
Curr Genet (1995) 28:242-247 �9 Springer-Verlag 1995
O R I G I N A L P A P E R
A. Baur �9 N. Chalwatzis �9 A. Buschinger F.K. Zimmermann
Mitochondrial DNA sequences reveal close relationships between social parasitic ants and their host species
Received: 15 March 1995
Abstrac t In the tribe Leptothoracini, the phylogenetic relationship of socially parasitic ants (Doronomyrmex kutteri, D. 9oesswaldi and Harpagoxenus sublaevis) and their host species Leptothorax acervorum has been controversial. Even more controversial is the relation- ship between the socially parasitic ant Chalepoxenus muellerianus and its host species Leptothorax unifas- ciatus, L. nigriceps, L. interruptus and L. recedens. On the basis of morphological, ecological and ethological criteria it has been argued that socially parasitic ants and their respective hosts always evolved from com- mon ancestors, and hence it has been postulated that these species should be included in common taxo- nomical groups. This would require the division of the tribe Leptothoracini into two subgroups, one compris- ing the subgenus Leptothorax (s. str.) and the other the subgenus Myrafant, together with their respective para- sitic genera. We have used the polymerase chain reac- tion (PCR) to compare a 360-bp sequence of the mitochondrial cytochrome b gene of 14 species belong- ing to the tribe Leptothoracini and an outgroup species Tetramorium impurum (Tetramoriini). The results gen- erally agree with the morphological studies which sug- gest that a common ancestral species differentiated into host and parasite species. This relationship is very obvious within the Leptothorax (s. str.) group but less pronounced in the species belonging to the Myrafant group. Leptothorax (Temnothorax) recedens shows a greater sequence divergence than the outgroup spe- cies T. impurum.
A. Baur �9 F.K. Zimmermann ( ~ ) Institut ffir Mikrobiologie, Technische Hochschule Darmstadt, Schnittspahnstrasse 10, D-64287 Darmstadt, Germany
N. Chalwatzis �9 A. Buschinger Institut ftir Zoologie, Technische Hochschule Darmstadt, Schnitt- spahnstrasse 3, D-64287 Darmstadt, Germany
Communicated by K. Wolf
K e y words L e p t o t h o r a c i n i - Social parasitism - Cytochrome b �9 mtDNA �9 PCR
Introduction
Social parasitism is a fascinating phenomenon ob- served in the Formicidae where it has evolved conver- gently several times in a number of subfamilies. Three types of true social parasitism, namely temporary para- sitism, dulosis (slavery), and inquilism are distinguished (Buschinger 1986; H611dobler and Wilson 1990). In all instances two or more species live together in one colony in such a way that the host species support the reproduction of the parasite. Morphological similarit- ies and the fundamental necessity of coordinated inter- actions and communication between the species led to the suggestion that a parasite and its respective host(s) always evolved from one common free-living ancestral species. The closest relatives of a parasitic species or genus should thus be found among its host-species group (Emery's rule; Le Masne 1956).
Two competing hypotheses exist on the mode of host-parasite speciation. The allopatric model suggests transient geographical isolation and, after speciation had occurred, the transition of one of the daughter species to parasitism of the other (Wilson 1971; HN1- dobler and Wilson 1990). The sympatric model (Bu- schinger 1970, 1990; Elmes 1978; Bourke and Franks 1991), suggests that a parasite evolves directly from its future host, gaining reproductive isolation through the assortative mating of sexuals which either remain close to the mother nest, or leave for swarming. The latter model fits best to the inquiline species which normally parasitize but one host species. Dulotic or slave-making ants, on the other hand, always have a range of several host species, though these, too, are close relatives of their respective slave-makers. In this group, however, an ex- tension of the host species range after the evolution of the parasitic life habit may sometimes have occurred.

243
Both inquilinism and slavery are common and have been extensively studied in the myrmicine tribe Lep- tothoracini. According to Buschinger (1981) this tribe comprises at least two very different groups (in the following: Leptothorax s. str.- and the Myrafant-group). Inquilinism and slavery have evolved several times independently in both groups (Buschinger 1990). In the present paper we have studied the free-living Lep- tothorax (s. str.) species L. acervorum and L. muscorum, and the inquilines Doronomyrmex kutteri and D. goes- swaIdi, both parasitizing exclusively L. acervorum. These species belong to the "Leptothorax s. str.-group". The "Myrafant-group" is represented by the free-living Leptothorax (Myrafant) unifasciatus, L. (M.) nigriceps and L. (M.) interruptus, and by the slave-maker Chalepoxenus muellerianus. The latter parasitizes a wide range of L. (M.) species including the three mentioned above. The second frequent host after L. unifasciatus is Leptothorax recedens, which is assigned by some authors to the subgenus Myrafant, whereas others place it in its own subgenus or even in the genus Temnothorax. As an outgroup species we chose Tet- ramorium impurum, belonging to the distantly related myrmicine tribe Tetramoriini.
We have started to employ DNA sequencing methods to supplement morphological and behavioral criteria in the study of the evolution of social parasitism (Baur et al. 1993). We sequenced a 1.2-kb segment of the 18 rRNA genes and found that they differed at less than 1% of the positions.
The mitochondrial gene for cytochrome b is ma- ternally inherited and contains sequences well conser- ved in the animal kingdom (Kocher et al. 1989; Thomas and Beckenbach 1989; Irwin et al. 1991; Avise et al. 1992; Baker et al. 1993; Beard et al. 1993; Crozier and Crozier 1993; Jermiin and Crozier 1994). Therefore, sections of this gene can be amplified from species of interest by the polymerase chain reaction (Mullis and Faloona 1987) and sequenced. We have used this ap- proach to investigate cytochrome b sequence polymor- phisms in the Leptothorax s. str. and the Myrafant group in an attempt to elucidate the phylogenetic rela- tionships between parasitic and host species. The re- sults indicate that some parasite systems could have arisen by a diversification of an ancestral species into host and parasite species. However, an evolutionary adapatation of a parasite to at least one distant host species, Leptothorax (Temnothorax) recedens, is sugges- ted for the slave-maker Chalepoxenus muellerianus.
Materials and methods
We used ant pupae for DNA extraction in order to reduce contami- nation through the gut content. The pupae were taken from laborat- ory stock colonies. For the main host species of the Leptothorax s. str. group, L. acervorum, we compared material from four geographi- cally distant populations: R (near Nuremberg, central Germany);
P (Pyrenees, France); S (a population with behavioral and slight morphological differences from near Teruel, Spain); J (Hokkaido, Japan). The inquilines D. ku~teri and D. goesswaldi were collected in the Alps (Valais, Switzerland); the slave-maker Harpagoxenus sub- laevis, and its second host species L. muscorum, in central Germany (near Nuremberg). The slave-making C. muellerianus and its host species L. (Myrafant) unifasciatus and L. recedens were collected in Northern Italy (Lago di Garda), the independent L. interruptus and L. nigriceps originated from France (Vercors). The outgroup species T. impurum was found in Switzerland (Valais).
Nucleic acid techniqes. High-molecular-weight DNA was prepared from pupae as previously described (Baur et al., 1993). The standard nucleic acid techniques of Sambrook et al. (1989) were followed. PCR primers for the amplification of a 360-bp fragment were de- signed using the published cytochrome b sequences of Drosophila yakuba (Clary and Wolstenholme 1985) and Apis mellifera (Crozier and Crozier 1993) in comparison with sequences from vertebrates (Kocher et al. 1989). The primer used for the amplification of the conserved region of the cytochrome b gene at the Y-end was GAA TTC ATT TAC CA(G/A) CTC CA(A/G) T(A/T) AAT ATT-3'. The primer for the T-end was 5' CGG GAT CCC TCA (G/A)(A/T)A TGA TAT TTG TCC TCA 3'. In addition to the expected cyto- chrome b gene sequences the primers contained the restriction sites EcoRI and BamHI respectively as indicated by the bases written in italics. One hundred nanograms of genomic DNA was amplified by PCR in a Biometra thermocycler in 35 cycles with 1.25 units of Taq polymerase (Boehringer Mannheim) following the protocol of Saiki et al. (1985) using a hot start. The cycles consisted of 45 s at 95 ~ for denaturation, followed by 45 s at 45 ~ for annealing and 45 s at 72~ for extension for five cycles, then 30 cycles in which the annealing temperature was elevated to 50 ~ We followed the rec- ommendations of Kwok and Higuchi (1989) to avoid false-positive amplification products. Amplified sequences were separated by agarose-gel electrophoresis, and the desired bands cut out and DNA extracted using the Geneclean-kit (Dianova, Hamburg). DNA was digested with restriction endonucleases generating cohesive ends and cloned into the vector pBIuescript II (Stratagene, Heidelberg). Escherichia coli strain SURE (Stratagene) was transformed by elec- troporation using a BioRad Genepulser. Plasmids were purified using the Qiagen kit (Diagen, Dfisseldorf). The cloned cytochrome b fragments were sequenced on both strands using the Sanger et al. (1977) 2-dideoxy chain-terminating method. DNA sequences were analyzed using the DNASIS computer program (Pharmacia).
Phylogenetic analysis. Was performed using the program package PHYLIP 3.5c (Felsenstein 1989, 1993). The sequences were aligned and trees constructed by the neighbor-joining method (Saitou and Nei 1987) and the maximum parsimony method (Eck and Dayhoff 1966; Kluge and Ferris 1969; Fitch 1971). Distances between DNA sequences were calculated according to Jukes and Cantor (1969) and bootstrap analysis was after Felsenstein (1985).
Results
All the PCR reactions yielded a single distinct band. Three different cloned fragments were sequenced for each species because of possible amplification errors during PCR (Saiki et al. 1985; Keohavong and Thilly 1989). We found two clones which differed in the num- ber of T-residues (one deletion and one insertion) in the positions between 88 and 95 in the PCR products of L. muscorum and L. unifasciatus. These insertion/ deletion differences caused frameshifts which led to completely altered amino-acid sequences in the
244
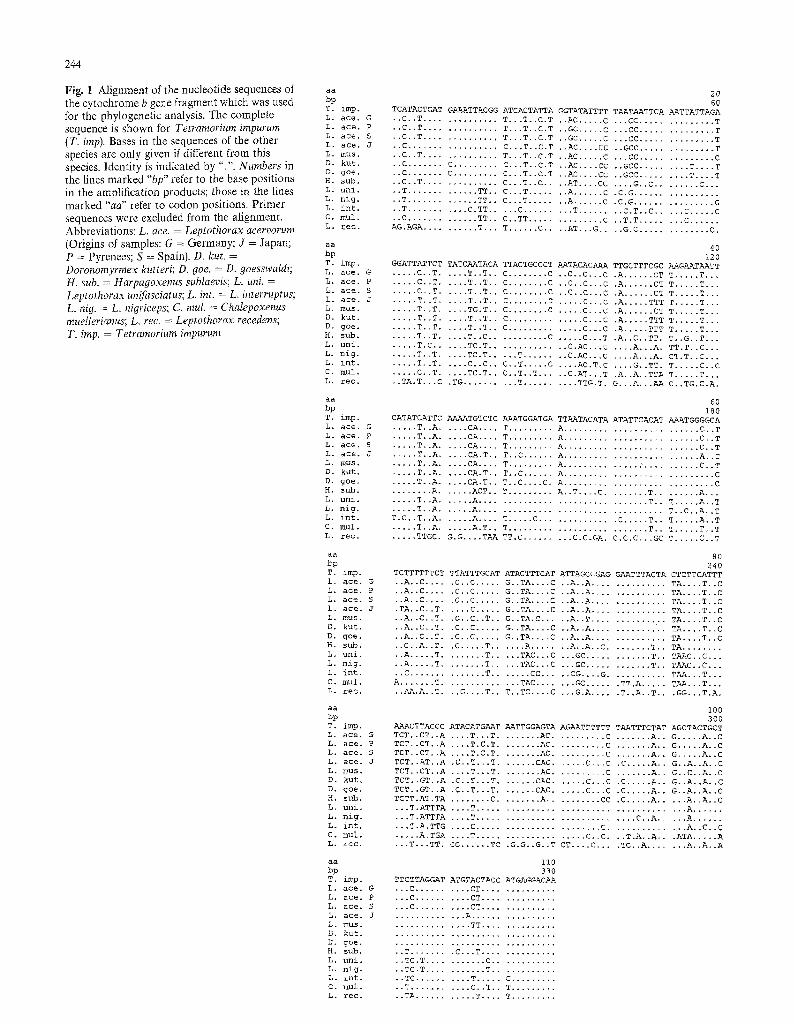
Fig, 1 Alignment of tbe nucleotide sequences of the cytochrome b gene fragment which was used for the phylogenetic analysis. The complete sequence is shown for Te~ramorium impurum (T. imp). Bases in the sequences of the other species are only given if different from this species. Identity is indicated by ".". Numbers in the lines marked "bp" refer to the base positions in the amplification products; those in the lines marked "aa" refer to codon positions, Primer sequences were excluded from the alignment, Abbreviations: L. ace. = Leptothorax acervorum (Origins of samples: G = Germany; 3 = Japan; P = Pyrenees; S = Spain)�9 D. kut. = Doronomyrmex kut~eri; D. goe. = D, goesswaldi; H. sub. = Harpagoxenus sublaevis; L. uni. = Leptothorax un~sc iams; L. int. = L. interruptus; L. nig. = L. nigriceps; C. mul. = Chatepoxenus mueIlerianus; L . rec. = Leptothorax recedens; T. imp. = T e t r a m o r i u m impurum
aa
bp
T. imp.
L. ace.
L. ace.
L. ace.
L. ace.
L. mus.
D. kut.
D. goe.
H. sub.
L. uni.
L. nig.
L. int.
C. mul.
L. rec.
aa
bp
T. imp.
L. ace. G
L. ace. P
L. ace. S
L. ace. J
L. mus.
D. kut.
D. goe.
K. sub.
L �9 un•
L. nig.
L, int.
C. mul.
L. rec.
aa
bp
T. imp.
L~ ace. G
L, ace. P
L. ace. S
L. ace. J
L. mus,
D. kut.
D. goe.
H, sub.
L. uni.
L. nig,
L. int.
C. mul.
L. rec.
aa
bp
T. imp.
L. ace. G
L. ace. P
L. ace. S
L. ace. J
L. mus.
D. kut.
D, goe.
H. sub.
L. uni.
L. nig.
L. int.
C. mul.
L. rec.
aa
bp
T. imp.
L. ace. G
L. ace. P
L. ace. S
L. ace. J
L, mus.
D, kut.
D. goe.
H. sub.
L. uni.
L. nig.
L. int,
C. mul.
L. rec.
aa
bp
T, imp.
L. ace. G
L. ace. P
L. ace. S
L. ace. J
L. mus.
D. kut.
D. goe.
H. sub.
L. uni.
L. nig~
L. int.
C. mul.
L. rec.
20
6O
TCATACTGAT GAAATTACGG ATCACTATTA GGTATATTTT TAATAATTCA AATTATTAGA
.C..T .............. T...T..C.T .AC ..... C ..,CC .............. T
.C..T .............. T...T..C.T .GC ..... C ...CC .............. T
.C..T .............. T...T..C.T .GC ..... C ...CC .............. T
.C ................. C...T..C.T .AC .... CC ..GCC .............. T
.C..T .............. T,..T..C.T .AC ..... C .,.CC .............. C
.C ....... C ......... C..,T.,C,T ,AC .... CC ..GCC ......... T .... T
.C ....... C ......... C.. .T..C.T .AC .... CC ..GCC ......... T .... T
.C..T ..............
.T ............. TT..
,T ............. TT. ,
�9 T ........... C.TT..
.C ............. TT..
AG .AGA .......... T...
GGATTATTCT
..... C..T.
..... C..T,
..... C..T.
..... T.,T.
..... T..T.
..... T..T.
..... T..T.
..... T..T.
..... T.C..
..... T..T.
..... T,.T.
..... C..T.
..TA.T..,C
C. . .T. .C. , .AT .... CC .... G. .C ........ C. . .
C...T ....... A ...... C .C.G ................
C, . .T ....... A ...... C .C.G ............... G
�9 . .C ......... T ........ C.T..C ..... C ..... C
C. .TT .............. C . ,T.T ........ C ......
T ...... C .... AT...G .... G.C ............. C.
4O
120
TATCAATACA TTACTGCCCT AATACACAAA TTGCTTTCGC AAGAATAATT
.... T. .T. . C ........ C . .C. .C. . .C .A ...... CT T ..... T. . ,
.... T. .T.. C ........ C ..C..C.-.C .A ...... CT T ..... T...
.... T..T.. C ........ C . .C. ,C., .C .A ...... CT T ..... T, ,.
.... T.,T.. C ........ C ..... C.. ,C ,A ..... TTT T ..... T. ,.
.... TG.T.. C ........ C ..... C.-.C ,A ...... CT T ..... T...
.... T..T.. C .............. C.-.C ,A ..... TTT T ..... T.. �9
.... T..T., C .............. C-..C .A ..... TTT T ..... T...
.... T..C ........... C ..... C...T .A..C..TT. T,.G.,T...
.... TC.T .............. C.AC...C .... A...A. TT.T..C...
.... TC.T..
.... C..C.,
.... TC.T..
�9 TG .......
CATATCATTC AAAATGTCTC
..... T..A ..... CA ....
..... T..A ..... CA ....
..... T..A ..... CA ....
..... T..A ..... CA.T �9 .
..... T..A . . . . . CA ....
..... T..A ..... CA.T..
..... T. .A ..... CA.T. �9
...T ........ C.AC.. ,C .... A...A. CT.T..C...
C. .T ..... C .... AC.T.C .... G. .TT. T ..... C. .C
C..T..T ..... C.AT...T .A..A..TTA T ..... T...
�9 .......... TTG.T. G...A...AA C, .TG.C.A.
6O
180
AAATGGATGA TTAATACATA ATATTCACAT AAATGGGGCA
T ......... A ......................... C..T
T ......... A ......................... C,.T
T ......... A ......................... C..T
T..C ...... A . . . . . . . . . . . . . . . . . . . . . . . . . A..C
T ......... A .............. : .......... C..T
T. �9 ...... A ............................ C
T..C .... C. A ............................ C
........ A ...... ACT.. T ......... A..T .... C ........ T ........ A, ..
..... T..A ...... A ............................... T.. T ..... A~.T
..... T..A ...... A .................................. T..C..A.,T
T.C, .T..A ...... A .... T ..... C .............. C ..... T,, T ..... A.,T
..... T..A ...... A.T., T .......................... T,. T ..... T..T
..... TTGC. G.G .... TAA TT.C ......... C.C.GA. C.C.C...GC T ..... C..T
8O
240
TCTTTTTTCT TTATTTGCAT ATACTTTCAT ATTAGCCGAG GAATTTACTA CTCTTCATTT
..A..C ..... C..C ..... G..TA .... C ,A..A .............. TA .... T..C
..A.,C ..... C..C ..... G.,TA .... C ,A..A .............. TA .... T..C
~.A..C ..... C..C ..... G..TA .... C .A..A .............. TA .... T.,C
.TA..C..T ..... C ..... G..TA .... C .A..A .............. TA .... T..C
..A..C..T. .C.,C..T.. G..TA.C .... A..T .............. TA .... T..C
..A..C..T, .C.,C ..... G..TA .... C ,A.~A .............. TA .... T..C
. .A..C..T, .C.,C ..... G..TA .... C ,A..A .............. TA .... T..C
..C..A..T. .C ..... T ...... A ....... A..A.,C ........ T.. TA ........
..A ..... T ........ T ..... TAC,..C ...GC ............ T.. TAAC,.C...
~,A ..... T ........ T ..... TAC...C .,.GC ............ T.. TAAC..C...
~.C .............. T ....... CC ..... CG .... G ........... TAA.. oT...
A ....... T .............. TAC ....... GC ...... TT.A ..... TAA...T...
..AA.A.~T �9 ..G .... T.. T..TC .... C ,,.G.A ..... T..A..T �9 .GG...T.A.
I00
3O0
AAACTTACCC ATACATGAAT AATTGGAGTA AGAATTTTTT TAATTTCTAT AGCTACTGCT
TCT..CT. �9 .... T...T ........ AC .......... C ....... A.. G ..... A..C
TCT..CT. �9 .... T.C.T ........ AC .......... C ....... A.. G ..... A,,C
TCT..CT, .A .... T.C.T ........ AC .......... C ....... A,, G ..... A.,C
TCT..AT..A .C �9 ....... CAC ...... C,.,C .C ..... A,, G..A, �9
TCT.. CT..A .... T...T ........ AC .......... C ....... A �9 G,,C. ~A..C
TCT. �9 .C..T...T ....... CAC ...... C. �9 .C ..... A,. G. �9
TCT..GT..A .C..T.-.T ....... CAC ...... C...C .C ..... A,. G..A..A..C
TCTT.AT.TA ........ C ........ A .......... CC .C ..... A ..... A �9 �9
�9 . . T �9 .... T ...................................... A ......
�9 . . T.ATTTA .... T ............................. C..A ..... A ......
�9 . , T.A. TTG .... C ....................... C .............. A. .C. -C
..... A.IGA .... T .................... C. -C. . .T.A. .A, - .ATA ..... A
�9 ..T.. .TT. CC ...... TC .G.G..G.,T CT .... C .... TC. �9 ....... A..A,.A
ii0
330
TTCTTAGGAT ATGTACTACC ATGAGGACAA
� 9 . . . . . . . . . . CT . . . . . . . . . . . . . .
�9 . .C . . . . . . . . . . CT . . . . . . . . . . . . . .
� 9 .......... CT ..............
............. A ................
.............. TT ..............
.TC.T . . . . . . . . . . . C . . . . . . . . . . . .
.TC.T . . . . . . . . . . . T . . . . . . . . . . . .
-TC .......... T ..... C .........
.T ........... C..T.. T .........
.TA ........... T .... T .........

predicted proteins and thus could be readily identified as amplification artifacts
The base sequences are shown in Fig. 1. Except for the two amplification artifacts mentioned above all amplified fragments were of the same length The AT content of the cloned mitochondrial gene fragments ranges from 66.9% for D. kutteri to 74.5% for C. muel- lerianus. This is close to the range also reported for D. yakuba, 73.9% (Clary and Wolstenholme 1985), A. mellifera, 80.0%, (Crozier and Crozier 1993) and the cytochrome b region of the ant Tetraponera rufoniger (Jermiin and Crozier 1994)
Using the code established for D. yakuba by Clary and Wolstenholme (1985), and applied to A. mellifera by Crozier and Crozier (1993), the base sequences of the amplification products of all 14 species could be trans- lated into an amino-acid sequence which is shown in Fig. 2
The DNA sequences were used to construct a phylogenetic tree with the neighbor-joining method employing the distances obtained by the method of Jukes and Cantor (1969) after 100 bootstrap replica- tions (Felsenstein 1985) (Fig 3 A). The branch lengths of this tree were calculated with the Fitch-Margoliash (1967) algorithm based on distances derived from the original data set (Jukes and Cantor 1969) (Fig. 3 B)
The species belonging to the Leptothorax s str.- group form a monophyletic group obtained in all boot- strap replications thus showing that the host species and their parasites D kutteri, D. goesswaldi and H. sublaevis derive from a common ancestor The two Doronomyrmex species and the Japanese Leptothorax acervorum isolate form a group separated from the European L. acervorum and L. muscorum
The two species L. unifasciatus and L. nigriceps ap- pear closely related whereas much lower bootstrap
245
values were obtained for the taxonomic positions of L. interruptus and C. muellerianus at the DNA level. However, the tree constructed on the basis of amino- acid sequences (using maximum parsimony) places all the Myrafant species into one group with good boot- strap values (Fig. 4). L. (Temnothorax) recedens did not group together with the Myrafant group either at the DNA or at the amino-acid sequence level. The se- quence divergences between this and the other species are unexpectedly high (Fig. 3 A, B).
Discussion
The phylogenetic problem to be solved was to test the hypothesis that parasitic species and their hosts evol- ved from common parental species groups This is obvious for the species within the Leptothorax s str.- group as documented by both the base (Fig. 1), and the predicted amino-acid, sequences (Fig. 2) and is also reflected in the dendrograms (Figs 3 and 4).
The L. acervorum isolates from Teruel and the Pyrenees are identical They are different, however, from the German, and still more different from the Japanese, isolates. Interestingly, the two Doronomyr- mex species share certain unique base and amino-acid positions with the Japanese isolate. This could indicate that the parasitic species originated in eastern Eurasia, though both are known as yet only from western Euro- pe, whereas a Harpagoxenus species (H zaisanicus, probably synonymous to H sublaevis) has been de- scribed from Mongolia H. sublaevis quite obviously deviated earlier from Leptothorax s. str than the Doronomyrmex species. This means that the ability to parasitize on L acervorum and/or L. muscorum evolved independently at different times.
Fig. 2 Alignment of the predicted amino-acid sequences. The complete sequence is given for ~ . a c e . G
L. ace. P
T impururn. Amino acids in the sequences of T . a c e . S the other species are only given where different ~. ace. J
L. mus. from that of this species. Identi ty is indicated D. kut. by . . . . The top numbers refer to the amino- o. goe. acid posit ions in the predicted section. H. sub.
L. uni.
Abbreviat ions are the same as in Fig. 1 L. nig. L. int.
C. mue.
L. rec.
T. imp.
L. ace. G
L. ace. P
L. ace. S
L. ace. J
L. mus.
D. kut.
D. goe.
H. sub.
L. uni.
L. nig.
L. int.
C. mue.
L, rec.
T. imp.
60
SYWWNYGSLL GLFLTIQIIS GFFLSIHYCP NTQLAFLSII HIMQNISNGW MMHNIHMNGA
.............. A ..................... F .......................
......................... V ..................................
�9 . .S.. ........ A...F ................. F .......................
�9 . .S .......... A...F ................. F ............ S ..........
.............. M ..................... SR ....... T ..... L ........
..... F ..... M..S ....... S..L ..... N .... Te ............ L ..... I...
..... F ..... M..S .... R ..... L ..... N .... TC ............ L ..... I...
..... F ........ L ................ NL...S... Y ......... L ..... I...
..... F ..... M..F .......... L ..... N .... Y ............. L ..... I...
.S...F ...... C.V .... T .I..AM ..... IDS..N.VN ..CRDVNy.. LLRTL.A...
........... M..M ...... L...M ....... I..A.M. ..I..V .... L .........
ii0
SFFFICMYIH MSRGIYYTSF SLSNTWLIGT SIFLISMATA FLGYVLpWGQ
......................... S ........................
......................... S ........................
L ........................................... E .....
:;2:111~[[ [[i[iiiiii i[iiiiiii~iiii~i[[[i iii;i[[i[[ . . . . . . . . T . I N . . . . . N . . K . F . . . M . . V . . . . . . . . . . . . . . . . . . . .
. . . . . . . . T . I A . . . . . N . . K . F . . . M . . V . . . . . . . . . . . . . . . . . . . .
. . . . . . . . S . I G . . . . . N . . K . I D . . M . . V . . . . . . . . . . . . . . . . . . . .
T . . . . . . . T . I A . . L . . N , . I < . i ' 4 , . . M , . V . . . . L . . M . . . . . . . . . . . .
. M . . V . I . L . I G . . M . . G . Y K F I P . . S V . V L . L F L T . . . . . M . . . . . . . .
. . . . . . . . F . I . . . . . . S . . I < . T H . , M . . V . . . . . . . . . . . . . . . . . . . .

246
A Leptethorax muscorum
~ Leptothorax acervorum - Germany Leptothorax acervorum - France Leptothorax aeervorum - Spain Leptothorax acervorum - Japan
Doronomyrmex geesswaldi Deronomyrme• kutteri Harpagoxenus sublaevis Chalepoxenus muellefianus Leptethorax interruptus
"'1 63 L,-]"- Leptothorax nigriceps I 100 I.. Leptothorax urlifasciatus
I --;:;22r;C22m B ,-- Leptothorax muscorum
"[iLeptothorax acervorum - Germany I --[iLeptothorax aeervorum - France
I I Leptothorax acervorum - Spain
L F Leptothorax acervorum - Japan
I "--LlDoronomyrmex goesswaldi
rl I I .I Chalepo• muellerianus
Li l Leptethorax interruptus
~ Leptothorax nigriceps
i !.- Leptethorax unifasciatus
I Tetramoriurn impurum Leptothorax recedens
Fig. 3 Phylogenetic tree, constructed with the Neighbor joining method. Distances between the DNA sequences were calculated with the Jukes and Cantor (1969) algorithm. T. irnpurum was used as an outgroup species. A the numbers at the branches are bootstrap values (100 replications). The branch lengths are mandatory. B the branch lengths were calculated with the Fitch-Margoliash algorithm based on distances of the original data set
Doronomyrmex and H. sublaevis do not share any particular amino acids except for those common to all the Leptothorax s. str. group. Thus a closer relationship between these two genera cannot be inferred.
The sequences obtained in the Myrafant group are rather confusing. C. muellerianus parasitizes on the three Leptothorax species, unifasciatus, interruptus and nigriceps. L.unifasciatus and nigriceps differ from each other at only three amino-acid positions and thus ap- pear closely related. L. interruptus surprisingly differs at 12 positions from the amino-acid sequence of the two previous species, though it is morphologically very similar to L. unifasciatus.
The fourth host of Chalepoxenus is L. (Temnothorax) recedens which differs from the three Leptothorax (Myrafant) and all the other species at 38 amino-acid positions. All the other species share the same amino acid at 23 of these positions. Consequently, C. muel- Ierianus parasitizes a spectrum quite different of host species.
~ Leptothorax muscorum
Leptothora• acervorum - Germany
Leptothorax acervorum - France Leptothorax acervorum - Spain
Leptothorax acervorum - Japan
Dorenomyrmex goesswaldi Doronomyrmex kutteri Harpagoxenus sublaevis Leptothorax interruptus Chalepoxenus muellerianus Leptothorax nigriceps Leptothorax unifasciatus
Leptothorax recedens -- Tetramorium impurum
Fig. 4 Phylogenetic tree based on the predicted protein sequences and constructed with maximum parsimony (PROTPARS). The numbers at the branches are bootstrap values (100 replications)
L. unifasciatus and nigriceps share common differ- ences to all other species, including Chalepoxenus, at four positions. However, Chalepoxenus shares three unique amino-acid positions with these two host spe- cies. This would indicate a relatively recent diversifica- tion into host and parasite species, and an origin of ChaIepoxenus from this group of Myrafant species.
L. (Temnothorax) recedens deviates so much at the amino-acid and DNA sequence levels from its parasite species that a close phylogenetic relationship is not obvious. Even the outgroup species T. impurum shows a higher level of sequence identity with ChaIepoxenus and its Myrafant host species. Apparently, C. muel- lerianus managed to extend its parasitic relationships beyond the relatives of its ancestral species to enslave a quite remotely related additional host.
References
Avise JC, Bowen BW, Lamb T, Meylan AB, Bermingham E (1992) Mitochondrial DNA evolution at a turtle's pace: evidence for low genetic variability and reduced microevolutionary rate in the Testudines. Mol Biol Evol 9:457-473
Baker CS, Perry A, Bannister JL, Weinright MT, Abernethy RB, Calambokidis J, Lien J, Lambertsen RH, Urbfin Ramirez J, Vasquez O, Clapman PJ, Alling A, O'Brien SJ Palumbi SR (1993) Abundant mitochondrial DNA variation and world-wide popu- lation structure in humpback wales. Proc Natl Acad Sci USA 90:8239-8243
Beard CB, Mills Hamm D, Collins (1993) The mitochondrial genome of the mosquito Anopheles 9ambiae: DNA sequence, genome organization, and comparisons with mitochondrial se- quences of other insects. Ins Mol Biol 2:103-124
Baur A, Buschinger A, Zimmermann FK (1993) Molecular cloning and sequencing of 18s rDNA fragments from six different ant species, Ins Soc 40:325-335
Bourke AFG, Franks NR (1991) Alternative adaptations, sympatric speciation and the evolution of parasitic, inquiline ants. Biol J Linn Soc 43:157 178.
Buschinger A (1970) Neue Vorstellungen zur Evolution des Sozial- parasitismus und der Dulosis bei Ameisen (Hym., Formicidae). Biol Zbl 88:273-299

247
Buschinger A (1974) Experimente und Beobachtungen zur Griin- dungund Entwicklung neuer Sozietfiten der sklavenhaltenden Ameise Harpagoxenus sublaevis (NyI.). Ins Soc 21:381-406
Buschinger (1981) Biological and systematic relationships of social- parasitic Leptothoracini from Europe and North America. In: Howse PE, C16ment J-L, (eds), Biosystematics of social insects. Academic Press, London New York, pp 211 222
Buschinger A (1986) Evolution of social parasitism in ants. Trends Ecol Evol 1:155-160
Buschinger A (1990) Sympatric speciation and radiative evolution of socially parasitic ants--heretic hypotheses and their factual background. Z Zool Syst Evolut-forsch 28:241 260
Clary DO, Wolstenholme DR (1985) The mitochondrial DNA mol- ecule of Drosophila yakuba: nucleotide sequence, gene organiza- tion, and genetic code. J Mol Evol 22:252 271
Crozier RH, Crozier YC (1993) The mitochondrial genome of the honeybee Apis mellifera: complete sequence and genome organ- ization. Genetics 133:97-117
Eck RV, Dayhoff MO (1966) Atlas of protein sequences and struc- ture. National Biomedical Research Foundation, Silver Spring, Maryland
Elmes GW (1978) A morphometric comparison of three closely related species of Myrmica (Formicidae) including a new species from England. Syst Entomol 3:131-145
Felsenstein J (1985) Coinfidence limits on phylogenies: an approach using the bootstrap. Evolution 39:783 791
Felsenstein J (1989) PHYLIP phylogeny inference package (ver- sion 3.2). Cladistics 5:164-166
Felsenstein J (1993) PHYLIP (phylogeny inference package) version 3.5c. Distributed by the author, Department of Genetics, Univer- sity of Washington, Seattle
Fitch WM (1971) Toward defining the course of evolution: min- imum change for a specified tree topology. Syst Zoo120:406-416.
Fitch WM, Margoliash E (1967) Construction of phylogenetic trees. Science 155:279-284
H611dobler B, Wilson EO (1990) The ants. The Belknap Press of Harvard University Press, Cambridge
Irwin IM, Kocher TD, Wilson AC (1991) Evolution of the cyto- chrome b gene of mammals. J Mol Evol 32:128-144
Jermiin LS, Crozier RH (1994) The cytochrome b region in the mitochondrial DNA of the ant Tetraponera rufoniger: sequence divergence in hymenoptera may be associated with nucleotide content. J Mol Evol 38:282-294
Jukes TH, Cantor CR (1969) Evolution of protein molecules. In: Munoro HN (ed), Mammalian protein metabolism. Academic Press, New York, pp 21 132
Keohavong P, Thilly WG (1989) Fidelity of DNA polymerase in DNA amplification. Proc Natl Acad Sci USA 86:9253 9257
Kluge AG, Farris JS (1969) Quantitative phyletics and the evolution of anurans. Syst Zool 18:1-32
Kocher TD, Thomas WK, Meyer A, Edwards SV, P/i/ibo S, Villab- lanca EX, Wilson AC (1989) Dynamics of mitochondrial DNA evolution in animals: amplification and sequencing with conser- ved primers. Proc Natl Acad Sci USA 86:6196 6200
Kwok S, Higuchi R (1989) Avoiding false positives with PCR. Nature 339:237 238
Le Masne G (1956) Recherches sur les fourmis parasites Plagiolepis grassei et l'Svolution des PlagioIepis parasites. C R Acad Sci Paris 243:673 675
Mullis KB Faloona KB (1987) Specific synthesis of DNA in vitro via polymerase chain reaction. Methods Enzymol 155:335 350
Saiki RK, Scharf S, Faloona F, Mullis KB, Horu GT, Erlich HA, Arnheim N (1985) Enzymatic amplification of 13-globin sequences and restriction-site analysis for diagnosis of sickle-cell anemia. Science 230:1350-1354
Saitou N, Nei M (1987) The neighbor-joining method: a new method for constructing phylogenetic trees. Mol Bio Evol 4:406 425
Sambrook J, Fritsch EF, Maniatis T (1989) Molecular cloning: a laboratory manual, 2nd edn. Cold Spring Harbour Laboratory, Cold Spring Harbor, New York
Sanger FS, Nicklen S, Coulson M (1977) DNA sequencing with chain-termination inhibitors. Proc Natl Acad Sci USA 162: 5463 5467
Thomas WK, Beckenbach AT (1989) Variation in salmonid mitochondrial DNA: evolutionary constraints and mechanisms of substitution. J Mol Evol 29:233-245
Wilson EO (1971) The insect societies.The Belknap Press of Harvard University Press, Cambridge
Recommended

















