
Pflanzenphysiologie 6: Blüten und Befruchtung
Auslösen der Blütenbildung
Identität des Apikalmeristems
Befruchtung und Embryogenese
Biologie I: Pflanzenphysiologie WS 2009/2010
Rüdiger HellHeidelberger Institut für Pflanzenwissenschaften
Copyright Hinweis:Das Copyright der in dieser Vorlesung genannten Lehrbücher oder reproduzierten Bilder wird anerkannt.Die Reproduktion dient reinen Lehrzwecken.
Tageslänge ist ein bestimmender Faktor derBlühinduktion
Nover/Weiler17-018
Abhängig von der Verbreitung reagieren viele Pflanzen mit Blütenbildung aufdie Tageslänge
Andere Induktionsfaktoren: Temperatur, Ernährung, Stress ( Notblüte )
Phytochrom ist an der Integration der Tageslänge beteiligt

Photoperiodische Steuerung des Blühens
Chrispeels and Bent
Es gibt typische Langtagpflanzen und Kurztagpflanzen
Arabidopsis thaliana ist eine Kurztagpflanze, die mindestens 9h Tageslängezur Blühinduktion benötigt
Molekularer Mechanismus der Blühinduktiondurch Tageslänge in Arabidopsis thaliana
CO = Constans;Transkriptionsfaktor erlaubtExpression von FT Gen imBlatt oberhalb kritischerTageslänge bzw. Proteinmenge
FT = Flowering locus T; FTProtein wandert aus Blatt überPhloem in das Apikalmeristem
FD Transkriptionsfaktor (bZIPFaktor: ausschließlich im Apexexprimiert). FD interagiert mit FT
Der FD/FT Komplex aktiviertSchlüsselgene (AP1 =Apetala1) um Blütenentwicklungzu induzieren
LFY (Leafy): Transkriptionsfaktorbenötigt für AP1 Expression
Blazquez (2005) Science 309:1024Corbesier et al. (2007) Science 316: 1030 + Tamaki et al. (2007) Science 316: 1033

Mutanten sind wichtige Werkzeuge zurFunktionsaufklärung von Entwicklungsprozessen
Buchanan 19.6, Nover/Weiler 17-006
phyB Mutante im LangtagNull-Mutationen in den GenenConstans (Co) undPhytochrom B (phyB) führenzu verspäteter Blütenbildung
Überproduktion von COresultiert in verfrühterBlütenbildung
35S::COim Kurztag
Pflanzenphysiologie 6: Blüten und Befruchtung
Auslösen der Blütenbildung
Identität des Apikalmeristems
Befruchtung und Embryogenese
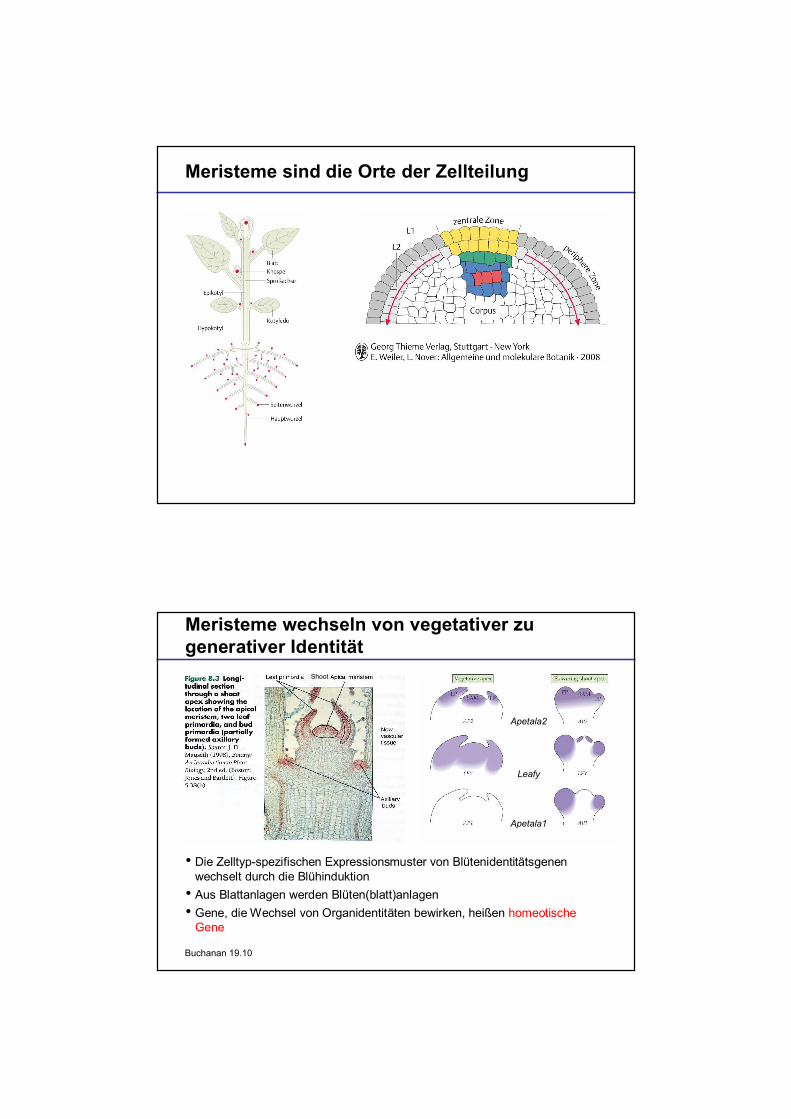
Meristeme sind die Orte der Zellteilung
Meristeme wechseln von vegetativer zugenerativer Identität
Buchanan 19.10
Die Zelltyp-spezifischen Expressionsmuster von Blütenidentitätsgenenwechselt durch die BlühinduktionAus Blattanlagen werden Blüten(blatt)anlagenGene, die Wechsel von Organidentitäten bewirken, heißen homeotischeGene
Shoot
Apetala2
Leafy
Apetala1
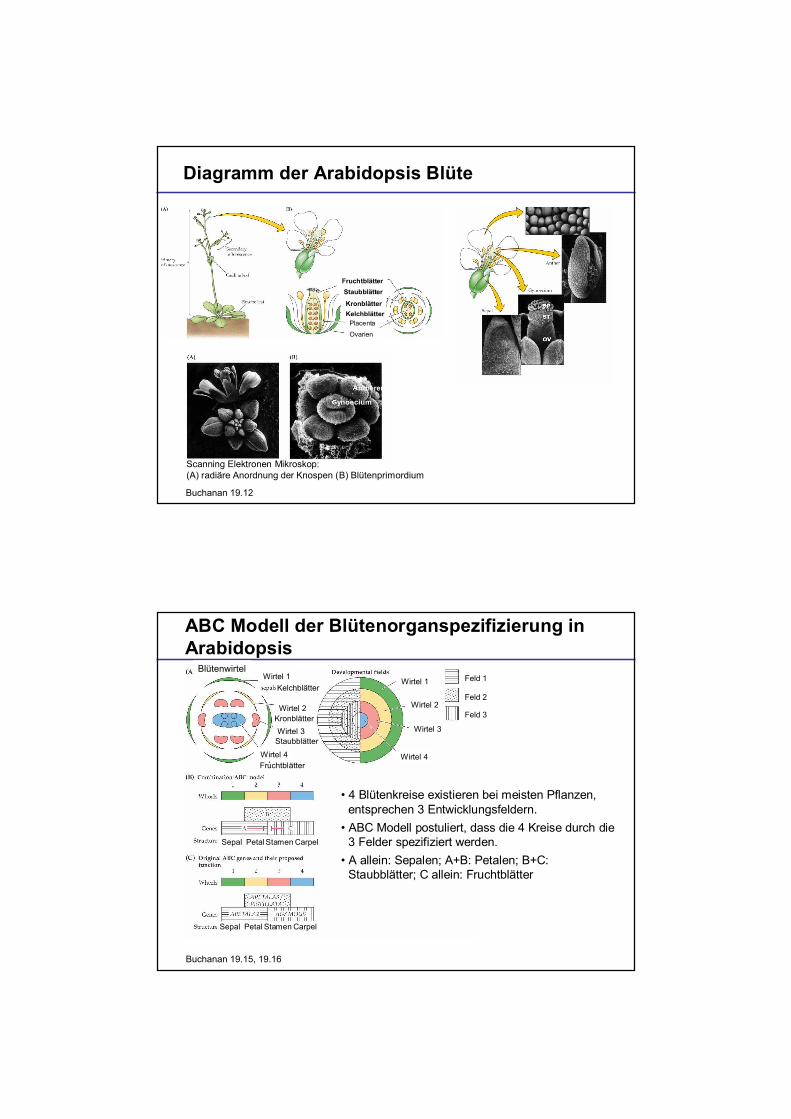
Diagramm der Arabidopsis Blüte
Scanning Elektronen Mikroskop:(A) radiäre Anordnung der Knospen (B) Blütenprimordium
Buchanan 19.12
ntheren
ynoecium
KelchblätterKronblätter
StaubblätterFruchtblätter
Ovarien
Placenta
ABC Modell der Blütenorganspezifizierung inArabidopsis
Buchanan 19.15, 19.16
Blütenwirtel
Kelchblätter
Kronblätter
Staubblätter
Fruchtblätter
4 Blütenkreise existieren bei meisten Pflanzen,entsprechen 3 Entwicklungsfeldern.ABC Modell postuliert, dass die 4 Kreise durch die3 Felder spezifiziert werden.A allein: Sepalen; A+B: Petalen; B+C:Staubblätter; C allein: Fruchtblätter
Wirtel 1
Wirtel 2
Wirtel 3
Wirtel 4
Wirtel 1
Wirtel 2
Wirtel 3
Wirtel 4
Feld 2
Feld 1
Feld 3
CarpelStamenSepal Petal
CarpelStamenSepal Petal
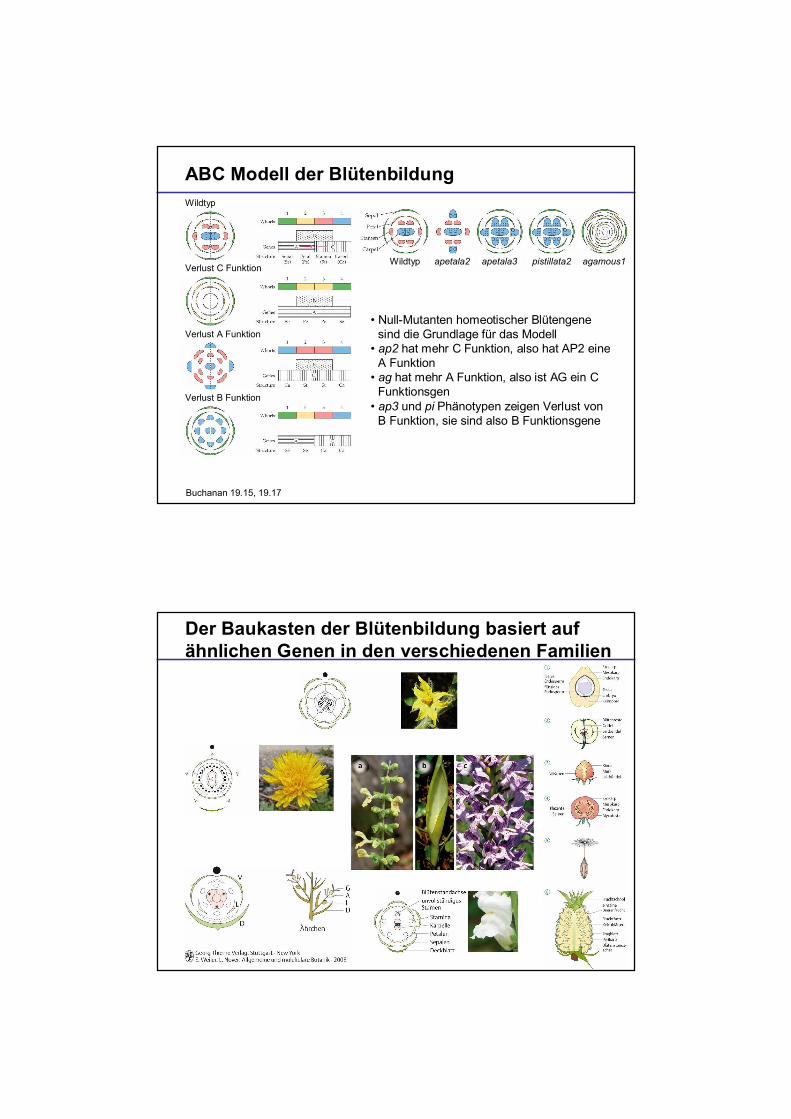
ABC Modell der Blütenbildung
Null-Mutanten homeotischer Blütengenesind die Grundlage für das Modellap2 hat mehr C Funktion, also hat AP2 eineA Funktionag hat mehr A Funktion, also ist AG ein CFunktionsgenap3 und pi Phänotypen zeigen Verlust vonB Funktion, sie sind also B Funktionsgene
Buchanan 19.15, 19.17
Wildtyp apetala2 apetala3 pistillata2 agamous1
Wildtyp
Verlust C Funktion
Verlust A Funktion
Verlust B Funktion
Der Baukasten der Blütenbildung basiert aufähnlichen Genen in den verschiedenen Familien

Pflanzenphysiologie 6: Blüten und Befruchtung
Auslösen der Blütenbildung
Identität des Apikalmeristems
Befruchtung und Embryogenese
Entwicklungsgang der dikotylen Angiospermen
Spermatophyten führen einen heterophasischen, heteromorphenGenerationswechsel durchMännlicher und weiblicher Gametophyt sind auf wenige Zellen beschränktund existieren nur als Teil des Spermatophyten
Weibl. Gametophyt
Männl. Gametophyt
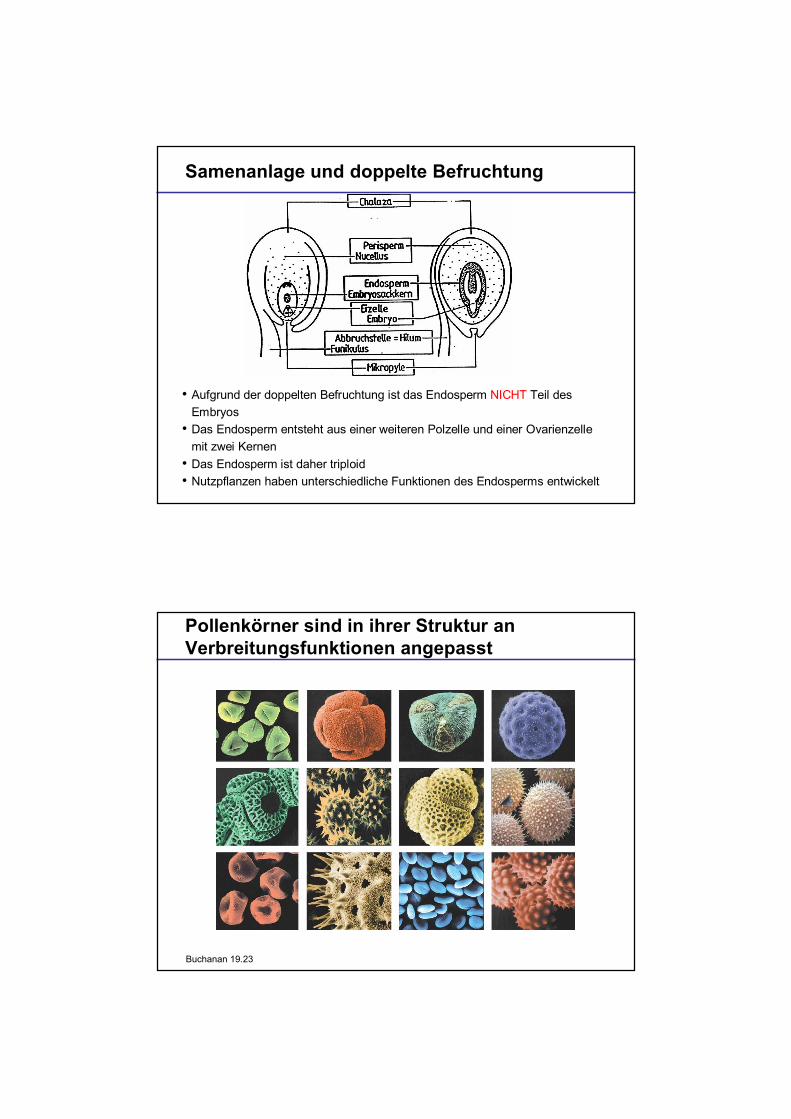
Samenanlage und doppelte Befruchtung
Aufgrund der doppelten Befruchtung ist das Endosperm NICHT Teil desEmbryosDas Endosperm entsteht aus einer weiteren Polzelle und einer Ovarienzellemit zwei KernenDas Endosperm ist daher triploidNutzpflanzen haben unterschiedliche Funktionen des Endosperms entwickelt
Pollenkörner sind in ihrer Struktur anVerbreitungsfunktionen angepasst
Buchanan 19.23
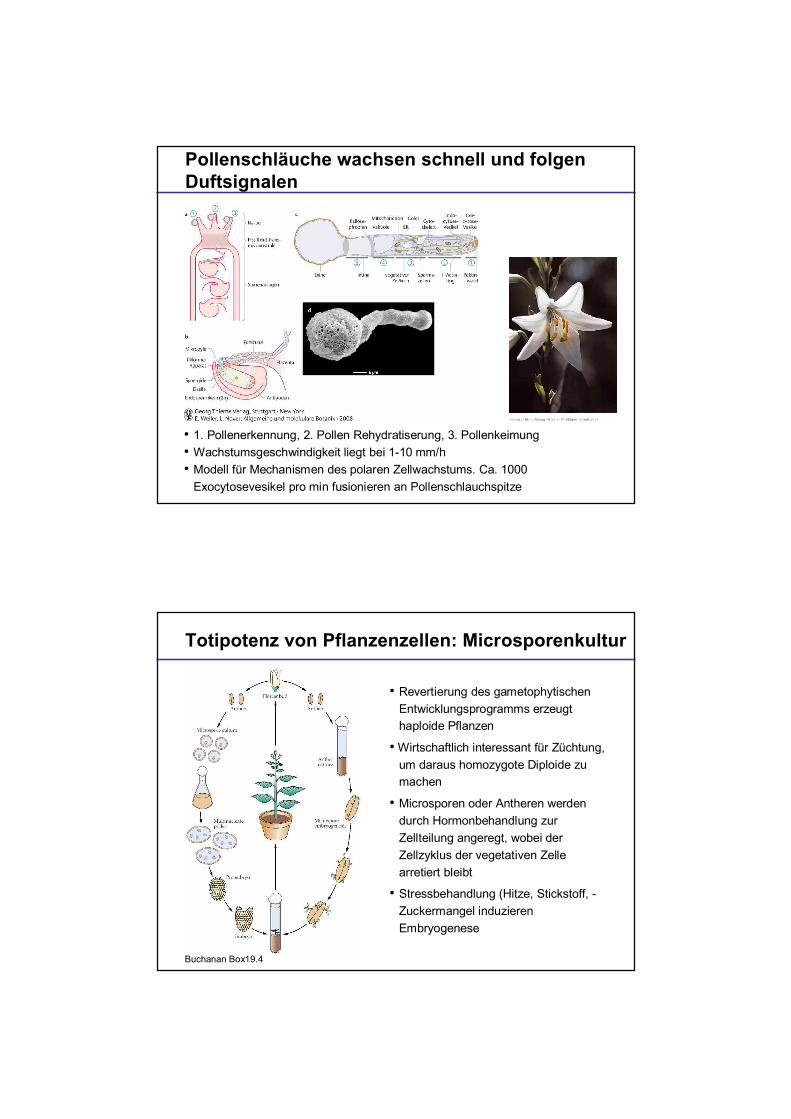
Pollenschläuche wachsen schnell und folgenDuftsignalen
1. Pollenerkennung, 2. Pollen Rehydratiserung, 3. PollenkeimungWachstumsgeschwindigkeit liegt bei 1-10 mm/hModell für Mechanismen des polaren Zellwachstums. Ca. 1000Exocytosevesikel pro min fusionieren an Pollenschlauchspitze
Totipotenz von Pflanzenzellen: Microsporenkultur
Revertierung des gametophytischenEntwicklungsprogramms erzeugthaploide Pflanzen
Wirtschaftlich interessant für Züchtung,um daraus homozygote Diploide zumachen
Microsporen oder Antheren werdendurch Hormonbehandlung zurZellteilung angeregt, wobei derZellzyklus der vegetativen Zellearretiert bleibt
Stressbehandlung (Hitze, Stickstoff, -Zuckermangel induzierenEmbryogenese
Buchanan Box19.4

Zusammenfassung
Externe Faktoren induzieren die Blütenbildung
Das Apikalmeristem wechselt von vegetativer zum generativer Identität
Das ABC Modell erklärt die homeotischen Prozesse der Blütenorgane
Die doppelte Befruchtung basiert auf einem heterophasischemGenerationswechsel
Blütenchromoplasten
Recommended

















