Structural Changes in the Primary Visual Cortex of Alzheimer’s Disease Model Mice As A Correlate to
Diminished Plasticity
Kiana Brown
May 2015

1
Structural Changes in the Primary Visual Cortex of Alzheimer’s Disease Model Mice As A Correlate to
Diminished Plasticity
An Honors Thesis Submitted to the Department of Biology
in partial fulfillment of the Honors Program STANFORD UNIVERSITY
By Kiana Brown
May 2015


3
Acknowledgments
I would like to thank my graduate student mentor George Vidal for teaching me the techniques used in my research, as well as for his support and assistance throughout my past year in the lab. I would also like to thank him for reading through many drafts of my thesis. I could not have completed this project without him. I would also like to thank Richie Sapp, who has been an outstanding peer and support through my time in the lab, and especially helpful in writing this thesis. I would like to thank Maja Djurisic for teaching me confocal microscopy, which has been central to imaging and obtaining results, as well as for general help throughout my time in the lab. I would like to thank Taeho Kim for his intellectual contributions that have prefaced my current work. I would also like to thank Dr. Carla Shatz, Principal Investigator, for the opportunity to conduct research in her lab and for her feedback and contributions to my research techniques and experimental design. She has been an amazing mentor and taught me a great deal about research in general. I would also like to thank the other members of the Shatz lab, Barbara Brott, Hanmi Lee, Kylie Chew, and Michael Birnbaum, for their support and help throughout my time in the lab. I would like to thank Thomas Joseph for his support in editing my thesis, as well as formatting figures. Nora Sotelo-Kury has been an invaluable resource in logistical and organizational support and I cannot thank her enough for that. Finally, I am grateful to Peggy Kemper and Christina Chechelski for their hard work on animal breeding and animal transfers of all the mice used in this study.

4
Table of Contents
Abstract…………………………………………………………………………………………...6 Introduction………………………………………………………………………………………7 Neural Plasticity…………………………………………………………………………...8 Molecular Controls of Plasticity…………………………………………………………..9
Structural Changes and Correlates of Plasticity………………………………………….11 Alzheimer’s Disease……………………………………………………………………..13
Materials and Methods………………………...……………………………………………….15 Mice……………………………………………………………………………………...15 Perfusions………………………………………………………………………………...15 Immunostaining………………………………………………………………………….15
Dendritic Spine Analysis………………………………………………………………...16 Results…………………………………………………………………………………………...16 Dendritic Spine Density of Juvenile Transgenic Mice Is Not Decreased………………..16 Dendritic Spine Density of Adult Transgenic Mice Is Not Decreased…………………..17 Discussion/Conclusions…………………………………………………………………………17 References……………………………………………………………………………………….21 Figures…………………………………………………………………………………………...23

5
List of Figures
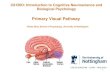
Figure A Schematic of Mouse Visual System
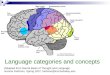
Figure B Schematic of APP/PS1;YFP-H+ Mouse Breeding
Figure C Western Blot Analysis of APP and Aβ Expression
Figure 1 Dendritic spine density remains unaltered in P30 APP/PS1 mice
Figure 2 P30 APP/PS1 mice show similar dendritic spine morphology as compared to wildtype
Figure 3 Dendritic spine density elevated in P74 APP/PS1 mice

6
Abstract
Amyloid-β (Aβ) accumulation has been implicated in Alzheimer’s disease pathogenesis.
Mice with transgenic expression of mutant amyloid precursor protein (APP) and presenilin-1
(PS1) accumulate Aβ and have therefore been used as models of the disease. Previous research
has shown that critical period ocular dominance (OD) plasticity is defective in these mice, which
indicate an early effect of Alzheimer’s on neural plasticity, even before the typical Aβ plaques
accumulate. In an attempt to determine the mechanism responsible for this defect, I investigated
layer 5 apical dendritic spine density and morphology in the primary visual cortex of juvenile
(P30) and adult (P74) APP/PS1 mice. We found that wildtype and APP/PS1 transgenic juvenile
and adult mice both displayed comparable spine densities and initially concluded that the critical
period ocular dominance plasticity defect observed in APP/PS1 transgenic mice is not associated
with a decrease in spine density in layer 5 apical dendrites. However, recent discoveries within
the lab have led us to believe that the APP/PS1;YFP-H+ mouse line used in these experiments
may have epigenetic or other modifications that would inhibit transcription of YFP and/or
APP/PS1. Further experiments should be conducted to verify the presence of Aβ accumulation in
this mouse line before continuing research.

7
Introduction
Neural plasticity refers to the brain’s ability to rewire pathways and synapses in response
to changes in behavior and environment, and to injury. The Shatz lab recently discovered that
specific molecules such as Major Histocompatibility Complex Class I (MHCI) (Corriveau et al,
1998) and its receptor, Paired Immunoglobulin-like receptor B (PirB) (Syken et al, 2006) appear
to act as “brakes” on forms of synaptic plasticity in the visual system, and may do so by
regulating synaptic density on pyramidal neurons of the cerebral cortex (Djurisic et al, 2013).
These molecules could therefore play a crucial role in understanding and treating Alzheimer’s
and other diseases that are characterized by loss of neural plasticity. Current mouse models for
Alzheimer’s disease express a transgenic, mutant form of amyloid precursor protein (APPswe)
and presenillin (PS1dE9), a subcomponent of the gamma secretase that cuts APP into its mature
form, amyloid-β (Aβ). The fibrillar form of this Aβ protein causes the amyloid plaques that are a
signature of Alzheimer’s pathology (Cerasoli et al, 2015). PirB is a high affinity receptor for Aβ
(Kim et al, 2013). Studies have already shown that APP/PS1 mice show an early form of
synaptic plasticity defect (William et al, 2012) that could possibly be restored. However, while
synaptic loss is an important characteristic of Alzheimer’s pathology, it is unknown whether
APP/PS1 mice exhibit an early loss of synapses, and whether blocking PirB function would
restore synapse number to normal levels. Therefore, monitoring structural and functional
synaptic changes in APP/PS1 mice is a crucial first step. If there is measurable synapse loss, next
steps are to determine whether PirB level modulation could act to restrict synaptic density early
on. If there is a structural correlate for the early plasticity deficits found in APP/PS1, and if this
correlate can be restored when blocking or removing PirB, then blocking the PirB-Aβ interaction
could become an extremely important early therapeutic.

8
Neural Plasticity
A crucial aspect of the brain is its ability to adapt according to sensory input and learning
(Feldman, 2009). Neural plasticity, the term used to describe these changes, is one of the major
fields of study within the field of neuroscience and is crucial for normal development of learning
and memory, the development of sensory maps, and recovery following injury. By uncovering
the mechanisms of neural plasticity, we can discover therapeutic techniques for disease such as
Alzheimer’s, which is neurodegenerative in nature and restricts various forms of neural plasticity
(William et al, 2012).
One method to study neural plasticity is through experience-driven or training-induced
changes in primary sensory cortex topographical maps. In the primary visual cortex, current
research focuses on ocular dominance plasticity and synaptic formation and loss (Bear &
Rittenhouse, 1999; Hensch, 2005; Levelt & Hübener, 2012). By studying the mouse visual
system through these methods, we have access to a highly adaptable model of neural plasticity,
as wiring is refined during critical periods of plasticity by interactions involving neural activity
and structural remodeling.
For example, wiring from the retina to the thalamic lateral geniculate nucleus (LGN)
occurs as spontaneous patterns of neural activity are sent to the brain from the eye, and is refined
by this activity. By blocking activity from retinal ganglion cells with injections of tetrodotoxin (a
voltage-gated sodium channel blocker) in utero or after birth through the critical period, normal
wiring is disrupted. Specifically, blocking activity from one retina induces projections from the
unmanipulated eye to expand into LGN territory normally belonging to the blocked eye (Penn, et
al, 1998). In the case of alterations to the synaptic plasticity of mice, these patterns get altered
and are often abolished or attenuated.

9
Another example of plasticity can be found
in the visual cortex. The visual cortex of normally
reared adult mice receives functional input from
both eyes. The monocular zone, where neurons are
driven solely by input from the contralateral eye,
encompasses a majority of the visual cortex. A
much smaller binocular zone also exists, receiving
input from both the ipsilateral and contralateral
eye. During development, larger areas of the visual cortex initially receive input bilaterally, but
ipsilateral eye input becomes restricted in an activity-dependent manner by the postnatal week
four, known as the critical period (Gordon & Styker, 1996). In the adult visual cortex, signals
from the eye are relayed mainly to the contralateral visual cortex, as well as a smaller binocular
zone of the ipsilateral cortex. This is called ocular dominance and can be altered in mice if the
relative ratio of activity being driven by both eyes is changed. Closing or removing one eye
during the critical period has been shown to shift ocular dominance towards the open functioning
eye (Antonini & Styker, 1993).
Molecular Controls of Plasticity Neural plasticity is under the control of many factors, including a number of molecules,
such as MHC I and its receptor, PirB. Originally thought to be located only in the immune
system, the Shatz lab has since discovered these molecules also regulate plasticity in the central
nervous system (CNS). In the immune system, MHCI molecules present antigens to cytotoxic T-
cells. There are over 50 different MHCI genes that encode MHCI proteins, which are cell-surface
ligands and must act through a receptor-mediated mechanism (Ploegh et al, 1981). In the
Figure A: Schematic of Mouse Visual system. Binocular Zone (BZ) receives input from both eyes (Datwani et al, 2009)

10
nervous system, an unbiased screen for activity-regulated genes in the LGN was performed
during layer-specific eye formation. This led to the discovery that MHCI mRNA is
downregulated in correlation with activity blockade (Corriveau et al, 1998). Subsequent studies
have shown that synaptic plasticity is dependent on cell surface expression of MHC I (Huh, et al
2000) and in mice lacking two specific MHCIs, H2-Kb and H2-Kb, ocular dominance plasticity
during the critical period and adulthood is enhanced. Together, these results suggest that
neuronal MHCI negatively regulates ocular dominance plasticity and is involved in the
regression and/or elimination of inappropriate synaptic connections. Known receptors for
immune MHCI ligands were not found to be expressed in the CNS, (Syken & Shatz, 2003) so a
screen to find a novel receptor expressed in the cortex was performed. This study led to the
identification of PirB as a MHCI receptor.
Separate studies have shown that MHCI is a physiological ligand for PirB, meaning
MHCI can bind and signal through PirB (Takai, 2005) The PirB molecule consists of six
extracellular immunoglobulin (IG)–like domains, a transmembrane domain, and four
intracellular immunoreceptor tyrosine-based inhibitory motif (ITIM) domains. Found in a
number of immune cells, including mast cells, B-cells, and macrophages, PirB aids in anti-
inflammation and recruiting phosphatases to decrease signaling in other pathways that
antagonize integrin and MAP Kinase cascade pathways (Maeda et al, 1999; Takaix, 2005). In
neurons, when ligands bind PirB, the ITIM domains are phosphorylated and PirB recruits
members of the SHP family of phosphatases (SHP-1,2), which sends an inhibitory signal to other
cellular pathways (Syken et al 2006). The subsequent pathways are currently undetermined.

11
Structural Changes and Correlates of Plasticity In addition to molecular controls of plasticity, structural changes are found to occur, both
through Arc mRNA induction and spine density measurements. Arc mRNA induction involves
exposing an eye to visual stimulation, which rapidly induces activity-regulated intermediate-
early gene Arc mRNA in visual cortex neurons driven by the stimulated eye. During monocular
enucleation (removal of one eye), inputs to the binocular zone driven by the remaining functional
eye expand. This is a measure of the strengthening of inputs from the open eye, which is a major
component of ocular dominance plasticity. By studying Arc mRNA induction in mice, it was
uncovered that mice lacking PirB display increased ocular dominance plasticity in the visual
cortex in both the critical period and adulthood as compared with wildtype (Syken et al, 2006).
A recently investigated mechanism for how ocular dominance plasticity occurs is by
identifying structural synapse changes that occur along with this shift in ocular dominance.
Inspired by Eric Knudsen’s experiments in barn owls, Hofer et al, 2009 studied dendritic spine
density. It was already known that temporary closure of one eye leads to ocular dominance
plasticity shifts that are reversed once normal vision is restored. When these same mice
experience a second monocular deprivation weeks later, the ocular dominance plasticity shifts
that occur are much more accelerated than the first, indicating that a structural remnant exists in
cortical circuits. By repeatedly imaging dendritic spine through the cycle of two monocular
deprivations, it was discovered that dendritic spine densities increase following the first
monocular deprivation, persist even after normal vision is restored, and allow for a more
accelerated ocular dominance shift when the mice undergo a second monocular deprivation.
Conclusions from the research noted that in the normal adult mouse visual cortex, monocular
deprivation will increase spine density and that this increase is stable and thought to be a
structural substrate for enhanced ocular dominance plasticity (Hofer et al, 2009).

12
In PirB deficient mice, Djurisic et al (2013) studied changes in dendritic spine density as
well as miniature excitatory postsynaptic current (mEPSC) frequency to determine that the
spines are indeed functional synapses. Imaging of apical dendrites of layer 5 pyramidal neurons
from wildtype and PirB germline knockout mice showed that PirB -/- mice have increased spine
density. This increase is insensitive to the effects of visual deprivation suggesting that PirB -/-
mice have already reached a ceiling spine density level, which cannot be further increased even
following monocular deprivation.
Following this study, the Shatz lab created sPirB, a soluble form of PirB that lacks the
intracellular domains necessary for PirB signaling. sPirB may act as decoy to endogenous
ligands (Bochner et al, 2014). In effect, sPirB blocks the function of endogenous PirB. This
approach was chosen as a proof-of-concept to achieve rapid and local disruption of PirB as a
therapeutic reagent.
Minipump infusion of sPirB during the critical period resulted in a marked increase in
ocular dominance plasticity, as assessed by Arc mRNA induction. Layer 5 pyramidal neurons
also showed increased spine density compared to wildtype, even in the absence of visual
deprivation (Bochner et al. 2014). Taken together with the increase in mEPSC frequency, these
results readily support the theory that sPirB infusion causes an increase in synaptic connectivity,
bolstering the role of PirB as a negative regulator of neural plasticity in the cortex.
To study PirB function in the hippocampus, an area of the brain intimately associated
with memory formation, LTP in in vitro hippocampal slice physiology was examined for PirB
KO mice (Djurisic et al, 2011; Sapp, 2013). Increased excitatory LTP in CA1-CA3 synapses
were identified. Further behavioral studies indicated that PirB KO mice display increased
acquisition and increased extinction in a hippocampal contextual and fear-conditioning paradigm.

13
Additionally, PirB KO mice showed improved learning in the Morris Water maze spatial
learning task (Djurisic et al, 2011). These results indicate a behavioral increase in learning speed
and strength.
Alzheimer’s Disease Although decreases in plasticity are important for stabilizing matured neural circuits by
studying ways to increase plasticity, we endeavor to find possible therapeutics that can be used in
treatments for neurodevelopmental and neurodegenerative diseases. PirB has been shown to be
an important regulator of neural connections and decreases in the protein, either through genetic
knockouts or decoy sPirB infusion, predictably lead to increased synaptic connectivity that
appears completely functional. Additionally, these physiological changes were correlated with
behavioral changes of increased speed and strength of learning.
Alzheimer’s Disease is a complex neurodegenerative disease that affects somewhere
between 3 and 4 million people and costs upwards of $1 billion annually in the United States
alone. Characterized mainly by dementia and memory loss symptoms, it is complex to treat
because of the array of molecular defects that can spur its pathology. Hallmarks of the disorder
include substantial cortical atrophy and upon silver stain examination, loss of neurons and
buildup of amyloid plaques (Selkoe & Lansbury, 1999). Amyloid β (Aβ) is a 36-43 amino acid
peptide formed from amyloid precursor protein (APP) being cleaved by specific enzymes.
Aggregation of misfolded Aβ leads to plaques, which are toxic to nerve cells. Soluble Aβ
oligomers are known to impair synaptic plasticity and cause the neuroplasticity loss prevalent in
Alzheimer’s.
An in vivo murine model of Alzheimer’s, which expressed mutant alleles of both amyloid
precursor protein (APPswe) and presenillin 1 (PSEN1ΔE9) showed very early loss of activity-
dependent plasticity. Aβ plaque deposition occurs at several months of age, long after the critical

14
period of visual cortical plasticity. (Garcia-Alloza et al, 2006 & Jankowksy et al, 2003).
However, normal ocular dominance plasticity was disrupted early on. (William et al, 2012). This
follows the progression thought to occur in human Alzheimer’s patents, where those with Aβ
plaques have already reached a late stage of the disease. In another study, PirB was shown to
regulate plasticity in Alzheimer’s mouse models (Kim et al, 2013). Both PirB and LilrB2, the
human homolog to PirB, are receptors for Aβ oligomers. Using biotinylated synthetic human
Aβ1-42 (Aβ42) peptides as monomers and oligomers, Aβ42 binding to human embryonic kidney
cells expressing PirB or control were tested. Oligo-Aβ42 was bound PirB-expressing cells six
times more than monomeric Aβ42 and found to be constantly associated with PirB protein in
both coimmunoistaining and coprecipitation experiments. This binding is specific to PirB, as
Aβ42 did not bind cells expressing mouse PirA1, mouse PirA4 or an isoform of rat PirB to any
appreciable degree.
Once it was known that PirB binds Aβ42, a more direct contribution of PirB to
Alzheimer’s pathology was sought. APP/PS1 transgenic (Tg) mice were crossed with PirB-/-
mice to generate littermates that were either PirB+/-Tg or PirB -/-Tg. At 9mo of age, recognition
memory was evaluated through novel object and novel place behavioral tests. (Kim et al, 2013)
While both tests revealed the expected impaired behaviors in PirB+/-Tg mice, these learning and
memory deficits were not apparent in mice lacking PirB (PirB -/-Tg). In summary, these results
suggest that PirB may act by negatively regulating plasticity and limiting PirB could be a method
of alleviating the effects of Alzheimer’s.
In order to elucidate how PirB plays a role in Alzheimer’s and how effective sPirB would
be as a potential therapeutic, the details of synaptic alteration need to be examined in more depth.
Since it is known that PirB-/- mice show increased layer 5 pyramidal neuron spine densities as

15
well as an increased area of Arc mRNA induction, the effect of Alzheimer’s on these structural
measures of plasticity should be examined. This project aims to identify and quantify changes in
APP /PS1 transgenic mouse spine density to characterize the development of Alzheimer’s.
Drawing from previous research that shows ocular dominance plasticity is defective in juvenile
transgenic mice, we hypothesize that APP/PS1 juvenile (P30) transgenic mice will also have
lower spine densities than wildtype littermates. Furthermore, this effect should become more
pronounced in adult (P74) mice as the disease has more time to develop.
Materials and Methods Mice APP/PS1 heterozygous transgenic mice (strain B6C3 Tg(APPswe,PSEN1dE9)85Dbo/J; Jan-kowsky et al., 2001) from The Jackson Laboratory were crossed into a PirB +/+ line, generating male mice in line 1 of the schematic below. All subsequent crosses were conducted following Figure B. Crosses generated APP/PS1 Tg+ and APP/PS1 Tg- mice, which were all YFP+ and were used to determine dendritic spine density at juvenile (P30) and adult (P74) ages. Juvenile females and males and adult males were used for experimentation. Perfusions Brains from APP/PS1 Tg+; (P30 and P74) were fixed via transcardial perfusion with 4% (wt/vol) paraformaldehyde in PBS. Coronal sections of 150 µm thickness were used for fluorescence microscopy of YFP+ neurons in P30 mice. Coronal sections of 50 µm were used for fluorescence miscropy of YFP+ neurons in P74 mice, to allow for immunostaining. All sections were cut on a vibratome (Leica VT1200S) and mounted on slides with Prolong Gold Antifade Reagent. Immunostaining Immunostaining was used to amplify the signal in P74 brains, but not P30 brains. All P74 brains analyzed were cut into 50 µm thick coronal sections, permeabilized in 0.2% Triton-X100, incubated in GFP primary antibody (NB600-308 from Novus Biologicals) at 1:500 concentration in 5% normal goat serum in 1x PBS, and then Alexa-488 conjugated goat anti-rabbit secondary antibody (A11008 from Life Technologies) at 1:500 concentration in 5% normal goat serum in 1x TBS.
B
Figure B: Schematic of APP/PS1;YFP-H+ Mouse Breeding. (Schematic by Taeho Kim, Shatz Lab, 2013)

16
Dendritic Spine Analysis Distal apical tufts of layer 5 neurons labeled with YFP were imaged with a confocal microscope (Leica Microsystems SP8) using a 63× oil immersion objective. Neurons with cell bodies in layer 5 were anatomically selected from the binocular zone of the visual cortex. High-resolution images of apical tufts and the primary dendritic shaft immediately below the main branching of the dendrite (roughly in layer 2) were taken using confocal microscopy. Initial neuron localization was done using a 10x objective and imaging was done using a 60x oil-immersion objective and 4x optical zoom. Z-stacks were taken to capture as much of the neuron as possible in three dimensions within the section. Spine density analysis was done blind to the genotype using FIJI, a build of ImageJ software. Densities were calculated manually using data from Cell Counter and Simple Neurite Tracer plugins to count spines and measure dendrite length. Results Layer 5 apical dendrites of APP/PS1 transgenic mice were imaged at P30 and P74 in an
effort to identify structural changes occurring in the brain of Alzheimer’s models before the
onset of typical behavioral phenotypes. Previous research had identified an ocular dominance
plasticity defect as early as P28 (William et al, 2012) and measuring changes in dendritic spines,
namely changes in spine density, provide a structural substrate for rapid synaptic changes (Hofer
et al, 2009). I hypothesized that spine densities in both P30 and P74 APP/PS1 mice would be
decreased compared to wildtype, correlating with the ocular dominance plasticity defect.
Dendritic Spine Density of Juvenile Transgenic Mice Is Not Decreased
Unexpectedly, results in juvenile (P30) mice have significant difference in spine density
between APP/PS1 transgenic and wildtype mice (Fig. 1B). This indicates that the critical period
ocular dominance plasticity defect observed in APP/PS1 transgenic mice is not associated with a
decrease in spine density in layer 5 apical dendrites in juvenile mice. Additionally, there is not
measurable change in spine morphology that could account for the ocular dominance plasticity
defect (Fig. 2A and B). It is possible that while morphology has not yet changed, physiology has
already been altered, causing the ocular dominance plasticity defect.
Previous research has shown that APP/PS1 mice have measurable synaptic plasticity
defects when measured using Arc mRNA induction (William et al, 2012) Determination of spine

17
density changes, which are a known structural correlate for rapid synaptic change, (Hofer et al,
2009) would illuminate early effects of Alzheimer’s on the brain and indicate sPirB as a potential
therapeutic. We hypothesized APP/PS1 mice would have decreased spine densities as compared
with wildtype as APP/PS1 mice display ocular dominance defects via Arc induction. While
wildtype mice demonstrated a normal expansion in the width of visual cortex responding to
ipsilateral eye input, APP/PS1 did not show this expansion (William et al, 2012).
Dendritic Spine Density of Adult Transgenic Mice Is Not Decreased
In order to gain a better understanding of Alzheimer’s disease progression, spine
densities were also measured in adulthood where structural changes were more likely to have
developed. P74 APP/PS1 transgenic mouse spine densities were compared with their wildtype
littermates as a control. Here as well, there was no significant difference in spine density
between the APP/PS1 mice and wildtype, although data did show an interesting trend not present
in juvenile mice (Fig. 3C). In general, this trend would indicate that individual spine densities for
P74 APP/PS1 mice were higher than those for P74 wildtype littermates. In summary, these
results are unexpected given our hypothesis that spine density would decrease and that this
would correlate to ocular dominance plasticity defects.
Discussion and Conclusions The spine densities of layer 5 pyramidal neurons were analyzed to identify structural
changes occurring in APP/PS1 mice in correlation with known ocular dominance plasticity
defects. As this defect is seen as early as the critical period, identification of structural changes
could provide a clue into Alzheimer’s disease progression and a target for early therapeutics. It
was hypothesized that spine densities measured would be lower in APP/PS1 mice compared to
wildtype, as this seems the most likely cause of plasticity defects. Results showed that neither
juvenile nor adult APP/PS1 mice displayed any significant change in spine density compared to

18
wildtype. These results seem to indicate that the critical period ocular dominance plasticity
defect previously seen were not associated with any changes in spine density of layer 5 apical
dendrites. The ocular dominance plasticity defect may then be due to physiological changes that
have occurred and would not be detectable via spine density measurements. This conclusion
would be inconsistent with other experiments that have indicated dendritic spines to be a
structural substrate for rapid synaptic change (Hofer et al, 2009).
P74 mice did show an interesting trend, where APP/PS1 generally had higher spine
densities than wildtype mice. As only 7 cells were analyzed for P74 APP/PS1 mice and 6 cells
for P74 wildtype, it is possible that with a larger population sample, data would be more
conclusive. If larger samples were measured and the trend found to be significant, it could have
important implications about the mechanism of Alzheimer’s disease development.
Primarily, it would suggest that Aβ could be causing problems with the brain’s ability to
form connections and wire correctly. As a method of compensating for this, increasing numbers
of spines are formed in an attempt to form new connections. In vivo time course imaging of
spines as well as a developmental study of mEPSC frequency and amplitude would help
illuminate whether new spines are being formed at any appreciable rate, if spine turnover has
increased, and if the spines are correlated with functional synapases. In the case of this
compensation model, it would be likely that while new spines are being formed, they are not all
able to wire correctly and are therefore not functional.
This trend towards increased spine density is also interesting in light of a known
association between Alzheimer’s and an elevated risk for seizures present in both mice and
humans (Amantneik et al, 2006). Many mouse models for Alzheimer’s exhibit abnormal
electroencephalogram activity, suggesting fundamental cognitive dysfunction (Born et al, 2014;

19
Palop & Mucke, 2010). If all spines counted are functional excitatory synapses, the high level of
neural excitation may be causing increased rates of epilepsy. Furthermore, when APP
overexpression was withheld to adulthood, epileptic activity was substantially delayed (Born et
al, 2014). If pruning defects during development are the cause of high spine density in adult
APP/PS1 mice, it would also explain this result, as APP overexpression after development would
not have the same effect on spine density.
In order to validate or disprove these hypotheses, further research would need to be
conducted. This would include in vivo imaging of spines over an extended time period in order to
determine how the “extra” spines occur and whether they are an artifact of pruning deficits or are
actually the appearance of new spines for another reason. Another next step would be to measure
mEPSCs from layer 5 pyramidal neurons in adult APP/PS1 mice and wildtype littermates as a
control (Djurisic et al, 2013). This would indicate whether the increase in spine density on the
apical dendrites is accompanied by an increase in functional excitatory synaptic inputs. If the
spines are functional excitatory synapses, further credence would be given to the theory that the
increased spine density in adult APP/PS1 mice could be responsible, at least in part, for the
increased risk for epilepsy associated with Alzheimer’s.
However, recent developments within the lab have indicated that the APP/PS1;YFP-H+
mouse line generated to measure spines in APP/PS1 mice may not have ocular dominance
plasticity defects at all (Richard W. Sapp, Shatz Lab, personal communication, April 28, 2015).
Recent measurements of ocular dominance plasticity in P30 APP/PS1;YFP-H+ mice appear to
match those of wildtype littermates. Since previous research has indicated that APP/PS1 mice
without the YFP transgene show ocular dominance plasticity defects, it is possible that the new
line generated for these sets of experiments has been epigenetically modified, and do not

20
transcribe the mutant APP or PS1 protein at high levels. Furthermore, YFP expression was
noticeably low in this line, leading us to immunostain sections for analysis, and in retrospect
these low YFP levels may have been an indication to study the expression of all transgenes in the
line, including APP and PS1.
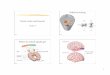
In order to determine the presence of APP
protein, a Western blot should be conducted similar to
that shown in Fig. C, where both APP and Aβ
expression were measured. Older APP/PS1;YFP-H+
mouse line brains can also be examined for the
presence of Aβ plaques, which would be a positive
indicator for APP/PS1 expression and reaffirm the line
as a valid model for Alzheimer’s disease.
In conclusion, current results indicate that ocular dominance plasticity defects previously
seen in Alzheimer’s model mice are not associated with changes in spine density as compared to
wildtype. Next steps are to first evaluate that our APP/PS1;YFP-H+ mouse line is a valid model
for Alzheimer’s disease or if APP and PS1 expression have been altered, changing Aβ
accumulation. In the case that these results are valid, research should extend to include more data
points, as well as time course measurements of spines and mEPSCs. Using this new information,
we could better characterize the early development of Alzheimer’s in mouse models, which may
provide new avenues for earlier and more targeted treatments to counteract the disease.
Fig. C Western Blot Analysis of APP and Aβ Expression (William et al, 2012)
C

21
References Amantneik JC, Hauser WA, DelCastillo-Castaneda C, Jacobs DM, Marder K, Bell K, Albert M,
Brandt J, Stern Y. (2006) Incidence and predictors of seizures in patients with Alzheimer’s disease. Epilepsia. 47(5), 867-872.
Antonini A, Styker MP. (1993) Rapid remodeling of axonal arbors in the visual cortex. Science. 260, 1819-1821.
Bear, MF, Rittenhouse CD. (1999). Molecular basis for induction of ocular dominance plasticity. J. Neurobiol. 41, 83–91.
Bochner DN. Sapp RW, Adelson JD, Zhang S, Lee H, Djurisic M, Syken J, Dan Y, Shatz CJ. (2014) Blocking PirB up-regulates spines and functional synapses to unlock visual cortical plasticity and facilitate recovery from amblyopia. Neurosci. 6(258).
Born HA, Kim JY, Savjani RR, Das P, Dabaghian YA, Guo Q, Yoo JW, Schuler DR, Cirrito JR, Zheng H, Golde TE, Noebels JL, Jankowsky JL. (2014) Genetic suppression of transgenic APP rescues hypersynchronous network activity in a mouse model of Alzheimer’s disease. J Neurosci. 34(11),3826-3840.
Cerasoli E, Ryadnov MG, Austen BM (2015) The elusive nature and diagnostics of misfolded Aβ oligomers. Frontiers in Chemistry. 3(17) 1-6.
Corriveau RA, Huh GS, Shatz CJ. (1998) Regulation of class I MHC gene expression in the developing and mature CNS by neural activity. Cell. 21, 505-520.
Datwani A, McConnell MJ, Kanold PO, Micheva KD, Busse B, Shamloo M, Smith SJ, Shatz CJ. (2009) Classical MHCI molecules regulate retinogeniculate refinement and limit ocular dominance plasticity. Neuron. 64(4), 463-470.
Djurisic M, Mann MM, Hubener M, Shatz CJ. Elevated spine density in PirB-/- mice as substrate for enhanced ocular dominance plasticity. In: Striate Cortex: Plasticity Nanosymposium, Neuroscience 2011, Washington D.C., November 13, 2011. Djurisic M, Vidal GS, Mann M, Aharon A, Kim T, Santos AF, Zuo Y, Hübener M, Shatz CJ.
(2013) PirB regulates a structural substrate for cortical plasticity. PNAS. 110(51), 20771-20776.
Feldman DE. (2009) Synaptic mechanisms for plasticity in neocortex. Annu Rev Neurosci. 32, 33-55.
Garcia-Alloza M, Robbins EM, Zhang-Nunes SX, Purcell SM, Betensky RA, Raju S, Prada C, Greenberg SM, Bacskai BJ, Frosch MP. (2006) Characterization of amyloid deposition in the APPswe/PS1dE9 mouse model of Alzheimer disease. Neurobiol Dis. 24, 516-524.
Gordon, JA. Stryker MP. (1996) Experience-dependent plasticity of binocular responses in the primary visual cortex of the mouse. J. Neurosci. 16, 3274–3286.
Hensch TK. (2005) Critical period mechanisms in developing visual cortex. Curr. Top. Dev. Biol. 69, 215–237.
Hofer SB, Mrsic-Flogel TD, Bonhoeffer T, Hübener M. (2009) Experience leaves a lasting structural trace in cortical circuits. Nature. 457, 313-317.
Huh GS, Boulanger LM, Du H, Riquelme PA, Brotz TM, Shatz CJ. (2000) Functional requirement for Class I MHC in CNS development and plasticity. Science. 290, 2155-2159.

22
Jankowsky JL, Fadale DJ, Anderson J, Xu GM, Gonzales V, Jenkins NA, Copeland NG, Lee MK, Younkin LH, Wagner SL, Younkin SG, Borchelt DR. (2004) Mutant presenilins specifically elevate the levels of the 42 residue β-amyloid peptide in vivo: evidence for augmentation of a 42-specific γ secretase. Hum Mol Genet. 13(2), 159-170.
Kim T, Vidal GS, Djurisic M, William CM, Birnbaum ME, Garcia KC, Hyman BT, Shatz CJ. (2013) Human LilrB2 is a β-amyloid receptor and its murine homolog PirB regulates synaptic plasticity in an Alzheimer’s model. Science. 341, 1399-1404.
Levelt CN, Hübener M. (2012) Critical-period plasticity in the visual cortex. Annu. Rev. Neurosci. 35, 309–330.
Maeda A, Scharenberg AM. Tsukada S, Bolen JB. Kinet JP. Kurosaki T. (1999) Paired immunoglobulin-like receptor B (PIR-B) inhibits BCR-induced activation of Syk and Btk by SHP-1. Oncogene. 18, 2291-2297.
Palop JJ, Mucke L. (2010) Amyloid-β-induced neuronal dysfunction in Alzheimer’s disease: from synapses towards neural networks. Nature Neuroscience. 13(7), 812-818.
Penn AA, Riquelme PA, Feller MB, Shatz CJ. (1998) Competition in retinogeniculate patterning driven by spontaneous activity. Science. 279, 2108-2112.
Ploegh HL, Orr HT, Strominger JL. (1981) Major histocompatibility antigens: the human (HLA-A, -B, -C) and murine (H-2K, H-2D) class I molecules. Cell. 24, 287-299.
Sapp RW. (2013) Acutely Disrupting the Function of PirB Enhances Ocular Dominance Plasticity. Undergraduate Thesis. Stanford University: USA.
Selkoe DJ, Lansbury PJ Jr. Alzheimer's Disease Is the Most Common Neurodegenerative Disorder. In: Siegel GJ, Agranoff BW, Albers RW, et al., Basic Neurochemistry: Molecular, Cellular and Medical Aspects. 6th edition. Philadelphia: Lippincott-Raven; 1999.
Syken J, Shatz CJ. (2003) Expression of T cell receptor β locus in central nervous system neurons. PNAS. 100(22) 13048-13053.
Syken J, GrandPre T, Kanold PO, Shatz CJ. (2006) PirB Restricts Ocular Dominance Plasticity in Visual Cortex. Science. 313, 1795-1800.
Takai T. (2005) Paired immunoglobulin-like receptors and their MHC class I recognition. Immunology. 155, 433-440.
Wiesel TN, Hubel DH. (1963) Effects of visual deprivation on morphology and physiology of cells in the cats lateral geniculate body. J. Neurophysiol. 26, 978–993 (1963).
William CM, Andermann ML, Goldey GJ, Roumis DK, Reid RC, Shatz CJ. (2012) Synaptic plasticity defect following visual deprivation in Alzheimer’s disease model for transgenic mice. J Neurosci. 32, 8004-8011.

23
Figures Figure 1: A) WT APP/PS1 B)
Figure 2: A) B)
Dendritic spine density remains unaltered in P30 APP/PS1 mice. 1A) Examples of apical tufts of YFP-labeled layer 5 pyramidal neurons in the binocular zone of the visual cortex. Scale bar represents 10 µm 1B) APP/PS1 transgene does not have an effect on dendritic spine densities in critical period-aged mice. N= 3/7, 3/11 animals/cells.
P30 APP/PS1 mice show similar dendritic spine morphology as compared to wildtypes. A) The distribution of dendritic spine length and width measurements for both wildtype and APP/PS1 critical period-age mice are strikingly similar. B) Histograms of frequency distribution illustrating that the proportion of dendritic spines of specific widths and lengths remain steady between wildtype and APP/PS1 P30 mice.

24
Figure 3: A) WT APP/PS1 B)
Dendritic spine density not significantly different P74 APP/PS1 mice, but trends towards an increase. 1A) Examples of apical tufts of YFP-labeled layer 5 pyramidal neurons in the binocular zone of the visual cortex. Scale bar represents 10 µm 1B) APP/PS1 transgenic mice do not have significantly different spine densities compared with wildtype. However, the data shows a trend towards elevated spine density for APP/PS1 mice. (p=0.11) N= 3/7, 3/6 animals/cells.
Recommended