
INSTITUTO POLITÉCNICO NACIONAL
CENTRO DE DESARROLLO DE PRODUCTOS BIÓTICOS
BÚSQUEDA Y ACEPTACIÓN DE LA PLANTA HOSPEDERA
EN Copitarsia decolora Guenée (LEPIDOPTERA: NOCTUIDAE)
TESIS
QUE PARA OBTENER EL GRADO DE:
DOCTORADO EN CIENCIAS EN
DESARROLLO DE PRODUCTOS BIÓTICOS
PRESENTA
HUMBERTO REYES PRADO
Yautepec, Morelos Enero, 2015


7
Autorizaci6n de uso de ohra
Instituto Polit6cnico NacionalPresente
Bajo protesta de decir verdad el que suscribe Humberto Reyes Prado (se anexa copiasimple de identificaci6n oficial), manifiesto ser autor y titular de los derechos morales ypatrimoniales de la obra titulada 'oBrfisqueda y aceptacirin de la planta hospedera enCopitarsia decolora Guen6e (Lepidoptera:Noctuidae)" en adelante "La Tesis" y de |acual se adjunta copia, por 1o que por medio del presente y con fundamento en el articulo 27fracci6n II, inciso b) de la Ley Federal del Derecho de Autor, otorgo a el InstitutoPolitdcnico Nacional, en adelante El IPN, autorizaci6n no exclusiva para comunicar yexhibir priblicamente total o parcialmente en medios digitales formato digital e impreso "LaTesis" por un periodo de un afio contado apartir de la fecha de la presente autoizaci6n,dicho periodo se renovard autom6ticamente en caso de no dar aviso expreso a "El IPN" desu terminaci6n.
En virtud de lo anterior, "El IPN" deberii reconocer en todo momento mi calidad de autorde"La Tesis".
Adicionalmente, y en mi calidad de autor y titular de los derechos morales y patrimonialesde "La Tesis", manifiesto que la misma es original y que la presente auiorizaci6n nocontraviene ninguna otorgada por el suscrito respecto de "La Tesis", por 1o que deslindo detoda responsabilidad a El IPN en caso de que el contenido de "La Tesis" o la autorizaci6nconcedida afecte o viole derechos autorales, industriales, secretos industriales, convenios ocontratos de confidencialidad o en general cualquier derecho de propiedad intelectual deterceros y asumo las consecuencias legales y econ6micas de cualquier demanda oreclamaci6n que puedan derivarse del caso.
Mexico, D. F., l0 de Diciembre de 2014
Atentamente
Humberto Reyes Prado

I NSTITUTo PoLrcEcl.ttco NAct aNALsEcRErA nin ne NvEsrteAct6tu yposGRADo
CARTA CES/ON DE DERECHOS
En la Ciudad de Yautepec, Morelos el dia l0 del mes de Diciembre del afio 2014, el que
suscribe Humberto Reyes Prado alumno del Programa de Doctorado en Ciencias en
Desarrollo de Productos Bi6ticos con nfmero de registro F101264, adscrito al Centro de
Desarrollo de Productos Bi6ticos, manifiesta que es autor intelectual del presente trabajo
de Tesis bajo la direcci6n del Dr. Angel Ren6 Arzuf{i Barrera y Dra. Norma Reyna
Robledo Quintos cede los derechos del trabajo intitulado "Btlsqueda y aceptaci6n de la
planta hospedera en Copitarsia decolora Guen6e (Lepidoptera:Noctuidae)", al Instituto
Polit6cnico Nacional para su difusi6n, con fines acaddmicos y de investigaci6n.
Los usuarios de la informaci6n no deben reproducir el contenido textual, grdficas o datos
del trabajo sin el permiso expreso del autor yio director del trabajo. Este puede ser obtenido
escribiendo a la siguiente direcci6n Carretera Yautepec-Jojutlao Km. 6, calle CEPROBI
No. 8, Col. San Isidroo Yautepec, Morelosn M6xico. C.P.62731, Apartado Postal 24.
Tel6fonos: (735) 394 20 200 3941896, (55) 57 29 69 00 f,xt. 82500 / 82505. Fax 82512,
82521, e-mail: [email protected] (http:\\rvww.ceprobi.ipn.mx). Si el permiso se otorga, el
usuario deber6 dar el agradecimiento correspondiente y citar la fuente del mismo.
Humberto Reyes Prado
Nombre del alumno

DECLARACION DE RESPONSABILIDAD
Con base en el articulo 57 fracci6n I del Reglamento de Estudios de Posgrado vigente y en
la Secci6n IV del C6digo de Etica del IPN, hacemos constar que el trabajo de tesis
u'Bfsqueda y aceptaci6n de la planta hospedera en Copitarcia decalora Guen6e
(tepidoptera:Noctuidae)" es responsabilidad del Dr. Angel Ren6 Arzuffi Barreran Dra.
Norma Reyna Robledo Quintos y llumberto Reyes Prado, y que ni los datos
experimentales ni el texto han sido usados para obtener otro grado acaddmico en el pafs o
en el extranjero. Cualquier colaboraci6n o cita textual fue declarada y reconocida en el
documento-
Yautepec, Morelos; a 10 de Diciembre de2Al4
ATENTAMENTE
Humberto Reyes PradoAlumno
Director de tesis Director de tesis

El presente trabajo se realizó en el laboratorio de Ecología Química de Insectos del
Departamento de Interacciones Planta-Insecto del Centro de Desarrollo de Productos
Bióticos del Instituto Politécnico Nacional bajo la dirección del Dr. Ángel René Arzuffi
Barrera y la Dra. Norma Reyna Robledo Quintos. Para la realización de los estudios se
contó con el apoyo económico de CONACyT (No. Becario 225566) y del Programa
Institucional de Formación de Investigadores de la Secretaría de investigación y Posgrado
(SIP) del IPN. La investigación fue realizada con el financiamiento otorgado a los
proyectos de la SIP (No. 20100397, 20110762, 20121192, 20130883 y 20141203).

AGRADECIMIENTOS
Al Instituto Politécnico Nacional (IPN), Consejo Nacional de Ciencia y Tecnología
(CONACYT) y al Programa Institucional de Formación de Investigadores (PIFI) del
Instituto Politécnico Nacional (IPN), por el apoyo financiero para realizar mis estudios de
Doctorado.
Al Dr. René Arzuffi Barrera y la Dra. Norma Robledo Quintos por la asesoría, la confianza,
el tiempo y la paciencia, muchas gracias.
A los profesores que revisaron mi tesis; el Dr. René Arzuffi Barrera, Dra. Norma Robledo
Quintos, Dra. Gabriela Trejo Tapia, Dr. Víctor Rogelio Castrejón Gómez, Dr. Federico
Castrejón Ayala, Dr. Alfredo Jiménez Pérez y Dra. Kalina Bermúdez Torres; gracias por
sus pertinentes sugerencias y comentarios que enriquecieron este trabajo de tesis.
Al Dr. Julio Rojas de ECOSUR, por aceptarme y permitirme trabajar en su laboratorio
durante la estancia de investigación.
A mi padres, mis hermanas y a Lety. Gracias por su apoyo incondicional. Estoy en deuda
con ustedes.

i
Contenido
Relación de cuadros
vi
Relación de figuras
vi
Resumen
ix
Abstract
x
I. Introducción general
1. Selección de la planta hospedera por noctuidos adultos
2
1.1. Búsqueda de la planta hospedera por adultos
3
1.2. Aceptación de la planta hospedera por adultos 5
2. Selección de la planta hospedera por larvas 7
3. Emisión de volátiles por las plantas 9
4. Factores que influencian la selección en adultos
12
5. Manejo de insectos plaga con semioquímicos 14
6. Copitarsia decolora como modelo de estudio
16
7. Objetivos
18
II. Atracción de hembras de C. decolora apareadas a compuestos volátiles de
la col
1. Introducción
20
2. Materiales y Métodos
21
2.1. Insectos
21
2.2. Plantas
22
2.3. Extracción de volátiles de la col sin daño y con daño
22
2.4. Respuesta de atracción en túnel de viento a extractos de col
23
2.5. Análisis estadístico
25

ii
3. Resultados
26
3.1. Respuesta de atracción en túnel de viento a volátiles de la col
26
4. Discusión 28
III. Identificación de los compuestos volátiles de la planta de col
1. Introducción
31
2. Materiales y Métodos
32
2.1. Insectos
32
2.2. Plantas
32
2.3. Extracción de compuestos volátiles de la col
32
2.4. Análisis químico e identificación de los compuestos
33
2.5. Respuesta electroantenográfica al extracto y a la mezcla de sintéticos
34
2.6. Respuesta de atracción en túnel de viento al extracto y a la mezcla de
sintéticos
34
2.7. Análisis estadístico
35
3. Resultados
35
3.1. Identificación de los compuestos volátiles
35
3.2. Respuesta electroantenográfica al extracto y a la mezcla de sintéticos
35
3.3. Respuesta de atracción en túnel de viento al extracto y a la mezcla de
sintéticos
37
4. Discusión
38

iii
IV. Factores que afectan a las hembras de C. decolora en la búsqueda de la
planta de col y a los machos en la búsqueda de pareja
1. Introducción
41
2. Materiales y Métodos
43
2.1. Insectos
43
2.1.1. Edad al apareamiento en hembras
43
2.1.2. Edad y estado de apareamiento en los machos 44
2.1.3. Acondicionamiento de los insectos para evaluar el efecto de pre-
exposición a los volátiles de col y a la feromona sexual
44
2.1.4. Efecto de pre-exposición a los volátiles de planta hospedera
y al extracto glandular de feromona sexual.
45
2.2. Plantas 45
2.3. Extracción de compuestos volátiles de la col 45
2.4. Extracción de feromona sexual 46
2.5. Respuesta de atracción en túnel de viento al extracto 46
2.6. Análisis estadístico 47
3. Resultados 47
3.1. Efecto de la edad al apareamiento en hembras 47
3.2. Edad y estado de apareamiento en los machos 48
3.3.- Efecto de la pre-exposición 51
3.3.1. Acondicionamiento de los insectos para evaluar el efecto de
pre-exposición
51
3.3.2. Pre-exposición de hembras apareadas a volátiles de col
53
3.3.3. Pre-exposición de machos vírgenes a feromona sexual 55

iv
4. Discusión
58
V. Preferencia de oviposición por C. decolora
1. Introducción 65
2. Materiales y métodos
67
2.1. Insectos
67
2.1.1. Preferencia de oviposición
67
2.2. Plantas
67
2.3. Determinación de preferencia de oviposición
67
2.4. Análisis de datos
68
3. Resultados
69
3.1. Preferencia de oviposición
69
4. Discusión
70
VI. Atracción de larvas de C. decolora a volátiles de la col
1. Introducción
74
2. Materiales y métodos
75
2.1. Insectos
75
2.1.1. Respuesta de larvas a plantas de col
75
2.2. Plantas
76
2.3. Determinación de la respuesta de larvas en plantas de col
76
2.4. Extracción e identificación de volátiles de col
76
2.5. Respuesta de larvas a extractos de col
76
2.6. Análisis de datos
77
3. Resultados
78
3.1. Respuesta de larvas a plantas de col
78

v
3.2. Respuesta de larvas a extractos de col
79
4. Discusión
80
VII. Discusión general
84
1.- Atracción de hembras de C. decolora a volátiles de col sin daño y con daño
2. Efecto del estado fisiológico y la experiencia sobre la respuesta a los volátiles
de planta hospedera y feromona sexual de C. decolora
3.- Preferencia de oviposición por C. decolora
4. Atracción de larvas de C. decolora a volátiles de la col
Conclusiones
84
85
87
88
91
Bibliografía 92
Anexos 112

vi
Relación de cuadros
Cuadro 1. Compuestos químicos identificados en CG-EM del extracto de col.
36
Relación de figuras
Figura 1. Eventos de búsqueda y aceptación de la planta hospedera por las
hembras.
4
Figura 2. Eventos de búsqueda y aceptación de la planta hospedera por las larvas.
8
Figura 3. Sistema de extracción de volátiles de plantas de col, en diferentes
condiciones.
23
Figura 4. Túnel de viento para evaluar la respuesta de hembras a volátiles de col.
25
Figura 5. Respuesta (%) de hembras apareadas a volátiles de col con diferentes
disolventes.
27
Figura 6. Respuesta (%) de hembras apareadas a volátiles de col sin daño y con
daño, en túnel de viento.
28
Figura 7. Despolarización (Promedio±EEM) provocada por el extracto de col
en antenas de hembras apareadas
37
Figura 8. Respuesta (%) de hembras apareadas a extracto de col y la mezcla de
sintéticos, en túnel de viento.
38
Figura 9. Respuesta (%) de hembras vírgenes y apareadas a extracto de col, en 48

vii
túnel de viento
Figura 10. Respuesta (%) de grupos de machos vírgenes a feromona sexual
49
Figura 11. Latencia de activación y latencia de aterrizaje (Q1 < Mediana < Q2)
por grupo de edad de machos vírgenes en respuesta al extracto glandular de
feromona sexual
50
Figura 12 Respuesta (%) de machos vírgenes y apareados a extracto glandular
de feromona sexual
51
Figura 13. Latencia de activación y latencia de aterrizaje (Q1 < Mediana < Q2) de
machos vírgenes y apareados en respuesta al extracto glandular de feromona
sexual
52
Figura 14. Respuesta (%) provocada por el extracto de col en machos y hembras.
53
Figura 15. Respuesta (%) provocada por el extracto glandular de feromona sexual
en machos y hembras en túnel de viento.
54
Figura 16. Respuesta (%) de hembras sin pre-exponer y pre-expuestas al extracto
de col en túnel de viento.
55
Figura 17. Latencia de activación y latencia de aterrizaje (Promedio ± EEM) de
hembras sin pre-exponer y pre-expuestas al extracto de col en el túnel de viento
56
Figura 18. Respuesta (%) de machos sin pre-exponer y pre-expuestos al extracto
glandular de feromona sexual en túnel de viento.
57

viii
Figura 19. Latencia de activación y latencia de aterrizaje (Promedio ± EEM) de
machos sin pre-exponer y pre-expuestos al extracto glandular de feromona sexual
en el túnel de viento.
58
Figura 20. Cámara para evaluar la preferencia de oviposición por hembras de C.
decolora.
68
Figura 21. Número de huevecillos (Promedio ± EEM) sobre plantas de col o
alrededor de ésta.
69
Figura 22. Olfatómetro en Y para evaluar la atracción de larvas neonatas. 78
Figura 23. Larvas neonatas (Promedio ± EEM) sobre plantas de col o alrededor de
ésta.
79
Figura 24. Respuesta de larvas neonatas a extractos volátiles de col con diferentes
disolventes y a la mezcla de sintéticos.
80

ix
Resumen
Copitarsia decolora es una plaga de plantas crucíferas, y la col es una de sus principales
plantas hospederas. El manejo de C. decolora puede incluir el uso de los volátiles de las
plantas hospederas como modificadores del comportamiento. Para C. decolora no se
han realizado estudios sobre los compuestos químicos involucrados en la atracción a la
planta hospedera, de algunos factores que afectan esta respuesta de atracción, además de
los factores que afectan a los machos en la búsqueda de pareja. Por lo anterior, se
propuso estudiar los compuestos volátiles de la col y de algunos factores bióticos que
afectan el comportamiento de atracción de C. decolora en la búsqueda y aceptación de
su planta hospedera y de pareja. Para lo cual, se obtuvieron extractos de volátiles de col
usando plantas sin daño y con daño, además de extractos glandulares de feromona
sexual. Las pruebas en olfatómetro mostraron que el extracto de col sin daño, del cual se
identificaron 9 compuestos volátiles, fue el que provocó más vuelos orientados hacia los
volátiles. Este extracto provocó la máxima respuesta de atracción en hembras apareadas
de 4-6 días de edad. Las hembras de 4 días de edad, se pre-expusieron a volátiles de la
col y se evaluó la atracción 2, 24 y 48h después, observándose una disminución de la
atracción en comparación con las hembras no pre-expuestas. Los machos de 4-7 días de
edad responden en mayor proporción a la feromona sexual, y al ser pre-expuestos a
dicho estímulo, se observó un efecto de inhibición a las 2 h y sensibilización a las 24 h.
Las hembras apareadas pusieron más huevecillos alrededor de la planta de col que sobre
ella. Las larvas neonatas al eclosionar a diferentes distancias de la planta de col, se
desplazan hacia la planta hospedera para su alimentación. El extracto de col sin daño
provocó la máxima atracción en las larvas neonatas, y aproximadamente la mitad de las
larvas evaluadas con la mezcla de compuestos identificados se dirigieron a dicho
estímulo. Esta investigación aporta conocimiento sobre la identidad de los compuestos
que participan en la atracción de hembras adultas de C. decolora, y posiblemente de
larvas, en la búsqueda de la planta hospedera. El estudio de algunos factores que afectan
esa búsqueda de hospedera por las hembras y de pareja por el macho. La oviposición
por las hembras ocurre alrededor de la planta y las larvas están involucradas en parte del
proceso de aceptación de la planta hospedera, ya que se desplazan hacia ella para
alimentarse.

x
Abstract
Copitarsia decolora is a pest of cruciferous plants and mainly cabbage is one of its host
plants. An alternative C. decolora management may include the use of host plant
volatiles and sex pheromone. For C. decolora studies have been conducted on the
chemical compounds involved in attracting the host, and the factors affecting this
response attraction, addition the factors affecting males in partner search Therefore, it
was proposed to study the volatiles of cabbage and sex pheromone and some biotic
factors that affect the behavior of attraction in the search C. decolora and acceptance of
their host plant and partner search. For this purpose, obtaining volatile extracts of
cabbage plants with damage and without damage was done, and sex pheromone gland
extracts. The extract of cabbage without damage was provoked longer flights to the
volatile and 9 volatile compounds were identified. This extract provoked maximum
response in females mated 4-6 days of age. Mated females of 4 days of age was pre-
exposed to volatiles from cabbage and attraction 2, 24 and 48h after evaluated, and it
was observed a decrease in the attraction compared with non-pre-exposed females.
Males 4-7 days of age respond most to the sex pheromone, and being pre-exposed to
said stimulus, an inhibition at 2 h was observed. Females prefer to lay eggs around the
cabbage plant, with a pattern of distribution of number of eggs up to 30 cm and then a
gradual decrease of eggs to cabbage plant. The neonate larvae to hatch at different
distances from the cabbage plant, moving toward her, taking part in the acceptance of
the host for food. The extract of cabbage without damage caused maximum attraction in
neonate larvae, and approximately half of the larvae tested the mixture identified
compounds were directed to this stimulus. This research provides knowledge about the
identity of the compounds involved in attracting adult females of C. decolora and
possibly larvae in search of the host plant. The study of some factors affecting this host
search by females and the male partner. Oviposition by females occurs around the plant
and the larvae are involved in part of the process of acceptance of the host plant, and
moving toward her to feed.

1
I. Introducción general
Los insectos herbívoros tales como las palomillas, discriminan olores entre
distintas plantas (De Bruyne y Baker, 2008), detectando el olor “específico” que le
permite ubicar a su planta hospedera en los procesos de oviposición y/o alimentación
(Visser, 1986; Bruce y Pickett, 2011). Muchas palomillas, representan un problema
económico en la agricultura causando pérdidas importantes en el rendimiento de sus
plantas hospederas (Tovar et al., 2007; Venette y Gould, 2006). Para el manejo de estos
insectos plaga se han utilizado insecticidas; pero éstos contaminan el ambiente y no son
específicos (Barrera et al., 2006). Los modificadores del comportamiento tales como las
feromonas y los volátiles de plantas hospederas pueden ser integrados en sistemas de
manejo de estos insectos (Schoonhoven et al., 2005). Para el empleo de compuestos
atrayentes en trampas y capturar hembras, primero se requiere hacer estudios de análisis
e identificación de los compuestos atractivos y los bioensayos de comportamiento para
conocer la respuesta del insecto a esos compuestos (Barrera et al., 2006).
El proceso de selección de la planta hospedera no sólo depende del estímulo
olfatorio, la recepción de volátiles y la respuesta a los mismos (Proffit et al., 2011),
también depende, entre otros factores, del estado fisiológico y la experiencia de la
hembra (Anton et al., 2007). Se conoce que la selección de la planta hospedera lo
realizan los adultos, específicamente las hembras; sin embargo, varios estudios han
demostrado que las hembras no siempre seleccionan la planta más apropiada, y que las
larvas recién emergidas pueden rechazar la planta sobre la que ellas eclosionaron y
seleccionar una planta más adecuada para su alimentación (Renwick y Chew, 1994).

2
En Copitarsia decolora (Guenée), se desconoce en gran parte el proceso de
selección de la planta hospedera, ya que no se sabe si existe una correlación adecuada
entre la selección de la hospedera por la hembra, y el desarrollo y supervivencia de las
larvas. En este trabajo, se estudian los compuestos volátiles emitidos por la planta
hospedera y algunos factores bióticos que están involucrados en la búsqueda y
aceptación de la planta hospedera por parte de hembras adultas y larvas neonatas, y de
pareja por lo machos de C. decolora.
1. Selección de la planta hospedera por noctuidos adultos
En los insectos herbívoros la búsqueda y aceptación de la planta hospedera
difiere entre las especies de acuerdo a si se alimentan u ovipositan en varias familias de
plantas hospederas (herbívoros generalistas) o en una o pocas especies de plantas
(herbívoros especialistas) (Bernays y Chapman, 1994; Rojas et al., 2003).
Los herbívoros generalistas utilizan estímulos comunes que están presentes en
sus plantas hospederas, para esto deben procesar la información visual, olfativa y táctil
proveniente de la planta hospedera, en contraste, los herbívoros especialistas utilizan
estímulos específicos de sus plantas hospederas, enfocándose en estímulos relevantes ya
sean visuales, olfativos o táctiles (Bernays y Chapman, 1994; Schoonhoven et al.,
2005). Sin embargo, esta clasificación no es estricta, ya que hay un gradiente entre
especies que se alimentan de un solo tipo de planta y aquellas que consumen varias
plantas. Además, la elección de una planta hospedera puede deberse a la combinación
de varios factores, es decir a una señal multimodal (Bernays y Chapman, 1994) más que
a un factor único (Marchioro y Foerster, 2014).

3
En general, el proceso de selección de la planta hospedera en insectos noctuidos
muestra una secuencia comportamental de modalidades sensoriales olfativas y
gustatorias, y en algunos casos, visuales y mecánicas (Bernays y Chapman, 1994; Rojas
y Wyatt, 1999a). Pero, el estímulo considerado más importante en el proceso de
selección de la planta hospedera para los noctuidos es el olfativo. Existe evidencia que
en algunas especies la atracción a la planta hospedera puede ser posible sólo con el
estímulo olfativo, sin el estímulo visual (Bernays y Chapman, 1994; Schoonhoven et
al., 2005).
En insectos noctuidos, las hembras son quienes seleccionan la planta hospedera
para su alimentación u oviposición, ya que esto determina la supervivencia de su
descendencia (Renwick y Chew 1994; Marchioro y Foerster, 2014).
1.1. Búsqueda de la planta hospedera por adultos
En las palomillas nocturnas, los estímulos olfativo y táctil son preponderantes
durante el proceso de selección de la planta hospedera (Rojas et al., 2003; Schoonhoven
et al., 2005). Los eventos conductuales que lleva a cabo una palomilla a la selección de
la planta hospedera son orientación, búsqueda, aterrizaje y evaluación al contacto, y
posteriormente la aceptación o el rechazo; lo anterior se puede resumir en búsqueda-
localización y aceptación de la planta hospedera (Thompson y Pellmyr, 1991; Renwick
y Chew 1994; Honda, 1995; Knolhoff y Heckel, 2014) (Figura 1).

4
Figura 1. Eventos de búsqueda y aceptación de la planta hospedera por las hembras (Esquema
modificado de Knolhoff y Heckel, 2014).
En la fase de búsqueda y localización de la planta hospedera, los compuestos
volátiles liberados por la planta son recibidos por quimiorreceptores (Leal, 2013)
localizados en las antenas del insecto (Keil, 1999), en donde las neuronas receptoras
olfatorias transmiten la información al lóbulo antenal, el cual procesa la información
olfatoria y genera una respuesta motora (Hildebrand, 1995). Estos compuestos pueden
ser percibidos a distancia de la planta hospedera y entre los más comunes están los
terpenos, alcoholes, aldehídos y ésteres (Dicke y van Loon, 2000). Todos éstos son de
bajo peso molecular y se volatilizan al ser expuestos al aire o a temperatura ambiente
porque tienen presiones de vapor altas (Kessler y Morrell, 2010).
Los insectos pueden detectar los volátiles de la planta hospedera entre los demás
volátiles del ambiente, el volátil o los volátiles “específicos” tienen un papel clave en la
ubicación de la planta hospedera, aún a la distancia. En la búsqueda de la planta
hospedera el insecto tiene que realizar movimientos orientados hacia la planta y tener
contacto con ella, aunque no siempre el insecto aterriza sobre la planta, pero al menos
éste debe permanecer en la proximidad de la planta, donde percibe otros estímulos con

5
la finalidad de seguir examinando sus características y aceptarla como sitio de
oviposición (Visser, 1986; Bernays y Chapman, 1994; Knolhoff y Heckel, 2014).
1.2. Aceptación de la planta hospedera por adultos
En la fase de aceptación de la planta hospedera, los insectos fitófagos utilizan
principalmente el estímulo químico (olfativo y táctil) para evaluar la calidad de la planta
hospedera como sitio de oviposición (Del Campo et al., 2001; Simmonds, 2001; Scheirs
y De Bruyn, 2002; Calas y Marion-Poll, 2007; Gripenberg et al., 2007a; Gripenberg et
al., 2007b). Para ello, la palomilla tiene receptores sensoriales en los tarsos, antenas,
probóscide y ovipositor (Leal, 2013) que le permitirán elegir una planta hospedera en
condiciones “óptimas” o con “calidad” para oviposición, es decir una planta con los
requerimientos nutricionales para la supervivencia y crecimiento para las larvas
(Awmack y Leather, 2002), y que además las larvas tengan la capacidad para
contrarrestar sus defensas químicas (Knolhoff y Heckel, 2014).
La presencia o ausencia de algunos metabolitos secundarios (estimulantes o
disuasivos de la oviposición) y nutrientes presentes en la planta hospedera, determinará
la aceptación de ésta por la palomilla (Bernays y Chapman, 1994; Honda, 1995;
Schoonhoven et al., 2005). Los compuestos estimulantes regularmente no son volátiles
(Simmonds, 2001; Calas y Marion-Poll, 2007), aunque existen ejemplos donde la
oviposición es estimulada por compuestos volátiles (Städler, 1974; Fatzinger y Merkel,
1985) o por una mezcla de ambos (Renwick et al., 2006).
La aceptación de la planta hospedera por el adulto está correlacionada con la
disponibilidad de la planta hospedera para el desarrollo y la supervivencia de su

6
progenie (Thompson y Pellmyr, 1991; Knolhoff y Heckel, 2014), aunque no siempre se
da esta correlación (Potter et al., 2012).
Cuando la preferencia de la hembra por la planta hospedera y el desarrollo larval
no coinciden, puede deberse a:
- El herbívoro generalista quede condicionado a la planta de la cual se
alimentó en su fase larvaria (principio de selección de Hopkins) (Barron,
2001).
- La experiencia tanto de adultos o las larvas sobre la planta hospedera
(Landolt y Molina, 1996; Cunningham et al., 1998b; Rojas y Wayt 1999b;
Papaj y Prokopy, 1989; Barron, 2001; Akhtar y Isman, 2003; Olsson et al.,
2006).
- La presencia de compuestos repelentes (Renwick y Chew, 1994).
- La presencia de parasitoides y depredadores por el daño al comienzo de la
oviposición (Meiners y Hilker, 2000; Hilker y Meiners, 2002; Hilker et al.,
2002).
- Cuando las hembras no son capaces de evaluar la “calidad” de la planta
correctamente (Jaenike, 1978).
- La presión de ovipositar por la carga de huevecillos (Jones, 1977; Doak et
al., 2006).
- La diversidad y distribución de vegetación alrededor de la planta hospedera
(Rausher y Papaj, 1983; Petzold, 2009).
- Cuando el desarrollo larval requiera moverse a otras plantas hospederas
(Bar-Yam y Morse, 2011).

7
- Las larvas por sí mismas eligen la planta hospedera para su alimentación
independientemente del sitio de oviposición elegido por la hembra
(Cunningham et al., 2011; Rivera y Burrack, 2012).
- Cuando las hembras prefieren ovipositar en plantas libres de enemigos
naturales que en plantas en condiciones “óptimas” para el crecimiento de las
larvas (Jeffries y Lawton, 1984).
2. Selección de la planta hospedera por larvas
Regularmente las palomillas herbívoras ovipositan sus huevecillos sobre la
planta hospedera, al emerger las larvas pueden quedarse o emigrar a otra planta
hospedera, en la mayoría de los herbívoros especialistas sus estados inmaduros se
alimentarán de la planta hospedera en la cual emergieron de huevo. En contraste con los
herbívoros generalistas que pueden poner sus huevecillos sobre la planta hospedera,
alrededor de ella o en otra planta que no sea su hospedera; es en estos casos cuando las
larvas, tienen opción de elegir la planta hospedera adecuada para su supervivencia y
crecimiento (Petzold, 2009; Knolhoff y Heckel, 2014) (Figura 2).
En la mayoría de los estudios donde se reporta el comportamiento de las larvas a
los volátiles de la planta hospedera, no identifican los compuestos químicos causantes la
atracción (Singh y Mullick, 2002; Zalucki et al., 2002; Castrejon et al., 2006). En
algunos casos las hembras y las larvas son atraídas por los mismos compuestos que
libera la planta hospedera (Rembold et al., 1991; Hartlieb y Rembold, 1996).

8
Figura 2. Eventos de búsqueda y aceptación de la planta hospedera por las larvas de lepidóptero
noctuido para su alimentación (Esquema modificado de Knolhoff y Heckel, 2014).
En las larvas de lepidópteros sigue siendo sumamente importante el estímulo
químico (olfativo y táctil), éstas se caracterizan por tener aproximadamente el 80% de
los quimiorreceptores en las antenas y los palpos maxilares (De Boer, 2006). Aunque en
algunos casos como en las larvas neonatas de Epyphias postvittana (Walker) usan la
señal visual (Harris y Foster, 1995) y química (Harris et al., 1999) para orientarse, y
encontrar la planta hospedera.
Además de los volátiles de la planta hospedera que afectan el comportamiento
locomotor de las larvas, también se han reportado que las plantas contienen inhibidores
de la alimentación (Landolt et al., 1998), pero ante esto último, las larvas pueden
modificar los compuestos e inhibir a los compuestos tóxicos o disuasivos de la
alimentación de la planta (Thompson y Pellmyr, 1991).

9
3. Emisión de volátiles por las plantas
Los compuestos químicos producidos por las plantas se clasifican según
modifiquen el comportamiento de los insectos (Bernays y Chapman, 1994) y pueden ser
repelentes o atrayentes de insectos en la búsqueda de la planta hospedera.
Por una parte, un atrayente es un compuesto o mezcla química que atrae al
insecto con movimientos orientados a la fuente de olor (Miller et al., 2009). Estos
compuestos atrayentes que liberan las plantas sin daño, sirven para que el insecto se
guíe hasta la planta hospedera, pero también se sabe que algunos volátiles atrayentes
son inducidos por el daño de larvas en la planta hospedera (Dicke y van Loon, 2000).
Por otra parte, un compuesto químico repelente provoca que el insecto realice
movimientos orientados para alejarse de la fuente de olor (De Moraes et al., 2001;
Miller et al., 2009). Estos compuestos son un mecanismo de defensa contra los
herbívoros alejándolos de la planta hospedera (Bernays y Chapman, 1994).
Los volátiles producidos por las plantas como respuesta al ataque de un
herbívoro son similares estructuralmente a los que liberan las plantas sin daño, sin
embargo, los herbívoros tienen la capacidad de distinguir los volátiles entre una planta
sin daño o con daño (Turlings y Tumlinson, 1992; Turlings et al., 1995). El daño
mecánico en las plantas ocasiona una respuesta volátil rápida y de corta duración, pero
cuando el herbívoro se alimenta de la planta, la respuesta es mayor en intensidad y
tiempo, además, las secreciones orales de los herbívoros inducen respuestas volátiles
específicas en las plantas (Turlings et al., 1998).
Son tres las principales rutas biosintéticas relacionadas con la producción de los
compuestos volátiles en plantas: la ruta de los ácidos grasos/lipoxigenasa que da origen
a los volátiles de hoja verde, la ruta del mevalonato/no mevalonato que da origen a los

10
terpenos, y la ruta del ácido shikímico/triptófano que da origen a los compuestos
aromáticos (Paré y Tumlinson, 1996; Paré y Tumlinson, 1999; Arimura et al., 2009;
Scala et al., 2013).
Los volátiles de hoja verde son compuestos de 6 carbonos, como aldehídos,
alcoholes y ésteres (Dudareva et al., 2004). Se liberan cuando la planta presenta daño
por herbívoros o sufren de estrés biótico/abiótico. El primer compuesto de la ruta
biosintética de los volátiles de hoja verde es el (Z)-3-hexenal, el cual se forma en el sitio
del daño inmediatamente después del ataque de herbívoros o daño mecánico (Arimura
et al., 2009). El (Z)-3-hexenal es reducido por una enzima alcohol deshidrogenasa a (Z)
-3-hexenol y este compuesto es convertido a (Z)-3-hexenil acetato por una enzima
alcohol aciltransferasa.
Algunos compuestos volátiles de hoja verde son sintetizados de novo y liberados
al momento, en horas, o días después del daño (Paré y Tumlinson, 1997a, b). En una
planta sin daño éstos son emitidos en cantidades traza y se emiten inmediatamente
después de la ruptura celular (Turlings et al., 1995; D’Auria et al., 2007).
Se han identificado compuestos volátiles característicos de grupos de plantas
hospederas, como los glucosinolatos de la familia Brassicaceae. Estas sustancias pueden
variar cualitativa y cuantitativamente entre genotipos de plantas, tejidos y estados
ontogénicos, por lo que la influencia en los insectos herbívoros puede ser variada. Los
glucosinolatos son defensas constitutivas, sus niveles están influenciados por factores
abióticos y bióticos que incluyen el daño provocado por el insecto (Pivnick et al., 1994;
Hopkins et al., 2009; Städler y Reifenrath, 2009).
En especies de plantas Brassica y Sinapsis, Tollsten y Bergström (1988)
identificaron compuestos como alcoholes, aldehídos y productos de degradación de

11
glucosinolatos. Los isotiocianatos estuvieron presentes en hojas maceradas, mientras
que los terpenos se identificaron en plantas completas cortadas desde el tallo (Tollsten y
Bergstrom, 1988).
En diversos estudios, se han identificado los compuestos volátiles de las plantas
y su efecto biológico. Para la identificación se ha usado la cromatografía de gases
acoplada a espectrometría de masas (CG-EM), una técnica suficientemente sensible
para identificar compuestos volátiles desconocidos en bajas concentraciones, además,
los registros en electroantenografía (EAG) han permitido confirmar la actividad
biológica a los compuestos más atractivos (Hartlieb y Rembold, 1996).
Por ejemplo, Fraser et al. (2003) en Manduca sexta (Linnaeus), analizaron
extractos de volátiles de plantas hospederas, encontrando 23 compuestos (alifáticos,
aromáticos y terpenoides) con actividad biológica e identificaron 15 de ellos en CG-
EM. Una mezcla de 8 compuestos comunes en sus plantas hospederas como el (Z)-3-
hexenil acetato, nonanal, decanal, fenil acetaldehído, metil salicilato, benzil alcohol,
geranil acetona, (E)-nerolidol) fue evaluada y encontraron que para las hembras, la
mezcla causó más atracción que los compuestos evaluados individualmente.
En otro estudio, Tasin et al. (2006) observaron que las hembras de Lobesia
botrana (Denis y Schiffermüller) son atraídas a una mezcla de 10 compuestos de planta
hospedera, aunque también se demostró que una combinación de tres compuestos, entre
ellos el compuesto más abundante el (E)-β-cariofileno, provocó el mismo efecto
atrayente.
La mezcla de compuestos atrayentes debe estar en una proporción tal y como la
liberan las plantas; aunque Najar-Rodríguez et al. (2010) reportan que la proporción de
compuestos de la mezcla puede variar un poco sin disminuir la atracción del insecto. De

12
manera general, se ha reportado que los insectos herbívoros responden a una mezcla de
compuestos más que a un solo compuesto (Finch y Collier, 2000; Bruce et al., 2005).
Compuestos de la familia Brassicaceae han sido reportados por Reddy y
Guerrero (2000), en extracciones de plantas de col dañadas mecánicamente se
identificaron 7 compuestos: 1-hexanol, hexanal, hexil acetato, (E)-2-hexenal, (Z)-3-
hexen-1-ol, (Z)-3-hexenil acetato y 1-hexen-3-ol, sin detectar isotiocianatos. En pruebas
de laboratorio, el (Z)-3-hexenil acetato fue el más atractivo para la palomilla Plutella
xylostella (Linnaeus), y en mezcla con feromona sexual aumentó las capturas de
hembras encampo.
Para Rojas (1999) el alil isotiocianato identificado en extractos de col estimuló
las hembras apareadas de M. brassica para que aterrizaran sobre la planta. También
identificó volátiles como el (E)-2-hexenal y el (Z)-3-hexenil acetato, los cuales
provocaron vuelos hacia la fuente de emisión.
4. Factores que influencian la selección en adultos
El proceso de búsqueda de la planta hospedera y de búsqueda de pareja no sólo
depende del estímulo olfatorio, la recepción del estímulo y la respuesta a los mismos
(Proffit et al., 2011), puede depender en gran medida de factores fisiológicos tales como
la edad de la hembra en la que se aparea (Pivnick et al., 1990; Browne, 1993), edad y
estado de apareamiento del macho (Roelofs, 1995) y de la experiencia a los volátiles de
hospedera (Pivnick et al., 1990; Petzold, 2009) y a la feromona sexual (Anton et al.,
2007).
El efecto de la edad al apareamiento se ha estudiado en algunas especies de
palomillas (Browne, 1993), por ejemplo Mechauber et al. (2002) mencionan que las

13
hembras de 4 días de edad apareadas tuvieron la máxima respuesta a los volátiles de su
planta hospedera, además, mostraron el ovipositor como señal de oviposición.
Para Rojas (1999a) e Ikeura et al. (2010) las hembras apareadas de M. brassicae
y Pieris rapae (Linnaeus) respectivamente, presentaron la máxima respuesta hacia los
volátiles de planta hospedera. En algunas palomillas las hembras vírgenes son
fuertemente atraídas a los volátiles de la planta hospedera (Saveer et al., 2012).
En el caso de los machos, en algunas especies de palomillas (Anton y Gadenne,
1999; Gadenne y Anton, 2000; Gadenne et al., 2001; Zhang et al., 2009; Soques et al.,
2010; Barrozo et al., 2010a, 2010b; 2011) la edad y el estado de apareamiento
modifican el proceso de recepción y procesamiento olfatorio de la feromona sexual.
La condición fisiológica en un insecto es fundamental para elegir la planta
hospedera y la pareja para apareamiento, sin embargo, su capacidad de aprendizaje les
permitirá adaptarse a las condiciones ambientales cambiantes (Landolt y Molina, 1996;
Olsson et al., 2006; Dukas, 2008; Dukas et al., 2012).
Para Matthews y Matthews (2010) el aprendizaje es un cambio adaptativo en el
comportamiento y ocurre como resultado de la experiencia o la práctica. En palomillas,
se ha documentado que el aprendizaje cambia la respuesta a las señales olfativas
relacionadas con la oviposición (Landolt y Molina, 1996; Cunningham et al., 1998b;
Rojas y Wayt, 1999b), la alimentación (Cunningham et al., 1998a), y la atracción sexual
(Anderson, 2003; Anderson et al., 2007).
Sin embargo, no es claro cuánto puede influir el aprendizaje en la respuesta a los
estímulos olfativos (Prokopy y Lewis, 1993). En este contexto, hay varios tipos de
aprendizaje en insectos herbívoros el aprendizaje asociativo y el no asociativo, éste
último involucra a la habituación y a la sensibilización (Papaj y Prokopy, 1989).

14
La habituación es probablemente la forma más común de aprendizaje en
insectos, es una disminución gradual de la capacidad de respuesta a un estímulo, por
exposición repetida al mismo (Daly y Figueredo, 2000; Dukas, 2008; Matthews y
Matthews, 2010). Este tipo de aprendizaje ha sido reportado en varias palomillas, en
estudios de pre-exposición a volátiles de planta hospedera (Rojas y Wayt, 1999b) como
para feromona sexual (Bartell y Lawrence, 1973, 1976, 1977; Bartell y Roelofs, 1973;
Figueredo y Baker, 1992; Daly y Figueredo, 2000).
En la sensibilización por pre-exposición al estímulo, hay un aumento en las
respuestas después de estimulaciones consecutivas, provocadas por cambios en la
respuesta de las neuronas postsinápticas (Landolt y Molina, 1996; Cunningham et al.,
1998b; Dukas, 2008; Matthews y Matthews, 2010). Por ejemplo, hembras de
Helicoverpa armígera (Boddie) pre-expuestas a volátiles de plantas hospederas y
machos de Spodoptera litoralis (Boisduval) pre-expuestos a feromona sexual,
mostraron una sensibilización en la posterior respuesta de atracción hacia los mismos
estímulos, respectivamente (Cunningham et al., 1998b; Anderson, 2003; Anderson et
al., 2007).
5. Manejo de insectos plaga con semioquímicos
El control o manejo de insectos plaga incluye diferentes métodos entre los que se
encuentra el desarrollo de técnicas basadas en la manipulación del comportamiento, lo
cual implica comprender la biología y ecología del insecto plaga, es decir, el estudio de
condicionantes fisiológicas y la relación con su comportamiento, que permitan
interpretar de manera adecuada los datos que un sistema de trampeo pueda
proporcionar.

15
En métodos alternativos de manejo para insectos plaga de cultivos agrícolas, se
pueden usar compuestos que median las interacciones dentro y entre especies como los
semioquímicos. Por un lado, está el uso de feromonas (compuestos que median las
interacciones entre organismos de una misma especie, la interacción resulta beneficiosa
tanto para el emisor como para el receptor) (Foster y Harris, 1997), las cuales se
emplean en el monitoreo y trampeo masivo, por ejemplo, en la interrupción del
apareamiento, donde la saturación del ambiente con moléculas de feromona sexual
inhibe los receptores olfatorios de los machos lo cual impide el encuentro con la hembra
para el apareamiento.
Por otro lado, están los compuestos llamados aleloquímicos que median
interacciones entre diferentes especies, como por ejemplo las kairomonas, que son
sustancias químicas que intervienen en las relaciones interespecíficas de los insectos, en
donde el organismo receptor se beneficia y el emisor se perjudica (Metclaf y Metclaf,
1992).
A pesar de que las feromonas de insectos han sido la base de la manipulación del
comportamiento de los insectos, los estudios de identificación química y la actividad de
la aplicación acerca de los volátiles de plantas hospederas y su potencial uso en la
protección de cultivos contra insectos plaga se ha incrementado (Rodriguez-Saona y
Stelinski, 2009).
Para la aplicación de semioquímicos es necesario comprender los factores que
afectan a la relación entre insectos y plantas. Por ejemplo, para el empleo de
aleloquímicos en un sistema de manejo, es importante investigar cómo los insectos
encuentran y eligen sus plantas hospederas (Rodriguez-Saona y Stelinski, 2009).

16
Además, la identificación de los estímulos químicos de atracción o repulsión que
participan en la selección de la planta hospedera para el proceso de oviposición y la
preferencia de las larvas en los insectos herbívoros polífagos, proporcionaría una
herramienta para diseñar trampas que atraigan hembras y/o larvas recién emergidas, con
la finalidad de disminuir el daño a los cultivos agrícolas.
Una estrategia de manejo de manipulación del comportamiento con
aleloquímicos es el método Push-Pull, en donde el cultivo es protegido mediante
compuestos que enmascaran su presencia o bien repelen al insecto y simultáneamente
compuestos atrayentes desvían al insecto plaga hacia zonas de captura o plantas trampa,
donde son eliminados (Cook et al., 2007).
El empleo de semioquímicos como los volátiles sintéticos de planta hospedera
de C. decolora, en conjunto con lo que se conoce para el manejo con feromona sexual
en esta especie (Rojas et al., 2006; Muñiz et al., 2007; Gómez et al., 2012), ofrece una
oportunidad para diseñar una estrategia de manejo para este insecto plaga que involucre
ambos estímulos olfativos. Tal estrategia permitiría monitorear C. decolora antes de que
afecte al cultivo, dando una gestión sostenible en la práctica de los sistemas agrícolas
que actualmente dependen en gran medida de las aplicaciones de insecticidas.
6. Copitarsia decolora como modelo de estudio
C. decolora conocida comúnmente como “gusano del corazón de la col” es una
plaga importante de plantas crucíferas en México, Centro y Sudamérica; es una especie
cuarentenada en los Estados Unidos de América (Venette y Goul, 2006). En México se
localiza principalmente en la col (Brassica oleracea var. capitata Linnaeus) (Suárez-
Vargas et al., 2006) presentándose todo el año, alimentándose del brote principal
ocasionándole deformación e inclusive la muerte de la planta (Tovar et al., 2007).

17
Para el manejo de C. decolora, se han utilizado insecticidas convencionales, el
manejo cultural, mediante prácticas agrícolas realizadas por los productores como
preparación del suelo, uso de invernaderos y eliminación de plantas afectadas (Cibrián,
1999).
Una alternativa para el manejo de este insecto, es el empleo de modificadores del
comportamiento, tales como los volátiles de planta hospedera, los atrayentes y los
repelentes. Al respecto, son escasos los estudios acerca del comportamiento de
búsqueda y la aceptación de la planta hospedera.
Se conoce que las larvas y las hembras de C. decolora, eligen plantas de col
entre diferentes plantas hospederas (Pérez, 2010; Acatitla, 2010). En estudios de
preferencia alimentaria de larvas neonatas hacia cinco especies de plantas hospederas,
las larvas se alimentaron significativamente más de plantas de col que de cualquier otra
planta evaluada (Perez, 2010). Pero no hay trabajos que precisen la identidad química
de los compuestos involucrados en la atracción de las hembras o larvas a la planta
hospedera.
Se sabe que la edad en la que las hembras de C. decolora alcanzan el máximo
potencial reproductivo posiblemente está relacionada con la máxima atracción a los
volátiles de planta hospedera (Rojas y Cibrian-Tovar, 1994; Callado-Galindo et al.,
2013), pero no se ha establecido la preferencia de oviposición sobre su planta hospedera
y el involucramiento de las larvas en la búsqueda y aceptación de la planta hospedera.
Por otra parte, en campo se ha evaluado la eficiencia de dispositivos en trampas con
feromona sexual sintética como posible alternativa de manejo al uso de insecticidas
sintéticos (Rojas et al., 2006; Muñíz et al., 2007; Gomez et al., 2012), pero se
desconoce el efecto de los diferentes factores ambientales (Dumont y McNeil, 1992)

18
sobre la respuesta de los machos a la feromona sexual. El conocimiento de la respuesta
hacia la feromona o a las plantas hospederas de las palomillas cuando se ven afectadas
por factores bióticos, ayudará a entender el proceso de atracción de los machos a las
trampas cebadas con feromona sexual saber si es posible la inclusión de volátiles de las
plantas en esa misma trampa diseñada para capturar machos y capturar hembras, lo que
generaría una herramienta de monitoreo o control más potente.
7. Objetivos
El objetivo de esta tesis fue generar y aportar conocimiento sobre el estudio de
los compuestos volátiles de col y de algunos factores bióticos que afectan el
comportamiento de atracción de Copitarsia decolora en la búsqueda y aceptación de su
planta hospedera, así como en la búsqueda de pareja por parte de los machos.
El estudio incluyó los siguiente capítulos; en el Capítulo II se estudió la
atracción de hembras de C. decolora a extractos de compuestos volátiles de hospedera,
se da a conocer el extracto de col con el disolvente que provocó la máxima atracción en
las hembras de C. decolora.
Posteriormente en el Capítulo III, se identificaron los compuestos volátiles del
extracto de col que fue más atractivo para las hembras, el tipo de compuestos atrayentes,
y la respuesta antenal y comportamental de las hembras al extracto y a la mezcla de
compuestos sintéticos identificados.
Para conocer si el estado fisiológico y la experiencia afectan la respuesta de
hembras a los volátiles de col en la búsqueda de planta hospedera y de machos a la
feromona sexual en la búsqueda de pareja, en el Capítulo IV, se estudió la edad de
apareamiento de las hembras, el efecto de la edad y el estado de apareamiento de los

19
machos, sobre las respuestas de atracción provocadas por extractos de col y extractos
glandulares de feromona sexual, respectivamente. Además, se estudió el efecto de la
pre-exposición de hembras apareadas a los volátiles de col y de machos vírgenes al
extracto glandular de feromona sexual sobre la respuesta de atracción a esos estímulos
químicos.
En el Capítulo V, se conoció la preferencia de oviposición por las hembras,
comparando como sitio de oviposición la planta de col o alrededor de ésta.
Finalmente en el Capítulo VI, se estudió la atracción de larvas neonatas a
compuestos volátiles de col, se investigó si las larvas se desplazan a la planta hospedera
y si son atraídas al mismo extracto de hospedera y a la misma mezcla de compuestos
sintéticos que atrae a las hembras.

20
II. Atracción de hembras de C. decolora apareadas a compuestos volátiles de la col
1. Introducción
Los olores que emiten las plantas, además de ser atrayentes para insectos
herbívoros, pueden ser también disuasivos de los procesos de oviposición y/o
alimentación (Bernays y Chapman, 1994; Renwick y Chew, 1994; Schoonhoven et al.,
2005).
Las plantas sin daño mantienen ciertos niveles de emisión de compuestos
volátiles orgánicos (Dudareva et al., 2004; Baldwin, 2010). Esta composición de los
volátiles que libera la planta hospedera puede variar cualitativa y cuantitativamente
(Bruce et al., 2005; Holopainen y Gershenzon, 2010; Bruce et al., 2010; Niinemets et
al., 2013) en condiciones de estrés abiótico, por ejemplo el daño mecánico (Gouinguene
y Turlings, 2002; Holopainen y Gershenzon, 2010) y biótico, por ejemplo por
herbívoros (Holopainen y Gershenzon, 2010).
En algunos estudios se ha observado que el daño mecánico no induce un
incremento en la emisión de volátiles (Bricchi et al., 2010), en cambio la emisión de
éstos aumenta drásticamente cuando los insectos herbívoros se alimentan de su planta
hospedera. Esta mezcla de compuestos volátiles emitidos por la planta en respuesta al
daño de herbívoros incluye a los llamados volátiles de hoja verde (Dudareva et al.,
2004; Hare, 2011). Por lo tanto, estos compuestos inducidos por herbivoría han sido
considerados como una forma de defensa indirecta en las plantas para combatir a los
herbívoros mediante la atracción de sus enemigos naturales (Turlings et al., 1995, De
Moraes et al., 1998; De Moraes et al., 2001; Heil et al., 2008; Howe y Jander, 2008;
Wu y Baldwin, 2010; Mumm y Dicke, 2010; Heil et al., 2012; Holopainen y Bland,
2013; Scala et al., 2013).

21
Los volátiles que liberan las plantas constituyen información química que puede
ser usada por algunas especies de palomillas que ovipositan en ellas, aun con el riesgo
de que sus huevecillos o larvas sean atacadas por depredadores o parasitoides (Loughrin
et al., 1995). El estudio de estas interacciones planta-insecto, han permitido entender
cómo las palomillas utilizan los diferentes tipos de volátiles y cómo modulan su
comportamiento en la búsqueda de la planta hospedera (Bernays y Chapman, 1994;
Shoonhoven et al., 2005).
En los adultos de C. decolora, son escasos los estudios acerca de los estímulos
olfativos involucrados en la búsqueda de la planta hospedera y su modificación por
efecto del daño en la planta. Por lo anterior, se propuso estudiar la atracción de hembras
apareadas a los extractos de volátiles de la col sin daño y con daño.
2. Materiales y Métodos
2.1. Insectos
Los insectos utilizados en los bioensayos se obtuvieron de la colonia de C.
decolora ubicada en el Centro de Desarrollo de Productos Bióticos-IPN en Yautepec,
Morelos, México; mantenida a 22°C (±3), 60 % (± 3) de HR y 12 horas luz: 12 horas
oscuridad, con la fotofase y escotofase invertidas respecto al ciclo de luz natural, para
realizar los bioensayos durante el día. Los estados inmaduros fueron alimentados con
una dieta para lepidópteros (Rojas et al., 2003). Se tuvo un intervalo de peso de las
pupas de 0.35 g a 0.45 de acuerdo a Callado-Galindo et al. (2013) para descartar la
influencia del mismo en la respuesta a los volátiles de col.
Los adultos se mantuvieron en jaulas de plexiglás (20x20x20cm) y se
alimentaron diariamente con solución azucarada al 50% colocada en un algodón, la cual
se cambió cada tercer día. En las jaulas de plexiglás se aparearon por jaula 30 machos y

22
30 hembras. En los bioensayos se utilizaron hembras apareadas de 4-5 días de edad
(n=10 por tratamiento).
2.2. Plantas
Las plantas de col se cultivaron en condiciones de invernadero a 20°C (±2), 60
% HR (± 2) y fotoperiodo 13 horas luz: 11 horas oscuridad. Estas se colocaron en
macetas de plástico (20 cm de alto por 25 cm de diámetro) conteniendo suelo estéril y se
requirieron de 30-40 días de crecimiento, antes de la floración y con un peso fresco
aproximado de las plantas de 120 g.
Para la colecta de volátiles se utilizaron plantas de col sin daño, con daño por
larvas (10 larvas de tercer estadio puestas en una planta durante media hora antes de la
extracción y retiradas al momento de la extracción) y con daño mecánico (10 cortes en
las hojas realizados 5 minutos antes de la extracción, cada corte de 1 cm).
2.3. Extracción de volátiles de la col sin daño y con daño
El sistema de colecta (Figura 3) consistió en colocar dentro de una cámara de
vidrio (30 cm de largo y 20 cm de diámetro) (modificado de Geervliet et al., 1997) una
planta de col sin daño o en diferentes condiciones de daño. Al sistema de colecta de
volátiles en cada planta se le adaptó en la entrada una placa de acrílico sujetando la
planta por el tallo y aislando el suelo, y en la salida de la cámara una micropipeta de
vidrio (13 cm largo × 0.6 cm diámetro externo) conteniendo 250 mg de material
adsorbente Super Q (Alltech Assoc, Inc., Deerfield, Illinois). A la cual se le conectó una
bomba de vacío (Welch® USA) con un flujo 1 L/min medido por un flujómetro (Cole
Palmer, Ev-03217-06, USA) para extraer los volátiles liberados por las plantas.
La colecta de volátiles se llevó a cabo durante tres días consecutivos usando una
planta/día durante las 3 primeras horas de la escotofase a 20°C y 60 % HR.

23
Posteriormente, para los extractos de plantas sin daño los compuestos capturados se
disolvieron en 3 mL de hexano (HPLC, JT Baker, EUA), diclorometano (HPLC, JT
Baker, EUA) o metanol (HPLC, JT Baker, EUA) Después, cada muestra se reconcentró
a 300 µL con corriente de nitrógeno y se almacenó a 4 ̊ C hasta su utilización en los
bioensayos. Para los extractos de plantas de col con daño se utilizó sólo hexano.
Figura 3. Sistema de extracción de volátiles de plantas de col en diferentes condiciones.
2.4. Respuesta de atracción en túnel de viento a extractos de col
El comportamiento de las palomillas hacia el estímulo olfativo (extracto de
volátiles de col sin daño con diferentes disolventes, daño por larva o daño mecánico) se
midió en un túnel de viento (Figura 4) de plexiglás (180x80x80 cm) equipado con un
extractor (Frecuency Inverter CFW-08 Software 4.1x, Mineapolis, USA) que generó

24
una corriente de aire a una velocidad de 40 cm/s y limpiado por un filtro de carbón
activado. La velocidad del viento se midió con un anemómetro (Sper Scientific 840003,
Taiwan) a 30 cm del piso y en el centro del túnel.
Los experimentos fueron realizados en las dos primeras horas de inicio de la
escotofase a 20°C (±3), 60 % (± 5) de humedad relativa (HR) y luz roja (3 focos rojos
de 20 watts). Las hembras fueron transferidas antes del inicio de la escotofase de su
cámara de cría al túnel de viento. En el extremo donde se ubica el extractor se colocó
una hembra y en el extremo opuesto se pusieron 10 µL (en estudios pre-eliminares, a 10
µL las palomillas presentaron una respuesta de atracción) del extracto de col sobre un
papel filtro Waltman No.1 (Whatman ® 2V, England) de 2 X 2 cm. Después de 20 s,
tiempo suficiente para que el disolvente se evaporara, la hembra fue liberada en el túnel
por 300 s y se registró su comportamiento. Después de cada bioensayo se limpió el
túnel de viento durante 300 s (flujo de viento sin estímulo) para eliminar cualquier
posible contaminación antes de continuar con los experimentos.
En el túnel de viento se registró cuando la hembra desplegó el ovipositor,
presentaba un vuelo orientado (vuelo mayor a los 150 cm en el túnel de viento) hacia el
estímulo (Modificado de Picimbon et al., 1997). No se incluyó el patrón de
comportamiento de aterrizaje, ya que en estudios pre-eliminares las hembras no
mostraron aterrizajes sobre el estímulo.
Por una parte, se compararon los datos de los patrones de comportamiento
mostrados por la hembra (desplegó el ovipositor y si presentó un vuelo orientado) hacia
el extracto de col sin daño con diferentes disolventes (Planta de col= PC, extracto de col
con hexano= EH, extracto de col con diclorometano= ED y extracto de col con
metanol= EM), la planta de col y con el control (Aire, hexano= H, diclorometano= D y

25
metanol= M). Por otra parte, se compararon los datos de los patrones de
comportamiento mostrados por la hembra (desplegó el ovipositor y vuelo orientado)
hacia el extracto de col sin daño con los mostrados por los extractos de col con daño por
larva y daño mecánico.
Figura 4. Túnel de viento para evaluar la respuesta de los insectos en diferentes condiciones a
los volátiles de col.
2.5. Análisis estadístico
Se realizó una prueba de χ² para probar si las respuestas de las hembras son
significativamente dependientes de los estímulos olfativos (p<0,05). Las pruebas se
realizaron con el programa estadístico Sigma Plot 11 (Systat Software Inc., Chicago,
Illinois).

26
3. Resultados
3.1. Respuesta de atracción en túnel de viento a volátiles de la col
No se encontraron diferencias en el patrón de vuelo orientado con extracto
hexánico en la atracción, comparando con una planta de col, pero si con respecto al
extracto con diclorometano (χ²= 7.521; gl= 1; p= 0.006) y el extracto con metanol (χ²=
72.521; gl= 1; p= 0.001) (Figura 5). Los grupos de insectos evaluados al control (aire,
hexano, diclorometano y metanol) no tuvieron ninguna respuesta.
El mayor porcentaje de hembras que desplegó el ovipositor se observó para la
planta de col y el extracto hexánico, aunque en esos grupos no hubo diferencia
significativa, pero si con el control (χ²= 82.881; gl= 1; p= 0.001), con el extracto con
diclorometano (χ²= 16.990; gl= 1; p= 0.001) y con el extracto de metanol (χ²= 82.881;
gl= 1; p= 0.001) (Figura 5).
El extracto hexánico de volátiles de col fue el que provocó la máxima respuesta
de atracción en las palomillas, debido a esto, el hexano fue el disolvente que se ocupó
para realizar las extracciones de volátiles de col sin daño o con daño.
El extracto de volátiles de col sin daño provocó una máxima respuesta de las
hembras apareadas de C. decolora, en comparación con los extractos de col con
diferentes tipos de daño. La diferencia significativa se observó en el patrón de
comportamiento de vuelo orientado (para daño por larva χ²= 16.990; gl= 1; p= 0.001 y
para daño mecánico χ²= 7.521; gl= 1; p= 0.006). Entre los tipos de daño en este patrón
de comportamiento no hubo diferencia significativa (Figura 6).

27
Tratamiento
PC EH ED EM
Re
sp
ue
sta
de
he
mb
ras a
vo
látile
s d
e c
ol (%
)
0
20
40
60
80
100
Vuelo orientado
Despliega ovipositor
b
c
c
a
bb
a
a
b
Figura 5. Respuesta (%) de hembras apareadas a una planta completa y a extractos volátiles de
col con diferentes disolventes, en túnel de viento. Planta de col= PC, extracto de col con
hexano= EH, extracto de col con diclorometano= ED y extracto de col con metanol= EM. Letras
diferentes sobre barras del mismo color indican valores significativamente diferentes al p<0.05
(χ²), n=10.
Para el patrón de comportamiento de desplegar el ovipositor hubo diferencia
significativa entre el extracto de col sin daño y daño por larva (χ²= 31.687; gl= 1; p=
0.001), sin daño y daño mecánico (χ²= 7.220; gl= 1; p= 0.007) y daño por larva y daño
mecánico (χ²= 8.595; gl= 1; p= 0.003) (Figura 6).

28
Condición del extracto de volátiles de col
Respuesta
de h
em
bra
s a
extr
acto
s d
e c
ol
(%)
0
20
40
60
80 Vuelo orientado
Despliega ovipositorb
a
a
a
c
b
Sin daño Daño por larva Daño mecánico
Figura 6. Respuesta de (%) de hembras apareadas a extractos volátiles de col sin daño y con
daño, en túnel de viento. Letras diferentes sobre barras del mismo color indican valores
significativamente diferentes al p<0.05 (χ²), n=10.
4. Discusión
Los resultados de este trabajo, muestran que entre los diferentes disolventes
utilizados para la extracción de volátiles de col, el extracto de col con hexano provocó la
máxima atracción en las hembras apareadas de C. decolora. Aunque las hembras de C.
decolora no presentaron el comportamiento de aterrizaje en la fuente de emisión,
podemos valorar el vuelo orientado hacia la fuente de olor como un indicador de
atracción (Visser, 1986). También, la hembra puede dar señales al momento de percibir
los volátiles de la planta hospedera, como es el desplegar el ovipositor como señal de
oviposición (Mechauber et al., 2002) como se observó en las hembras de C.decolora.

29
En otras especies de palomillas, se ha encontrado que solamente con
determinados disolventes se extraen los compuestos que estimulan a las hembras para la
búsqueda de planta hospedera, como es el caso de E. postvittana, los extractos de planta
hospedera con metanol y diclorometano no atrajeron a las hembras, pero si con pentano
(Foster y Howard, 1998).
En cuanto al efecto del daño en las plantas, se observó que el extracto de plantas
de col sin daño provocó mayor atracción en las hembras de C. decolora que aquellos
contenidos en extractos de plantas con los dos tipos de daño, a diferencia de lo que
ocurre en otras especies de palomillas como Helicoverpa armígera (Hübner) (Hartlieb y
Rembold, 1996), M. brassicae (Rojas, 1999) y P. xylostella (Reddy y Guerrero, 2000)
las cuales presentan mayor respuesta a extractos de su planta hospedera con daño.
Las hembras de C. decolora, tuvieron menor porcentaje de respuesta a los
extractos de col con daño por larvas que al daño mecánico. El daño en las plantas, ya
sea ocasionado por la alimentación de herbívoros, daño mecánico, o por la oviposición
(Wu y Baldwin, 2010), desencadena reacciones enzimáticas que cambian la
composición de la emisión de los volátiles (Agelopoulos y Pickett, 1998; Van Loon y
Dicke, 2001; Shiojiri et al., 2006) respecto a la condición sin daño, lo que afecta el
comportamiento de los insectos (Dicke et al., 1990; Schoonhoven et al., 2005;
Agelopoulos y Pickett, 1998).
Cuando las plantas presentan daño por herbívoros, hay incremento en la emisión
de compuestos volátiles y algunos de éstos se emiten hasta varias horas después del
daño (Paré y Tumlinson, 1998). Otros volátiles son liberados de manera inmediata al
daño tales como los volátiles de hoja verde, que pueden ser compuestos atractivos
(Turlings et al., 1995) o repulsivos a los insectos herbívoros (Allmann et al., 2013).

30
Los volátiles de hoja verde proveen información de la localización exacta del
ataque del herbívoro (Scala et al., 2013). Entre estos compuestos se han reportado
alcoholes, aldehídos y ésteres los cuales son sintetizados y emitidos, en cuestión de
segundos, a partir de ácidos grasos insaturados (Turlings et al., 1995; Paré y Tumlinson,
1997a; Gang et al., 2001; D’Auria et al., 2007; Arimura et al., 2009).
El tipo de volátiles que emite la planta hospedera guía a la palomilla en el
proceso de búsqueda (Späthe et al., 2013), en este sentido, C. decolora presentó mayor
vuelo orientado hacia extractos de col sin daño y menor hacia los extractos de col con
daño, aunque no se observaron aterrizajes en ningún tipo de estímulo. Normalmente las
hembras de noctuidos prefieren una planta hospedera en condiciones “óptimas” para el
crecimiento y desarrollo de su descendencia (Schoonhoven et al., 2005) como sucede en
otras palomillas (Landolt, 1993; De Moraes et al., 2001; Kessler y Baldwin, 2001;
Huang et al., 2009).
Cuando una hembra aterriza en una planta independientemente de su condición
“óptima” para el desarrollo larval, puede ser debido a que selecciona la planta hospedera
de la cual se alimentó en su etapa de larva (Barron, A. 2001).
Otra explicación a la correlación negativa entre la preferencia de la hembra por
la planta hospedera y el desarrollo larval es que en ocasiones las hembras prefieren
ovipositar en plantas libres de enemigos naturales que en plantas en condiciones
“óptimas” para el crecimiento de las larvas (Jeffries y Lawton, 1984). También, pueden
existir compuestos repelentes que alejen a la hembra de la planta hospedera (Renwick y
Chew, 1994).

31
III. Identificación de los compuestos volátiles de la planta de col
1. Introducción
La respuesta comportamental de orientación hacia la fuente de olor de los
insectos herbívoros en la búsqueda de la planta hospedera, depende de una mezcla de
compuestos más que a un compuesto por sí solo (Dicke y van Loon, 2000).
La identificación de los compuestos atractivos para los herbívoros es parte
integral del uso y la manipulación de los semioquímicos como componentes
importantes de una estrategia de manejo de plagas (Shoonhoven et al., 2005). Para esto,
la actividad antenal a compuestos identificados y el estudio comportamental en túnel de
viento debe tener una correlación positiva en función de la atracción de los insectos
herbívoros.
Es importante que la mezcla de compuestos atrayentes esté en una proporción tal
como la liberan las plantas; aunque Najar-Rodríguez et al. (2010) reportan que la
proporción de compuestos de la mezcla puede variar un poco sin disminuir la atracción
de la misma. Algunas palomillas como H. armígera son atraídas a las mezclas de
compuestos en la misma proporción que extractos de su planta hospedera (Rembold et
al., 1991; Hartlieb y Rembold, 1996).
La mezcla de compuestos como (Z)-3-hexenil acetato, nonanal, decanal, fenil
acetaldehído, metil salicilato, benzil alcohol, geranil acetona y (E)-nerolidol provocó en
las hembras de M. sexta mayor atracción que los compuestos evaluados individualmente
(Fraser et al., 2003). En otras palomillas, una combinación de tres compuestos volátiles
de hospedera, tienen el mismo efecto atrayente que la mezcla de 10 compuestos (Tasin
et al., 2006).

32
Algunos compuestos de la familia Brassicaceae han sido reportados por Reddy y
Guerrero (2000), como el (Z)-3-hexenil acetato. Este compuesto fue el más atractivo
para la palomilla P. xyllostella, y en mezcla con feromona sexual aumentó las capturas
de hembras en campo. El (Z)-3-hexenil acetato, junto con el (E)-2-hexenal identificado
en extractos de col estimuló las hembras apareadas de M. brassicae provocando vuelos
hacia la fuente de emisión (Rojas, 1999).
En C. decolora, no se han identificado los estímulos que la atraen a la planta de
col, por lo que en el presente estudio, se identificaron los compuestos de la planta de col
que provocan la atracción de las hembras apareadas, para ello, se evaluó la respuesta
antenal de las hembras apareadas a los compuestos identificados y la atracción de las
hembras a la mezcla de compuestos sintéticos en túnel de viento.
2. Materiales y Métodos
2.1. Insectos
Los insectos utilizados en los bioensayos se obtuvieron, mantuvieron y se
aparearon como lo descrito en el Capítulo II, en la Sección 2.1.
Para los bioensayos de electroantenografía (n=12) y de túnel de viento (n=10)
por tratamiento) se ocuparon las antenas de hembras apareadas de 4-5 días de edad.
2.2. Plantas
Para la extracción de volátiles se utilizaron plantas de col sin daño, las que se
obtuvieron y mantuvieron como lo descrito en el Capítulo II, en la Sección 2.2.
2.3. Extracción de compuestos volátiles de la col
Los extractos de plantas de col sin daño se realizaron como se describe en el
Capítulo II, en la Sección 2.3, con la variante de colocar dentro de la cámara de vidrio

33
sólo plantas de col sin daño. Además, los compuestos capturados se extrajeron con
hexano.
2.4. Análisis químico e identificación de los compuestos
Para el análisis e identificación del perfil químico del extracto de volátiles de
col, el cual fue más atractivo para las hembras apareadas en los bioensayos en túnel de
viento, se utilizó un cromatógrafo de gases (CG) (HP 6890) acoplado a un
espectrómetro de masas (EM) (HP 5972) (Agilent, EUA). Se utilizó una columna no
polar HP 5 MS (30 m, 250 µm de diámetro interno y 0.25 µm de espesor de película)
(Agilent, Palo Alto, CA). La temperatura inicial del horno fue de 60°C la que se
incrementó 4°C/min hasta 220°C (Tollsten y Bergstrom, 1988). El gas acarreador fue
helio a un flujo constante de 1 mL/min. La temperatura del inyector fue de 225°C y del
auxiliar de 250°, se utilizó el inyector en modo Splitless durante 18 s. El EM se trabajó
en ionización electrónica (70 EV), en modo SCAN y en un intervalo de masas de 35 a
550 UMA.
La identificación de los compuestos se llevó a cabo considerando sus tiempos de
retención e índices de retención (Kovats) (Kovats, 1958) y comparando los espectros de
masas con los de la biblioteca espectral (NIST/EPA/NIH. 2002) y por comparación con
estándares sintéticos (Sigma Aldrich®, Toluca, México). Se inyectaron alícuotas del
extracto de col de 2 µL.
Los compuestos químicos 6-metil-5-hepten-2-ona (pureza≥ 99 %), (Z)-3-hexenil
acetato (pureza≥ 98 %), limoneno (pureza≥ 98 %) nonanal (pureza≥ 98 %) decanal
(pureza≥ 98 %) tetradecano (pureza≥ 99.5 %) pentadecano (pureza≥ 98 %) hexadecano
(pureza≥ 99 %) y heptadecano (pureza≥ 99 %) fueron obtenidos de (Sigma Aldrich®,
Toluca, México).

34
2.5. Respuesta electroantenográfica al extracto y a la mezcla de sintéticos
El análisis y medición de las respuestas en electroantenografía (EAG) se efectuó
en un equipo Syntech (Kirchzarteng, Alemania). Una vez que la antena del insecto fue
desprendida, se montó entre dos electrodos metálicos, el electrodo registrador se colocó
en la parte distal del funículo, y el electrodo de referencia en el segmento de la base de
la antena. A ambos electrodos se les agregó gel conductor Syntech para facilitar el
registro de la respuesta. La señal generada por la antena fue trasmitida a un amplificador
IDAC-2 y observada en un monitor con el software EAG. La estimulación de la antena
fue por 1s con flujo de aire de 0.5 L/min transportado a la antena por una bomba
(Stimulus controler SC-55) con un flujo constante de aire puro humidificado (0.7
L/min).
Para la aplicación del estímulo, cada uno de los compuestos químicos
identificados del extracto de col y de la mezcla de compuestos identificados del extracto
de col, fueron colocados sobre piezas de papel Whatman No. 1 (1 x 0.5 cm). En cada
experimento se dejó evaporar el disolvente por 20 s, entre cada estímulo fue de 120 s y
entre cada antena fue de 300 s. En estos bioensayos, se consideraron los valores de
despolarización de las antenas, descontando las despolarizaciones provocadas por el
disolvente. Los compuestos fueron el 2 µL (6-metil-5-hepten-2-ona (14.58 pg/µL), (Z)-
3-hexenil acetato (12.35 pg/µL), limoneno (156.66 pg/µL) nonanal (148.89 pg/µL)
decanal (39.45 pg/µL) tetradecano (17.45 pg/µL) pentadecano (16.78 pg/µL)
hexadecaneo (18.11 pg/µL) y heptadecano (64.46 pg/µL).
2.6. Respuesta de atracción en túnel de viento al extracto y a la mezcla de sintéticos
Se utilizó el túnel de viento con las mismas condiciones descritas en la Sección
2.4 del Capítulo II, con la variante de que el estímulo utilizado para medir la atracción

35
de las hembras fue el extracto de col sin daño y la mezcla de compuestos sintéticos
incluyendo el 6-metil-5-hepten-2-ona (72.90 pg/µL), (Z)-3-hexenil acetato (61.77
pg/µL), limoneno (783.30 pg/µL) nonanal (744.45 pg/µL) decanal (197.23 pg/µL)
tetradecano (87.27 pg/µL) pentadecano (83.92 pg/µL) hexadecano (90.57 pg/µL) y
heptadecano (322.32 pg/µL) contra el extracto donde se identificaron los compuestos
químicos. La mezcla de compuestos sintéticos se preparó con la proporción de cada
compuesto similar al que se encontró en el extracto de col.
2.7. Análisis estadístico
Los datos obtenidos de la despolarización provocada en la antena de las hembras
por el extracto de col, mezcla y compuestos individuales sintéticos, fueron analizados
por medio de un análisis de varianza unifactorial para medidas repetidas (ANOVA MR),
después de la transformación de datos (lnx) y para las comparaciones entre grupos se
hizo una prueba de Tukey.
Los datos de los patrones de comportamiento mostrados por las hembras en túnel
de viento, se analizaron con una prueba de χ². Las pruebas se realizaron con el programa
estadístico Sigma Plot 11 (Systat Software Inc., Chicago, Illinois).
3. Resultados
3.1. Identificación de los compuestos volátiles
En el análisis del extracto de col en CG-EM se identificaron 9 compuestos, una
cetona, un éster, un terpeno, 2 aldehídos y 4 hidrocarburos (Cuadro 1).
3.2. Respuesta electroantenográfica al extracto y a la mezcla de sintéticos
El extracto de col y la mezcla de compuestos químicos sintéticos provocaron una
despolarización significativamente mayor en las antenas de las hembras apareadas en
comparación con los compuestos químicos evaluados individualmente (F10/131=17.322,

36
p=0.001), aunque la mezcla de compuestos sintéticos provocó mayor despolarización
que el (Z)-3-hexenil acetato, no hubo diferencia significativa entre ellos (Figura 7).
Cuadro 1. Compuestos químicos identificados en CG-EM del extracto de col
Compuesto químico Clase Peso
molecular
Tiempo de
retención
Índice de
Kovats
6-metil-5-hepten-2-ona Cetona 126.104 7.66 693
(Z)-3-Hexenil acetato Éster 142.09 8.145 1004
Limoneno Terpeno 136.125 8.81 1017
Nonanal Aldehído 142.135 11.38 1057
Decanal Aldehído 156.151 14.79 1203
Tetradecano Alcano 198.234 21.20 1400
Pentadecano Alcano 212.25 24.1 1500
Hexadecano Alcano 226.266 27.23 1600
Heptadecano Alcano 240.281 30.03 1700

37
Figura 7. Despolarización (Promedio ± EEM) provocada por el extracto de col en antenas de
hembras apareadas. Letras diferentes indican valores que son significativamente diferentes al p
< 0.05 (Tukey), n = 12.
3.3. Respuesta de atracción en túnel de viento al extracto y a la mezcla de sintéticos
Por un lado, en el patrón de vuelo orientado el extracto de volátiles de col sin
daño provocó mayor porcentaje de atracción en las hembras apareadas que la mezcla de
compuestos químicos sintéticos equivalente, pero esta diferencia no fue significativa.
Por otro lado, hubo diferencia significativa en mostrar el ovipositor entre los dos tipos
de estímulos (χ²= 16.990; gl= 1; p= 0.001) (Figura 8).
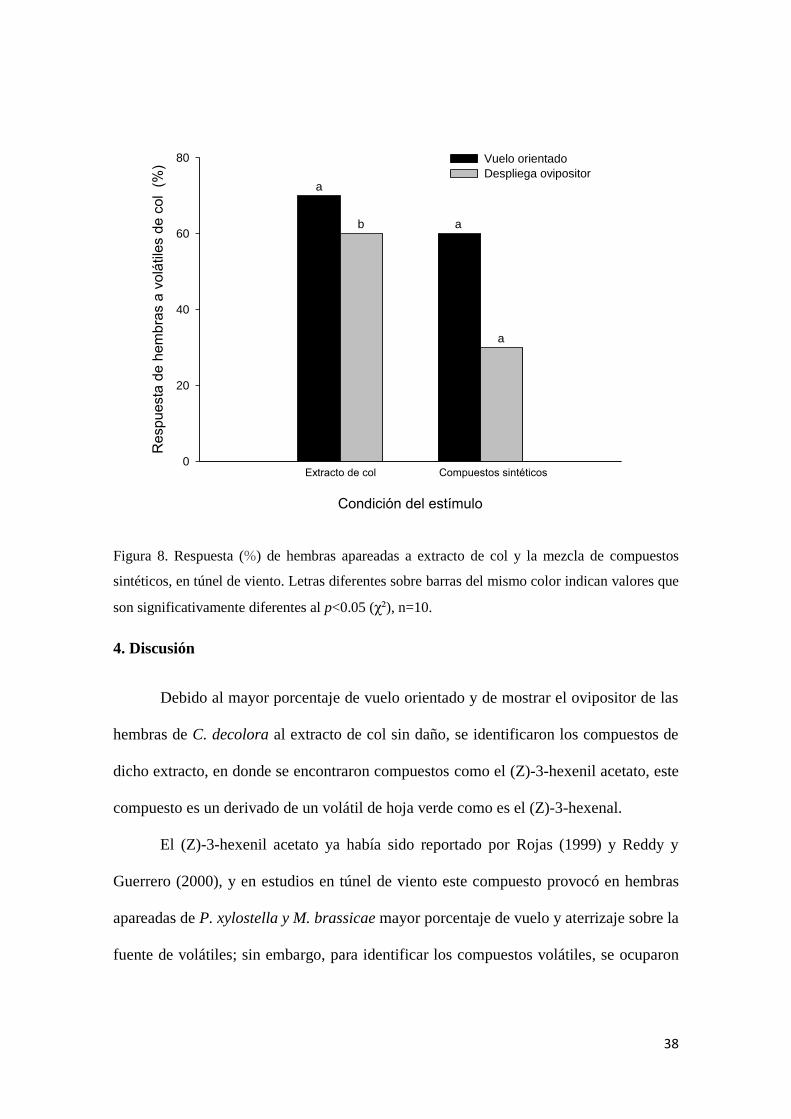
38
Condición del estímulo
Respuesta
de h
em
bra
s a
volá
tile
s d
e c
ol (
%)
0
20
40
60
80 Vuelo orientado
Despliega ovipositora
a
a
b
Extracto de col Compuestos sintéticos
Figura 8. Respuesta (%) de hembras apareadas a extracto de col y la mezcla de compuestos
sintéticos, en túnel de viento. Letras diferentes sobre barras del mismo color indican valores que
son significativamente diferentes al p<0.05 (χ²), n=10.
4. Discusión
Debido al mayor porcentaje de vuelo orientado y de mostrar el ovipositor de las
hembras de C. decolora al extracto de col sin daño, se identificaron los compuestos de
dicho extracto, en donde se encontraron compuestos como el (Z)-3-hexenil acetato, este
compuesto es un derivado de un volátil de hoja verde como es el (Z)-3-hexenal.
El (Z)-3-hexenil acetato ya había sido reportado por Rojas (1999) y Reddy y
Guerrero (2000), y en estudios en túnel de viento este compuesto provocó en hembras
apareadas de P. xylostella y M. brassicae mayor porcentaje de vuelo y aterrizaje sobre la
fuente de volátiles; sin embargo, para identificar los compuestos volátiles, se ocuparon

39
extractos de hojas de col cortadas, a diferencia del extracto de la planta col sin daño,
ocupada en este estudio.
En este trabajo, el (Z)-3-hexenil acetato estuvo presente en los extractos en
plantas sin daño; algunos compuestos volátiles inducidos por daño presentan similitud
estructural con los volátiles liberados cuando no hay daño, lo que indica que sus vías
biosintéticas son semejantes (Geervliet et al., 1997).
En otras brasicáceas como Arabidopsis, el (Z) -3-hexenal se forma segundos
después de ser dañada la hoja, seguido por la formación del (Z)-3-hexenol y el (Z)-3-
hexenil acetato (D’Auria et al., 2007). El producto principal en hojas dañadas de
Arabidopsis es el (Z)-3-hexenal, mientras que el (Z) -3-hexenol y el (Z)-3-hexenil
acetato se producen en hojas parcialmente dañadas (Matsui et al., 2012). Las plantas sin
daño aledañas a las plantas que están siendo atacadas por herbívoros, pueden
incrementar la emisión de volátiles de defensa en respuesta a señales que emiten las
plantas dañadas (Dicke et al., 2003). Por ejemplo, en plantas sin daño de Arabidopsis,
se incrementó la síntesis de (Z)-3-hexenil acetato y (Z)-3-hexenol, cuando estas plantas
fueron expuestas a los volátiles de plantas de Arabidopsis con daño (Ozawa et al.,
2013).
La respuesta antenal y en túnel de viento para el extracto de col fue alta y similar
a la mezcla de compuestos sintéticos elaborada con las proporciones de compuestos
encontradas en el extracto. El (Z)-3-hexenil acetato por sí solo provocó una
despolarización alta pero no mayor al de la mezcla de compuestos. En túnel de viento,
las hembras desplegaron menos el ovipositor al exponerlas a la mezcla de compuestos
que al extracto de col, posiblemente porque en la mezcla no existen compuestos que
estímulen el proceso de oviposición (Dicke y van Loon, 2000).

40
La atracción de palomillas a la mezcla de compuestos en la misma proporción
que se encuentran en los extractos de su planta hospedera, ha sido reportado por
(Rembold et al., 1991; Hartlieb y Rembold, 1996). Se conoce que el reconocimiento de
la planta hospedera por medio del olfato puede deberse a un solo compuesto o bien a
una mezcla específica (Finch y Collier, 2000; Bruce et al., 2005; Tasin et al., 2006;
Riffell et al., 2009), lo que sugiere que los insectos tienen receptores olfativos
específicos para detectar diversos compuestos volátiles de plantas.
Cuando los insectos herbívoros detectan compuestos de plantas hospederas y no
hospederas, la proporción de volátiles emitidos por la planta hospedera (llamada mezcla
especifica) es vital en la señal olfatoria (Bruce et al., 2005). En este caso, la
especificidad dependería de una proporción particular de los compuestos más que de un
solo compuesto (Fraser et al., 2003; Bruce et al., 2005).
Se ha visto en algunas palomillas, que formular atrayentes con algunos
compuestos tiene el mismo efecto de atracción que la mezcla completa (Tasin et al.,
2006b; Cha et al., 2008; Lu et al., 2012). Para C. decolora, en otros experimentos de
túnel de viento, se podrán evaluar mezclas con menor número compuestos volátiles
identificados pero sin disminuir el efecto atractivo para las hembras.

41
IV. Factores que afectan a las hembras en la búsqueda de la planta de col y a los
machos en la búsqueda de pareja
1. Introducción
La búsqueda de la planta hospedera para oviposición o alimentación por parte de
las hembras y la búsqueda de pareja por parte de los machos en las palomillas, está
mediada principal por el olfato. El proceso de búsqueda implica una secuencia
comportamental guiada por estímulos químicos como volátiles de planta hospedera o
feromona sexual (Visser 1986; Roelofs 1995; Proffit et al., 2011). Sin embargo, su
condición fisiológica (edad y estado de apareamiento) será fundamental para realizar los
procesos de búsqueda de planta hospedera y de pareja, pero además, su capacidad de
aprendizaje les permitirá adaptarse a las condiciones ambientales cambiantes (Browne,
1993; Landolt y Molina, 1996; Olsson et al., 2006; Anton et al., 2007; Dukas, 2008;
Dukas et al., 2012).
Las palomillas hembras apareadas son más atraídas a los volátiles de sus plantas
hospederas que cuando son vírgenes. Por ejemplo, hembras apareadas de M. sexta de 3
días de edad son más atraídas a su planta hospedera que hembras vírgenes o apareadas
de otras edades (Mechauber et al., 2002). En otras palomillas, las hembras vírgenes son
atraídas a sus plantas hospederas pero en menor grado que las hembras apareadas
(Pivnick et al., 1990; Rojas, 1999; Ikeura et al., 2010).
En el caso de la atracción del macho por parte de la hembra, en algunas especies
como P. xilostella (Zhang et al., 2009), Agrotis ipsilon (Hufnagel) (Anton y Gadenne,
1999; Gadenne y Anton, 2000; Gadenne et al., 2001; Barrozo et al., 2010a, 2010b;
2011), Heliothis virescens (Fabricius) y Heliothis subflexa (Guenée) (Soques et al.,

42
2010) la edad y el estado de apareamiento pueden modificar el proceso de recepción y
procesamiento olfatorio del macho a la feromona sexual.
En cuanto al aprendizaje en palomillas, se ha documentado que éste cambia la
respuesta a las señales olfativas relacionadas con la atracción sexual (Anderson, 2003;
Anderson et al., 2007), oviposición (Landolt y Molina, 1996; Cunningham et al.,
1998b; Rojas y Wayt, 1999b) y alimentación (Cunningham et al., 1998a).
Algunos de estos estudios mencionados muestran que la pre-exposición a
feromona (Bartell y Lawrence, 1973, 1976, 1977; Bartell y Roelofs, 1973; Figueredo y
Baker, 1992; Daly y Figueredo, 2000) y a volátiles de planta hospedera (Rojas y Wayt,
1999b) provoca una reducción de la respuesta de atracción; pero puede en otros casos,
provocar un aumento en la respuesta a feromona sexual (Anderson, 2003; Anderson et
al., 2007) o a volátiles de planta hospedera para oviposición (Landolt y Molina, 1996;
Cunningham et al., 1998b).
En C. decolora, estudios acerca de factores bióticos que afecten la búsqueda de
la planta hospedera por las hembras o búsqueda de pareja por el macho, son escasos.
Por una parte, se sabe que en pruebas de elección entre varias plantas hospederas, las
hembras eligen plantas de col (Pérez, 2010; Acatitla, 2010). Además, se han realizado
estudios acerca del comportamiento reproductivo, donde las hembras de un intervalo de
edad alcanzan su mayor potencial reproductivo, entendido como fecundidad, fertilidad y
dinámica de oviposición (Rojas y Cibrian-Tovar, 1994; Callado-Galindo et al., 2013).
Por otra parte, aunque ya se han empleado dispositivos en trampas con feromona
sexual sintética como alternativa de manejo al uso de insecticidas sintéticos (Rojas et
al., 2006; Muñíz et al., 2007; Gomez et al., 2012), no se ha estudiado el efecto de los

43
factores ambientales (Dumont y McNeil, 1992) y fisiológicos sobre la respuesta de los
machos a la feromona sexual.
Conocer como la respuesta hacia la feromona o a las plantas hospederas de las
palomillas se ve afectada por el estado fisiológico y la experiencia de hembras y
machos, ayudará a entender el proceso de atracción de los machos a las trampas cebadas
con feromona sexual y decidir sobre la inclusión de volátiles de las plantas en esa
misma trampa diseñada para capturar machos y capturar hembras, lo que generaría una
herramienta de monitoreo o control más potente.
En este capítulo de estudió la edad al apareamiento de las hembras, el efecto de la edad
y la condición de apareamiento de los machos, sobre las respuestas de atracción
provocadas por extractos de col y extractos glandulares de feromona sexual. También,
se estudió el efecto de la pre-exposición de hembras apareadas a los volátiles de plantas
de col y de machos vírgenes al extracto glandular de feromona sexual sobre la respuesta
de atracción a esos estímulos químicos.
2. Materiales y Métodos
2.1. Insectos
Los insectos utilizados en los bioensayos se obtuvieron y mantuvieron como lo
descrito en el Capítulo II, en la Sección 2.1.
2.1.1. Edad al apareamiento en hembras
Para estudiar el efecto de la edad al apareamiento en las hembras, las pupas se
separaron por sexo en jaulas de 20x20x20 cm hasta su emergencia como adultos para su
apareamiento. Se utilizaron 10 adultos por edad: hembras de 2, 4, 6 y 8 días de edad
apareadas y se compararon con hembras vírgenes de sus respectivas edades. Los grupos

44
de hembras apareadas fueron de 2-8 días de edad vírgenes (dv) y apareadas (da)
(Callado-Galindo et al., 2013).
Después del apareamiento, cada hembra se separó individualmente en frascos de
20 cm de altura x 10 cm de diámetro y se registró la hora final de apareamiento para
sincronizar el tiempo y medir su respuesta al extracto de volátiles de col. Para
comprobar si hubo apareamiento, se disectó la bursa copulatrix y se observó la
presencia de espermatóforos.
2.1.2. Edad y estado de apareamiento en los machos
Para estudiar el efecto de la edad en los machos se utilizaron grupos de 2-3, 4-5,
6-7, 8-9 y 10-11 días de edad (n=10, para cada intervalo de edad) y para el efecto del
apareamiento, la respuesta de los machos a los extractos fue medida 24 y 48 horas
después del apareamiento (24 h-dap y 48 h-dap).
2.1.3. Acondicionamiento de los insectos para evaluar el efecto de pre-exposición a
los volátiles de col y a la feromona sexual.
Para este estudio, se utilizaron hembras y machos de 4-6 días de edad, ya que se
observó previamente que su respuesta al extracto de volátiles de col y al extracto
glandular de feromona sexual no cambio con la edad, respectivamente. Una vez
controlada la edad y para que el estado de apareamiento y el sexo no interfiriera en el
efecto del aprendizaje sobre la posterior respuesta a los estímulos olfativos, se evaluaron
machos vírgenes, machos apareados, hembras vírgenes y hembras apareadas al extracto
de volátiles de col y al extracto glandular de feromona sexual (n=10, para cada grupo de
insectos).
Aquellos grupos de insectos que presentaron una atracción significativamente
mayor en vuelo orientado hacia el estímulo (en el caso de las hembras a volátiles de col)

45
o aterrizaje en la fuente de emisión (en el caso de los machos a feromona sexual), fueron
utilizados para los experimentos del efecto de pre-exposición.
2.1.4. Efecto de pre-exposición a los volátiles de planta hospedera y al extracto
glandular de feromona sexual.
Para el efecto de pre-exposición al extracto de volátiles de planta hospedera al
extracto glandular de feromona sexual, se utilizaron los machos y las hembras que
presentaron una respuesta máxima al estímulo de los experimentos de la sección
anterior. Así, las hembras apareadas fueron pre-expuestas a 10 µl del extracto de col y
los machos vírgenes fueron pre-expuestos a 3 (EH) del extracto glandular de feromona
sexual, ambos durante 10s.
Posteriormente, los insectos se evaluaron a corto plazo (2 horas después de la
pre-exposición=2h-DP) y a largo plazo (24 horas después de la pre-exposición= 24h-DP
y 48 horas después de la pre-exposición= 48h-DP). Se hicieron 10 repeticiones para cada
tiempo evaluado. En el análisis de datos del efecto a corto y largo plazo, los insectos pre-
expuestos se compararon con insectos sin pre-exponer (SP) de edades iguales (4-6 días
de edad).
2.2. Plantas
Las plantas de col sin daño se utilizaron para la extracción de volátiles, se
obtuvieron y mantuvieron como lo descrito en el Capítulo II, en la sección 2.2.
2.3. Extracción de compuestos volátiles de la col
Los volátiles de plantas de col sin daño se obtuvieron como lo descrito en el
Capítulo II, en la Sección 2.3.
El análisis para confirmar el perfil químico de los extractos de volátiles de col y
poder utilizarlos en los bioensayos, se realizó de acuerdo a lo descrito en el Capítulo III,

46
en la sección 2.4. En todos los extractos de volátiles de col utilizados en los experimentos, se
verificó que los compuestos químicos aparecieran constantemente.
2.4. Extracción de feromona sexual
La glándula productora de feromona sexual de hembras vírgenes de 4-5 días de
edad, fue disectada de acuerdo a Rojas et al. (2006). Estas glándulas fueron colocadas en
viales de vidrio de 2 ml con un 1 ml de diclorometano como disolvente, durante 10 min.
El sobrenadante se refrigeró a -4ºC hasta su análisis químico o su utilización en los
bioensayos. Cada extracto tenía una concentración de 3 equivalentes hembra (EH) por
cada 5 μl.
2.5. Respuesta de atracción en túnel de viento al extracto
Se utilizó el túnel de viento con las mismas condiciones descritas en el Capítulo
II, en la Sección. Se utilizó como estímulo el extracto de col o el extracto glandular de
feromona sexual. En el acondicionamiento de los machos o las hembras que se pre-
expondrían a los volátiles de col, para las hembras se registró sólo el patrón de
comportamiento de vuelo orientado hacia el estímulo y para los machos cuando
aterrizaban.
En los experimentos de edad al apareamiento y pre-exposición, se registró
cuando la hembra desplegó el ovipositor o presentó un vuelo orientado (vuelo mayor a
los 150 cm en el túnel de viento) hacia el estímulo; además se registró la latencia de
activación (movimiento de antenas y de alas) y la latencia de aterrizaje o vuelo
orientado hasta la fuente de olor.
En el caso de los experimentos de edad, el estado de apareamiento y pre-
exposición se registró cuando el macho desplegó los “claspers”, el aterrizaje sobre el
estímulo, la latencia de activación y aterrizaje.

47
2.6. Análisis estadístico
Los datos de los patrones de comportamiento mostrados por las hembras y los
machos en el túnel de viento fueron analizados con una prueba de χ².
Para las hembras, los datos de latencia de activación y latencia a la fuente de olor
se analizaron por medio de un análisis de varianza unifactorial para medidas repetidas
(ANOVA MR), después de la transformación de datos (lnx) y para las comparaciones
entre grupos se hizo una prueba de Tukey.
Para los machos, los datos de latencia de activación y aterrizaje para edad y
estado de apareamiento, fueron analizados por una prueba de Kruskal-Wallis y la
comparación entre grupos con una prueba de Tukey.
Las pruebas se realizaron con el programa estadístico Sigma Plot 11 (Systat
Software Inc., Chicago, Illinois).
3. Resultados
3.1. Efecto de la edad al apareamiento en hembras
Las hembras apareadas de C. decolora de 4da y 6da fueron más atraídas por el
extracto de volátiles de col en comparación con las otras edades de hembras vírgenes o
apareadas, no se encontró diferencia significativa entre los grupos de edades de 4da y
6da, pero si con 6dv (χ²= 52.769; gl= 1; p= 0.001), con 4dv y 8da (χ²= 31.687; gl= 1; p=
0.001) y con 2da (χ²= 16.990; gl= 1; p= 0.001) (Figura 9).
Para el patrón de comportamiento de desplegar el ovipositor no hubo diferencia
significativa entre los grupos de 4da y 6da, todos los demás grupos de hembras no
desplegaron el ovipositor (Figura 9).
No se encontró diferencia significativa entre los grupos de hembras apareadas
para la latencia de activación y latencia de vuelo orientado.

48
Edad y condición de las hembras
2da 4dv 4da 6dv 6da 8da
Re
sp
ue
sta
a e
xtr
acto
s d
e c
ol (
%)
0
10
20
30
40
50
60
70
Vuelo orientado
Despliega ovipositor
a
ab
b
c c
a
a
ab
a
Figura 9. Respuesta (%) de hembras vírgenes y apareadas a extracto de col, en túnel de viento.
Hembras de días de edad vírgen (dv) y hembra de días de edad apareada (da). Letras diferentes
sobre barras del mismo color indican valores que son significativamente diferentes al p<0.05,
(χ²), n=10.
3.2. Edad y estado de apareamiento en los machos
En el túnel de viento, se observó que la atracción de los machos al extracto fue
mayor a los 4-5 días y después disminuyó conforme incrementó la edad (Figura 10). Los
machos de 4-5 y de 6-7 días de edad no presentaron diferencias significativas, pero si
con los otros grupos en los patrones de comportamiento de aterrizaje para 2-3 y 8-9 días
de edad (χ²= 52.769; gl= 1; p= 0.001) y para 10-11 días de edad (χ²= 72.521; gl= 1; p=

49
0.001) y mostrar “claspers” para 2-3 días de edad (χ²= 20.056; gl= 1; p= 0.001) y para 8-
9 y 10-11 días de edad (χ²= 8.526; gl= 1; p= 0.004) (Figura 10.
Edad de los machos (días)
2-3 4-5 6-7 8-9 10-11
Re
sp
ue
sta
a e
xtr
acto
de
fe
rom
on
a s
exu
al (
%)
0
20
40
60
80
100
Aterrizaje
Despliega claspers
a
a
a
b
b
b
b
a a
a
Figura 10. Respuesta (%) de grupos de machos vírgenes a extracto glandular de feromona
sexual, en túnel de viento. Letras diferentes sobre barras del mismo color indican valores que
son significativamente diferentes al p<0.05 (χ²), n=10.
Por otra parte, los machos de 4-5 días de edad presentaron una latencia de
activación y aterrizaje significativamente menores que aquellos mostrados por los
machos de 2-3, 8-9 y 10-11 días de edad (H= 24.167, H= 28.577, p= 0.001), pero no
con aquellos de 6-7 días (Figura 11).

50
Edad de los machos (días)
Late
ncia
de a
ctivació
n (
s)
0
10
20
30
40
50
Late
ncia
de a
terr
izaje
(s
)
0
100
200
300
8-92-3 4-5 6-7 10-11
bc
a
ab
c
c
a
ab
b bb
Figura 11. Latencia de activación y latencia de aterrizaje (Q1 < Mediana < Q2) por grupo de
edad de machos vírgenes en respuesta al extracto glandular de feromona sexual, en el túnel de
viento. Letras diferentes sobre barras del mismo color indican valores que son
significativamente diferentes al p < 0.05 (Tukey), n = 10
Los machos vírgenes presentaron las mayores respuestas de atracción al extracto
que los machos evaluados 24 y 48 h después de apareados. Se encontraron diferencias en
el comportamiento de aterrizaje para los machos 24 h-dap (χ²= 18.484; gl= 1; p= 0.001)
y para los machos 48 h-dap (χ²= 48.505; gl= 1; p= 0.001) (Figura 12).
Se observó que el apareamiento tiene un efecto tanto en la latencia de activación
y aterrizaje en la fuente de olor; los grupos de 24 h-dap y 48 h-dap presentaron
diferencias significativas respecto al control (H= 14.885, H= 17.867, p= 0.001), pero no
entre ellos. Los machos de los grupos 24 h-dap y 48 h-dap emplearon más tiempo para

51
activarse y aterrizar en la fuente de olor que los del grupo de machos vírgenes (Figura
13).
Condición de los machos
Vírgen 24 h-dap 48 h-dap
Respuesta
a e
xtr
acto
de f
ero
mona s
exual
(%)
0
20
40
60
80
100
Aterrizaje
Despliega claspers
c
b
a a
a
a
Figura 12. Respuesta (%) de machos vírgenes y apareados a extracto glandular de feromona
sexual, en túnel de viento. Los machos apareados fueron evaluados 24 y 48 horas después del
apareamiento (24 h dap y 48 h-dap, respectivamente). Letras diferentes sobre barras del mismo
color indican valores que son significativamente diferentes al p<0.05 (χ²), n=10.
3.3. Efecto de la pre-exposición
3.3.1. Acondicionamiento de los insectos para evaluar el efecto de pre-exposición
En el experimento de condicionamiento para saber cuáles de los grupos de
insectos considerados responden más a los volátiles de col y evaluar el efecto de pre-
exposición, la respuesta máxima de atracción se observó en el patrón de comportamiento

52
vuelo orientado en hembras apareadas, en comparación con las hembras vírgenes (χ²=
16.990; gl= 1; p= 0.001), los machos apareados (χ²= 52.769; gl= 1; p= 0.001) y los
machos vírgenes (χ²= 31.687; gl = 1; p= 0.001) (Figura 14).
Condición de los machos
Late
ncia
de a
ctivació
n (
s)
0
10
20
30
40
50
Late
ncia
de a
terr
izaje
(s)
0
50
100
150
200
250
300
Vírgen 24 h-dap 48 h-dap
a
b
b
a
b b
Figura 13. Latencia de activación y latencia de aterrizaje (Q1 < Mediana < Q2) de machos
vírgenes y apareados en respuesta al extracto glandular de feromona sexual, en el túnel de
viento. Los machos apareados fueron evaluados 42 h y 48 h después del apareamiento (24 h dap
y 48 h-dap, respectivamente). Letras diferentes sobre barras del mismo color indican valores
que son significativamente diferentes al p < 0.05 (Tukey), n = 10.
Para los machos, en el patrón de comportamiento aterrizaje se observó una
respuesta significativamente mayor en los machos vírgenes en comparación con los

53
machos apareados (χ²= 36.214: gl= 1; p= 0.001), con las hembras vírgenes (χ²= 96.182;
gl= 1; p= 0.001) y con las hembras apareadas (χ²= 124.820; gl= 1; p= 0.001) (Figura 15).
Respuesta
a e
xtr
acto
de c
ol
(%)
0
10
20
30
40
50
60
70
a
ab
b
c
Condición de machos y hembras
Macho virgen
Macho apareado
Hembra apareada
Hembra virgen
Figura 14. Respuesta (%) provocada por el extracto de col en machos y hembras en túnel de
viento. Letras diferentes indican valores que son significativamente diferentes al p<0.05 (χ²), n
= 10.
3.3.2. Pre-exposición de hembras apareadas a volátiles de col
Las hembras SP presentaron diferencias significativas en el patrón de
comportamiento vuelo orientado en comparación con las hembras 2h-DP (χ²= 52.769;
gl= 1; p= 0.001) y las hembras 24 h-DP y 48h-DP (χ²= 7.220; gl= 1; p= 0.001). Además,
para el patrón de comportamiento despliega ovipositor, entre las hembras SP y 48h-DP
no mostraron diferencias, pero con las hembras 2h-DP (χ²= 11.281; gl= 1; p=0.001) y
24h-DP (χ²= 7.521; gl= 1; p= 0.006) si hubo diferencia (Figura 16).

54
Macho virgen
Macho apareado
Hembra apareada
Hembra virgen
Respuesta
a e
xtr
acto
de f
ero
mona
sexual (
%)
0
20
40
60
80
100
a
a
b
c
Condición de machos y hembras
Figura 15. Respuesta (%) provocada por el extracto glandular de feromona sexual en machos
y hembras en túnel de viento. Letras diferentes indican valores que son significativamente
diferentes al p<0.05 (χ)², n = 10.
Respecto a la latencia de tiempo de activación, todas las hembras apareadas se
activaron, pero ninguna aterrizó en la fuente de olor. Para el tiempo de activación, se
observaron diferencias significativas en las hembras 2h-DP en comparación con las
hembras SP, las hembras 24h-DP y las hembras 48h-DP (F3/39= 20.573; n= 10; p=
0.001). En el tiempo de arribo a vuelo largo se observaron diferencias significativas en
las hembras SP con las hembras 2h-DP y las hembras 48h-DP (F3/39= 33.687; n= 10; p=

55
0.001). Entre las hembras pre-expuestas hay diferencia significativa en las hembras 2h-
DP con las hembras 24h-DP y 48h-DP (F3/39= 33.687; n= 10; p= 0.001) (Figura 17).
Condición de las hembras
Re
sp
ue
sta
a e
xtr
acto
de
co
l (
%)
0
10
20
30
40
50
60
70 Vuelo orientado
Despliega ovipositor
c
a
b b
b
a
c
b
SP 2h-DP 24h-DP 48h-DP
Figura 16. Respuesta (%) de hembras sin pre-exponer y pre-expuestas al extracto de col
en túnel de viento. A corto plazo (2 horas después de la pre-exposición=2h-DP) y a largo plazo
(24 horas después de la pre-exposición= 24h-DP y 48 horas después de la pre-exposición= 48h-
DP). Letras diferentes sobre barras del mismo color indican valores que son significativamente
diferentes al p<0.05 (χ)², n = 10.
3.3.3. Pre-exposición de machos vírgenes a feromona sexual
Los machos 2h-DP mostraron diferencias significativas respecto a los machos SP (χ²=
72.521; gl= 1; p= 0.001), los machos 24h-DP (χ²= 30.420; gl= 1; p= 0.001) y con los

56
machos 48h-DP (χ²= 16.990; gl= 1; p= 0.001) en el patrón de comportamiento de
aterrizaje. En éste patrón de comportamiento, no se observó diferencias entre los machos
24h-DP y los machos 48h-DP. La mayoría de los machos en diferentes condiciones
evaluados en el túnel de viento mostraron los “claspers” (Figura 18).
Condición de las hembras
La
ten
cia
de
activa
ció
n (
s)
0
10
20
30
40
50
La
ten
cia
a v
ue
lo o
rie
nta
do
(s
)0
50
100
150
200
250
300
350
SP 2h-DP 24h-DP 48h-DP
c
ba
a
a
aba
b
Figura 17. Latencia de activación y latencia de aterrizaje (Promedio ± EEM) de hembras sin
pre-exponer y pre-expuestas al extracto de col en el túnel de viento. A corto plazo (2 horas
después de la pre-exposición=2h-DP) y a largo plazo (24 horas después de la pre-exposición=
24h-DP y 48 horas después de la pre-exposición= 48h-DP). Letras diferentes sobre barras del
mismo color indican valores que son significativamente diferentes al p < 0.05 (Tukey), n = 10.
Para el tiempo de activación, los machos SP mostraron diferencias significativas
con los machos 2h-DP y con los machos 24h-DP (F3/39= 65.722; n= 10; p= 0.001). En

57
los machos pre-expuestos se observó diferencias en 2h-DP con 24h-DP y 48h-DP (F3/39=
65.722; n= 10; p= 0.001), además, los machos 24h-DP presentaron diferencias con los
machos 48 h-DP (F3/39= 65.722; n= 10; p= 0.001). Respecto a la latencia de aterrizaje, se
encontraron diferencias entre los machos 24h-DP con los machos SP, los machos 24h-
DP y los machos 48h-DP (F3/39= 12.349; n= 10; p= 0.001) (Figura 19).
Condición de los machos
SP 2h-DP 24h-DP 48h-DPRespu
esta
a e
xtr
acto
de
fero
mon
a s
exua
l
(%)
0
20
40
60
80
100
120
Aterrizaje
Despliega claspers
c
a
a
a
b
a
b
a
Figura 18. Respuesta (%) de machos sin pre-exponer y pre-expuestos al extracto glandular de
feromona sexual en túnel de viento. A corto plazo (2 horas después de la pre-exposición=2h-DP)
y a largo plazo (24 horas después de la pre-exposición= 24h-DP y 48 horas después de la pre-
exposición= 48h-DP). Letras diferentes sobre barras del mismo color indican valores que son
significativamente diferentes al p<0.05 (χ²), n = 10.

58
b
c
a
b
aa
a
b
Condición de los machos
Late
ncia
de a
ctivació
n (
s)
0
10
20
30
40
Late
ncia
de a
terr
izaje
(s)
0
50
100
150
200
250
300
350
SP 2h-DP 24h-DP 48h-DP
aa
a
b
b
c
a
b
Figura 19. Latencia de activación y latencia de aterrizaje (Promedio ± EEM) de machos sin pre-
exponer y pre-expuestos al extracto glandular de feromona sexual en el túnel de viento. A corto
plazo (2 horas después de la pre-exposición=2h-DP) y a largo plazo (24 horas después de la pre-
exposición= 24h-DP y 48 horas después de la pre-exposición= 48h-DP). Letras diferentes sobre
barras del mismo color indican valores que son significativamente diferentes al p < 0.05
(Tukey), n = 10.
4. Discusión
Los estudios en túnel de viento con las hembras de C. decolora mostraron que la
edad de apareamiento es un factor que afecta la respuesta comportamental a los volátiles
de col, y que los individuos apareados de diferentes edades presentan diferencias en el
comportamiento de atracción. Las hembras vírgenes de C. decolora tuvieron una mínima
respuesta a los volátiles de col como sucede en otras palomillas (Browne, 1993).

59
En C. decolora posiblemente una hembra virgen a la edad de 4 días está enfocada
en la búsqueda de pareja para el apareamiento y no en la búsqueda de la planta
hospedera. Una hembra después del apareamiento, tiene como prioridad la búsqueda de
la planta hospedera para oviposición. Es decir, la hembra realiza movimientos orientados
hacia la planta hospedera para tener contacto con ella aunque no siempre se da en esta
última acción de aterrizaje (Visser, 1986).
En C. decolora las hembras apareadas de 4-6 días de edad, fueron las que
tuvieron una atracción máxima a los volátiles de col, como ocurre en otras palomillas
apareadas con respuesta máxima en otras edades (Ikeura et al., 2010). Por ejemplo en M.
sexta al cuarto día de su emergencia, es la edad en la que se aparean en mayor
proporción y en la que son más atraídas a los volátiles de su planta hospedera
(Mechauber et al., 2002) y para P. xylostella la respuesta se incrementa del día 2 hasta
los 7 días de edad (Pivnick et al., 1990).
La edad en la cual las hembras apareadas de C. decolora alcanzaron la máxima
respuesta de atracción a los volátiles de col, está relacionada con el máximo potencial
reproductivo de la hembra alcanzado a los 4 días de edad, ya que se ha observado que a
otra edad su fecundidad y fertilidad disminuye (Callado-Galindo et al., 2013).
El apareamiento en los insectos debe ocurrir en un intervalo de edad determinado
ya que la fisiología tanto de la hembra como del macho cambia con el tiempo (Jiménez-
Pérez y Wang, 2003), como consecuencia de un retraso en el apareamiento las hembras
reducen su potencial reproductivo, ya que influye sobre la fecundidad y fertilidad de los
huevecillos, como sucede en otras palomillas (Rogers y Marti, 1996; Jiménez-Pérez y
Wang, 2003).

60
La edad de apareamiento, en la que alcanzan la madurez sexual las hembras,
aumenta su adecuación biológica y les permite aumentar su fecundidad y fertilidad, lo
anterior relacionado con la “calidad” de apareamiento que le ofrece el macho (tiempo de
apareamiento, tamaño del espermatóforo y cantidad de espermatóforos transferidos del
macho, tamaño del insecto) (Gillott, 2003).
El apareamiento para la hembra es un cambio fisiológico que influencia
fuertemente la respuesta a los estímulos, como en este caso los volátiles de planta
hospedera. Este cambio sensorial permite a las hembras modular su comportamiento a
una serie de estímulos (Mowrey y Portman, 2012) y responder oportunamente a las
condiciones ambientales presentes.
En C. decolora, los primeros días después de su emergencia, la respuesta de
atracción de los machos es baja e incrementa en relación a la edad hasta alcanzar su
máximo los días 4-5 y 6-7 días, para luego disminuir conforme aumenta la edad. En
algunas otras especies de palomillas, como por ejemplo en los machos de A. ipsilon, la
respuesta a la feromona sexual también incrementó a partir de los 3 días de edad
(Gadenne et al., 1993).
En otras palomillas como Spodoptera frugiperda (Smith) la edad óptima es
alcanzada a los 2 días de edad (Rogers y Marti, 1994), mientras que para Spodoptera
exigua (Hübner) se alcanza entre 1-2 días de edad (Rogers y Marti, 1996). Al parecer,
los machos jóvenes (2-3 días de edad) de C. decolora, no han alcanzado la madurez
sexual para responder a la feromona sexual como sucede en machos recién emergidos de
A. ipsilon (Gadenne et al., 1993; Duportets et al., 1998). En este sentido, la respuesta
olfatoria aumenta conforme los machos alcanzan la madurez sexual y después disminuye
conforme éstos son más longevos (Evenden y Gries, 2008).

61
El apareamiento en los machos de C. decolora está asociado con una
disminución de la respuesta comportamental, como sucede en machos apareados de A.
ípsilon (Gadenne et al., 2001). En C. decolora, esta disminución en la respuesta a la
feromona puede estar relacionada con el hecho de que los machos se aparean una sola
vez por noche (Rojas y Cibrián, 1994) y en total pueden aparearse de 2 a 3 veces de por
vida (Castrejón-Gómez et al., 2000).
Posiblemente los machos de C. decolora necesitan recuperarse del gasto de
energía realizado en el apareamiento y volver a almacenar esperma (Gillott, 2003). En
ese periodo de inhibición los machos pueden enfocarse en la búsqueda de alimento para
restablecer las reservas en sus glándulas accesorias para la producción de
espermatóforos y volver a aparearse, como se ha observado que ocurre en A. ipsilon
(Barrozo et al., 2011).
El efecto de la experiencia previa de las hembras apareadas sometidas 10s al
extracto de col, fue suficiente para que se produjera en éstas un cambio en su
sensibilidad de respuesta. Las hembras apareadas después de una breve exposición a
volátiles de col, tuvieron a las 2h un efecto inhibitorio de respuesta a los mismos,
posiblemente por una saturación de sus receptores, sin llegar a una facilitación o
potenciación de la respuesta olfatoria a las 24h. Se ha documentado que el ciclo
circadiano del insecto puede tener efecto sobre el aprendizaje olfativo (Decker et al.,
2007) y los niveles de hormonas (Bloch et al., 2013), entonces, el que se dé un efecto de
inhibición a las 2h, posiblemente es porque el ciclo circadiano de las hembras apareadas
de C. decolora no coincida con la puesta de huevecillos inmediatamente después de
haberse apareado.

62
Si la atracción a la planta hospedera está relacionada con la carga de huevecillos
(Minkenberg et al., 1992), entonces esto coincide con la restauración de la respuesta de
las hembras apareadas de C. decolora a las 24 y 48h después de haberse pre-expuesto a
volátiles de col, ya que se ha observado que las hembras ovipositan la mayor cantidad de
huevecillos los primeros días después de haberse apareado (Callado-Galindo et al.,
2013). Además, para la latencia de activación y latencia a vuelo orientado se observó
que en las hembras pre-expuestas y evaluadas a las 24 y 48h, se restaura la respuesta
olfatoria al nivel del que muestra las hembras sin pre-exponer.
Por una parte, en estudios con otras palomillas, se ha documentado un efecto
inhibitorio de atracción y aterrizaje en las hembras por pre-exposición a los volátiles de
planta hospedera (Rojas y Wyatt, 1999a). Por otra parte, la pre-exposición a esos
estímulos puede provocar un aumento en la respuesta a volátiles de planta hospedera
para oviposición (Landolt y Molina, 1996; Cunningham et al., 1998b). Para estas
palomillas Rojas y Wyatt (1999a), Landolt y Molina (1996) y Cunningham et al. (1998b)
mencionan que el efecto está relacionado con la habituación o sensibilización de la
respuesta olfatoria.
Dado que la habituación se da por estímulos repetidos, entonces la reducción de
la respuesta a las 2h en las hembras de C. decolora no parece debida a un efecto de
habituación a nivel de sistema nervioso central, sino a una inhibición o posiblemente a
una adaptación sensorial, es decir, a una reducción en la sensibilidad a nivel periférico
(Dukas, 2008). Los mecanismos neurobiológicos de la sensibilidad en la respuesta
olfatoria a volátiles de hospedera han sido explicados muy poco o no han sido
explicados a nivel periférico y central del sistema nervioso (Cunningham et al., 1998b;
Dukas, 2008).

63
Posiblemente, la sensibilidad a volátiles de planta hospedera en el insecto tenga
un mecanismo parecido al que ocurre con los machos a feromona sexual, con un cambio
en las concentraciones de hormonas o neurotransmisores o aminas biogénicas, por
ejemplo, serotonina (Kloppenburg et al., 1999), octopamina (Pophof, 2000) y hormona
juvenil (Anton y Gadenne, 1999).
En cuanto a los machos pre-expuestos al extracto glandular de feromona sexual,
se produjo en éstos un cambio en su sensibilidad de respuesta, aunque la mayoría de los
machos pre-expuestos y sin pre-exponer a feromona sexual mostraron los “claspers”, se
observó en el patrón de aterrizaje una disminución a corto plazo (2h-AP) y un
incrementó a largo plazo (24h-AP y 48h-AP).
Por una parte, una reducción en la respuesta comportamental por efecto de la
experiencia previa a feromona ha sido observado en otras palomillas (Bartell y
Lawrence, 1973, 1976; Bartell y Roelofs, 1973; Kuenen y Baker 1981; Figueredo y
Baker, 1992; Daly y Figueredo, 2000; Judd et al., 2005; Stelinski et al., 2004, 2006a, b;
Trimble et al., 2012) en donde la pre-exposición a la feromona sexual causo un efecto
inhibidor en su respuesta comportamental, tal efecto, está relacionado con la habituación
de la respuesta olfatoria. Sin embargo, una comparación directa con los trabajos
mencionados y éste no puede hacerse, debido a que usaron diferentes tiempos de pre-
exposición y de medición, así como distintas dosis de feromona sexual y repetición de
ésta.
La respuesta a las 2h en los machos de C. decolora posiblemente no fue debida a
un efecto de habituación, ya que ésta se da por estímulos repetidos y en este trabajo no
se hicieron, tal vez exista una reducción en la sensibilidad a nivel periférico, como

64
sucedió en otras palomillas (Rumbo y Vicrers 1997; Stelinski et al., 2005; Trimble y
Marshall, 2010; D´Errico et al., 2013).
Por otra parte, se ha documentado en otras palomillas, un incremento en la
sensibilidad a la feromona sexual (Anderson et al., 2003; Stelinski et al., 2004; 2006b;
Anderson et al., 2007), ya que los machos después de una inhibición a las 2h
incrementaron su respuesta a las 24 h como se observó en el patrón de aterrizaje,
además, para el tiempo de activación se observó que la respuesta se potencia aún más
que en un macho sin pre-exponer.
Asimismo, se observó que en un macho pre-expuesto y medido a las 48 h se
restauró la respuesta olfatoria a nivel de un macho sin pre-exponer. En otras palomillas,
algunos de los mecanismo neurobiológicos de la sensibilidad en la respuesta olfatoria a
feromona han sido explicados a nivel del sistema nervioso periférico y central (Guerrieri
et al., 2012). Es decir, la sensibilidad a feromona cambia por las concentraciones de
hormonas o neurotransmisores o aminas biogénicas, por ejemplo, serotonina
(Kloppenburg et al., 1999), octopamina (Pophof, 2000) y hormona juvenil (Anton y
Gadenne, 1999).
La respuesta de atracción a los volátiles de planta hospedera y a los compuestos
feromonales de C. decolora, no sólo depende de su naturaleza química, hay una
plasticidad neural inducida por el estado fisiológico y la experiencia, que les permite a
las hembras y a los machos modular su respuesta a los volátiles de planta hospedera y a
la feromona sexual.

65
V. Preferencia de oviposición por C. decolora
1. Introducción
El proceso de selección de planta hospedera por el adulto está correlacionado
con la disponibilidad de la planta hospedera para el desarrollo y supervivencia de su
progenie (Thompson y Pellmyr, 1991; Knolhoff y Heckel, 2014). Sin embargo, no
siempre se observa una correlación positiva entre la elección de la planta hospedera para
oviposición por la hembra y el desarrollo larval (Potter et al., 2012), lo que puede variar
en función de variables ecológicas y presiones de selección (Janz, 2008).
La preferencia por una planta hospedera puede estar influenciada por factores
como la experiencia tanto de adultos o las larvas sobre la planta hospedera (Landolt y
Molina, 1996; Cunningham et al., 1998b; Rojas y Wayt 1999b; Papaj y Prokopy, 1989;
Barron, 2001; Akhtar y Isman, 2003; Olsson et al., 2006), la presencia de parasitoides y
depredadores por el daño al comienzo de la oviposición (Meiners y Hilker, 2000; Hilker
y Meiners, 2002; Hilker et al., 2002) y cuando las hembras no son capaces de evaluar la
“calidad” de la planta correctamente (Jaenike, 1978).
Otros factores son la presión de ovipositar por la carga de huevecillos (Jones,
1977; Doak et al., 2006), la diversidad y distribución de vegetación alrededor de la
planta hospedera (Rausher y Papaj, 1983; Petzold, 2009), cuando el desarrollo larval
requiera moverse a otras plantas hospederas (Bar-Yam y Morse, 2011), las larvas por sí
mismas eligen la planta hospedera para su alimentación independientemente del sitio de
oviposición elegido por la hembra (Cunningham et al., 2011; Rivera y Burrack, 2012),
cuando las hembras prefieren ovipositar en plantas libres de enemigos naturales que en
plantas en condiciones “óptimas” para el crecimiento de las larvas (Jeffries y Lawton,
1984).

66
La aceptación o rechazo de la planta hospedera puede estar regulado por la
presencia de estimulantes o disuasivos de la oviposición (Honda, 1995). Los
compuestos estimulantes regularmente no son volátiles (Simmonds, 2001; Calas et al.,
2007), aunque existen ejemplos donde la oviposición es estimulada por compuestos
volátiles (Städler, 1974; Fatzinger y Merkel, 1985) o por una mezcla de ambos
(Renwick et al., 2006).
Por una parte, se menciona que la elección de la planta hospedera la hace la
hembra (Gripenberg et al., 2010; Refsnider y Janzen, 2010; García-Robledo y Horvitz,
2012), es decir, cuando hay una planta hospedera que es apropiada para el desarrollo de
la larvas (Shikano et al., 2010) y también cuando el sitio de oviposición es mal elegido
y hay una alta mortalidad de larvas (Groenteman et al., 2006). En el proceso de
oviposición, la hembra puede modificar su comportamiento reduciendo el número de
huevecillos puestos sobre la planta hospedera, si esta no se encuentra en condiciones
“óptimas” para el desarrollo de su descendencia (Thompson y Pellmyr, 1991; Knolhoff
y Heckel, 2014).
En C. decolora, los estudios acerca de la aceptación de la planta hospedera son
escasos. Se sabe que las hembras ovipositan durante toda su vida, sin embargo a los 4
días de edad alcanzan su mayor potencial reproductivo, reflejado en su fecundidad y
fertilidad (Callado-Galindo et al., 2013). En estudios pre-eliminares de oviposición
sobre plantas de col por hembras de C. decolora se observó que éstas no ovipositan o lo
hacen muy poco sobre la planta. Por tal motivo, en este estudio se investigó en hembras
apareadas de C. decolora, la preferencia de oviposición sobre la planta de col o
alrededor de ésta.

67
2. Materiales y métodos
2.1. Insectos
Los insectos utilizados en los bioensayos se obtuvieron y mantuvieron como lo
descrito en el Capítulo II, en la Sección 2.1.
2.1.1. Preferencia de oviposición
Para aparear a los insectos, 30 machos y 30 hembras de 4 días de edad se
colocaron en una cámara de plexiglás (20x20x20 cm) para que estos se aparearan,
después de lo cual, cada hembra se separó individualmente en frascos de 20 cm de altura
x 10 cm de diámetro hasta su utilización en el bioensayo. Posteriormente, la hembra
apareada fue introducida en la cámara bioclimática con la planta de col sin daño y a las
24 horas después de haber introducido la hembra apareada, se registró el número de
huevecillos puestos en la planta de col o alrededor de ésta. Al final del registro del
número de huevecillos, se verificó que las hembras hubieran sido apareadas, de lo
contrario estas fueron excluidas de los análisis. Se ocuparon 12 hembras, una por cada
repetición.
2.2. Plantas
Las plantas de col sin daño fueron las que se utilizaron para la extracción de
volátiles, se obtuvieron y mantuvieron como lo descrito en el Capítulo II, en la Sección
2.2.
2.3. Determinación de preferencia de oviposición
Para evaluar la preferencia de oviposición sobre la planta de col o alrededor de la
planta, se utilizó una jaula de plexiglás (100x80x60 cm), a 20°C (±2), 60 % HR (± 2). A
cada maceta con la planta de col utilizada en el bioensayo, se le adaptó una placa de
acrílico sujetando el tallo y aislando el suelo. Asimismo, para el registro del número de

68
huevecillos, se marcaron círculos a partir de la base de la maceta cada 5 cm hasta los 30
cm (Figura 20).
Figura 20. Dispositivo para evaluar la preferencia de oviposición por hembras de C. decolora.
Distancias de la planta de col: a) 5 cm, b) 10 cm, c) 15 cm, d) 20 cm, e) 25 cm y f) 30 cm.
2.4. Análisis de datos
Los datos de preferencia de oviposición sobre col y en cada una de las distancias alrededor de
col, fueron analizados por medio de un análisis de varianza unifactorial para medidas repetidas
(ANOVA MR), y para las comparaciones entre grupos se hizo una prueba de Tukey.

69
3. Resultados
3.1. Preferencia de oviposición
Las hembras de C. decolora ovipositan un mayor número de huevecillos
alrededor de la planta de col que sobre ésta. El incremento en el número de huevecillos
puestos fue desde la planta de col hasta los 30 cm. Se observó que sobre la planta de col
hasta los 15 cm a su alrededor no hay diferencia significativa en la cantidad de
huevecillos, pero de los 20 a los 30 cm de distancia de la planta de col la diferencia en el
número de huevecillos es significativa entre ellos y respecto a los encontrados sobre la
col y hasta los 15 cm (F= 185.190; gl= 6,83; n= 12; p= 0.001) (Figura 21).
Distancia de la planta de col (cm)
Planta de col 5 10 15 20 25 30
Nú
me
ro d
e h
ue
ve
cill
os
(Pro
me
dio
± E
EM
)
0
10
20
30
40
50
60
70
a
b
a
aa
d
c
[------------------------------------------------------------]
Figura 21. Número de huevecillos (Promedio ± EEM) sobre plantas de col o alrededor de
ésta. Letras diferentes indican valores significativamente diferentes al p< 0.05 (Tukey), n = 12.

70
4. Discusión
Las hembras de C. decolora ovipositaron mayor cantidad de huevecillos
alrededor de la planta de col, y en menor cantidad sobre la planta.
Algunas palomillas como Cydia pomonella (Linnaeus) ponen sus huevecillos a
20 cm de su planta hospedera y al emerger las larvas son atraídas por el olor de la planta
hospedera, aunque para C. pomonella se ha observado una mayor atracción a su planta
hospedera cuando esta presenta daño por conespecíficos (Landolt et al., 2000).
Debido a que se encontraron huevecillos sobre la planta de col, esto hace pensar
que la hembras de C. decolora en un momento dado aterrizaron sobre la col, en este
sentido,
según la teoría de oviposición “óptima” las hembras al momento de colocar sus
huevecillos, seleccionan aquellas plantas hospederas donde las larvas puedan alcanzar
un máximo desarrollo (Jaenike, 1978). Posiblemente, las hembras de C. decolora no
sean capaces de evaluar la calidad de la planta para poner la totalidad de huevecillos
sobre la planta de col.
Los pocos huevecillos que pusieron las hembras de C. decolora sobre la planta
de col, se encontraron en el centro de la col llamado “cogollo”, posiblemente sea la zona
menos expuesta al ataque de enemigos naturales y la que más consumen las larvas,
evitando lo que pasa con Helicoverpa zea (Boddie), que pone sus huevecillos en una
zona de su planta hospedera con una elevada depredación de los mismos, aunque
necesaria para la nutrición de las larvas (Takeuchi et al., 2009).
La hipótesis de que la hembra siempre elige la planta hospedera en condiciones
“óptimas” para que las larvas alcancen su máximo desarrollo larval, es aplicable en su
mayoría a herbívoros especialistas, mientras que en los herbívoros generalistas como C.

71
decolora, muchas veces no se cumple, ya que estos insectos pueden “equivocarse” al
elegir la planta hospedera. Es cuando las larvas tienen que elegir otra planta hospedera
“óptima” en términos de desarrollo larval. (Jaenike, 1978; Barron, 2009).
La preferencia de elegir una planta hospedera no está determinada sólo por
máximo desarrollo larval, otras variables ecológicas y presiones de selección pueden
estar influenciando la elección de la planta hospedera, por ejemplo el evitar
depredadores y parasitoides.
La distribución de huevecillos alrededor de la planta de col por hembras de C.
decolora hace pensar en la posibilidad elige varias plantas de modo que exista una
distribución de larvas dentro y entre las plantas. De este modo maximiza la dispersión
larval, teniendo un espacio libre de enemigos naturales (Jeffries y Lawton, 1984).
Entre menos larvas haya en una planta, serán insuficientes para estimular la
respuesta defensiva de la planta. Al menos por un tiempo el sitio de oviposición estará
libre de enemigos naturales, con la llegada de más larvas a la planta la emisión de
volátiles se incrementa, y con ello la llegada de parasitoides y depredadores que son
capaces de identificar los volátiles que libera la planta por el daño (Jeffries y Lawton,
1984; Singer et al., 2004).
En el proceso de oviposición los insectos herbívoros, utilizan el estímulo táctil
sobre la planta hospedera para reconocer los estímulantes de la oviposición e influenciar
la elección de hospedera, como ocurre en algunas palomillas (Schoonhoven y van Loon,
2002; Rojas et al., 2003; Lee et al., 2006), aunque existen casos en donde los
estimulantes de la oviposición son volátiles como los isotiocianatos (Jallow et al., 1999;
Renwick et al., 2006). En este sentido, posiblemente C. decolora reconozca algunos
volátiles estímulantes de la oviposición, pero que al inicio de éste proceso la planta de

72
col emite compuestos volátiles que atraen a los depredadores y parasitoides de sus
huevecillos; sin embargo, en el presente estudio no se observó si la hembra empezó a
ovipositar sobre la planta inmediatamente después de colocarla en la jaula.
Un factor fisiológico que afecta la preferencia del adulto al elegir la planta
hospedera es la carga de huevecillos (Javois y Tammaru, 2006). Aquellas hembras que
están muy cargadas de huevecillos regularmente no discriminan entre plantas
hospederas en comparación con las que producen pocos huevecillos que discriminan los
sitios de oviposición (Jones, 1977; Jaenike, 1990; Doak et al., 2006). Por ejemplo, en un
estudio con Pieris rapae (Linnaeus) se encontró que las hembras con un mayor carga de
huevecillos tienden a reducir la longitud de sus vuelos e incrementar sus giros, lo que
lleva a aceptar plantas que deberían ser rechazadas cuando las hembras no tienen tal
carga de huevecillos (Jones, 1977).
Otro factor importante en el proceso de selección de la hospedera y que puede
ser determinante, es el aprendizaje o experiencia previa de los adultos al reconocer los
volátiles de la planta hospedera (Landolt y Molina, 1996; Cunningham et al., 1998b;
Rojas y Wayt, 1999b) o en el caso de las larvas, donde la experiencia de sus larvas al
haberse alimentado de una hospedera puede influir en la preferencia de oviposición de
las hembras de la siguiente generación (Papaj y Prokopy, 1989; Barron, 2001; Akhtar y
Isman, 2003; Olsson et al., 2006). Los insectos que se utilizaron en este estudio se
alimentaron con una dieta para lepidópteros, sería necesario conocer si la dieta contiene
compuestos químicos de la planta de col, para descartar la atracción de las hembras a la
planta de col por el efecto de la experiencia previa de las larvas con el alimento.
En conclusión, las hembras prefieren ovipositar alrededor de la planta de la col,
con un patrón de distribución de número de huevecillos máximo a los 30 cm, ésta

73
distancia concuerda con lo observado en cultivos de plantas de col en campo, ya que las
plantas en un mismo surco están separadas entre ellas a los 25-30 cm. En este trabajo se
observó una disminución gradual de puesta de huevecillos desde los 30 cm hasta la
planta de col.
El entender la dinámica de oviposición por las hembras podría ser de utilidad en
alternativas de manejo, con compuestos químicos empleados para la interrupción del
comportamiento de oviposición.

74
VI. Atracción de larvas de C. decolora a volátiles de la col
1. Introducción
La elección de un sitio adecuado para oviposición es fundamental por parte del
insecto adulto (Bar-Yam y Morse, 2011), pero para las hembras generalistas de
lepidoptera parecen no siempre elegir el sitio de oviposición “óptimo” (Potter et al.,
2012).
Cuando a las hembras no les es fácil elegir una planta hospedera de calidad
nutricional para su descendencia, el distribuir los huevecillos fuera de la planta
hospedera o en una planta no hospedera puede ser una estrategia para maximizar su
adecuación biológica (Hopper, 1999; Zalucki et al., 2002; Diamond y Kingsolver, 2010;
Refsnider y Janzen, 2010). En este caso las larvas necesitan moverse a otras plantas
hospederas para alcanzar su máximo desarrollo (Bar-Yam y Morse, 2011) o cuando las
larvas eligen la planta hospedera para su alimentación independientemente del sitio de
oviposición elegido por la hembra (Cunningham et al., 2011; Rivera y Burrack, 2012)
Se ha documentado que desde el momento en que las larvas participan
activamente en la búsqueda y aceptación de la planta hospedera (Gripenberg et al.,
2010), le quitan presión selectiva de hospedera al adulto (Pöykkö, 2006; Cunningham et
al., 2011; Rivera y Burrack, 2012; Bora et al., 2013). En algunos casos, los estados
larvales responden a los mismos compuestos que los adultos en la búsqueda y
aceptación de su planta hospedera (Rembold et al., 1991; Huang y Renwick, 1994;
Hartlieb y Rembold, 1996; Miles et al., 2005).
En pruebas de doble elección para evaluar la preferencia alimentaria C.
decolora, hacia cinco especies de plantas hospederas, las larvas se desplazaron

75
significativamente más a plantas de col que a las otras plantas evaluadas (Pérez, 2010).
Puede ser que las larvas son las que se desplacen hasta la planta hospedera para su
alimentación, posiblemente guiadas por los mismos compuestos volátiles que atraen a la
hembra adulta. En este sentido, se evaluó la atracción de las larvas neonatas de C.
decolora a la planta de col completa y a extractos de compuestos volátiles de col.
2. Materiales y métodos
2.1. Insectos
Los huevecillos y las larvas neonatas utilizados en los bioensayos se obtuvieron
y mantuvieron como lo descrito en el Capítulo II, en la Sección 2.1.
2.1.1. Respuesta de larvas a plantas de col
Hembras apareadas de C. decolora se colocaron en jaulas de plexiglás
(20x20x20 cm)) cubriendo la entrada con una malla de tela, después de que las hembras
ovipositaron sobre la malla, esta se cortó en secciones donde hubiera 10 huevecillos. En
cada bioensayo, se utilizaron 50 huevecillos de 5 días después de la oviposición en la
última fase de su desarrollo embrionario (color negro característico de la última fase),
estos se distribuyeron en grupos de 10 huevecillos a 30 cm de la base de la maceta con
la planta de col (n=12 repeticiones, cada repetición con 50 huevecillos).
El registro de número de larvas neonatas presentes en la planta de col o
alrededor de ésta se realizó a las 2, 6, 12, 18 y 24 horas después de haber colocado los
huevecillos en la cámara bioclimática, entre cada registro se retiraron las larvas
contadas. Después de cada prueba se contó el número de larvas, se cambió la planta de
col para que no interfiriera el efecto del daño por larva en la planta. Todos los registros
fueron con plantas de col intactas.

76
2.2. Plantas
Las plantas de col sin daño se obtuvieron y mantuvieron como lo descrito en el
Capítulo II, en la Sección 2.2.
Para los bioensayos de presencia de larvas se sincronizó el fotoperíodo invertido
para las larvas y las plantas de col, con la finalidad de realizar los experimentos durante
el día.
2.3. Determinación de la respuesta de larvas en plantas de col
Para el registro del número de larvas sobre y alrededor de la planta de col, se
utilizó la cámara descrita en el Capítulo V, en la Sección 2.3.
2.4. Extracción e identificación de volátiles de col
Se utilizaron plantas de col sin daño, se utilizó el sistema de extracción de
volátiles descrito en el Capítulo II, en la Sección 2.3. Los compuestos capturados en
Super Q, se extrajeron con hexano, diclorometano o metanol.
El análisis para confirmar el perfil químico de los extractos de volátiles de col y
poder utilizarlos en los bioensayos en olfatómetro en Y, se realizó de acuerdo a lo
descrito en el Capítulo III, en la Sección 2.4. En todos los extractos de volátiles de col
utilizados en los experimentos, se verificó que los compuestos químicos atractivos
aparecieran constantemente.
2.5. Respuesta de larvas a extractos de col
La respuesta de las larvas neonatas de C. decolora a los diferentes extractos de
col se evaluó en pruebas de una elección en un olfatómetro de vidrio tipo Y (Figura 22),
similar al utilizado por Vander Meer et al. (1988), donde se comparó la atracción de las
larvas por el extracto (extracto de col con hexano, diclorometano o metanol) contra un
control (aire), además, se comparó el extracto más atractivo para las larvas contra la

77
mezcla de compuestos sintéticos atractiva para las hembras (compuestos sintéticos
identificados en el Capítulo III, en la Sección 2.6).
En cada cámara del olfatómetro Y, se pasó una corriente de aire con un flujo de
200 mL/min, medido por unos flujómetros (Cole Palmer, Ev-03217-06, USA). Para cada
bioensayo, en el tubo principal del olfatómetro, se introdujo una larva neonata (al menos
dos horas de edad después de la emergencia), y se observó su comportamiento durante
300 s. Durante este tiempo, se consideró una elección de la larva, cuando esta alcanzó
alguna de las cámaras, con la fuente de olor o el control.
Para cada bioensayo, en un papel filtro Waltman No.1 (Whatman ® 2V,
England) de 1 cm de largo por 0.5 cm de ancho, se colocaron 10 µL de cada uno de los
extractos de col o la mezcla de compuestos. Todos los bioensayos se realizaron en el
primer tercio de la escotofase a 20°C (±3), 60 % (± 5) de HR y luz roja (40 watts).
Después de cada prueba se invirtió el olfatómetro para eliminar cualquier sesgo.
2.6. Análisis de datos
Los datos de presencia de larvas, sobre col y en cada una de las distancias
alrededor de col, fueron analizados por medio de un análisis de varianza unifactorial
para medidas repetidas (ANOVA MR), y para las comparaciones entre grupos se hizo
una prueba de Tukey.
Los datos de las respuestas de una elección de las larvas neonatas, se analizaron
con una prueba de χ². Las pruebas se realizaron con el programa estadístico Sigma Plot
11 (Systat Software Inc., Chicago, Illinois).

78
Figura 22. Representación esquemática de los componentes del sistema con olfatómetro en Y
para evaluar la atracción de larvas neonatas a diferentes extractos de col y a la mezcla de
compuestos sintéticos.
3. Resultados
3.1. Respuesta de larvas a plantas de col
Los registros del número de larvas presentes sobre la planta de col o alrededor de
ésta, se realizaron a partir de las 12 h, ya que hasta ese tiempo se detectó la presencia de
larvas. A diferencia de la puesta de huevecillos, existe una tendencia del incremento de
número de larvas presentes desde los 30 cm hasta la planta de col. Se observó en los
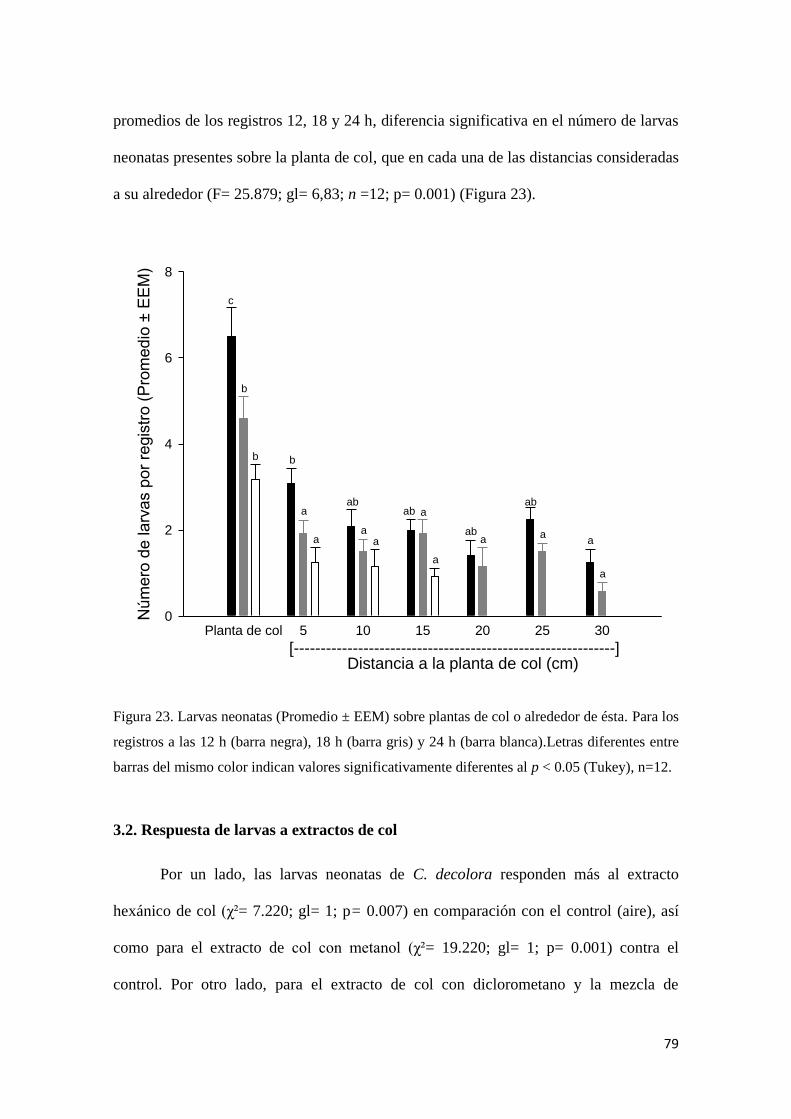
79
promedios de los registros 12, 18 y 24 h, diferencia significativa en el número de larvas
neonatas presentes sobre la planta de col, que en cada una de las distancias consideradas
a su alrededor (F= 25.879; gl= 6,83; n =12; p= 0.001) (Figura 23).
Distancia a la planta de col (cm)
Planta de col 5 10 15 20 25 30
Nú
me
ro d
e la
rva
s p
or
regis
tro
(P
rom
ed
io ±
EE
M)
0
2
4
6
8
[------------------------------------------------------------]
ab
aa
a
aab
b
b
a
aab
a
ab
aa
a
b
c
Figura 23. Larvas neonatas (Promedio ± EEM) sobre plantas de col o alrededor de ésta. Para los
registros a las 12 h (barra negra), 18 h (barra gris) y 24 h (barra blanca).Letras diferentes entre
barras del mismo color indican valores significativamente diferentes al p < 0.05 (Tukey), n=12.
3.2. Respuesta de larvas a extractos de col
Por un lado, las larvas neonatas de C. decolora responden más al extracto
hexánico de col (χ²= 7.220; gl= 1; p= 0.007) en comparación con el control (aire), así
como para el extracto de col con metanol (χ²= 19.220; gl= 1; p= 0.001) contra el
control. Por otro lado, para el extracto de col con diclorometano y la mezcla de

80
compuestos sintéticos atractiva para las hembras no tuvieron deferencia significativa en
comparación con el control (Figura 24).
Figura 24. Respuesta de larvas neonatas a extractos volátiles de col con diferentes disolventes y
a la mezcla de sintéticos, en olfatómetro Y. Estadísticamente significativo * * (p < 0.007) * (p <
0.001), no estadísticamente significativo (ns), no responde (NR), χ², n=15.
4. Discusión
En este estudio, la mayoría de las larvas que emergieron en la periferia migraron
hacia la planta de col. Posiblemente si se hubiera hecho un registro de la presencia de
larvas a las 36 h, todas las larvas contadas estarían sobre la col, ya que en el registro del
número de larvas a las 24 h no se observaron larvas en las distancias de 20 a 30 cm. En
ocasiones las larvas emergidas en la periferia de la planta hospedera, no se desplazan
-100 -60 -20 20 60 100
% de elección
Extracto metanólico
Extracto diclorometanólico
Extracto hexánico
ns
ns
**
100 60 20
Estímulo Control (Aire) % de NR
Mezcla de compuestos
*
13.33 33.33 6.66 13.33

81
hacia la planta hospedera, como sucede en las larvas de Phthorimaea operculella (Zell),
donde sólo el 50 % localizan un sitio adecuado de alimentación (Varela, 1988).
Cuando las larvas participan activamente en la búsqueda y aceptación de la
hospedera, le quitan presión selectiva de hospedera al adulto (Gripenberg et al., 2010),
presentando una alta movilidad dentro o entre las plantas para elegir el sitio adecuado
para su alimentación (Pöykkö, 2006; Cunningham et al., 2011; Rivera y Burrack, 2012).
Parece ser que las larvas de C. decolora participan activamente en la búsqueda y
aceptación de la planta hospedera, la hipótesis de que el adulto hembra “sabe” más al
momento de elegir una planta hospedera en condiciones “óptimas”, puede no cumplirse
en C. decolora. Sin embargo, puede ser que la elección de la planta hospedera por el
adulto, no está determinada sólo por el futuro desarrollo de la larva, sino que existan
otras variables ecológicas y presiones de selección, como la presencia de depredadores
y parasitoides (Paré y Tumlinson, 1999; Potter et al., 2012).
Se sabe que los volátiles inducidos por el ataque de herbívoros pueden ser
atractivos para las larvas de conespecíficos u otra especie, entonces, las larvas además
de vencer las defensas de las plantas tendrán la competencia por el alimento (Landolt et
al., 2000; Carroll et al., 2006, 2008).
Dada la alta movilidad hacia la planta de col por las larvas neonatas de C.
decolora, posiblemente detecten los mismos compuestos que son atractivos para la
hembra. Así lo demuestran estudios con los estados inmaduros de H. armígera, C.
pomonella y P. rapae, las cuales son atraídas a los mismos compuestos a los que
responden las hembras (Rembold et al., 1991; Huang y Renwick, 1994; Hartlieb y
Rembold, 1996; Miles et al., 2005), puede ser que las larvas tengan neuronas olfativas

82
que responden a diferentes olores, sin estar especializadas a moléculas específicas
(Rharrabe et al., 2014).
En las pruebas de una elección, el extracto hexánico de col provocó más
atracción en larvas neonatas de C. decolora. Un estudio similar pero con plantas
dañadas, se realizó con larvas de H. armígera, donde los extractos hexánicos de varias
plantas hospederas provocaron en las larvas mayor atracción en comparación con los
extractos metánolicos (Singh y Mullick, 2002).
Posiblemente, el realizar experimentos donde se evalué la respuesta de las larvas
neonatas a extractos con daño en las plantas u otro tipo de extracción como lavado de
hojas o macerado, se pueda identificar los compuestos que detectan las larvas cuando
tocan la hoja.
En algunos trabajos, han identificado los compuestos involucrados en la
atracción de las larvas neonatas a los volátiles de su planta hospedera, como en C.
pomonella, el compuesto (E,E)-α-farneseno de frutos dañados es atrayente de larvas
(Landolt et al., 2000).
Cuando las larvas disponen de opciones de planta hospedera y se desplazan a la
más adecuada para su alimentación, existen factores que afectan la mortalidad de las
larvas como el microclima, y los riesgos de depredación y parasitismo. Además, en
condiciones naturales la respuesta única a un determinado estímulo en particular,
difícilmente conducirá al insecto en la búsqueda de alimento o sitio de oviposición, sino
que es muy probable que deban estar en combinación con otra serie de estímulos
involucrados en estos procesos, tales como los estímulos visuales y táctiles (Harris y
Foster, 1995; Harris et al., 1995).

83
Este tipo de estudios, permite conocer la dinámica del comportamiento y
supervivencia de las larvas para aumentar su adecuación biológica. En este sentido, una
interrupción de la orientación larval, es decir, atracción de las larvas a trampas, sería
posible como estrategia de manejo para este insecto plaga (Cook et al., 2007).

84
VII. Discusión general
1.- Atracción de hembras de C. decolora a volátiles de col sin daño y con daño
En este estudio, el extracto de col sin daño fue el que provocó la máxima
atracción en las palomillas en comparación con los extractos de plantas con daño.
Después de conocer el extracto que provocó mayor atracción en las hembras apareadas
de C. decolora, se identificaron los compuestos de dicho extracto. Entre los compuestos
identificados, se encontró el (Z)-3-hexenil acetato, compuesto que individualmente
provocó la máxima despolarización antenal, y que en otras palomillas provocó un alto
porcentaje de vuelo y aterrizaje sobre la fuente de volátiles (Rojas, 1999; Reddy y
Guerrero, 2000).
El (Z)-3-hexenil acetato ha sido identificado de extractos de plantas en donde el
método de extracción implica el corte de hojas (Rojas, 1999; Reddy y Guerrero, 2000),
lo cual puede interpretarse como daño en la planta. Algunos compuestos volátiles
inducidos por daño presentan similitud estructural con los volátiles liberados cuando no
hay daño, lo que indica que sus vías biosintéticas son semejantes (Geervliet et al.,
1997).
Los compuestos identificados del extracto de col sin daño y evaluados en
mezcla, provocaron en las hembras un porcentaje de vuelo orientado al estímulo similar
al que tuvieron con el extracto ya mencionado. Tal reconocimiento puede deberse a un
solo compuesto presente en la mezcla o la mezcla completa (Finch y Collier, 2000).
Probablemente el evaluar individualmente cada compuesto identificado en túnel de
viento, complementaría este estudio, ya que posiblemente el (Z)-3-hexenil acetato por sí
solo, provoque las máximas respuestas en los vuelos orientados, como ocurre en P.
xylostella y M. brassicae.

85
También, se pueden identificar los compuestos del extracto de col con daño para
conocer los compuestos responsables de la disminución de la atracción y evaluar
posibles respuestas de repelencia. La repelencia de herbívoros y la atracción de
enemigos naturales a plantas dañadas tienen aplicación directa en el campo del control
de insectos plagas de cultivos agrícolas.
2. Efecto del estado fisiológico y la experiencia sobre la respuesta a los volátiles de
planta hospedera y feromona sexual de C. decolora
En insectos noctuidos el estudio del estímulo químico es importante en la
búsqueda y aceptación de la planta hospedera por parte de la hembra y en la búsqueda de
pareja por el macho; sin embargo, estudios sobre la biología del insecto plaga permitirá
diseñar estrategias de manejo adecuadas con las características de cada especie.
Las hembras de C. decolora de 4-6 días de edad apareadas tuvieron una respuesta
máxima los volátiles de col. Este intervalo de edad está relacionado con el máximo
potencial reproductivo de la hembra, según lo observado por Callado-Galindo et al.
(2013), ya que a otra edad su fecundidad y fertilidad disminuye. La edad adecuada de
apareamiento en las hembras aumenta su adecuación biológica y les permite un mayor
éxito en su fecundidad y fertilidad, como sucede en otras palomillas (Rogers y Marti,
1996; Jiménez-Pérez y Wang, 2003)
Respecto a los machos de C. decolora, a los de 4-7 días de edad alcanzaron la
máxima atracción al extracto glandular de feromona sexual, la respuesta olfatoria
aumenta conforme los machos alcanzan la madurez sexual y después disminuye
conforme éstos son más longevos (Evenden y Gries, 2008). Asimismo, la respuesta de
atracción disminuyó después de que los machos se aparearon. Posiblemente necesitan
recuperarse del gasto de energía realizado en el apareamiento y volver a almacenar

86
esperma (Gillott, 2003). En ese periodo de inhibición los machos pueden enfocarse en la
búsqueda de alimento para restablecer las reservas en sus glándulas accesorias para la
producción de espermatóforos y volver a aparearse, como se ha observado que ocurre en
A. ipsilon (Barrozo et al., 2011).
Otro factor importante que se estudió en este trabajo, fue la experiencia previa de
hembras apareadas a volátiles de planta hospedera como de machos a la feromona
sexual.
Una breve exposición de las hembras apareadas a volátiles de col y de los
machos vírgenes a feromona sexual, provocó a las 2h un efecto inhibitorio de la
respuesta olfatoria posiblemente por una saturación de sus receptores o adaptación
sensorial, es decir una reducción de la sensibilidad a nivel periférico, como sucede en
otras palomillas (Rumbo y Vicrers 1997; Cunningham et al., 1998b; Stelinski et al.,
2005; Dukas, 2008; Trimble y Marshall, 2010; D´Errico et al., 2013).
Las hembras después de que se aparean buscan un sitio para oviposición,
entonces, el que se dé un efecto de inhibición a las 2h, posiblemente es porque el ciclo
circadiano de las hembras apareadas de C. decolora no coincide con la puesta de
huevecillos inmediatamente después de haberse apareado (Decker et al., 2007).
Posiblemente, la restauración de la respuesta olfatoria en las hembras, está relacionada a
la presión de carga de huevecillos, ya que se ha reportado que las hembras ovipositan la
mayor cantidad de huevecillos los primeros días después de haberse apareado (Callado-
Galindo et al., 2013), esto coincide la restauración de la respuesta olfatoria a las 24 y 48
horas.
Para C. decolora, probablemente en otros estudios de pre-exposición a volátiles
de planta hospedera en las hembras y de feromona sexual en los machos, en donde se

87
incluyan diferentes tiempos de pre-exposición, dosis de estímulo y tiempos de medición
de la respuesta olfatoria después de haber pre-expuestos a los insectos, se pueda conocer
si la inhibición se mantiene y hay un proceso de habituación, como sucede en otras
palomillas (Rojas y Wyatt, 1999b; Landolt y Molina, 1996; Cunningham et al., 1998b).
C. decolora como muchos otros insectos, tiene una plasticidad neural, inducida
por la experiencia que les permite a las hembras y a los machos modular su respuesta a
los volátiles de la planta hospedera y a la feromona sexual (Anton et al., 2007).
3.- Preferencia de oviposición por C. decolora
El que las hembras ovipositaran muy pocos huevecillos sobre la planta de col,
posiblemente sea porque ellas no son capaces de evaluar la calidad de la planta para
poner la totalidad de huevecillos sobre la planta de col (Jaenike, 1978). Es entonces
cuando las larvas tienen que elegir otra planta hospedera “óptima” en términos de
desarrollo larval. (Jaenike, 1978; Barron, 2009). Sin embargo, la preferencia de elegir
una planta hospedera no siempre está determinada por el futuro desarrollo larval, otras
variables ecológicas y presiones de selección pueden estar influenciando la elección de
la planta hospedera, por ejemplo el evitar depredadores y parasitoides.
Posiblemente en el campo C. decolora distribuya sus huevecillos en varias
plantas para maximizar la dispersión larval, teniendo un espacio libre de enemigos
naturales (Jeffries y Lawton, 1984) al menos por un tiempo, mientras la planta
hospedera libera los compuestos volátiles que atraen a los enemigos naturales del
herbívoro.
En otras palomillas, la puesta de huevecillos sobre las plantas, provocó la
liberación de volátiles atrayentes de parasitoides (Meiners y Hilker, 2000; Hilker y
Meiners, 2002; Hilker et al., 2002) En este sentido, posiblemente C. decolora reconozca

88
algunos volátiles estimulantes de la oviposición, pero al inicio de éste proceso la planta
de col emite compuestos volátiles que atraerán a los depredadores y parasitoides de sus
huevecillos; sin embargo, en el presente estudio no se observó si la hembra empezó a
ovipositar sobre la planta inmediatamente después de colocarla en la jaula.
Como ya se mencionó anteriormente, C. decolora produce una gran cantidad de
huevecillos durante toda su vida (Callado-Galindo et al., 2013), debido a esto,
posiblemente las hembras de C. decolora no discriminen entre plantas hospederas para
oviposición, como sucede con las hembras que producen pocos huevecillos y que
discriminan los sitios de oviposición (Jones, 1977; Jaenike, 1990; Doak et al., 2006).
4. Atracción de larvas de C. decolora a volátiles de la col
En las larvas neonatas de C. decolora se observó una alta movilidad hacia la
planta de col, parece ser que ellas detectan los mismos compuestos que son atractivos
para el adulto. En estudios con los estados larvales de otras palomillas, se ha observado
que son atraídas a los mismos compuestos a los que responden las hembras (Rembold et
al., 1991; Huang y Renwick, 1994; Hartlieb y Rembold, 1996; Miles et al., 2005).
Posiblemente, el realizar experimentos donde se evalué la respuesta de las larvas
neonatas a extractos con daño en las plantas u otro tipo de extracción como lavado de
hojas o macerado, se pueda identificar ese tipo de compuestos que detectan las larvas
cuando tocan la hoja.
En C. decolora, dada la alta movilidad de las larvas, probablemente sean ellas
quienes elijan la planta hospedera, quitándole presión selectiva de hospedera al adulto
(Gripenberg et al., 2010). Será necesario estudiar el comportamiento de larvas de
diferentes estadios para ver qué tipos de estímulos usan para elegir la planta hospedera.
Ya que es probable que deban estar en combinación con otra serie de estímulos

89
involucrados en estos procesos, tales como los estímulos visuales y táctiles (Harris y
Foster, 1995; Harris et al., 1995). Además, habría que estudiar la posibilidad de que las
larvas no permanecen alimentándose en la misma planta sino que se desplacen en busca
de otras plantas hospederas (Bernays et al., 2004).
De acuerdo a la hipótesis de que la hembra “reconoce”, al momento de elegir
una planta hospedera, que tiene las condiciones “óptimas” para el desarrollo larval. Para
C. decolora tal vez ni la hembra ni la larva “reconocen” esas condiciones al momento
de elegir la planta hospedera; se podría pensar que el adulto y la larva participan en el
proceso de elección adecuándose a la distribución y abundancia relativa de las
potenciales hospederas para maximizar su adecuación biológica, y que la selección final
de la planta hospedera está en función de otras variables ecológicas y presiones de
selección.
La información básica de este trabajo contribuye al conocimiento del estímulo
químico olfativo que causa la atracción de las hembras de C. decolora, la influencia que
tienen algunos factores en la atracción a la planta hospedera por las hembras, y de
búsqueda de pareja por los machos. Además, el estudio de la aceptación de la planta
hospedera por adultos y larvas neonatas.
Los estudios acerca del uso de volátiles de planta hospedera, de feromona sexual
y de la biología de la especie en relación a la respuesta a esos volátiles, deben incluir a
los estados inmaduros, formulaciones con los compuestos sintéticos y los dispositivos
de liberación a emplearse, a fin de proporcionar alternativas que puede integrarse en el
manejo de insectos plaga (Cook et al., 2007). Tales alternativas de manejo pueden
incluir a la feromona sexual y volátiles de planta hospedera, para capturar ambos sexos

90
en trampas en el monitoreo de poblaciones o para trampeo masivo, en combinación con
insecticidas o como atrayentes de depredadores y parasitoides.

91
Conclusiones
El extracto de col sin daño provocó la respuesta máxima de atracción en las
hembras apareadas.
Se identificaron 9 compuestos del extracto de col: una cetona, un éster, un
terpeno, 2 aldehídos y 4 hidrocarburos, de los cuales el (Z)-3-hexenil acetato fue el que
provocó la máxima despolarización antenal. La mezcla de compuestos en proporción
similar a lo encontrado en el extracto, provocó igual porcentaje de atracción en las
hembras.
Las hembras apareadas de 4-6 días de edad, presentaron la máxima respuesta de
atracción al extracto de compuestos volátiles de col y una breve pre-exposición de las
hembras a volátiles de col, causó un efecto de inhibición en la respuesta olfatoria.
Los machos vírgenes de 4-7 días de edad, presentaron la máxima respuesta de
atracción al extracto glandular de feromona sexual y una breve pre-exposición de los
machos a feromona sexual, causó un efecto de inhibición a corto plazo (2h) y una
sensibilización a largo plazo (24h y 48 h).
Las hembras prefieren ovipositar alrededor de la planta de la col, con un patrón
de distribución de número de huevecillos máximo a los 30 cm y de ahí una disminución
gradual de huevecillos hasta la planta de col.
Las larvas neonatas al eclosionar a diferentes distancias de la planta de col, se
desplazan hacia ella, participando en la aceptación de la hospedera para su alimentación.
El extracto hexánico de col sin daño provocó la máxima atracción en las larvas
neonatas, y un aproximado del 50% de las larvas evaluadas a la mezcla de compuestos
identificados se dirigieron a dicho estímulo.

92
Bibliografía
Acatitla, T. C. 2010. Copitarsia decolora Guenée: Su preferencia por brócoli, col y
coliflor, su caracterización molecular y de tres de sus himenópteros parasitoides.
Tesis (Doctor en Ciencias, especialista en Entomología y Acarología). Colegio de
Postgraduados.
Agelopoulos, N. G., y J. A. Pickett. 1998. Headspace analysis in chemical ecology:
effects of different sampling methods on ratios of volatile compounds present in
headspace samples. J. Chem. Ecol. 24: 1161-1172.
Akhtar, Y., y M. Isman. 2003. Larval exposure to oviposition deterrents alters
subsequent oviposition behavior in generalist, Trichoplusia ni and specialist,
Plutella xylostella moths. J. Chem. Ecol. 29: 1853-1870.
Allmann, S., A. Späthe, S. Bisch-Knaden, M. Kallenbach, A. Reinecke, S. Sachse, I. T.
Baldwin, B. S. Hansson, y M. Dicke. 2013. Feeding-induced rearrangement of
green leaf volatiles reduces moth oviposition. eLife 2.
Anderson, P., M. M. Sadek, y B. S. Hansson. 2003. Pre-exposure modulates attraction
to sex pheromone in a moth. Chem. Senses 28: 285-291.
Anderson, P., B. S. Hansson, U. Nilsson, Q. Han, M. Sjoholm, N. Skals, y S. Anton.
2007. Increased behavioral and neuronal sensitivity to sex pheromone after brief
odor experience in a moth. Chem. Senses 32: 483-91.
Anton, S., y C. Gadenne. 1999. Effect of juvenile hormone on the central nervous
processing of sex pheromone in an insect. Proc. Natl. Acad. Sci. U.S.A. 96:
5764-7.
Anton, S., M. C. Dufour, y C. Gadenne. 2007. Plasticity of olfactory-guided behaviour
and its neurobiological basis: lessons from moths and locusts. Entomol. Exp.
App. 123: 1-11.
Arimura, G., K. Matsui, y J. Takabayashi. 2009. Chemical and molecular ecology of
herbivore-induced plant volatiles: proximate factors and their ultimate functions.
Plant Cell Physiol. 50: 911-23.
Awmack, C. S., y S. R. Leather. 2002. Host plant quality and fecundity in herbivorous
insects. Annu. Rev. Entomol. 47: 817-844.
Baldwin, I. T. 2010. Plant volatiles. Curr. Biol. 20: 392-397.

93
Bar-Yam, S., y D. H. Morse. 2011. Host-plant choice behavior at multiple life-cycle
stages: the roles of mobility and early growth in decision-making. Ethology 117:
508-519.
Bartell, R. J., y L. A. Lawrence. 1973. Reduction in responsiveness of males of
Epiphyas postvittana (Lepidoptera) to sex pheromone following previous brief
pheromonal exposure. J. Insect. Physiol. 19: 845-855.
Bartell, R. J., y W. L. Roelofs. 1973. Inhibition of sexual response in males of the moth
Argyrotaenia velutinana by brief exposures to synthetic pheromone or its
geometrical isomer. J. Insect. Physiol. 19: 655-661.
Bartell, R. J., y L. A. Lawrence. 1976. Reduction in sexual responsiveness of male
light-brown apple moth following previous brief pheromonal exposure is
concentration dependent. Aust. J. Entomol. 15: 236-236.
Bartell, R. J., y L. A. Lawrence. 1977. Reduction in responsiveness of male light-brown
apple moths, Epiphyas postvittana, to sex pheromone following pulsed. Physiol.
Entomol. 2: 89-95.
Barrera, J., P. Montoya, y J. Rojas. 2006. Bases para la aplicación de trampas y
atrayentes. Simposio sobre trampas y atrayentes en detección, monitoreo y
control de plagas de importancia económica. J. F Barrera & P. Montoya (eds.).
Sociedad Mexicana de Entomología y el Colegio de la Frontera Sur, Manzanillo,
Colima, México pp: 1-16.
Barron, A. 2001. The Life and Death of Hopkins' Host-Selection Principle. J. Insect
Behav. 14: 725-737.
Barrozo, R. B., D. Jarriault, X. Simeone, C. Gaertner, C. Gadenne, y S. Anton. 2010a.
Mating-induced transient inhibition of responses to sex pheromone in a male
moth is not mediated by octopamine or serotonin. J. Exp. Biol. 213: 1100-1106.
Barrozo, R. B., C. Gadenne, y S. Anton. 2010b. Switching attraction to inhibition:
Mating-induced reversed role of sex pheromone in an insect. J. Exp. Biol. 213:
2933-2939.
Barrozo, R. B., D. Jarriault, N. Deisig, C. Gemeno, C. Monsempes, P. Lucas, C
Gadenne, y S. Anton. 2011. Mating-induced differential coding of plant odour
and sex pheromone in a male moth. Eur. J. Neurosci. 33: 1841-1850.

94
Bernays, E. A., y R. F. Chapman. 1994. Host-plant selection by phytophagous insects.
Chapman & Hall, New York. 312 pp.
Bernays, E. A., M. S. Singer, y D. Rodrigues. 2004. Foraging in nature: foraging
efficiency and attentiveness in caterpillars with different diet breadths. Ecol.
Entomol. 29: 389-397.
Bloch, G., E. Hazan, y A. Rafaeli. 2013. Circadian rhythms and endocrine functions in
adult insects. J. Insect Physiol. 59: 56-69.
Bora, D. S., B. Deka, y A. Sen. 2013. Host plant selection by larvae of the muga Silk
Moth, Antheraea assamensis, and the role of the antenna and maxillary palp. J.
Insect Sci. 13: 1-13.
Bricchi, I., M. Leitner, M. Foti, A. Mithofer, W. Boland, y M. E. Maffei. 2010. Robotic
mechanical wounding (MecWorm) versus herbivore-induced responses: early
signaling and volatile emission in Lima bean (Phaseolus lunatus L.). Planta 232:
719-29.
Browne, L. B. 1993. Physiologically induced changes in resource-oriented behavior.
Annu. Rev. Entomol. 38: 1-23.
Bruce, T. J., y J. A. Pickett. 2011. Perception of plant volatile blends by herbivorous
insects--finding the right mix. Phytochemistry 72: 1605-11.
Bruce, T. J., L. J. Wadhams, y C. M. Woodcock. 2005. Insect host location: a volatile
situation. Trends Plant Sci. 10: 269-74.
Calas, D., A. Berthier, y F. Marion-Poll. 2007. Do european corn borer females detect
and avoid laying eggs in the presence of 20-hydroxyecdysone? J. Chem. Ecol.
33: 1393-404.
Callado-Galindo, M., P. Villa-Ayala, F. Castrejón-Ayala, y A. Jiménez-Pérez. 2013.
Effect of age, body weight and multiple mating on Copitarsia decolora
(Lepidoptera: Noctuidae) reproductive potential and longevity. J. Insect Behav.
26:860-872.
Carroll, M. J., E. A. Schmelz, y P. E. Teal. 2008. The attraction of Spodoptera
frugiperda neonates to cowpea seedlings is mediated by volatiles induced by
conspecific herbivory and the elicitor inceptin. J. Chem. Ecol. 34: 291-300.

95
Carroll, M. J., E. A. Schmelz, R. L. Meagher, y P. E. Teal. 2006. Attraction of
Spodoptera frugiperda larvae to volatiles from herbivore-damaged maize
seedlings. J. Chem. Ecol. 32: 1911-24.
Castrejón-Gómez, V. R., J. Cibrián-Tovar, J. Romero-Napoles, y M. Camino-Lavin.
2000. Mating frequency in wild females of Copitarsia consueta (Lepidoptera:
Noctuidae). Fla. Entomol. 83: 374-376.
Castrejon, F., A., Virgen, y J. C. Rojas. 2006. Influence of chemicals cues from host
plants on the behavior of neonate Estigmene acrea larvae (Lepidoptera:
Noctuidae). Environ. Entomol. 35: 700-707.
Cha, D. H., S. Nojima, S. P. Hesler, A. Zhang, C. E. Linn, Jr., W. L. Roelofs, y G. M.
Loeb. 2008. Identification and field evaluation of grape shoot volatiles attractive
to female grape berry moth (Paralobesia viteana). J. Chem. Ecol. 34: 1180-9.
Cibrián, T. J. 1999. Feromonas y su importancia en el manejo de plagas. Hortalizas,
Plagas y Enfermedades. Ed. Trillas, México. pp: 435-460.
Cook, S. M., Z. R. Khan, y J. A. Pickett. 2007. The use of push-pull strategies in
integrated pest management. Ann. Rev. Entomol. 52: 375–400.
Cunningham, J. P., S. A. West, y D. J. Wright. 1998a. Learning in the nectar foraging
behaviour of Helicoverpa armigera. Ecol. Entomol. 23: 363-369.
Cunningham, J. P., M. F. A. Jallow, D. J. Wright, y M. P. Zalucki. 1998b. Learning in
host selection in Helicoverpa armigera (Hubner) (Lepidoptera: Noctuidae).
Anim. Behav. 55: 227-34.
Cunningham, J. P., C. L. Lange, G. H. Walter, y M. P. Zalucki. 2011. Host location
behaviour in the desert caterpillar, Heliothis punctifera. Entomol. Exp. App.
141: 1-7.
D'Auria, J. C., E. Pichersky, A. Schaub, A. Hansel, y J. Gershenzon. 2007.
Characterization of a BAHD acyltransferase responsible for producing the green
leaf volatile (Z)-3-hexen-1-yl acetate in Arabidopsis thaliana. Plant J. 49: 194-
207.
D'Errico, G., N. Faraone, G. Rotundo, A. De Cristofaro, y R. M. Trimble. 2013. Sensory
adaptation of antennae and sex pheromone-mediated flight behavior in male
Oriental Fruit Moths (Leptidoptera: Tortricidae) after prolonged exposure to

96
single and tertiary blends of synthetic sex pheromone. Environ. Entomol. 42:
548-557.
Daly, K. C., y A. J. Figueredo. 2000. Habituation of sexual response in male Heliothis
moths. Physiol. Entomol. 25: 180-190.
De Boer, G. 2006. The role of the antennae and maxillary palps in mediating food
preference by larvae of the tobacco hornworm, Manduca sexta. Entomol. Exp.
App. 119: 29-38.
De Bruyne, M., y T. C. Baker. 2008. Odor Detection in Insects: Volatile Codes. J.
Chem. Ecol. 34: 882-897.
De Moraes, C. M., M. C. Mescher, y J. H. Tumlinson. 2001. Caterpillar-induced
nocturnal plant volatiles repel conspecific females. Nature 410: 577-580.
De Moraes, C. M., W. J. Lewis, P. W. Pare, H. T. Alborn, y J. H. Tumlinson. 1998.
Herbivore-infested plants selectively attract parasitoids. Nature 393: 570-573.
De Vos, M., V. R. Van Oosten, R. M. Van Poecke, J. A. Van Pelt, M. J. Pozo, M. J.
Mueller, A. J. Buchala, J. P. Metraux, L. C. Van Loon, M. Dicke, y C. M.
Pieterse. 2005. Signal signature and transcriptome changes of Arabidopsis
during pathogen and insect attack. Mol. Plant Microbe Interact. 18: 923-37.
Decker, S., S. Mc Connaughey, y T. L. Page. 2007. Circadian regulation of insect
olfactory learning. Proc. Natl. Acad. Sci. USA 104: 15905-15910.
Del Campo, M. L., C. I. Miles, F. C. Schroeder, C. Mueller, R. Booker, y J. A.
Renwick. 2001. Host recognition by the tobacco hornworm is mediated by a
host plant compound. Nature 411: 186-189.
Diamond, S. E., y J. G. Kingsolver. 2010. Fitness consequences of host plant choice: a
field experiment. Oikos 119: 542-550.
Dicke, M., y J. J. A. van Loon. 2000. Multitrophic effects of herbivore-induced plant
volatiles in an evolutionary context. Entomol. Exp. App. 97: 237-249.
Dicke, M., T. A. Van Beek, M. A. Posthumus, N. Ben Dom, H. Van Bokhoven, y A. De
Groot. 1990. Isolation and identification of volatile kairomone that affects
acarine predatorprey interactions Involvement of host plant in its production. J.
Chem. Ecol. 16: 381-396.
Dicke, M., A. A. Agrawal, y J. Bruin. 2003. Plants talk, but are they deaf? Trends Plant
Sci. 8:403-405.

97
Doak, P., P. Kareiva, y J. Kingsolver. 2006. Fitness consequences of choosy oviposition
for a time-limited butterfly. Ecology 87: 395-408.
Dudareva, N., E. Pichersky, y J. Gershenzon. 2004. Biochemistry of plant volatiles.
Plant Physiol. 135: 1893-902.
Dukas, R. 2006. Learning in the context of sexual behaviour in insects. Anim. Biol. 56:
125-142.
Dukas, R. 2008. Evolutionary biology of insect learning. Annu. Rev. Entomol. 53: 145-
60.
Dukas, R., Z. Durisko, y L. Dukas. 2012. Learning in the context of sexual behaviour
and danger in female and male Drosophila pseudoobscura. Anim. Behav. 83: 95-
101.
Dumont, S. y J. N. McNeil. 1992. Responsiveness of Pseudaletia unipuncta
(Lepidoptera:Noctuidae) males, maintained as adults under different temperature
and photoperiodic conditions, to female sex pheromone. J. Chem. Ecol.18: 1797-
1807.
Duportets, L., M. C. Dufour, F. Couillaud, y C. Gadenne. 1998. Biosynthetic activity of
corpora allata growth of sex accessory glands and mating in the male moth
Agrotis ipsilon. J. Exp. Biol. 201: 2425-2432.
Evenden, M. L. y R. Gries. 2008. Plasticity of male response to sex pheromone depends
on physiological state in a long-lived moth. Anim. Behav. 75: 663-672.
Fatzinger, C. W., y E. P. Merkel. 1985. Oviposition and feeding preferences of the
southern pine coneworm (Lepidoptera: Pyralidae) for different host-plant
materials and observations on monoterpenes as an oviposition stimulant. J
Chem. Ecol. 11: 689-99.
Figueredo, A. J., y T. C. Baker. 1992. Reduction of the response to sex pheromone in
the oriental fruit moth,Grapholita molesta (Lepidoptera: Tortricidae) following.
J. Insect Behav. 5: 347-363.
Finch, S., y R. H. Collier. 2000. Host-plant selection by insects – a theory based on
‘appropriate/inappropriate landings’ by pest insects of cruciferous plants.
Entomol. Exp.App. 96: 91-102.
Foster, S. P., y M. O. Harris. 1997. Behavioral manipulation methods for insect pest-
management. Annu. Rev. Entomol. 42: 123–46.

98
Foster, S. P., y A. J. Howard. 1998. Influence of stimuli from Camellia japonica on
ovipositional behavior of generalist herbivore Epiphyas postvittana. J. Chem.
Ecol. 24: 1251-1275.
Fraser, A. M., W. Mechaber, y J. Hildebrand. 2003. Electroantennographic and
behavioral responses of the sphinx moth Manduca sexta to host plant headspace
volatiles. J. Chem. Ecol. 29-234-239.
Gadenne, C., M. Renou, y L. Sreng. 1993. Hormonal control of pheromone
responsiveness in the male black cutworm Agrotis ipsilon. Experientia 49: 721-
724.
Gadenne, C. y S. Anton. 2000. Central processing of sex pheromone stimuli is
differentially regulated by juvenile hormone in a male moth. J. Insect Physiol.
46: 1195-1206.
Gadenne. C., M. C. Dufour, y S. Anton. 2001. Transient post-mating inhibition of
behavioural and central nervous responses to sex pheromone in an insect. Proc.
R. Soc. Lond. B 268: 1631-1635.
Gang, D. R., J. Wang, N. Dudareva, K. H. Nam, J. E. Simon, E. Lewinsohn, y E.
Pichersky. 2001. An investigation of the storage and biosynthesis of
phenylpropenes in sweet basil. Plant Physiol. 125: 539-55.
García-Robledo, C., y C. C. Horvitz. 2012. Parent–offspring conflicts, “optimal bad
motherhood” and the “mother knows best” principles in insect herbivores
colonizing novel host plants. Ecol. Evol. 2: 1446-1457.
Geervliet, J. B. F., M. A. Posthumus, L. E. M. Vet, y M. Dicke 1997. Comparative
analysis of headspace volatiles from different caterpillar-infested or uninfested
food plants of Pieris species. J. Chem. Ecol. 23; 2935-2954.
Gillott, C. 2003. Male accessory gland secretions: Modulators of female reproductive
physiology and behavior. Annu. Rev. Entomol. 48: 163-184.
Gómez, O., E. Malo, S. Arellano, y J. Rojas. 2012. Pheromone trap for monitoring
Copitarsia decolora (Lepidoptera:Noctuidae) activity in cruciferous crops in
Mexico. Fla. Entomol. 95: 602-609.
Gouinguene, S. P., y T. C. Turlings. 2002. The effects of abiotic factors on induced
volatile emissions in corn plants. Plant Physiol. 129: 1296-307.

99
Gripenberg, S., J. P. Salminen, y T. Roslin. 2007a. A tree in the eyes of a moth temporal
variation in oak leaf quality and leaf miner performance. Oikos 116: 592–600.
Gripenberg, S., E. Morriën, A. Cudmore, J. P. Salminen, y T. Roslin. 2007b. Resource
selection by female moths in a heterogeneous environment: what is a poor girl to
do? J. Anim. Ecol. 76: 854-865.
Gripenberg, S., P. J. Mayhew, M. Parnell, y T. Roslin. 2010. A meta-analysis of
preference–performance relationships in phytophagous insects. Ecol. Lett. 13:
383-393
Groenteman, R., M. Guershon, y M. Coll. 2006. Effects of leaf nitrogen content on
oviposition site selection, offspring performance, and intraspecific interactions
in an omnivorous bug. Ecol. Entomol. 31: 155-161.
Guerrieri, F., C. Gemeno, C. Monsempes, S. Anton, E. Jacquin-Joly, P. Lucas, y J.-M.
Devaud. 2012. Experience-dependent modulation of antennal sensitivity and
input to antennal lobes in male moths (Spodoptera littoralis) pre-exposed to sex
pheromone. J. Exp. Biol. 215: 2334-2341.
Hare, J. D. 2011. Ecological role of volatiles produced by plants in response to damage
by herbivorous insects. Annu. Rev. Entomol. 56: 161-80.
Harris, M. O. y S. P. Foster. 1995. Behavior and integration. pp. 3-46 En: Cardé, R. T.
and W. J. Bell (eds.). Chemical Ecology of Insects 2. Chapman & Hall, New
York.
Harris, M. O., M. Sandanayake, y S. P. Foster. 1999. Chemical stimuli from apple
influence the behavior of neonate caterpillars of the generalist herbivore,
Epiphyas postvittana. J. Chem. Ecol. 25: 1717-1738.
Harris, M. O., S. P. Foster, T. Bittar, K. Ekanayake, K. Looij, y A. Howard. 1995.
Visual behaviour of neonate larvae of the light brown apple moth. Entomol.
Exp. App.77: 323-334.
Hartlieb, E., y H. Rembold. 1996. Behavioral response of female Helicoverpa
(Heliothis) armigera HB. (Lepidoptera: Noctuidae) moths to synthetic
pigeonpea (Cajanus cajan L.) kairomone. J. Chem. Ecol. 22: 821-837.
Heil, M., U. Lion, y W. Boland. 2008. Defense-inducing volatiles: in search of the
active motif. J. Chem. Ecol. 34: 601-604.

100
Heil, M., E. Ibarra-Laclette, R. M. Adame-Álvarez, O. Martínez., E. Ramírez-Chávez, J.
Molina-Torres, y L. Herrera-Estrella. 2012. How plants sense wounds:
damaged-self recognition is based on plant-derived elicitors and induces
octadecanoid signaling. PLoS One 7: e30537.
Hildebrand, J. 1995. Analysis of chemical signals by nervous systems. En: Chemical
Ecology. The Chemistry of Biotic Interaction. T. Eisner y J. Meinwald (eds).
National Academy Press, Washington, D. C. pp: 161-181.
Hilker, M., y T. Meiners. 2002. Induction of plant responses to oviposition and feeding
by herbivorous arthropods: a comparison, pp. 181-192. In J. Nielsen, C. Kjar
and L. Schoonhoven (eds.), Proceedings of the 11th International Symposium on
Insect-Plant Relationships, vol. 57. Springer Netherlands.
Hilker, M., O. Rohfritsch, y T. Meiners. 2002. The plant’s response towards insect egg
deposition, pp. 205-233, en Hilker, M., & Meiners, T. (eds.), Chemoecology of
Insect Eggs and Egg Deposition. Blackwell Verlag, Berlin.
Holopainen, J. K., y J. Gershenzon. 2010. Multiple stress factors and the emission of
plant VOCs. Trends Plant Sci. 15: 176-184.
Holopainen, J. K., y J. D. Blande. 2013. Where do herbivore-induced plant volatiles go?
Front Plant Sci. 4: 185.
Honda, K. 1995. Chemical basis of differential oviposition by lepidopterous insects.
Arch. Insect Biochem. Physiol. 30: 1-23.
Hopkins, R. J., N. M. van Dam, y J. J. van Loon. 2009. Role of glucosinolates in insect-
plant relationships and multitrophic interactions. Annu. Rev. Entomol. 54:57-83.
Hopper, K. R. 1999. Risk-spreading and bet-hedging in insect population biology.
Annu. Rev. Entomol. 44: 535-60.
Howe, G. A., y G. Jander. 2008. Plant immunity to insect herbivores. Annu. Rev. Plant
Biol. 59: 41-66.
Huang, C. H., F. M. Yan, J. A. Byers, R. J. Wang, y C. R. Xu. 2009. Volatiles induced
by the larvae of the Asian corn borer (Ostrinia furnacalis) in maize plants affect
behavior of conspecific larvae and female adults. Insect Sci. 16: 311-320.
Huang, X., y J. A. A. Renwick. 1994. Relative activities of glucosinolates as oviposition
stimulants forPieris rapae andP. napi oleracea. J. Chem. Ecol. 20: 1025-1037.

101
Ikeura, H., F. Kobayashi, y Y. Hayata. 2010. How do Pieris rapae search for
Brassicaceae host plants? Biochem. Syst. Ecol.38: 1199-1203.
Jaenike, J. 1978. On optimal oviposition behavior in phytophagous insects. Theor.
Popul. Biol. 14: 350-356.
Jaenike, J. 1990. Host specialization in phytophagous insects. Ann. Rev. Ecol. Syst. 21:
243-273.
Jallow, M. F., M. P. Zalucki, y G. P. Fitt. 1999. Role of chemical cues from cotton in
mediating host selection and oviposition behaviour in Helicoverpa armigera
(Hübner) (Lepidoptera: Noctuidae). Aust. J. Entomol. 38: 359-366.
Janz, N. 2008. Evolutionary ecology of oviposition strategies, pp. 349-376,
Chemoecology of insect eggs and egg deposition. Blackwell Publishing Ltd.
Javois, J. y T. Tammaru. 2006. The effect of egg load on readiness to accept a low-
quality host plant is weak and age dependent in a geometrid moth. Ecol.
Entomol. 31: 597–600.
Jeffries, M. J., y J. H. Lawton. 1984. Enemy free space and the structure of ecological
communities. Biol J Linnean Soc, 23:269–286.
Jiménez-Pérez, A. y Q. Wang. Effect of mating delay on the reproductive performance
of Cnephasia jactatana (Lepidoptera: Tortricidae). J. Econ. Entomol. 96: 592-
598.
Jones, E. E. 1977. Movement patterns and egg distribution in cabbage butterflies. J.
Anim. Ecol. 46: 195-212.
Judd, G. J. R., Gardiner, M. G. T., DeLury, N. C. y G. Karg. 2005. Reduced antennal
sensitivity, behavioural response, and attraction of male codling moths, Cydia
pomonella, to their pheromone (E,E)-8,10-dodecadien-1-ol following various
pre-exposure regimes Entomol. exp. appl. 114: 65-78.
Keil, T. A. 1999. Morphology and development of the peripheral olfactory organs. pp.
5-48 In Hansson BS [ed.], Insect Olfaction. Springer, Berlin, Germany.
Kessler, A., y I. T. Baldwin. 2001. Defensive function of herbivore-induced plant
volatile emissions in nature. Science 291: 2141-2144.
Kessler, A., y K. Morrell. 2010. Plant volatile signalling: multitrophic interactions in the
headspace En: The chemistry and biology of volatiles. Ed. Andreas Herrmann
Wiley. 95-112.

102
Kovats, E. 1958. Gaschromatographische charakterisierung organischer Verbindungen.
Teil 1. Retentionsindices aliphatischer Halogenide, Alkohole, Aldehyde, und
Ketone. Helv. Chim. Acta 41:1915–1932
Kloppenburg, P., D. Ferns, y A. R. Mercer. 1999. Serotonin enhances central olfactory
neuron responses to female sex pheromone in the male sphinx moth manduca
sexta. J. Neurosci. 19: 8172-81.
Knolhoff, L. M., y D. G. Heckel. 2014. Behavioral assays for studies of host plant
choice and adaptation in herbivorous insects. Annu. Rev. Entomol. 59: 263-278.
Landolt, P. J. 1993. Effects of host plant leaf damage on cabbage looper moth attraction
and oviposition. Entomol. Exp. Appl. 67: 79-85.
Landolt, P. J., y O. Molina. 1996. Host-finding by cabbage looper moths (Lepidoptera:
Noctuidae): Learning of host odor upon contact with host foliage. J. Insect
Behav. 9: 899-908.
Landolt, P. J., R. W. Hofstetter, y P. S. Chapman. 1998. Neonate codling moth larvae
(Lepidoptera: Tortricidae) orient anemotactically to odor of immature apple
fruit. Pan Pac. Entomol. 74:140–49.
Landolt, P., J. Brumley, C. Smithhisler, L. Biddick, y R. Hofstetter. 2000. Apple fruit
infested with codling moth are more attractive to neonate codling moth larvae
and possess increased amounts of (E,E)-α-farnesene. J. Chem. Ecol 26: 1685-
1699.
Leal, W. S. 2013. Odorant reception in insects: roles of receptors, binding proteins, and
degrading enzymes. Annu. Rev. Entomol. 58: 373-91.
Lee, H. S., T. Hieu, y Y. J. Ahn. 2006. Oviposition-stimulating activity of (E)-capsaicin
identified in Capsicum annuum fruit and related compounds towards
Helicoverpa assulta (Lepidoptera: Noctuidae). Chemoecology 16: 153-157.
Loughrin, J., A. Manukian, R. Heath, y J. Tumlinson. 1995. Volatiles emitted by
different cotton varieties damaged by feeding beet armyworm larvae. J. Chem.
Ecol. 21: 1217-1227.
Lu, P. F., L. Q. Huang, y C. Z. Wang. 2012. Identification and field evaluation of pear
fruit volatiles attractive to the oriental fruit moth, Cydia molesta. J. Chem. Ecol.
38: 1003-16.

103
Major, I. T., y C. P. Constabel. 2006. Molecular analysis of poplar defense against
herbivory: comparison of wound- and insect elicitor-induced gene expression.
New Phytol. 172: 617-35.
Marchioro, C. A., y L. A. Foerster. 2014. Preference—Performance linkage in the
diamondback moth, Plutella xylostella, and implications for its management. J.
Insect Sci. 14: 1-14.
Matsui, K., K. Sugimoto, J. Mano, R. Ozawa, y J. Takabayashi. 2012. Differential
metabolisms of green leaf volatiles in injured and intact parts of a wounded leaf
meet distinct ecophysiological requirements. PLoS ONE 7:e36433.
Matthews, R. y J. Matthews. 2010. Insect Behavior. Programming and Integrating
Behavior. Chapter 2. Second Edition. Springer Dordrecht Heidelberg London
New York p. 341-387.
Mechauber, W., C. Capaldo, y J. Hildebrand. 2002. Behavioral responses of adult
female tobacco hornworms, Manduca sexta, to host plant volatiles change with
age and mating status. J. Insect Sci. 2:5-8.
Meiners, T., y M. Hilker. 2000. Induction of plant synomones by oviposition of a
phytophagous insect. J. Chem. Ecol. 26: 221-232.
Metcalf, R. L., y E. R. Metcalf. 1992. Plant kairomones in insect ecology and control.
London, Chapman and Hall. 168 p.
Miles, C., M. D. Campo, y J. A. Renwick. 2005. Behavioral and chemosensory
responses to a host recognition cue by larvae of Pieris rapae. J. Comp. Physiol.
A. 191: 147-155.
Miller, J., P. Siegert, F. Amino, y E. Walker. 2009. Designation of chemicals in terms of
the locomotor responses they elicit from insects: an update of Dethier et al.
(1960). J. Econ. Entomol. 102: 2056-2060.
Minkenberg, O. P., M. Tatar, y J. A. Rosenheim. 1992. Egg load as a major source of
variability in insect foraging and oviposition behavior. Oikos: 134-142.
Mowrey, W. R., y D. S. Portman. 2012. Sex differences in behavioral decision-making
and the modulation of shared neural circuits. Biol. Sex. Differ. 3: 8.
Mumm, R., y M. Dicke. 2010. Variation in natural plant products and the attraction of
bodyguards involved in indirect plant defense. Can. J. Zool. 88: 628-667.

104
Muñiz-Reyes, E., J Cibrián-Tovar, J. Rojas-Leon, O. Díaz-Gómez, J. Valdés-Carrasco,
y N. Bautista-Martínez. 2007. Capturas de Copitarsia decolora (Lepidoptera:
Noctuidae) en trampas cebadas con diferentes proporciones de feromona sexual
Agrociencia 41: 575-581.
Najar-Rodriguez, A. J., C. G. Galizia, J. Stierle, y S. Dorn. 2010. Behavioral and
neurophysiological responses of an insect to changing ratios of constituents in
host plant-derived volatile mixtures. J. Exp. Biol. 213: 3388-97.
Olsson, P. O. C., O. Anderbrant, y C. Löfstedt. 2006. Experience influences oviposition
behaviour in two pyralid moths, Ephestia cautella and Plodia interpunctella.
Anim. Behav. 72: 545-551.
Ozawa, R., K. Shiojiri, K. Matsui, y J. Takabayashi. 2013. Intermittent exposure to
traces of green leaf volatiles triggers the production of (Z)-3-hexen-1-yl acetate
and (Z)-3-hexen-1-ol in exposed plants. Plant Signal Behav. 8:e27013.
Papaj, D. R., y R. J. Prokopy. 1989. Ecological and evolutionary aspects of learning in
phytophagous insects. Annu. Rev. Entomol. 34: 315-350.
Paré, P. W., y J. H. Tumlinson. 1996. Plant volatile signals u¿in response to herbivore
feeding. Behabioral Ecology Symposium 95. Fla. Entomol. 79: 93-103.
Paré, P. W., y J. H. Tumlinson. 1997a. Induced synthesis of plant volatiles. Nature 385:
30-31.
Paré, P. W., y J. H. Tumlinson. 1997b. De Novo Biosynthesis of Volatiles Induced by
Insect Herbivory in Cotton Plants. Plant Physiol 114: 1161-1167.
Paré, P. W., y J. H. Tumlinson. 1998. Cotton volatiles synthesized and released distal to
the site of insect damage. Phytochemistry 47: 521-526.
Paré, P. W., y J. H. Tumlinson. 1999. Plan volatiles as a defense against insect
herbivores. Plant Physiol. 121: 325-331.
Pérez, C. 2010. Respuesta de atracción y preferencia alimentaria de larvas de Copitarsia
decolora (Guenée) (Lepidoptera: Noctuidae) hacia plantas hospederas. Tesis:
Maestría en Ciencias en Manejo Agroecológico de Plagas y Enfermedades.
Instituto Politécnico Nacional.
Petzold, J. L. 2009. Interactions between a specialist and generalist moth and their host
plants, pp. 169, Plant Biology. North Carolina State University, Raleigh, NC.

105
Picimbon, J., C. Gadenne, M. Becard, L. Clement, y L. Sreng. 1997. Sex pheromone of
the french black cutworm moth, Agrotis ipsilon (Lepidoptera: Noctuidae)
identification and regulation of multicomponent blend. J. Chem. Ecol. 23: 211-
229.
Pivnick, K.A., B. J. Jarvis, G. P. Slater, C. Gillott, y E. Underhill. 1990. Attraction of
the diamondback moth (Lepidoptera: Plutellidae) to volatiles of oriental
mustard: the influence of age, sex, and prior exposure to mates and host plants.
Environ. Entomol. 19:704-709.
Pivnick, K. A., B. J. Jarvis, y G. P. Slater. 1994. Identification of olfactory cues used in
host-plant finding by diamondback moth, Plutella xylostella (Lepidoptera:
Plutellidae). J. Chem. Ecol. 20:1407-1427.
Pophof, B. 2000. Octopamine modulates the sensitivity of silkmoth pheromone receptor
neurons. J. Comp. Physiol. A. 186: 307-13.
Potter, K. A., J. Bronstein, y G. Davidowitz. 2012. Choice of oviposition sites by
Manduca sexta and its consequences for egg and larval performance. Entomol.
Exp. App. 144: 286-293.
Pöykkö, H. 2006. Females and larvae of a geometrid moth, Cleorodes lichenaria, refer
a lichen host that assures shortest larval period. Environ. Entomol. 35: 1669-
1676.
Proffit, M., G. Birgersson, M. Bengtsson, R. Reis Jr., P. Witzgall, y E. Lima. 2011.
Attraction and oviposition of Tuta absoluta females in response to tomato leaf
volatiles. J. Chem. Ecol. 37:565-74.
Prokopy, R., y W. J. Lewis. 1993. Application of learning to pest management. pp. 308-
342. In D. Papaj and A. Lewis (eds.), Insect Learning. Springer US.
Rausher, M. D., y D. R. Papaj. 1983. Demographic consequences of descrimination
among conspecific host plants by battus philenor butterflies. Ecology 64: 1402-
1410.
Reddy, G. V., y A. Guerrero. 2000. Behavioral responses of the diamondback moth,
Plutella xylostella, to green leaf volatiles of Brassica oleracea subsp. capitata. J.
Agric. Food Chem. 48: 6025-9.

106
Refsnider, J. M., y F. J. Janzen. 2010. Putting eggs in one basket: ecological and
evolutionary hypotheses for variation in oviposition-site choice. Ann. Rev.
Ecol. Evol. Syst. 41: 39-57.
Rembold, H., A. C. Köhne, y A. Schroth. 1991. Behavioral response of Heliothis
armigera Hb. (Lep., Noctuidae) moths on a synthetic chickpea (Cicer arietinum
L.) kairomone. J. App. Entomol. 112: 254-262.
Renwick, J. A., M. Haribal, S. Gouinguene, y E. Stadler. 2006. Isothiocyanates
stimulating oviposition by the diamondback moth, Plutella xylostella. J. Chem.
Ecol. 32: 755-66.
Renwick, J. A., y F. S. Chew. 1994. Oviposition Behavior in Lepidoptera. Ann. Rev.
Entomol. 39: 377-400.
Rharrabe, K., E. Jacquin-Joly, y F. Marion-Poll. 2014. Electrophysiological and
behavioral responses of Spodoptera littoralis caterpillars to attractive and
repellent plant volatiles. Front. Ecol. Evol. 2:1-9
Riffell, J. A., H. Lei, T. A. Christensen, y J. G. Hildebrand. 2009. Characterization and
coding of behaviorally significant odor mixtures. Curr. Biol. 19: 335-40.
Rivera, M. J., y H. J. Burrack. 2012. Host utilization is mediated by movement of pre-
feeding Phthorimaea operculella larvae in the Nicotiana tabacum
agroecosystem. Entomol. Exp. App. 145: 153-161.
Rodriguez-Saona, C., y L. Stelinski. 2009. IPM, Behaviour-Modifying Strategies in
IPM: Theroy and Practice. In R. Peshin and A. K. Dhawan (Eds.), Integrated
Pest Management: Volume 1: Innovation-Development Process (1st ed., p. 689).
Springer.
Roelofs, W. L. 1995. Chemistry of sex attraction. Proc. Natl. Acad. Sci. U.S.A. 92: 44-
9.
Rogers, C. E. y O. G. Jr. Marti. 1994. Reproductive potential of once-mated moths of
fall armyworm (Lepidoptera: Noctuidae). Fla. Entomol. 77: 402-410.
Rogers, C. E. y O. G. Jr. Marti.1996. Beet armyworm (Lepidoptera: Noctuidae): Effects
of age at first mating on reproductive potential. Fla. Entomol. 79: 343-352
Rojas, J. C. y J. Cibrián-Tovar. 1994. Reproductive behavior of Copitarsia consueta
(Lepidoptera: Noctuidae): mating frequency, effect of age on mating, and

107
influence of delayed mating on fecundity and egg fertility. Pan-Pac. Entomol 70:
276-282.
Rojas, J. C. 1999. Electrophysiological and behavioral responses of the cabbage moth to
plant volatiles. J. Chem. Ecol. 25: 1867-1883.
Rojas, J. C., y T. D. Wyatt. 1999a. Role of visual cues and interaction with host odour
during the host-finding behaviour of the cabbage moth. Entomol. Exp. Appl. 91:
59-65.
Rojas, J. C., y T. D. Wyatt. 1999b. The role of pre- and post-imaginal experience in the
host-finding and oviposition behaviour of the cabbage moth. Physiol. Entomol.
24: 83-89.
Rojas, J. C., A. Virgen, y L. Cruz-López. 2003. Chemical and tactile cues influencing
oviposition of a generalist moth, Spodoptera frugiperda (Lepidoptera:
Noctuidae). Environ. Entomol. 32: 1386-1392.
Rojas, J. C., L. Cruz-Lopez, E. A. Malo, O. Diaz-Gomez, G. Calyecac, y J. Cibrian-
Tovar. 2006. Identification of the sex pheromone of Copitarsia decolora
(Lepidoptera: Noctuidae). J. Econ. Entomol. 99: 797-802.
Rumbo, E. R. y R. A. Vickers. 1997. Prolonged adaptation as possible mating
disruption mechanism in oriental fruit moth Cydia (=Grapholita) molesta. J.
Chem. Ecol. 23: 445-457.
Saveer, A. M., S. H. Kromann, G. Birgersson, M. Bengtsson, T. Lindblom, A.
Balkenius, B. S. Hansson, P. Witzgall, P. G. Becher, y R. Ignell. 2012. Floral to
green: mating switches moth olfactory coding and preference. Proc. Biol. Sci.
279: 2314-22.
Scala, A., S. Allmann, R. Mirabella, M. A. Haring, y R. C. Schuurink. 2013. Green leaf
volatiles: a plant's multifunctional weapon against herbivores and pathogens. Int.
J. Mol. Sci. 14: 17781-811.
Scheirs, J., y L. De Bruyn. 2002. Integrating optimal foraging and optimal oviposition
theory in plant–insect research. Oikos 96: 187-191.
Schoonhoven, L. M., J. Van Loon, y M. Dicke. 2005. Insect-plant biology. 2ed.Oxford
University Press, Nueva York. 421p.

108
Shikano, I., Y. Akhtar, y M. Isman. 2010. Relationship between adult and larval host
plant selection and larval performance in the generalist moth, Trichoplusia ni.
Arth. Plant Int. 4: 197-205.
Shiojiri, K. Kishimoto, R. Ozawa, S. Kugimiya, S. Urashimo, G. Arimura, J. Horiuchi,
T. Nishioka, K. Matsui, y J. Takabayashi. 2006. Changing green leaf volatile
biosynthesis in plants: An approach for improving plant resistance against both
herbivores and pathogens. Proc. Natl. Acad. Sci. 103: 16672-16676.
Simmonds, M. S. 2001. Importance of flavonoids in insect--plant interactions: feeding
and oviposition. Phytochemistry 56: 245-52.
Singh, A. K., y S. Mullick. 2002. Leaf volatiles as attractants for neonate Helicoverpa
armigera Hbn. (Lep., Noctuidae) larvae. J. App. Entomol. 126: 14-19.
Singer, M. S., D. Rodríguez, J. O. Stireman, y Y. Carriére. 2004. Roles of food quality
and enemy-free space in host use by a generalist insect herbivore. Ecology
85:2747-2753.
Soques, S., G. M. Vazquez, C. M. Grozinger, y F. Gould. 2010. Age and mating status
do not affect transcript levels of odorant receptor genes in male antennae of
Heliothis virescens and Heliothis subflexa. J. Chem. Ecol. 36: 1226-1233.
Spathe, A., A. Reinecke, A. Haverkamp, B. S. Hansson, y M. Knaden. 2013. Host plant
odors represent immiscible information entities - blend composition and
concentration matter in hawkmoths. PLoS One 8: e77135.
Städler, E. 1974. Host plant stimuli affecting oviposition behavior of the eastern spruce
budworm. Entomol. Exp. App. 17: 176-188.
Städler, E., y K. Reifenrath. 2009. Glucosinolates on the leaf surface perceived by insect
herbivores: review of ambiguous results and new investigations. Phytochem.
Rev. 8:207-225.
Stelinski, L. L., L. J. Gut, K. J. Vogel, yJ. R. Miller. 2004. Behaviors of naïve vs.
pheromone-exposed leafroller moths in plumes from high-dosage pheromone
dispensers in a sustained-flight wind tunnel: implications for mating disruption
of these species. J. Insect Behav. 17: 533-554.
Stelinski, L. L., K. J. Vogel, L. J. Gut, y J. R. Miller. 2005. Seconds-long preexposures
to pheromone from rubber septum or polyethelene tube dispensers alters

109
subsequent behavioral responses of male grapholita molesta (Lepidoptera:
Tortricidae) in a sustained-flight tunnel. Environ. Entomol. 34: 696-704.
Stelinski, L. L., A. L. Il’ichev, y L. J. Gut. 2006a. Antennal and behavioral responses of
virgin and mated oriental fruit moth (Lepidoptera:Tortricidae) females to their
sex pheromone. Ann. Entomol. Soc. Am. 99: 898-904.
Stelinski, L. L., L. J. Gut, y J. R. Miller. 2006b. Orientational behaviors and EAG
responses of male codling moth after exposure to synthetic sex pheromone from
various dispensers. J. Chem. Ecol. 32: 1527-38.
Suárez-Vargas, A. D., N. Bautista-Martínez, J. Valdez-Carrasco, A. Angulo-Ormeño, R.
Alatorre-Rosas, J. Vera-Graziano, A. Equihua-Martínez, y V. Manuel Pinto.
2006. Fluctuación poblacional de Copitarsia decolora (Gueéne) y su asociación
con crucíferas comerciales Agrociencia 40: 501-509.
Takeuchi, H., M. P. Zalucki, y M. J. Furlong. 2009. Crocidolomia pavonana larval
foraging: behavior and feeding site preferences on cabbage, Brassica oleracea.
Entomol. Exp. App. 133: 154-164.
Tasin, M., A. C. Bäckman, M. Bengtsson, N. Varela, C. Ioriatti, y P. Witzgall. 2006.
Wind tunnel attraction of grapevine moth females, Lobesia Botrana, to natural
and artificial grape odour. Chemoecology 16: 87-92.
Thompson, J. N., y O. Pellmyr. 1991. Evolution of oviposition behavior and host
preference in lepidoptera. Ann. Rev. Entomol. 36: 65-89.
Tollsten, L., y G. Bergström. 1988. Headspace volatiles of whole plants and macerated
plant parts of Brassica and Sinapis. Phytochemistry 27: 2073-2077.
Tovar H., N. Bautista., G. Vera, A. Suárez, y S. Ramírez. 2007. Fluctuación poblacional
y parasitismo de larvas de Copitarsia decolora (Guenée), Plutella xylostella L. y
Trichoplusia ni (Hübner) (Lepidoptera) en Brassica oleracea. Act. Zool. Mex.
23: 183-196.
Trimble, R. M. 2012. Sexual behavior of Grapholita molesta and Choristoneura
rosaceana (Lepidoptera:Tortricidae) in a flight tunnel after prolonged exposure
to the aerial concentration of pheromone previously measured in orchards
treated with pheromone for mating disruption. Environ. Entomol. 41: 1481-
1493.

110
Trimble, R. M. y D. B. Marshall. 2010. Differences in the relationship between sensory
adaptation of antennae and concentration of aerial pheromone in the oriental
fruit moth and obliquebanded leafroller (Lepidoptera: Tortricidae): implications
for the role of adaptation in sex pheromone mediated mating disruption of these
species. Environ. Entomol. 39: 625-632.
Turlings, T. C., y J. H. Tumlinson. 1992. Systemic release of chemical signals by
herbivore-injured corn. Proc. Natl. Acad. Sci. U. S. A. 89: 8399-402.
Turlings, T. C., J. H. Loughrin, P. J. Mc Call, U. S. Röse, W. J. Lewis, y J. H.
Tumlinson. 1995. How caterpillar-damaged plants protect themselves by
attracting parasitic wasps. Proc. Natl. Acad. Sci. USA 92: 4169-4174.
Turlings, T., U. Lengwiler, M. Bernasconi, y D. Wechsler. 1998. Timing of induced
volatile emissions in maize seedlings. Planta 207: 146-152.
Tovar, H., N. Bautista, G. Vera, A. Suárez, y S. Ramírez. 2007. Fluctuación poblacional
y parasitismo de larvas de Copitarsia decolora (Guenée), Plutella xylostella L. y
Trichoplusia ni (Hübner) (Lepidoptera) en Brassica oleracea. Act. Zool. Mex. 23:
183-196.
Vander Meer, M. R. K., F. Alvarez, y C. S. Lofgren. 1988. Isolation of the trail
recruitment pheromone of Solenopsis invicta. J. Chem. Ecol. 14: 825–838.
Van Loon, J. J. A., y M. Dicke. 2001. Sensory ecology of arthropods utilizing plant
infochemicals. pp. 253-270. En: Barth, F. G. and A. Schmid (eds.). Ecology of
Sensing. Springer Verlag, Berlin.
Varela, L. G. y E. A. Bernays. 1988. Behavior of newly hatched potato tuber moth
larvae, Phthorimaea operculella Zell. (Lepidoptera: Gelechiidae), in relation to
their host plants. J. Insect Behav. 1:261-275.
Venette, R., y J. Gould. 2006. A pest risk assessment for Copitarsia spp., Insects
associated with importation of commodities into the United States. Euphytica
148: 165-183.
Visser, J. H. 1986. Host Odor Perception in Phytophagous Insects. Annu. Rev. Entomol.
31: 121-144.
Voelckel, C., W. W. Weisser, y I. T. Baldwin. 2004. An analysis of plant-aphid
interactions by different microarray hybridization strategies. Mol. Ecol. 13:
3187-95.

111
Wu, J., y I. T. Baldwin. 2010. New insights into plant responses to the attack from
insect herbivores. Annu. Rev. Genet. 44: 1-24.
Zalucki, M. P., A. R. Clarke, y S. B. Malcolm. 2002. Ecology and behavior of first
instar larval Lepidoptera. Annu. Rev. Entomol. 47:361–393.
Zhang, Z. C., M. Q. Wang, y G. Zhang. 2009. Molecular cloning and expression of
pheromone-binding protein1 from the diamondback moth, Plutella xylostella.
Entomol. Exp. App. 133: 136-145.

ANEXOS
Carta de aceptación del artículo científico.
De: Waldemar Klassen [[email protected]]
Enviado: sábado, 01 de noviembre de 2014 14:32
Para: Angel Rene Arzuffi Barrera
CC: Epsky, Nancy
Asunto: Your manuscript- RES 8 14-039 Effect of Male Age and Mating on Response to the Sex
Pheromone of Copitarsia decolora 11 01 14 EDITED
Dr. René Arzuffi;
Dr. Nancy Epsky, Associate Editor, has recommended that your manuscript be accepted for
publication in the Florida Entomologist.
I concur and I will include it in the March 2015 issue.
Please proof the attached version and make needed changes with "track changes".
Also please email me tiff or jpeg images of the 4 figures.
Best regards,
Waldemar Klassen, Editor

Running Head: Reyes et al.: Copitarsia decolora Sex Pheromone
Please address correspondence to:
René Arzuffi
Km. 8.5 Carretera Yautepec-Jojutla Yautepec, Morelos. C.P. 62731, México
Phone: 55(735)3942020, ext 82511
Fax: ext 82509
Email: [email protected]
Effect of male age and mating status on response to female sex
pheromone of Copitarsia decolora (Lepidoptera: Noctuidae)
Humberto Reyes, René Arzuffi* and Norma Robledo
CEPROBI, Instituto Politécnico Nacional, Km. 8.5 Carretera Yautepec-Jojutla Yautepec,
Morelos C.P. 62731, México
*Corresponding author: [email protected]

Abstract
The effects of male age and mating status on the behavioral response of Copitarsia
decolora (Guenée) males to the glandular extract of the female sex pheromone were studied
by conducting wind tunnel bioassays and electroantennography (EAG). To study the effect
of age, 2-3, 4-5, 6-7, 8-9, 10-11 day old males were used to measure attraction in the wind
tunnel. To study the effect of age on virgin male antenna depolarization, individual males
3-, 6- and 9-days old were used. The effect of male mating status was studied with 4-6 day
old males and subsequent pheromone response was measured after a period of 24 and 48 h.
Both male age and mating status significantly affected male response to the female sex
pheromone. In the wind tunnel, male attraction to the extract reached its maximum in males
4-7 days old and then decreased as moth age increased. The glandular extract provoked a
significantly greater depolarization of the antennae of 6 day old males than in 3 and 9 day
old males. In the wind tunnel, virgin males were more attracted to female extracts than
mated males. Significantly greater depolarization was observed in the antennae of virgin
than mated males.
Key Words: Copitarsia decolora, sex pheromone, age, mating, wind tunnel,
elecroantennography
Resumen
Se estudió el efecto de la edad del macho y el estado de apareamiento sobre la respuesta de
comportamiento de machos de Copitarsia decolora (Guenée) al extracto glandular de la

feromona sexual de la hembra, mediante bioensayos en túnel de viento y
electroantenografía (EAG). Para medir la atracción en túnel de viento se utilizaron machos
de 2-3, 4-5, 6-7, 8-9, 10-11 días de edad y para estudiar el efecto de la edad sobre la
despolarización de sus antenas, se utilizaron individuos de 3, 6 y 9 días de edad. Para
estudiar el efecto del apareamiento se utilizaron machos de 4-6 días de edad y su respuesta
a la feromona se midió 24 y 48 horas después. Ambos, la edad del macho y el estado de
apareamiento, afectan de manera significativa la respuesta de los machos a la feromona
sexual de las hembras. En el túnel de viento, se observó que la atracción de los machos al
extracto fue mayor a los 4-7 días y después disminuyó conforme incrementó la edad. El
extracto glandular provocó una despolarización significativamente mayor en las antenas de
los machos de 6 días de edad que en los machos de 3 y 9 días de edad. En túnel de viento
los machos no apareados fueron más atraídos al extracto glandular de las hembras que los
machos apareados. Se observó una despolarización significativamente mayor en las antenas
de los machos no apareados que en los machos apareados.
Palabras Clave: Copitarsia decolora, feromona sexual, edad, apareamiento, túnel de viento,
electroantennografía
Males moths use sex pheromones emitted by females to locate a mate. However,
male response can be affected by numerous factors, including the physiological state of the
insect (Anton et al. 2007; Evenden & Gries 2008) and experience (Anton et al. 2007).
Pheromones are detected by olfactory receptor neurons (ORN) housed within cuticular

sensilla mainly on the antennae (Keil 1999). The axons of ORNs project via the antennal
nerve into the antennal lobe (AL). There they make synaptic contact with intrinsic AL
neurons, the local interneurons, and with AL projection neurons, which transfer information
to higher brain centers (de Belle & Kanzaki 1999). The AL consists of a species-specific
number of glomeruli (Hansson &Anton 2000). Male moths have large numbers of olfactory
receptor neurons tuned to single compounds of the sex pheromone produced by females,
and large glomeruli, called the macroglomerular complex, are entirely dedicated to the
processing signals pertaining to these sex pheromones (Hansson & Anton 2000).
For some moth species such as Plutella xylostella (L.) (Zhang et al. 2009), Agrotis
ipsilon (Hufnagel) (Anton & Gadenne 1999; Gadenne & Anton 2000; Gadenne et al. 2001;
Barrozo et al. 2010a, 2010b, 2011), Heliothis virescens (F.) and Heliothis subflexa
(Guenée) (Soques et al. 2010) it has been demonstrated that age and mating status can
modify olfactory reception and sexual pheromone processing in males. In these species, the
male response to the female sex pheromone is regulated by hemolymph titers of juvenile
hormone (JH) (Gadenne et al. 1993; Duportets et al. 1998; Anton & Gadenne 1999;
Gadenne & Anton 2000), and by neuromodulators that act on ORNs and AL neurons
(Anton et al. 2007).
Copitarsia decolora (Guenée) (Lepidoptera: Noctuidae) is an important pest of
cruciferous crops in Mexico (Suarez-Vargas et al. 2006) and subject to quarantine in the
United States (Venette & Gould 2006). Rojas et al. (2006) identified the sex pheromone of
C. decolora and, although synthetic sex pheromones have been used as an alternative to
synthetic insecticides (Rojas et al 2006; Muñiz-Reyes et al. 2007; Barrientos-Hernández et
al. 2011; Diaz-Gomez et al. 2012), the effect of environmental (Dumont & McNeil 1992)
and physiological factors on male response to the sex pheromone have not been studied.

However several factors, including of age and physiological status of males, should be
evaluated before the pheromone compounds can be used effectively for C. decolora
management. The efficiency of traps baited with pheromone could be affected importantly
by these 2 factors.
This study examined the effect of age and male mating status on male attraction and
antennal responses to sex pheromone extracts of C. decolora females. Understanding the
effect of the factors on olfactory guided behavior of C. decolora males will contributes to
improving current pheromone-based methods for the management of this pest species.
Materials and Methods _____
INSECTS
Insects used in the bioassays were obtained from a C. decolora colony at the Centro
de Desarrollo de Productos Bióticos, National Polytechnic Institute (IPN) in Yautepec,
Morelos, Mexico. The insects were maintained at 25 ± 3 °C, 60 ± 5% RH and 12:12 h L:D,
with the photophase and scotophase reversed with respect to the natural light cycle in order
to allow pheromone extraction and bioassays during the day. Larvae were fed with a special
artificial diet for lepidopterans (Cibrián-Tovar & Sugimoto 1992). Adults were fed with a
50% sucrose solution placed on cotton wool, which was replaced every 3rd day.
PREPARATION OF SEX PHEROMONE EXTRACT

The gland that produces the sex pheromone, situated between the seventh and ninth
abdominal segment (Rojas et al. 1995), was dissected from virgin 3-5 day old females that
presented sexual calling behavior during the final third of the scotophase (Rojas et al.
2006). Sixty glands were placed in 2 mL glass vials containing 1 mL of dichloromethane as
a solvent for 10 min. The supernatant was concentrated to 100 µL under a nitrogen flow.
Each extract had a concentration of 3 female equivalents (FE) for each 5 μL. The gland
extracts was stored at -4 ºC until chemical analysis or use in the bioassays.
ANALYSIS AND IDENTIFICATION OF VOLATILE COMPOUNDS
Two µL of extract were injected into a gas chromatograph (GC) (HP 6890) coupled
with a mass spectrometer (MS) (HP 5972) (Agilent, USA). The samples were analysed
using a non-polar HP 5MS column (30 m long, 250 µm internal diameter and 0.25 µm film
thickness; Agilent Technologies, Santa Clara, California). The initial oven temperature was
60 °C, increasing 15 °C/min until it reached 280 °C. The carrier gas was helium at a
constant flow of 1 mL/min. The injector temperature was 225 °C and the auxiliary was 280
°C; the injector functioned in splitless mode for 0.30 min. The MS functioned by electronic
ionization (70 EV), in SCAN mode and at a mass interval of 35 to 550 AMU. Compound
identification was carried out by considering retention time (RT) and by comparing mass
spectra to spectral libraries (NIST/EPA/NIH 2002) and synthetic standards. The standards
(Z)-9-tetradecenylacetate (Z9-14:Ac) and (Z)-9-tetradecenol (Z9-14:OH), with 98% purity
were obtained from Sigma Aldrich (Toluca, Mexico). In all of the extracts used in the
bioassays, the presence of Z9-14:Ac and Z9-14:OH in similar proportions to those reported
by Rojas et al. (2006) were verified.

WIND TUNNEL BIOASSAYS
In the wind tunnel bioassays, volatile chemicals from the glandular extract were
carried to the insects by a current of air (0.4 m/s) produced by a fan (Siemens 1RA3055-
4YK31, México). The experiments were conducted at 25 ± 3 °C, 60 ± 5% RH. In
preliminary studies the following concentrations were tested: 3, 1.2, 0.12, 0.012 and 0.0012
FE. The 3FE concentration caused the greatest attraction response and landing, so this was
used in both experiments (wind tunnel and EAG). 5 µL (3 FE) of glandular extract was
individually placed on filter paper (2.0 × 2.0 cm, Whatman no. 1), then left for 20 s at the
same height as the air flow entrance to allow the solvent to evaporate. A single male was
released at this height but at the opposite end of the tunnel. Each moth was tested for
response within 300 s and with one test per moth.
The following types of behavior were measured in the wind tunnel: no response,
short flight (flight less than 100 cm), long flight (flight more than 100cm but less than 150
cm), and landing on the source. The number of males that displayed their claspers was also
recorded. Furthermore, latency to activation (antenna and wing movement and displaying
claspers) and latency to landing on the scent source were recorded.
ELECTROANTENNOGRAPHY
The measurement and analysis of the EAG responses were conducted using Syntech
EAG equipment (Kirchzarteng, Germany). Once the antenna of the insect was dissected, it
was mounted between 2 silver (Ag) electrodes and conductor gel was added. The signal

generated by the antenna was transmitted to an IDAC-2 amplifier and was observed on a
monitor with software for EAG recording and analysis. Antenna stimulation continued
for1s with an air flow of 0.5 L/min transported to the antenna by a pump (stimulus
controller SC-55) with a constant flow of humidified purified air (0.7 L/min). For stimulus
application, 5 µL (3 FE) of glandular extract of the sex pheromone were placed on filter
paper (1 × 0.5 cm, Whatman No. 1). In each experiment the solvent was allowed to
evaporate for 20 s and the interval between each stimulus was 120 s. In these bioassays, the
antenna polarization response to solvent was subtracted from response to glandular extract
before analysis. Each antenna was used only once.
TREATMENTS
To study the effect of age, the pupae were separated by sex. After emergence, adults
were placed in plastic containers (20 x 20 x 20 cm) to obtain single sex groups of known
age. In the wind tunnel bioassays, virgin males that were 2-3, 4-5, 6-7, 8-9 and 10-11 days
old were used, with 10 moths tested per group. For EAG response, virgin males that were
3, 6 and 9 days old were tested, with 6 moths tested for each age. Selection of the age
groups for EAG tests was based on results of wind tunnel tests.
For tests of the effect of mating, virgin males and females, 4-5 days after
emergence, were individually paired and held in plastic containers (200 mL). The pairs
were observed for mating and time when mating ended was recorded. Male responses in
wind tunnel bioassays and EAG to the extracts were measured 24 and 48 h after mating. All
of the evaluated mated males in both wind tunnel (n = 10) and EAG (n = 6) were 5-7 days
old. The depolarization values obtained from the groups of mated males were compared

against a group of virgin 4-6 day old males. The females from each mated pair were
dissected and the presence of the spermatophore used to confirm that matings were
successful.
STATISTICAL ANALYSIS
The behaviors of the males in the wind tunnel were analysed by the Chi-square
test with Yates correction. Regarding the latency of the responses observed in the wind
tunnel, the median and interquartile range (Q1 to Q3) were determined for each group. The
interquartile range is little affected by extreme scores, so it is a good measure of spread for
skewed distributions. The latency data were analysed by a Kruskal-Wallis test and Tukey’s
test (P = 0.05) was used for comparison of means.
The data obtained from the depolarization in the male antennae were analysed by
ANOVA. Comparisons of means were carried out using a Tukey's test. Sigma Plot 11
(Systat Software Inc., Chicago, Illinois) was used for all statistical analyses.
Results _____
EFFECT OF AGE
Male attraction to the extract was greatest for 4-5 and 6-7 day old males and then
decreased with age in wind tunnel tests (Table 1). These groups were the most responsive,
although there was no difference between these 2 groups. There were also significant

differences between 4-5, 6-7 and 2-3 (Chi square = 52.769; df = 1; P < 0.001), 8-9 (Chi
square = 52.769; df = 1; P < 0.001) and 10-11 day old males (Chi square = 72.521; df = 1;
P < 0.001) regarding flight behavior patterns when landing on the scent source. In
addition, there were significant differences between 4-7 and 2-3 day old (Chi square =
20.056; df = 1; P < 0.001), 8-9 and 10-11 day old males (Chi square = 8.526; df = 1; P =
0.004) for displaying claspers (Table 1).
The 4-5 and 6-7 day old males presented activation and landing latencies
significantly less than those displayed by 2-3, 8-9 and 10-11 day old males (H = 28.557, df
= 4, P < 0.001 and H = 24.167, df = 4, P < 0.001; respectively) (Fig.1).
The EAG responses by age group are shown in Fig. 2. The extract elicited a
significantly greater depolarization in antennae of 6 day old males than in 3 and 9 day old
males (F = 9.965; df = 2,17; P = 0.002).
EFFECT OF MATING
In the wind tunnel bioassays, the virgin males were more attracted (both long flight
and landing) than males evaluated 24 and 48 h after mating (AM). Differences were found
in source landing behavior for 24 AM males (Chi square =18.484; df = 1; P < 0.001) and
48 AM males (Chi square = 48.505; df = 1; P < 0.001); as well as in long flight behavior
for 24 AM males (Chi square = 11.281; df = 1; P < 0.001) and 48 AM males (Chi square
=36.214; df = 1; P < 0.001) (Table 2).
Mating had an effect on both activation and landing latencies (activation latency H
= 17.867, df = 2, P < 0.001; landing latency H = 14.885, df = 2, P < 0.001). The males from
the 24 AM and 48 AM groups spent more time in activation and landing on the scent

source than the virgin males group (Fig. 3). Similarly, a significantly greater depolarization
was observed EAG responses of virgin males than mated males (24 AM and 48 AM
groups) (F = 9.814; df = 2, 17; P = 0.002) (Fig. 4).
Discussion ___
In this study, we found that antennal sensitivity to the sex pheromone by male C.
decolora moths varies in relation to age, reaching a maximum at 6 days. Similar changes in
antennal sensitivity due to age have been observed in other species of moths such as A.
ipsilon (Gadenne et al. 1993), Spodoptera litoralis (Boisduval) (Martel et al. 2009) and
Mamestra brassica (L.) (Pophof 2000). In these species, this has been attributed to the
maturation of the ORNs (Martel et al. 2009), changes in hemolymph titers of the juvenile
hormone (Gadenne et al. 1993; Gadenne & Anton 2000) and neuromodulation of biogenic
amines that act on ORNs sensitivity (Pophof 2000; Grosmaitre et al. 2001). These 3
mechanisms could explain the results observed in C. decolora as well; however studies are
needed on these variables for this species.
In C. decolora, the observed changes in antennal sensitivity to the sex pheromone
correspond to observed behavioral changes. During the initial days after emergence, male
attraction response is low and it then begins to increase with age until reaching its
maximum level at days 4-5 and 6-7, subsequently decreasing with age. In some species of
moths, such as A. ipsilon, male response to the sex pheromone starts to increase in 3 day
old individuals (Gadenne et al. 1993). In other moths such as Spodoptera frugiperda
(Smith), the optimal age is 2 days (Rogers & Marti 1994), while for Spodoptera exigua

(Hübner) it is reached at 1-2 days (Rogers & Marti 1996). It seems that young C. decolora
males (2-3 days old) do not have sufficient sexual maturity to respond to the sex
pheromone, and this also seems to be the case regarding recently emerged A. ipsilon males
(Gadenne et. al 1993; Duportets et al. 1998). Across species, the olfactory response
increases until the males reach sexual maturity, and then decreases (Evenden & Gries
2008).
Mating in C. decolora males was associated with a decrease in behavioral
responses, similar to those observed in A. ipsilon mated males (Gadenne et al. 2001). In A.
ipsilon, mating triggers the release of neuromodulators that have the ability to modulate
antennal lobe neurons in response to female sex pheromones and affect the development of
the sex accessory glands, but antennal sensitivity is not affected (Duportets et al. 1998;
Gadenne et al. 2001). In contrast, in C. decolora the antennal sensitivity is affected by
mating. The decrease in pheromone response could be related to the fact that C. decolora
males do not mate more than once during the night (Rojas & Cibrián-Tovar 1994) and can
mate a total of 2 to 3 times during their lifetime (Castrejón-Gómez et al. 2000). This
inhibition in the antennal and behavioral responses, combined with the effect of age, could
represent a mechanism for temporally separated matings.
Copitarsia decolora males possibly need to recover from increased energy
expenditure during mating and store sperm again (Gillott 2003). During this period of
inhibition, the males can focus on searching for food to re-establish reserves in their
accessory glands to allow sperm production and therefore mate again, as observed in A.
ipsilon (Barrozo et al. 2011).
The attraction response of male C. decolora to pheromone not only depends on the
chemical composition (type of compounds and their relative proportions) (Rojas et al.

2006), but is also influenced by insect age and mating status, which affect the sensory input
or processing that generates the necessary locomotion pattern to land on the stimulation
source (Daly et al. 2007; Anton et al. 2007; Lemmen & Evenden 2009). In C. decolora
males, neural plasticity induced by the physiological state allows males to modulate their
response to the sex pheromone, which must be considered before that synthetic pheromone
is used to manage this insect.
Acknowledgments ___
R. Arzuffi and N. Robledo are COFAA and EDI, IPN fellows. H. Reyes would like
to thank CONACYT and IPN for a scholarship. We are also very grateful to SIP, IPN
(Projects 20121192 and 20130883).
References Cited _
Anton S, Gadenne, C. 1999. Effect of juvenile hormone on the central nervous processing
of sex pheromone in an insect. Proceedings of the National Academy of Science,
USA 96: 5764-5767.
Anton S, Dufour MC, Gadenne C. 2007. Plasticity of olfactory guide behaviour and its
neurobiological basis: lessons from moths and locusts. Entomologia Experimentalis
et Applicata 123:1-11.

Barrientos-Hernández I, Hernández-Ramírez G, Cibrián-Tovar J. 2011. Tasa de liberación
de la feromona sexual de Copitarsia decolora (Lepidoptera: Noctuidae) y capturas
de machos con trampas en brócoli. Entomotropica 26: 47-58.
Barrozo RB, Jarriault D, Simeone X, Gaertner C, Gadenne C, Anton S. 2010a. Mating-
induced transient inhibition of responses to sex pheromone in a male moth is not
mediated by octopamine or serotonin. Journal of Experimental Biology 213: 1100-
1106.
Barrozo RB, Gadenne C, Anton S. 2010b. Switching attraction to inhibition: Mating-
induced reversed role of sex pheromone in an insect. Journal of Experimental
Biology 213: 2933-2939.
Barrozo RB, Jarriault D, Deisig N, Gemeno C, Monsempes C, Lucas P, Gadenne C, Anton
S. 2011. Mating-induced differential coding of plant odour and sex pheromone in a
male moth. European Journal of Neuroscience 33: 1841-1850.
Castrejón-Gómez VR, Cibrián-Tovar J, Romero-Napoles J, Camino-Lavin M. 2000.
Mating frequency in wild females of Copitarsia consueta (Lepidoptera: Noctuidae).
Florida Entomologist 83: 374-376.
Cibrián-Tovar J, Sugimoto A. 1992. Elaboración de una dieta artificial para la cría de
Copitarsia consueta Walker (Lepidoptera:Noctuidae), pp. 416 In Martínez CG,
Lastras AR, Valle JM., Medellín PM., Hernández JG., Loza RJM., García RL
[eds.], Memorias del XXVII Congreso Nacional de Entomología. Sociedad
Mexicana de Entomología. México.
Daly KC, Carrell LA, Mwilaria E. 2007. Detection versus perception: physiological and
behavioral analysis of olfactory sensitivity in the moth (Manduca sexta). Behavioral
Neuroscience 121: 794-807.

De Belle JS, Kanzaki R. 1999. Protocerebral olfactory processing. pp. 243–282 In Hansson
BS [ed.], Insect Olfaction. Springer, Berlin, Germany.
Diaz-Gomez O, Malo EA, Patiño-Arellano SA, Rojas JC. 2012. Pheromone trap for
monitoring Copitarsia decolora (Lepidoptera:Noctuidae) activity in cruciferous
crops in Mexico. Florida Entomologist 95: 602-609.
Dumont S, McNeil JN. 1992. Responsiveness of Pseudaletia unipuncta (Lepidoptera:
Noctuidae) males, maintained as adults under different temperature and
photoperiodic conditions, to female sex pheromone. Journal of Chemical Ecology
18: 1797-1807.
Duportets L, Dufour MC, Couillaud F, Gadenne C. 1998. Biosynthetic activity of corpora
allata growth of sex accessory glands and mating in the male moth Agrotis ipsilon.
Journal of Experimental Biology 201: 2425-2432.
Evenden ML, Gries R. 2008. Plasticity of male response to sex pheromone depends on
physiological state in a long-lived moth. Animal Behavior 75: 663-672.
Gadenne C, Anton S. 2000. Central processing of sex pheromone stimuli is differentially
regulated by juvenile hormone in a male moth. Journal of Insect Physiology 46:
1195-1206.
Gadenne C, Renou M, Sreng L. 1993. Hormonal control of pheromone responsiveness in
the male black cutworm Agrotis ipsilon. Experientia 49: 721-724.
Gadenne C, Dufour MC, Anton S. 2001. Transient post-mating inhibition of behavioural
and central nervous responses to sex pheromone in an insect. Proceedings of the
Royal Society of London B 268: 1631-1635.
Gillott C. 2003. Male accessory gland secretions: Modulators of female reproductive
physiology and behavior. Annual Review of Entomology 48: 163-184.

Grosmaitre X, Marion-Poll F, Renou M. 2001. Biogenic amines modulate olfatory receptor
neurons firing activity in Mamestra brassicae. Chemical Senses 26: 653-661.
Hansson BS, Anton S. 2000. Function and morphology of the antennal lobe: New
developments. Annual Review of Entomology 45: 203–231.
Keil TA. 1999. Morphology and development of the peripheral olfactory organs. pp. 5-48
In Hansson BS [ed.], Insect Olfaction. Springer, Berlin, Germany.
Lemmen J, Evenden M. 2009. Peripheral and behavioral plasticity of pheromone response
and its hormonal control in a long-lived moth. Journal of Experimental Biology 212:
2000-2006.
Martel V, Anderson P, Hansson BS, Schlyter F. 2009. Peripheral modulation of olfaction
by physiological state in the Egyptian leafworm Spodoptera littoralis
(Lepidoptera:Noctuidae). Journal of Insect Physiology 55: 793-797.
Muñiz-Reyes E, Cibrián-Tovar J, Rojas-Leon J, Díaz-Gómez O, Valdés-Carrasco J,
Bautista-Martínez N. 2007. Capturas de Copitarsia decolora (Lepidoptera:
Noctuidae) en trampas cebadas con diferentes proporciones de feromona sexual.
Agrociencia 41: 575-581.
NIST/EPA/NIH. 2002. Mass spectral library. Mass Spectral Library with Search Program
(Data Version: NIST05, Software Version 2.0), USA.
Pophof B. 2000. Octopamine modulates the sensitivity of silk moth pheromone receptor
neurons. Journal of Comparative Physiology A 186: 307-313.
Rogers CE, Marti OG. Jr. 1994. Reproductive potential of once-mated moths of fall
armyworm (Lepidoptera: Noctuidae). Florida Entomologist 77: 402-410.
Rogers CE, Marti OG. Jr. 1996. Beet armyworm (Lepidoptera: Noctuidae): Effects of age
at first mating on reproductive potential. Florida Entomologist 79: 343-352.

Rojas JC, Cibrián-Tovar J. 1994. Reproductive behavior of Copitarsia consueta
(Lepidoptera: Noctuidae): mating frequency, effect of age on mating, and influence
of delayed mating on fecundity and egg fertility. Pan-Pacific Entomologist 70: 276-
282.
Rojas JC, Valdez-Carrasco J, Cibrian-Tovar J. 1995. Morphological evidence for possible
sites of production of the female sex pheromone of Copitarsia consueta
(Lepidoptera: Noctuidae). Florida Entomologist 78: 360-363.
Rojas JC, Cruz-Lopez C, Malo EA, Díaz-Gómez O, Calyecac G, Cibrian-Tovar J. 2006.
Identification of the sex pheromone of Copitarsia decolora. Journal of Economic
Entomology 99: 797-802.
Soques S, Vazquez GM, Grozinger CM, Gould F. 2010. Age and mating status do not
affect transcript levels of odorant receptor genes in male antennae of Heliothis
virescens and Heliothis subflexa. Journal of Chemical Ecology 36: 1226-1233.
Suarez-Vargas AD, Bautista-Martinez N, Valdez-Carrasco J, Angulo-Ormeño A, Alatorre-
Rosas R, Vera-Graziano J, Equihua-Martinez A, Manuel-Pinto V. 2006. Fluctuación
poblacional de Copitarsia decolora (Gueéne) y su asociación con crucíferas
comerciales. Agrociencia 40: 501-509.
Venette CR, Gould JR. 2006. A pest risk assessment for Copitarsia spp.: Insects associated
with importation of commodities into the United States. Euphytica 148: 165-183.
Zhang ZC, Wang MQ, Zhang G. 2009. Molecular cloning and expression of pheromone-
binding protein1 from the diamondback moth, Plutella xylostella. Entomologia
Experimentalis et Applicata 133: 136-145.

Table 1. Response (percentage) by age group of virgin Copitarsia decolora males to
female sex pheromone extract (3fe) in wind tunnel bioassays.
Behavior
Age (days)
2-3 4-5 6-7 8-9 10-11
No response 20 b 0 a 0 a 10 b 50 b
Short flight 30 b 0 a 10 a 30 b 10 a
Long flight 10 a 10 a 10 a 20 a 10 a
Landed 40 a 90 b 80 b 40 a 30 a
Claspers displayed 80 a 100 b 100 b 90 a 90 a
Percentages followed by the same letter within a behavior are not significantly different
(Chi-square, n = 10, P < 0.05).

Table 2. Response (percentage) of virgin and mated Copitarsia decolora males to female
sex pheromone extract (3FE) in wind tunnel bioassays. Mated males were tested 24 h and
48 h after mating (AM).
Behavior
Condition
Virgin Mated, 24 h AM Mated, 48 h AM
Short flight 10 a 20 a 20 a
Long flight 10 a 30 b 50 c
Landing 80 c 50 b 30 a
Claspers displayed 100 a 100 a 100 a
Percentages in each row followed by the same letter within a behavior are not significantly
different (Chi-square, n = 10, P < 0.05).
FIGURE CAPTIONS
Fig. 1. Activation latencies (black bars) and landing latencies (gray bars) (Q1 < Median
< Q2) by age group of virgin males in response to a glandular extract of female sex
pheromone (3FE) in wind tunnel bioassays. Bars within a behavior headed by the same
letter are not significantly different (Tukey's mean separation test, n = 10, P < 0.05).
Fig. 2. Depolarization (mean ± SEM) of virgin male antennae in response to a glandular
extract of female sex pheromone (3FE) by male age. Bars headed by the same letter are not
significantly different (Tukey's mean separation test, n = 6, P < 0.05).

Fig.3. Activation latencies (black bars) and landing latencies (gray bars) (Q1 < Median <
Q2) of virgin and mated Copitarsia decolora males to female sex pheromone extract (3FE)
in wind tunnel bioassays. Mated males were tested 24 h and 48 h after mating (24 AM and
48 AM, respectively). Bars within a behavior headed by the same letter are not significantly
different (Tukey's mean separation test, n = 10, P < 0.05).
Fig. 4. Depolarization (mean ± SEM) of antennae in response to a glandular extract of
female sex pheromone (3FE) of virgin and mated males. Mated males were tested 24 h and
48 h after mating (24 AM and 48 AM, respectively). Bars headed by the same letter are not
significantly different (Tukey's mean separation test, n = 6, P < 0.05).

Fig. 1. Activation latencies (black bars) and landing latencies (gray bars) (Q1 < Median <
Q2) by age group of virgin males in response to a glandular extract of female sex
pheromone (3FE) in wind tunnel bioassays. Bars within a behavior headed by the same
letter are not significantly different (Tukey's mean separation test, n = 10, P < 0.05).

Fig. 2. Depolarization (mean ± SEM) of virgin male antennae in response to a glandular
extract of female sex pheromone (3FE) by male age. Bars headed by the same letter are not
significantly different (Tukey's mean separation test, n = 6, P < 0.05).

Fig.3. Activation latencies (black bars) and landing latencies (gray bars) (Q1 < Median <
Q2) of virgin and mated Copitarsia decolora males to female sex pheromone extract (3FE)
in wind tunnel bioassays. Mated males were tested 24 h and 48 h after mating (24 AM and
48 AM, respectively). Bars within a behavior headed by the same letter are not significantly
different (Tukey's mean separation test, n = 10, P < 0.05).
Fig. 4. Depolarization (mean ± SEM) of antennae in response to a glandular extract of
female sex pheromone (3FE) of virgin and mated males. Mated males were tested 24 h and
48 h after mating (24 AM and 48 AM, respectively). Bars headed by the same letter are not
significantly different (Tukey's mean separation test, n = 6, P < 0.05).
Recommended

















