheidendes Indiz dafür, dass der Ionenkanal der Haupttransportweg für die
Malatakkumulation in M. crystallinum ist. Bisher wurde der vakuoläre Malattransport in
diesen Pflanzen nur in indirekten Untersuchungen über eine Malatstimulierbarkeit der
Ansäuerung des Tonoplastenvesikellumens durch die V-ATPase charakterisiert (Pfeifer,
1999; Lüttge et al., 2000). Die in diesen Studien aufgestellte Hypothese eines Carrier-
vermittelten Mechanismus des Malatinflux basierte auf niedrigen Wechselzahlen des
gemessenen Transports. Dennoch diskutierten diese Autoren, dass eine relativ niedrige
Transportrate auch auf die Präparationsbedingungen zurückzuführen sein kann. Diese
Ansicht bestätigten die vorliegenden Patch-clamp-Studien, in denen eine strenge
Regulation der Aktivität des Malatkanals nachgewiesen wurde (siehe Kapitel 4.2).
Zusammenfassend wurde durch die im Rahmen der vorliegenden Arbeit durchgeführten
immunologischen und elektrophysiologischen Untersuchungen gezeigt, dass eine
differentielle Speicherfunktion der Vakuolen von M. crystallinum beim Salzstress-CAM
auf unterschiedlichen Transporteigenschaften der entsprechenden Vakuolentypen beruhte:
der höheren Menge von Na+-H+-Antiporter-Protein in der Membran neutraler
salzspeichernder Vakuolen und dem aktiven Malatkanal in der Membran säurehaltiger
Vakuolen.
Diskussion 96
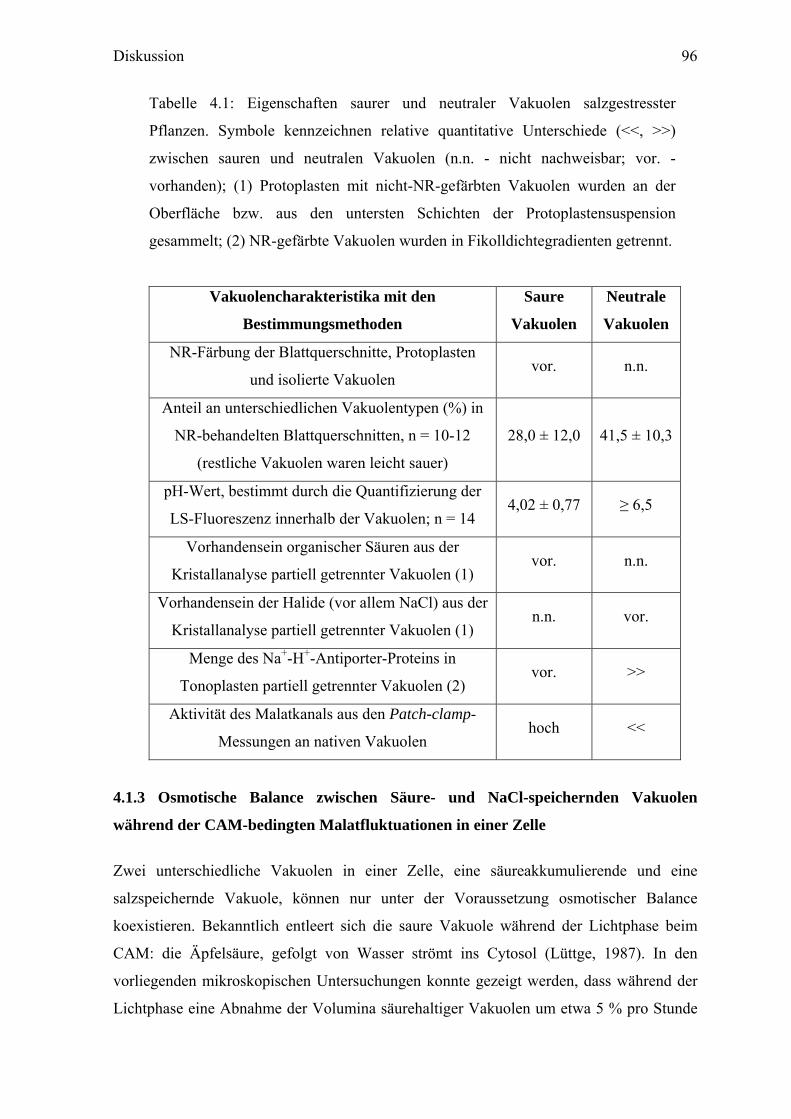
Tabelle 4.1: Eigenschaften saurer und neutraler Vakuolen salzgestresster
Pflanzen. Symbole kennzeichnen relative quantitative Unterschiede (<<, >>)
zwischen sauren und neutralen Vakuolen (n.n. - nicht nachweisbar; vor. -
vorhanden); (1) Protoplasten mit nicht-NR-gefärbten Vakuolen wurden an der
Oberfläche bzw. aus den untersten Schichten der Protoplastensuspension
gesammelt; (2) NR-gefärbte Vakuolen wurden in Fikolldichtegradienten getrennt.
Vakuolencharakteristika mit den
Bestimmungsmethoden
Saure
Vakuolen
Neutrale
Vakuolen
NR-Färbung der Blattquerschnitte, Protoplasten
und isolierte Vakuolen vor. n.n.
Anteil an unterschiedlichen Vakuolentypen (%) in
NR-behandelten Blattquerschnitten, n = 10-12
(restliche Vakuolen waren leicht sauer)
28,0 ± 12,0 41,5 ± 10,3
pH-Wert, bestimmt durch die Quantifizierung der
LS-Fluoreszenz innerhalb der Vakuolen; n = 14 4,02 ± 0,77 ≥ 6,5
Vorhandensein organischer Säuren aus der
Kristallanalyse partiell getrennter Vakuolen (1) vor. n.n.
Vorhandensein der Halide (vor allem NaCl) aus der
Kristallanalyse partiell getrennter Vakuolen (1) n.n. vor.
Menge des Na+-H+-Antiporter-Proteins in
Tonoplasten partiell getrennter Vakuolen (2) vor. >>
Aktivität des Malatkanals aus den Patch-clamp-
Messungen an nativen Vakuolen hoch <<
4.1.3 Osmotische Balance zwischen Säure- und NaCl-speichernden Vakuolen
während der CAM-bedingten Malatfluktuationen in einer Zelle
Zwei unterschiedliche Vakuolen in einer Zelle, eine säureakkumulierende und eine
salzspeichernde Vakuole, können nur unter der Voraussetzung osmotischer Balance
koexistieren. Bekanntlich entleert sich die saure Vakuole während der Lichtphase beim
CAM: die Äpfelsäure, gefolgt von Wasser strömt ins Cytosol (Lüttge, 1987). In den
vorliegenden mikroskopischen Untersuchungen konnte gezeigt werden, dass während der
Lichtphase eine Abnahme der Volumina säurehaltiger Vakuolen um etwa 5 % pro Stunde

Diskussion 97
stattfand. Dies bestätigte wiederum die Ansicht, dass nur saure Vakuolen an der
nächtlichen Malatakkumulation beteiligt waren. Neutrale Vakuolen, die zusammen mit
sauren Vakuolen in den untersuchten Zellen vorlagen, zeigten hingegen eine proportionale
Volumenzunahme. Die antiparallelen und proportionalen Volumenänderungen saurer und
neutraler Vakuolen spiegelten offensichtlich den Ausgleich der Wasserpotentiale zwischen
diesen Vakuolenkompartimenten wieder, deren Gleichgewicht bei der Remobilisierung des
Osmotikums Malats aus sauren Vakuolen, wenn in neutralen Vakuolen NaCl zurückhalten
wird, gestört war. Eine ähnliche kooperative Umgestaltung koexistierender Vakuolentypen
als Antwort auf Veränderungen im Stoffwechsel- oder / und Wasserstatus wurde auch für
Aleuronzellen mit lytischen Vakuolen und PSVs und Pulvinizellen mit Tannin- und
Wasservakuolen beschrieben (Swanson und Jones, 1996; Swanson et al., 1998; Fleurat-
Lessard et al., 1997).
Die hier beobachtete rapide Volumenvergrößerung salzspeichernder Kompartimente in der
Lichtphase könnte durch das Einbringen von neuem Membranmaterial in neutrale
Vakuolen erklärt werden. An transgenen Hefe-Kulturen wurde nachgewiesen, dass die
NaCl-Akkumulation bereits in Prävakuolen stattfindet, die nach einer Fusionierung mit
reifen Vakuolen diese mit gespeichertem Salz beladen (Nass und Rao, 1998). Diese
Autoren stellten die Hypothese auf, dass der osmotisch bedingte Wasserinflux in die
Prävakuolen diese schwellen lässt, was eine Destabilisierung ihrer Membran mit einer
nachfolgenden Fusion mit salzspeichernden reifen Vakuolen hervorruft. Einen ähnlichen
Vorgang könnte man sich in Zellen von M. crystallinum mit den an Volumen
zunehmenden neutralen Vakuolen vorstellen.
4.2 Regulation des Malatkanals Die Inkubation der vakuolären Seite isolierter Tonoplasten in einer Messlösung führte zu
einer Aktivierung des Kanals auch in den Membranen von Vakuolen, in denen in der
vacuolar-attached-Konfiguration (mit dem natürlichen Vakuolenmilieu) kein Kanalstrom
zu registrieren war: z.B. in neutralen und leicht sauren Vakuolen aus C3- oder
salzgestressten CAM-Pflanzen. Dies ist nur so zu verstehen, dass der Kanal in allen
Vakuolentypen präsent war, aber einer Aktivitätskontrolle durch das vakuoläre Milieu
unterlag. Die Suche nach entsprechenden Regulationsmechanismen wurde auf einige
wenige Faktoren beschränkt, die offensichtlich in sauren und neutralen Vakuoleninhalten
verschieden waren, nämlich die Konzentration von Malat, die vakuoläre Azidität und die
damit verbundene Konzentration von freiem Kalzium sowie luminales Ered. Weder das

Diskussion 98
Applizieren von 10 mM oder 100 mM Mal2- auf der vakuolären Seite des Tonoplasten
(Daten sind nicht gezeigt) bei einem konstanten pH-Wert, noch eine Variierung des
vakuolären pH-Wertes oder der Konzentration von freiem Ca2+ konnten die Malatströme
signifikant beeinflussen (siehe Kapitel 3.7.4.1 und 3.7.4.2). Dieser Sachverhalt stand in
guter Übereinstimmung mit Ergebnissen aus Untersuchungen von Malatkanälen in anderen
Pflanzenspezies (Iwasaki et al., 1992; Cerana et al., 1995; Hafke, 2001; Pantoja und Smith,
2002) und schloss aus, dass weder der unterschiedliche luminale pH-Wert, noch die Ca2+-
Konzentration den Malatkanal in sauren und neutralen Vakuolen primär regulieren.
Vakuoläres Redoxpotential und die Aktivität des Malatkanals In Experimenten mit isolierten Tonoplastenarealen neutraler Vakuolen trat die Aktivierung
des Malatkanals nach einer Lag-Phase von mehreren Minuten (10-15 min) auf (siehe
Kapitel 3.7.3). Dies war ein Indiz dafür, dass eine langsame Modifizierung des
Kanalproteins, nicht aber ein schneller Austausch der Lösung auf der vakuolären Seite des
Tonoplasten während des Übergangs zwischen den vacuolar-attached- und inside out-
Konfigurationen diese Aktivierung verursachte. Der zeitliche Verlauf des Effekts wies eine
Ähnlichkeit mit dem präparationsbedingten Aktivitätsrückgang von SV-Kanälen und der
V-ATPase in Abwesenheit von Reduktionsmitteln auf (Feng und Forgac, 1992 a / b;
Carpaneto et al., 1999). Für die V-ATPase wurde nämlich nachgewiesen, dass die
erwähnte Inhibierung infolge einer Oxidation der funktionellen Cystein-Reste der zum
Cytosol gerichteten katalytischen Untereinheit A (Feng und Forgac, 1992 a / b; Feng und
Forgac, 1994) und der Stiel-Untereinheit E auftrat (Tavakoli et al., 2001).
Die vorliegenden Ered-Messungen haben gezeigt, dass die beobachtete Aktivierung des
Malatkanals tatsächlich an einer Verschiebung des Ered der Kanal-Umgebung liegen
könnte, da die verwendeten Messlösungen ein wesentlich positiveres Ered im Vergleich
zum Ered des natürlichen Vakuoleninhaltes, insbesondere eines neutralen Vakuolenlumens
aufwiesen (siehe Kapitel 3.8).
Nützliche Werkzeuge für Studien von Redoxeffekten sind SH-modifizierende Substanzen
mit unterschiedlicher Fähigkeit zur Membranpermeation. Die Wirkung dieser für SH-
Gruppen hoch affinen Substanzen auf den Redoxzustand eines Proteins besteht darin, dass
sie das Verhältnis zwischen den –S-S- und SH-Gruppen des Proteins verändern (Roth et
al., 1983). Reduktionsmittel, wie DTT, Mercaptoethanol oder GSH weisen gegenüber den
SH-modifizierenden Substanzen eine entgegengesetzte Wirkung auf und können die

Diskussion 99
Effekte dieser Reagenzien in vielen Fällen umkehren oder verhindern (Bertl und Slayman,
1990; Feng und Forgac a / b, 1992; Kourie, 1998; Carpaneto et al., 1999; Oba et al., 2002).
In Anwesenheit der nicht-membrangängigen SH-Reagenzien PCMBS und DTNB auf der
vakuolären Seite des Tonoplasten wurde in den vorliegenden Untersuchungen ein
Aktivitätsanstieg des Malatkanals am Tonoplasten von M. crystallinum gemessen (siehe
Kapitel 3.7.4.3). Hingegen bewirkte das Reduktionsmittel Mercaptoethanol auf der
luminalen Seite des Tonoplasten einen Rückgang der Kanalaktivität. Diese Ergebnisse
lieferten direkte Hinweise auf eine Redoxempfindlichkeit des vakuolären malatselektiven
Kanals von M. crystallinum und wiesen darauf hin, dass die redoxempfindliche Stelle des
Kanals zum Vakuolenlumen hin exponiert ist.
Luminales Ered und die Anzahl der aktiven Malatkanäle Eine Redox-Modifikation von Transportproteinen ist von großer Bedeutung bei der
Feinregulation der Membrantransportkapazität und deren Anpassung an die aktuellen
Bedürfnisse der Zelle. In Membranen tierischer Zellen wurden viele Ionenkanäle gefunden,
die eine Redoxregulation aufweisen, indem oxidierende bzw. reduzierende SH-Reagenzien
im Cytosol die Permeabilität, das Gating oder die Deaktivierung eines Kanals modifizieren
(Kourie, 1998). Am Beispiel von Ryanodin-Rezeptor mit einer Ca2+-Kanal-Aktivität am
Sarkolemma wurde auch eine Wirkung des nicht-cytosolischen Ered gezeigt (Feng et al.,
2000). Die Hinweise auf die Redoxregulation der Transportsysteme in Pflanzen wurden in
Studien von Lucas und Alexander (1980), Lichter et al. (1981) und Thiel (1994) am
Plasmalemma von Algen-Zellen erhalten. Eine Redoxregulation von Tonoplastenkanälen
wurde bisher nur in Studien von Bertl und Slayman, (1990) an Vakuolen aus Hehe und von
Carpaneto et al. (1999) an höheren Pflanzen erforscht. In diesen Untersuchungen wurde
eine Redoxabhängigkeit der Offenwahrscheinlichkeit und der Kalziumempfindlichkeit von
kationselektiven Kanälen nachgewiesen. Die hier nachgewiesene Redoxsensitivität des
Malatstroms am Tonoplasten von M. crystallinum wirft ebenso die Frage nach
Mechanismen dieser Regulation. Da in Anwesenheit der SH-Reagenzien keine
Verschiebung des Aktivierungspotentials des Malatstroms zu beobachten war (siehe
Kapitel 3.7.4.3), schien der Redoxzustand der Kanalproteine die Anzahl von aktiven
Malatkanälen ohne Modulierung ihrer Potentialabhängigkeit zu kontrollieren. Ein
ähnlicher Regulationsmechanismus wird für vergleichbare redoxempfindliche
chloridselektive Kanäle (chloride intracellular channels CLICs) aus Endomembranen
tierischer Zellen vermutet (Redhead et al., 1992; Harrop et al., 2001; Warton et al., 2002;

Diskussion 100
Littler et al., 2003). Bei diesem Mechanismus wird unter oxidierenden Bedingungen die
Ausbildung gemischter Disulfide zwischen CLICs und GSH bzw. intramolekularer
Disulfid-Brücken begünstigt. Dies kann zum Einbau von im Cytosol befindlichen CLIC-
Monomeren in die Membran oder zur Ausbildung kanalkompetenter Tetramere führen.
4.2.1 Saurer Vakuoleninhalt und Redoxregulation des Malatkanals Im Falle der luminalen Redoxregulation des Malatkanals am Tonoplasten von M.
crystallinum kann man spekulieren, dass Ered des natürlichen Vakuoleninhalts von der
Konzentration der Äpfelsäure abhängen könnte. Erstens wurde ein aktiver Malatkanal nur
in malatspeichernden Vakuolen registriert. Zweitens zeigte Ered des nächtlichen Presssaftes
aus HL-gestressten CAM-Pflanzen eine gute Übereinstimmung mit dem
Mittelpunktpotential des Malat-OAA-Paars bei pH 4,0. Ein Zusammenhang zwischen der
möglichen malatbedingten positiven Verschiebung des Ered im Vakuolenlumen und dem
Redoxzustand des Malatkanals im Tonoplasten von M. crystallinum folgt aus einem
Vergleich der Mittelpunktpotentiale entsprechender Redoxpaare: das Mittelpunktpotential
des Malat-OAA-Paars beträgt -166 mV und liegt positiver als das Mittelpunktpotential des
eventuellen Cystein-Cystin-Paars des Kanalproteins von - 340 mV (Rauen, 1964). Daher
ist es nachvollziehbar, dass mit dem Malat-OAA-Redoxsystem eine Oxidation von SH-
Gruppen des Kanals thermodynamisch möglich ist. Man kann sich aber auch eine
Redoxregulation des Malatkanals durch eine Kette gekoppelter Redoxreaktionen
vorstellen, u. a. mit der Beteiligung des GSH-GSSG-Paars, das z.B. im ER-Lumen
tierischer Zellen Ered bestimmt (Hwang et al., 1992).
Hinsichtlich der Säureakkumulation in Vakuolen kann abgeleitet werden, dass alleine eine
Erniedrigung des vakuolären pH-Werts infolge der CAM-bedingten Ansäuerung zu einer
Positivierung des luminalen Ered führen könnte. In Experimenten mit einer direkten
Beladung des Cytoplasmas von Chara-Zellen mit schwacher Säure, die einen Abfall des
cytosolischen pH-Wertes um 0,5 Einheiten verursachte, konnte eine Positivierung des Ered
um 20 mV gemessen werden (Thiel, 1994). Die nächtliche Vakuolenansäuerung bei CAM-
Pflanzen von M. crystallinum beträgt mehr als zwei pH-Einheiten in saurem pH-Bereich.
Für ein System mit einer niedrigen Pufferkapazität wie den Vakuoleninhalt (Roth et al.,
1983) kann man daher erwarten, dass es durch die Ansäuerung des Lumens tatsächlich zu
einer starken Positivierung des Ered in der Vakuole kommt.

Diskussion 101
4.2.2 Regulation des Malatkanals in C3-Pflanzen Die Aktivität des Malatkanals war in leicht sauren und neutralen Vakuolen, die für C3-
Pflanzen charakteristisch sind, sehr gering (siehe Kapitel 3.7.1), obwohl die Kanaldichte
im Tonoplasten dieser Pflanzengruppe keinen Unterschied gegenüber den sauren Vakuolen
aus CAM-Pflanzen aufwies (siehe Kapitel 3.7.3). Dies bedeutet, dass die Expression des
Malatkanals in M. crystallinum unabhängig von der CAM-Induktion ist. Auf diesen
Sachverhalt weisen auch Untersuchungen von Pantoja und Smith (2002) an K.
daigremontiana, bei denen gleiche Stromdichten im Tonoplasten aus jungen Blättern ohne
nächtliche Ansäuerung und aus voll entwickelten Blättern mit CAM-bedingter
Malatakkumulation registriert wurden.
In den wenigen sauren Vakuolen, die in Blättern von M. crystallinum im C3-Zustand zu
finden waren, konnte ähnlich wie in sauren Vakuolen aus CAM-Pflanzen eine hohe
Malatkanalaktivität registriert werden. Dies unterstreicht wiederum, dass die hohe
Malatkanalaktivität am Tonoplasten unabhängig vom Stoffwechselmodus der jeweiligen
Pflanze (C3 oder CAM) mit der Akkumulation von organischen Säuren und der damit
verbundenen Positivierung des luminalen Ered korreliert. Man kann daher vermuten, dass
das Kanalabschalten in leicht sauren und neutralen Vakuolen der C3-Pflanzen ähnlich wie
in neutralen Vakuolen salzgestresster Pflanzen durch ein reduzierendes Ered des Lumens
zustande kam.
4.2.3 Regulation des Malatkanals beim Salzstress-CAM In M. crystallinum beim Salzstress-CAM, wo zwei Vakuolentypen, eine neutrale,
salzakkumulierende Vakuole und eine saure, malatakkumulierende Vakuole in einer Zelle
koexistieren, sind Kontrollmechanismen des tonoplastidären Malattransports nicht-
cytosolischen Ursprungs erforderlich, um die Malatkanalaktivität in beiden Vakuolentypen
differentiell zu regulieren. In dieser Situation übernimmt vakuoläres Ered die Rolle des
luminalen Regulationsfaktors des Malatkanals. Der redoxsensitive Malatkanal wird in
neutralen Vakuolen mit negativerem Ered abgeschaltet und in sauren Vakuolen mit
positiverem Ered hingegen aktiviert (siehe Kapitel 3.8). Andererseits werden tonoplastidäre
Transportsysteme u. a. auch den Malatkanal auch direkt, unabhängig von luminalem Ered
von NaCl bzw. Äpfelsäure beeinflusst, die in hohen Konzentrationen in Vakuolen
vorliegen. Viele Aspekte, die aus der Akkumulation dieser Substanzen resultieren,
sprechen daher gegen eine gemeinsame Akkumulation von NaCl und Äpfelsäure innerhalb
eines Vakuolenkompartiments:

Diskussion 102
Der Malatkanal unterliegt einer Hemmung durch eine hohe Konzentration von
vakuolärem Chlorid (siehe Kapitel 3.7.2.3). Deswegen kann kein Malatinflux
durch den Malatkanal in NaCl-speichernde Vakuolen stattfinden.
Der Chlorid-Influx in die Vakuole wird vermutlich durch einen passiven
Kanalmechanismus vermittelt und ist nicht elektroneutral (Kaestner und Sze,
1987; Pope und Leigh, 1987; Plant et al., 1994; Barbier-Brygoo et al., 2000,
Wissing und Smith, 2000). Aus diesen Gründen kann man von einer
Konkurrenz um das elektrische Potential zwischen Malat- und Chlorid-
transportierenden Systemen ausgehen, sollten sie in ein und derselben Membran
vorliegen.
Man geht davon aus, dass der Rücktransport des Malats ins Cytosol über eine
passive Diffusion der ungeladenen Äpfelsäure durch die lipophile
Tonoplastenphase abläuft (Lüttge und Smith, 1984; Kliemchen et al., 1993).
Folglich ist eine Protonierung des Malats in der Vakuole für diurnale
Malatfluktuationen von großer Bedeutung. Beim elektroneutralen Na+-H+-
Antiport wird der Protonengradient über den Tonoplasten erniedrigt, der bei
einer Protonierung des gespeicherten Malats benötigt wird (Spickett et al.,
1993; Barkla et al., 1995).
Der Chloridinflux wird durch einen sauren luminalen pH-Wert vermindert
(Wissing und Smith, 2000) und kann daher nicht in säurespeichernden
Vakuolen stattfinden.
Diurnale Malatfluktuationen beim CAM erfordern eine Regulation der
Protonenpumpenaktivität im Tag-Nacht-Rhythmus. Ihre Depression während
der Lichtphase kann im Falle einer gemeinsamen Akkumulation von NaCl und
Malat in einer Vakuole zu einem Austritt toxischen Natriums ins Cytoplasma
führen.
4.3 Tonoplasten unterschiedlicher Dichte und Vakuolen mit Malat- oder
Salzakkumulation
Die Ergebnisse immunologischer Untersuchungen von Saccharosedichtefraktionen
salzgestresster Pflanzen von M. crystallinum führten zum Schluss, dass bei schweren
Dichtegradientenfraktionen von 44 / 49 % (w/w) Saccharose tatsächlich eine zweite
Population von Tonoplastenvesikeln vorliegt, zusätzlich zur bereits beschriebenen
Tonoplastenfraktion bei ca. 25 % (w/w) Saccharose.

Diskussion 103
Hinweise auf die Tonoplasten unterschiedlicher Dichte wurden bereits für andere
Pflanzenspezies und Gewebetypen berichtet und zwar im Zusammenhang mit der Existenz
unterschiedlicher Vakuolentypen in einer Zelle. In Kotyledonen von Pisum sativum
wurden zwei Tonoplastenpopulationen, welche die Membranen der lytischen Vakuole und
der PSVs repräsentieren, in leichten Gradientenfraktionen von 20 / 26 % (w/w) Saccharose
bzw. in schweren Fraktionen von 36 / 45 % (w/w) Saccharose gefunden (Hoh et al., 1995).
Bereits in früheren Untersuchungen von Emig (1999) wurden kontinuierliche
Saccharosedichtegradienten von mikrosomalen Membranfraktionen aus salzgestressten
Pflanzen von M. crystallinum unter dem Aspekt der Verteilung der tonoplastentypischen
Proteine V-ATPase und V-PPase analysiert. Die Verteilungsmuster bei diesen
Untersuchungen zeigten neben deutlichen Markierungen von V-ATPase-UE im Bereich
von 25% (w/w) Saccharose kombiniert mit einer hohen ATP-Hydrolyse-Aktivität, auch
einen zweiten Bereich bei höheren Saccharosedichten. Dieser Sachverhalt wurde zunächst
auf einen Lokalisierung des Enzyms in Endomembranvesikeln oder im Plasmalemma
zurückgeführt, obwohl die hohe Enzymaktivität der V-ATPase in diesen Membranen nicht
überzeugend zu erklären war. Auch eine relativ hohe V-PPase-Aktivität wurde in den
Fraktionen mit hoher Dichte von Emig (1999) nachgewiesen. In der vorliegenden Arbeit
konnten zusätzlich zu den Antiseren gegen die V-ATPase und V-PPase zwei weitere
Antiseren erfolgreich eingesetzt werden, die gegen den vakuolären Na+-H+-Antiporter und
das γ-TIP gerichtet waren. Die Immunmarkierungsversuche haben eindeutig ein
Markierungsmaximum dieser tonoplasttypischen Proteine bei hohen Dichten von 44 / 49 %
(w/w) Saccharose gezeigt, was gemeinsam mit den Ergebnissen der Immunmarkierungen
der V-ATPase und der V-PPase sowie den hohen Aktivitäten der beiden Protonenpumpen
in diesem Dichtebereich als Nachweis für die Existenz einer zweiten Tonoplastenfraktion
gewertet wird (siehe Kapitel 3.9 und Tab. 4.2).
Aus den Ergebnissen der vergleichenden immunologischen Markierung des Na+-H+-
Antiporters in HL- und salzgestressten Pflanzen, wurde geschlossen, dass sich die
Tonoplasten der säurespeichernden und der salzspeichernden Vakuolen von M.
crystallinum in zwei hier angesprochenen Dichtebereichen anreichern. Während sich die
Membranen der säurespeichernden Vakuolen in dem Dichtebereich von 25 / 34 % (w/w)
Saccharose anreichern, fraktionieren die Membranen der salzspeichernden Vakuolen, die
ähnlich wie partiell getrennte neutrale, salzspeichernde Vakuolen durch eine deutlich
höhere Menge des Na+-H+-Antiporter-Proteins charakterisiert sind, bei einer höheren
Dichte von 44 / 49 % (w/w) Saccharose. In Membranpräparaten HL-gestresster Pflanzen,

Diskussion 104
konnte zwar auch bei hohen Dichten eine Immunmarkierung des Na+-H+-Antiporter-
Proteins beobachtet werden, aber diese Markierungen sowie auch die Markierungen des �-
TIP, der V-ATPase und V-PPase und ihre Hydrolyseaktivitäten waren im Vergleich zu
salzgestressten Pflanzen nur schwach ausgeprägt, was mit dem niedrigen Anteil neutraler
Vakuolen in HL-gestressten Pflanzen korrelierte.
Die Zuordnung der leichten und schweren Tonoplasten zu Säure- bzw. NaCl-speichernden
Vakuolen kann weitere Besonderheiten in der Proteinausrüstung dieser Vakuolentypen
aufdecken (siehe Tab. 4.2 und nachfolgende Diskussion).
Tabelle 4.2: Vergleich der Proteinmenge der getesteten tonoplastidären Proteine
sowie der Aktivitäten der V-PPase und V-ATPase in leichten und schweren
Dichtefraktionen, isoliert aus HL-gestressten und salzbehandelten Pflanzen.
Symbole kennzeichnen Gleichheit (=) und relative quantitative Unterschiede (>, >>,
<) zwischen den Saccharosedichten innerhalb einer Versuchsgruppe sowie
zwischen den salzgestressten Pflanzen gegenüber den HL-gestressten Pflanzen (n.b.
- nicht bestimmt; vor. - vorhanden).
HL-gestresste
CAM-Pflanzen
Salzgestresste
CAM-Pflanzen
Tonoplasten-
fraktionen,
% (w/w)
Saccharose
leicht
(20 / 34 %)
schwer
(44 / 49 %)
leicht
(25 / 34 %)
schwer
(44 / 49 %)
Na+-H+-Antiporter = = > >>
VM23 = = < >>
UE der V-ATPase = = = >>
V-ATP-Hydrolyse = = > >>
V-PPase n.b. n.b. vor. >> (Doppelbande)
PPi-Hydrolyse = = = >>
Tonoplastenaquaporin In salzgestressten Pflanzen war die VM23-Markierungsintensität und somit die Menge
dieses Proteins in den Tonolastenfraktionen mit hoher Dichte deutlich größer als in der
Tonoplastenvesikelpopulation bei ca. 25 % (w/w) Saccharose. Diese Beobachtung lässt

Diskussion 105
vermuten, dass sich möglicherweise die Tonoplasten der säurespeichernden und
salzspeichernden Vakuolen in ihrer Wasserleitfähigkeit unterscheiden. Nach
Immunmarkierungsversuchen von Fleurat-Lessard et al. (1997) zeigten die in ihrer
Wasserleitfähigkeit unterschiedliche Tannin- und Wasservakuolen der Pulvini-Zellen von
Mimosa pudica auch deutliche Unterschiede in der Dichte des VM23-Proteins.
Die unterschiedlichen VM23-Proteingehalte in den Membranen der säurespeichernden und
salzspeichernden Vakuolen könnten andererseits durch eine differentielle Sortierung der
Membranproteine, darunter der Aquaporintypen zustande kommen. In früheren
Untersuchungen wurde gezeigt, dass den spezifischen vakuolären Kompartimenten mit
lytischer, autofager oder proteinspeichernder Funktion unterschiedliche TIP-Isoformen,
nämlich α-, γ- δ-TIPs und DIP (dark-induced protein) zuzuordnen sind (Hoh et al., 1995;
Paris et al., 1996; Jauh et al., 1998; Neuhaus und Rogers, 1998).
Die im Rahmen der vorliegenden Arbeit durchgeführten Versuche mit einem anderen
Aquaporintyp α-TIP (Jauh et al., 1999) schlugen fehl, da nur eine sehr schwache oder gar
keine Markierung im passenden Molmassenbereich von 25 kDa zu beobachten war (Daten
nicht gezeigt). Man kann hier vermuten, dass der für die Samengewebe typischen α-TIP
entweder nicht spezifisch für Mesephyll war oder in einem olygomerisierten Zustand
vorlag und daher keine typische Bande aufwies.
Ein Molmassenvergleich des von M. crystallinum VM23-markierten Polypeptids von 23
kDa mit bekannten Proteinen aus der Aquaporinfamilie von M. crystallinum lieferte den
Hinweis, dass es sich um MIP-B handelt (Barkla et al., 1999; Kirch et al., 2000). Diese
Autoren berichteten über eine komplexe Verteilung von MIP-B in Dichtegradienten: die
von MIP-B markierten Banden waren sowohl in der tonoplastentypischen Fraktion mit
niedrigen Dichten, als auch in den dichtesten Fraktionen von 45 % (w/w) Saccharose zu
finden, wie es auch in der vorliegenden Arbeit mit der VM23-Markierung der Fall war.
Darüber hinaus zeigten die immunocytologischen Untersuchungen von Kirch et al. (2000)
eine wesentlich stärkere MIP-B-Markierung in Zellen, die Leitbündel umringen. Den
vorliegenden mikroskopischen Untersuchungen folgend haben gerade diese Zellen große,
neutrale, salzakkumulierende Vakuolen, und man kann vermuten, dass ihre Tonoplasten in
schweren Dichtegradienten mit der stärksten VM23-Markierung zu fraktionieren waren.
V-ATPase beim HL- und Salzstress-CAM Interessanterweise zeigten die bei einer Dichte von 43 / 49 % (w/w) Saccharose
angereicherten Membranvesikel, die nach der hier aufgestellten Hypothese die Tonoplasten

Diskussion 106
salzspeichernder Vakuolen repräsentieren, im Vergleich zu der Tonoplastenfraktion von
25 % (w/w) Saccharose eine sehr hohe ATP-Hydrolyse-Aktivität der V-ATPase (siehe
Kapitel 3.9.2.3 und Tab. 4.2), was auch in den Untersuchungen von Emig (1999) in
ähnlicher Weise beobachtet wurde. Diese hohe V-ATPase-Aktivität entsprach auch einer
Anreicherung aller V-ATPase-UE in diesem hohen Dichtebereich (siehe Kapitel 3.9.2.2).
V-PPase beim Salzstress-CAM Frühere Studien haben gezeigt, dass die tonoplastidäre V-PPase nur in jüngeren Pflanzen
von M. crystallinum aktiv ist und beim Übergang zum CAM verschwindet (Bremberger et
al., 1988; Bremberger und Lüttge, 1992). Diese Untersuchungen wurden jedoch mit
Tonoplastenfraktionen durchgeführt, die auf einem 25 % (w/w) Saccharosekissen
angereichert wurden. Die vorliegenden Ergebnisse stimmen insoweit mit dem bisherigen
Bild überein, als nur eine geringe Rate der PPi-Hydrolyse in Tonoplastenfraktionen
niedriger Dichte in salzbehandelten CAM-Pflanzen, sowie in allen Fraktionen aus HL-
gestressten CAM-Pflanzen gemessen werden konnte (siehe Kapitel 3.9.2.5).
Die hohe V-PPase-Aktivität in Membranpräparaten hoher Dichte salzgestresster CAM-
Pflanzen von M. crystallinum, die im Rahmen der vorliegenden Untersuchungen
festgestellt wurde, ist ein unerwartetes Ergebnis. Interessanterweise findet man jedoch in
den Untersuchungen von Emig (1999) an Membranpräparaten von salzbehandelten
Pflanzen von M. crystallinum ein ähnliches Phänomen der V-PPase-Dichtefraktionierung.
Die hohe V-PPase Aktivität und Proteinmenge bei Dichten von 41 / 43 % (w/w)
Saccharose wurde in der erwähnten Arbeit durch die vorübergehende Lokalisierung der V-
PPase in Endomembranvesikeln während des Membranflusses auf dem Weg zum
Tonoplasten oder am Plasmalemma erklärt. Nach den jetzt vorliegenden Ergebnissen und
den Immunmarkierungen des Na+-H+-Antiporters und VM23 in diesen Fraktionen muss
man jedoch davon ausgehen, dass die bei hohen Saccharosedichten zu fraktionierende V-
PPase in der hier angesprochenen zweiten Tonoplastenfraktion vorliegt, die vermutlich
ihren Ursprung in den Membranen der neutralen Vakuolen hat. Obwohl spekulativ,
erscheint es sinnvoll, über die Bedeutung der hohen Aktivität der V-PPase in schweren
Tonoplasten salzgestresster CAM-Pflanzen bezüglich des ionischen Stresses
nachzudenken.
Über den Einfluss von NaCl-Stress auf die V-PPase liegen zur Zeit in der Literatur
kontroverse Angaben vor. In Studien von Nakamura et al. (1992) nahm die Aktivität der
V-PPase in Wurzeln von Vigna mungo während einer kurzzeitigen Salzbehandlung von

Diskussion 107
wenigen Stunden und bei einer Erhöhung der cytosolischen Na+-Konzentration ab. Wang
et al. (2001) berichteten über einen Rückgang der PPi-Hydrolyse im Mesophyll der
halophytischen Pflanze Suaeda salsa bei einer Behandlung mit 400 mM NaCl, obwohl ein
moderater Salzstress mit 100 mM NaCl die V-PPase stimulierte. In Untersuchungen von
Membranen aus salzadaptierten Zellkulturen von Daucus carota durch Colombo und
Cerana (1993) und von Acer pseudoplatanus durch Zingarelli et al. (1994) konnte
hingegen ein Anstieg der V-PPase-Aktivität als Antwort auf eine Salzadaptation
nachgewiesen werden. Die erwähnten Studien haben eine interessante Gemeinsamkeit. In
den Experimenten mit Tonoplastenfraktionen, die bei geringeren Saccharosedichten
angereichert wurden, wurde eine NaCl-Inhibierung der V-PPase festgestellt (Nakamura et
al., 1992; Wang et al., 2001); hingegen wurde eine Stimulierung der PPi-Hydrolyse unter
Salzstress in Studien mit mikrosomalen Fraktionen beobachtet, d.h. mit gemischten
Zellmembranfraktionen, die u. a. auch die schweren Membranenfraktionen enthielten
(Colombo und Cerana, 1993; Zingarelli et al., 1994). Der Einsatz von Membranmaterial
mit unterschiedlicher Qualität könnte möglicherweise die widersprüchlichen Ergebnisse
der früheren Untersuchungen zur Bedeutung der V-PPase in Pflanzen unter Salzstress
erklären. Studien von Gaxiola et al. (1999, 2001) unterstreichen auch, dass die V-PPase in
der Salzsequestrierung in der Vakuole eine entscheidende Rolle spielt. Diese Autoren
berichteten, dass die Salztoleranz salzsensitiver Hefezellen durch die Überexpression von
Na+- und Cl--Transporterproteinen nur in einer Kombination mit der Überexpression der
V-PPase erreicht werden konnte. In transgenen Pflanzen von Arabidopsis thaliana führte
eine Überexpression der V-PPase auch ohne der Überexpression der Na+- und Cl--
Transporterproteinen bereits zu einer erheblichen Salz- und Trockenheittoleranz (Gaxiola
et al., 2001). Die hohe V-PPase-Aktivität zusammen mit dem Vorkommen des Na+-H+-
Antiporters in schweren Tonoplasten aus salzgestressten CAM-Pflanzen von M.
crystallinum in der vorliegenden Arbeit standen mit diesen Beobachtungen in guter
Übereinstimmung.
4.4 Diurnale Malatfluktuationen und konstante NaCl-Sequestrierung Aus der Ergebnissen der vorliegenden Arbeit und früheren Untersuchungen, sowie der hier
aufgestellten Hypothese über die Dichteunterschiede der Tonoplasten
malatakkumulierender und salzspeichernder Vakuolen ließ sich ein komplexes Bild über
die Regulation des Malat- und NaCl-Transports am Tonoplasten saurer und neutraler
Vakuolen in einer Zelle konstruieren (siehe Abb. 4.1).

Diskussion 108
Malattransport Der malatselektive Einwärtskanal ist im Tonoplasten beider Vakuolentypen vorhanden.
Dennoch kommt es infolge der luminalen Redoxregulation und Inhibierung durch
vakuoläres Chlorid zu einem Abschalten des Kanals in salzspeichernden Vakuolen;
hingegen ist der Malatkanal in säureakkumulierenden Vakuolen aktiviert.
Die Regulation des Malatkanals im Tag-Nacht-Rhythmus wird durch zwei Faktoren
bewirkt: den cytosolischen pH-Wert, der im diurnalen Rhythmus oszilliert (Hafke et al.,
2001 / 2003), und den vakuolären Redoxstatus. Dabei kann man sich das folgende
Szenario für eine malatakkumulierende Vakuole vorstellen:
Die nächtliche Aktivität der V-ATPase führt in Malat-akkumulierenden Vakuolen zu einer
Hyperpolarisierung des Tonoplasten und zu einer Ansäuerung des Vakuolenlumens mit
einer damit verbundenen, positiven Verschiebung des vakuolären Redoxpotentials. Die
negative Membranspannung einerseits und das oxidierende Vakuolenmilieu anderseits
aktivieren malatselektive Einwärtskanäle. Sie vermitteln einen elektrophoretischen
Malatinflux in die Vakuole in Form von Malat2-, das bei einem leicht alkalischen
cytosolischen pH-Wert dominiert. Obwohl Malat in der Vakuole akkumuliert wird, bleibt
der ins Cytosol gerichtete chemische Gradient für der doppelt negativ geladenen
Malationen, d.h. der einzigen Kanal-permeierenden Form des Malats, über den
Tonoplasten niedrig, da im sauren Vakuolenlumen die Malationen protoniert werden (pKa1
= 3,4; pKa2 = 5,1) und in Folge dessen als Substrate für den Malatkanal nicht mehr zu
Verfügung stehen (Hafke, 2001; Hafke et al., 2003). Ohne eine starke Erhöhung des
Membranpotentials wird eine weitere Konzentrierung der Äpfelsäure in der Vakuole
möglich. Die luminale Protonierung des Malats stellt damit eine Trapping-Reaktion dar,
die den Malatinflux gegen den Malatkonzentrationsgradienten begünstigt (Wink, 1993).
Die Akkumulierung der Äpfelsäure führt zu einer weiteren Positivierung des vakuolären
Ered. In Folge dessen werden immer mehr Malatkanäle aktiv und die
Äpfelsäurespeicherung wird intensiviert.
Am Tag sollte die oben erwähnte positive Rückkopplung zwischen der V-ATPase und dem
Malatkanal unterbrochen werden, um eine unerwünschte Äpfelsäure-Akkumulation in der
Vakuole zu unterbinden. Möglicherweise kann eine solche Herunterregulation des
vakuolären Malattransports in der Lichtphase durch das cytosolische Ered gesteuert werden.
Am Tag führt die Photosynthese bei geschlossenen Stomata in CAM-Pflanzen,
insbesondere unter Salzstress, zu einer Anreicherung reaktiver Sauerstoffspezies und einer
damit verbundenen Positivierung des Redoxpotentials im Cytosol (Miszalski et al., 1998).
Diskussion 109
Darüber hinaus wurde anhand direkter intrazellulärer Redoxmessungen im Licht und im
Dunkeln unter Zugabe von Photosyntheseinhibitoren nachgewiesen, dass allein die
photosynthetische Aktivität der Chloroplasten eine Oxidation des Cytoplasmas hervorruft
(Thiel, 1994). Außerdem kann eine für die Lichtphase des CAM charakteristische
Ansäuerung des Cytosols ebenso mit einer positiven Verschiebung des cytosolischen
Redoxpotentials einhergehen (Thiel, 1994; Hafke et al., 2001). Eine solche Positivierung
des cytosolischen Redoxpotentials kann die Aktivität der redoxempfindlichen V-ATPase
(Feng und Forgac, 1992 a / b) und somit auch die positive Interaktion zwischen den
Protonenpumpen und dem Malatkanal beeinträchtigen.
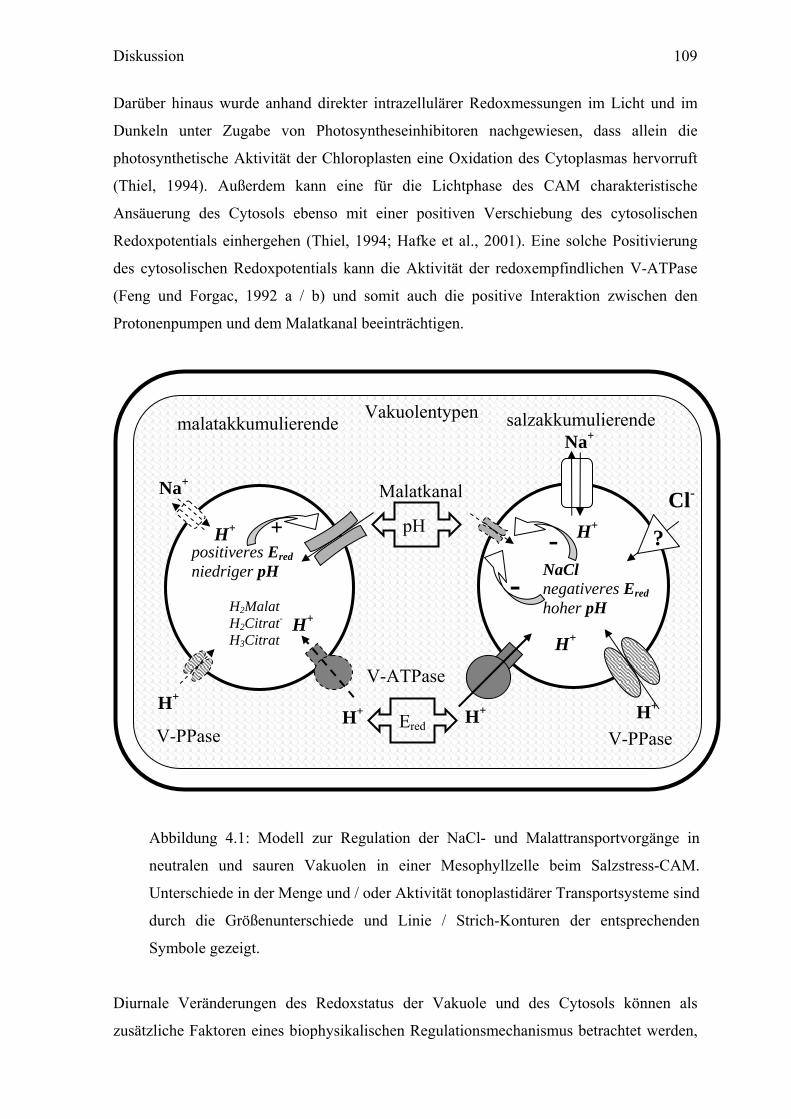
Abbildung 4.1: Modell zur Regulation der NaCl- und Malattransportvorgänge in
neutralen und sauren Vakuolen in einer Mesophyllzelle beim Salzstress-CAM.
Unterschiede in der Menge und / oder Aktivität tonoplastidärer Transportsysteme sind
durch die Größenunterschiede und Linie / Strich-Konturen der entsprechenden
Symbole gezeigt.
Diurnale Veränderungen des Redoxstatus der Vakuole und des Cytosols können als
zusätzliche Faktoren eines biophysikalischen Regulationsmechanismus betrachtet werden,
H+
Malatkanal
H+
V-PPase
Na+
H+
Ered
pH+
-- ?
Cl-
H2Malat H2Citrat- H3Citrat
H+
H+
positiveres Ered niedriger pH
H+
NaCl negativeres Ered hoher pH
V-ATPase
Na+
H+
H+
V-PPase
malatakkumulierende salzakkumulierende Vakuolentypen

Diskussion 110
der für den endogenen zirkadianen CAM-Rhythmus verantwortlich ist (Neff et al., 1998;
Lüttge, 2000). So kann man aus Studien von Dodd et al. (2002) und von Slesak et al.
(2003) ableiten, dass die nächtliche Äpfelsäureakkumulation von Faktoren abhängig sein
kann, die wie das cytosolische Redoxpotential an Lichtreaktionen gekoppelt sind. Die
Autoren berichteten nämlich über eine Hemmung der CAM-typischen Oszillationen von
C4-Säuren nach Einsetzen von Dauerlicht oder nach einer Inhibierung der
Sauerstoffevolution.
NaCl-Transport Die Präsenz des Na+-H+-Antiporters im Tonoplasten neutraler Vakuolen in hoher
Abundanz ist für ihre salzakkumulierende Funktion entscheidend. Demnach sollte man
eine Sortierung der Transportproteine in unterschiedlichen Vakuolentypen von M.
crystallinum postulieren, wie es für PSVs und lytische Vakuolen bereits nachgewiesen
wurde (Nakamura und Matsuoka, 1993; Neuhaus und Rogers, 1998). Neben der
Regulation über die Menge des in die Membran eingebauten Na+-H+-Antiporters könnte
auch eine Regulation der Na+-Transportaktivität erfolgen, wie es in Untersuchungen von
Natriumtransportraten in transgenen Pflanzen von A. thaliana nachgewiesen wurde
(Yamaguchi et al., 2003). Die Autoren spekulieren über Änderungen von Na+-
Transportraten infolge von Konformationsänderungen des zum Vakuolenlumen
exponierten C-Terminus des Na+-H+-Antiporter-Proteins, die vom vakuolären pH-Wert
oder von einer Glykosilierung verursacht werden könnten.
Unter der Annahme, dass die Membranfraktion bei hohen Saccharosedichten tatsächlich
die Tonoplasten salzspeichernder Vakuolen repräsentiert, zeigen salzspeichernde Vakuolen
im Vergleich zu malatakkumulierenden Vakuolen höhere Aktivitäten der V-ATPase und
der V-PPase. Daraus lässt sich folgern, dass die NaCl-Speicherung möglicherweise im
Vergleich zur Malatakkumulation eine verstärkte Energetisierung des Vakuolentransportes
erfordert. Die Pumpenaktivität in salzspeichernden Vakuolen sollte im Tagesgang konstant
sein, um der Diffusion von Na+ und Cl- aus der Vakuole entgegen zu wirken und somit
eine NaCl-Vergiftung des Cytosols zu verhindern, während in Malat-speichernden
Vakuolen diurnale Änderungen der Pumpenaktivität auftreten, die durch das cytosolischen
Ered kontrolliert werden (siehe oben). Gegenüber der V-ATPase zeichnet sich die V-PPase
durch eine niedrige Empfindlichkeit gegenüber Oxidation aus (Hoffman-Thoma und
Willenbrink, 1993). Tagsüber könnte daher eine in salzspeichernden Vakuolen aktive V-
PPase die notwendige Energetisierung des Tonoplasten für die NaCl-Akkumulation

Diskussion 111
aufrechterhalten. Die V-PPase in der Membran salzspeichernder Vakuolen stellt somit eine
sinnvolle Ergänzung zu der in der Lichtphase gedrosselten V-ATPase dar.

Zusammenfassung 112
5 Zusammenfassung In photosynthetisch aktivem Blattgewebe der halophytischen und C3-CAM-intermediären
Pflanze Mesembryanthemum crystallinum L. konnten erstmals Vakuolen unterschiedlicher
Charakteristika, vor allem bezüglich der Azidität (saure und neutrale Vakuolen),
identifiziert werden. Der Vergleich der Anteile beider Vakuolentypen in Pflanzen, in denen
der CAM-Photosynthesemodus durch Salz- oder Hochlichtstress induziert wurde, zeigte,
dass die CAM-bedingte Malatakkumulation in sauren Vakuolen und die NaCl-Speicherung
unter Bedingungen hoher Salinität in neutralen Vakuolen stattfindet. Auch die Analyse der
vakuolären Inhaltsstoffe wies auf separate Vakuolen für Speicherung von organischen
Säuren und Haliten hin.
Die malatakkumulierenden und salzspeichernden Vakuolen in einer Zelle zeigen im Laufe
des Tages ein dynamisches Verhalten, das durch antiparallele Volumenänderungen dieser
Vakuolentypen charakterisiert ist. Während der Oszillation des Malatgehalts sorgen diese
Volumenfluktuationen für eine osmotische Balance zwischen den beiden Vakuolentypen.
In Patch-clamp-Studien konnte in der Membran beider Vakuolentypen aus CAM-Pflanzen,
wie auch in Vakuolen aus C3-Planzen ein malatselektiver, einwärts gleichrichtender
Anionenkanal detektiert werden, der bezüglich seiner Eigenschaften dem Malatkanal der
CAM-Pflanze Kalanchoë daigremontiana ähnelt. Es wurde nachgewiesen, dass die
Aktivität des Malatkanals durch den Redoxzustand des Vakuolenlumens reguliert wird, mit
dem Ergebnis, dass der Kanal nur in sauren malatakkumulierenden Vakuolen aktiv ist.
Eine partiell angereichte Fraktion neutraler Vakuolen aus salzgestressten CAM-Pflanzen
zeigte eine höhere Abundanz des Na+-H+-Antiporterproteins im Vergleich zu der Fraktion
saurer Vakuolen. Dies bestätigte die Rolle neutraler Vakuolen in der NaCl-Akkumulation.
Weiterhin wurden im Blattgewebe zwei Populationen von Tonoplastenvesikeln
unterschiedlicher isopycnischer Dichte nachgewiesen. Anhand vergleichender
immunologischer Studien der tonoplastidären Markerproteine Na+-H+-Antiporter, γ-TIP,
V-ATPase und V-PPase, sowie Aktivitätsmessungen der tonoplastidären Protonenpumpen
in mit Salz- und Hochlicht gestressten CAM-Pflanzen, wurden Hinweise dafür erhalten,
dass die Tonoplastenpopulation mit hoher Dichte von salzspeichernden Vakuolen stammt.
Die Ergebnisse weisen auf eine funktionelle Kopplung der primär aktiven (V-ATPase und
V-PPase) und sekundär aktiven (Malatkanal und Na+-H+-Antiporter) Transportsysteme am
Tonoplasten hin. Dabei spielen der Redoxpotential und Azidität des vakuolären Lumens
und des Cytosols eine wichtige Rolle bei Regulation und Koordination tonoplastidärer
Transportsysteme.

Literaturverzeichnis 113
6 Literaturverzeichnis Adams, P., Nelson, D.E., Yamada, S., Chamara, W., Jensen, R.G., Bohnert, H.J., Griffith, H.
(1998): Growth and development of Mesembryanthemum crystallinum (Aizoaceae).
New Phytol. 138, 171-190.
Adams, P., Thomas, J.C., Vernon, D.M., Bohnert, H.J., Jensen, R.G. (1992): Distinct cellular
and organismic responses to salt stress. Plant Cell Physiol. 33, 1215-1223.
Aharon, G. S., Apse, M. P., Duan, S., Hua, X., Blumwald, E. (2003): Characterization of a
family of vacuolar Na+/H+ antiporters in Arabidopsis thaliana. Plant and Soil 253,
245-256.
Ames, B.N. (1966): Assay of inorganic phosphate, total phosphate and phosphatases. Meth.
Enzymol. 8, 115-118.
Amtmann, A., Sanders, D. (1997): A unified procedure for the correction of liquid junction
potentials in patch clamp experiments on endo- and plasma membranes. J. Exp. Botany
48, 361-364.
Apse, M.P., Aharon, G.S., Snedden, W.A., Blumwald E. (1999): Salt tolerance conferred by
overexpression of a vacuolar Na+/H+ antiporter in Arabidopsis. Science 285, 1256-
1258.
Barbier-Brygoo, H., Vinauger, M., Colcombet, J., Ephritikhine, G., Frachisse, J.M., Maurel,
C. (2000): Anion channels in higher plants: functional characterization, molecular
structure and physiological role. Biochim. Biophys. Acta 1465, 199-218.
Barkla, B.J., Blumwald, E. (1991): Identification of a 170-kDa protein associated with the
vacuolar Na+/H+ antiport of Beta vulgaris. Proc. Natl. Acad. Sci. U S A 88, 11177-
11181.
Barkla, B.J., Vera-Estrella, R., Camacho-Emiterio, J., Pantoja, O. (2002): Na+/H+ exchange in
the halophyte Mesembryanthemum crystallinum is associated with cellular sites of Na+
storage. Func. Plant Biology 29, 1017 – 1024.
Barkla, B.J., Vera-Estrella, R., Pantoja, O., Kirch, H.H., Bohnert, H.J. (1999): Aquaporin
localization - how valid are the TIP and PIP labels? Trends Plant Sci. 4, 86-88.
Barkla, B.J., Zingarelli, L., Blumwald, E., Smith, J.A.C. (1995): Tonoplast Na+/H+ antiport
activity and its energization by the vacuolar H+-ATPase in the halophytic plant
Mesembryanthemum crystallinum L. Plant Physiol. 109, 549-556.
Becker, A., Canut, H., Lüttge, U., Maeshima, M., Marigo, G., Ratajczak, R. (1995):
Purification and immunological comparison of the tonoplast H+-pyrophosphatase from

Literaturverzeichnis 114
cells of Catharanthus roseus and leaves from Mesembryanthemum crystallinum
performing C3-photosynthesis and the obligate CAM-plant Kalanchoe daigremontiana.
J. Plant Physiol.146, 88-94.
Bertl, A., Blumwald, E., Coronado, R., Eisenberg, R., Findlay, G., Gradmann, D., Hille, B.,
Kohler, K., Kolb, H.A., Mac Robbie, E., Meissner, G., Miller, C., Neher, E., Palde, P.,
Pantoja, O., Sanders, D., Schroeder, J., Slayman, C., Spanswick, R., Walker, A.,
Williams, A. (1992): Electrical measurements on endomembranes. Science 258, 873-
874.
Bertl, A., Slayman, C.L. (1990): Cation-selective channels in the vacuolar membrane of
Saccharomyces: dependence on calcium, redox state, and voltage. Proc. Natl. Acad.
Sci. U S A 87, 7824-7828.
Blumwald, E., Aharon, G.S., Apse, M.P. (2000): Sodium transport in plant cells. Biochim.
Biophys. Acta 1465, 140-151.
Blumwald, E., Poole, R.J. (1985): Na+/H+ antiport activity in isolated tonoplast vesicels from
storage tissue of Beta vulgaris. Plant Physiol. 78, 163-167.
Bohnert, H.J., Cushman, J. C. (2000): The ice plant cometh: Lessons in abiotic stress
tolerance. J. Plant Growth Regul. 19, 334 – 346.
Bremberger, C, Haschke, H.-P., Lüttge, U. (1988): Separation and purification of the
tonoplast ATPase and pyrophosphatase from plants with constitutive and inducible
crassulacean acid metabolism. Planta 175, 465-470.
Bremberger, C., Lüttge, U. (1992): Dynamics of tonoplast proton pumps and other tonoplast
proteins of Mesembryanthemum crystallinum during the induction of crassulacean acid
metabolism. Planta 188, 575-580.
Broetto, F., Lüttge, U., Ratajczak, R. (2002): Influence of light intensity and salt-treatment on
mode of photosynthesis and enzymes of the antioxidative response system of
Mesembryanthemum crystallinum. Funct. Plant Biol. 29, 13–23.
Brüggemann, L., Pottosin, I., Schönknecht, G. (1999 a): Selectivity of the fast activating
vacuolar cation channel. J. Exp. Bot. 50, 873-876.
Brüggemann, L., Pottosin, I., Schönknecht, G. (1999 b): Cytoplasmic magnesium regulates
the fast activating vacuolar cation channel. J. Exp. Bot. 50, 1547-1552.
Buser, C., Matile, P. (1977): Malic acid in vacuoles from Bryophyllum leaf cells. Z.
Pflanzenphysiol. 82, 462-466.
Buser-Suter, C., Wiemken, A., Matile, P. (1982): A malic acid permease in isolated vacuoles
of a crassulaceen acid metabolism plant. Plant Physiol. 69, 456-459.

Literaturverzeichnis 115
Carpaneto, A., Cantu, A.M., Gambale, F. (1999): Redox agents regulate ion channel activity
in vacuoles from higher plant cells. FEBS Lett. 442, 129-132.
Cerana, R., Giromini, L., Colombo, R. (1995): Malate regulated channels permeable to anions
in vacuoles of Arabidopsis thaliana. Aust. J. Plant Physiol. 22, 115-121.
Chauhan, S., Forsthoefel, N., Ran, Y., Quigley, F., Nelson, D.E., Bohnert, H.J. (2000):
Na+/myo-inositol symporters and Na+/H+-antiport in Mesembryanthemum
crystallinum. Plant J. 24, 511-522.
Cheffings, C.M., Pantoja, O., Ashcroft, F.M., Smith, J.A.C. (1997): Malate transport and
vacuolar ion channels in CAM plants. J. Exp. Bot. 48, 623-631.
Colombo, R., Cerana, R. (1993): Enhanced activity of tonoplast pyrophosphatase in NaCl-
grown cells of Daucus carota. J. Plant Physiol.142, 216-229.
Cushman, J.C. (2001): Crassulacean acid metabolism. A plastic photosynthetic adaptation to
arid environments. Plant Physiol.127, 1439-1448.
Cushman, J.C., Borland, A.M. (2002): Induction of Crassulacean acid metabolism by water
limitation. Plant Cell Environ. 25, 295-310.
Darley, C.P., Davies, J.M., Sanders, D. (1995): Chill-induced changes in the activity and
abundance of the vacuolar proton-pumping pyrophosphatase from mung bean
hypocotyls. Plant Physiol.109, 659-665.
Davies, D.D. (1986): The fine control of cytosolic pH. Physiol. Plant. 67, 702-706.
Di Sansebastiano, G.P., Paris, N., Marc-Martin, S., Neuhaus, J.M. (1998): Specific
accumulation of GFP in a non-acidic vacuolar compartment via a C-terminal
propeptide-mediated sorting pathway. Plant J.15, 449-457.
Di Sansebastiano, G.P., Paris, N., Marc-Martin, S., Neuhaus, J.M. (2001): Regeneration of a
lytic central vacuole and of neutral peripheral vacuoles can be visualized by green
fluorescent proteins targeted to either type of vacuoles. Plant Physiol.126, 78-86.
Dodd, A.N., Griffiths, H., Taybi, T., Cushman J.C., Borland A.M. (2002): Integrating diel
starch metabolism with the circadian and environmental regulation of Crassulacean
acid metabolism in Mesembryanthemum crystallinum. Planta 216, 789-797.
Dröse, S., Altendorf, K. (1997): Bafilomycins and concanamycins as inhibitors of V-ATPases
and P-ATPases. J. Exp. Biol. 200, 1-8.
Dröse, S., Bindsei, K.U., Bowman, E.J., Siebers, A., Zeeck, A., Altendorf, K. (1993):
Inhibitory effect of modified bafilomycins and concanamycins on P- and V-type
adenosinetriphosphatases. Biochemistry 32, 3902-3906.

Literaturverzeichnis 116
Emig, I. (1999): Intrazelluläre Verteilung der V-ATPase in der C3-CAM intermediären
Pflanze Mesembryanthemum crystallinum L. Dissertation. TU-Darmstadt.
Feng, W., Liu, G., Allen, P.D., Pessah, I.N. (2000): Transmembrane redox sensor of
ryanodine receptor complex. J. Biol. Chem. 275, 35902-35907.
Feng, Y., Forgac, M. (1992 a): A novel mechanism for regulation of vacuolar acidification. J.
Biol. Chem. 267, 19769-19772.
Feng, Y., Forgac, M. (1992 b): Cysteine 254 of the 73-kDa A subunit is responsible for
inhibition of the coated vesicle H+-ATPase upon modification by sulfhydryl reagents.
J. Biol. Chem. 267, 5817-5822.
Feng, Y., Forgac, M. (1994): Inhibition of vacuolar H+-ATPase by disulfide bond formation
between cysteine 254 and cysteine 532 in subunit A. J. Biol. Chem. 269, 13224-13230.
Fleurat-Lessard, P., Frangne, N., Maeshima, M., Ratajczak, R., Bonnemain, J.L., Martinoia,
E. (1997): Increased expression of vacuolar aquaporin and H+-ATPase related to motor
cell function in Mimosa pudica L. Plant Physiol. 114, 827-834.
Forgac, M. (1989): Structure and function of vacuolar class of ATP-driven proton pumps.
Physiol. Rev. 69, 765-769.
Garbarino, J., DuPont, F.M. (1989): Rapid induction of Na+ / H+ exchange activity in barley
root tonoplast. Plant Physiol. 89, 1-4.
Gaxiola, R.A., Li, J., Undurraga, S., Dang, L.M., Allen, G.J., Alper, S.L., Fink, G.R. (2001):
Drought- and salt-tolerant plants result from overexpression of the AVP1 H+-pump.
Proc. Natl. Acad. Sci. U S A 98, 11444-11449.
Gaxiola, R.A., Rao, R., Sherman, A., Grisafi, P., Alper, S.L., Fink, G.R. (1999): The
Arabidopsis thaliana proton transporters, AtNHX1 and AVP1, can function in cation
detoxification in yeast. Proc. Natl. Acad. Sci. U S A 96, 1480-1485.
Gerlach, D.(1976): Das Lichtmikroskop. Eine Einführung in Funktion, Handhabung und
Spezialverfahren für Mediziner und Biologen. Georg Thieme Verlag Stuttgart.
Hafke, J.B. (2001): Biophysikalische und physiologische Charakterisierung eines Malatkanals
aus dem Tonoplasten von Kalanchoe daigremontiana. Dissertation. TU-Darmstadt.
Hafke, J.B., Hafke, Y., Smith, J.A.C., Lüttge, U., Thiel, G. (2003): Vacuolar malate uptake is
mediated by an anion-selective inward rectifier. Plant J. 35, 116-128.
Hafke, J.B., Neff, R., Hütt, M-T., Lüttge, U., Thiel, G. (2001): Day-to-night variations of
cytoplasmic pH in a crassulacean acid metabolism plant. Protoplasma 216, 164-170.

Literaturverzeichnis 117
Hamill, O.P., Marty, A., Neher, E., Sakmann, B., Sigworth, F.J. (1981): Improved patch-
clamp techniques for high-resolution current recording from cells and cell-free
membrane patches. Pflügers Arch. 391, 85-100.
Harrop, S.J., DeMaere, M.Z., Fairlie, W.D., Reztsova, T., Valenzuela, S.M., Mazzanti, M.,
Tonini, R., Qiu, M.R., Jankova, L., Warton, K., Bauskin, A.R., Wu, W.M., Pankhurst,
S., Campbell, T.J., Breit, S.N., Curmi, P.M. (2001): Crystal structure of a soluble form
of the intracellular chloride ion channel CLIC1 (NCC27) at 1.4-Å resolution. J. Biol.
Chem. 276, 44993-50000.
Haschke, H.P., Bremberger, C., Lüttge, U. (1989): Transportproteins in plants with
crassulacean acid metabolism: immunological charaterisation of ATPase subunits. In:
Dainty, J., De Michelis, M.J., Marré, E., Rasi-Caldogno, F. (eds.) Plant membrane
transport: the current position. Elsevier Science Publishers, Amsterdam-New-York-
Oxford, 149-154.
Haugland, P. (1996): Handbook of fluorescent probes and research chemicals. Spence, M.T.Z
(ed.). Molecular Probes, Inc., Eugene.
Hedrich, R., Neher, E. (1987): Cytoplasmatic calcium regulates voltage-dependent ion
channels. Nature 329, 833-836.
Herman, E.M., Li, X., Su, R.T., Larsen, P., Hsu, H.-T., Sze, H. (1994): Vacuolar-type H+-
ATPases are associated with the endoplasmic reticulum and provacuoles of root cells.
Plant Physiol. 106, 1313-1324.
Higuchi, T., Suga, S., Tsuchiya, T., Hisada, H., Morishima, S., Okada, Y., Maeshima, M.
(1998): Molecular cloning, water channel activity and tissue specific expression of two
isoforms of radish vacuolar aquaporin. Plant Cell Physiol. 39, 905-913.
Hille, B. (1992): Ionic channels of excitable membranes. Sinauer Associates, Sunderland,
MA.
Hoffman-Thoma, G., Willenbrink, J. (1993): Differential sensitivities towards inhibitors and
SH-compounds of H+-ATPase and pyrophosphatase associated with intact vacuoles
from sweet sorghum (Sorghum bicolor [L.] Moench) stem parenchyma. J. Plant
Physiol.141, 681-689.
Hoh, B., Hinz, G., Jeong, B.K., Robinson, D.G. (1995): Protein storage vacuoles form de
novo during pea cotyledon development. J. Cell Sci. 108, 299-310.
Holwerda, B.C., Galvin, N.J., Baranski T.J., Rogers, J.C. (1990): In vitro processing of
aleurain, a barley vacuolar thiol protease. Plant Cell 2, 1091-1106.

Literaturverzeichnis 118
Holwerda, B.C., Padgett, H.S., Rogers, J.C. (1992): Proaleurain vacuolar targeting is
mediated by short contiguous peptide interactions. Plant Cell 4, 307-338.
Hurley, D., Taiz, L. (1989): Immunological localization of the vacuolar H+-ATPase in maize
root tip cells. Plant Physiol. 89, 391-395.
Hwang, C., Sinskey, A.J., Lodish, H.F. (1992): Oxidized redox state of glutathione in the
endoplasmatic reticulum. Science 257, 1496-1502.
Iwasaki, I., Arata, H., Kijima, H., Nishimura, M. (1992): Two types of ion channels involved
in the malate ion transport across the tonoplast of a crassulacean acid metobolism plant.
Plant Physiol. 98, 1494-1497.
Jauh, G.Y., Phillips, T.E., Rogers, J.C. (1999): Tonoplast intrinsic protein isoforms as markers
for vacuolar functions. Plant Cell 11, 1867-1882.
Jiang, L., Phillips, T.E., Hamm, C.A., Drozdowicz Y.M., Rea, P.A., Maeshima, M., Rogers,
S.W. (2001): The protein storage vacuole: a unique compound organelle. J. Cell Biol.
155, 991-1002.
Jiang, L., Rogers, J.C. (1998) Integral membrane protein sorting to vacuoles in plant cells:
evidence for two pathways. J. Cell Biol. 143, 1183-1199.
Jochem, P., Lüttge, U. (1987): Proton transporting enzymes at the tonoplast of leaf cells of the
CAM plant Kalanchoe daigremontiana I. The ATPase. Plant Physiol. 129, 251-268.
Kaestner, K.H., Sze, H. (1987): Potential-dependent anion transport in tonoplast vesicles from
oat roots. Plant Physiol. 83, 483-489.
Kirch, H.H., Vera-Estrella, R., Golldack, D., Quigley, F., Michalowski, C.B., Barkla, B.J.,
Bohnert, H.J. (2000): Expression of water channel proteins in Mesembryanthemum
crystallinum. Plant Physiol. 123, 111-124.
Kliemchen, A., Schomburg, M., Galla, H.J., Lüttge, U., Kluge, M. (1993): Phenotypic
changes in the fluidity of the tonoplast membrane of crassulacean-acid-metabolism
plants in response to temperature and salinity stress. Planta 189, 403-409.
Kluge, C., (2002): Der Multiproteinkomplex der vakuolären H+-ATPase höherer Pflanzen:
Klonierung, Expression und subzelluläre Lokalisation der Untereinheiten.
Dissertation. Universität Bielefeld.
Kourie, J.I. (1998): Interaction of reactive oxygen species with ion transport mechanisms. J.
Physiol. 275, 1-24.
Krisch, R. (1999): Untersuchungen zur proteolytischen Prozessierung der B-Untereinheit der
V-ATPase von Mesembryanthemum crystallinum L. Diplomarbeit, TU-Darmstadt.

Literaturverzeichnis 119
Lämmli, U.K. (1970): Cleavage of structural proteins during the assembly of the head of
bacteriophage T4. Nature 227, 680-685.
Lichter, F., Lucas, W.J., Spanswick, R.M. (1981): Effect of sulfhydryl reagents on the
biophysical properties of the plasmalemma of Chara corallina. Plant Physiol. 68, 899-
904.
Littler, D.R., Harrop, S.J., Fairlie, W.D., Brown, L.J., Pankhurst, G.J., Pankhurst, S.,
DeMaere, M.Z., Campbell, T.J., Bauskin, A.R., Tonini, R., Mazzanti, M., Breit, S.N.,
Curmi, P.M. (2003): The intracellular chloride ion channel protein CLIC1 undergoes a
redox-controlled structural transition. J. Biol. Chem. [Epub ahead of print].
Lucas, W.J., Alexander, J.M. (1980): Sulfhydryl group involvement in plasmalemma
transport of HCO3- and OH- in Chara corallina. Plant Physiol. 65, 274-280.
Lüttge, U. (1987): Carbon dioxide and water demand: crassulacean acid metabolism (CAM),
a versatile ecological adaptation exemplifying the need for intergration in
ecophysiological work. New Phytol. 106, 593-629.
Lüttge, U. (1988): Day-night changes of citric acid levels in crassulacean acid metabolism:
phenomenon and eophysiological significance. Plant, Cell and Env. 11, 445-451.
Lüttge, U. (1993): The role of crassulacean acid metabolism (CAM) in the adaptation of
plants to salinity. New Phytol. 125, 59-71.
Lüttge, U. (2000): The tonoplast functioning as the master switch for circadian regulation of
crassulacean acid metabolism. Planta 211, 761-769.
Lüttge, U., Ball, E. (1980): 2 H+:1 malate2- stoichiometry during crassulacean acid
metabolism is unaffected by lipophilic cations. Plant, Cell and Env. 3, 195-200.
Lüttge, U., Fischer-Schliebs, E., Ratajczak, R. (2001): The H+-pumping V-ATPase of higher
plants: a versatile “eco-enzyme” in response to environmental stress. Cell Biol. Mol.
Lett. 6, 356-361.
Lüttge, U., Fischer-Schliebs, E., Ratajczak, R., Kramer, D., Berndt, E., Kluge, M. (1995 a):
Functioning of the tonoplast in vacuolar C-storage and remobilization in crassulacean
acid metabolism. J. Exp. Bot. 46, 1377-1388.
Lüttge, U., Fischer-Schliebs, E., Steudle, E. (1978): Membrane potentials and salt distribution
in epidermal bladders and photosynthetic tissue of Mesembryanthemum crystallinum L.
Plant Cell Environ. 1, 121-129.
Lüttge, U., Pfeifer, T., Fischer-Schliebs, E., Ratajczak, R. (2000): The role of vacuolar
malate-transport capacity in crassulacean acid metabolism and nitrate nutrition. Higher

Literaturverzeichnis 120
malate-transport capacity in ice plant after crassulacean acid metabolism-induction and
in tobacco under nitrate nutrition. Plant Physiol. 124, 1335-1348.
Lüttge, U., Ratajczak, R. (1997): The physiology, biochemistry and molecular biology of the
plant vacuolar ATPase. In: The Plant Vacuole. Leigh, R.A., Sanders, D. (Eds.),
Advances in Botanical Research 25, 253-296.
Lüttge, U., Ratajczak, R., Rausch, T., Rockel, B. (1995 b): Stress response of tonoplast
proteins: an example for molecular ecophysiology and the search for eco-enzymes. Acta
Bot. Neerl. 44, 343-362.
Lüttge, U., Smith, J.A.C. (1984): Mechanism of passive malic-acid efflux from vacuoles of
the CAM-plant Kalanchoe daigremontiana. J. Membr. Biol. 47, 401-422.
Maeshima, M. (1992): Characterisation of major integral protein of vacuolar membrane. Plant
Physiol. 98, 1248-1254.
Maeshima, M. (2000): Vacuolar H+-pyrophosphatase. Biochim. Biophys. Acta 1465, 37-51.
Maeshima, M., Yoshida, S. (1989): Purification and properties of the vacuolar membrane
proton-translocating inorganic pyrophosphatase from mung-bean. J. Biol. Chem. 264,
20068-20073.
Marigo, G., Bouyssou, H., Laborie, D. (1988): Evidence for a malate transport into vacuoles
isolated from Catharanthus roseus cells. Bot. Acta 101, 187-191.
Marquardt-Jarczyk, G., Lüttge, U. (1990 a): V-PPase-activated ATP-dependent H+-transport
at the tonoplast of mesophyll cells of the CAM plant Kalanchoe daigremontiana. Bot.
Acta 103, 203-213.
Marquardt-Jarczyk, G., Lüttge, U. (1990 b): Anion transport at the tonoplast of mesophyll
cells of the CAM plant Kalanchoe daigremontiana. J. Plant Physiol. 136, 129-136.
Martinoia, E., Massonneau, A., Frangne, N. (2000): Transport processes of solutes across the
vacuolar membrane of higher plants. Plant Cell Physiol. 41, 1175-1186.
Martinoia, E., Rentsch, D. (1994): Malate compartmentation-responses to a complex
metabolism. Ann. Rev. Plant Physiol. Plant Mol. Biol. 45, 447-467.
Martinoia, E., Vogt, E., Rentsch, D., Amrhein, M. (1991): Functional reconstitution of the
malate carrier of barley mesophyll vacuoles in liposomes. Biochim. Biophys. Acta
1062, 271-278.
Matsuoka, K., Higuchi, T., Maeshima, M., Nakamura, K.A. (1997): Vacuolar-type H+-
ATPase in a nonvacuolar organelle is required for the sorting of soluble vacuolar
protein precursors in tobacco cells. Plant Cell 9, 533-546.

Literaturverzeichnis 121
Miedema, H., Balderas, E., Pantoja, O. (2000): Current oscillations under voltage-clamp
conditions: an interplay of series resistance and negative slope conductance. J. Membr.
Biol. 173, 31-37.
Miedema, H., Pantoja, O. (2001): Anion modulation of the slowly activating vacuolar
channel. J. Membr. Biol. 183, 137-145.
Miszalski, Z., Slesak, I., Niewiadomska, E., Baczek, R., Lüttge, U., Ratajczak, R. (1998):
Subcellular localization and stress responses of superoxide dismutase isoforms from
leaves in the C3-CAM intermediate halophyte Mesembryanthemum crystallinum L. Plant
Cell Environ. 21, 169-179.
Möllering, H. (1974): Metabolite: Substanzen im Citronensäure-Cyclus. L-Malat:
Bestimmung mit Malat-Dehydrogenase und Glutamat-Oxalacetat-Transaminase.
Methoden der enzymatischen Analyse. Bergmeyer, J.W. (Hrsg.) 2, 1636-1639.
Nakamura, K., Matsuoka, K. (1993): Protein targeting to the vacuole in plant cells. Plant
Physiol. 101, 1-5.
Nakamura, Y., Kasamo. K., Shimosato, N., Sakata, M., Ohta, E. (1992): Stimulation of the
extrusion of protons and H+-ATPase activities with the decline in pyrophosphatase
activity of the tonoplast in intact mung bean roots under high-NaCl stress and its
regulation to external levels of Ca2+ ions. Plant Cell Pysiol. 33, 139-149.
Nass, R., Rao, R. (1998): Novel localization of a Na+/H+ exchanger in a late endosomal
compartment of yeast. Implications for vacuole biogenesis. J. Biol. Chem. 273, 21054-
21060.
Neff, R., Blasius, B., Beck, F., Lüttge, U. (1998): Thermodynamics and energetics of the
tonoplast membrane operating as a hysteresis switch in an oscillatory model of
Crassulacean acid metabolism. J. Membr. Biol. 165, 37-43.
Neubert, K. (2000): Untersuchungen zu tonoplastidären Wasserkanalproteinen bei
Mesembryanthemum crystallinum L. Diplamarbeit. TU-Darmstadt
Neuhaus, J.M., Roger, J.C. (1998): Sorting of proteins to vacuoles in plant cells. Plant Mol.
Biol. 38, 127-144.
Nishida, K., Tominaga, O. (1987): Energy-dependent uptake of malate into vacuoles isolated
from CAM-plant Kalanchoe daigremontiana. J. Plant Physiol. 127, 385-393.
Oba, T., Murayama, T., Ogawa, Y. (2002): Redox states of type 1 ryanodine receptor alter
Ca2+ release channel response to modulators. Am J. Physiol. Cell Physiol. 282, 684-
692.

Literaturverzeichnis 122
Oberbeck, K., Drucker, M., Robinson, D.G. (1994): V-ATPase and pyrophosphatase in
endomembranes of maize roots. J. Exp. Bot. 45, 235-244.
Osmond, C.B. (1978): Crassulacean acid metabolism: a curiosity in context. Ann. Rev. Plant
Physiol. 29, 379-414.
Pantoja, O., Gelli, A., Blumwald, E. (1992): Characterization of vacuolar malate and K+
channels under physiological conditions. Plant Physiol. 100, 1137-1141.
Pantoja, O., Smith, J.A.C. (2002): Sensitivity of the plant vacuolar malate channel to pH, Ca2+
and anion-channel blockers. J. Membr. Biol. 186, 31-42.
Paris, N., Stanley, C.M., Jones, R.L., Rogers J.C. (1996): Plant cells contain two functionally
distinct vacuolar compartments. Cell 85, 563-572.
Pfeifer, T. (1999): Vergleichende physiologische Untersuchungen zum Malattransport am
Tonoplasten verschiedener Pflanzen und Reinigung des Malattransport-Proteins aus
Kalanchoe daigremontiana. Dissertation Fachbereich Biologie. TU Darmstadt.
Plant, P.J., Gelli, A., Blumwald, E. (1994): Vacuolar chloride regulation of an anion-selective
tonoplast channel. J. Membr. Biol. 140, 1-12.
Pope, A.J., Leigh, R.A. (1987): Some characteristics of anion transport at the tonoplast of oat
roots, determined from the effects of anions on pyrophosphate-dependent proton
transport. Planta 172, 91-100.
Popov, N., Schmitt, M., Schulzek, S., Matthies, H. (1975): Eine störungsfreie Mikromethode
zur Bestimmung des Proteingehaltes in Gewebehomogenaten. Acta Biol. Med. Ger. 34,
1441-1446.
Ratajczak, R. (2000): Structure, function and regulation of the plant vacuolar H+-translocating
ATPase. Biochim. Biophys. Acta 1465, 17-36.
Ratajczak, R., Hinz, G., Robinson, D.G. (1999): Localization of pyrophosphatase in
membranes of cauliflower inflorescence cells. Planta 208, 205-211.
Ratajczak, R., Kemna, I., Lüttge, U. (1994 a): Characteristics, partial purification and
reconstitution of the vacuolar malate transporter of the CAM plant Kalanchoe
daigremontiana Hamet et Perrier de la Bathie. Planta 195, 226-236.
Ratajczak, R., Richter, J., Lüttge, U. (1994 b): Adaptation of the tonoplast V-type H+-ATPase
of Mesembryanthemum crystallinum to salt stress, C3-CAM-transition and plant age.
Plant Cell Environ. 17, 1101-1112.
Rauen, H.M. (1964): Biochemisches Taschenbuch. Zweiter Teil. Zweite Ausgabe. Springer-
Verlag Berlin, Göttingen, Heidelberg, New York, 140-141.

Literaturverzeichnis 123
Rea, P.A., Poole, R.J. (1993): Vacuolar H+-translocating pyrophosphatase. Ann. Rev. Plant
Physiol. Plant Mol. Biol. 44, 157-180.
Redhead, C.R., Edelman, A.E., Brown, D., Landry, D.W., al-Awqati, Q. (1992): A ubiquitous
64-kDa protein is a component of a chloride channel of plasma and intracellular
membranes. Proc. Natl. Acad. Sci. U S A 89, 3716-3720.
Robinson, D.G., Haschke, H.P., Hinz, G., Hoh, B., Maeshima, M., Marty, F. (1996):
Immunological detection of tonoplast polypeptides in the plasma membrane of pea
cotyledones. Planta 198, 95-103.
Roth, Z., Chayen, N., Dickstein, S. (1983): The involvement of intracellular redox-state and
pH in the metabolic control of stimulus-responce coupling. Int. Rev. Cytol. 85, 39-61.
Sanders, D., Brownlee,C., Harper, J.F. (1999): Communicating with calcium. Plant Cell 11,
691-706.
Schulz-Lessdorf, B., Hedrich, R. (1995): Protons and calcium modulate SV-type channels in
the vacuolar-lysosomal compartment-channel interaction with calmodulin inhibitors.
Planta 197, 655-671.
Slesak, I., Karpinska, B., Surowka, E., Miszalski, Z., Karpinski, S. (2003): Redox changes in
the chloroplast and hydrogen peroxide are essential for regulation of C3-CAM
transition and photooxidative stress responses in the facultative CAM plant
Mesembryanthemum crystallinum L. Plant Cell Physiol. 44, 573-581.
Spickett, C.M., Smirnoff, N., Ratcliffe, R.G. (1993): An in vivo nuclear magnetic resonance
investigation of ion transport in maize (Zea mays) and Spartina anglica roots during
exposure to high salt concentrations. Plant Physiol.102, 629-638.
Steiger, S., Pfeifer, T., Ratajczak, R., Martinoia, E., Lüttge, U. (1997): The vacuolar malate
transporter of Kalanchoe daigremontiana: A 32 kDa polypeptide. J. Plant Physiol.
151, 137-141.
Struve, I., Lüttge, U. (1987): Characteristics of MgATP-dependent electrogenic proton
transport in tonoplast vesicles of the facultative crassulacean-acid-metabolism plant
Mesembryanthemum crystallinum L. Planta 170, 111-120.
Struve, I., Lüttge, U. (1988): Biochemical and immunological properties of solubilized
tonoplast ATPase of the facultative CAM plant Mesembryanthemum crystallinum in
the C3 and CAM states. Bot. Acta 101, 39-44.
Struve, I., Weber, A., Lüttge, U., Ball, E., Smith, J.A.C. (1985): Increased vacuolar ATPase
activity correlated with CAM induction in Mesembryanthemum crystallinum and
Kalanchoe blossfeldiana cv. Tom Thumd. J. Plant Physiol. 117, 451-468.

Literaturverzeichnis 124
Swanson, S.J., Bethke, P.C., Jones, R.L. (1998): Barley aleurone cells contain two types of
vacuoles. Characterization of lytic organelles by use of fluorescent probes. Plant Cell
10, 685-698.
Swanson, S.J., Jones, R.L. (1996): Gibberellic acid induces vacuolar acidification in barley
aleurone. Plant Cell 8, 2211-2221.
Taiz, L. (1992): The plant vacuole. J. Exp. Biol. 172, 113-122.
Tavakoli, N., Kluge, C., Golldack, D., Mimura, T., Dietz, K.J. (2001): Reversible redox
control of plant vacuolar H+-ATPase activity is related to disulfide bridge formation in
subunit E as well as subunit A. Plant J. 28, 51-59.
Taybi, T., Cushman, J.C. (1999): Signaling events leading to crassulacean acid metabolism
induction in the common ice plant. Plant Physiol. 121, 545-556.
Taybi, T., Patil, S., Chollet, R., Cushman, J.C. (2000): A minimal serine/threonine protein
kinase circadianly regulates phosphoenolpyruvate carboxylase activity in crassulacean
acid metabolism-induced leaves of the common ice plant. Plant Physiol. 4, 1471-1482.
Thiel, G. (1994): Redox-state of intact Nitella cells: dependency on intracellular pH and
photosynthesis. Protoplasma 179, 26-33.
Thiel, G., Weise, R. (1999): Auxin augments conductance of K+ inward rectifier in maize
coleoptile protoplasts. Planta 208, 38–45.
Ullrich, C.I., Köhler, K., Baier, M., Förster, B., Hartung, W. (1990): Neutral red as a redox
dye induces K+ efflux and current-voltage changes in Eremosphaera, Lemna and
guard cells. Bot. Acta 103, 214-221.
Vera-Estrella, R., Barkla, B.J., Bohner, H.J., Pantoja, O. (1999): Salt stress in
Mesembryanthemum crystallinum L. cell suspensions activates adaptive mechanisms
similar to those observed in the whole plant. Planta 207, 426-435.
Wang, B., Lüttge, U., Ratajczak, R. (2001): Effects of salt treatment and osmotic stress on V-
ATPase and V-PPase in leaves of the halophyte Suaeda salsa. J. Exp. Bot. 52, 2355-
2365.
Ward, J.M., Schroeder, J.L. (1994): Calcium-activated K+-channels and calcium-induced
calcium release by slow vacuolar ion channels in guard cell vacuoles implicated in the
control of stomata closure. Plant Cell 6, 669-683.
Warton, K., Tonini, R., Fairlie, W.D., Matthews, J.M., Valenzuela, S.M., Qiu, M.R., Wu,
W.M., Pankhurst, S., Bauskin, A.R., Harrop, S.J., Campbell, T.J., Curmi, P.M., Breit,
S.N., Mazzanti, M. (2002): Recombinant CLIC1 (NCC27) assembles in lipid bilayers
via a pH-dependent two-state process to form chloride ion channels with identical

Literaturverzeichnis 125
characteristics to those observed in Chinese hamster ovary cells expressing CLIC1. J.
Biol. Chem. 19, 26003-26011.
White, P.J., Smith, J.A.C. (1989): Proton and anion transport at the tonoplast in crassulacean-
acid-metabolism plants: specifity of the malate-influx system in Kalanchoe
daigremontiana. Planta 179, 265-274.
White, P.J., Smith, J.A.C. (1992): Malate-dependent proton transport in tonoplast vesicles
isolated from orchid leaves correlated with the expression of crassulacean acid
metabolism. J. Plant Physiol. 139, 533-538.
Wink, M. (1993): The plant vacuole: a multifunctional compartment. J. Exp. Bot. 44, 231-
246.
Winter, K., Gademann, R. (1991): Daily changes in CO2 and water vapour exchange,
chlorophyll fluorescence, and leaf water relations in the halophyte
Mesembryanthemum crystallinum during the induction of Crassulacean acid
metabolism in response of high NaCl salinity. Plant Physiol. 95, 768-776.
Winter, K., Smith, J.A.C. (eds.) (1996): Crassulacean acid metabolism. Biochemistry,
ecophysiology and evolution. Springer-Verlag Berlin.
Wissing, F., Smith, J.A.C. (2000): Vacuolar chloride transport in Mesembryanthemum
crystallinum L. measured using the fluorescent dye lucigenin. J. Membr. Biol. 177,
199-208.
Yamaguchi, T., Apse, M.P., Shi, H., Blumwald, E. (2003): Topological analysis of a plant
vacuolar Na+/H+ antiporter reveals a luminal C terminus that regulates antiporter
cation selectivity. Proc. Natl. Acad. Sci. USA 14, 100, 12510-12515.
Zhigang, A., Low, R., Rausch, T., Lüttge, U., Ratajczak, R. (1996): The 32 kDa tonoplast
polypeptide Di is associated with the V-ATPase of Mesembryanthemum crystallinum
L. in the CAM state: a proteolytically processed subunit B? FEBS Letters 389, 314-
318.
Zingarelli, L., Anzani, P., Lado, P. (1994): Enhanced K+ stimulated pyrophosphatase activity
in NaCl adapted cells of Acer pseudoplatanus. Physiol. Plant. 91, 510-516.

Eidesstattliche Erklärung
Ich erkläre hiermit an Eides statt, dass ich die vorliegende Dissertation selbständig und nur
mit den angegebenen Hilfsmitteln angefertigt und bisher noch keinen Promotionsversuch
unternommen habe.
Svetlana Epimashko
Darmstadt, den 5.05.2004

Lebenslauf Name Geburtstag Geburtsort Familienstand Staatsangehörigkeit Schulbildung 1981-1985 1985-1989 1989-1991 1990-1991 Studium 1991-1996 1996 1996-1998 08/1998-09/1998 10/1998-07/1999 seit 06/2000 seit 01/2002
Svetlana Epimashko 12.01.1974 Minsk, Weißrussland verheiratet weißrussisch Grundschule in Minsk Gesamtschule in Minsk mit den Leistungskursen Physik und Mathematik Abschluss: Oberstufenschule in Minsk mit Leistungskursen in Biologie und Chemie Zusätzliche Abitur-Kurse in Biologie und Chemie an der Weißrussischen Staatlichen Universität in Minsk Studium an der Weißrussischen Staatlichen Universität in Minsk, Fachbereich Biologie mit dem Grundstudium in Chemie, Physik und Mathematik Staatsexamen in Biologie und Chemie Diplomarbeit am Lehrstuhl für Physiologie und Biochemie der Pflanzen an der Weißrussischen Staatlichen Universität in Minsk bei Prof. Dr. V.M. Yurin mit dem Thema: „Wirkung niedriger Kalium-Konzentrationen auf die elektro-physiologischen Eigenschaften des Plasmalemmas von Nitella flexilis“ Abschluss: Diplom Wissenschaftliche Mitarbeiterin am Lehrstuhl für Physiologie und Biochemie der Pflanzen an der Weißrussischen Staatlichen Universität in Minsk mit dem Thema: „Wirkung extremer Kalium-Konzentrationen auf die Transportsysteme am Plasmalemma von Nitella flexilis“ Sprachkurs in Konstanz, Deutschland Zehnmonatiges Stipendium vom DAAD für wissenschaftliche Fortbildung, Gastwissenschaftlerin an der Technischen Universität Darmstadt Institut für Botanik, in der Arbeitsgruppe von Prof. Dr. Ulrich Lüttge, Promotionsstudium mit dem Landes Hessen Stipendium am Institut für Botanik der Technischen Universität Darmstadt Stipendiatin des Graduiertenkollegs 340 „Kommunikation in biologischen Systemen: Vom Molekül zum Organismus in seiner Umwelt.“
Recommended

















