
Contents lists available at ScienceDirect
Carbohydrate Polymers
journal homepage: www.elsevier.com/locate/carbpol
Extremophilic exopolysaccharides: A review and new perspectives onengineering strategies and applications
Jia Wanga,d, David R. Salema,b,c,⁎, Rajesh K. Sania,c,d,⁎
a Department of Chemical and Biological Engineering, South Dakota School of Mines and Technology, Rapid City, SD 57701, USAbDepartment of Materials and Metallurgical Engineering, South Dakota School of Mines and Technology, Rapid City, SD 57701, USAc Composite and Nanocomposite Advanced Manufacturing – Biomaterials Center (CNAM-Bio Center), Rapid City, SD 57701, USAd BuG ReMeDEE consortium, South Dakota School of Mines and Technology, Rapid City, SD 57701, USA
A R T I C L E I N F O
Keywords:ExtremophileExopolysaccharideExopolysaccharide propertyExopolysaccharide biosynthesisExopolysaccharide application
A B S T R A C T
Numerous microorganisms inhabiting harsh niches produce exopolysaccharides as a significant strategy tosurvive in extreme conditions. The exopolysaccharides synthesized by extremophiles possess distinctive char-acteristics due to the varied harsh environments which stimulate the microorganisms to produce these biopo-lymers. Despite many bioprocesses have been designed to yield exopolysaccharides, the production of exopo-lysaccharides by extremophiles is inefficient compared with mesophilic and neutrophilic exopolysaccharideproducers. Meanwhile, the industrial development of novel extremophilic exopolysaccharides remains con-strained due to the lack of exploration. In this review, we summarize the structure and properties of variousexopolysaccharides produced by extremophiles, and also discuss potential metabolic and genetic engineeringstrategies for enhanced yield and modified structure of extremophilic exopolysaccharides. Special focus is givento the applications of extremophilic exopolysaccharides in the areas of biomedicine, food industry, and bio-materials via nano-techniques, casting and electrospinning.
1. Introduction
In the past few decades, extremophilic microorganisms and some oftheir metabolites were reported in light of their particular biosyntheticmechanisms, functions, and properties which can permit the strains tobe habitant in extreme niches. Among all the products from ex-tremophiles, exopolysaccharides (EPSs) have led to significant interestdue to the increasing demand for natural polymers in various industrialfields. EPSs are high molecular weight carbohydrate biopolymers,composed of sugar residues, and are secreted by microorganisms intothe surrounding environment, providing certain properties and func-tions useful to the microorganisms (Nicolaus, Kambourova, & Oner,2010; Poli, Anzelmo, & Nicolaus, 2010). The EPS molecular chains havea broad range of molecular weights, and different microorganisms cansynthesize a wide variety of EPSs with a diverse range of functions, suchas intercellular signal transduction, molecular recognition, protectionagainst predation, adhesion, biofilm formation, construction of a com-fortable extracellular environment, and pathogenic processes (Morielloet al., 2003; Nicolaus et al., 1999). Some of the EPSs with valuablephysicochemical properties have already been utilized in industry. Forinstance, among all the reported EPSs, xanthan gum has been most
studied during the past several decades and applied in a variety of in-dustrial areas. In addition to xanthan gum, dextran and gellan gum arecurrently being used in the food industry (Donot, Fontana, Baccou, &Schorr-Galindo, 2012; Rehm, 2010). Bacterial polysaccharides possess agreat diversity of properties that may not be found in more traditionalpolymers of plant origin. Several EPSs have also demonstrated them-selves as useful materials without the environmental disadvantagesassociated with synthetic polymers (Chawla, Bajaj, Survase, & Singhal,2009; Freitas, Alves, & Reis, 2011; Guezennec, 2002).
Currently, it is widely accepted that extremophilic microorganismswill provide a valuable resource for exploitation in novel biotechnolo-gical processes, including synthesis of unique EPSs (Bhalla, Bansal,Kumar, Bischoff, & Sani, 2013; Nicolaus et al., 2010). The environmentsthat extremophiles inhabit are obviously more inhospitable than theenvironmental pressures inducing common mesophilic and neutrophilicmicrobes to secrete their EPSs. Extremophiles have to adapt to hostileenvironments through unique mechanisms, and the biosynthesis ofEPSs is one of their vital survival mechanisms. Extremophilic micro-organisms inhabiting different extreme environments have been re-cognized as promising producers of EPSs, and the examination of EPSproduction by extremophiles (thermophiles, halophiles, alkaliphiles,
https://doi.org/10.1016/j.carbpol.2018.10.011Received 2 August 2018; Received in revised form 20 September 2018; Accepted 4 October 2018
⁎ Corresponding authors at: Department of Chemical and Biological Engineering, South Dakota School of Mines and Technology, Rapid City, SD 57701, USA.E-mail addresses: [email protected] (D.R. Salem), [email protected] (R.K. Sani).
Carbohydrate Polymers 205 (2019) 8–26
Available online 09 October 20180144-8617/ © 2018 Elsevier Ltd. All rights reserved.
T

psychrophiles, and acidophiles) has revealed an abundance of novelproperties that may have strong potential in industrial applications(Fig. 1).
Although more and more novel extremophiles have been isolated,and their unique EPSs characterized, the research depth of ex-tremophilic EPSs is still not comparable with EPSs from mesophilic orneutrophilic microorganisms with regard to biosynthetic pathways,regulatory mechanisms, and engineering strategies. It is necessary tomake a comprehensive summarization concerning the structures andcharacteristics of the recently described extremophilic EPSs, which canprovide crucial fundamentals for further exploitation of engineeringstrategies to obtain tailor-made extremophilic EPSs with desired yieldand functions. The targeted cultivation of extremophilic bacteriathrough metabolic and genetic engineering will eventually pave theway for industrial level applications of extremophilic EPSs.
This article reviews the EPSs produced by various kinds of ex-tremophilic bacteria, including an inventory of extremophilic EPSs ofindustrial interest, as well as promising engineering strategies forhigher yield or modified molecular structure of extremophilic EPSs.Moreover, the recent advances in the actual and potential applicationsof EPSs produced by extremophilic bacteria are presented.
2. EPSs produced by different extremophile types
2.1. EPSs produced by thermophiles
Elevated temperature generally increases the rate of most chemicalreactions and improves cumulative production in a given time frame.Thus, thermophiles can be of commercial value in the synthesis ofimportant compounds, and are of growing interest to many sectors ofindustry. Although EPS production is lower than most of the meso-philes, the uncommonly short fermentation process, which is usually nomore than 24 h, makes thermophiles important contenders as com-mercially competitive EPS producers (Kambourova et al., 2009;Radchenkova et al., 2013; Yildiz et al., 2014). The thermophilic strainscan also typically minimize environmental contamination from meso-philic microbial growth, reduce operational maintenance cost, andimprove the efficiency of substrate utilization.
Marine hot springs, terrestrial hot springs, and deep sea thermalvents have been demonstrated as the primary habitats that promotethermophilic microbial organisms, and the majority of EPSs producedby thermophiles have been located in these types of environments.Several thermophilic bacteria in hot marine shallow vents or marine hotsprings have been shown to produce large amounts of EPSs (Mancaet al., 1996; Moriello et al., 2003; Nicolaus et al., 2002; Nicolaus,Moriello, Lama, Poli, & Gambacorta, 2004). These environments aretypically characterized by their high temperature, high pressure, andtoxic, high inorganic or metal concentrations. Thermophilic
microorganisms can survive in high temperatures, and their EPS pro-duction has been a proposed adaptation mechanism to enable theirsurvival in these extreme conditions.
The growth media for thermophiles, containing sugars as carbonand energy sources, have always been considered a primary target to beoptimized for maximum production of EPSs. Disaccharides such asmaltose, lactose and sucrose are the optimized carbon source for mostof thermophilic bacteria for EPS production. Besides chemical compo-sition and molecular weight, thermophilic EPSs have been character-ized mostly in terms of thermostability. The highest decompositiontemperature of 280 °C is from an EPS produced by Geobacillus tepida-mans (Table 1). The summarized data suggest that the type of sugarsubunits present in the EPS may affect their thermostability. Themodification of monomer sugars or some other residues in EPSs can beutilized to find out the active sites for certain functions (e.g., thermo-stability) of EPSs. Although a relatively unexplored area with a sparsedatabase, there is already significant evidence that EPSs from thermo-philes possess a broad range of interesting properties for industrialapplications (Nicolaus et al., 2004, 2010). The literature to date in-dicates that further screening and systematic investigation of EPSsproduced by thermophiles, in conjunction with advances in under-standing the biochemistry of microbial EPS synthesis, will result in thediscovery of novel biopolymers of commercial importance.
2.2. EPSs produced by psychrophiles
Psychrophiles can be isolated from Antarctic, Arctic, or deep-seasediment, and they predominate in marine ecosystems (Ewert &Deming, 2013; Li, Zhou, Zhang, Wang, & Zhu, 2008; Nevot, Deroncele,Montes, & Mercade, 2008; Nichols, Bowman, & Guezennec, 2005). TheEPSs from psychrophilic marine bacteria are generally carboxylatedpolysaccharides, and the carboxyl groups confer a net negative chargeand acidic properties to the EPSs at the pH of seawater (pH around 8)(Caruso et al., 2017; Casillo, Parrilli et al., 2017). The negative chargeof psychrophilic EPSs can also be attributed to the phosphate groups(Corsaro et al., 2004; Llamas et al., 2010). In the marine environment,bacterial EPSs are essential in the production of aggregates, adhesion tosurface, biofilm formation and sequestering of nutrients, and provideprotection and ecosystem stability. Due to their polyanionic property,psychrophilic EPSs can accumulate cations such as metal ions, andmetal binding offers a potential ecological role for these biopolymers.Extracellular polysaccharides strengthen the psychrophiles’ ability tocompete and survive in changing environmental conditions by alteringthe physical and biogeochemical micro-environment around the cells(Nichols, Bowman et al., 2005). The EPSs of psychrophiles in a coldmarine environment should possess the capability to protect the mi-croorganisms from not only the low temperature but also the relativelyhigh salinity (Caruso et al., 2018). Therefore, the EPSs secreted by the
Fig. 1. The EPS from different kinds of extremophiles and potential applications.
J. Wang et al. Carbohydrate Polymers 205 (2019) 8–26
9
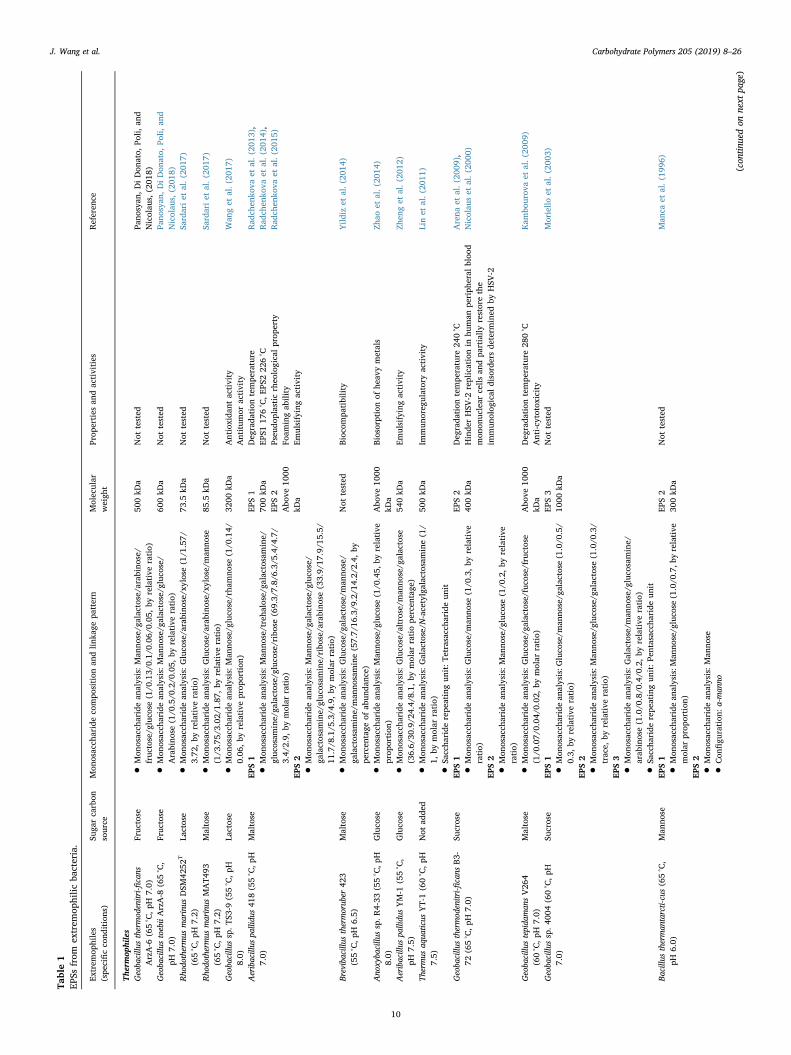
Table1
EPSs
from
extrem
ophilic
bacteria.
Extrem
ophiles
(spe
cificco
nditions)
Suga
rcarbon
source
Mon
osacch
arideco
mpo
sition
andlin
kage
pattern
Molecular
weigh
tProp
erties
andactivities
Referen
ce
Thermop
hiles
Geoba
cillu
sthermod
enitri-fi
cans
ArzA-6
(65°C,p
H7.0)
Fruc
tose
•Mon
osacch
aridean
alysis:Man
nose/g
alactose/arabino
se/
fruc
tose/g
luco
se(1/0
.13/
0.1/
0.06
/0.05,
byrelative
ratio)
500kD
aNot
tested
Pano
syan
,DiDon
ato,
Poli,
and
Nicolau
s,(201
8)Geoba
cillu
stoebiiArzA-8
(65°C,
pH7.0)
Fruc
tose
•Mon
osacch
aridean
alysis:Man
nose/g
alactose/g
luco
se/
Arabino
se(1/0
.5/0
.2/0
.05,
byrelative
ratio)
600kD
aNot
tested
Pano
syan
,DiDon
ato,
Poli,
and
Nicolau
s,(201
8)Rho
dothermus
marinus
DSM
4252
T
(65°C,p
H7.2)
Lactose
•Mon
osacch
aridean
alysis:Gluco
se/arabino
se/xylose(1/1
.57/
3.72
,byrelative
ratio)
73.5
kDa
Not
tested
Sardariet
al.(20
17)
Rho
dothermus
marinus
MAT4
93(65°C,p
H7.2)
Maltose
•Mon
osacch
aridean
alysis:Gluco
se/arabino
se/xylose/man
nose
(1/3
.75/
3.02
/1.87,
byrelative
ratio)
85.5
kDa
Not
tested
Sardariet
al.(20
17)
Geoba
cillu
ssp.T
S3-9
(55°C,p
H8.0)
Lactose
•Mon
osacch
aridean
alysis:M
anno
se/g
luco
se/rha
mno
se(1/0
.14/
0.06
,byrelative
prop
ortion
)32
00kD
aAntioxida
ntactivity
Antitum
oractivity
Wan
get
al.(20
17)
Aeribacillus
pallidu
s41
8(55°C,p
H7.0)
Maltose
EPS1
EPS1
700kD
aEP
S2
Abo
ve10
00kD
a
Deg
rada
tion
tempe
rature
EPS1
176°C,E
PS222
6°C
Pseu
doplasticrheo
logicalprop
erty
Foam
ingab
ility
Emulsifyingactivity
Rad
chen
kova
etal.(20
13),
Rad
chen
kova
etal.(20
14),
Rad
chen
kova
etal.(20
15)
•Mon
osacch
aridean
alysis:Man
nose/treha
lose/g
alactosamine/
gluc
osam
ine/ga
lactose/gluc
ose/ribo
se(69.3/
7.8/
6.3/
5.4/
4.7/
3.4/
2.9,
bymolar
ratio)
EPS2
•Mon
osacch
aridean
alysis:Man
nose/g
alactose/g
luco
se/
galactosam
ine/gluc
osam
ine/ribo
se/arabino
se(33.9/
17.9/1
5.5/
11.7/8
.1/5
.3/4
.9,by
molar
ratio)
Brevibacillus
thermorub
er42
3(55°C,p
H6.5)
Maltose
•Mon
osacch
aridean
alysis:Gluco
se/g
alactose/m
anno
se/
galactosam
ine/man
nosamine(57.7/
16.3/9
.2/1
4.2/
2.4,
bype
rcen
tage
ofab
unda
nce)
Not
tested
Bioc
ompa
tibility
Yild
izet
al.(20
14)
Ano
xyba
cillu
ssp.R
4-33
(55°C,p
H8.0)
Gluco
se•M
onosacch
aridean
alysis:M
anno
se/g
luco
se(1/0
.45,
byrelative
prop
ortion
)Abo
ve10
00kD
aBiosorptionof
heav
ymetals
Zhao
etal.(20
14)
Aeribacillus
pallidu
sYM-1
(55°C,
pH7.5)
Gluco
se•M
onosacch
aridean
alysis:Gluco
se/altrose/m
anno
se/g
alactose
(36.6/
30.9/2
4.4/
8.1,
bymolar
ratiope
rcen
tage
)54
0kD
aEm
ulsifyingactivity
Zhen
get
al.(20
12)
Thermus
aqua
ticus
YT-1(60°C,p
H7.5)
Not
adde
d•M
onosacch
aridean
alysis:Galactose/N
-acetylgalactosamine(1/
1,by
molar
ratio)
•Sacch
ariderepe
atingun
it:T
etrasaccha
ride
unit
500kD
aIm
mun
oreg
ulatoryactivity
Linet
al.(20
11)
Geoba
cillu
sthermod
enitri-fi
cans
B3-
72(65°C,p
H7.0)
Sucrose
EPS1
EPS2
400kD
aDeg
rada
tion
tempe
rature
240°C
Hinde
rHSV
-2replicationin
human
periph
eral
bloo
dmon
onuc
lear
cells
andpa
rtially
restorethe
immun
olog
ical
disordersde
term
ined
byHSV
-2
Arena
etal.(20
09),
Nicolau
set
al.(20
00)
•Mon
osacch
aridean
alysis:Gluco
se/m
anno
se(1/0
.3,b
yrelative
ratio)
EPS2
•Mon
osacch
aridean
alysis:Man
nose/g
luco
se(1/0
.2,b
yrelative
ratio)
Geoba
cillu
stepida
man
sV26
4(60°C,p
H7.0)
Maltose
•Mon
osacch
aridean
alysis:Gluco
se/g
alactose/fuc
ose/fruc
tose
(1/0
.07/
0.04
/0.02,
bymolar
ratio)
Abo
ve10
00kD
aDeg
rada
tion
tempe
rature
280°C
Anti-cytotoxicity
Kam
bourov
aet
al.(20
09)
Geoba
cillu
ssp.4
004(60°C,p
H7.0)
Sucrose
EPS1
EPS3
1000
kDa
Not
tested
Moriello
etal.(20
03)
•Mon
osacch
aridean
alysis:G
luco
se/m
anno
se/g
alactose
(1.0/0
.5/
0.3,
byrelative
ratio)
EPS2
•Mon
osacch
aridean
alysis:M
anno
se/g
luco
se/g
alactose
(1.0/0
.3/
trace,
byrelative
ratio)
EPS3
•Mon
osacch
aridean
alysis:Galactose/m
anno
se/g
luco
samine/
arab
inose(1.0/0
.8/0
.4/0
.2,by
relative
ratio)
•Sacch
ariderepe
atingun
it:P
entasaccha
ride
unit
Bacillu
stherman
tarcti-cus(65°C,
pH6.0)
Man
nose
EPS1
EPS2
300kD
aNot
tested
Man
caet
al.(19
96)
•Mon
osacch
aridean
alysis:M
anno
se/g
luco
se(1.0/0
.7,b
yrelative
molar
prop
ortion
)EP
S2
•Mon
osacch
aridean
alysis:Man
nose
•Con
figu
ration
:α-m
anno
(con
tinuedon
next
page)
J. Wang et al. Carbohydrate Polymers 205 (2019) 8–26
10
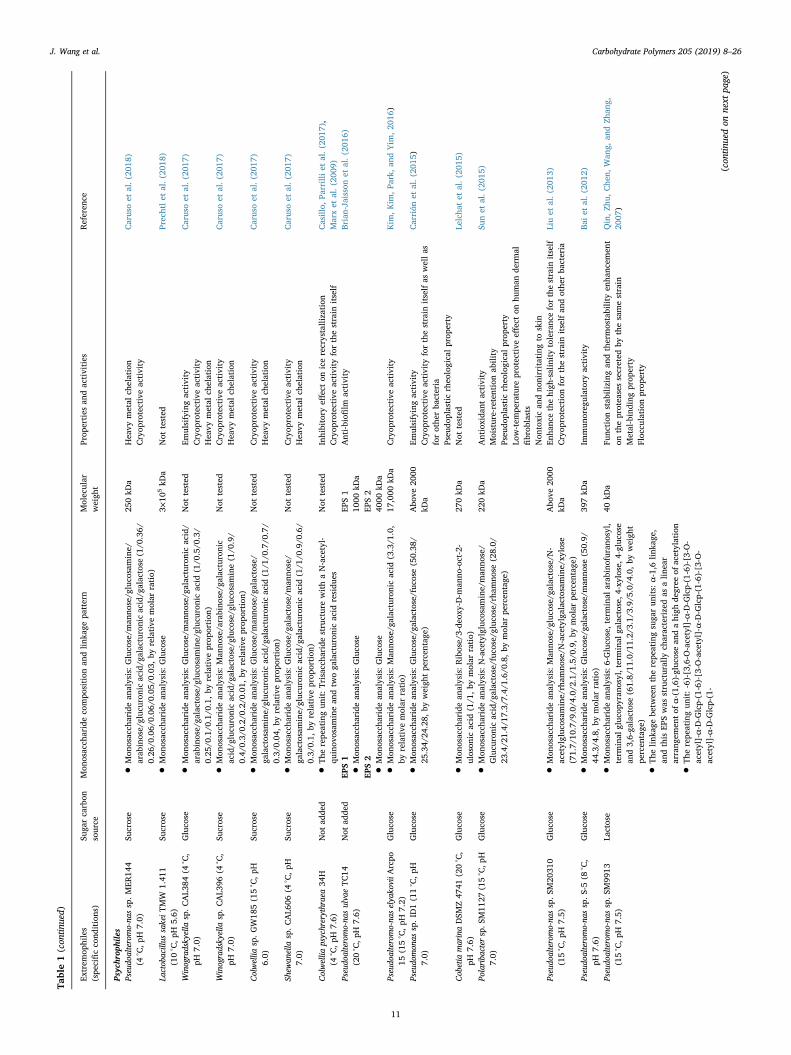
Table1(con
tinued)
Extrem
ophiles
(spe
cificco
nditions)
Suga
rcarbon
source
Mon
osacch
arideco
mpo
sition
andlin
kage
pattern
Molecular
weigh
tProp
erties
andactivities
Referen
ce
Psyc
hrop
hiles
Pseudo
alteromo-na
ssp.M
ER14
4(4
°C,p
H7.0)
Sucrose
•Mon
osacch
aridean
alysis:Gluco
se/m
anno
se/g
luco
samine/
arab
inose/gluc
uron
icacid/g
alacturonicacid/g
alactose
(1/0
.36/
0.26
/0.06/
0.06
/0.05/
0.03
,by
relative
molar
ratio)
250kD
aHeavy
metal
chelation
Cryop
rotectiveactivity
Carusoet
al.(20
18)
Lactobacillus
sakeiT
MW
1.41
1(10°C,p
H5.6)
Sucrose
•Mon
osacch
aridean
alysis:Gluco
se3×
105kD
aNot
tested
Prechtlet
al.(20
18)
Winogradsky
ella
sp.C
AL3
84(4
°C,
pH7.0)
Gluco
se•M
onosacch
aridean
alysis:Gluco
se/m
anno
se/g
alacturonicacid/
arab
inose/ga
lactose/gluc
osam
ine/gluc
uron
icacid
(1/0
.5/0
.3/
0.25
/0.1/0
.1/0
.1,by
relative
prop
ortion
)
Not
tested
Emulsifyingactivity
Cryop
rotectiveactivity
Heavy
metal
chelation
Carusoet
al.(20
17)
Winogradsky
ella
sp.C
AL3
96(4
°C,
pH7.0)
Sucrose
•Mon
osacch
aridean
alysis:Man
nose/arabino
se/g
alacturonic
acid/g
lucu
ronicacid/g
alactose/g
luco
se/g
luco
samine(1/0
.9/
0.4/
0.3/
0.2/
0.2/
0.01
,by
relative
prop
ortion
)
Not
tested
Cryop
rotectiveactivity
Heavy
metal
chelation
Carusoet
al.(20
17)
Colwellia
sp.G
W18
5(15°C,p
H6.0)
Sucrose
•Mon
osacch
aridean
alysis:Gluco
se/m
anno
se/g
alactose/
galactosam
ine/gluc
uron
icacid/g
alacturonicacid
(1/1
/0.7/0
.7/
0.3/
0.04
,byrelative
prop
ortion
)
Not
tested
Cryop
rotectiveactivity
Heavy
metal
chelation
Carusoet
al.(20
17)
Shew
anella
sp.C
AL6
06(4
°C,p
H7.0)
Sucrose
•Mon
osacch
aridean
alysis:Gluco
se/g
alactose/m
anno
se/
galactosam
ine/gluc
uron
icacid/g
alacturonicacid
(1/1
/0.9/0
.6/
0.3/
0.1,
byrelative
prop
ortion
)
Not
tested
Cryop
rotectiveactivity
Heavy
metal
chelation
Carusoet
al.(20
17)
Colwellia
psychrerythraea34
H(4
°C,p
H7.6)
Not
adde
d•T
herepe
atingun
it:T
risaccha
ride
structurewithaN-acetyl-
quinov
osam
inean
dtw
oga
lacturon
icacid
residu
esNot
tested
Inhibitory
effecton
icerecrystallization
Cryop
rotectiveactivity
forthestrain
itself
Casillo,
Parrilliet
al.(20
17),
Marxet
al.(20
09)
Pseudo
alteromo-na
sulvaeTC
14(20°C,p
H7.6)
Not
adde
dEP
S1
EPS1
1000
kDa
EPS2
4000
kDa
Anti-biofi
lmactivity
Brian-Jaissonet
al.(20
16)
•Mon
osacch
aridean
alysis:Gluco
seEP
S2
•Mon
osacch
aridean
alysis:Gluco
sePseudo
alteromo-na
selya
koviiA
rcpo
15(15°C,p
H7.2)
Gluco
se•M
onosacch
aridean
alysis:Man
nose/g
alacturonicacid
(3.3/1
.0,
byrelative
molar
ratio)
17,000
kDa
Cryop
rotectiveactivity
Kim
,Kim
,Park,
andYim
,201
6)
Pseudo
mon
assp.ID1(11°C,p
H7.0)
Gluco
se•M
onosacch
aridean
alysis:Gluco
se/g
alactose/fuc
ose(50.38
/25
.34/
24.28,
byweigh
tpe
rcen
tage
)Abo
ve20
00kD
aEm
ulsifyingactivity
Cryop
rotectiveactivity
forthestrain
itselfas
wellas
forothe
rba
cteria
Pseu
doplasticrheo
logicalprop
erty
Carrión
etal.(20
15)
Cobetia
marinaDSM
Z47
41(20°C,
pH7.6)
Gluco
se•M
onosacch
aridean
alysis:Ribose/3-de
oxy-D-m
anno
-oct-2-
uloson
icacid
(1/1
,bymolar
ratio)
270kD
aNot
tested
Lelcha
tet
al.(20
15)
Polariba
cter
sp.S
M11
27(15°C,p
H7.0)
Gluco
se•M
onosacch
aridean
alysis:N-acetylgluco
samine/man
nose/
Glucu
ronicacid/g
alactose/fuc
ose/gluc
ose/rham
nose
(28.0/
23.4/2
1.4/
17.3/7
.4/1
.6/0
.8,by
molar
percen
tage
)
220kD
aAntioxida
ntactivity
Moisture-retentionab
ility
Pseu
doplasticrheo
logicalprop
erty
Low-tem
perature
protective
effecton
human
derm
alfibrob
lasts
Non
toxican
dno
nirritatingto
skin
Sunet
al.(20
15)
Pseudo
alteromo-na
ssp.S
M20
310
(15°C,p
H7.5)
Gluco
se•M
onosacch
aridean
alysis:Man
nose/g
luco
se/g
alactose/N
-acetylgluc
osam
ine/rham
nose/N
-acetylgalactosamine/xy
lose
(71.7/
10.7/9
.0/4
.0/2
.1/1
.5/0
.9,by
molar
percen
tage
)
Abo
ve20
00kD
aEn
hanc
ethehigh
-salinitytoleranc
eforthestrain
itself
Cryop
rotectionforthestrain
itselfan
dothe
rba
cteria
Liuet
al.(20
13)
Pseudo
alteromo-na
ssp.S
-5(8
°C,
pH7.6)
Gluco
se•M
onosacch
aridean
alysis:Gluco
se/g
alactose/m
anno
se(50.9/
44.3/4
.8,b
ymolar
ratio)
397kD
aIm
mun
oreg
ulatoryactivity
Baiet
al.(20
12)
Pseudo
alteromo-na
ssp.S
M99
13(15°C,p
H7.5)
Lactose
•Mon
osacch
aridean
alysis:6
-Gluco
se,terminal
arab
inofuran
osyl,
term
inal
gluc
opyran
osyl,terminal
galactose,
4-xy
lose,4
-gluco
sean
d3,6-ga
lactose(61.8/
11.0/1
1.2/
3.1/
3.9/
5.0/
4.0,
byweigh
tpe
rcen
tage
)
•The
linka
gebe
tweentherepe
atingsuga
run
its:
α-1,6lin
kage
,an
dthis
EPSwas
structurally
characterizedas
alin
ear
arrang
emen
tofα
-(1,6)-gluco
sean
dahigh
degree
ofacetylation
•The
repe
atingun
it:-6)-[3,6-O-acetyl]-α-D
-Glcp-(1-6)-[3-O
-acetyl]-α-D-G
lcp-(1-6)-[3-O
-acetyl]-α-D
-Glcp-(1-6)-[3-O
-acetyl]-α-D-G
lcp-(1-
40kD
aFu
nction
stab
ilizing
andthermostabilityen
hanc
emen
ton
theproteasessecreted
bythesamestrain
Metal-binding
prop
erty
Floc
culation
prop
erty
Qin,Z
hu,C
hen,
Wan
g,an
dZh
ang,
2007
)
(con
tinuedon
next
page)
J. Wang et al. Carbohydrate Polymers 205 (2019) 8–26
11
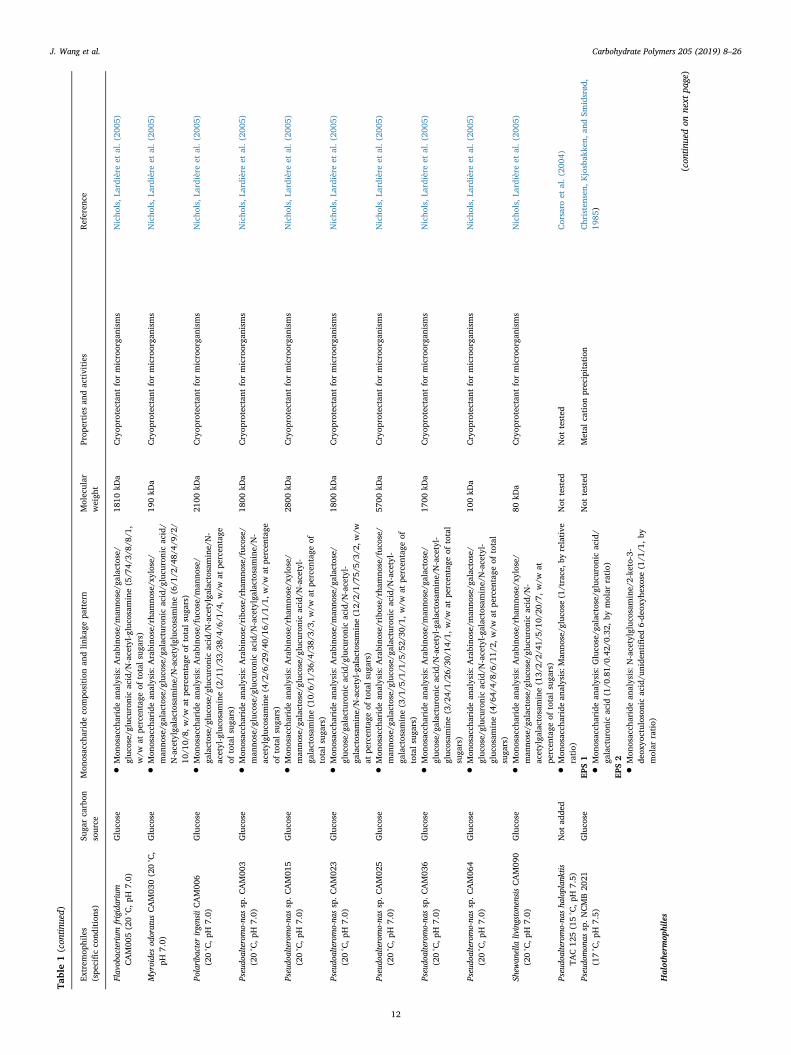
Table1(con
tinued)
Extrem
ophiles
(spe
cificco
nditions)
Suga
rcarbon
source
Mon
osacch
arideco
mpo
sition
andlin
kage
pattern
Molecular
weigh
tProp
erties
andactivities
Referen
ce
Flavobacterium
frigidarium
CAM00
5(20°C,p
H7.0)
Gluco
se•M
onosacch
aridean
alysis:Arabino
se/m
anno
se/g
alactose/
gluc
ose/gluc
uron
icacid/N
-acetyl-g
luco
samine(5/7
4/3/
8/8/
1,w/w
atpe
rcen
tage
oftotalsuga
rs)
1810
kDa
Cryop
rotectan
tformicroorga
nism
sNicho
ls,L
ardièreet
al.(20
05)
Myroidesod
oratus
CAM03
0(20°C,
pH7.0)
Gluco
se•M
onosacch
aridean
alysis:Arabino
se/rha
mno
se/xylose/
man
nose/g
alactose/g
luco
se/g
alacturonicacid/g
lucu
ronicacid/
N-acetylgalactosamine/N-acetylgluco
samine(6/1
/2/4
8/4/
9/2/
10/1
0/8,
w/w
atpe
rcen
tage
oftotalsuga
rs)
190kD
aCryop
rotectan
tformicroorga
nism
sNicho
ls,L
ardièreet
al.(20
05)
Polariba
cter
irgensiiCAM00
6(20°C,p
H7.0)
Gluco
se•M
onosacch
aridean
alysis:Arabino
se/fuc
ose/man
nose/
galactose/gluc
ose/gluc
uron
icacid/N
-acetylgalactosamine/N-
acetyl-gluco
samine(2/1
1/33
/38/
4/6/
1/4,
w/w
atpe
rcen
tage
oftotalsuga
rs)
2100
kDa
Cryop
rotectan
tformicroorga
nism
sNicho
ls,L
ardièreet
al.(20
05)
Pseudo
alteromo-na
ssp.C
AM00
3(20°C,p
H7.0)
Gluco
se•M
onosacch
aridean
alysis:Arabino
se/ribose/rham
nose/fuc
ose/
man
nose/g
luco
se/g
lucu
ronicacid/N
-acetylgalactosamine/N-
acetylgluc
osam
ine(4/2
/6/2
9/40
/16/
1/1/
1,w/w
atpe
rcen
tage
oftotalsuga
rs)
1800
kDa
Cryop
rotectan
tformicroorga
nism
sNicho
ls,L
ardièreet
al.(20
05)
Pseudo
alteromo-na
ssp.C
AM01
5(20°C,p
H7.0)
Gluco
se•M
onosacch
aridean
alysis:Arabino
se/rha
mno
se/xylose/
man
nose/g
alactose/g
luco
se/g
lucu
ronicacid/N
-acetyl-
galactosam
ine(10/
6/1/
36/4
/38/
3/3,
w/w
atpe
rcen
tage
oftotalsuga
rs)
2800
kDa
Cryop
rotectan
tformicroorga
nism
sNicho
ls,L
ardièreet
al.(20
05)
Pseudo
alteromo-na
ssp.C
AM02
3(20°C,p
H7.0)
Gluco
se•M
onosacch
aridean
alysis:Arabino
se/m
anno
se/g
alactose/
gluc
ose/ga
lacturon
icacid/g
lucu
ronicacid/N
-acetyl-
galactosam
ine/N-acetyl-g
alactosamine(12/
2/1/
75/5
/3/2
,w/w
atpe
rcen
tage
oftotalsuga
rs)
1800
kDa
Cryop
rotectan
tformicroorga
nism
sNicho
ls,L
ardièreet
al.(20
05)
Pseudo
alteromo-na
ssp.C
AM02
5(20°C,p
H7.0)
Gluco
se•M
onosacch
aridean
alysis:Arabino
se/ribose/rham
nose/fuc
ose/
man
nose/g
alactose/g
luco
se/g
alacturonicacid/N
-acetyl-
galactosam
ine(3/1
/5/1
/1/5
/52/
30/1
,w/w
atpe
rcen
tage
oftotalsuga
rs)
5700
kDa
Cryop
rotectan
tformicroorga
nism
sNicho
ls,L
ardièreet
al.(20
05)
Pseudo
alteromo-na
ssp.C
AM03
6(20°C,p
H7.0)
Gluco
se•M
onosacch
aridean
alysis:Arabino
se/m
anno
se/g
alactose/
gluc
ose/ga
lacturon
icacid/N
-acetyl-g
alactosamine/N-acetyl-
gluc
osam
ine(3/2
4/1/
26/3
0/14
/1,w/w
atpe
rcen
tage
oftotal
suga
rs)
1700
kDa
Cryop
rotectan
tformicroorga
nism
sNicho
ls,L
ardièreet
al.(20
05)
Pseudo
alteromo-na
ssp.C
AM06
4(20°C,p
H7.0)
Gluco
se•M
onosacch
aridean
alysis:Arabino
se/m
anno
se/g
alactose/
gluc
ose/gluc
uron
icacid/N
-acetyl-g
alactosamine/N-acetyl-
gluc
osam
ine(4/6
4/4/
8/6/
11/2
,w/w
atpe
rcen
tage
oftotal
suga
rs)
100kD
aCryop
rotectan
tformicroorga
nism
sNicho
ls,L
ardièreet
al.(20
05)
Shew
anella
livingstonensisCAM09
0(20°C,p
H7.0)
Gluco
se•M
onosacch
aridean
alysis:Arabino
se/rha
mno
se/xylose/
man
nose/g
alactose/g
luco
se/g
lucu
ronicacid/N
-acetylga
lactosam
ine(13/
2/2/
41/5
/10/
20/7
,w/w
atpe
rcen
tage
oftotalsuga
rs)
80kD
aCryop
rotectan
tformicroorga
nism
sNicho
ls,L
ardièreet
al.(20
05)
Pseudo
alteromo-na
sha
loplan
ktis
TAC12
5(15°C,p
H7.5)
Not
adde
d•M
onosacch
aridean
alysis:M
anno
se/g
luco
se(1/trace,b
yrelative
ratio)
Not
tested
Not
tested
Corsaro
etal.(20
04)
Pseudo
mon
assp.N
CMB20
21(17°C,p
H7.5)
Gluco
seEP
S1
Not
tested
Metal
cation
precipitation
Christensen
,Kjosbak
ken,
andSm
idsrød
,19
85)
•Mon
osacch
aridean
alysis:Gluco
se/g
alactose/g
lucu
ronicacid/
galacturon
icacid
(1/0
.81/
0.42
/0.32,
bymolar
ratio)
EPS2
•Mon
osacch
aridean
alysis:N-acetylgluco
samine/2-ke
to-3-
deox
yoctuloson
icacid/u
nide
ntified
6-de
oxyh
exose(1/1
/1,by
molar
ratio)
Halothe
rmop
hiles
(con
tinuedon
next
page)
J. Wang et al. Carbohydrate Polymers 205 (2019) 8–26
12
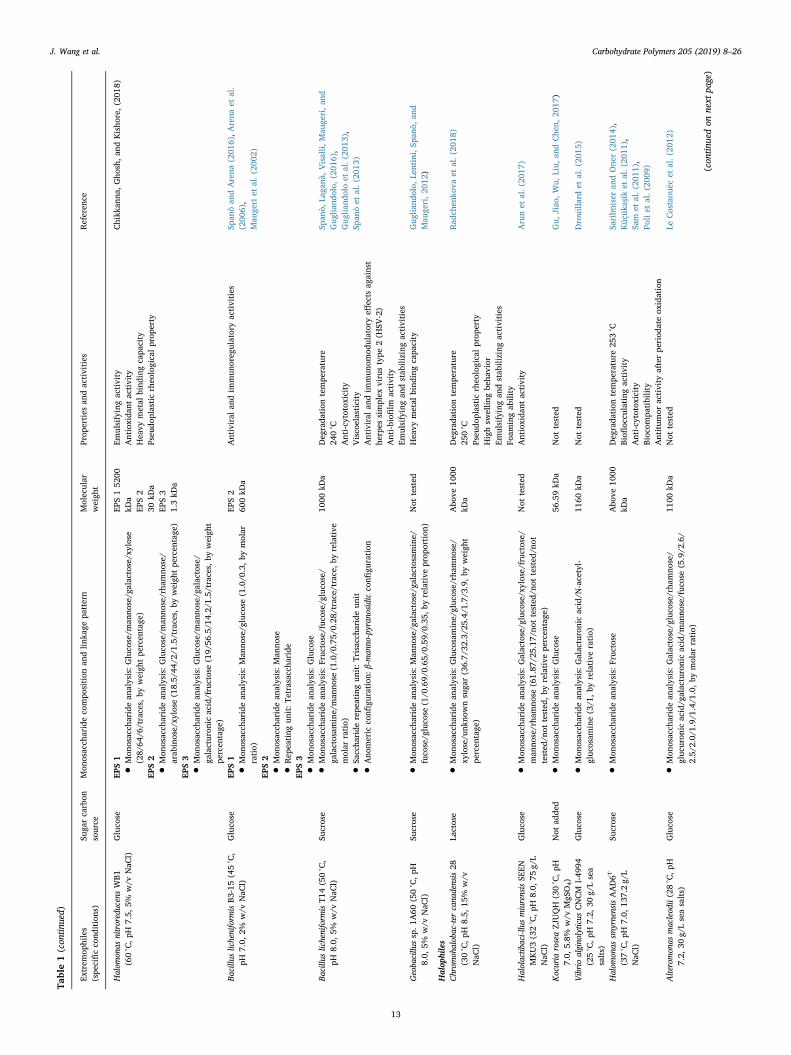
Table1(con
tinued)
Extrem
ophiles
(spe
cificco
nditions)
Suga
rcarbon
source
Mon
osacch
arideco
mpo
sition
andlin
kage
pattern
Molecular
weigh
tProp
erties
andactivities
Referen
ce
Halom
onas
nitroreducensWB1
(60°C,p
H7.5,
5%w/v
NaC
l)Gluco
seEP
S1
EPS152
00kD
aEP
S2
30kD
aEP
S3
1.3kD
a
Emulsifyingactivity
Antioxida
ntactivity
Heavy
metal
bind
ingcapa
city
Pseu
doplasticrheo
logicalprop
erty
Chikk
anna
,Gho
sh,a
ndKisho
re,(201
8)
•Mon
osacch
aridean
alysis:Gluco
se/m
anno
se/g
alactose/xylose
(28/
64/6
/traces,
byweigh
tpe
rcen
tage
)EP
S2
•Mon
osacch
aridean
alysis:Gluco
se/m
anno
se/rha
mno
se/
arab
inose/xy
lose
(18.5/
44/2
/1.5/traces,by
weigh
tpe
rcen
tage
)EP
S3
•Mon
osacch
aridean
alysis:Gluco
se/m
anno
se/g
alactose/
galacturon
icacid/fructose(19/
56.5/1
4.2/
1.5/
traces,b
yweigh
tpe
rcen
tage
)Ba
cillu
slicheniform
isB3
-15(45°C,
pH7.0,
2%w/v
NaC
l)Gluco
seEP
S1
EPS2
600kD
aAntiviral
andim
mun
oreg
ulatoryactivities
Span
òan
dArena
(201
6),A
rena
etal.
(200
6),
Mau
geri
etal.(20
02)
•Mon
osacch
aridean
alysis:Man
nose/g
luco
se(1.0/0
.3,by
molar
ratio)
EPS2
•Mon
osacch
aridean
alysis:Man
nose
•Rep
eating
unit:T
etrasaccha
ride
EPS3
•Mon
osacch
aridean
alysis:Gluco
seBa
cillu
slicheniform
isT1
4(50°C,
pH8.0,
5%w/v
NaC
l)Su
crose
•Mon
osacch
aridean
alysis:Fruc
tose/fuc
ose/gluc
ose/
galactosam
ine/man
nose
(1.0/0
.75/
0.28
/trace/trace,b
yrelative
molar
ratio)
•Sacch
ariderepe
atingun
it:T
risaccha
ride
unit
•Ano
meric
confi
guration
:β-m
anno
-pyran
osidic
confi
guration
1000
kDa
Deg
rada
tion
tempe
rature
240°C
Anti-cytotoxicity
Visco
elasticity
Antiviral
andim
mun
omod
ulatoryeff
ects
against
herpes
simplex
virustype
2(H
SV-2)
Anti-biofi
lmactivity
Emulsifyingan
dstab
ilizing
activities
Span
ò,La
ganà
,Visalli,
Mau
geri,a
ndGug
liand
olo,
(201
6),
Gug
liand
oloet
al.(20
13),
Span
òet
al.(20
13)
Geoba
cillu
ssp.1
A60
(50°C,p
H8.0,
5%w/v
NaC
l)Su
crose
•Mon
osacch
aridean
alysis:Man
nose/g
alactose/g
alactosamine/
fuco
se/g
luco
se(1/0
.69/
0.65
/0.59/
0.35
,byrelative
prop
ortion
)Not
tested
Heavy
metal
bind
ingcapa
city
Gug
liand
olo,
Lentini,Sp
anò,
and
Mau
geri,2
012)
Halop
hiles
Chrom
ohalobac-te
rcana
densis28
(30°C,p
H8.5,
15%
w/v
NaC
l)
Lactose
•Mon
osacch
aridean
alysis:Gluco
samine/gluc
ose/rham
nose/
xylose/u
nkno
wnsuga
r(36.7/
32.3/2
5.4/
1.7/
3.9,
byweigh
tpe
rcen
tage
)
Abo
ve10
00kD
aDeg
rada
tion
tempe
rature
250°C
Pseu
doplasticrheo
logicalprop
erty
Highsw
ellin
gbe
havior
Emulsifyingan
dstab
ilizing
activities
Foam
ingab
ility
Rad
chen
kova
etal.(20
18)
Halolactib
aci-llusmiurensisSE
ENMKU3(32°C,p
H8.0,
75g/
LNaC
l)
Gluco
se•M
onosacch
aridean
alysis:Galactose/g
luco
se/xylose/fruc
tose/
man
nose/rha
mno
se(61.87
/25.17
/not
tested
/not
tested
/not
tested
/not
tested
,byrelative
percen
tage
)
Not
tested
Antioxida
ntactivity
Arunet
al.(20
17)
Kocuria
roseaZJ
UQH
(30°C,p
H7.0,
5.8%
w/v
MgS
O4)
Not
adde
d•M
onosacch
aridean
alysis:Gluco
se56
.59kD
aNot
tested
Gu,
Jiao
,Wu,
Liu,
andChe
n,20
17)
Vibrioalgino
lyticus
CNCM
I-49
94(25°C,p
H7.2,
30g/
Lsea
salts)
Gluco
se•M
onosacch
aridean
alysis:Galacturonicacid/N
-acetyl-
gluc
osam
ine(3/1
,byrelative
ratio)
1160
kDa
Not
tested
Drouilla
rdet
al.(20
15)
Halom
onas
smyrnensisAAD6T
(37°C,p
H7.0,
137.2g/
LNaC
l)
Sucrose
•Mon
osacch
aridean
alysis:Fruc
tose
Abo
ve10
00kD
aDeg
rada
tion
tempe
rature
253°C
Biofl
occu
lating
activity
Anti-cytotoxicity
Bioc
ompa
tibility
Antitum
oractivity
afterpe
riod
ateox
idation
Sarilm
iser
andOne
r(201
4),
Küç
ükaşik
etal.(20
11),
Sam
etal.(20
11),
Poliet
al.(20
09)
Alteromon
asmacleod
ii(28°C,p
H7.2,
30g/
Lseasalts)
Gluco
se•M
onosacch
aridean
alysis:Galactose/g
luco
se/rha
mno
se/
gluc
uron
icacid/g
alacturonicacid/m
anno
se/fuc
ose(5.9/2
.6/
2.5/
2.0/
1.9/
1.4/
1.0,
bymolar
ratio)
1100
kDa
Not
tested
LeCostaou
ëcet
al.(20
12)
(con
tinuedon
next
page)
J. Wang et al. Carbohydrate Polymers 205 (2019) 8–26
13
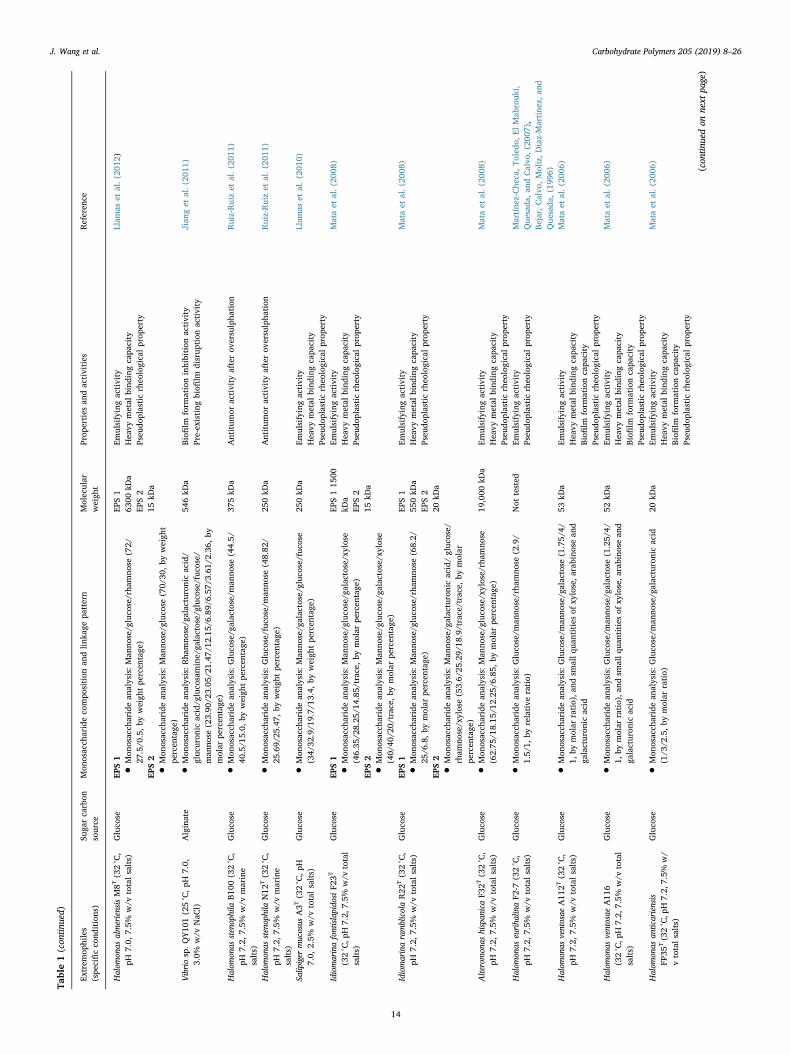
Table1(con
tinued)
Extrem
ophiles
(spe
cificco
nditions)
Suga
rcarbon
source
Mon
osacch
arideco
mpo
sition
andlin
kage
pattern
Molecular
weigh
tProp
erties
andactivities
Referen
ce
Halom
onas
almeriensisM8T
(32°C,
pH7.0,
7.5%
w/v
totalsalts)
Gluco
seEP
S1
EPS1
6300
kDa
EPS2
15kD
a
Emulsifyingactivity
Heavy
metal
bind
ingcapa
city
Pseu
doplasticrheo
logicalprop
erty
Llam
aset
al.(20
12)
•Mon
osacch
aridean
alysis:Man
nose/g
luco
se/rha
mno
se(72/
27.5/0
.5,b
yweigh
tpe
rcen
tage
)EP
S2
•Mon
osacch
aridean
alysis:Man
nose/g
luco
se(70/
30,by
weigh
tpe
rcen
tage
)Vibriosp.Q
Y10
1(25°C,p
H7.0,
3.0%
w/v
NaC
l)Alginate
•Mon
osacch
aridean
alysis:Rha
mno
se/g
alacturonicacid/
gluc
uron
icacid/g
luco
samine/ga
lactose/gluc
ose/fuco
se/
man
nose
(23.90
/23.05
/21.47
/12.15
/6.89/
6.57
/3.61/
2.36
,by
molar
percen
tage
)
546kD
aBiofi
lmform
ationinhibition
activity
Pre-existing
biofi
lmdisrup
tion
activity
Jian
get
al.(20
11)
Halom
onas
stenophila
B100
(32°C,
pH7.2,
7.5%
w/v
marine
salts)
Gluco
se•M
onosacch
aridean
alysis:Gluco
se/g
alactose/m
anno
se(44.5/
40.5/1
5.0,
byweigh
tpe
rcen
tage
)37
5kD
aAntitum
oractivity
afterov
ersulpha
tion
Ruiz-Ruizet
al.(20
11)
Halom
onas
stenophila
N12
T(32°C,
pH7.2,
7.5%
w/v
marine
salts)
Gluco
se•M
onosacch
aridean
alysis:Gluco
se/fuc
ose/man
nose
(48.82
/25
.69/
25.47,
byweigh
tpe
rcen
tage
)25
0kD
aAntitum
oractivity
afterov
ersulpha
tion
Ruiz-Ruizet
al.(20
11)
Salip
iger
mucosus
A3T
(32°C,p
H7.0,
2.5%
w/v
totalsalts)
Gluco
se•M
onosacch
aridean
alysis:Man
nose/g
alactose/g
luco
se/fuc
ose
(34/
32.9/1
9.7/
13.4,by
weigh
tpe
rcen
tage
)25
0kD
aEm
ulsifyingactivity
Heavy
metal
bind
ingcapa
city
Pseu
doplasticrheo
logicalprop
erty
Llam
aset
al.(20
10)
Idiomarinafontislapido
siF2
3T
(32°C,p
H7.2,
7.5%
w/v
total
salts)
Gluco
seEP
S1
EPS115
00kD
aEP
S2
15kD
a
Emulsifyingactivity
Heavy
metal
bind
ingcapa
city
Pseu
doplasticrheo
logicalprop
erty
Mataet
al.(20
08)
•Mon
osacch
aridean
alysis:Man
nose/g
luco
se/g
alactose/xylose
(46.35
/28.25
/14.85
/trace,by
molar
percen
tage
)EP
S2
•Mon
osacch
aridean
alysis:Man
nose/g
luco
se/g
alactose/xylose
(40/
40/2
0/trace,
bymolar
percen
tage
)IdiomarinaramblicolaR22
T(32°C,
pH7.2,
7.5%
w/v
totalsalts)
Gluco
seEP
S1
EPS1
550kD
aEP
S2
20kD
a
Emulsifyingactivity
Heavy
metal
bind
ingcapa
city
Pseu
doplasticrheo
logicalprop
erty
Mataet
al.(20
08)
•Mon
osacch
aridean
alysis:Man
nose/g
luco
se/rha
mno
se(68.2/
25/6
.8,b
ymolar
percen
tage
)EP
S2
•Mon
osacch
aridean
alysis:M
anno
se/g
alacturonicacid/gluc
ose/
rham
nose/xylose(53.6/
25.29/
18.9/trace/trace,by
molar
percen
tage
)Alteromon
ashispan
icaF3
2T(32°C,
pH7.2,
7.5%
w/v
totalsalts)
Gluco
se•M
onosacch
aridean
alysis:Man
nose/g
luco
se/xylose/rham
nose
(62.75
/18.15
/12.25
/6.85,
bymolar
percen
tage
)19
,000
kDa
Emulsifyingactivity
Heavy
metal
bind
ingcapa
city
Pseu
doplasticrheo
logicalprop
erty
Mataet
al.(20
08)
Halom
onas
euriha
linaF2
-7(32°C,
pH7.2,
7.5%
w/v
totalsalts)
Gluco
se•M
onosacch
aridean
alysis:Gluco
se/m
anno
se/rha
mno
se(2.9/
1.5/
1,by
relative
ratio)
Not
tested
Emulsifyingactivity
Pseu
doplasticrheo
logicalprop
erty
Martíne
z-Che
ca,To
ledo
,ElMab
rouk
i,Que
sada
,an
dCalvo
,(20
07),
Bejar,
Calvo
,Moliz,D
iaz-Martine
z,an
dQue
sada
,(199
6)Halom
onas
ventosae
A11
2T(32°C,
pH7.2,
7.5%
w/v
totalsalts)
Gluco
se•M
onosacch
aridean
alysis:Gluco
se/m
anno
se/g
alactose
(1.75/
4/1,
bymolar
ratio),a
ndsm
allq
uantitiesof
xylose,a
rabino
sean
dga
lacturon
icacid
53kD
aEm
ulsifyingactivity
Heavy
metal
bind
ingcapa
city
Biofi
lmform
ationcapa
city
Pseu
doplasticrheo
logicalprop
erty
Mataet
al.(20
06)
Halom
onas
ventosae
A11
6(32°C,p
H7.2,
7.5%
w/v
total
salts)
Gluco
se•M
onosacch
aridean
alysis:Gluco
se/m
anno
se/g
alactose
(1.25/
4/1,
bymolar
ratio),a
ndsm
allq
uantitiesof
xylose,a
rabino
sean
dga
lacturon
icacid
52kD
aEm
ulsifyingactivity
Heavy
metal
bind
ingcapa
city
Biofi
lmform
ationcapa
city
Pseu
doplasticrheo
logicalprop
erty
Mataet
al.(20
06)
Halom
onas
anticariensis
FP35
T(32°C,p
H7.2,
7.5%
w/
vtotalsalts)
Gluco
se•M
onosacch
aridean
alysis:Gluco
se/m
anno
se/g
alacturonicacid
(1/3
/2.5,b
ymolar
ratio)
20kD
aEm
ulsifyingactivity
Heavy
metal
bind
ingcapa
city
Biofi
lmform
ationcapa
city
Pseu
doplasticrheo
logicalprop
erty
Mataet
al.(20
06)
(con
tinuedon
next
page)
J. Wang et al. Carbohydrate Polymers 205 (2019) 8–26
14
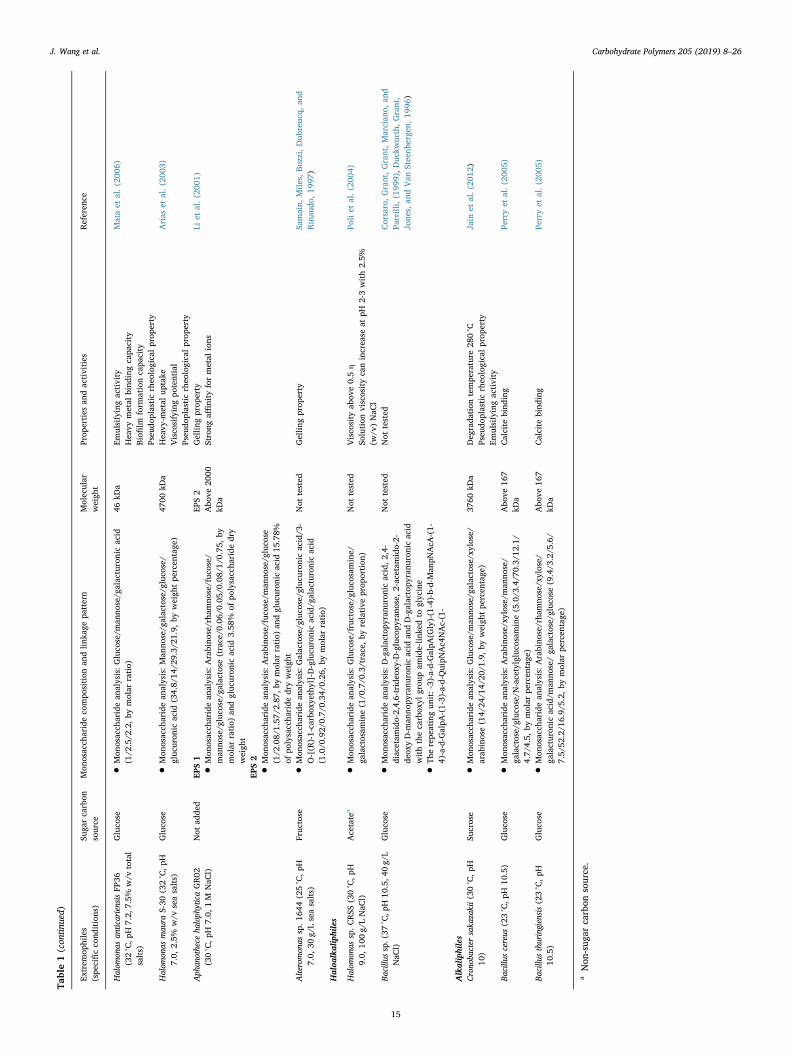
Table1(con
tinued)
Extrem
ophiles
(spe
cificco
nditions)
Suga
rcarbon
source
Mon
osacch
arideco
mpo
sition
andlin
kage
pattern
Molecular
weigh
tProp
erties
andactivities
Referen
ce
Halom
onas
anticariensisFP
36(32°C,p
H7.2,
7.5%
w/v
total
salts)
Gluco
se•M
onosacch
aridean
alysis:Gluco
se/m
anno
se/g
alacturonicacid
(1/2
.5/2
.2,by
molar
ratio)
46kD
aEm
ulsifyingactivity
Heavy
metal
bind
ingcapa
city
Biofi
lmform
ationcapa
city
Pseu
doplasticrheo
logicalprop
erty
Mataet
al.(20
06)
Halom
onas
mau
raS-30
(32°C,p
H7.0,
2.5%
w/v
seasalts)
Gluco
se•M
onosacch
aridean
alysis:Man
nose/g
alactose/g
luco
se/
gluc
uron
icacid
(34.8/
14/2
9.3/
21.9,by
weigh
tpe
rcen
tage
)47
00kD
aHeavy
-metal
uptake
Visco
sifyingpo
tential
Pseu
doplasticrheo
logicalprop
erty
Arias
etal.(20
03)
Aph
anothece
haloph
yticaGR02
(30°C,p
H7.0,
1M
NaC
l)Not
adde
dEP
S1
EPS2
Abo
ve20
00kD
a
Gellin
gprop
erty
Strong
affinity
formetal
ions
Liet
al.(20
01)
•Mon
osacch
aridean
alysis:Arabino
se/rha
mno
se/fuc
ose/
man
nose/g
luco
se/g
alactose
(trace/0
.06/
0.05
/0.08/
1/0.75
,by
molar
ratio)
andgluc
uron
icacid
3.58
%of
polysaccha
ride
dry
weigh
tEP
S2
•Mon
osacch
aridean
alysis:Arabino
se/fuc
ose/man
nose/g
luco
se(1/2
.08/
1.57
/2.87,
bymolar
ratio)
andgluc
uron
icacid
15.78%
ofpo
lysaccha
ride
dryweigh
tAlteromon
assp.1
644(25°C,p
H7.0,
30g/
Lseasalts)
Fruc
tose
•Mon
osacch
aridean
alysis:G
alactose/g
luco
se/g
lucu
ronicacid/3
-O-[(R
)-1-carbox
yethyl]-D-glucu
ronicacid/g
alacturonicacid
(1.0/0
.92/
0.7/
0.34
/0.26,
bymolar
ratio)
Not
tested
Gellin
gprop
erty
Samain,
Mile
s,Bo
zzi,Dub
reuc
q,an
dRinau
do,1
997)
Haloa
lkaliphiles
Halom
onas
sp.C
RSS
(30°C,p
H9.0,
100g/
LNaC
l)Acetate
a•M
onosacch
aridean
alysis:Gluco
se/fructose/gluc
osam
ine/
galactosam
ine(1/0
.7/0
.3/trace,by
relative
prop
ortion
)Not
tested
Visco
sity
abov
e0.5η
Solution
viscositycanincrease
atpH
2-3with2.5%
(w/v
)NaC
l
Poliet
al.(20
04)
Bacillu
ssp.(37
°C,p
H10
.5,4
0g/
LNaC
l)Gluco
se•M
onosacch
aridean
alysis:D-galactopy
ranu
ronicacid,2
,4-
diacetam
ido-2,4,6-trideo
xy-D
-gluco
pyrano
se,2-acetam
ido-2-
deox
yD-m
anno
pyranu
ronicacid
andD-galactopy
ranu
ronicacid
withthecarbox
ylgrou
pam
ide-lin
kedto
glycine
•The
repe
atingun
it:-3)-a-d-G
alpA
(Gly)-(1-4)-b-d-Man
pNAcA
-(1-
4)-a-d-G
alpA
-(1-3)-a-d-Q
uipN
Ac4
NAc-(1-
Not
tested
Not
tested
Corsaro,G
rant,G
rant,M
arcian
o,an
dPa
rrilli,(199
9),D
uckw
orth,Grant,
Jone
s,an
dVan
Steenb
erge
n,19
96)
Alkaliphiles
Crono
bacter
saka
zakii(30
°C,p
H10
)Su
crose
•Mon
osacch
aridean
alysis:Gluco
se/m
anno
se/g
alactose/xylose/
arab
inose(14/
24/1
4/20
/1.9,by
weigh
tpe
rcen
tage
)37
60kD
aDeg
rada
tion
tempe
rature
280°C
Pseu
doplasticrheo
logicalprop
erty
Emulsifyingactivity
Jain
etal.(20
12)
Bacillu
scereus
(23°C,p
H10
.5)
Gluco
se•M
onosacch
aridean
alysis:Arabino
se/xylose/man
nose/
galactose/gluc
ose/N-acetylgluco
samine(5.0/3
.4/7
0.3/
12.1/
4.7/
4.5,
bymolar
percen
tage
)
Abo
ve16
7kD
aCalcite
bind
ing
Perryet
al.(20
05)
Bacillu
sthuringiensis(23°C,p
H10
.5)
Gluco
se•M
onosacch
aridean
alysis:Arabino
se/rha
mno
se/xylose/
galacturon
icacid/m
anno
se/ga
lactose/gluc
ose(9.4/3
.2/5
.6/
7.5/
52.2/1
6.9/
5.2,
bymolar
percen
tage
)
Abo
ve16
7kD
aCalcite
bind
ing
Perryet
al.(20
05)
aNon
-sug
arcarbon
source.
J. Wang et al. Carbohydrate Polymers 205 (2019) 8–26
15

microorganisms in a marine environment may provide both cryopro-tection and a buffering effect against low temperature and high salinitysimultaneously. The secondary molecular structure analysis of psy-chrophilic EPS indicates that a pseudohelicoidal structure may be ad-vantageous for the inhibition of ice recrystallization (Casillo, Parrilliet al., 2017). Moreover, the decoration by amino acid motifs onto themonosaccharide moieties was speculated to endow a structural equili-brium between hydrophilic and hydrophobic regions in the EPS mole-cule, and thus contribute to the inhibitory effect on ice crystal devel-opment (Casillo, Ståhle et al., 2017). The sulphate moieties inpsychrophilic EPSs may also play a significant role against extremelycold environments (Nichols, Guezennec et al., 2005; Nichols, Bowmanet al., 2005). The physical, rheological, and chemical properties of EPSscan be influenced by the length of the polymer chain, and the highmolecular weights of EPSs from psychrophiles provide greater oppor-tunity for complex entanglement of polymer chains and intramolecularassociations, which may contribute to the tertiary structure and en-hance the physical behavior of the EPSs in their environment (Nichols,Bowman et al., 2005). Besides, the EPSs with higher molecular weightalso possess better water binding capacity than EPSs with lower mo-lecular weight (Prechtl, Wefers, Jakob, & Vogel, 2018).
Normally, psychrophilic EPS production can be inhibited by a re-latively elevated temperature, in the region of 20 °C and above (Nichols,Garon, Bowman, Raguenes, & Guezennec, 2004, 2005). The contents ofmonosaccharide components in psychrophilic EPSs can be modifiedthrough change of temperature, and some of the monosaccharides andother residues in EPSs from psychrophiles may help to confer ad-vantageous cryoprotectant properties. For example, the uronic acidcontent in the EPSs produced by Pseudoalteromonas sp. CAM025 at−2 °C and 10 °C was significantly higher than that at 20 °C; and themonosaccharide compositions were also found to differ among the EPSsharvested at −2 °C, 10 °C, and 20 °C (Nichols, Bowman et al., 2005).The psychrotolerant strain Lactobacillus sakei TMW 1.411 produceddextran with less branching and higher molecular weight at 10 °C thanthe dextran produced at 30 °C (Prechtl et al., 2018). At temperaturesbelow the optimum temperature for cell growth, the psychrophiles werestimulated to produce excessive EPSs (Marx, Carpenter, & Deming,2009; Nevot et al., 2008; Nichols et al., 2004). This is consistent withthe fact that EPS production is one of the main mechanisms to protectextremophiles and enable them to survive in extreme conditions.Therefore, output of EPS for each cell can be enhanced with the dete-rioration of environmental conditions in a certain range, albeit the cellgrowth may sharply decrease. Enhancing net EPS production may thusinvolve identifying the optimal trade-off between increased EPS pro-duction per cell and reduced cell count.
In several former studies, the stabilization effect of psychrophilicEPS for protease against thermal denaturation was confirmed (Huston,Methe, & Deming, 2004; Junge, Eicken, Swanson, & Deming, 2006;Marx et al., 2009), which indicates that psychrophilic biopolymers canbe applied to the stabilization of industrially promising enzymes used inunfavorable conditions. In future research on psychrophilic EPSs, it isrecommended that significant insights may be found by comparing thestructure and function of EPSs from different culture conditions, inorder to reveal what kind of structure can be more advantageous forprotection and stabilization effects.
2.3. EPSs produced by halophiles
Moderately halophilic bacteria are defined as those which growoptimally in media containing 5–20% (w/v) salts, and they constitutethe most important eubacteria group living in hypersaline habitats(Ollivier, Caumette, Garcia, & Mah, 1994; C. Qian et al., 2018). Mosthalophilic EPSs are heteropolysaccharides, and mannose and glucoseare the most common monosaccharide moieties in halophilic EPSs(Table 1). So far, the research focus for halophilic EPSs properties hasbeen emulsifying activity, gelling properties, heavy metal binding
capacity, and rheological properties, with existing and potential ap-plications in a range of industrial fields, such as utilization as a sub-stitute for xanthan gum in the food industry.
Changes in salinity affects the biosynthesis of halophilic EPSs,especially the ratio for each type of monosaccharide composition. Toprotect the microorganism from increasing salinity, the content of somemonosaccharide components in EPS may need to be modified in orderto maintain its functions. For the EPS obtained from strain Aphanothecehalophytica GR02, the proportions of galactose and rhamnose decreasedwhen the NaCl concentration in the medium was elevated from 0.5 to2.0 M; in contrast, the proportions of arabinose and glucose increasedwith NaCl concentration. Meanwhile, the monosaccharides present inthe EPS at different salinities stayed the same (P. Li, Liu, & Xu, 2001).This indicates that the increase of glucose and arabinose, and the de-crease of galactose and rhamnose in the EPS secreted by Aphanothecehalophytica GR02 may be advantageous to its survival in a high salinityenvironment. Mata et al. (2006) mentioned that for the strain Halo-monas ventosae A112T, its EPS incorporated a significant quantity ofsulphate. Sulphate is not commonly found in mesophilic EPSs; however,it has been observed in the EPSs excreted by microorganisms living insaline habitats. In addition, the EPSs from halophiles usually containsignificant amounts of uronic acids. The high viscosity of the EPS so-lution at acidic pH and the gelification capacity may be due to the highuronic acid content (Béjar, Llamas, Calvo, & Quesada, 1998). EPSs withhigh concentrations of charged components (e.g. uronic acids) oftenform gels in the presence of metal ions and have enormous potential forremoving toxic metal from polluted environments and wastewater as analternative to other physical and chemical methods.
2.4. EPSs produced by acidophiles
Acidophiles are extremophiles which inhabit a low pH environment,usually less than pH 3 for optimum growth. Some of the acidophilescannot grow at all in a neutral pH condition (Baker-Austin & Dopson,2007; Johnson, 1998; Johnson, Joulian, d’Hugues, & Hallberg, 2008).Both natural and artificial acidic niches can occur in the biosphere, suchas a sulfidic mine area or a marine volcanic vent. The acidic environ-ments usually include the presence of sulphur, sulphide, and theiroxidates. Pyrite is one of the main acidic niches for acidophiles. Theseareas are quite toxic due to high concentrations of various heavy metalsulphides, but they are rich in valuable metals, such as Fe, Cu, Co, Al,Mg, Zn, and Mn (Dopson, Baker-Austin, Koppineedi, & Bond, 2003; Jiaoet al., 2010; Johnson et al., 2008; Nicolaus et al., 2010).
Compared with the research for other kinds of extremophilic EPSs,the acidophilic EPSs have not been studied sufficiently to reveal theirfermentation process, molecular structure, or properties. Usually EPSsfrom acidophiles are considered as bioproducts generated in anotherbioprocessing technology such as a bioleaching process. For acid-ophiles, the genome analysis cannot identify ubiquitous DNA adapta-tions for growth in an extremely low pH environment (Baker-Austin &Dopson, 2007). On the other hand, the EPSs produced by acidophilesmay play a protective role against stress conditions related to the lowpH and presence of metals. Acidophilic EPS biosynthesis can be in-hibited by increased temperature during the bioleaching process, andthe inhibited EPS production may have been related to the loss ofbioleaching efficiency observed in the reactor when the temperaturewas increased (d’Hugues et al., 2008). This phenomenon indicates thatthe acidophilic EPSs protecting acidophiles from an acidic environmentare not able to protect them against a relatively high temperaturecondition, unlike thermophilic EPSs. Therefore, it is of significant in-terest to explore the acidophilic EPSs for functional diversity elucida-tion through molecular level structure and comparison among acid-ophilic and other extremophilic EPSs as models.
Some acidophilic EPSs were discovered during the study of extra-cellular polymeric substances, which are one of the major componentsin biofilms, and they mainly consist of EPSs, proteins, and nucleic acids
J. Wang et al. Carbohydrate Polymers 205 (2019) 8–26
16

(Flemming & Wingender, 2010; Moreno-Paz, Gómez, Arcas, & Parro,2010; Subramanian, Yan, Tyagi, & Surampalli, 2010; Vu, Chen,Crawford, & Ivanova, 2009). The extracellular polymeric substancescontaining acidophilic EPSs are usually generated by mixed culturesduring the bioleaching process. Bioleaching uses the oxidation ability ofbacteria to dissolve metal sulphides in order to facilitate the extractionand recovery of precious metals from primary ores and concentrates.The involved microbial consortia are mainly composed of acidophilic,autotrophic iron-oxidizing, and sulphur-oxidizing bacteria (Michelet al., 2009). In Zeng’s report (Zeng et al., 2010), an acidophilic mixedculture was able to produce extracellular polymeric substances duringthe bioleaching process, and Acidithiobacillus caldus and Leptospirillumferriphilum were considered as the dominant microorganisms in themixed culture. The extracellular polymeric substance had protein,polysaccharide, fatty acid, and ferric ion as its main components.Rhamnose, fucose, xylose, mannose, glucose, and uronic acids were thecomponents of the polysaccharide which could be considered to comefrom the EPS excreted by the mixed culture during the bioleachingprocess. The percentages of these components varied at different sam-pling time during bioleaching, while the presence of these componentsremained stable. A pure culture, Thiobacillus ferrooxidans, was alsocarried in the bioleaching process, and the monosaccharide units of thecarbohydrate in the extracellular polymeric substance were rhamnose,fucose, xylose, mannose, glucose, and glucuronic acid. This composi-tion varied greatly when T. ferrooxidans was grown in a differentmedium containing iron (II) sulphate, pyrite, or sulfur as the solidsubstrate (Gehrke, Telegdi, Thierry, & Sand, 1998).
2.5. EPSs produced by alkaliphiles
The alkaliphiles are microorganisms that grow optimally or verywell at pH values above 9, often between 10 and 12, but cannot grow orgrow slowly at near-neutral pH values (Horikoshi, 1999). Soda lakesand deserts represent the most stable, naturally occurring alkaline en-vironments which can be found all over the world (Rees, Grant, Jones,& Heaphy, 2004). The enzymes isolated from alkaliphiles, includingalkaline proteases, amylases, cellulases, and lipase, have been appliedin various industrial sectors such as the detergent industry (Ito et al.,1998). As with other kinds of extremophiles, the alkaliphiles produceEPSs as metabolic products. So far, certain functions of EPSs from al-kaliphiles have been partially studied (Table 1), but more research onmolecular structure, properties, and the biosynthesis pathway of alka-liphilic EPSs are necessary to improve scientific understanding and toenable targeted industrial applications.
Alkaliphilic EPSs are functional for the attachment of the associatedmicrobial strains to a certain matrix. For example, the binding strengthto calcite was found to be due to the chemical properties of the EPSssecreted by two natural alkaliphiles isolated from biofilms on historiclimestone. Meanwhile, these two alkaliphilic EPSs could also contributeto calcite dissolution in the biofilm development process (Perry et al.,2005). Unlike most other extremophiles, for which sugar is the optimalcarbon source for EPS production, the most efficient carbon source forEPS production of the haloalkalophilic strain Halomonas sp. CRSS wasacetate. The growth conditions strongly influenced the cumulativeproduction, relative fractions of different monosaccharides, andmonomer compositions of the EPS from Halomonas sp. CRSS (Poli et al.,2004).
3. Metabolic and genomic engineering of extremophilic EPSs
Extremophilic EPSs have increasing significance in material andbiomedical applications that require a more profound understanding ofthe metabolic pathways and biosynthetic mechanisms of EPS in order tocontrol the production process and molecular structure, and hence thephysiochemical properties. The development of engineered EPS-pro-ducing strains can also reduce their exceptionally expensive production
costs, allowing extremophilic EPSs to compete in the biopolymermarket. Several biopolymers from mesophiles and neutrophiles, such ascellulose, alginate, gellan, and sphingan have already been profoundlystudied for the enzymes and genes involved in their biosyntheticpathways (Ates, 2015; Schmid, Sieber, & Rehm, 2015). However, theinformation about metabolic pathways and functional assignment ofgene clusters for extremophilic EPS biosynthesis is still limited. Re-search focusing on genetically modified strains capable of producinghighly improved levels of extremophilic EPS is also necessary since,compared to mesophilic and neutrophilic EPS-producing strains, ex-tremophilic bacteria are relatively inefficient at producing EPSs.
3.1. Engineering strategies in EPS biosynthetic pathways for improved EPSproduction and modified molecular weight
EPS biosynthesis is highly associated with catabolic processes ofoxidation and does not interfere with other anabolic bioprocesses(Chawla et al., 2009). As a carbon and energy intensive process, thebiosynthesis of extremophilic EPSs usually requires the recruitment ofnucleoside diphosphate saccharides (NDP-sugars) as precursors, glyco-syltransferases (GTs) for assembly, and membrane proteins for thetransfer of repeat units across cell envelope. Generally, the EPS bio-synthetic pathway starts from glycolysis of simple sugar for cytosolicformation of the NDP-sugar precursors; then the monosaccharides aresequentially transported from nucleotide-sugar donors to activated lipidcarriers and assembled as repeating units of polysaccharide throughGTs. Finally, the EPS needs to be exported to an extracellular en-vironment. Based on the general biosynthetic pathway of EPS, the genesinvolved can be organized into three functional types: (1) genes in-volved in NDP-sugar synthesis, (2) genes coding for GTs required forbiosynthesis of EPS repeating unit, (3) genes encoding proteins forpolymerization and export (Ates, 2015).
During the first phase of EPS biosynthesis, the NDP-sugars representthe interface between primary and secondary metabolism (Ates, Arga,& Oner, 2013). A bottleneck is the low level of activated NDP-sugarprecursors which can be exploited as design space through metabolicengineering to alter the expression of enzymes involved in the centralmetabolism for supplying nucleotide-sugar precursors. The higher EPSproducing mutant demonstrated that the specific activities of phos-phoglucomutase, UDP-glucose pyrophosphorylase, UDP-glucose dehy-drogenase, and UDP-galactose-4-epimerase were higher than those inthe wild-type strain (Fig. 2), indicating these enzymes involved in NDP-sugar synthesis can be potential targets for enhancement of EPS pro-duction (Li et al., 2010; Welman, Maddox, & Archer, 2006; Zhu et al.,2014). Although it is still relatively nascent for extremophilic bacteriato be applied as intact platforms for metabolic engineering, a group ofextremophiles have already been metabolically engineered for en-hanced biofuel or enzyme production due to the recent expansion of thegenetic systems and tools for extremophiles (Lin & Xu, 2013; Zeldeset al., 2015). In EPS-producing bacteria, the sugar substrates are eitherconverted into EPS synthesis or cell mass by alternative intermediarymetabolic routes. The rerouting of the carbon flux through the aug-mentation of a critical enzyme at the principal branch point to NDP-sugar synthesis was considered as a strategy to enhance the EPS pro-duction of several mesophilic bacteria. The homologous over-expres-sion of phosphoglucomutase in Sphingomonas sanxanigenens strain re-sulted in a 17% increase in EPS production (Huang et al., 2013).However, the flow of carbon towards the synthesis of EPS by Sphingo-monas sp. strain S7 was manipulated by augmenting the cellularphosphoglucomutase activity with additional genes, and no significantincrease in EPS yield was observed (Thorne, Mikolajczak, Armentrout,& Pollock, 2000). The over-expression of UDP-glucose pyropho-sphorylase involved in the synthesis of UDP-glucose also had negligibleeffect on EPS productivity. Meanwhile, the inactivation of glucose-6-phosphate dehydrogenase could not divert carbon flow toward EPSsynthesis (Sá-Correia et al., 2002). On the other hand, the simultaneous
J. Wang et al. Carbohydrate Polymers 205 (2019) 8–26
17
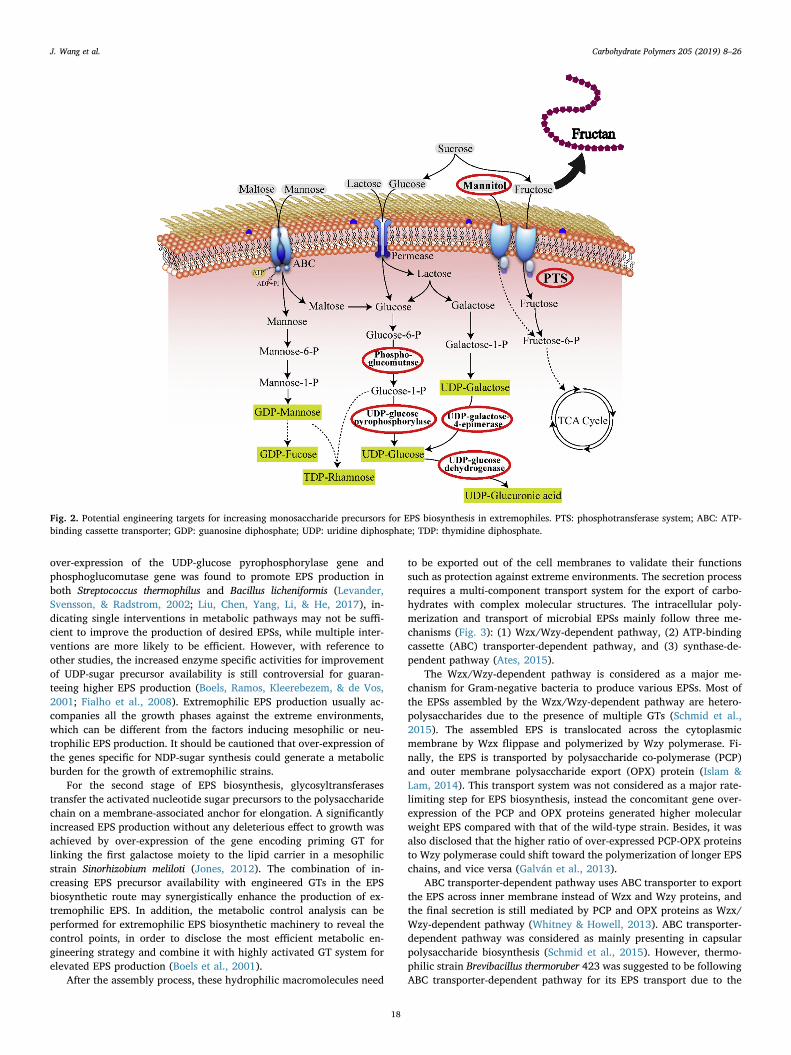
over-expression of the UDP-glucose pyrophosphorylase gene andphosphoglucomutase gene was found to promote EPS production inboth Streptococcus thermophilus and Bacillus licheniformis (Levander,Svensson, & Radstrom, 2002; Liu, Chen, Yang, Li, & He, 2017), in-dicating single interventions in metabolic pathways may not be suffi-cient to improve the production of desired EPSs, while multiple inter-ventions are more likely to be efficient. However, with reference toother studies, the increased enzyme specific activities for improvementof UDP-sugar precursor availability is still controversial for guaran-teeing higher EPS production (Boels, Ramos, Kleerebezem, & de Vos,2001; Fialho et al., 2008). Extremophilic EPS production usually ac-companies all the growth phases against the extreme environments,which can be different from the factors inducing mesophilic or neu-trophilic EPS production. It should be cautioned that over-expression ofthe genes specific for NDP-sugar synthesis could generate a metabolicburden for the growth of extremophilic strains.
For the second stage of EPS biosynthesis, glycosyltransferasestransfer the activated nucleotide sugar precursors to the polysaccharidechain on a membrane-associated anchor for elongation. A significantlyincreased EPS production without any deleterious effect to growth wasachieved by over-expression of the gene encoding priming GT forlinking the first galactose moiety to the lipid carrier in a mesophilicstrain Sinorhizobium meliloti (Jones, 2012). The combination of in-creasing EPS precursor availability with engineered GTs in the EPSbiosynthetic route may synergistically enhance the production of ex-tremophilic EPS. In addition, the metabolic control analysis can beperformed for extremophilic EPS biosynthetic machinery to reveal thecontrol points, in order to disclose the most efficient metabolic en-gineering strategy and combine it with highly activated GT system forelevated EPS production (Boels et al., 2001).
After the assembly process, these hydrophilic macromolecules need
to be exported out of the cell membranes to validate their functionssuch as protection against extreme environments. The secretion processrequires a multi-component transport system for the export of carbo-hydrates with complex molecular structures. The intracellular poly-merization and transport of microbial EPSs mainly follow three me-chanisms (Fig. 3): (1) Wzx/Wzy-dependent pathway, (2) ATP-bindingcassette (ABC) transporter-dependent pathway, and (3) synthase-de-pendent pathway (Ates, 2015).
The Wzx/Wzy-dependent pathway is considered as a major me-chanism for Gram-negative bacteria to produce various EPSs. Most ofthe EPSs assembled by the Wzx/Wzy-dependent pathway are hetero-polysaccharides due to the presence of multiple GTs (Schmid et al.,2015). The assembled EPS is translocated across the cytoplasmicmembrane by Wzx flippase and polymerized by Wzy polymerase. Fi-nally, the EPS is transported by polysaccharide co-polymerase (PCP)and outer membrane polysaccharide export (OPX) protein (Islam &Lam, 2014). This transport system was not considered as a major rate-limiting step for EPS biosynthesis, instead the concomitant gene over-expression of the PCP and OPX proteins generated higher molecularweight EPS compared with that of the wild-type strain. Besides, it wasalso disclosed that the higher ratio of over-expressed PCP-OPX proteinsto Wzy polymerase could shift toward the polymerization of longer EPSchains, and vice versa (Galván et al., 2013).
ABC transporter-dependent pathway uses ABC transporter to exportthe EPS across inner membrane instead of Wzx and Wzy proteins, andthe final secretion is still mediated by PCP and OPX proteins as Wzx/Wzy-dependent pathway (Whitney & Howell, 2013). ABC transporter-dependent pathway was considered as mainly presenting in capsularpolysaccharide biosynthesis (Schmid et al., 2015). However, thermo-philic strain Brevibacillus thermoruber 423 was suggested to be followingABC transporter-dependent pathway for its EPS transport due to the
Fig. 2. Potential engineering targets for increasing monosaccharide precursors for EPS biosynthesis in extremophiles. PTS: phosphotransferase system; ABC: ATP-binding cassette transporter; GDP: guanosine diphosphate; UDP: uridine diphosphate; TDP: thymidine diphosphate.
J. Wang et al. Carbohydrate Polymers 205 (2019) 8–26
18
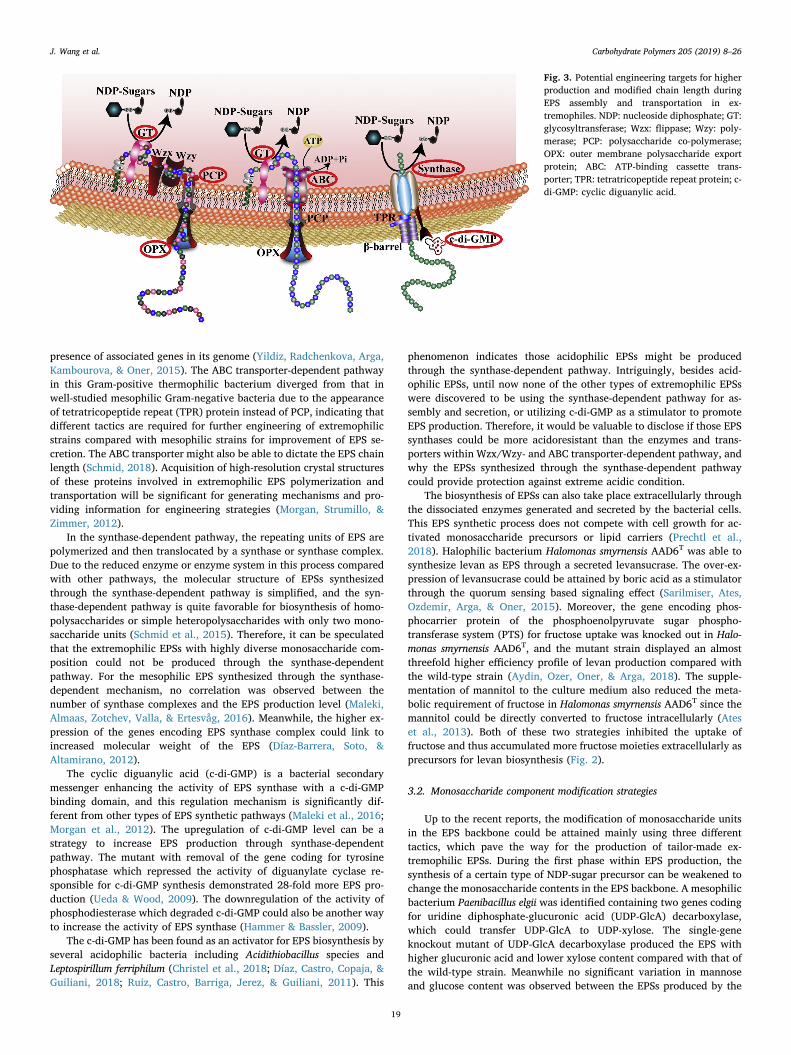
presence of associated genes in its genome (Yildiz, Radchenkova, Arga,Kambourova, & Oner, 2015). The ABC transporter-dependent pathwayin this Gram-positive thermophilic bacterium diverged from that inwell-studied mesophilic Gram-negative bacteria due to the appearanceof tetratricopeptide repeat (TPR) protein instead of PCP, indicating thatdifferent tactics are required for further engineering of extremophilicstrains compared with mesophilic strains for improvement of EPS se-cretion. The ABC transporter might also be able to dictate the EPS chainlength (Schmid, 2018). Acquisition of high-resolution crystal structuresof these proteins involved in extremophilic EPS polymerization andtransportation will be significant for generating mechanisms and pro-viding information for engineering strategies (Morgan, Strumillo, &Zimmer, 2012).
In the synthase-dependent pathway, the repeating units of EPS arepolymerized and then translocated by a synthase or synthase complex.Due to the reduced enzyme or enzyme system in this process comparedwith other pathways, the molecular structure of EPSs synthesizedthrough the synthase-dependent pathway is simplified, and the syn-thase-dependent pathway is quite favorable for biosynthesis of homo-polysaccharides or simple heteropolysaccharides with only two mono-saccharide units (Schmid et al., 2015). Therefore, it can be speculatedthat the extremophilic EPSs with highly diverse monosaccharide com-position could not be produced through the synthase-dependentpathway. For the mesophilic EPS synthesized through the synthase-dependent mechanism, no correlation was observed between thenumber of synthase complexes and the EPS production level (Maleki,Almaas, Zotchev, Valla, & Ertesvåg, 2016). Meanwhile, the higher ex-pression of the genes encoding EPS synthase complex could link toincreased molecular weight of the EPS (Díaz-Barrera, Soto, &Altamirano, 2012).
The cyclic diguanylic acid (c-di-GMP) is a bacterial secondarymessenger enhancing the activity of EPS synthase with a c-di-GMPbinding domain, and this regulation mechanism is significantly dif-ferent from other types of EPS synthetic pathways (Maleki et al., 2016;Morgan et al., 2012). The upregulation of c-di-GMP level can be astrategy to increase EPS production through synthase-dependentpathway. The mutant with removal of the gene coding for tyrosinephosphatase which repressed the activity of diguanylate cyclase re-sponsible for c-di-GMP synthesis demonstrated 28-fold more EPS pro-duction (Ueda & Wood, 2009). The downregulation of the activity ofphosphodiesterase which degraded c-di-GMP could also be another wayto increase the activity of EPS synthase (Hammer & Bassler, 2009).
The c-di-GMP has been found as an activator for EPS biosynthesis byseveral acidophilic bacteria including Acidithiobacillus species andLeptospirillum ferriphilum (Christel et al., 2018; Díaz, Castro, Copaja, &Guiliani, 2018; Ruiz, Castro, Barriga, Jerez, & Guiliani, 2011). This
phenomenon indicates those acidophilic EPSs might be producedthrough the synthase-dependent pathway. Intriguingly, besides acid-ophilic EPSs, until now none of the other types of extremophilic EPSswere discovered to be using the synthase-dependent pathway for as-sembly and secretion, or utilizing c-di-GMP as a stimulator to promoteEPS production. Therefore, it would be valuable to disclose if those EPSsynthases could be more acidoresistant than the enzymes and trans-porters within Wzx/Wzy- and ABC transporter-dependent pathway, andwhy the EPSs synthesized through the synthase-dependent pathwaycould provide protection against extreme acidic condition.
The biosynthesis of EPSs can also take place extracellularly throughthe dissociated enzymes generated and secreted by the bacterial cells.This EPS synthetic process does not compete with cell growth for ac-tivated monosaccharide precursors or lipid carriers (Prechtl et al.,2018). Halophilic bacterium Halomonas smyrnensis AAD6T was able tosynthesize levan as EPS through a secreted levansucrase. The over-ex-pression of levansucrase could be attained by boric acid as a stimulatorthrough the quorum sensing based signaling effect (Sarilmiser, Ates,Ozdemir, Arga, & Oner, 2015). Moreover, the gene encoding phos-phocarrier protein of the phosphoenolpyruvate sugar phospho-transferase system (PTS) for fructose uptake was knocked out in Halo-monas smyrnensis AAD6T, and the mutant strain displayed an almostthreefold higher efficiency profile of levan production compared withthe wild-type strain (Aydin, Ozer, Oner, & Arga, 2018). The supple-mentation of mannitol to the culture medium also reduced the meta-bolic requirement of fructose in Halomonas smyrnensis AAD6T since themannitol could be directly converted to fructose intracellularly (Ateset al., 2013). Both of these two strategies inhibited the uptake offructose and thus accumulated more fructose moieties extracellularly asprecursors for levan biosynthesis (Fig. 2).
3.2. Monosaccharide component modification strategies
Up to the recent reports, the modification of monosaccharide unitsin the EPS backbone could be attained mainly using three differenttactics, which pave the way for the production of tailor-made ex-tremophilic EPSs. During the first phase within EPS production, thesynthesis of a certain type of NDP-sugar precursor can be weakened tochange the monosaccharide contents in the EPS backbone. A mesophilicbacterium Paenibacillus elgii was identified containing two genes codingfor uridine diphosphate-glucuronic acid (UDP-GlcA) decarboxylase,which could transfer UDP-GlcA to UDP-xylose. The single-geneknockout mutant of UDP-GlcA decarboxylase produced the EPS withhigher glucuronic acid and lower xylose content compared with that ofthe wild-type strain. Meanwhile no significant variation in mannoseand glucose content was observed between the EPSs produced by the
Fig. 3. Potential engineering targets for higherproduction and modified chain length duringEPS assembly and transportation in ex-tremophiles. NDP: nucleoside diphosphate; GT:glycosyltransferase; Wzx: flippase; Wzy: poly-merase; PCP: polysaccharide co-polymerase;OPX: outer membrane polysaccharide exportprotein; ABC: ATP-binding cassette trans-porter; TPR: tetratricopeptide repeat protein; c-di-GMP: cyclic diguanylic acid.
J. Wang et al. Carbohydrate Polymers 205 (2019) 8–26
19

single-gene knockout mutant and wild-type strain (Li et al., 2015).Another genetic engineering strategy targets the genes coding for
glycosyltransferases responsible for monomer assembly. The condi-tional reduction of the activity of the priming GT involved in attachingthe first monosaccharide unit to the lipid carrier in mesophilic bac-terium Lactobacillus rhamnosus could decrease the amount of repeatingunit modules available for polymerase, leading to premature chaintermination; thus, shorter EPS chains were secreted (Bouazzaoui &LaPointe, 2006). The removal of gene encoding a non-priming GT wasable to block the addition of the undesired monosaccharide unit ontothe EPS polymer chain. In a current study concerning the EPS biosyn-thetic machinery in a mesophilic strain Paenibacillus polymyxa, thestate-of-the-art CRISPR-Cas9 genome editing tool was applied for geneknockout strategy in order to modify its EPS monomer composition.The EPS variants with altered monosaccharide distribution and rheo-logical behavior from the wild-type EPS were obtained by disruption ofthe gene coding for one of the non-priming glycosyltransferases withinits EPS biosynthetic system (Rütering et al., 2017).
The heterologous expression of the exopolysaccharide gene clusterto the recombinant strain also leads to generation of the EPS with dif-ferent monosaccharide composition from the native strain. The genecluster with functional regions coding for EPS biosynthesis regulatoryprotein, glycosyltransferases, EPS chain-length determinator, poly-merase and transporter from a thermotolerant bacterium Streptococcusthermophilus Sfi6 was expressed heterologously in a non-EPS-producingstrain Lactococcus lactis MG1363. The transferred EPS biosyntheticsystem could utilize the NDP-sugars generated in the host strainthrough its house-keeping genes as building blocks for EPS production.For the recombinant EPS, the N-acetylgalactosamine (GalNAc) moietiesin the backbone was replaced by galactose residues, since the hoststrain was unable to synthesize the corresponding UDP-GalNAc pre-cursor. Meanwhile, the recombinant EPS also lacked the galactose side-chain compared with the wild-type EPS (Stingele et al., 1999). Thisheterologous strategy requires that the polymerases and transportersfrom the native strain can recognize the recombinant EPS chain withoutstrong exclusive selectivity. Many extremophilic bacteria are able togenerate two or three EPSs with various molecular weight andmonomer distributions (Table 1). Therefore, the polymerization andexport system from the extremophiles with multi-EPS producing cap-ability may not be strictly specific towards a single type of EPS back-bone, and this infidelity provides a further degree of flexibility inproducing recombinant extremophilic EPS in an engineered strainthrough heterologous genetic engineering techniques.
3.3. Genome annotation for extremophilic EPS biosynthetic system
The genome annotation for extremophilic strains can be a powerfultool to disclose the essential genes associated with EPS biosynthesis andhence gain more insight about the biological mechanisms of EPS pro-duction, serving as the starting point to develop genetic and metabolicengineering strategies to optimize EPS production and modify EPSmonomer composition to fit industrial and biomedical requirements.Currently, a preliminary model of the EPS biosynthesis mechanism wassuccessfully proposed for a thermophilic bacterium Brevibacillus ther-moruber 423 through whole-genome analysis (Yildiz et al., 2015). Thegenome annotation of halophilic strain Halomonas smyrnensis AAD6T
revealed the presence of a Pel exopolysaccharide gene cluster in itsgenome, indicating its capacity to produce Pel EPS besides being alevan producer (Diken et al., 2015). The genome analysis was alsocarried for acidophilic bacterium Leptospirillum ferriphilum, which de-monstrated cellulose and Pel EPS synthetic genes involved in the syn-thase-dependent pathway (Christel et al., 2018). With a relativelylarger genome size, the psychrophile Phormidesmis priestleyi BC1401contained both gene clusters following the scheme of Wzx/Wzy-de-pendent and ABC transporter-dependent EPS export systems (Chrismas,Barker, Anesio, & Sánchez-Baracaldo, 2016). This is quite intriguing,
and further study for EPS characterization and transcriptomics is highlyrecommended to elucidate the EPS biosynthetic and regulatory me-chanism in this psychrophilic bacterium.
4. Recent progress in the application of extremophilic EPSs
4.1. Biomedical application
4.1.1. Antitumor and immunoregulatory effectCurrently, accumulated evidence has demonstrated that ex-
tremophilic EPSs have a broad spectrum of biological activities, such asanti-cancer, anti-oxidant and immunoregulatory properties, which canbe promising for biomedical applications. The anti-cancer efficacy hasalready been recognized for the polysaccharides generated by fungi,algae, and plants (Zong, Cao, & Wang, 2012). Extremophilic EPSs canhardly be highly cytotoxic against malignant proliferating mammaliancells like chemotherapeutic drugs. Instead, extremophilic EPSs mayinduce apoptosis in tumor cells via coupling specific surface receptors(Ruiz-Ruiz et al., 2011). The thermophilic EPSs from Geobacillus sp.TS3-9 was found to significantly inhibit the proliferation of hepatomacarcinoma cell in a dose-dependent manner in vitro (Wang et al., 2017).Further study of antiproliferation effects on non-tumor cells is requiredto identify its antitumor specificity. Furthermore, the addition of non-sugar functional groups onto extremophilic EPS molecules may gen-erate improved biological activities (e.g. antitumor activity) comparedwith native EPSs. An oversulphated EPS produced by a halophilicbacterium Halomonas stenophila B100 could specifically induce apop-tosis of leukemia cells from peripheral blood, and the addition of sul-phate moieties to native EPS was considered to enhance its anti-proliferative efficacy (Ruiz-Ruiz et al., 2011). Another halophilic EPSlevan was modified through periodate oxidation to harbor aldehydegroups, and the aldehyde-activated levan derivatives showed bothbiocompatibility to non-tumor cells and anti-cancer activity againstseveral human tumor cell lines in vitro. The antitumor efficacy wasconfirmed to be enhanced by increasing the oxidation degree of the EPS(Sarilmiser & Oner, 2014).
EPSs can also inhibit tumor progression through the im-munoenhancement effect (Zong et al., 2012). The EPS from a thermo-philic bacterium Thermus aquaticus YT-1 was proved to be an im-munomodulator which stimulated macrophage cells to produce thecytokines tumor necrosis factor alpha (TNF-α) and interleukin 6 (IL-6).This thermophilic EPS also induced macrophages to release nitric oxide(NO) as inflammatory mediator. Toll-like receptor 2 (TLR2) expressedon the immune cell surface was confirmed as the natural receptor of thisthermophilic EPS (Lin et al., 2011). A psychrophilic EPS produced byPseudoalteromonas sp. strain S-5 showed a similar stimulation effect onmacrophages to secrete TNF-α and NO. Meanwhile, it could also en-hance the phagocyte function of macrophage cells (Bai et al., 2012).The stimulation of the production of TNF-α and NO indicates a role ofthese EPSs in activation of macrophages into the M1 subtype (classi-cally activated macrophages), which can suppress tumor growth, me-tastasis and angiogenesis (Chen et al., 2018). Thus, these extremophilicEPSs have high potential for application in macrophage-mediated im-mune therapy for cancer treatment.
Several extremophilic EPSs were found to be non-cytotoxic fornormal cell lines. The heteropolysaccharide produced by thermophilicbacterium Brevibacillus thermoruber strain 423 demonstrated high bio-compatibility to a monkey kidney fibroblast cell line (Yildiz et al.,2014). Levan secreted by halophile Halomonas smyrnensis AAD6T alsoshowed high biocompatibility and affinity with non-cancerous celllines. The lack of branch structure in this halophilic levan molecule wasconsidered as crucial for the absence of cytotoxic activity (Küçükaşiket al., 2011; Poli et al., 2009). The EPSs from halothermophilic bacteriaBacillus licheniformis strain T14 and B3-15 were non-cytotoxic towardhuman peripheral blood mononuclear cells (PMBC) at maximum con-centration of 400 and 300 μg/ml, respectively (Arena et al., 2006;
J. Wang et al. Carbohydrate Polymers 205 (2019) 8–26
20

Gugliandolo, Spanò, Lentini, Arena, & Maugeri, 2013). Additionally,thermophilic EPS synthesized by Geobacillus thermodenitrificans strainB3-72 was also non-toxic to PMBC cells at concentration of 300 μg/mlor below (Arena et al., 2009). Psychrophilic EPS of Pseudoalteromonassp. S-5 had no cytotoxic effect under 500 μg/ml against murine peri-toneal macrophages (Bai et al., 2012). These non-cytotoxic ex-tremophilic EPSs are highly promising for application as a biocompa-tible carrier to conjugate antineoplastic drug and targeting ligand suchas monoclonal antibody. This three-phase antitumor drug conjugatewould attain the specific delivery of non-selective cytotoxic drugs totumor tissue with enhanced antitumor efficacy and selectivity(Asamoah-Asare, Zhang, & Chen, 2013; Dragojevic, Ryu, & Raucher,2015). The biodegradability of these extremophilic EPSs must beidentified in order to make in vivo test and future clinical trial feasible.
The immunoregulatory effect of several extremophilic EPSs alsoleads to antiviral activity. The EPSs from halothermophilic strainBacillus licheniformis strain B3-15, Bacillus licheniformis strain T14, andthermophile Geobacillus thermodenitrificans strain B3-72 decreasedherpes simplex virus type 2 (HSV-2) replication in PMBC through sti-mulating the expression of different proinflammatory cytokines in-volved in the immune surveillance toward virus infection, indicating apotential application as therapy in herpes virus infection and im-munocompromised host (Arena et al., 2006, 2009; Spanò & Arena,2016).
4.1.2. Antioxidant effectAntioxidant activity leads to scavenging reactive oxygen species
(ROS), which generate oxidative stress to neuronal cells and are deeplyassociated with chronic and degenerative diseases such as neurode-generative disorders (Xu, Bi, & Wan, 2016). Polysaccharides have beenshown to play a crucial role as natural antioxidants for the preventionof oxidative damage in the human body (Wang et al., 2013). Ex-tremophilic EPSs are usually non-pathogenic and their high biodiversityoffers various biotechnological activities including antioxidancy. Ha-lophilic EPS isolated from Halolactibacillus miurensis, and thermophilicEPS from Geobacillus sp. strain TS3-9 both demonstrated dose-depen-dent scavenging activity against DPPH (2,2-diphenyl-1-picrylhydrazyl),hydroxyl and superoxide free radicals (Arun et al., 2017; Wang et al.,2017). Additionally, the EPS produced by halothermophilic bacteriumHalomonas nitroreducens WB1 had antioxidant properties to scavengehydroxyl and DPPH radicals (Chikkanna, Ghosh, & Kishore, 2018). Ahyper-branched psychrophilic EPS secreted by Polaribacter sp. SM1127showed substantially higher antioxidant activity than that of hyaluronicacid, an industrial annexing agent for scavenging radicals (Sun et al.,2015). These extremophilic EPSs with remarkable antioxidative capa-city may be efficacious in the treatment of neurodegenerative diseases.Furthermore, they can be studied as natural dietary antioxidants for theinactivation of oxyradicals and reduction of the incidence rate of neu-rodegenerative disease.
4.2. Food application
The microbial polysaccharides which are able to stabilize emulsionsbetween water and hydrophobic compounds have potential as naturalemulsifiers in the food industry (Freitas et al., 2009). Bioemulsifiershave the advantages of biodegradability, low toxicity, selectivity andenvironmental compatibility over artificial products (Mata et al., 2008).The emulsifying activity has been found to be common in extremophilicEPSs, including those produced by thermophiles, psychrophiles, halo-philes, and alkaliphiles; and they are all heteropolysaccharides(Table 1). Among those extremophilic EPSs, EPS from psychrophilicbacterium Pseudomonas sp. ID1 had a higher emulsifying activity thanxanthan gum and arabic gum for several food oils (Carrión, Delgado, &Mercade, 2015).
In industrial processes, emulsifiers may be exposed to extremes oftemperature, environmental pH and salinity (Freitas et al., 2009). An
important property of bioemulsifiers from extremophiles is their highemulsion stability over a wide range of temperature, pH and salinity(Zheng et al., 2012). For example, in Arias’ report, neither the viscositynor the pseudoplasticity of mauran (a halophilic EPS) solution was af-fected by the presence of salts, sugar, surfactants, lactic acid, changes inpH, or freezing and thawing (Arias et al., 2003). The minor content ofprotein in those extremophilic EPSs might be essential for the emulsi-fying activity (Llamas, Amjres, Mata, Quesada, & Béjar, 2012). Thepresence of uronic acid and acetyl group in EPS was also considered tocontribute to its emulsifying capacity (Caruso et al., 2017; Jain, Mody,Mishra, & Jha, 2012; Mata et al., 2006).
Pseudoplastic rheological behavior is another common propertyamong those extremophilic EPSs with emulsifying activity (Table 1).High pseudoplasticity is an attractive rheological characteristic in di-verse types of food formulations, such as sauce, dairy, cake, saladdressing, syrup, and pudding (Bahram Parvar & Razavi Seyed, 2012;Han et al., 2014). For the development of foodstuffs, the pseudoplasticproperty of EPS is advantageous to generate comfortable sensoryproperties such as mouth feel and flavor release. It is also useful for foodprocesses, such as mixing, pouring and pumping with different opera-tive shear rates (Han et al., 2014).
Based on current extremophilic EPS research, it remains difficult forthe newly discovered extremophilic EPSs to displace the commercia-lized biopolymers such as xanthan and gellan in the food industry, dueto the costly production processes of extremophilic EPSs using puresugar as substrate. Inexpensive and renewable substrates need to beused for EPS production, such as molasses which can be a feasiblesubstitute for sucrose (Küçükaşik et al., 2011; Sam et al., 2011). Acontinuous cultivation technique with economic potential for industrialscale production also showed a much higher efficiency for the fer-mentation of extremophilic EPS in comparison with batch cultures(Radchenkova et al., 2015).
4.3. Biomaterial application
4.3.1. EPS-based nanoparticlesNanotechnology is an indispensable discipline in the modern
pharmaceutical field for a variety of applications. The non-ideal bio-degradability and biocompatibility of the synthetic nanoparticlesusually impair renal excretion and induce many unacceptable side ef-fects. During the constant lookout for novel products with improvedpharmaceutical functions, the extremophilic EPSs have received in-creasing attention as alternatives to synthetic polymers in the produc-tion and modification of nanoparticles. Their valuable biological char-acteristics and simplicity of chemical modifications make them strongcandidates for nanoparticle application (Raveendran et al., 2014).
Extremophilic EPSs can be applied to nanoparticle technology intwo ways: one is to utilize EPS to form nanoparticles directly, and theother is to use EPS to encapsulate nanoparticles made from anothermaterial. The extremophilic EPSs with negatively charged groups couldperform as polyelectrolytes, which allow them to self-assemble withpositively charged biomolecules to form biodegradable nanoparticlesthrough polyelectrolyte complexation. As a positively charged biopo-lymer, chitosan is often used for polyelectrolyte nanoparticle formationwith negatively charged EPSs (Deepak, Pandian, Sivasubramaniam,Nellaiah, & Sundar, 2015; Karlapudi et al., 2016). Halophilic EPSmauran-chitosan hybrid nanoparticles, produced through an ionic-ge-lation method, manifested stable drug release and biocompatibilitywhen used for antitumor drug encapsulation (Raveendran, Pouloseet al., 2013). Another halophilic EPS levan, produced by Halomonassmyrnensis AAD6T, was investigated for nanoparticle formation throughself-assembly, and its suitability was affirmed as a nanocarrier for de-livery of peptides and proteins (Sezer, Kazak, Öner, & Akbuğa, 2011).
Application of EPSs in the coating or stabilization of chemicallysynthetic nanoparticles also has important potential. Some syntheticnanoparticles possess irreplaceable properties, such as a
J. Wang et al. Carbohydrate Polymers 205 (2019) 8–26
21

photoluminescence for imaging, but their toxicity in the human bodyneeds to be minimized through suitable modification. Polysaccharidecoated nanoparticles have already been shown to possess rapid uptakeand internalization through the endocytosis effect compared with un-coated nanoparticles, and the cellular toxicity of EPS-coated nano-particles was also notably reduced (Banerjee & Bandopadhyay, 2016).Extremophilic EPSs can therefore be applied as a passivation agent toimprove the biocompatibility of nanoparticles, and make them morefeasible for pharmaceutical applications (Raveendran et al., 2014). Forinstance, quantum dots (QDs) are nanocrystals with a photoluminescentproperty and applied as preferred imaging agents in biological tissuesfor clinical diagnose (Deepagan et al., 2012; Raveendran et al., 2014).However, QDs such as ZnS nanocrystals are synthesized from toxicchemicals in order to maintain their imaging property. These types ofQDs are hydrophobic and water-insoluble, which hinder their applica-tion in the medical field. One solution to overcome these drawbacks isto stabilize QDs with a capping agent, and extremophilic EPSs can be astabilizing agent for QDs to improve their cellular acceptance. In astudy, the stabilization of ZnS-Mn QDs using halophilic EPS mauranwas highly successful in imparting a biocompatible and safe mode ofcellular imaging under in vitro conditions. Anionic EPSs are able to bindwith nanoparticles having positive charge. Additionally, the acetylgroups present in EPSs can bring more positively charged ions to thevicinity of binding sites, thus allowing stronger binding (Raveendranet al., 2014).
4.3.2. EPS-based filmsAdhesive and biocompatible films are an attractive way to offer
fixation to tissues either externally, for wound healing, or internally asa surgical sealant. For the purpose of medical adhesives, such as a burndressing, drug delivery, and implantation, the films are required to keepgood long-term performance on skin or in biological fluids withouttriggering a pathological process (Costa et al., 2013). The inherentfunctions of extremophilic EPSs are to provide adhesion and protectionto bacteria in an extreme environment. Therefore, naturally derivedEPS-based films confer sufficient cohesive strength and maintain abiocompatible response to cells and tissues. The negative charge allowsEPSs to be adsorbed by electrostatic self-assembly and sequential for-mation onto multilayer film. The EPS levan produced by halophilicbacterium Halomonas smyrnensis AAD6T was applied by electrostaticadsorption to construct multilayer film, which demonstrated a pro-mising enhancement of live cell adhesive property. The extremophilicEPS-based film surface, having better biocompatibility, provides a va-luable means to explore novel types of cell-material interactions,leading to an understanding of how to promote or inhibit specific cel-lular responses when contacting bio-based materials (Costa et al.,2013). Moreover, the halophilic EPS levan was mixed with chitosan andpolyethylene oxide (PEO) to produce hybrid films through a solventcasting method. This ternary blend film showed better biocompatiblebehavior compared with chitosan-PEO binary film (Bostan et al., 2014).Another method applied for thin film production using halophilic EPSlevan was matrix-assisted pulsed laser evaporation (MAPLE). This na-nostructured film was also able to sustain cell adhesion and prolifera-tion (Sima et al., 2011).
4.3.3. EPS-based materials through electrospinningElectrospinning is a versatile and relatively cost-effective technique
to fabricate a large variety of soluble or fusible synthetic and naturalpolymers into continuous fibers with diameters in the submicron tonanometer range (Salem, 2007). The electrospinning method has al-ready been applied in many technological fields, and it enables theproduction of novel biomaterials using naturally occurring biopolymerswith complex molecular structures (Torres-Giner, Ocio, & Lagaron,2008). Due to the natural properties of extremophilic EPSs, the mate-rials electrospun from those EPSs are considered sustainable, bio-compatible, biodegradable, and non-toxic. Additionally, the high water-
solubility of EPSs avoids using toxic solvents or additives during elec-trospinning process, making EPS-based electrospun materials excellentcandidates for biomedical engineering applications. However, it can bedifficult to generate neat EPS fibers by electrospinning, since EPS so-lutions tend to have high surface tension, non-ideal viscosity, and ex-cessively strong charge density due to the anionic nature of EPSs(Santos et al., 2014; Torres-Giner et al., 2008). The addition of a hy-drophilic co-polymer as a carrier agent is one way to circumvent thelimitations from those bio-polyelectrolyte (e.g. EPS) solutions(Vashisth, Pruthi, Singh, & Pruthi, 2014). For example, polyvinyl al-cohol (PVA), a biocompatible and water-soluble polymer, can beblended with EPS, reducing the repulsive force from the negativelycharged EPS solution and allowing the generation of uniform nanofi-bers by electrospinning (Qian et al., 2016; Santos et al., 2014; Vashisthet al., 2014). The halophilic EPS mauran was blended with PVA andelectrospun to generate a scaffold with continuous, uniform nanofibers.The mauran-based nanofiber was able to boost cellular adhesion, mi-gration, proliferation, and differentiation of mammalian cells in vitro.The polyanionic nature of extremophilic EPSs increases the negativecharge accumulation on the surface of the scaffold, which is helpful forprotein adsorption and the ability to enhance cellular attachment. Anexcellent property of mauran is that it can keep the same viscosityunder a high concentration of salt and sugar, or under extreme pHvalues; and the stable viscosity of a mauran solution is highly ad-vantageous in obtaining stable electrospinning conditions (Raveendran,Dhandayuthapani et al., 2013). In future studies, those extremophilicEPSs with heavy metal adsorption capability may also be blended withPVA and then electrospun onto a basal microfiltration membrane forwater filtration applications (Santos et al., 2014).
5. Conclusion and prospective
It is now widely considered that extremophilic microorganismsprovide a valuable resource, not only for the elucidation of bioprocessesin extreme environments, but also for the exopolysaccharides theyproduce, which have a valuable range of physicochemical propertiesand highly promising commercial applications. To realize the full valueof these biopolymers, it will be necessary to gain insight into themodulation of EPS biosynthesis in extremophiles. The genome anno-tation and construction of EPS biosynthetic pathways will be of sig-nificant importance in determining the kind of monosaccharide unitsthat can be incorporated into the structures of the EPSs, and how theyare incorporated; as well as how the compositions of monosaccharidesin the EPSs can be affected. Metabolic and genetic engineering willenable the development of effective strategies to successfully enhanceEPS production and engineer EPS properties via tailoring their chemicalcomposition and structure. Applications of extremophilic EPSs in bio-medical, food and pollution-mitigation products can be foreseen basedon their known biodegradability, biocompatibility, non-toxicity, andchemical functionality. It can be anticipated that more extremophilicEPSs from different harsh environments will be discovered, character-ized, and optimized via evolving bioengineering protocols, and thatthey will be employed in high value-added industries with potential forstrong, sustainable growth.
Acknowledgements
This research was supported by the National Science Foundation inthe form of BuG ReMeDEE initiative (Award # 1736255). Authors alsoacknowledge the financial support in the form of CNAM/Bio Centreprovided by the South Dakota Governor’s Office of EconomicDevelopment. Research support from the Department of Chemical andBiological Engineering at the South Dakota School of Mines andTechnology is gratefully acknowledged.
J. Wang et al. Carbohydrate Polymers 205 (2019) 8–26
22

References
Arena, A., Gugliandolo, C., Stassi, G., Pavone, B., Iannello, D., Bisignano, G., ... Maugeri,T. L. (2009). An exopolysaccharide produced by Geobacillus thermodenitrificans strainB3-72: Antiviral activity on immunocompetent cells. Immunology Letters, 123(2),132–137.
Arena, A., Maugeri, T. L., Pavone, B., Iannello, D., Gugliandolo, C., & Bisignano, G.(2006). Antiviral and immunoregulatory effect of a novel exopolysaccharide from amarine thermotolerant Bacillus licheniformis. International Immunopharmacology, 6(1),8–13.
Arias, S., del Moral, A., Ferrer, M. R., Tallon, R., Quesada, E., & Bejar, V. (2003). Mauran,an exopolysaccharide produced by the halophilic bacterium Halomonas maura, with anovel composition and interesting properties for biotechnology. Extremophiles, 7(4),319–326.
Arun, J., Selvakumar, S., Sathishkumar, R., Moovendhan, M., Ananthan, G., Maruthiah,T., ... Palavesam, A. (2017). In vitro antioxidant activities of an exopolysaccharidefrom a salt pan bacterium Halolactibacillus miurensis. Carbohydrate Polymers, 155,400–406.
Asamoah-Asare, J., Zhang, Y., & Chen, Y. (2013). Antibody conjugated polymeric pro-drugs the future for cancer therapy. International Journal of Advanced Biotechnologyand Bioengineering, 1(1), 1–17.
Ates, O. (2015). Systems biology of microbial exopolysaccharides production. Frontiers inBioengineering and Biotechnology, 3, 200.
Ates, O., Arga, K. Y., & Oner, E. T. (2013). The stimulatory effect of mannitol on levanbiosynthesis: Lessons from metabolic systems analysis of Halomonas smyrnensisAAD6T. Biotechnology Progress, 29(6), 1386–1397.
Aydin, B., Ozer, T., Oner, E. T., & Arga, K. Y. (2018). The genome-based metabolic sys-tems engineering to boost levan production in a halophilic bacterial model. OMICS: AJournal of Integrative Biology, 22(3), 198–209.
Bahram Parvar, M., & Razavi Seyed, M. A. (2012). Rheological interactions of selectedhydrocolloid–sugar–milk–emulsifier systems. International Journal of Food Science &Technology, 47(4), 854–860.
Bai, Y., Zhang, P., Chen, G., Cao, J., Huang, T., & Chen, K. (2012). Macrophage im-munomodulatory activity of extracellular polysaccharide (PEP) of Antarctic bac-terium Pseudoaltermonas sp.S-5. International Immunopharmacology, 12(4), 611–617.
Baker-Austin, C., & Dopson, M. (2007). Life in acid: pH homeostasis in acidophiles. Trendsin Microbiology, 15(4), 165–171.
Banerjee, A., & Bandopadhyay, R. (2016). Use of dextran nanoparticle: A paradigm shiftin bacterial exopolysaccharide based biomedical applications. International Journal ofBiological Macromolecules, 87, 295–301.
Bejar, V., Calvo, C., Moliz, J., Diaz-Martinez, F., & Quesada, E. (1996). Effect of growthconditions on the rheological properties and chemical composition of Volcaniellaeurihalina exopolysaccharide. Applied Biochemistry and Biotechnology, 59(1), 77–86.
Béjar, V., Llamas, I., Calvo, C., & Quesada, E. (1998). Characterization of exopoly-saccharides produced by 19 halophilic strains of the species Halomonas eurihalina.Journal of Biotechnology, 61(2), 135–141.
Bhalla, A., Bansal, N., Kumar, S., Bischoff, K. M., & Sani, R. K. (2013). Improved lig-nocellulose conversion to biofuels with thermophilic bacteria and thermostable en-zymes. Bioresource Technology, 128, 751–759.
Boels, I. C., Ramos, A., Kleerebezem, M., & de Vos, W. M. (2001). Functional analysis ofthe Lactococcus lactis galU and galE genes and their impact on sugar nucleotide andexopolysaccharide biosynthesis. Applied and environmental microbiology, 67(7),3033–3040.
Bostan, M. S., Mutlu, E. C., Kazak, H., Sinan Keskin, S., Oner, E. T., & Eroglu, M. S. (2014).Comprehensive characterization of chitosan/PEO/levan ternary blend films.Carbohydrate Polymers, 102, 993–1000.
Bouazzaoui, K., & LaPointe, G. (2006). Use of antisense RNA to modulate glycosyl-transferase gene expression and exopolysaccharide molecular mass in Lactobacillusrhamnosus. Journal of Microbiological Methods, 65(2), 216–225.
Brian-Jaisson, F., Molmeret, M., Fahs, A., Guentas-Dombrowsky, L., Culioli, G., Blache, Y.,... Ortalo-Magné, A. (2016). Characterization and anti-biofilm activity of extracellularpolymeric substances produced by the marine biofilm-forming bacteriumPseudoalteromonas ulvae strain TC14. Biofouling, 32(5), 547–560.
Carrión, O., Delgado, L., & Mercade, E. (2015). New emulsifying and cryoprotectiveexopolysaccharide from Antarctic Pseudomonas sp. ID1. Carbohydrate Polymers, 117,1028–1034.
Caruso, C., Rizzo, C., Mangano, S., Poli, A., Di Donato, P., Finore, I., ... Giudice, A. L.(2017). Production and biotechnological potentialities of extracellular polymericsubstances from sponge-associated Antarctic bacteria. Applied and environmental mi-crobiology, 84(4), e01624–01617.
Caruso, C., Rizzo, C., Mangano, S., Poli, A., Di Donato, P., Nicolaus, B., ... Lo Giudice, A.(2018). Extracellular polymeric substances with metal adsorption capacity producedby Pseudoalteromonas sp. MER144 from Antarctic seawater. Environmental Science andPollution Research, 25(5), 4667–4677.
Casillo, A., Parrilli, E., Sannino, F., Mitchell, D. E., Gibson, M. I., Marino, G., ... Corsaro,M. M. (2017). Structure-activity relationship of the exopolysaccharide from a psy-chrophilic bacterium: A strategy for cryoprotection. Carbohydrate Polymers, 156,364–371.
Casillo, A., Ståhle, J., Parrilli, E., Sannino, F., Mitchell, D. E., Pieretti, G., ... Corsaro, M. M.(2017). Structural characterization of an all-aminosugar-containing capsular poly-saccharide from Colwellia psychrerythraea 34H. Antonie van Leeuwenhoek, 110(11),1377–1387.
Chawla, P. R., Bajaj, I. B., Survase, S. A., & Singhal, R. S. (2009). Microbial cellulose:Fermentative production and applications. Food Technology & Biotechnology, 47(2),107–124.
Chen, M., Li, Y., Liu, Z., Qu, Y., Zhang, H., Li, D., ... Liu, M. (2018). Exopolysaccharidesfrom a Codonopsis pilosula endophyte activate macrophages and inhibit cancer cellproliferation and migration. Thoracic Cancer, 9(5), 630–639.
Chikkanna, A., Ghosh, D., & Kishore, A. (2018). Expression and characterization of apotential exopolysaccharide from a newly isolated halophilic thermotolerant bacteriaHalomonas nitroreducens strain WB1. PeerJ, 6, e4684.
Chrismas, N. A. M., Barker, G., Anesio, A. M., & Sánchez-Baracaldo, P. (2016). Genomicmechanisms for cold tolerance and production of exopolysaccharides in the Arcticcyanobacterium Phormidesmis priestleyi BC1401. BMC Genomics, 17(1), 533.
Christel, S., Herold, M., Bellenberg, S., El Hajjami, M., Buetti-Dinh, A., Pivkin, I. V., ...Dopson, M. (2018). Multi-omics reveals the lifestyle of the acidophilic, mineral-oxi-dizing model species Leptospirillum ferriphilumT. Applied and environmental micro-biology, 84(3), e02091–02017.
Christensen, B. E., Kjosbakken, J., & Smidsrød, O. (1985). Partial chemical and physicalcharacterization of two extracellular polysaccharides produced by marine, periphyticPseudomonas sp. Strain NCMB 2021. Applied and environmental microbiology, 50(4),837–845.
Corsaro, M. M., Grant, W. D., Grant, S., Marciano, C. E., & Parrilli, M. (1999). Structuredetermination of an exopolysaccharide from an alkaliphilic bacterium closely relatedto Bacillus spp. European Journal of Biochemistry, 264(2), 554–561.
Corsaro, M. M., Lanzetta, R., Parrilli, E., Parrilli, M., Tutino, M. L., & Ummarino, S.(2004). Influence of growth temperature on lipid and phosphate contents of surfacepolysaccharides from the Antarctic bacterium Pseudoalteromonas haloplanktis TAC125. Journal of bacteriology, 186(1), 29–34.
Costa, R. R., Neto, A. I., Calgeris, I., Correia, C. R., Pinho, A. C. M., Fonseca, J., ... Mano, J.F. (2013). Adhesive nanostructured multilayer films using a bacterial exopoly-saccharide for biomedical applications. Journal of Materials Chemistry B, 1(18),2367–2374.
d’Hugues, P., Joulian, C., Spolaore, P., Michel, C., Garrido, F., & Morin, D. (2008).Continuous bioleaching of a pyrite concentrate in stirred reactors: Population dy-namics and exopolysaccharide production vs. bioleaching performance.Hydrometallurgy, 94(1-4), 34–41.
Deepagan, V. G., Sarmento, B., Menon, D., Nascimento, A., Jayasree, A.,Sreeranganathan, M., ... Rangasamy, J. (2012). In vitro targeted imaging and deliveryof camptothecin using cetuximab-conjugated multifunctional PLGA-ZnS nano-particles. Nanomedicine, 7(4), 507–519.
Deepak, V., Pandian, S. R. K., Sivasubramaniam, S. D., Nellaiah, H., & Sundar, K. (2015).Synthesis of polyelectrolyte nanoparticles from anticancer exopolysaccharide isolatedfrom probiotic Lactobacillus acidophilus. Research Journal of Microbiology, 10(5),193–204.
Díaz, M., Castro, M., Copaja, S., & Guiliani, N. (2018). Biofilm formation by the acid-ophile bacterium Acidithiobacillus thiooxidans involves c-di-GMP pathway and Pelexopolysaccharide. Genes, 9(2), 113.
Díaz-Barrera, A., Soto, E., & Altamirano, C. (2012). Alginate production and alg8 geneexpression by Azotobacter vinelandii in continuous cultures. Journal of IndustrialMicrobiology & Biotechnology, 39(4), 613–621.
Diken, E., Ozer, T., Arikan, M., Emrence, Z., Oner, E. T., Ustek, D., ... Arga, K. Y. (2015).Genomic analysis reveals the biotechnological and industrial potential of levan pro-ducing halophilic extremophile, Halomonas smyrnensis AAD6T. SpringerPlus, 4(1),393.
Donot, F., Fontana, A., Baccou, J. C., & Schorr-Galindo, S. (2012). Microbial exopoly-saccharides: Main examples of synthesis, excretion, genetics and extraction.Carbohydrate Polymers, 87(2), 951–962.
Dopson, M., Baker-Austin, C., Koppineedi, P. R., & Bond, P. L. (2003). Growth in sulfidicmineral environments: Metal resistance mechanisms in acidophilic micro-organisms.Microbiology, 149(Pt 8), 1959–1970.
Dragojevic, S., Ryu, S. J., & Raucher, D. (2015). Polymer-based prodrugs: Improvingtumor targeting and the solubility of small molecule drugs in cancer therapy.Molecules, 20(12), 21750–21769.
Drouillard, S., Jeacomine, I., Buon, L., Boisset, C., Courtois, A., Thollas, B., ... Helbert, W.(2015). Structure of an amino acid-decorated exopolysaccharide secreted by a Vibrioalginolyticus strain. Marine Drugs, 13(11), 6723–6739.
Duckworth, A., Grant, W., Jones, B., & Van Steenbergen, R. (1996). Phylogenetic diversityof soda lake alkaliphiles. FEMS microbiology ecology, 19(3), 181–191.
Ewert, M., & Deming, J. W. (2013). Sea ice microorganisms: Environmental constraintsand extracellular responses. Biology, 2(2), 603–628.
Fialho, A. M., Moreira, L. M., Granja, A. T., Popescu, A. O., Hoffmann, K., & Sa-Correia, I.(2008). Occurrence, production, and applications of gellan: Current state and per-spectives. Applied Microbiology and Biotechnology, 79(6), 889–900.
Flemming, H. C., & Wingender, J. (2010). The biofilm matrix. Nature ReviewsMicrobiology, 8(9), 623–633.
Freitas, F., Alves, V. D., Carvalheira, M., Costa, N., Oliveira, R., & Reis, M. A. M. (2009).Emulsifying behaviour and rheological properties of the extracellular polysaccharideproduced by Pseudomonas oleovorans grown on glycerol byproduct. CarbohydratePolymers, 78(3), 549–556.
Freitas, F., Alves, V. D., & Reis, M. A. (2011). Advances in bacterial exopolysaccharides:From production to biotechnological applications. Trends in Biotechnology, 29(8),388–398.
Galván, E. M., Ielmini, M. V., Patel, Y. N., Bianco, M. I., Franceschini, E. A., Schneider, J.C., ... Ielpi, L. (2013). Xanthan chain length is modulated by increasing the avail-ability of the polysaccharide copolymerase protein GumC and the outer membranepolysaccharide export protein GumB. Glycobiology, 23(2), 259–272.
Gehrke, T., Telegdi, J., Thierry, D., & Sand, W. (1998). Importance of extracellularpolymeric substances from Thiobacillus ferrooxidans for bioleaching. Applied and en-vironmental microbiology, 64(7), 2743–2747.
Gu, D., Jiao, Y., Wu, J., Liu, Z., & Chen, Q. (2017). Optimization of EPS production and
J. Wang et al. Carbohydrate Polymers 205 (2019) 8–26
23

characterization by a halophilic bacterium, Kocuria rosea ZJUQH from Chaka SaltLake with response surface methodology. Molecules, 22(5), 814.
Guezennec, J. (2002). Deep-sea hydrothermal vents: A new source of innovative bacterialexopolysaccharides of biotechnological interest? Journal of Industrial Microbiology andBiotechnology, 29(4), 204–208.
Gugliandolo, C., Lentini, V., Spanò, A., & Maugeri, T. L. (2012). New bacilli from shallowhydrothermal vents of Panarea Island (Italy) and their biotechnological potential.Journal of Applied Microbiology, 112(6), 1102–1112.
Gugliandolo, C., Spanò, A., Lentini, V., Arena, A., & Maugeri, T. L. (2013). Antiviral andimmunomodulatory effects of a novel bacterial exopolysaccharide of shallow marinevent origin. Journal of Applied Microbiology, 116(4), 1028–1034.
Hammer, B. K., & Bassler, B. L. (2009). Distinct sensory pathways in Vibrio cholerae El Torand classical biotypes modulate cyclic dimeric GMP levels to control biofilm for-mation. Journal of bacteriology, 191(1), 169–177.
Han, P.-p., Sun, Y., Wu, X.-y., Yuan, Y.-j., Dai, Y.-j., & Jia, S.-r. (2014). Emulsifying,flocculating, and physicochemical properties of exopolysaccharide produced by cy-anobacterium Nostoc flagelliforme. Applied Biochemistry and Biotechnology, 172(1),36–49.
Horikoshi, K. (1999). Alkaliphiles: Some applications of their products for biotechnology.Microbiology and molecular biology reviews, 63(4), 735–750.
Huang, H., Li, X., Wu, M., Wang, S., Li, G., & Ma, T. (2013). Cloning, expression andcharacterization of a phosphoglucomutase/phosphomannomutase from sphingan-producing Sphingomonas sanxanigenens. Biotechnology Letters, 35(8), 1265–1270.
Huston, A. L., Methe, B., & Deming, J. W. (2004). Purification, characterization, andsequencing of an extracellular cold-active aminopeptidase produced by marine psy-chrophile Colwellia psychrerythraea strain 34H. Applied and environmental micro-biology, 70(6), 3321–3328.
Islam, S. T., & Lam, J. S. (2014). Synthesis of bacterial polysaccharides via the Wzx/Wzy-dependent pathway. Canadian Journal of Microbiology, 60(11), 697–716.
Ito, S., Kobayashi, T., Ara, K., Ozaki, K., Kawai, S., & Hatada, Y. (1998). Alkaline de-tergent enzymes from alkaliphiles: Enzymatic properties, genetics, and structures.Extremophiles, 2(3), 185–190.
Jain, R. M., Mody, K., Mishra, A., & Jha, B. (2012). Isolation and structural character-ization of biosurfactant produced by an alkaliphilic bacterium Cronobacter sakazakiiisolated from oil contaminated wastewater. Carbohydrate Polymers, 87(3),2320–2326.
Jiang, P., Li, J., Han, F., Duan, G., Lu, X., Gu, Y., ... Yu, W. (2011). Antibiofilm activity ofan exopolysaccharide from marine bacterium Vibrio sp. QY101. PLOS ONE, 6(4),e18514.
Jiao, Y., Cody, G. D., Harding, A. K., Wilmes, P., Schrenk, M., Wheeler, K. E., ... Thelen, M.P. (2010). Characterization of extracellular polymeric substances from acidophilicmicrobial biofilms. Applied and environmental microbiology, 76(9), 2916–2922.
Johnson, D. B. (1998). Biodiversity and ecology of acidophilic microorganisms. FEMSmicrobiology ecology, 27(4), 307–317.
Johnson, D. B., Joulian, C., d’Hugues, P., & Hallberg, K. B. (2008). Sulfobacillus benefacienssp. nov., an acidophilic facultative anaerobic firmicute isolated from mineral bio-leaching operations. Extremophiles, 12(6), 789.
Jones, K. M. (2012). Increased production of the exopolysaccharide succinoglycan en-hances Sinorhizobium meliloti 1021 symbiosis with the host plant Medicago truncatula.Journal of bacteriology, 194(16), 4322–4331.
Junge, K., Eicken, H., Swanson, B. D., & Deming, J. W. (2006). Bacterial incorporation ofleucine into protein down to -20 degrees C with evidence for potential activity in sub-eutectic saline ice formations. Cryobiology, 52(3), 417–429.
Kambourova, M., Mandeva, R., Dimova, D., Poli, A., Nicolaus, B., & Tommonaro, G.(2009). Production and characterization of a microbial glucan, synthesized byGeobacillus tepidamans V264 isolated from Bulgarian hot spring. CarbohydratePolymers, 77(2), 338–343.
Karlapudi, A. P., Kodali, V. P., Kota, K. P., Shaik, S. S., Sampath Kumar, N. S., & Dirisala,V. R. (2016). Deciphering the effect of novel bacterial exopolysaccharide-based na-noparticle cream against Propionibacterium acnes. 3. Biotech, 6(1), 35.
Kim, S. J., Kim, B.-G., Park, H. J., & Yim, J. H. (2016). Cryoprotective properties andpreliminary characterization of exopolysaccharide (P-Arcpo 15) produced by theArctic bacterium Pseudoalteromonas elyakovii Arcpo 15. Preparative Biochemistry andBiotechnology, 46(3), 261–266.
Küçükaşik, F., Kazak, H., Güney, D., Finore, I., Poli, A., Yenigün, O., ... Öner, E. T. (2011).Molasses as fermentation substrate for levan production by Halomonas sp. AppliedMicrobiology and Biotechnology, 89(6), 1729–1740.
Le Costaouëc, T., Cérantola, S., Ropartz, D., Ratiskol, J., Sinquin, C., Colliec-Jouault, S., ...Boisset, C. (2012). Structural data on a bacterial exopolysaccharide produced by adeep-sea Alteromonas macleodii strain. Carbohydrate Polymers, 90(1), 49–59.
Lelchat, F., Cérantola, S., Brandily, C., Colliec-Jouault, S., Baudoux, A.-C., Ojima, T., ...Boisset, C. (2015). The marine bacteria Cobetia marina DSMZ 4741 synthesizes anunexpected K-antigen-like exopolysaccharide. Carbohydrate Polymers, 124, 347–356.
Levander, F., Svensson, M., & Radstrom, P. (2002). Enhanced exopolysaccharide pro-duction by metabolic engineering of Streptococcus thermophilus. Applied and environ-mental microbiology, 68(2), 784–790.
Li, H., Xu, H., Li, S., Xu, H., Guo, C., Ying, H., ... Ouyang, P. (2010). Strain improvementand metabolic flux modeling of wild-type and mutant Alcaligenes sp. NX-3 forsynthesis of exopolysaccharide welan gum. Biotechnology and Bioprocess Engineering,15(5), 777–784.
Li, O., Qian, C., Zheng, D., Wang, P., Liu, Y., Jiang, X., ... Wu, X. (2015). Two UDP-glucuronic acid decarboxylases involved in the biosynthesis of a bacterial exopoly-saccharide in Paenibacillus elgii. Applied Microbiology and Biotechnology, 99(7),3127–3139.
Li, P., Liu, Z., & Xu, R. (2001). Chemical characterisation of the released polysaccharidefrom the cyanobacterium Aphanothece halophytica GR02. Journal of Applied Phycology,
13(1), 71–77.Li, W., Zhou, W., Zhang, Y., Wang, J., & Zhu, X. (2008). Flocculation behavior and me-
chanism of an exopolysaccharide from the deep-sea psychrophilic bacteriumPseudoalteromonas sp. SM9913. Bioresource Technology, 99(15), 6893–6899.
Lin, L., & Xu, J. (2013). Dissecting and engineering metabolic and regulatory networks ofthermophilic bacteria for biofuel production. Biotechnology Advances, 31(6),827–837.
Lin, M., Yang, Y., Chen, Y., Hua, K., Lu, C., Sheu, F., ... Wu, S. (2011). A novel exopo-lysaccharide from the biofilm of Thermus aquaticus YT-1 induces the immune re-sponse through Toll-like receptor 2. Journal of Biological Chemistry, 286(20),17736–17745.
Liu, P., Chen, Z., Yang, L., Li, Q., & He, N. (2017). Increasing the bioflocculant productionand identifying the effect of overexpressing epsB on the synthesis of polysaccharideand γ-PGA in Bacillus licheniformis. Microbial Cell Factories, 16(1), 163.
Liu, S., Chen, X., He, H., Zhang, X., Xie, B., Yu, Y., ... Zhang, Y. (2013). Structure andecological roles of a novel exopolysaccharide from the Arctic sea ice bacteriumPseudoalteromonas sp. strain SM20310. Applied and environmental microbiology, 79(1),224–230.
Llamas, I., Amjres, H., Mata, J. A., Quesada, E., & Béjar, V. (2012). The potential bio-technological applications of the exopolysaccharide produced by the halophilicbacterium Halomonas almeriensis. Molecules, 17(6), 7103–7120.
Llamas, I., Mata, J. A., Tallon, R., Bressollier, P., Urdaci, M. C., Quesada, E., ... Béjar, V.(2010). Characterization of the exopolysaccharide produced by Salipiger mucosus A3T,a halophilic species belonging to the Alphaproteobacteria, isolated on the SpanishMediterranean seaboard. Marine Drugs, 8(8), 2240–2251.
Maleki, S., Almaas, E., Zotchev, S., Valla, S., & Ertesvåg, H. (2016). Alginate biosynthesisfactories in Pseudomonas fluorescens: Localization and correlation with alginate pro-duction level. Applied and environmental microbiology, 82(4), 1227–1236.
Manca, M. C., Lama, L., Improta, R., Esposito, E., Gambacorta, A., & Nicolaus, B. (1996).Chemical composition of two exopolysaccharides from Bacillus thermoantarcticus.Applied and environmental microbiology, 62(9), 3265–3269.
Martínez-Checa, F., Toledo, F. L., El Mabrouki, K., Quesada, E., & Calvo, C. (2007).Characteristics of bioemulsifier V2-7 synthesized in culture media added of hydro-carbons: Chemical composition, emulsifying activity and rheological properties.Bioresource Technology, 98(16), 3130–3135.
Marx, J. G., Carpenter, S. D., & Deming, J. W. (2009). Production of cryoprotectant ex-tracellular polysaccharide substances (EPS) by the marine psychrophilic bacteriumColwellia psychrerythraea strain 34H under extreme conditions. Canadian Journal ofMicrobiology, 55(1), 63–72.
Mata, J. A., Béjar, V., Bressollier, P., Tallon, R., Urdaci, M., Quesada, E., ... Llamas, I.(2008). Characterization of exopolysaccharides produced by three moderately halo-philic bacteria belonging to the family Alteromonadaceae. Journal of AppliedMicrobiology, 105(2), 521–528.
Mata, J. A., Béjar, V., Llamas, I., Arias, S., Bressollier, P., Tallon, R., ... Quesada, E. (2006).Exopolysaccharides produced by the recently described halophilic bacteriaHalomonas ventosae and Halomonas anticariensis. Research in Microbiology, 157(9),827–835.
Maugeri, T. L., Gugliandolo, C., Caccamo, D., Panico, A., Lama, L., Gambacorta, A., ...Nicolaus, B. (2002). A halophilic thermotolerant Bacillus isolated from a marine hotspring able to produce a new exopolysaccharide. Biotechnology Letters, 24(7),515–519.
Michel, C., Beny, C., Delorme, F., Poirier, L., Spolaore, P., Morin, D., ... d’Hugues, P.(2009). New protocol for the rapid quantification of exopolysaccharides in con-tinuous culture systems of acidophilic bioleaching bacteria. Applied Microbiology andBiotechnology, 82(2), 371–378.
Moreno-Paz, M., Gómez, M. J., Arcas, A., & Parro, V. (2010). Environmental tran-scriptome analysis reveals physiological differences between biofilm and planktonicmodes of life of the iron oxidizing bacteria Leptospirillum spp. in their natural mi-crobial community. BMC Genomics, 11(1), 404.
Morgan, J. L. W., Strumillo, J., & Zimmer, J. (2012). Crystallographic snapshot of cel-lulose synthesis and membrane translocation. Nature, 493, 181–186.
Moriello, V. S., Lama, L., Poli, A., Gugliandolo, C., Maugeri, T. L., Gambacorta, A., ...Nicolaus, B. (2003). Production of exopolysaccharides from a thermophilic micro-organism isolated from a marine hot spring in flegrean areas. Journal of IndustrialMicrobiology and Biotechnology, 30(2), 95–101.
Nevot, M., Deroncele, V., Montes, M. J., & Mercade, E. (2008). Effect of incubationtemperature on growth parameters of Pseudoalteromonas antarctica NF and its pro-duction of extracellular polymeric substances. Journal of Applied Microbiology, 105(1),255–263.
Nichols, C. M., Garon, S., Bowman, J. P., Raguenes, G., & Guezennec, J. (2004).Production of exopolysaccharides by Antarctic marine bacterial isolates. Journal ofApplied Microbiology, 96(5), 1057–1066.
Nichols, C. M., Bowman, J. P., & Guezennec, J. (2005). Effects of incubation temperatureon growth and production of exopolysaccharides by an Antarctic sea ice bacteriumgrown in batch culture. Applied and environmental microbiology, 71(7), 3519–3523.
Nichols, C. M., Guezennec, J., & Bowman, J. P. (2005). Bacterial exopolysaccharides fromextreme marine environments with special consideration of the southern ocean, seaice, and deep-sea hydrothermal vents: A review. Marine Biotechnology, 7(4), 253–271.
Nichols, C. M., Lardière, S. G., Bowman, J. P., Nichols, P. D., Gibson, J. A. E., &Guézennec, J. (2005). Chemical characterization of exopolysaccharides fromAntarctic marine bacteria. Microbial Ecology, 49(4), 578–589.
Nicolaus, B., Kambourova, M., & Oner, E. T. (2010). Exopolysaccharides from ex-tremophiles: From fundamentals to biotechnology. Environmental Technology, 31(10),1145–1158.
Nicolaus, B., Lama, L., Esposito, E., Manca, M. C., Improta, R., Bellitti, M. R., ...Gambacorta, A. (1999). Haloarcula spp able to biosynthesize exo- and endopolymers.
J. Wang et al. Carbohydrate Polymers 205 (2019) 8–26
24

Journal of Industrial Microbiology and Biotechnology, 23(6), 489–496.Nicolaus, B., Lama, L., Panico, A., Moriello, V. S., Romano, I., & Gambacorta, A. (2002).
Production and characterization of exopolysaccharides excreted by thermophilicbacteria from shallow, marine hydrothermal vents of flegrean ares (Italy). Systematicand Applied Microbiology, 25(3), 319–325.
Nicolaus, B., Moriello, V. S., Lama, L., Poli, A., & Gambacorta, A. (2004). Polysaccharidesfrom extremophilic microorganisms. Origins of life and evolution of the biosphere,34(1), 159–169.
Nicolaus, B., Panico, A., Manca, M. C., Lama, L., Gambacorta, A., Maugeri, T. L., ...Caccamo, D. (2000). A thermophilic Bacillus isolated from an Eolian shallow hy-drothermal vent able to produce exopolysaccharides. Systematic and AppliedMicrobiology, 23(3), 426–432.
Ollivier, B., Caumette, P., Garcia, J.-L., & Mah, R. (1994). Anaerobic bacteria from hy-persaline environments. Microbiological reviews, 58(1), 27–38.
Panosyan, H., Di Donato, P., Poli, A., & Nicolaus, B. (2018). Production and character-ization of exopolysaccharides by Geobacillus thermodenitrificans ArzA-6 andGeobacillus toebii ArzA-8 strains isolated from an Armenian geothermal spring.Extremophiles, 22(5), 725–737.
Perry, T. D., Klepac-Ceraj, V., Zhang, X. V., McNamara, C. J., Polz, M. F., Martin, S. T., ...Mitchell, R. (2005). Binding of harvested bacterial exopolymers to the surface ofcalcite. Environmental Science & Technology, 39(22), 8770–8775.
Poli, A., Anzelmo, G., & Nicolaus, B. (2010). Bacterial exopolysaccharides from extrememarine habitats: Production, characterization and biological activities. Marine Drugs,8(6), 1779–1802.
Poli, A., Kazak, H., Gürleyendağ, B., Tommonaro, G., Pieretti, G., Öner, E. T., ... Nicolaus,B. (2009). High level synthesis of levan by a novel Halomonas species growing ondefined media. Carbohydrate Polymers, 78(4), 651–657.
Poli, A., Moriello, V. S., Esposito, E., Lama, L., Gambacorta, A., & Nicolaus, B. (2004).Exopolysaccharide production by a new Halomonas strain CRSS isolated from salinelake Cape Russell in Antarctica growing on complex and defined media. BiotechnologyLetters, 26(21), 1635–1638.
Prechtl, R. M., Wefers, D., Jakob, F., & Vogel, R. F. (2018). Cold and salt stress modulateamount, molecular and macromolecular structure of a Lactobacillus sakei dextran.Food Hydrocolloids, 82, 73–81.
Qian, C., Li, W., Li, H., Ou, D., Zhu-Ge, Y., & Liu, Y. (2018). Responses of genes for theuptake of glycine betaine in Virgibacillus halodenitrificans PDB-F2 under NaCl stress.International Biodeterioration & Biodegradation, 132, 192–199.
Qian, Y., Qi, M., Zheng, L., King, W. M., Lv, L., & Ye, F. (2016). Incorporation of rutin inelectrospun pullulan/PVA nanofibers for novel UV-resistant properties. Materials,9(7), 504.
Qin, G., Zhu, L., Chen, X., Wang, P. G., & Zhang, Y. (2007). Structural characterizationand ecological roles of a novel exopolysaccharide from the deep-sea psychrotolerantbacterium Pseudoalteromonas sp. SM9913. Microbiology, 153(Pt 5), 1566–1572.
Radchenkova, N., Boyadzhieva, I., Atanasova, N., Poli, A., Finore, I., Di Donato, P., ...Kambourova, M. (2018). Extracellular polymer substance synthesized by a halophilicbacterium Chromohalobacter canadensis 28. Applied Microbiology and Biotechnology,102(11), 4937–4949.
Radchenkova, N., Panchev, I., Vassilev, S., Kuncheva, M., Dobreva, S., & Kambourova, M.(2015). Continuous cultivation of a thermophilic bacterium Aeribacillus pallidus 418for production of an exopolysaccharide applicable in cosmetic creams. Journal ofApplied Microbiology, 119(5), 1301–1309.
Radchenkova, N., Vassilev, S., Martinov, M., Kuncheva, M., Panchev, I., Vlaev, S., ...Kambourova, M. (2014). Optimization of the aeration and agitation speed ofAeribacillus palidus 418 exopolysaccharide production and the emulsifying propertiesof the product. Process Biochemistry, 49(4), 576–582.
Radchenkova, N., Vassilev, S., Panchev, I., Anzelmo, G., Tomova, I., Nicolaus, B., ...Kambourova, M. (2013). Production and properties of two novel exopolysaccharidessynthesized by a thermophilic bacterium Aeribacillus pallidus 418. AppliedBiochemistry and Biotechnology, 171(1), 31–43.
Raveendran, S., Girija, A. R., Balasubramanian, S., Ukai, T., Yoshida, Y., Maekawa, T., ...Kumar, D. S. (2014). Green approach for augmenting biocompatibility to quantumdots by extremophilic polysaccharide conjugation and nontoxic bioimaging. ACSSustainable Chemistry & Engineering, 2(7), 1551–1558.
Raveendran, S., Dhandayuthapani, B., Nagaoka, Y., Yoshida, Y., Maekawa, T., & SakthiKumar, D. (2013). Biocompatible nanofibers based on extremophilic bacterial poly-saccharide, Mauran from Halomonas maura. Carbohydrate Polymers, 92(2),1225–1233.
Raveendran, S., Poulose, A. C., Yoshida, Y., Maekawa, T., & Kumar, D. S. (2013). Bacterialexopolysaccharide based nanoparticles for sustained drug delivery, cancer che-motherapy and bioimaging. Carbohydrate Polymers, 91(1), 22–32.
Rees, H. C., Grant, W. D., Jones, B. E., & Heaphy, S. (2004). Diversity of Kenyan soda lakealkaliphiles assessed by molecular methods. Extremophiles, 8(1), 63–71.
Rehm, B. H. (2010). Bacterial polymers: Biosynthesis, modifications and applications.Nature Reviews Microbiology, 8(8), 578–592.
Ruiz, L. M., Castro, M., Barriga, A., Jerez, C. A., & Guiliani, N. (2011). The extremophileAcidithiobacillus ferrooxidans possesses a c‐di‐GMP signalling pathway that could playa significant role during bioleaching of minerals. Letters in Applied Microbiology, 54(2),133–139.
Ruiz-Ruiz, C., Srivastava, G. K., Carranza, D., Mata, J. A., Llamas, I., Santamaría, M., ...Molina, I. J. (2011). An exopolysaccharide produced by the novel halophilic bac-terium Halomonas stenophila strain B100 selectively induces apoptosis in human Tleukaemia cells. Applied Microbiology and Biotechnology, 89(2), 345–355.
Rütering, M., Cress, B. F., Schilling, M., Rühmann, B., Koffas, M. A. G., Sieber, V., ...Schmid, J. (2017). Tailor-made exopolysaccharides—CRISPR-Cas9 mediated genomeediting in Paenibacillus polymyxa. Synthetic Biology, 2(1) ysx007-ysx007.
Sá-Correia, I., Fialho, A. M., Videira, P., Moreira, L. M., Marques, A. R., & Albano, H.
(2002). Gellan gum biosynthesis in Sphingomonas paucimobilis ATCC 31461: Genes,enzymes and exopolysaccharide production engineering. Journal of IndustrialMicrobiology and Biotechnology, 29(4), 170–176.
Salem, D. R. (2007). Electrospinning of nanofibers and the charge injection method. In P.J. Brown, & K. Stevens (Eds.). Nanofibers and nanotechnology in textiles (pp. 3–21).Woodhead Publishing.
Sam, S., Kucukasik, F., Yenigun, O., Nicolaus, B., Oner, E. T., & Yukselen, M. A. (2011).Flocculating performances of exopolysaccharides produced by a halophilic bacterialstrain cultivated on agro-industrial waste. Bioresource Technology, 102(2),1788–1794.
Samain, E., Miles, M., Bozzi, L., Dubreucq, G., & Rinaudo, M. (1997). Simultaneousproduction of two different gel-forming exopolysaccharides by an Alteromonas strainoriginating from deep sea hydrothermal vents. Carbohydrate Polymers, 34(4),235–241.
Santos, C., Silva, C. J., Büttel, Z., Guimarães, R., Pereira, S. B., Tamagnini, P., ... Zille, A.(2014). Preparation and characterization of polysaccharides/PVA blend nanofibrousmembranes by electrospinning method. Carbohydrate Polymers, 99, 584–592.
Sardari, R. R., Kulcinskaja, E., Ron, E. Y., Bjornsdottir, S., Friethjonsson, O. H.,Hreggviethsson, G. O., ... Karlsson, E. N. (2017). Evaluation of the production ofexopolysaccharides by two strains of the thermophilic bacterium Rhodothermusmarinus. Carbohydrate Polymers, 156, 1–8.
Sarilmiser, H. K., & Oner, E. T. (2014). Investigation of anti-cancer activity of linear andaldehyde-activated levan from Halomonas smyrnensis AAD6T. Biochemical EngineeringJournal, 92, 28–34.
Sarilmiser, H. K., Ates, O., Ozdemir, G., Arga, K. Y., & Oner, E. T. (2015). Effective sti-mulating factors for microbial levan production by Halomonas smyrnensis AAD6T.Journal of Bioscience and Bioengineering, 119(4), 455–463.
Schmid, J. (2018). Recent insights in microbial exopolysaccharide biosynthesis and en-gineering strategies. Current opinion in biotechnology, 53, 130–136.
Schmid, J., Sieber, V., & Rehm, B. (2015). Bacterial exopolysaccharides: Biosynthesispathways and engineering strategies. Frontiers in Microbiology, 6, 496.
Sezer, A. D., Kazak, H., Öner, E. T., & Akbuğa, J. (2011). Levan-based nanocarrier systemfor peptide and protein drug delivery: Optimization and influence of experimentalparameters on the nanoparticle characteristics. Carbohydrate Polymers, 84(1),358–363.
Sima, F., Mutlu, E. C., Eroglu, M. S., Sima, L. E., Serban, N., Ristoscu, C., ... Mihailescu, I.N. (2011). Levan nanostructured thin films by MAPLE assembling. Biomacromolecules,12(6), 2251–2256.
Spanò, A., & Arena, A. (2016). Bacterial exopolysaccharide of shallow marine vent originas agent in counteracting immune disorders induced by herpes virus. Journal ofImmunoassay and Immunochemistry, 37(3), 251–260.
Spanò, A., Gugliandolo, C., Lentini, V., Maugeri, T. L., Anzelmo, G., Poli, A., ... Nicolaus,B. (2013). A novel EPS-producing strain of Bacillus licheniformis isolated from ashallow vent off Panarea Island (Italy). Current Microbiology, 67(1), 21–29.
Spanò, A., Laganà, P., Visalli, G., Maugeri, T. L., & Gugliandolo, C. (2016). In vitro anti-biofilm activity of an exopolysaccharide from the marine thermophilic Bacillus li-cheniformis T14. Current Microbiology, 72(5), 518–528.
Stingele, F., Vincent, S. J. F., Faber, E. J., Newell, J. W., Kamerling, J. P., & Neeser, J.-R.(1999). Introduction of the exopolysaccharide gene cluster from Streptococcus ther-mophilus Sfi6 into Lactococcus lactis MG1363: Production and characterization of analtered polysaccharide. Molecular Microbiology, 32(6), 1287–1295.
Subramanian, S. B., Yan, S., Tyagi, R. D., & Surampalli, R. Y. (2010). Extracellularpolymeric substances (EPS) producing bacterial strains of municipal wastewatersludge: Isolation, molecular identification, EPS characterization and performance forsludge settling and dewatering. Water Research, 44(7), 2253–2266.
Sun, M.-L., Zhao, F., Shi, M., Zhang, X.-Y., Zhou, B.-C., Zhang, Y.-Z., ... Chen, X.-L. (2015).Characterization and biotechnological potential analysis of a new exopolysaccharidefrom the Arctic marine bacterium Polaribacter sp. SM1127. Scientific Reports, 5,18435.
Thorne, L., Mikolajczak, M. J., Armentrout, R. W., & Pollock, T. J. (2000). Increasing theyield and viscosity of exopolysaccharides secreted by Sphingomonas by augmentationof chromosomal genes with multiple copies of cloned biosynthetic genes. Journal ofIndustrial Microbiology and Biotechnology, 25(1), 49–57.
Torres-Giner, S., Ocio, M. J., & Lagaron, J. M. (2008). Development of active anti-microbial fiber-based chitosan polysaccharide nanostructures using electrospinning.Engineering in Life Sciences, 8(3), 303–314.
Ueda, A., & Wood, T. K. (2009). Connecting quorum sensing, c-di-GMP, Pel poly-saccharide, and biofilm formation in Pseudomonas aeruginosa through tyrosinephosphatase TpbA (PA3885). PLOS Pathogens, 5(6), e1000483.
Vashisth, P., Pruthi, P. A., Singh, R. P., & Pruthi, V. (2014). Process optimization forfabrication of gellan based electrospun nanofibers. Carbohydrate Polymers, 109,16–21.
Vu, B., Chen, M., Crawford, R. J., & Ivanova, E. P. (2009). Bacterial extracellular poly-saccharides involved in biofilm formation. Molecules, 14(7), 2535–2554.
Wang, H., Liu, Y. M., Qi, Z. M., Wang, S. Y., Liu, S. X., Li, X., ... Xia, X. C. (2013). Anoverview on natural polysaccharides with antioxidant properties. Current MedicinalChemistry, 20(23), 2899–2913.
Wang, L., Zhang, H., Yang, L., Liang, X., Zhang, F., & Linhardt, R. J. (2017). Structuralcharacterization and bioactivity of exopolysaccharide synthesized by Geobacillus sp.TS3-9 isolated from radioactive radon hot spring. Advances in Biotechnology andMicrobiology, 4(2), 1–8.
Welman, A. D., Maddox, I. S., & Archer, R. H. (2006). Metabolism associated with raisedmetabolic flux to sugar nucleotide precursors of exopolysaccharides in Lactobacillusdelbrueckii subsp. bulgaricus. Journal of Industrial Microbiology and Biotechnology,33(5), 391.
Whitney, J. C., & Howell, P. L. (2013). Synthase-dependent exopolysaccharide secretion
J. Wang et al. Carbohydrate Polymers 205 (2019) 8–26
25

in Gram-negative bacteria. Trends in Microbiology, 21(2), 63–72.Xu, X., Bi, D., & Wan, M. (2016). Characterization and immunological evaluation of low-
molecular-weight alginate derivatives. Current Topics in Medicinal Chemistry, 16(8),874–887.
Yildiz, S. Y., Anzelmo, G., Ozer, T., Radchenkova, N., Genc, S., Di Donato, P., ...Kambourova, M. (2014). Brevibacillus themoruber: A promising microbial cell factoryfor exopolysaccharide production. Journal of Applied Microbiology, 116(2), 314–324.
Yildiz, S. Y., Radchenkova, N., Arga, K. Y., Kambourova, M., & Oner, E. T. (2015).Genomic analysis of Brevibacillus thermoruber 423 reveals its biotechnological andindustrial potential. Applied Microbiology and Biotechnology, 99(5), 2277–2289.
Zeldes, B. M., Keller, M. W., Loder, A. J., Straub, C. T., Adams, M. W. W., & Kelly, R. M.(2015). Extremely thermophilic microorganisms as metabolic engineering platformsfor production of fuels and industrial chemicals. Frontiers in Microbiology, 6, 1209.
Zeng, W., Qiu, G., Zhou, H., Liu, X., Chen, M., Chao, W., ... Peng, J. (2010).
Characterization of extracellular polymeric substances extracted during the bio-leaching of chalcopyrite concentrate. Hydrometallurgy, 100(3-4), 177–180.
Zhao, S., Cao, F., Zhang, H., Zhang, L., Zhang, F., & Liang, X. (2014). Structural char-acterization and biosorption of exopolysaccharides from Anoxybacillus sp. R4-33isolated from radioactive radon hot spring. Applied Biochemistry and Biotechnology,172(5), 2732–2746.
Zheng, C., Li, Z., Su, J., Zhang, R., Liu, C., & Zhao, M. (2012). Characterization andemulsifying property of a novel bioemulsifier by Aeribacillus pallidus YM‐1. Journal ofApplied Microbiology, 113(1), 44–51.
Zhu, P., Chen, X., Li, S., Xu, H., Dong, S., Xu, Z., ... Zhang, Y. (2014). Screening andcharacterization of Sphingomonas sp. mutant for welan gum biosynthesis at an ele-vated temperature. Bioprocess and Biosystems Engineering, 37(9), 1849–1858.
Zong, A., Cao, H., & Wang, F. (2012). Anticancer polysaccharides from natural resources:A review of recent research. Carbohydrate Polymers, 90(4), 1395–1410.
J. Wang et al. Carbohydrate Polymers 205 (2019) 8–26
26
Recommended

















