
• Frecuencias génicas y genotípicas
• Equilibrio de Hardy-Weinberg
• Factores que alteran el equilibrio

• La aplicación de los principios de las genética mendeliana
a las poblaciones, sirvieron para explicar tanto el origen de
la variación (mutación), como el mecanismo de herencia
(segregación de alelos).
• Consideramos la evolución como una consecuencia de los
cambios en los alelos y en las frecuencias alélicas de las
poblaciones a lo largo del tiempo.
• Esta unión de la genética de poblaciones con la teoría de la
selección natural dio lugar a una nueva visión del proceso
evolutivo.
Inicio de la genética de poblaciones
2

• Además de la selección natural, otras fuerzas —la mutación, la
migración y la deriva— alteran individual y colectivamente las
frecuencias alélicas, y provocan una divergencia evolutiva que puede
terminar conduciendo a la especiación o la formación de nuevas
especies.
• Las poblaciones genéticamente diferenciadas pueden continuar
existiendo, extinguirse, volver a unirse unas con otras o continuar
divergiendo hasta que se vuelven reproductivamente aisladas.
• Las poblaciones reproductivamente aisladas se consideran especies
separadas.
• Los cambios genéticos dentro de las poblaciones pueden modificar
una especie a lo largo del tiempo, transformarla en otra especie
• o hacer que se divida en dos o más especies.
Inicio de la genética de poblaciones
3

Especiación
4

La variación genética está presente en la mayoría de poblaciones y especies
• Una población es un grupo de individuos que pertenece a la misma
especie, que viven en un área geográfica definida y que se emparejan
o pueden potencialmente llegar a hacerlo.
• La información genética portada por los miembros de una población
constituye el acervo génico de esa población.
• Una población que esté bien adaptada a su entorno debe ser
altamente homocigótica
• El alelo más favorable de cada locus estará presente con una alta
frecuencia
5

Sin embargo:
La mayoría de las poblaciones de plantas y animales revela muchas
similitudes fenotípicas.
Una amplia serie de evidencias indica que, en realidad, la mayoría de las
poblaciones contienen un alto grado de heterocigosidad.
La variación genética está presente en la mayoría de poblaciones y especies
6

Detección de la variación genética mediante selección artificial
Un ejemplo muy evidente de este tipo de prueba es el perro doméstico.
La amplia variedad de tamaños, colores y comportamientos que podemos ver
en las distintas razas de perro son el resultado de los efectos de la selección
sobre la variación genética presente en los lobos silvestres. 7
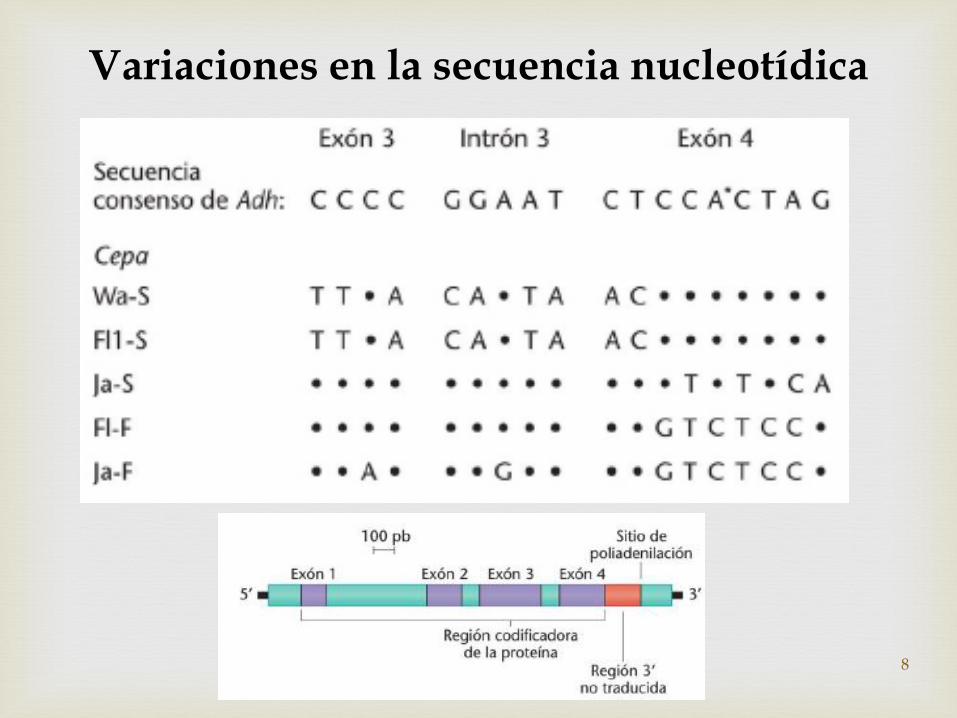
Variaciones en la secuencia nucleotídica
8

Explicación del alto nivel de variación genética en las poblaciones
El hallazgo de que las poblaciones albergan una considerable variación genética,
cambio la idea que la selección tendería a favorecer un único alelo óptimo en cada
locus y que, como resultado, las poblaciones tendrían altos niveles de
homocigosidad
La teoría neutralista de la evolución molecular.
• Postula que las mutaciones que conducen a sustituciones de aminoácidos suelen
ser perjudiciales, existiendo solo una fracción muy pequeña de ellas que pueden
ser favorables. Estas se preservan o se eliminan de la población,
respectivamente, mediante selección natural.
• Algunas mutaciones son neutrales, es decir, son funcionalmente equivalentes al
alelo al que sustituyen. La frecuencia de estos alelos neutrales en una población
estará determinada por la tasa de mutación y por la deriva génica aleatoria y no
por selección.9

La ley de Hardy-Weinberg describe las frecuencias alélicas y las frecuencias genotípicas de las
poblaciones
• Las poblaciones son dinámicas; se expanden y se contraen debido a las
tasas de natalidad y mortalidad, a las migraciones o al contacto con otras
poblaciones.
• A menudo, algunos individuos de una población tendrán más
descendientes que otros, contribuyendo con una mayor fracción de sus
alelos a la siguiente generación.
• Por tanto, la reproducción diferencial dentro de una población puede
llevar, con el tiempo, a cambios en las frecuencias alélicas y genotípicas
de las generaciones subsiguientes
10

La ley de Hardy-Weinberg describe las frecuencias alélicas y las frecuencias genotípicas de las poblaciones
El cálculo de las frecuencias alélicas y las frecuencias fenotípicas en la
población, y la determinación de cómo varían esas frecuencias entre esa
generación y la siguiente son dos elementos clave de la genética de
poblaciones.
La relación entre las proporciones relativas de alelos dentro del acervo
génico y las frecuencias de los diferentes genotipos de una población fue
descrita, mediante un modelo matemático desarrollado
independientemente por el matemático británico Godfrey H. Hardy y el
médico alemán Wilhelm Weinberg.
Este modelo, denominado ley de Hardy-Weinberg 11

La ley de Hardy-Weinberg describe las frecuencias alélicas y las frecuencias genotípicas de las poblaciones
Ley de Hardy-Weinberg, describe lo que pasa con los alelos y los genotipos, en
una población «ideal» que sea:
• Infinitamente grande• Que los emparejamientos que se produzcan sean aleatorios• Que no esté sujeta a ninguna fuerza evolutiva, como la mutación, la
migración o la selección.
Bajo estas condiciones, el modelo de Hardy-Weinberg hace dos predicciones:
1. Las frecuencias de los alelos en el acervo génico no varían con el
tiempo.
2. Si examinamos dos alelos de un locus, A y a, después de una generación
de emparejamiento aleatorio, las frecuencias de los genotipos AA:Aa:aa
dentro de la población pueden calcularse mediante la fórmula
p2 + 2pq + q2 = 1
donde p es la frecuencia del alelo A y q es la frecuencia del alelo a. 12

• Una población que satisfaga estos criterios
• Donde dos alelos de un locus concreto, den como resultado
las frecuencias genotípicas predichas
• Se dice que está en equilibrio de Hardy-Weinberg
La ley de Hardy-Weinberg describe las frecuencias alélicas y las frecuencias genotípicas de las poblaciones
El modelo de Hardy-Weinberg utiliza
• Los principios mendelianos de segregación,
• Junto con teoría simple de probabilidades
• Para explicar las relaciones entre las frecuencias alélicas y
genotípicas de una población 13

Demostración:• Consideremos un único locus autosómico con dos alelos A y
a• La frecuencia de A= 0.7 y a=0.3• Obsérvese que 0.7 + 0.3 = 1 (consideramos todos los alelos)• Los individuos se emparejan aleatoriamente• La P de un gameto femenino contenga A es 0.7 y La P de un
gameto masculino contenga A es 0.7• La P de que ambos gametos contengan A es 0.7 x 0.7 = 0.49
• El genotipo se presentara en 49%• La P de que se forme un cigoto
• Aa es de 0.7 x 0.3 = 0.21• aA es de 0.3 x 0.7 = 0.21• La frecuencia del genotipo será 0.21 + 0.21 = 0.42 = 42%
• La P que se forme un cigoto aa es 0.3 x 0.3 = 0.09 = 9%• Comprobando: 0.49 + 0.42 + 0.09 = 1
14

De acuerdo con las suposiciones de la ley de Hardy-Weinberg
• Todos los genotipos tienen idénticas tasas de supervivencia y de reproducción.
• Por tanto la siguiente generación, todos los genotipos contribuirán por igual al
nuevo acervo génico.
• Los individuos AA constituyen el 49% de la población, y mantendrán esta
proporción.
• los individuos Aa constituyen el 42% de la población, así que predecimos que
sus gametos constituirán el 42% del nuevo acervo génico.
• La mitad (0,5) de estos gametos portará el alelo A. Por tanto, la frecuencia
del alelo A en el acervo génico será 0,49 + 0,42(0,5) = 0,7.
• Los individuos aa constituyen el 9% (0.09) de la población, así que sus gametos
constituirán el 9% del nuevo acervo génico
• Por tanto el alelo a su frecuencia en la población será 0.09 + 0,42(0,5) = 0.3
• Los resultados comprueban el equilibrio 0.7 + 0.3 = 1
15

Calculo de las frecuencias genotípicas utilizando uncuadro
16

17

Las suposiciones en las que esta basada la población teórica descrita por la ley de Hardy-Weinberg
18
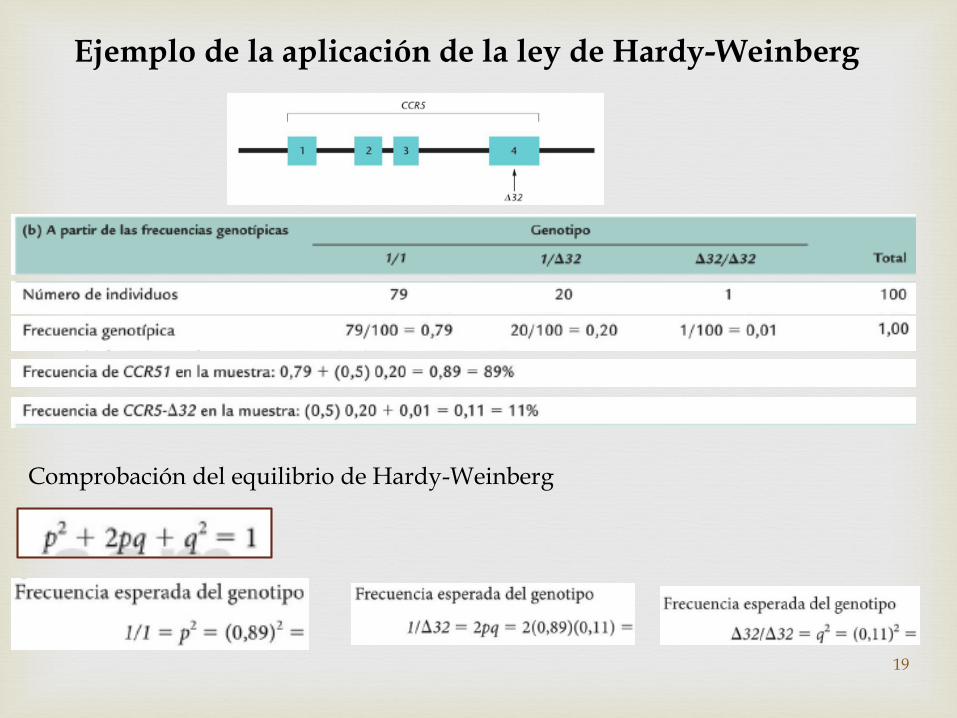
Ejemplo de la aplicación de la ley de Hardy-Weinberg
19
Comprobación del equilibrio de Hardy-Weinberg

Calculo de las frecuencias para múltiples alelos en poblaciones de Hardy-Weinberg
• En los humanos tenemos el grupos sanguíneo ABO
• El locus I, tiene tres alelos: IA, IB e i
• Produciendo seis combinaciones posibles: IAIA; IB i; ii; IAIB; IA i; IBIB.
En una población muestreado, se observaron las siguientes frecuencias de los grupos sanguíneos: A = 0.53, B = 0.13 y O = 0.26
20

Solución:Puesto que alelo i es recesivo, la frec. Del grupo sanguíneo O será:
r2 = 0.26; r = 0.26 ; r= 0.51
Utilizando r podemos calcular la Frec. Alélicas de IA e IB
La frecuencia del genotipo IA IA esta representada por p2 y la del genotipo IA i por 2pr.
Por tanto la frecuencia combinada del grupo sanguíneo A y O estará dada por:p2 + 2pr +r2 = 0.53 + 0.26
Factorizando
(p + r)2= 0.79; P + r = 0.79; P = 0.89 –r; P= 0.89 – 0.51 P = 0.38
Habiendo calculado p y r, que son las frecuencias del alelo IA e i, podemos calcular la frecuencia del alelo IB
p + q + r= 1; q = 1 – p –r; q = 1 -0.38 – 0.51; q = 0.11 21

22
Calculo de las frecuencias genotípicas
q

Calculo de la frecuencia de los heterocigóticos
La fibrosis quística, un carácter autosómico recesivo tiene una incidencia de
aproximadamente 1/2500 = 0,0004 en las personas de origen norte-europeo.
Su frecuencia dentro de la población esta representada por q2
Por tanto la frecuencia del alelo recesivo se calcula así:
q2= √0.0004
Puesto que p + q = 1, entonces la frecuencia de p es:
p = 1 – q
P = 1- 0.02 = 0.98
La frecuencia de los heterocigóticos es 2pq, por tanto2pq = 2(0.98)(0.02) = 0.04 o 4% o 1/25
23
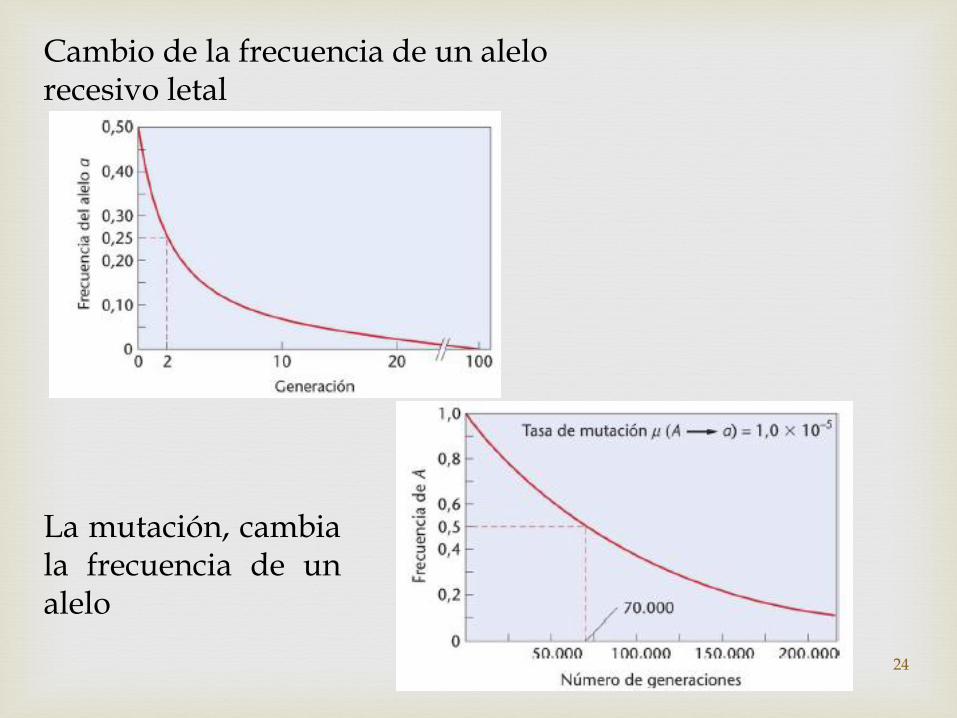
Cambio de la frecuencia de un alelo recesivo letal
La mutación, cambiala frecuencia de unalelo
24
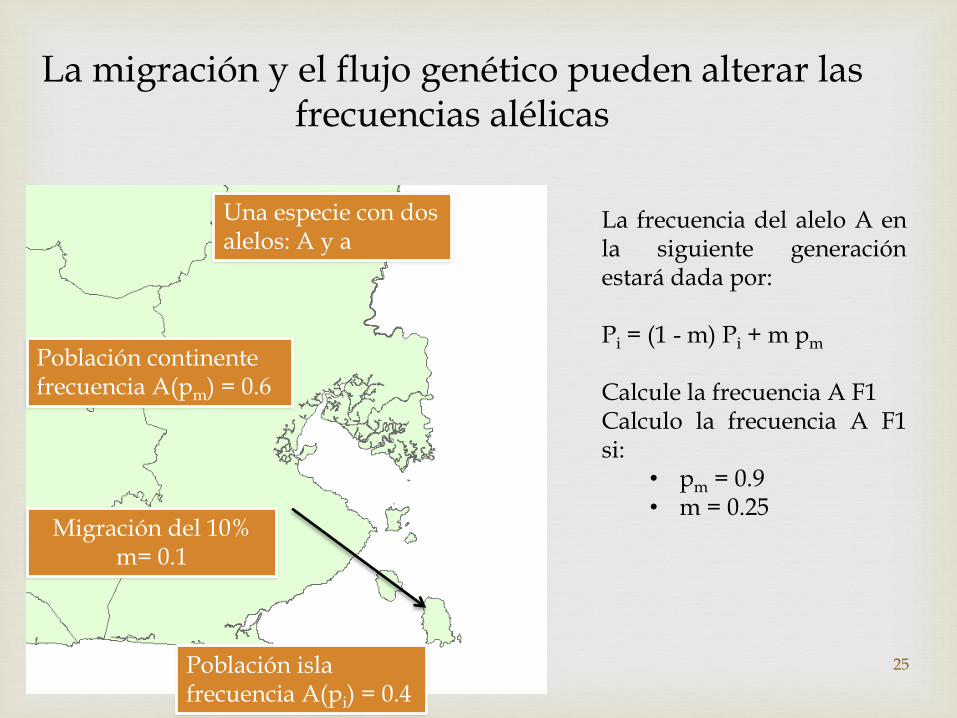
La migración y el flujo genético pueden alterar las frecuencias alélicas
Una especie con dos alelos: A y a
Población continente frecuencia A(pm) = 0.6
Población isla frecuencia A(pi) = 0.4
Migración del 10% m= 0.1
La frecuencia del alelo A enla siguiente generaciónestará dada por:
Pi = (1 - m) Pi + m pm
Calcule la frecuencia A F1Calculo la frecuencia A F1si:
• pm = 0.9• m = 0.25
25
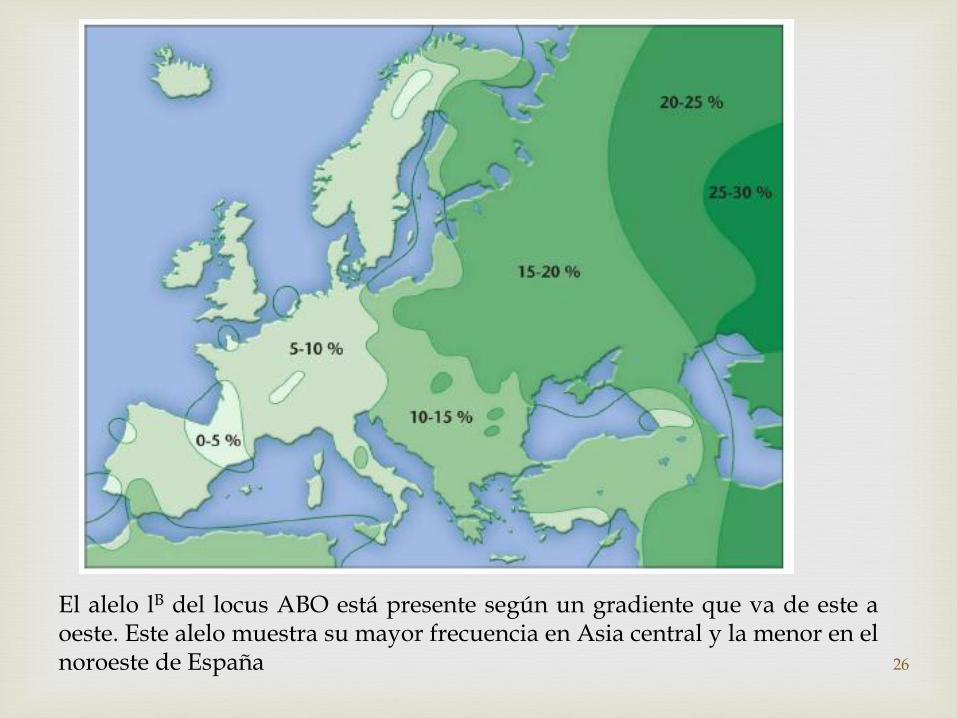
El alelo lB del locus ABO está presente según un gradiente que va de este aoeste. Este alelo muestra su mayor frecuencia en Asia central y la menor en elnoroeste de España 26

La deriva genética provoca cambios aleatorios en las frecuencias alélicas en las poblaciones pequeñas
• En las poblaciones de pequeño tamaño, pueden producirse
fluctuaciones aleatorias significativas de las frecuencias alélicas por
mero azar.
• El grado de fluctuación aumenta cuando el tamaño de la población
disminuye, una situación que se conoce con el nombre de deriva
genética.
• Además de por el pequeño tamaño de la población, la deriva puede
producirse debido al denominado efecto fundador
27

Efectos fundador en poblaciones humanas
Albinismo.
En la tribu de los Navajo, el albinismo se presenta en una frecuencia entre 1
de 1,500 - 2,000,
la población blanca (1 entre 36.000)
Los afroamericanos (1 entre 10.000)
Para indagar este fenómeno se investigaron distintas poblaciones nativas
de Norteamérica.
No se encontró el alelo de deleción en otras poblaciones (34 individuos)
Por tanto el alelo mutante pudo haber surgido de un único individuo que
formaba parte de los fundadores de la tribu navaja
28

29
Recommended

















