

Lactation Physiology(part 1)
By: A. Riasi (PhD in Animal Nutrition &
Physiology)
فیزیولوژی تولید و ترشح شیر

At the end of this section student will be able to reply
What is the supportive system in udder?
What is the forstenberg’s rosette in papilla mammae?
What are the improtant parts of a secretory cells?
Which barriers are between the circulatory primary substance and milk?
What are the transporter systems in mammary epithelial cells?
Where is the lactose production site in the alveolar epithelial cells?
What are the pathways for milk fat globule transit and secretion from
mammary epithelial cells?
How is the de novo fatty acid synthesis in mammary cells?
What is the milk fat depression syndrome?

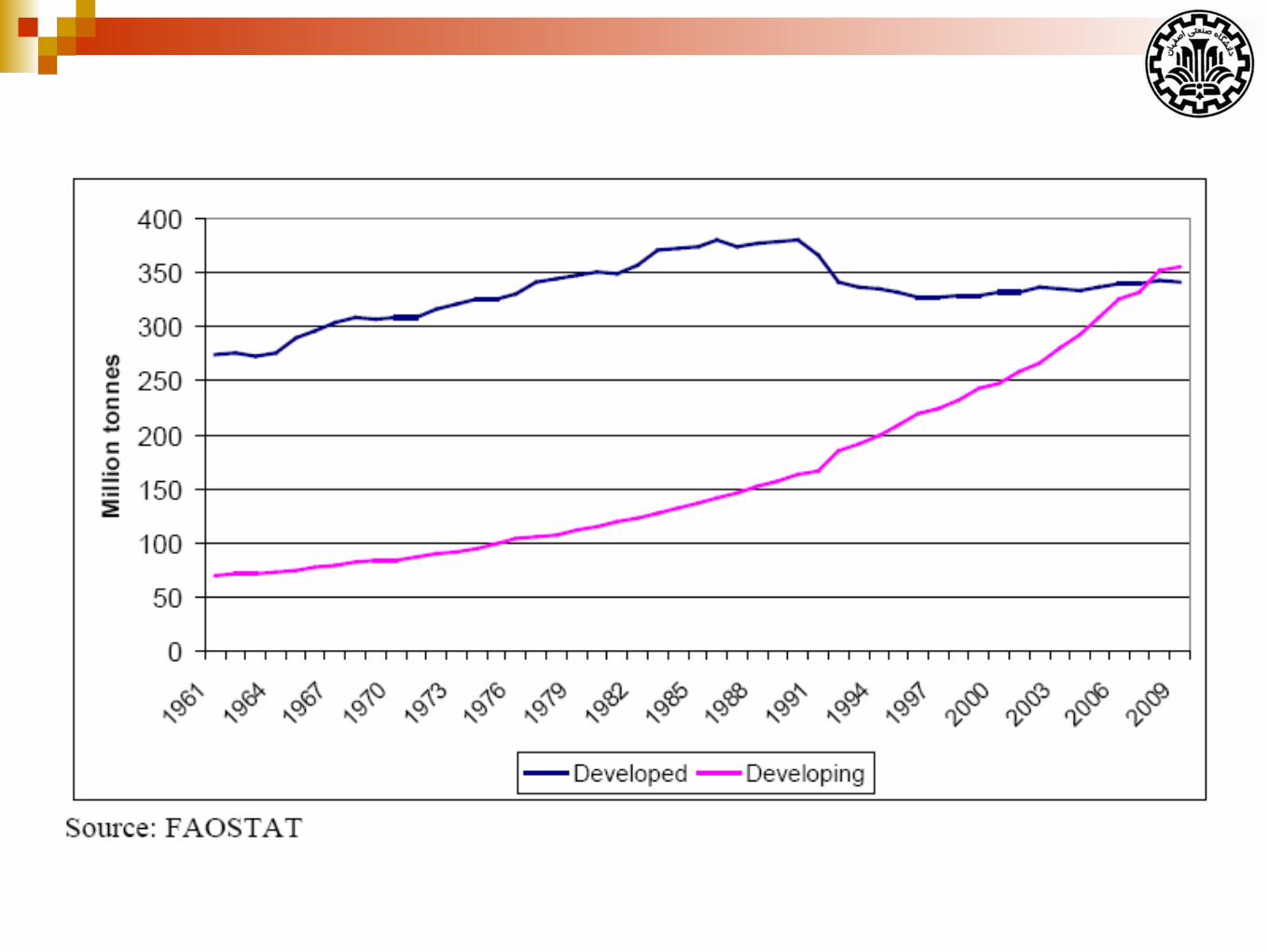

What is a mammary gland?
Is modified sweat gland.
Serves a reproductive function; nourishment of the neonate.
Can repeatedly undergo growth, functional differentiation, and
regression.
Relies on same endocrine (hormonal) support for development
and function.
Example: gonadal steroids, prolactin, etc.
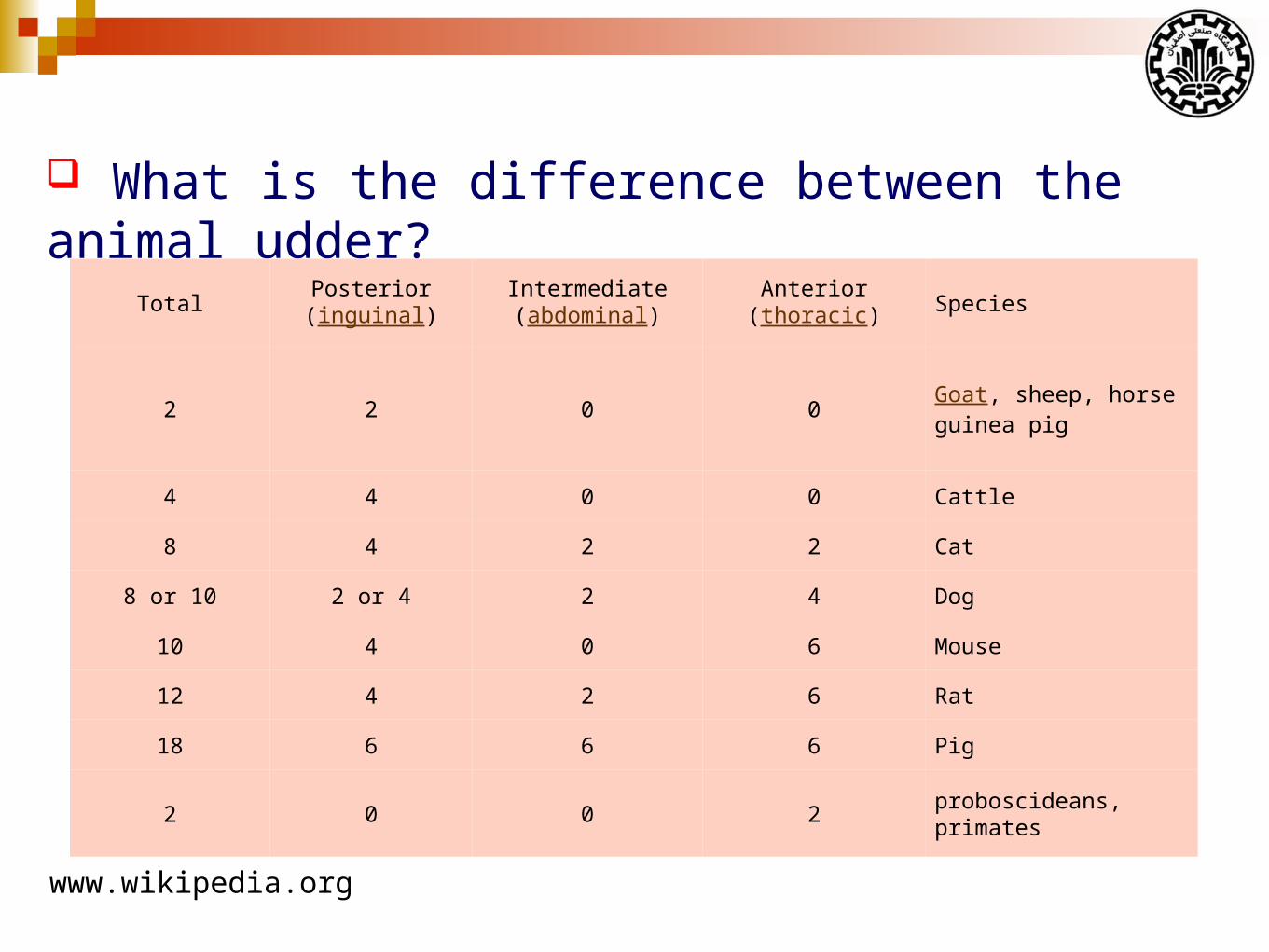
What is the difference between the animal udder?
SpeciesAnterior
(thoracic)Intermediate(abdominal)
Posterior(inguinal)
Total
Goat, sheep, horseguinea pig
0022
Cattle0044
Cat2248
Dog422 or 48 or 10
Mouse60410
Rat62412
Pig66618
proboscideans, primates2002
www.wikipedia.org

What is the difference between the animal udder?
Cow: Four glands and four teats
Sheep and goats: Two glands and two teats
Sow: 12-14 teats and two glands per teat.
Mare: Four glands and only two teats.

The udder is a complex system
A supportive system.
A secretory system composed of epithelial cells.
A duct system for storage and conveyance of milk.
Blood, lymph, and nerve systems.
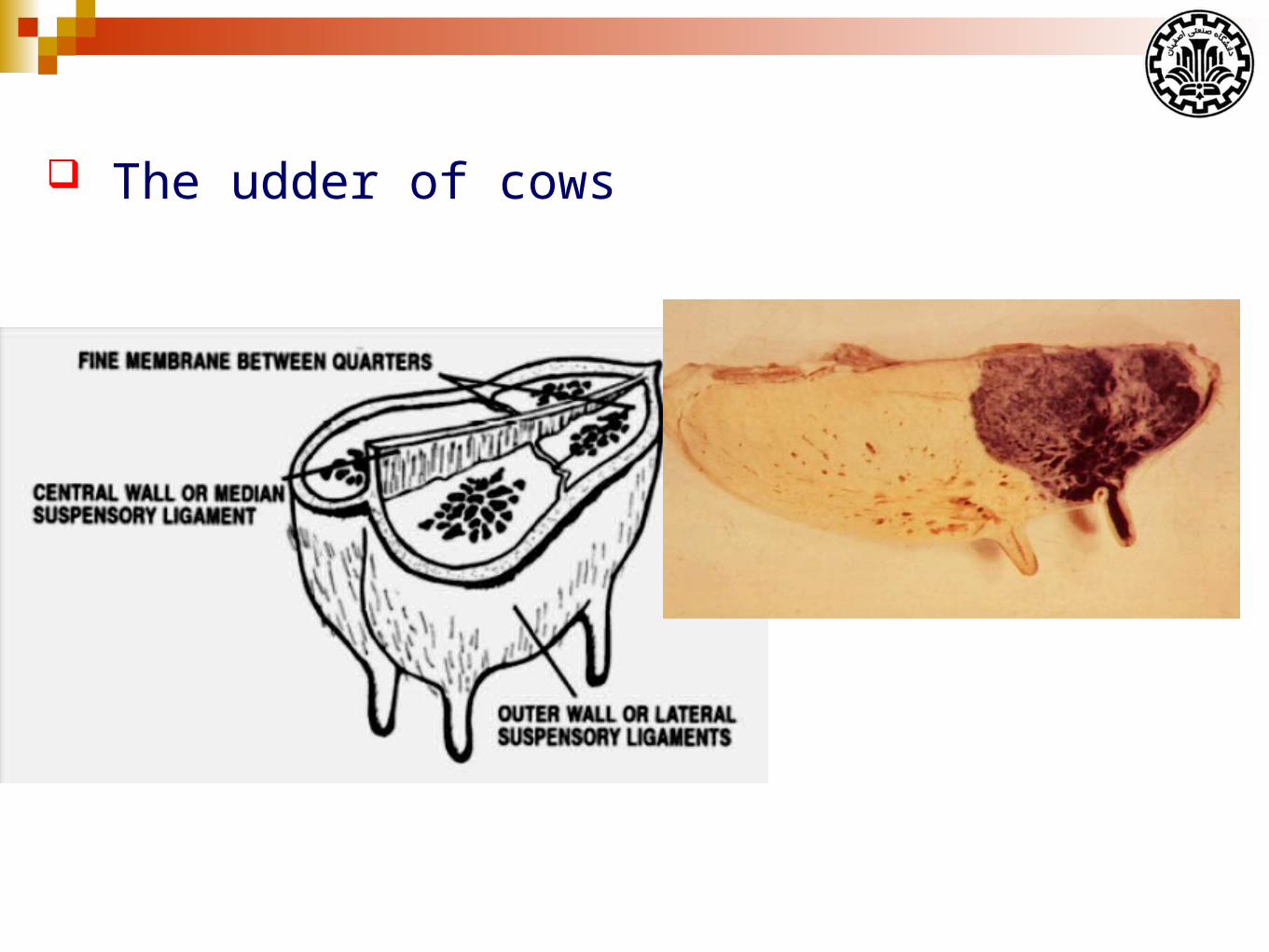
The udder of cows

The weight of empty cows udder is about 12-30 kg.
The udder weight is affected by:
Age
Stage of lactation
Amount of milk in the udder
Inherited differences among cows

The supportive system of udder

There are seven tissues that provide support for the udder:
Skin (covering the gland is only of very minor support)
Superficial fascia or Areolar subcutaneous tissue
Coarse areolar or cordlike tissue
Subpelvic tendon
Superficial layers of lateral suspensory ligament
Deep lateral suspensory ligament
Median Suspensory Ligament
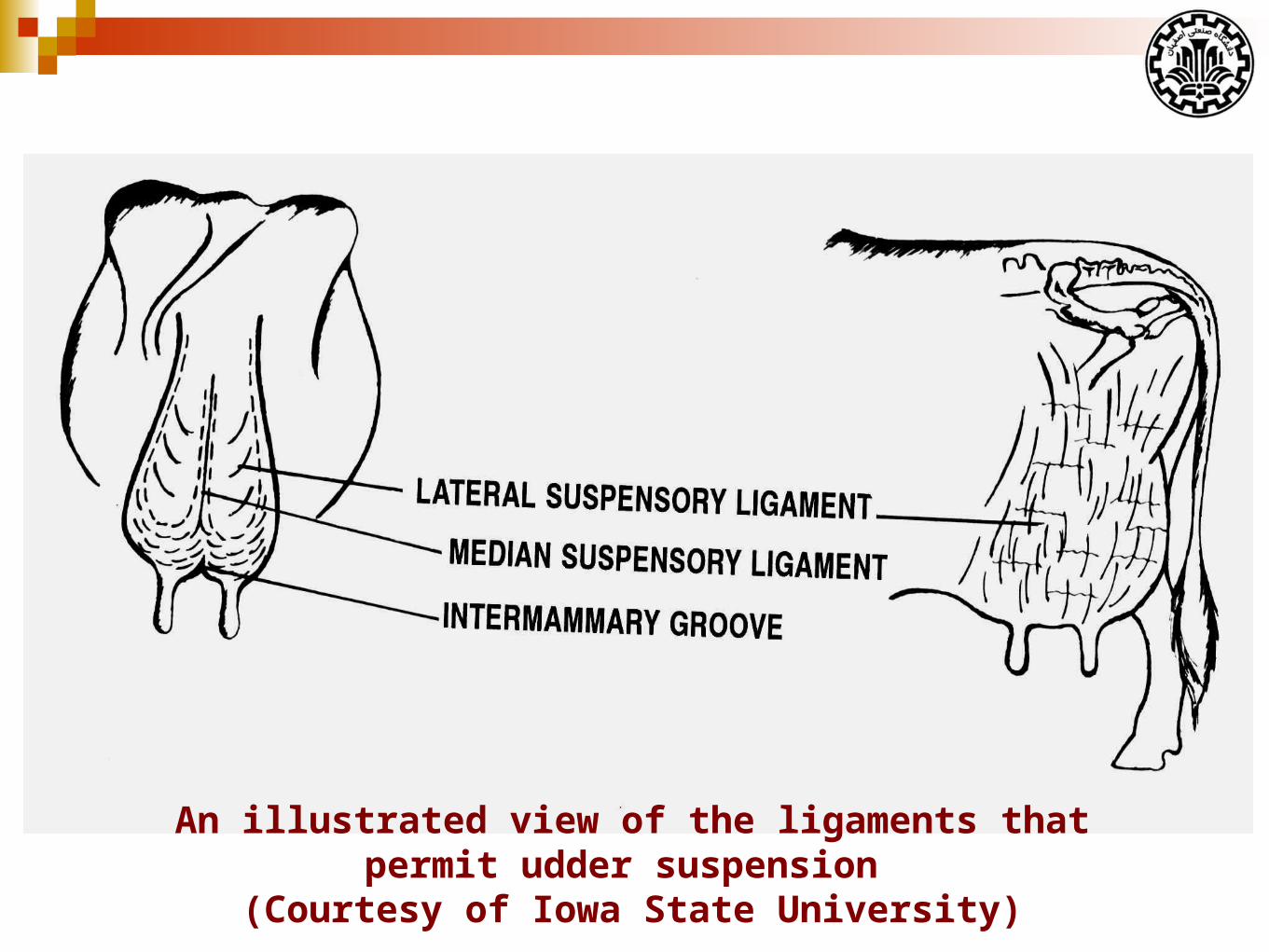
An illustrated view of the ligaments that permit udder suspension (Courtesy of Iowa State University)
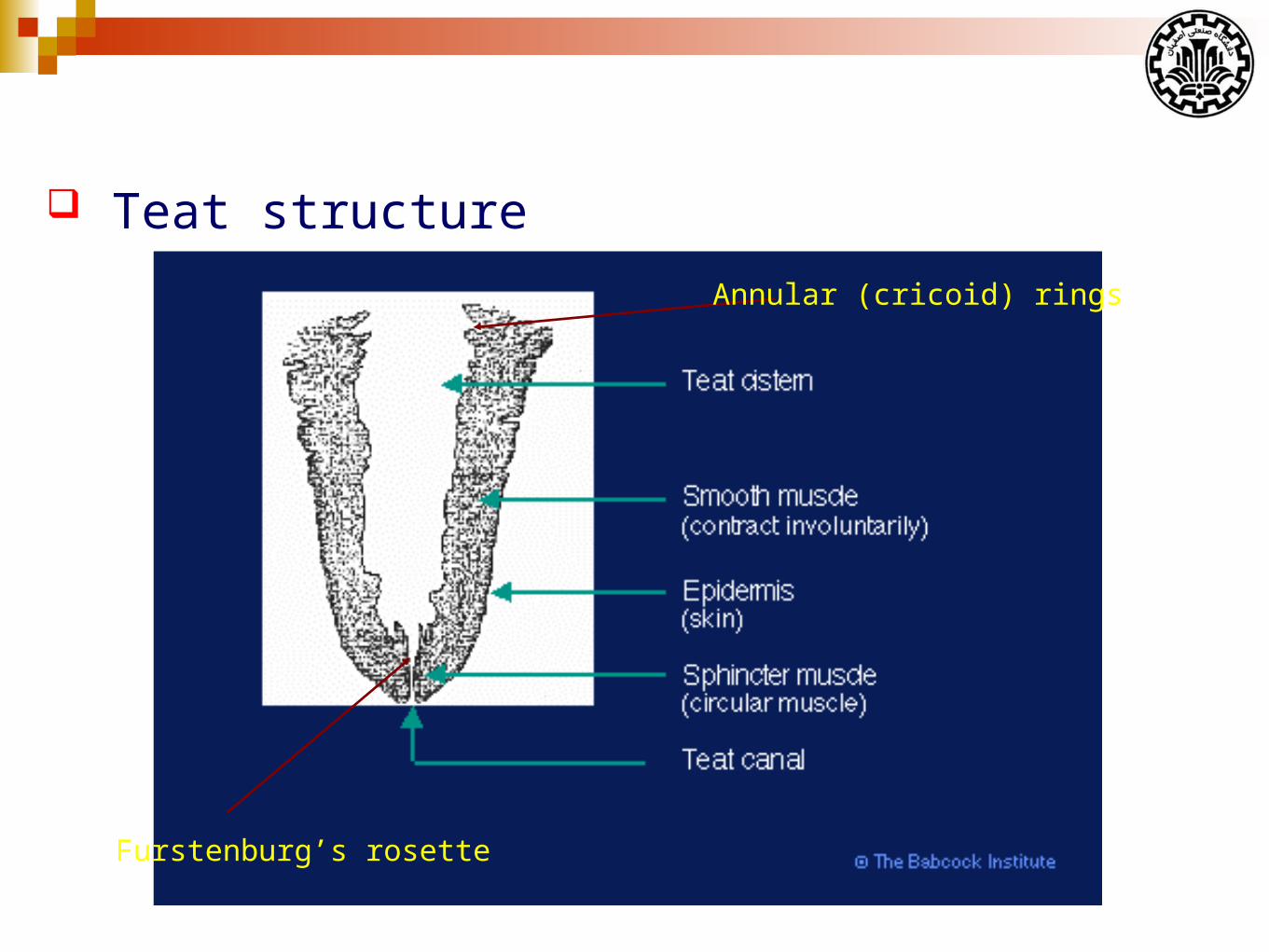
Teat structure
Furstenburg’s rosette
Annular (cricoid) rings

Interior anatomy of the Mammary Gland
The interior structure of mammary gland:
Connective tissue (Stroma)
Ductular system
Secretory tissue
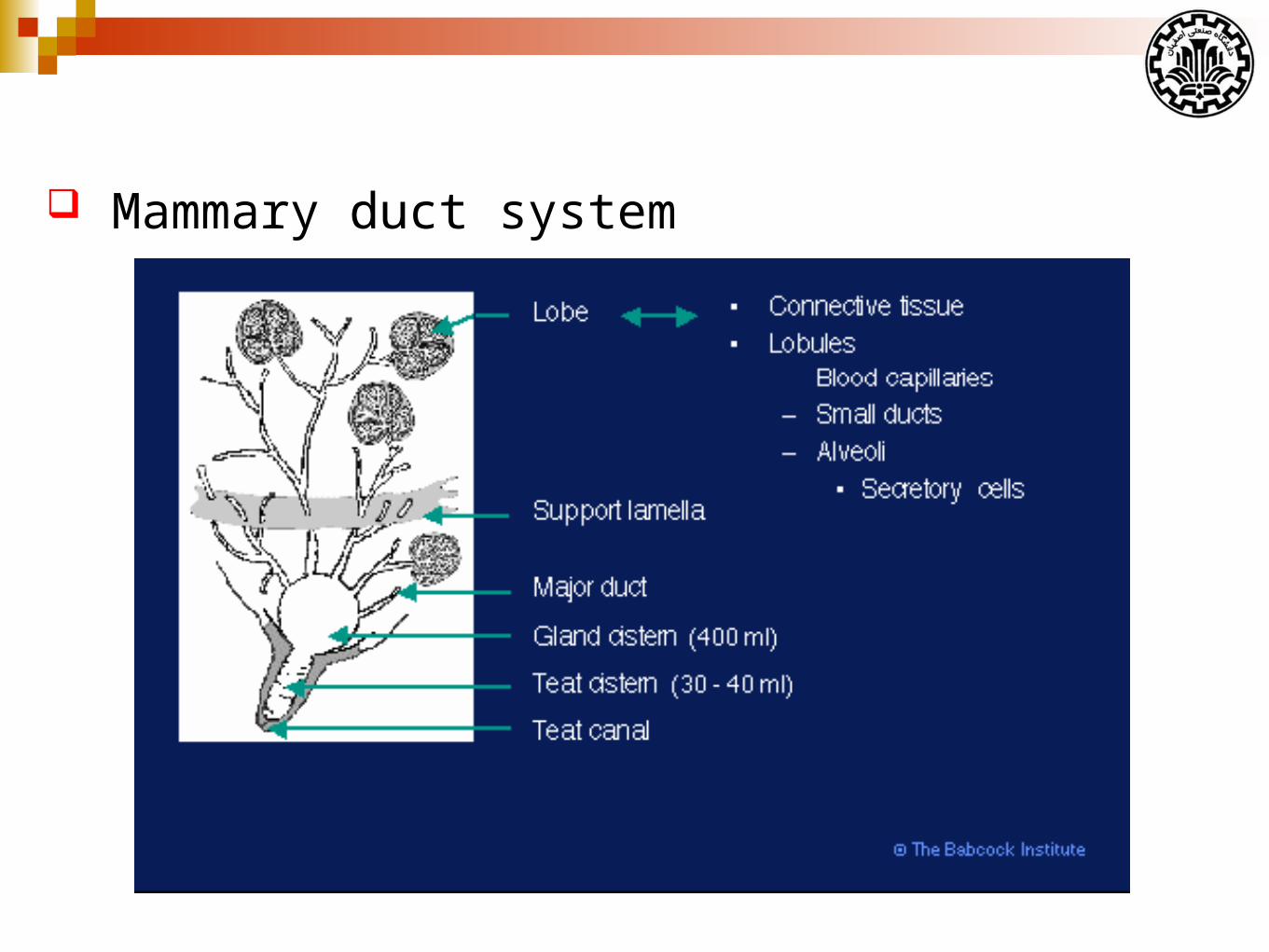
Mammary duct system
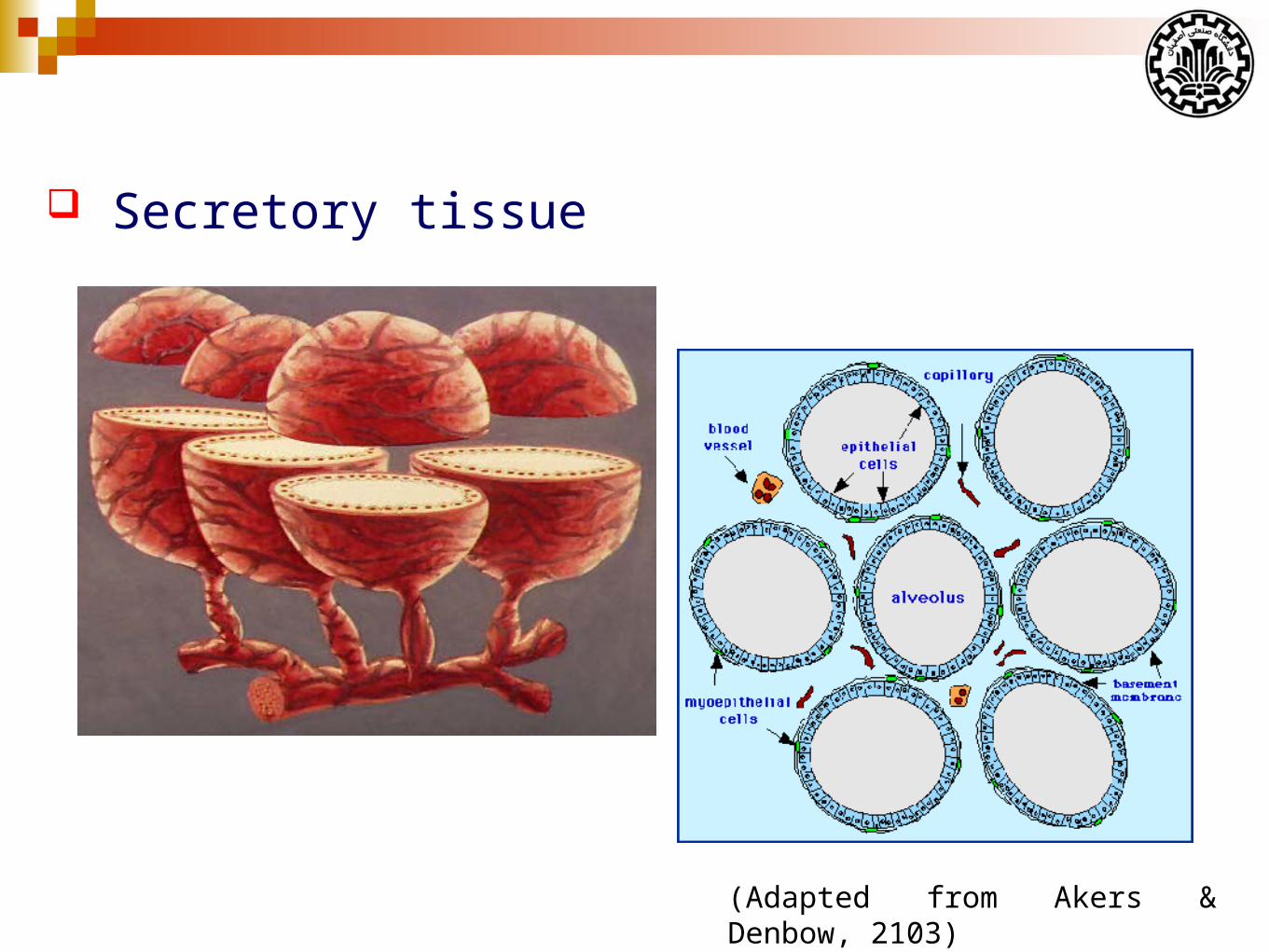
Secretory tissue
(Adapted from Akers & Denbow, 2103)
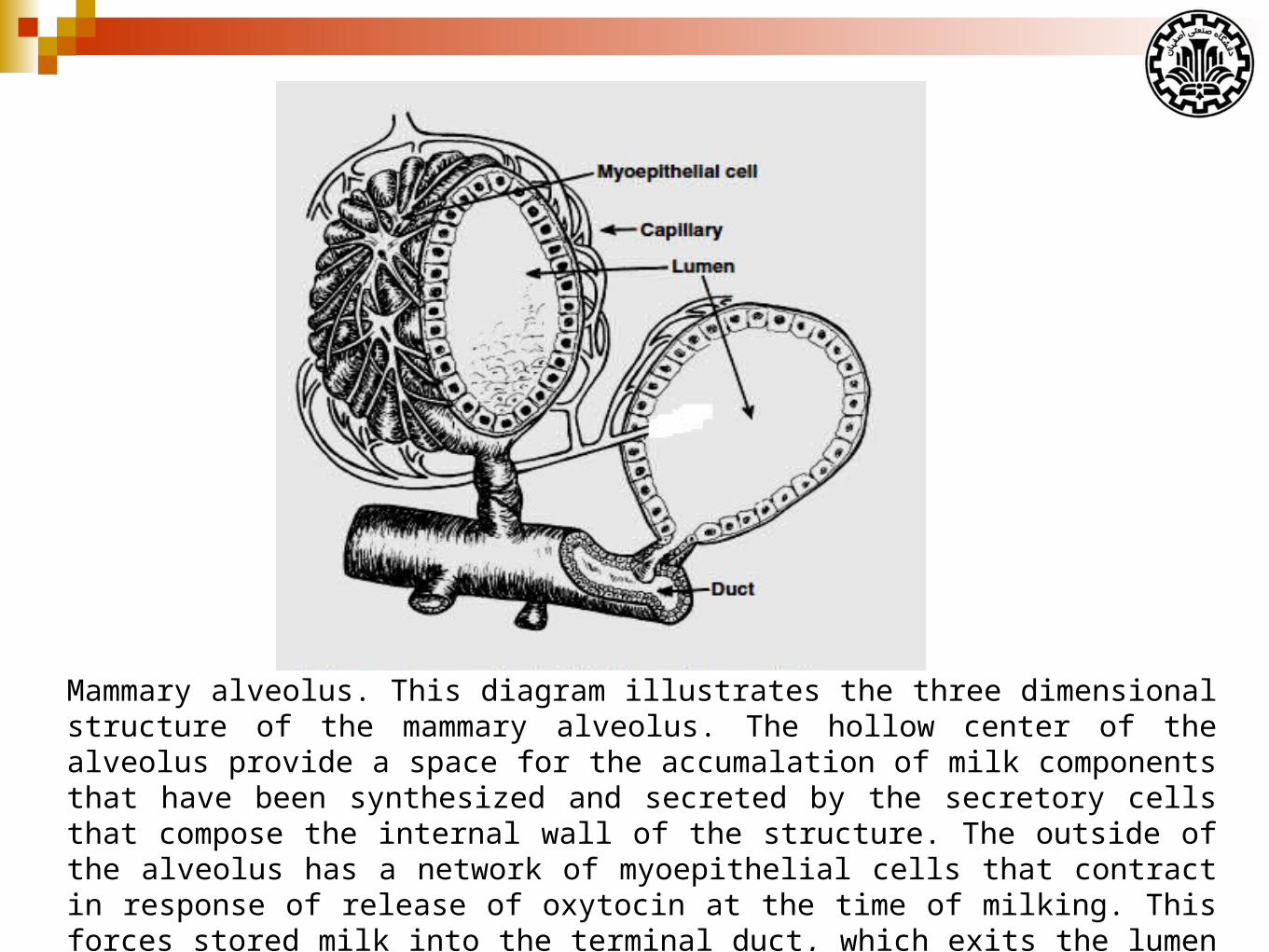
Mammary alveolus. This diagram illustrates the three dimensional structure of the mammary alveolus. The hollow center of the alveolus provide a space for the accumalation of milk components that have been synthesized and secreted by the secretory cells that compose the internal wall of the structure. The outside of the alveolus has a network of myoepithelial cells that contract in response of release of oxytocin at the time of milking. This forces stored milk into the terminal duct, which exits the lumen the alveolus. The milk progresses through larger ducts to be emptied at the nipple or teat end. (Adapted from Akers & Denbow, 2103)
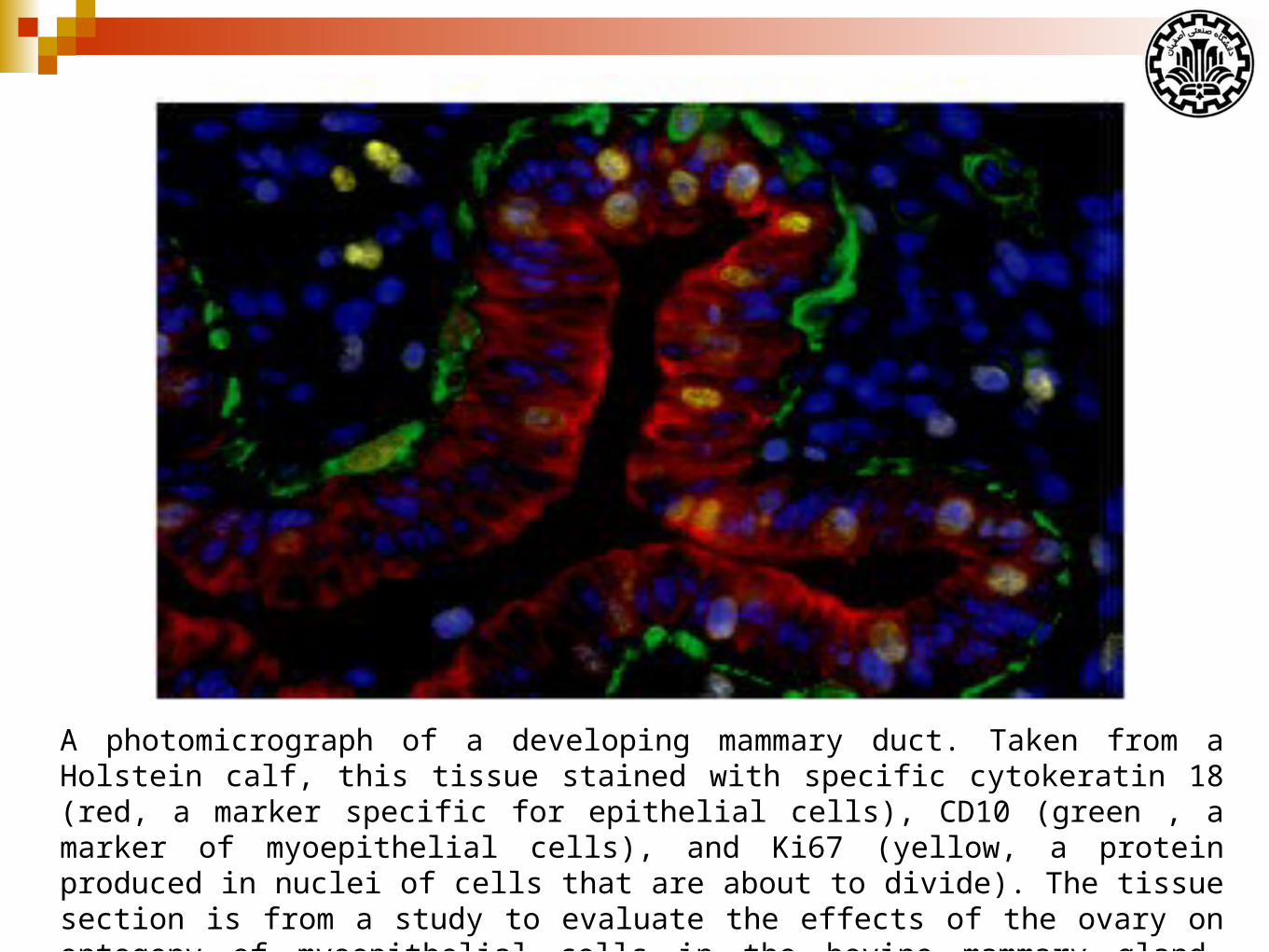
A photomicrograph of a developing mammary duct. Taken from a Holstein calf, this tissue stained with specific cytokeratin 18 (red, a marker specific for epithelial cells), CD10 (green , a marker of myoepithelial cells), and Ki67 (yellow, a protein produced in nuclei of cells that are about to divide). The tissue section is from a study to evaluate the effects of the ovary on ontogeny of myoepithelial cells in the bovine mammary gland. (Adapted from Akers & Denbow, 2103)

Secretory tissue
A lactating secretory cell is the basic unit of milk synthesis
Milk precursors are taken from the blood into the cell
The secretory cell have two kind of junctions with neighbor cells:
Tight junction around the apical portion
Gap junction in lateral portion
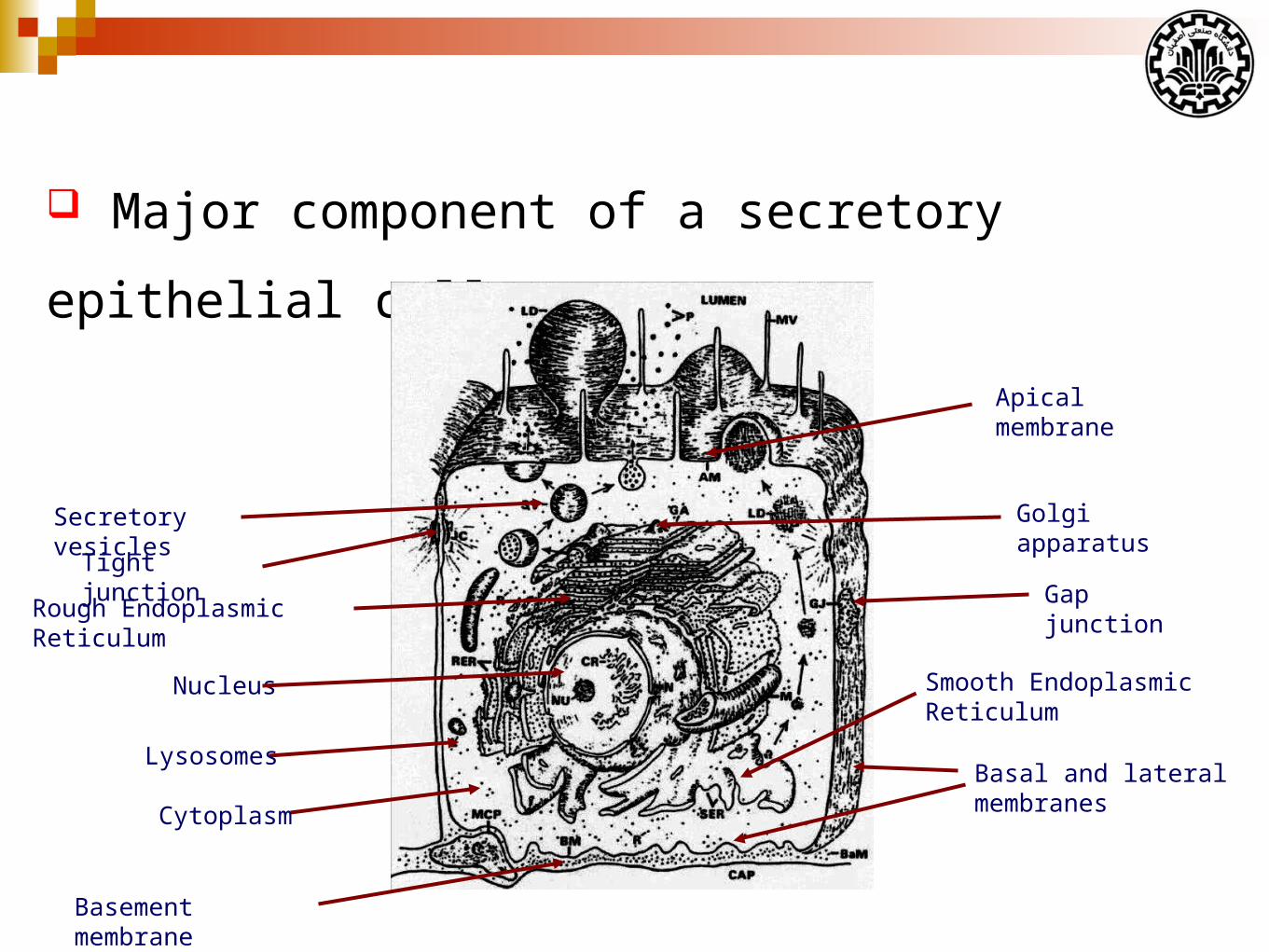
Major component of a secretory epithelial cell
Nucleus
Rough Endoplasmic Reticulum
Golgi apparatusSecretory vesicles
Lysosomes
Cytoplasm
Tight junctionGap junction
Basal and lateral membranes
Apical membrane
Basement membrane
Smooth Endoplasmic Reticulum

Milk synthesis and secretion
The product of mammary gland depends on two mode of
secretion:
Apocrine
Merocrine
Other components are derived by passage of soluble molecules
across (transcellular) and sometimes between (paracellular) the
cells.

Milk synthesis and secretion
Physically, milk is a complex solution of:
salts,
carbohydrates,
miscellaneous compounds with dispersed proteins and protein
aggregates,
casein micelles, and
fat globules.
Milk osmolarity generally equals blood (~300 mOsm) and has a
pH between 6.2 and 7.0.

Milk synthesis and secretion
Function of the mammary gland during established lactation is
closely linked with a number of hormones, growth factors, and
local tissue regulators.
Along with mammary cell-specific constituents, milk contains a
myriad of minor components.
Many of these molecules are important nutrients or regulators of
the neonate.

Milk synthesis and secretion
Molecules are transported into the milk by several possible
routes.
Mammary epithelial cells are able to maintain substantial
gradients for Na+, K+, and Cl− ions across the cell membrane.
Concentrations of Na+ inside (~ 43 mM) the cells are typically
lower than outside (150 mM)
The gradient for K+ is the opposite (143 mM inside compared
with 4.5 mM outside).
Concentration of Cl− is higher inside the cells.

Milk synthesis and secretion
Milk is a rich source of calcium.
The calcium in the milk exists as:
Casein-bound calcium
Calcium associated with various inorganic anions
For example, citrate and phosphate
Free calcium

Milk synthesis and secretion
The rate of calcium influx into the cell is matched by a
corresponding uptake of calcium by cellular organelles.
An ATP-dependent calcium pump on Golgi membranes
The uptake of Ca by the epithelial cells probably dependent to:
Parathyroid hormone-related protein
1,25-(OH)2 vitamin D3.

Precursors of Milk
Precursors of milk come from the bloodstream and the primary
substrates extracted from blood are:
Glucose
Amino acids
Fatty acids
Minerals
Acetate *
βHB *

Precursors of Milk
Several materials in milk come unchanged from the blood:
Minerals
Hormones
Immunoglobulins

Synthesis of milk proteins
There are several specific systems for amino acids absorption
through the basal membrane.
Inside the cell, amino acids are covalently bound together to
form proteins at the polysomes (Poly-ribosomes).
Proteins sythesized at RER include:
Casein
β-lactoglobulin
α-lactalbumin
Membrane bound proteins
Membrane boding enzymes

Synthesis of milk proteins
Synthesized proteins are transferred the golgi apparatues (GA).
Casein is secreted as micelle, which is formed in the GA from:
Casein molecules
Calcium
Phosphorus
A: a submicelle; B: protruding chain; C: Calcium phosphate; D: κ-casein; E: phosphate groups

Synthesis of milk lactose
Glucose enters the cells via the basolateral membrane by
specific transport system.
Some glucose is converted to galactose in the cell.
Both glucose and galactose enter the GA and react resulting in the
formation of lactose.

Synthesis of milk fat
The sources of milk FA:
Blood FA
De novo FA
Glycerol
Monoacylglyceride (MAG)
Acetate and *
β-hydroxybutyrate *
Milk fat triglycerides are synthesized on the smooth
endoplasmic reticulum and form small droplet.

Synthesis of milk fat
The protein coat on the milk fat globule membrane comprises:
Mainly butyrophilin (BTN) *
Xanthine oxidoreductase (XDH) *
Adipophilin (ADPH)**
Mucin 1
CD36
Periodic acid/Schiff
PAS III
FABP
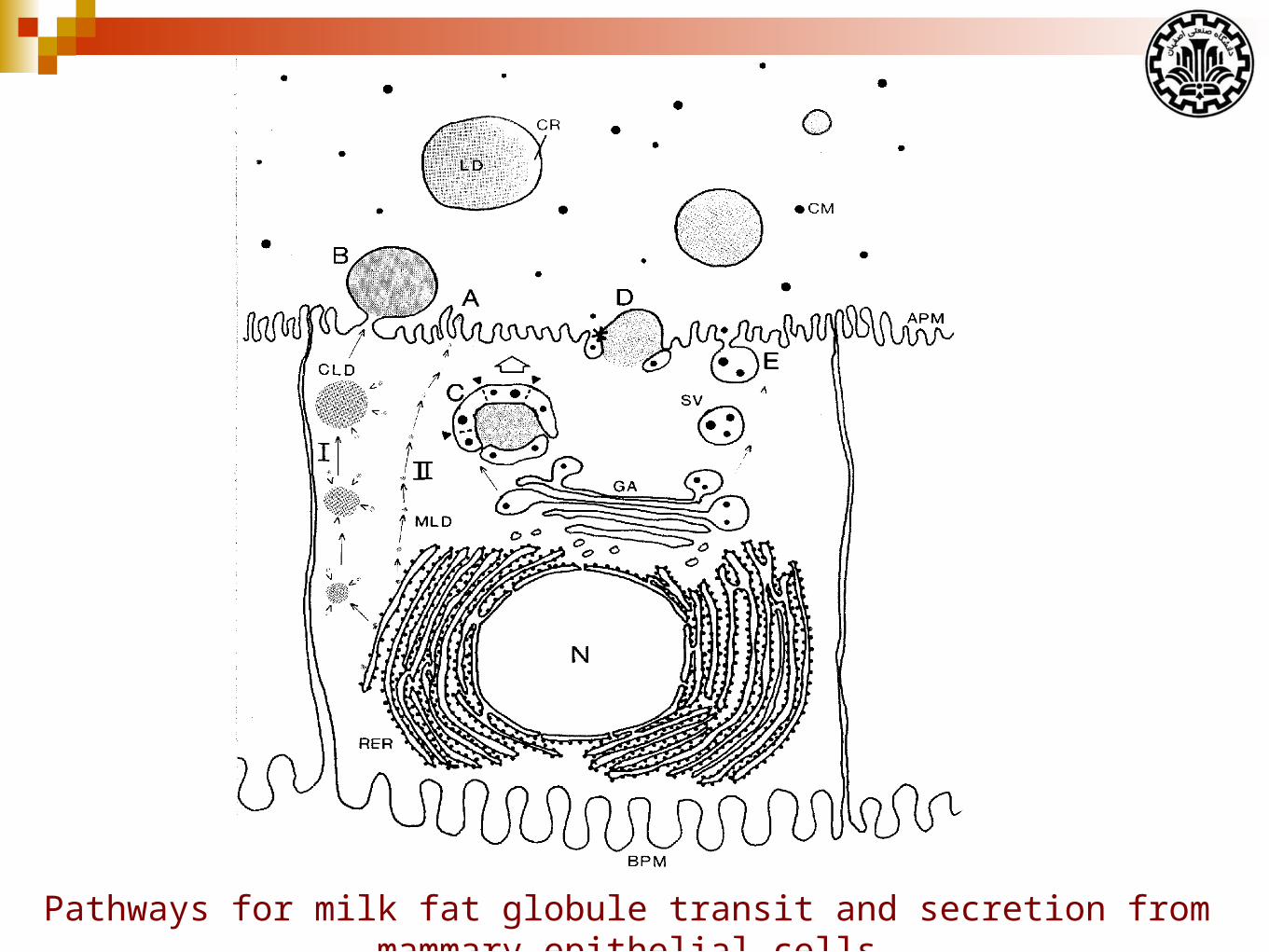
Pathways for milk fat globule transit and secretion from mammary epithelial cells

Synthesis of milk fat
The properties of milk fat:
Milk fat composed of different fatty acids:
Short chains (4-8 C)
Medium chains (10-14 C)
Long chains (≥16 C)

Synthesis of milk fat
The properties of milk fat:
TAG (more than 95% of milk fat)
DAG (2%)
Phospholipids (1%)
Cholesterol (0.5%)
FFA (0.1%)
Ether lipid, Fat soluble vitamns., etc.

Synthesis of milk fat
The properties of milk fat:
Saturated FAs (~70%)
Palmitic acid
Myristic acid
Stearic acid
Monounsaturated FA (~25%)
Oleic acid
Vaccenic acis
Polyunsaturated FA (~5%)

Synthesis of milk fat
There are two sources of FA for milk fat synthesis:
The de novo FA synthesis in mammary epithelial cells
Short chain (4-8 C)
Medium chain (10-14 C)
~ 50% of 16 C
Preformed FA uptake from blood circulation
~ 50% of 16 C
> 16 C

De novo fatty acid synthesis
In ruminants, the substrates for de novo FA synthesis in
mammary epithelial cells are:
Acetate produced by rumen fermentation
β- hydroxybutyrate produced by the rumen epithelium

Preformed fatty acid uptake
Long-chain FA taken up by the mammary gland are imported
from plasma:
Released from circulating lipoproteins by lipoprotein lipase
NEFA bound to albumin
There is evidence showing that the membrane transport of long-
chain FA is a facilitated process.
Some factors might play a role in FA uptake and transport:
Cluster of differentiation 36 (CD36)
Fatty acid binding protein 3 (FABP3)
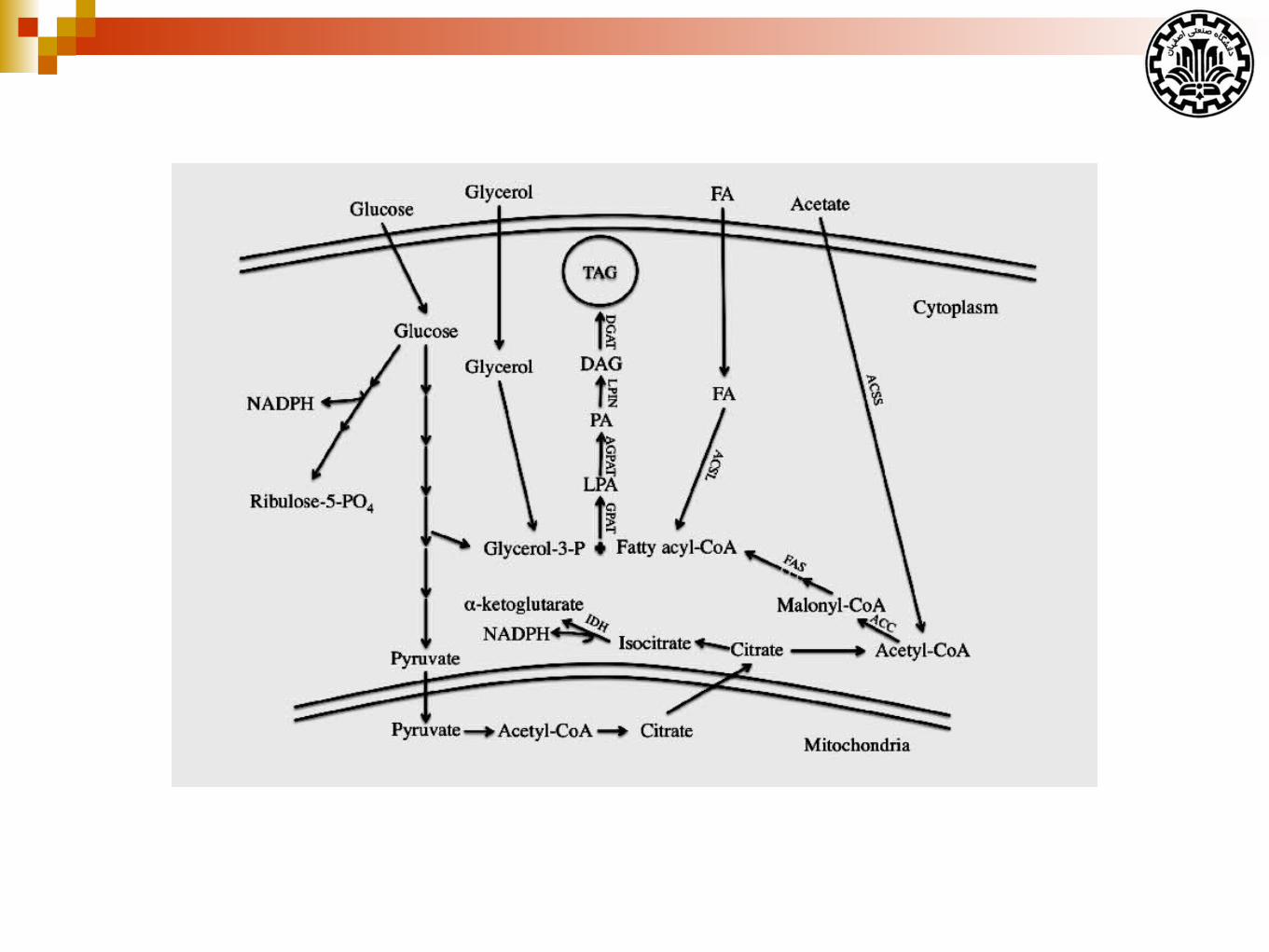

Properties of milk TAG
Fatty acids are not esterified randomly to the sn-1, -2, and -3
positions of glycerol backbone.
The distribution of FA is dependent on the distinct binding
affinities of the acyltransferase enzymes for substrate FA.
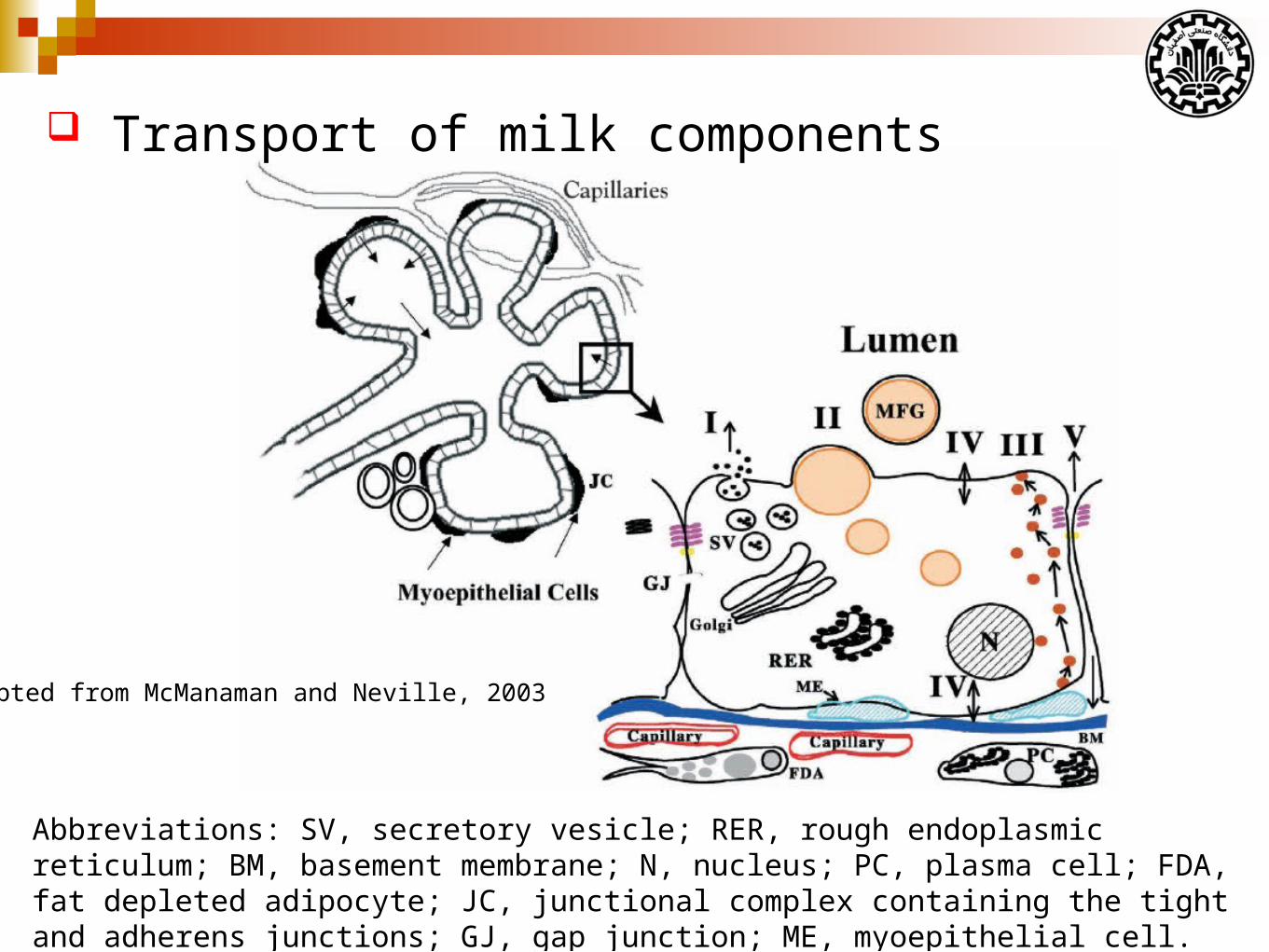
Adapted from McManaman and Neville, 2003
Abbreviations: SV, secretory vesicle; RER, rough endoplasmic reticulum; BM, basement membrane; N, nucleus; PC, plasma cell; FDA, fat depleted adipocyte; JC, junctional complex containing the tight and adherens junctions; GJ, gap junction; ME, myoepithelial cell.
Transport of milk components

Transport of milk components
Pathway I depicts exocytotic for:
Protein secretion by alveolar cells
Water
Lactose
Oligosaccharides
Phosphate
Calcium
Citrate

Transport of milk components
Pathway II depicts milk fat secretion.
Milk lipids, primarily triacylgycerides and phospholipids, are
synthesized in the smooth endoplasmic reticulum in the basal region
of the cell.
Newly synthesized lipid molecules form cytoplasmic lipid
droplets and are secreted by a unique budding process (MFGs).

Transport of milk components
Milk fat globule membrane is known to contain numerous
enzymes, including oxidases, reductases and hydrolases with
relatively high specific activities.
In particular milk fat globule membranes are highly enriched in
the purine oxidizing enzyme xanthine oxidoreductase (XOR).

Transport of milk components
Pathway III depicts transcytotic pathways for transport of
proteins and other macromolecules.
Transcytotic secretion of immunoglobulin A in rabbit mammary
glands has been shown to occur.
Prolactin and transferrin transcytosis have been detected
Transfer of labeled low-density lipoprotein (LDL) from blood to
milk has been reported.
Considering that xenobiotic agents, including carcinogens and some
drugs, can bind to and be transported by lipoproteins.

Transport of milk components
Pathway IV depicts transport of:
monovalent and polyvalent ions
glucose
amino acids

Transport of milk components
Ion transport: Transporters or channels for sodium, potassium and
chloride have been identified on the basal and apical plasma
membranes of alveolar cells.
Phosphate and iodide transporters appear to be limited to the basal
membrane.

Transport of milk components
Glucose transport: Glucose transport systems have been detected
in the mammary gland at both the apical and basal plasma
membrane, and on Golgi and secretory vesicle membranes.
Two distinct glucose transport mechanisms have been identified
in the mammary gland:
GLUT1 transporter mechanism
A sodium dependent glucose transporter

Transport of milk components
Amino acid transport: Both sodium-dependent and sodium
independent amino acid transport mechanisms analogous to those
found in other organs have been demonstrated at the basolateral
component of the mammary epithelium.
Other agents: The presence of higher than expected
concentrations of certain drugs in milk have raised the possibility
that alveolar cells may have active transport mechanisms for such
compounds.

Transport of milk components
Pathway V depicts transport the paracellular pathway for direct,
bi-directional, extracellular movement of both low-molecular-weight
substances and macromolecular solutes.
This pathway is closed during lactation in humans and most other
species by the presence of very tight-junction.

Milk fat depression (MFD)
Several theories have been proposed to explain the physiology
behind this reduction in fat synthesis.
Lower production of acetic and butyric acids in the rumen caused
less fat production in mammary gland.
The greater proportionate production in rumen increases the blood
insulin, which partitions nutrients away from the mammary gland.
A more current theory is that the combination of high grain and
high unsaturated fatty acids in the diet causes the microorganisms in
the rumen to produce more trans fatty acids.

Milk fat depression (MFD)
Avoiding milk fat depression
Proper cooling of cows
Control the amount of polyunsaturated fatty acids in the diet
Balance dietary carbohydrates
Buffer and alkalinizing agents
Ionophores
Feeding Management
Recommended

















