Embed Size (px)
Citation preview


FBW
3-11-2015
Wim Van Criekinge

BPC 2015
*** ERGRO ***

BPC 2015
*** ERGRO *** 1. Longest English word where first three
letters are identical to the last three
2. English word where longest stretch of letters
are identical at beginning and at the end
3. In Dutch ?
4. Any other language
5. Biological relevance ?
Send before 1st of december to
Longest one wins, if same size first to submit

Phylogenetics
Introduction
Definitions
Species concept
Examples
The Tree-of-life
Phylogenetics Methodologies
AlgorithmsDistance Methods
Maximum Likelihood
Maximum Parsimony
Rooting
Statistical Validation
Conclusions
Orthologous genes
Horizontal Gene Transfer
Phylogenomics
Practical Approach: PHYLIP
Weblems

Phylogeny (phylo =tribe + genesis)
Phylogenetic trees are about visualising evolutionary relationships. They reconstruct the pattern of events that have led to the distribution and diversity of life.
The purpose of a phylogenetic tree is to illustrate how a group of objects (usually genes or organisms) are related to one another
Nothing in Biology Makes Sense Except in the Light of Evolution. Theodosius Dobzhansky (1900-1975)
What is phylogenetics ?

Trees
• Diagram consisting of branches and nodes
• Species tree (how are my species related?)
– contains only one representative from each
species.
– all nodes indicate speciation events
• Gene tree (how are my genes related?)
– normally contains a number of genes from a
single species
– nodes relate either to speciation or gene
duplication events
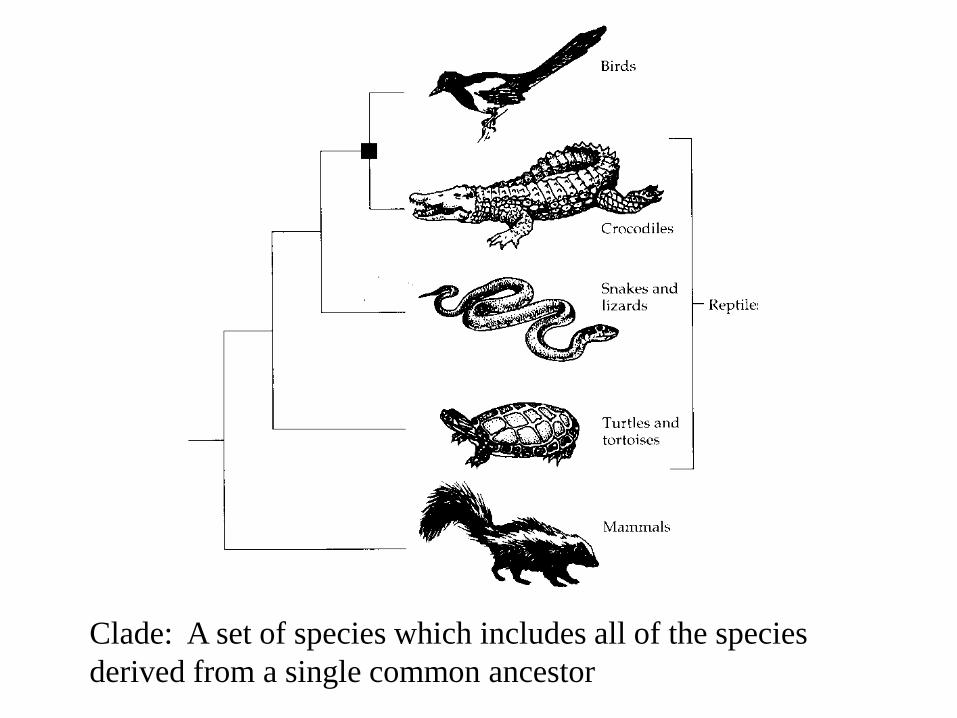
Clade: A set of species which includes all of the species
derived from a single common ancestor

Species Concepts from Various Authors
D.A. Baum and K.L. Shaw - Exclusive groups of organisms, where an exclusive group is one whose members are all more closely related to
each other than to any organisms outside the group.
J. Cracraft - An irreducible cluster of organisms, diagnosably distinct from other such clusters, and within which there is a parental pattern of
ancestry and descent.
Charles Darwin - "From these remarks it will be seen that I look at the term species, as one arbitrarily given for the sake of convenience to a set
of individuals closely resembling each other, and that it does not essentially differ from the term variety, which is given to less distinct and
more fluctuating forms. The term variety, again, in comparison with mere individual differences, is also applied arbitrarily, and for mere
convenience sake" (Origin of Species, 1st ed., p. 108).
T. Dobzhansky - The largest and most inclusive reproductive community of sexual and cross-fertilizing individuals which share a common gene
pool. And later...Systems of populations, the gene exchange between which is limited or prevented by reproductive isolating mechanisms.
M. Ghiselin - The most extensive units in the natural economy, such that reproductive competition occurs among their parts.
D.M. Lambert - Groups of individuals that define themselves by a specific mate recognition system.
J. Mallet - Identifiable genotypic clusters recognized by a deficit of intermediates, both at single loci and at multiple loci.
E. Mayr - Groups of actually or potentially interbreeding natural populations which are reproductively isolated from other such groups.
C.D. Michener - A group of organisms not itself divisible by phenetic gaps resulting from concordant differences in character states (except for
morphs - such as sex, age, or caste), but separated by such phenetic gaps from other such units.
H.E.H. Patterson - That most inclusive population of individual biparental organisms which share a common fertilization system.
G.G. Simpson - A lineage of populations evolving with time, separately from others, with its own unique evolutionary role and tendencies.
P.H.A. Sneath and R.R. Sokal - The smallest (most homogeneous) cluster that can be recognized upon some given criterion as being distinct
from other clusters.
A.R. Templeton - The most inclusive population of individuals having the potential for phenotypic cohesion through intrinsic cohesion
mechanisms (genetic and/or demographic - i.e. ecological -exchangeability).
E.O. Wiley - A single lineage of ancestor-descendant populations which maintains its identity from other such lineages and which has its own
evolutionary tendencies and historical fate.
S. Wright - A species in time and space is composed of numerous local populations, each one intercommunicating and intergrading with others.

Species
I. Definitions:
Species = the basic unit of classification
> Three different ways to recognize species:

Definitions:
> Three different ways to recognize species:
1) Morphological species = the smallest group that is
consistently and persistently distinct (Clusters in
morphospace)
species are recognized initially on the basis of
appearance; the individuals of one species look
different from the individuals of another
Plant Species

Definitions:
> Three different ways to recognize species:
2) Biological species = a set of interbreeding or
potentially interbreeding individuals that are
separated from other species by reproductive
barriers
species are unable to interbreed
Species

Definitions:
> Three different ways to recognize species:
3) Phylogenetic species = the boundary between
reticulate (among interbreeding individuals) and
divergent relationships (between lineages with no
gene exchange)
Species
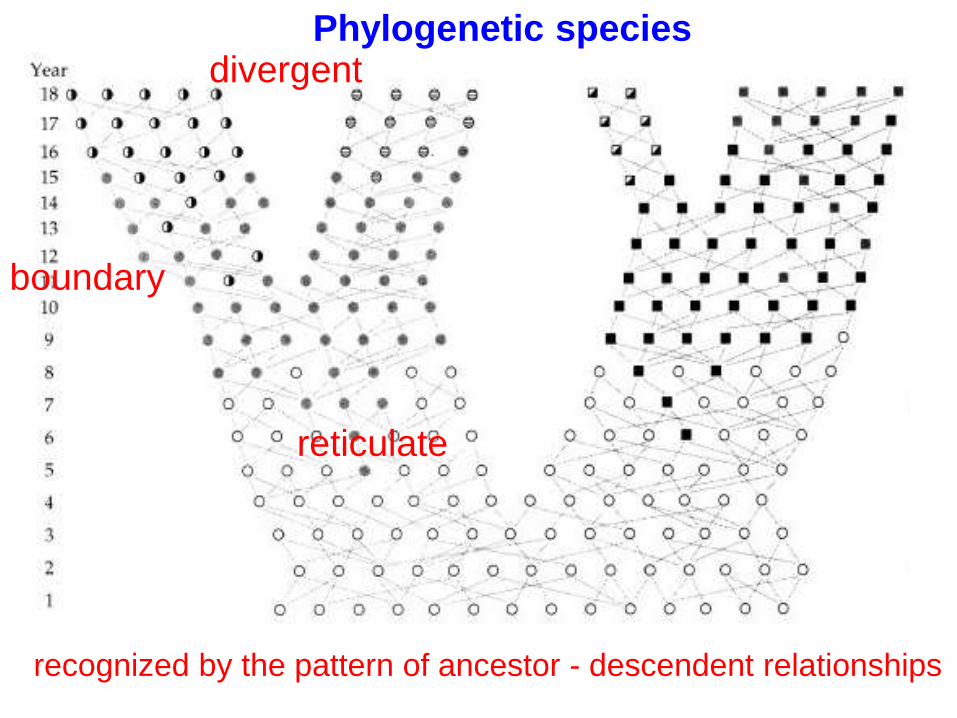
reticulate
divergentPhylogenetic species
recognized by the pattern of ancestor - descendent relationships
boundary

Definitions:
> Three different ways to recognize species:
4) Phylogenomics species = ability to transmit (and
maintain) a (stable) gene pool
Adresses the Anopheles genome topology
variations
Species

• In the tree to the left, A and B share the most recent
common ancestry. Thus, of the species in the tree,
A and B are the most closely related.
• The next most recent common ancestry is C with
the group composed of A and B. Notice that the
relationship of C is with the group containing A
and B. In particular, C is not more closely related to
B than to A. This can be emphasized by the
following two trees, which are equivalent to each
other:
Branching Order in a Phylogenetic Tree
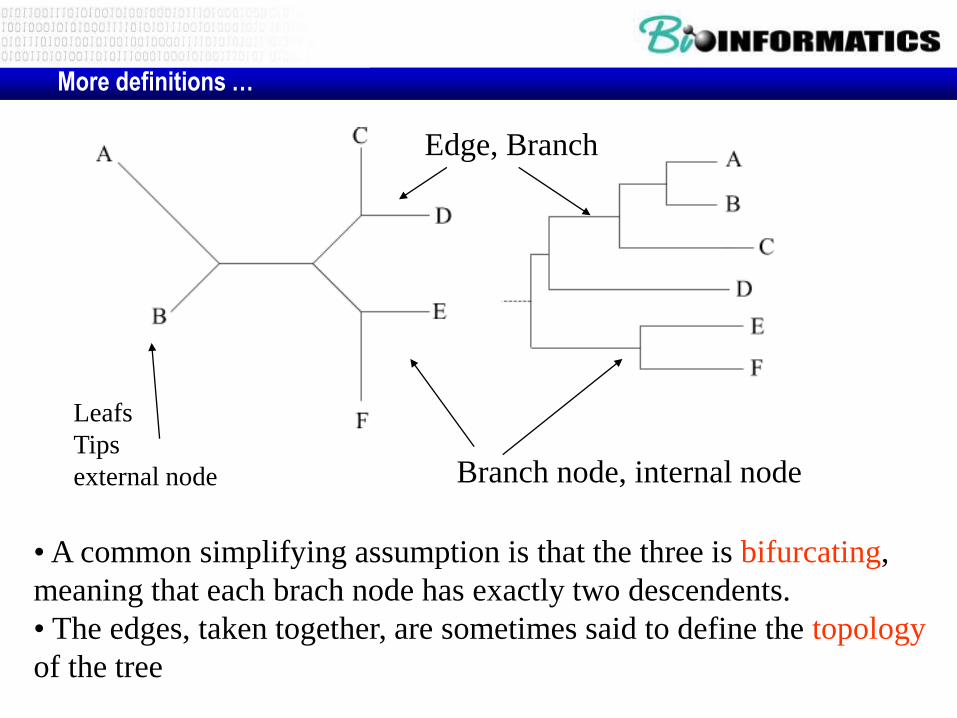
• A common simplifying assumption is that the three is bifurcating,
meaning that each brach node has exactly two descendents.
• The edges, taken together, are sometimes said to define the topology
of the tree
More definitions …
Branch node, internal node
Edge, Branch
Leafs
Tips
external node
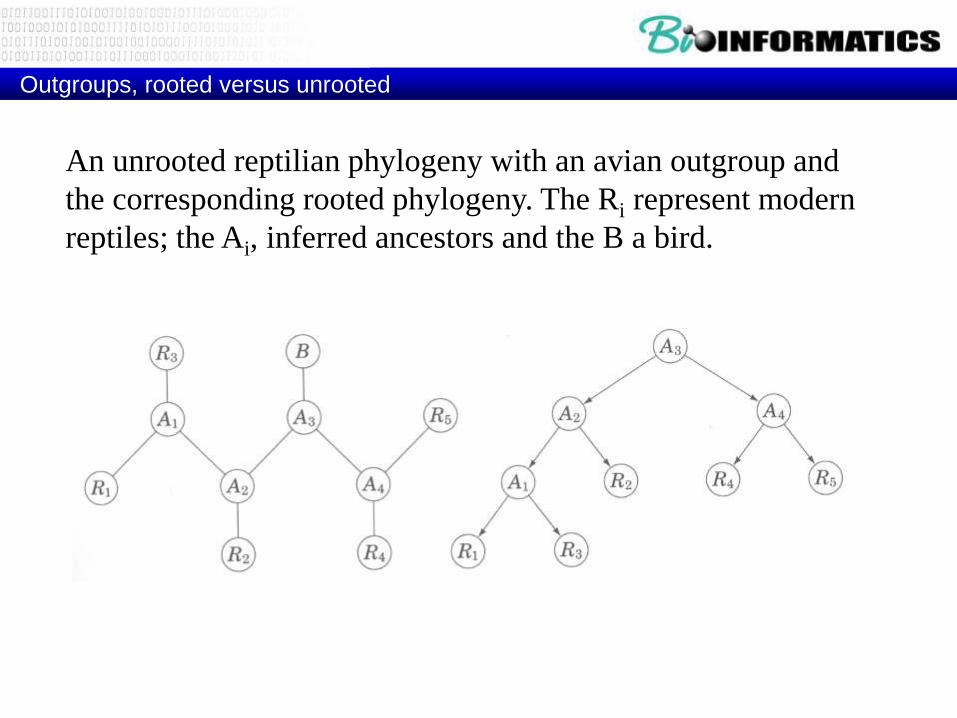
Outgroups, rooted versus unrooted
An unrooted reptilian phylogeny with an avian outgroup and
the corresponding rooted phylogeny. The Ri represent modern
reptiles; the Ai, inferred ancestors and the B a bird.
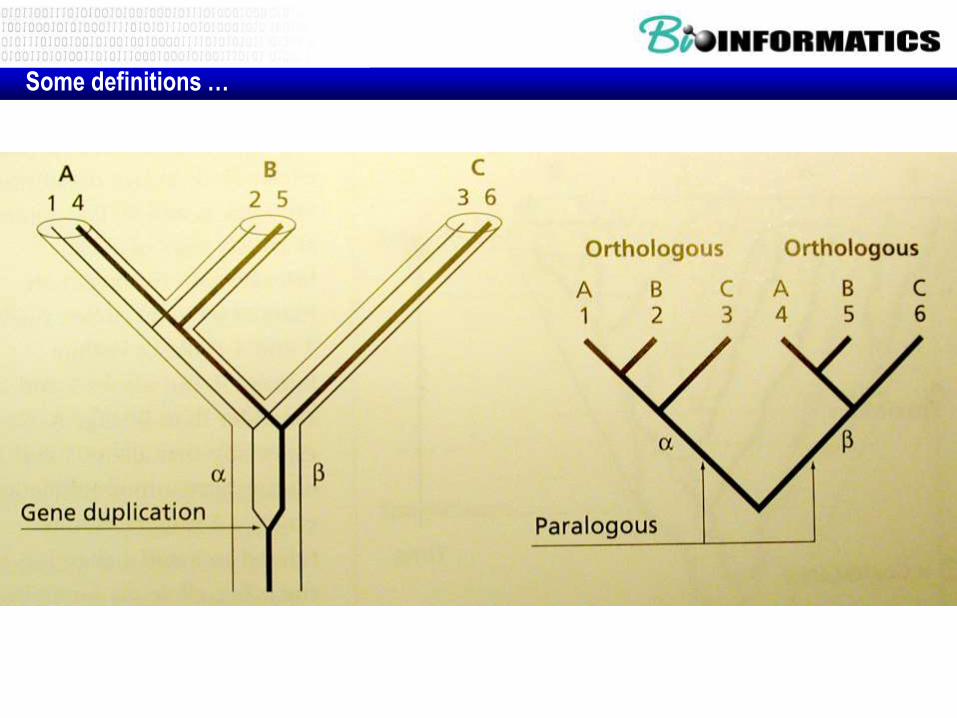
Some definitions …

Phylogenetic methods may be used to
solve crimes, test purity of products, and
determine whether endangered species
have been smuggled or mislabeled:
– Vogel, G. 1998. HIV strain analysis debuts in
murder trial. Science 282(5390): 851-853.
– Lau, D. T.-W., et al. 2001. Authentication of
medicinal Dendrobium species by the internal
transcribed spacer of ribosomal DNA. Planta
Med 67:456-460.
Examples


– Epidemiologists use phylogenetic methods to
understand the development of pandemics,
patterns of disease transmission, and
development of antimicrobial resistance or
pathogenicity:
• Basler, C.F., et al. 2001. Sequence of the 1918
pandemic influenza virus nonstructural gene (NS)
segment and characterization of recombinant viruses
bearing the 1918 NS genes. PNAS, 98(5):2746-2751.
• Ou, C.-Y., et al. 1992. Molecular epidemiology of HIV
transmission in a dental practice. Science
256(5060):1165-1171.
• Bacillus Antracis:
Examples

• Conservation biologists may use these techniques to determine which populations are in greatest need of protection, and other questions of population structure: – Trepanier, T.L., and R.W. Murphy. 2001. The Coachella Valley
fringe-toed lizard (Uma inornata): genetic diversity and phylogenetic relationships of an endangered species. Mol Phylogenet Evol 18(3):327-334.
– Alves, M.J., et al. 2001. Mitochondrial DNA variation in the highly endangered cyprinid fish Anaecypris hispanica: importance for conservation. Heredity 87(Pt 4):463-473.
• Pharmaceutical researchers may use phylogenetic methods to determine which species are most closely related to other medicinal species, thus perhaps sharing their medicinal qualities: – Komatsu, K., et al. 2001. Phylogenetic analysis based on 18S
rRNA gene and matK gene sequences of Panax vietnamensisand five related species. Planta Med 67:461-465.
Examples
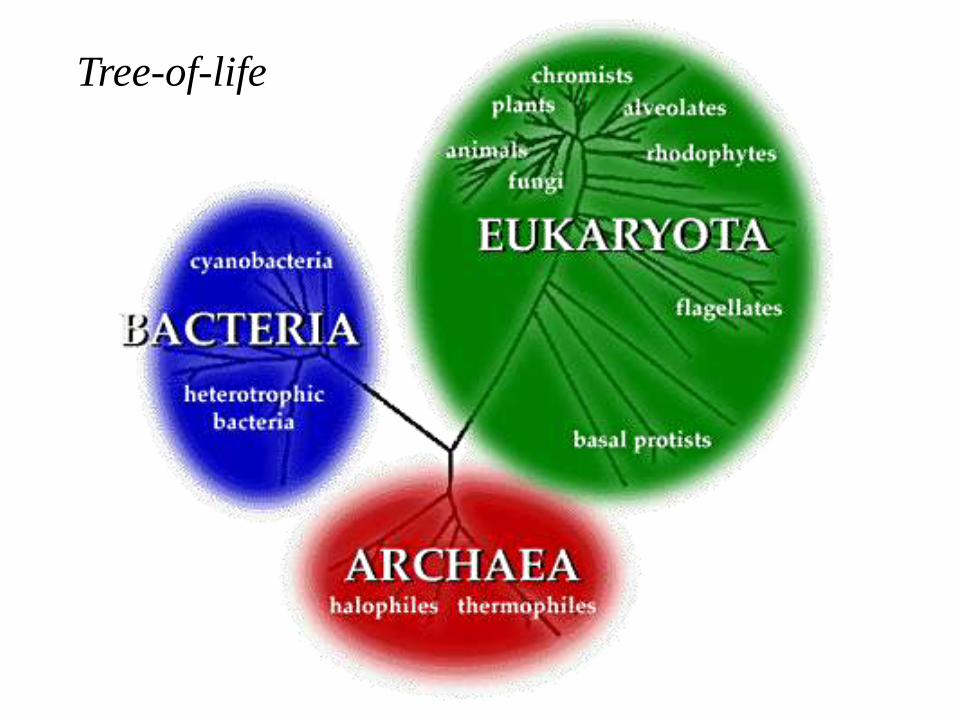
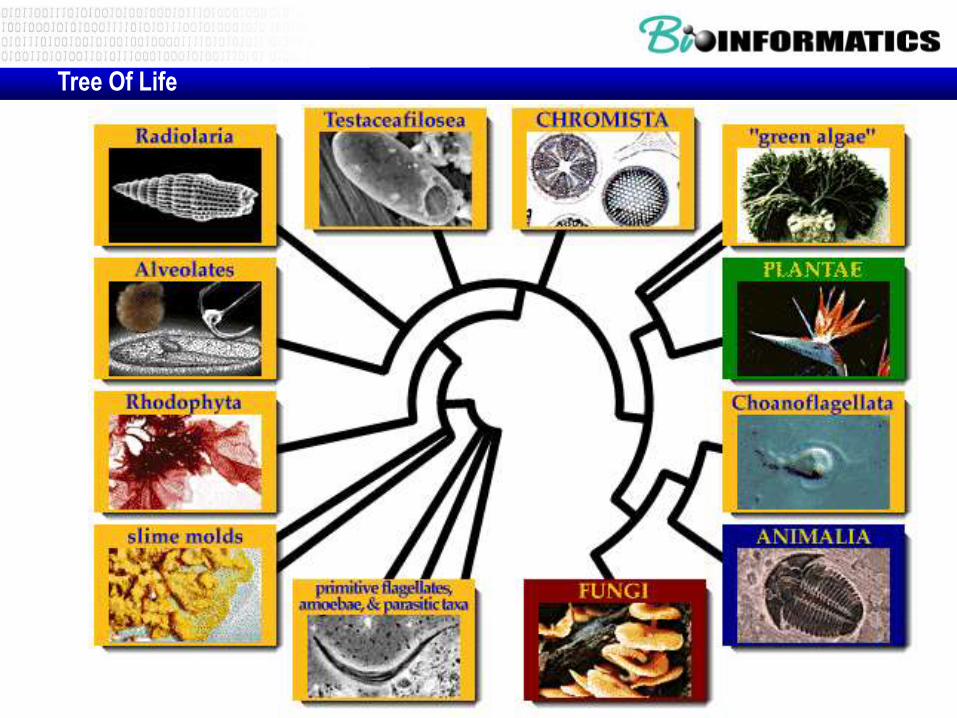
Tree-of-life

Origin of the Universe 15 billion yrs
Formation of the Solar System 4.6 "
First Self-replicating System 3.5 "
Prokaryotic-Eukaryotic Divergence 2.0 "
Plant-Animal Divergence 1.0 "
Invertebrate-Vertebrate Divergence 0.5 "
Mammalian Radiation Beginning 0.1 "
Some Important Dates in History
Tree Of Life
Tree Of Life
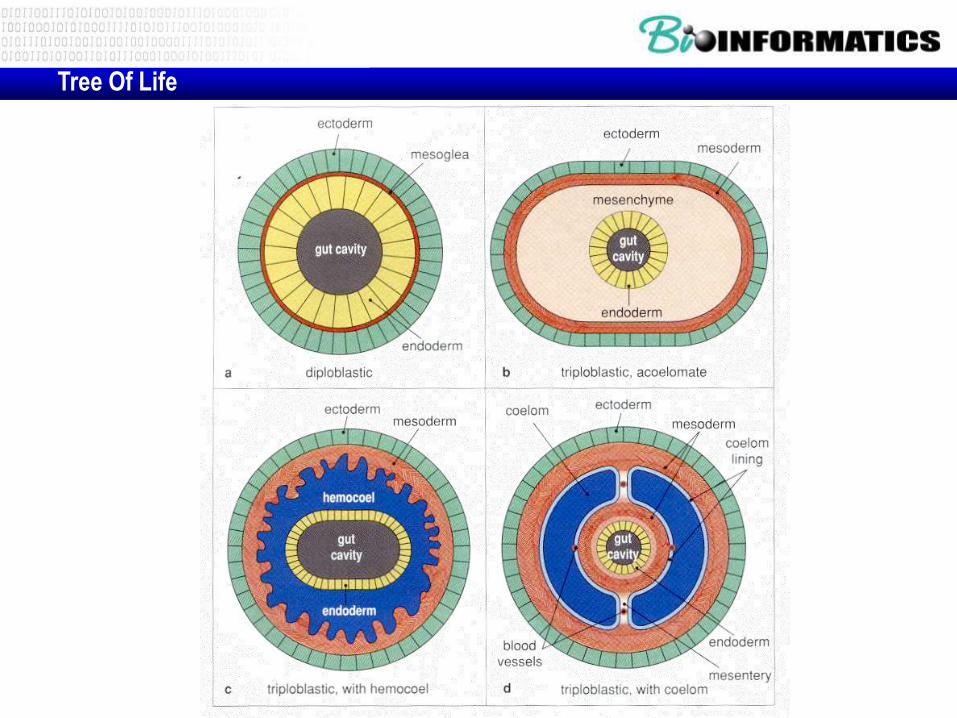
Tree Of Life
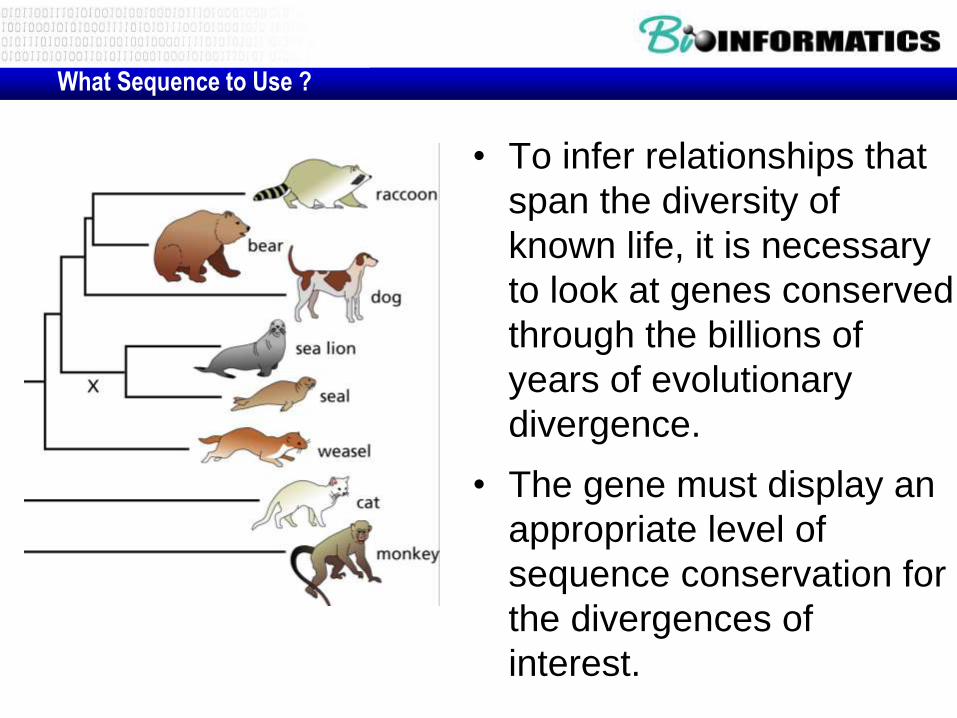
What Sequence to Use ?
• To infer relationships that
span the diversity of
known life, it is necessary
to look at genes conserved
through the billions of
years of evolutionary
divergence.
• The gene must display an
appropriate level of
sequence conservation for
the divergences of
interest.
.

• If there is too much change, then
the sequences become
randomized, and there is a limit to
the depth of the divergences that
can be accurately inferred.
• If there is too little change (if the
gene is too conserved), then
there may be little or no change
between the evolutionary
branchings of interest, and it will
not be possible to infer close
(genus or species level)
relationships.
What Sequence to Use ?

Carl Woese
recognized the full potential of rRNA sequences as a measure of phylogenetic relatedness. He initially used an RNA sequencing method that determined about 1/4 of the nucleotides in the 16S rRNA (the best technology available at the time). This amount of data greatly exceeded anything else then available. Using newer methods, it is now routine to determine the sequence of the entire 16S rRNA molecule. Today, the accumulated 16S rRNA sequences (about 10,000) constitute the largest body of data available for inferring relationships among organisms.
Ribosomal RNA Genes and Their Sequences

An example of genes in this
category are those that define
the ribosomal RNAs (rRNAs).
Most prokaryotes have three
rRNAs, called the 5S, 16S and
23S rRNA.
What Sequence to Use ?
Namea Size (nucleotides) Location
5S 120 Large subunit of ribosome
16S 1500 Small subunit of ribosome
23S 2900 Large subunit of ribosome
a The name is based on the rate that the
molecule sediments (sinks) in water.
Bigger molecules sediment faster than small ones.

The extraordinary conservation of rRNA genes can
be seen in these fragments of the small subunit
rRNA gene sequences from organisms spanning
the known diversity of life:
human ...GTGCCAGCAGCCGCGGTAATTCCAGCTCCAATAGCGTATATTAAAGTTGCTGCAGTTAAAAAG...
yeast ...GTGCCAGCAGCCGCGGTAATTCCAGCTCCAATAGCGTATATTAAAGTTGTTGCAGTTAAAAAG...
Corn ...GTGCCAGCAGCCGCGGTAATTCCAGCTCCAATAGCGTATATTTAAGTTGTTGCAGTTAAAAAG...
Escherichia coli ...GTGCCAGCAGCCGCGGTAATACGGAGGGTGCAAGCGTTAATCGGAATTACTGGGCGTAAAGCG...
Anacystis nidulans ...GTGCCAGCAGCCGCGGTAATACGGGAGAGGCAAGCGTTATCCGGAATTATTGGGCGTAAAGCG...
Thermotoga maratima ...GTGCCAGCAGCCGCGGTAATACGTAGGGGGCAAGCGTTACCCGGATTTACTGGGCGTAAAGGG...
Methanococcus vannielii ...GTGCCAGCAGCCGCGGTAATACCGACGGCCCGAGTGGTAGCCACTCTTATTGGGCCTAAAGCG...
Thermococcus celer ...GTGGCAGCCGCCGCGGTAATACCGGCGGCCCGAGTGGTGGCCGCTATTATTGGGCCTAAAGCG...
Sulfolobus sulfotaricus ...GTGTCAGCCGCCGCGGTAATACCAGCTCCGCGAGTGGTCGGGGTGATTACTGGGCCTAAAGCG...
Ribosomal RNA Genes and Their Sequences
Other genes …

• Rate of evolution = rate of mutation
• rate of evolution for any macromolecule is
approximately constant over time (Neutral
Theory of evolution)
• For a given protein the rate of sequence
evolution is approximately constant across
lineages. Zuckerkandl and Pauling (1965)
• This would allow speciation and duplication
events to be dated accurately based on
molecular data
Molecular Clock (MC)
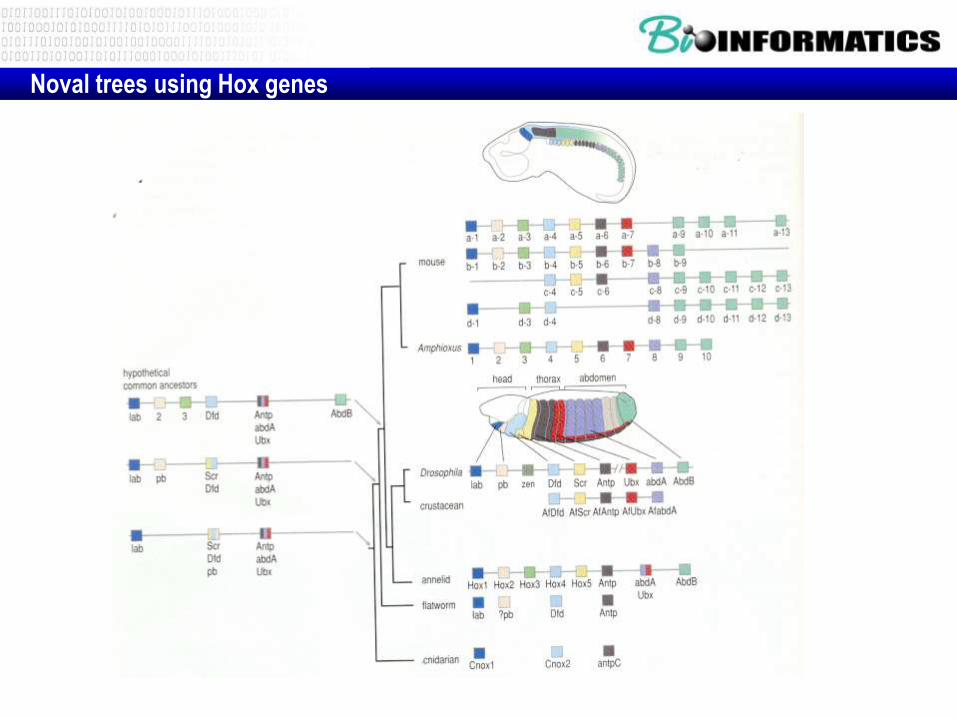
Noval trees using Hox genes
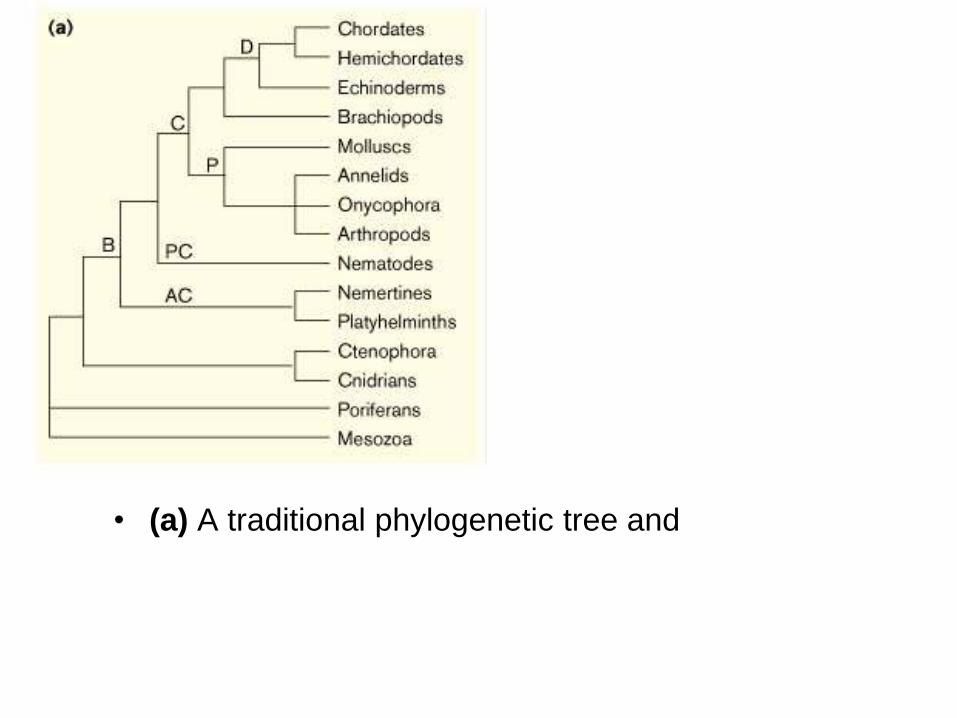
• (a) A traditional phylogenetic tree and
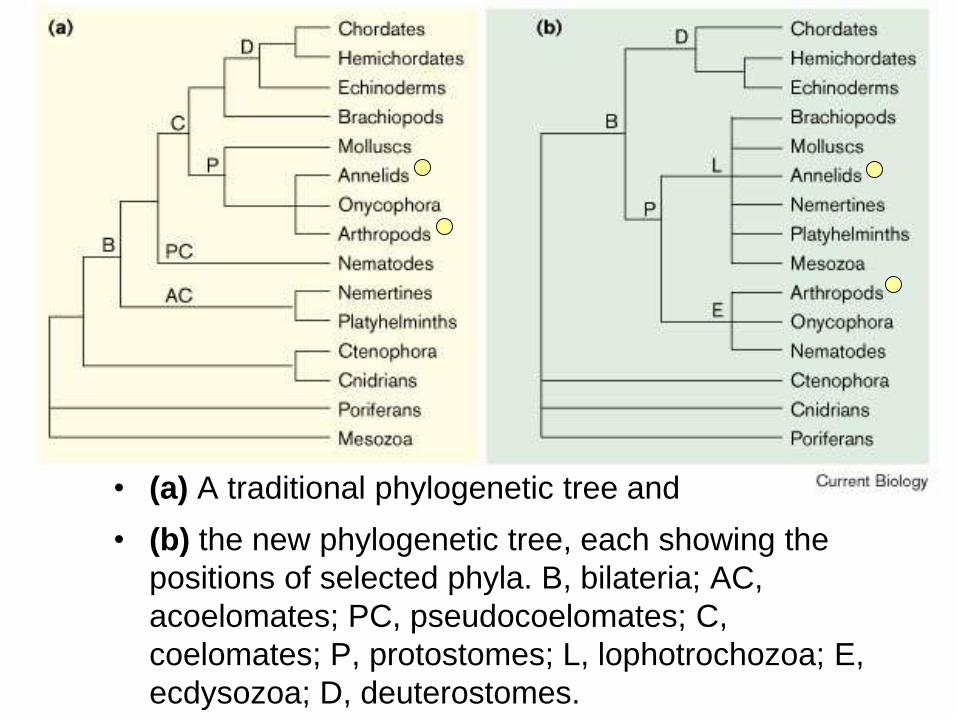
• (a) A traditional phylogenetic tree and
• (b) the new phylogenetic tree, each showing the
positions of selected phyla. B, bilateria; AC,
acoelomates; PC, pseudocoelomates; C,
coelomates; P, protostomes; L, lophotrochozoa; E,
ecdysozoa; D, deuterostomes.

• Local and approximate molecular
clocks more reasonable
– one amino acid subst. 14.5 My
– 1.3 10-9 substitutions/nucleotide site/year
– Relative rate test (see further)
• ((A,B),C) then measure distance between
(A,C) & (B,C)
Molecular Clock (MC)

Rate of Change Theoretical Lookback Time
(PAMs / 100 myrs) (myrs)
Pseudogenes 400 45
Fibrinopeptides 90 200
Lactalbumins 27 670
Lysozymes 24 850
Ribonucleases 21 850
Haemoglobins 12 1500
Acid proteases 8 2300
Cytochrome c 4 5000
Glyceraldehyde-P dehydrogenase2 9000
Glutamate dehydrogenase 1 18000
PAM = number of Accepted Point Mutations per 100 amino acids.
Proteins evolve at highly different rates

Phylogenetics
Introduction
Definitions
Species concept
Examples
The Tree-of-life
Phylogenetics Methodologies
AlgorithmsDistance Methods
Maximum Likelihood
Maximum Parsimony
Rooting
Statistical Validation
Conclusions
Orthologous genes
Horizontal Gene Transfer
Phylogenomics
Practical Approach: PHYLIP
Weblems
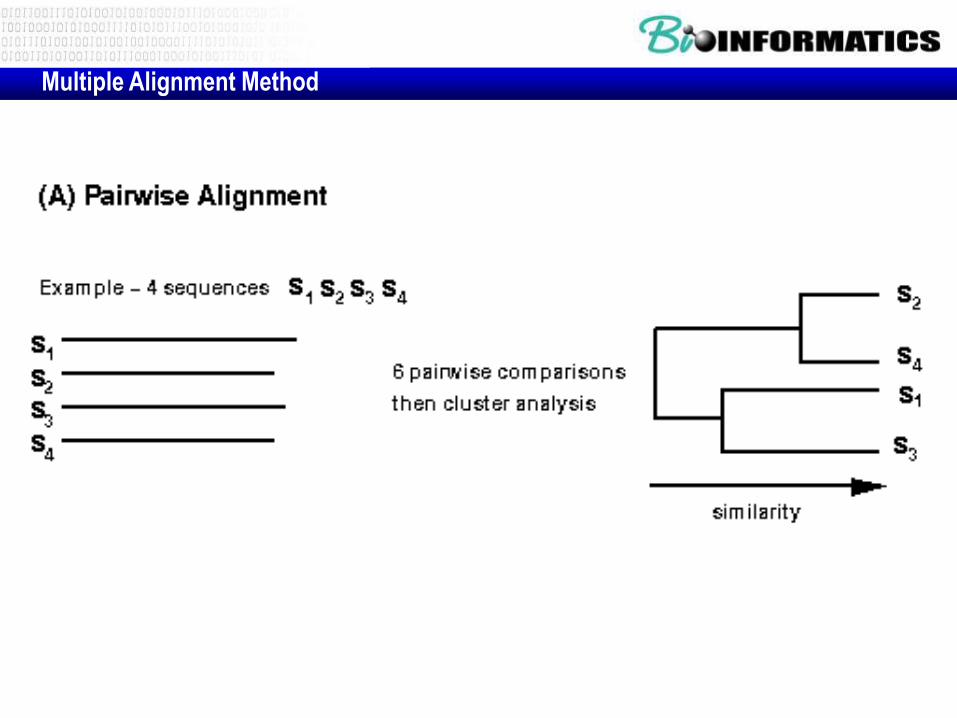
Multiple Alignment Method

• align
• select method (evolutionary
model)
–Distance
–ML
–MP
• generate tree
• validate tree
4-steps


Some definitions …

• Convert sequence data into a
set of discrete pairwise distance
values (n*(n-1)/2), arranged into
a matrix. Distance methods fit a
tree to this matrix.
• The phylogenetic topology tree
is constructed by using a cluster
analysis method (like upgma or
nj methods).
Distance matrix methods (upgma, nj, Fitch,...)


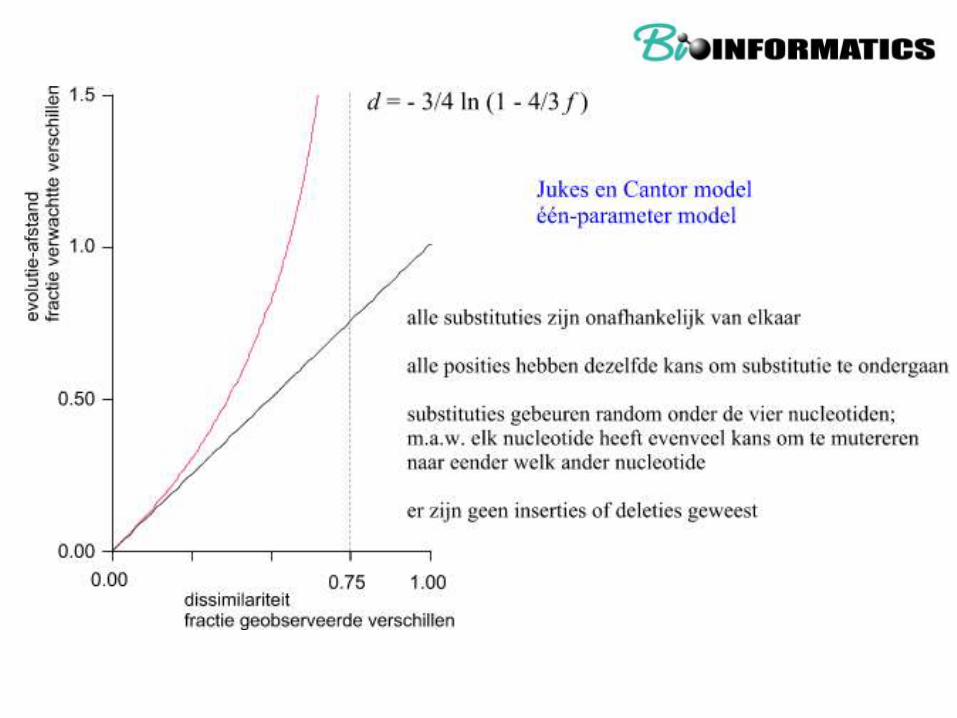
Distance matrix methods (upgma, nj, Fitch,...)

Distance matrix methods (upgma, nj, Fitch,...)
CGT

Distance matrix methods (upgma, nj, Fitch,...)
Since we start with A,p(A)=1

Distance matrix methods (upgma, nj, Fitch,...)
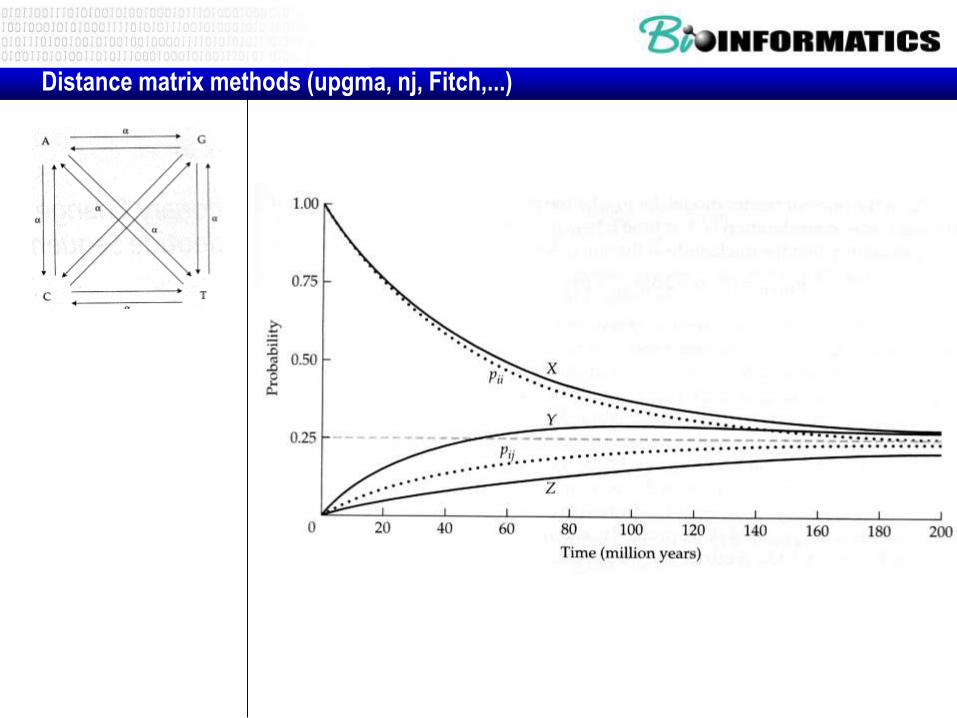
D=evolutionary distance ~ tijd
F = dissimilarity ~ (1 – PX(t))
F ~ 1 – d
Distance matrix methods (upgma, nj, Fitch,...)


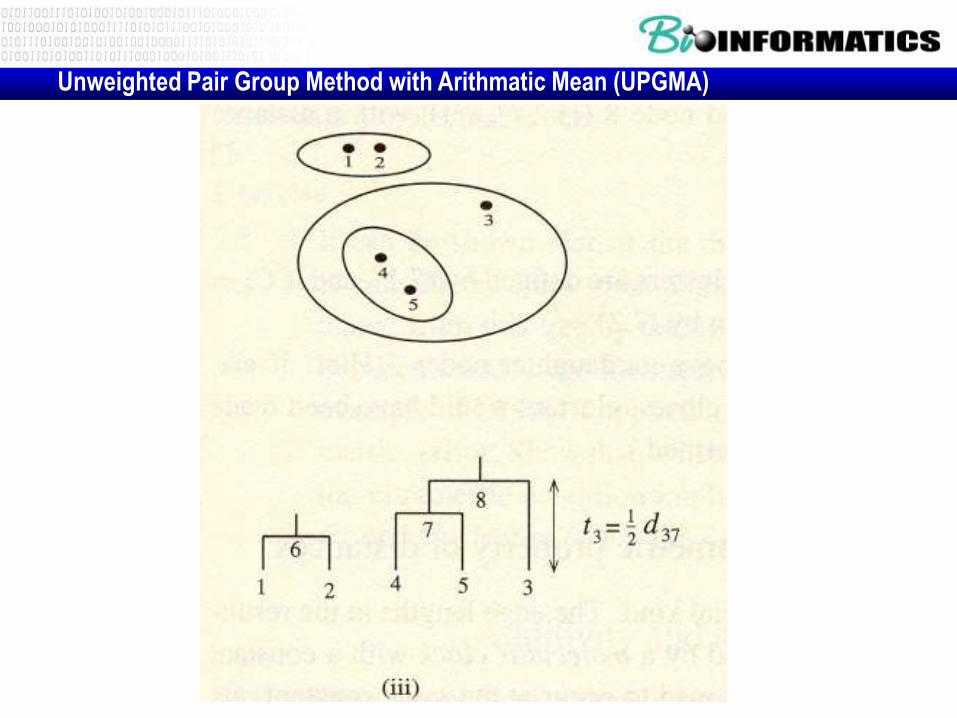
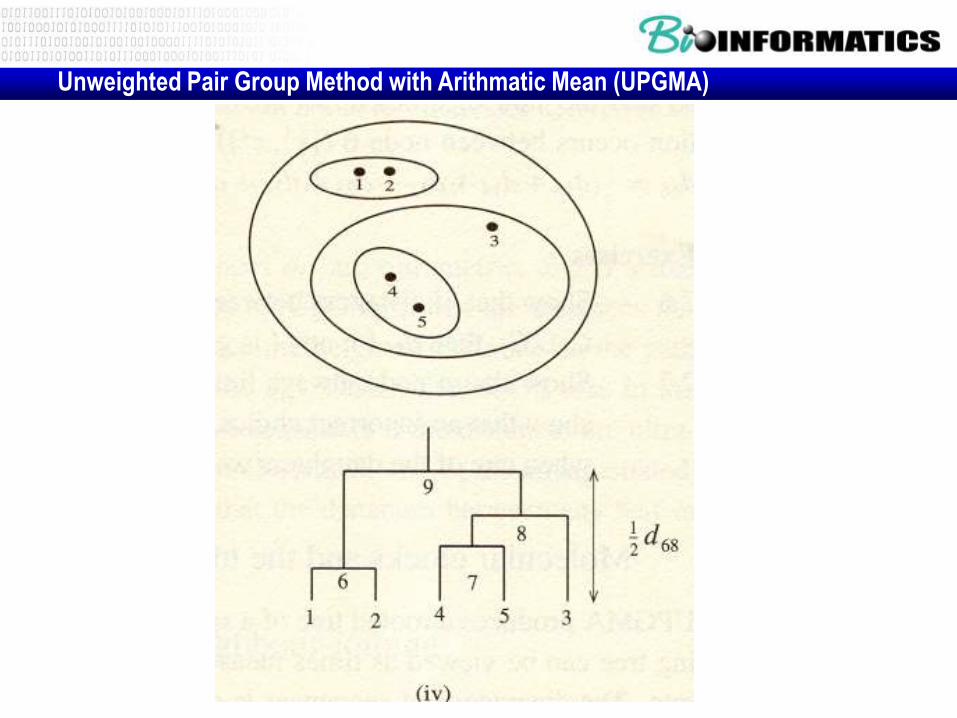
Unweighted Pair Group Method with Arithmatic Mean (UPGMA)

Unweighted Pair Group Method with Arithmatic Mean (UPGMA)
Unweighted Pair Group Method with Arithmatic Mean (UPGMA)
Unweighted Pair Group Method with Arithmatic Mean (UPGMA)
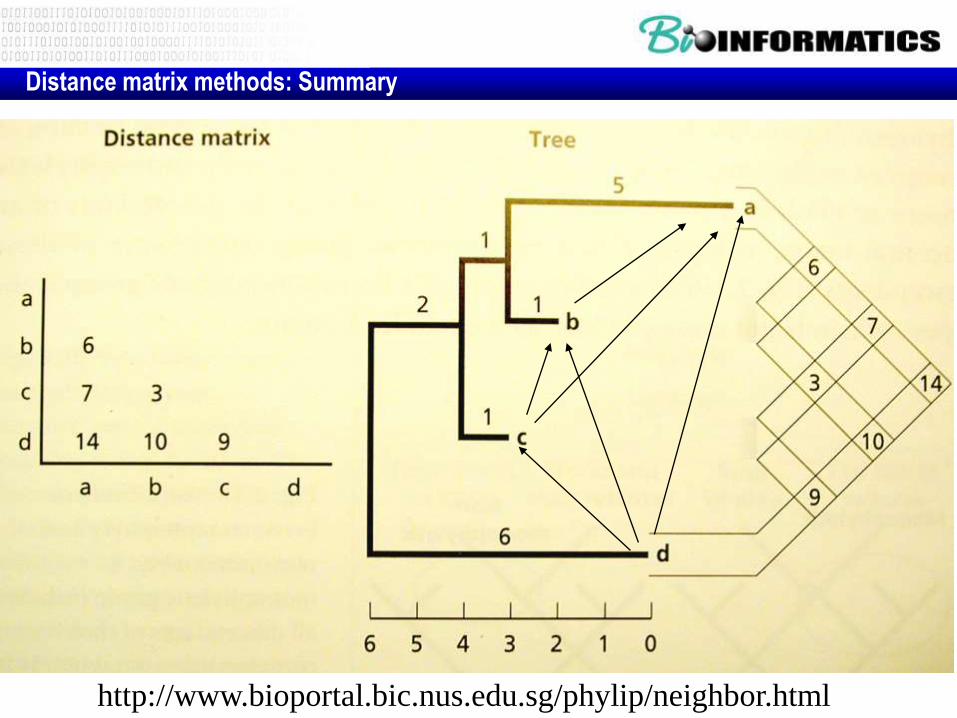
Distance matrix methods: Summary
http://www.bioportal.bic.nus.edu.sg/phylip/neighbor.html

• The phylogeny makes an estimation of the distance for each pair as the sum of branch lengths in the path from one sequence to another through the tree.
easy to perform ;
quick calculation ;
fit for sequences having high similarity scores ;
• drawbacks :
the sequences are not considered as such (loss of information) ;
all sites are generally equally treated (do not take into account differences of substitution rates ) ;
not applicable to distantly divergent sequences.
Distance matrix methods (upgma, nj, Fitch,...)
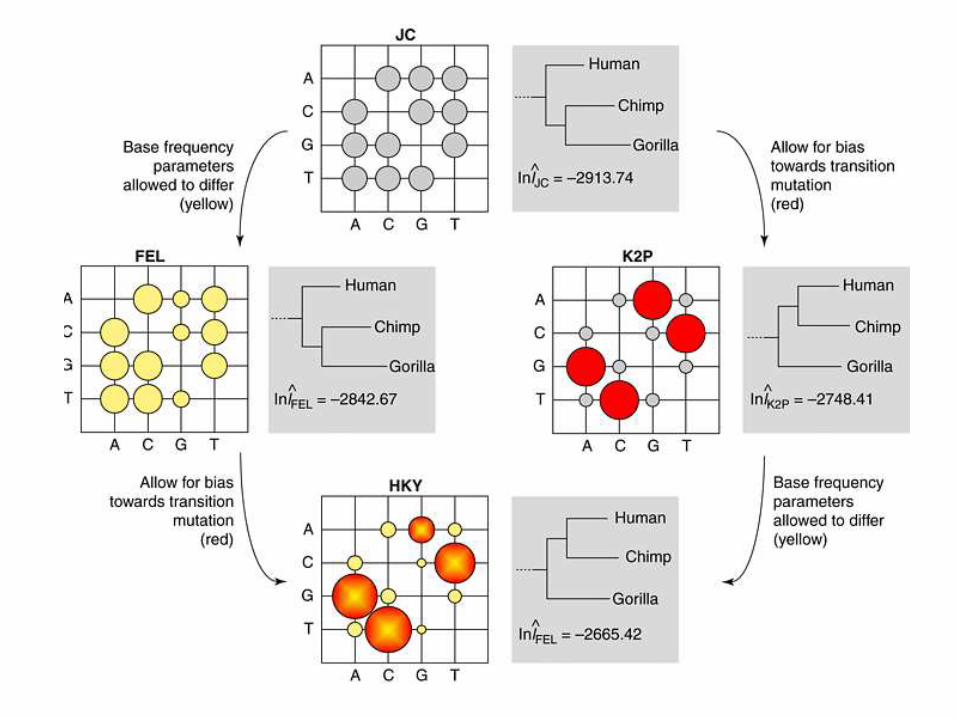


• In this method, the bases
(nucleotides or amino acids) of all
sequences at each site are
considered separately (as
independent), and the log-likelihood
of having these bases are computed
for a given topology by using a
particular probability model.
• This log-likelihood is added for all
sites, and the sum of the log-
likelihood is maximized to estimate
the branch length of the tree.
Maximum likelihood
Maximum likelihood

• This procedure is repeated for all possible topologies, and the topology that shows the highest likelihood is chosen as the final tree.
• Notes :
ML estimates the branch lengths of the final tree ;
ML methods are usually consistent ;
ML is extented to allow differences between the rate of transition and transversion.
• Drawbacks
need long computation time to construct a tree.
Maximum likelihood
Maximum likelihood


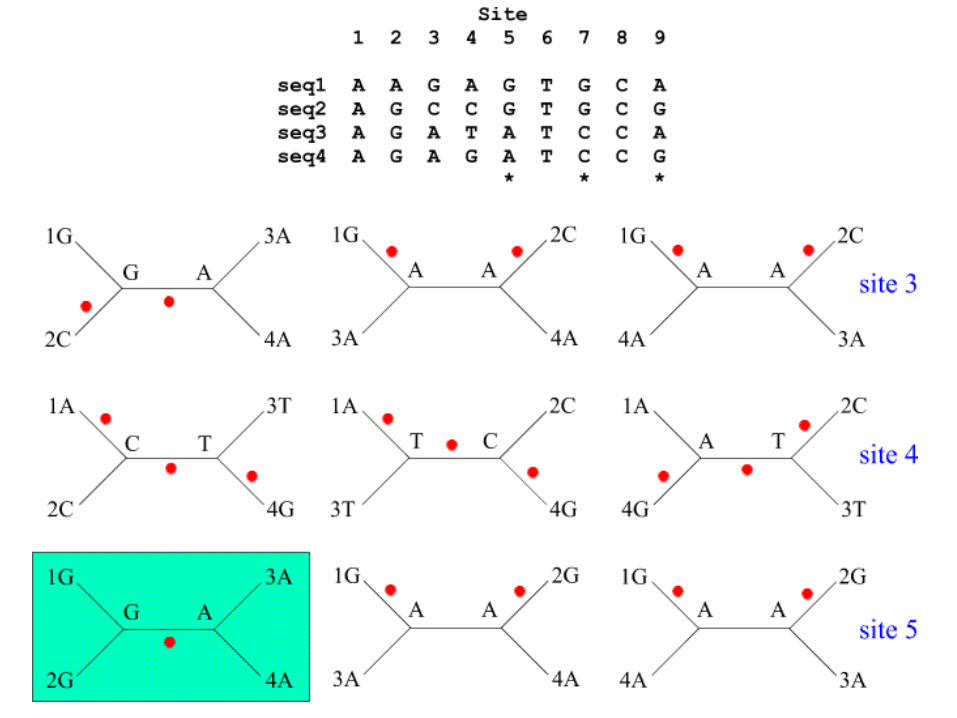
Parsimony criterion
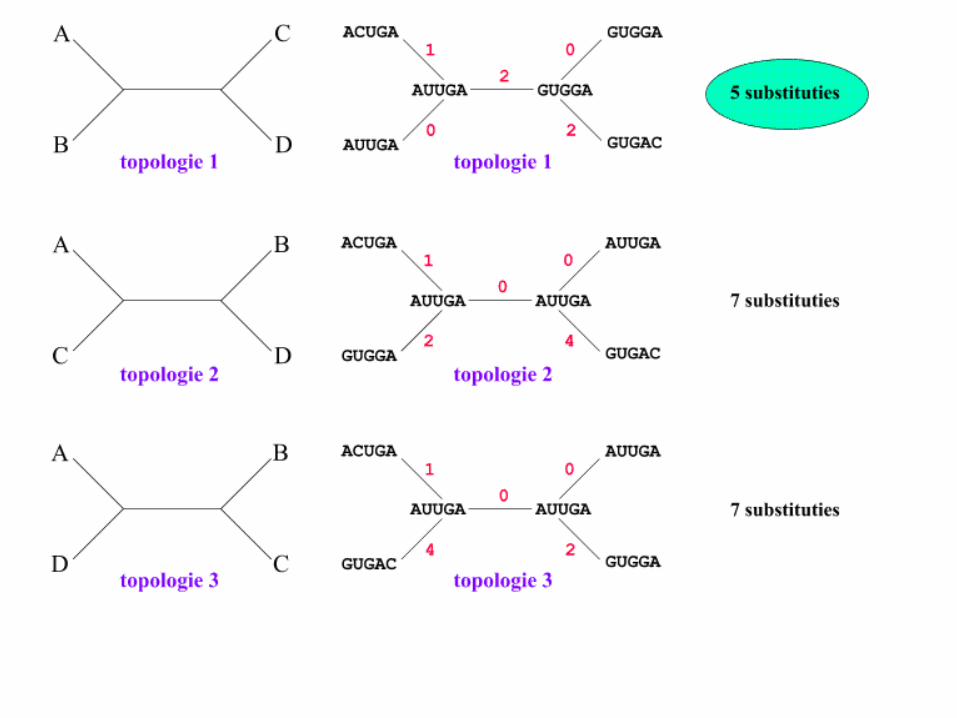
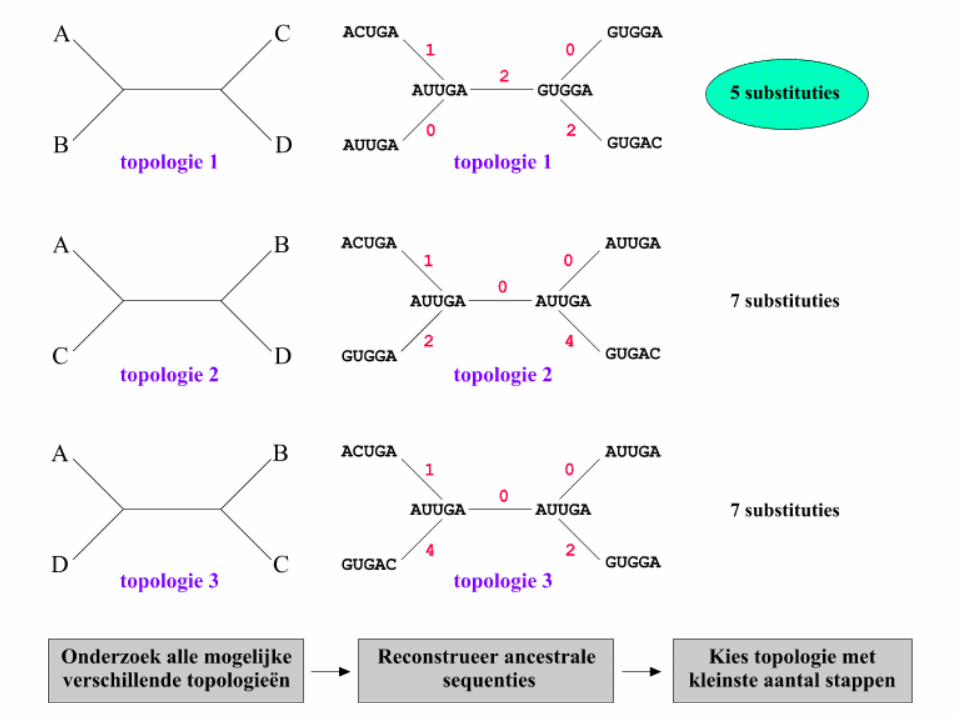
• It consists of determining the minimum number of changes (substitutions) required to transform a sequence to its nearest neighbor.
Maximum Parsimony
• The maximum parsimony algorithm searches for the minimum number of genetic events (nucleotide substitutions or amino-acid changes) to infer the most parsimonious tree from a set of sequences.
Maximum Parsimony

Maximum Parsimony
Occam’s Razor
Entia non sunt multiplicanda praeter necessitatem.William of Occam (1300-1349)
The best tree is the one which requires the least number of
substitutions

• The best tree is the one which needs the
fewest changes.
– If the evolutionary clock is not constant, the
procedure generates results which can be
misleading ;
– within practical computational limits, this
often leads in the generation of tens or more
"equally most parsimonious trees" which
make it difficult to justify the choice of a
particular tree ;
– long computation time to construct a tree.
Maximum Parsimony
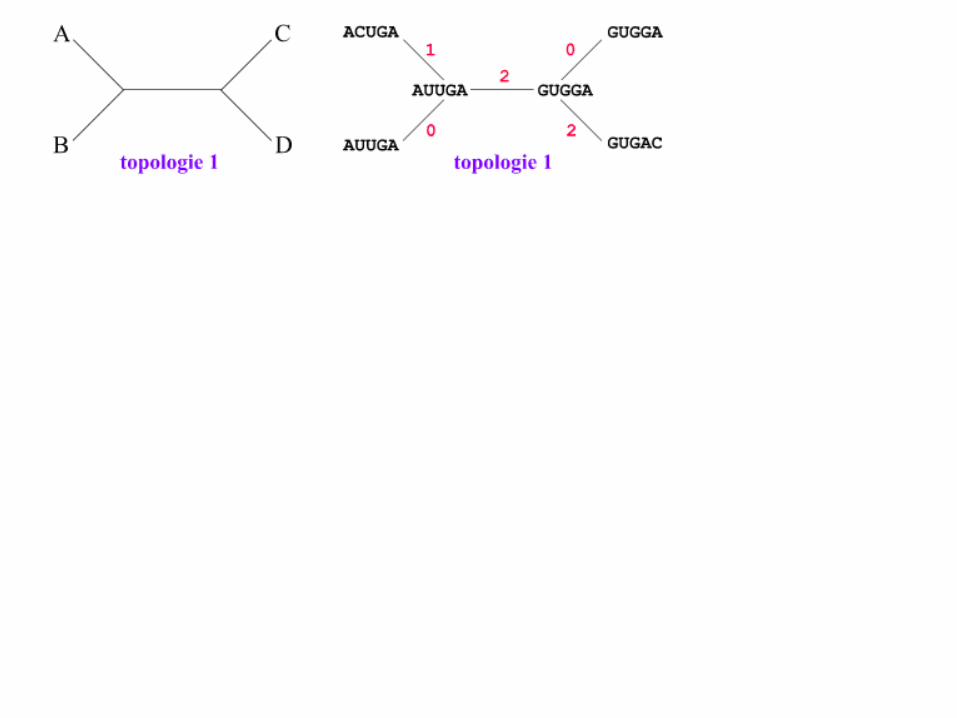

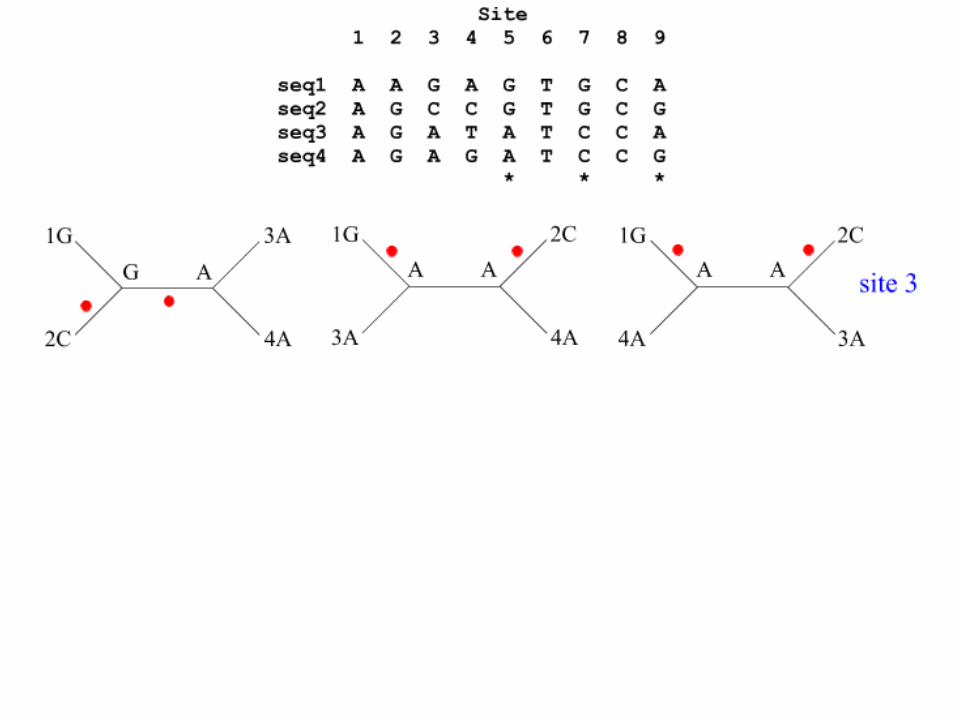
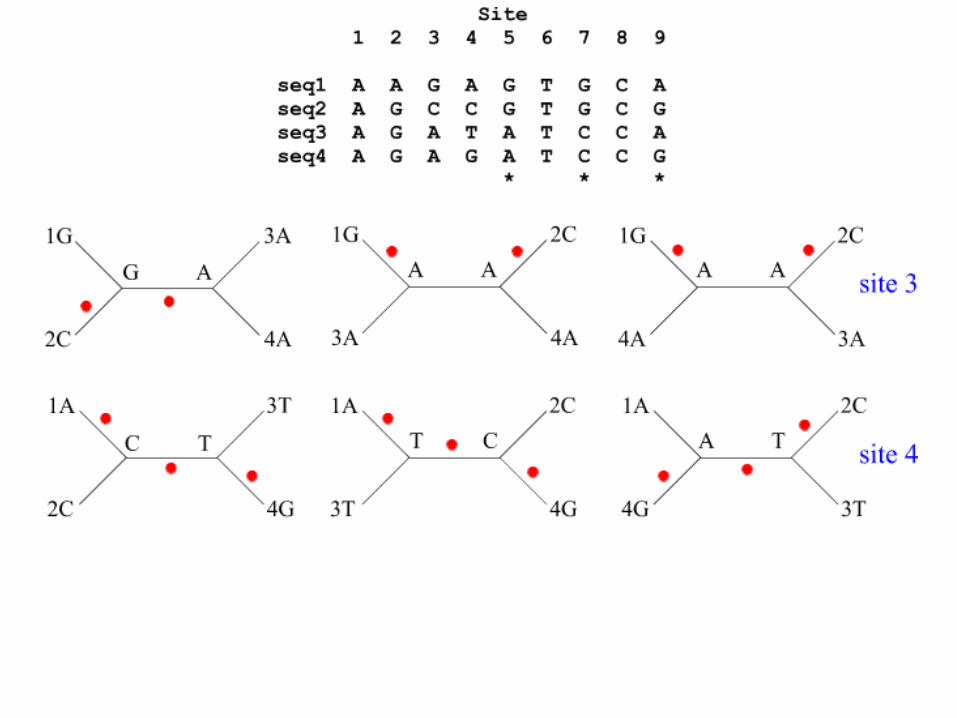
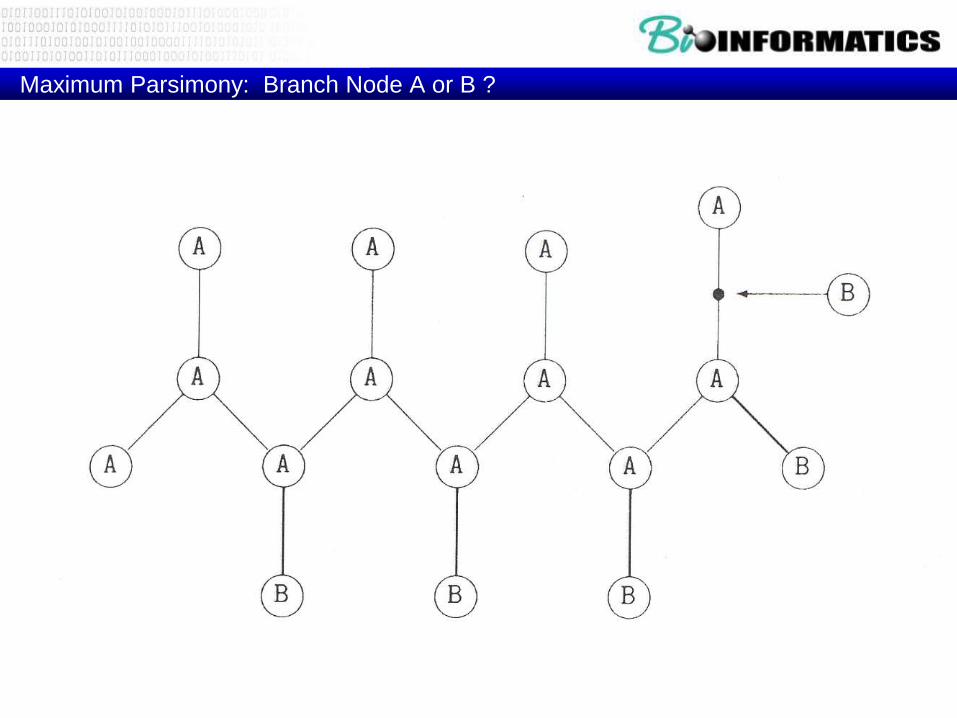
Maximum Parsimony: Branch Node A or B ?
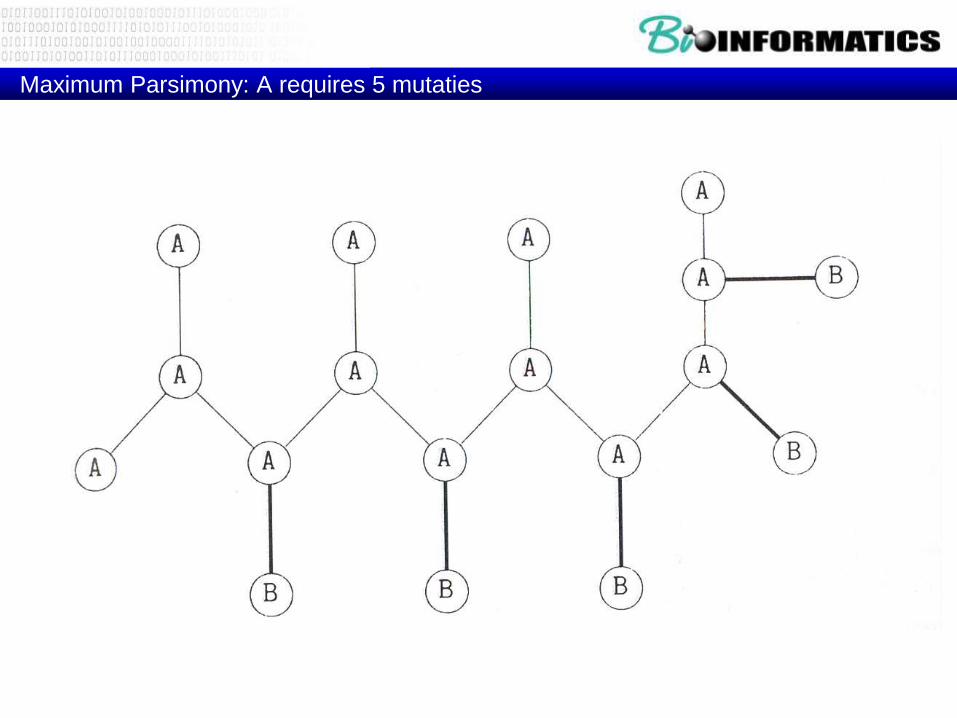
Maximum Parsimony: A requires 5 mutaties
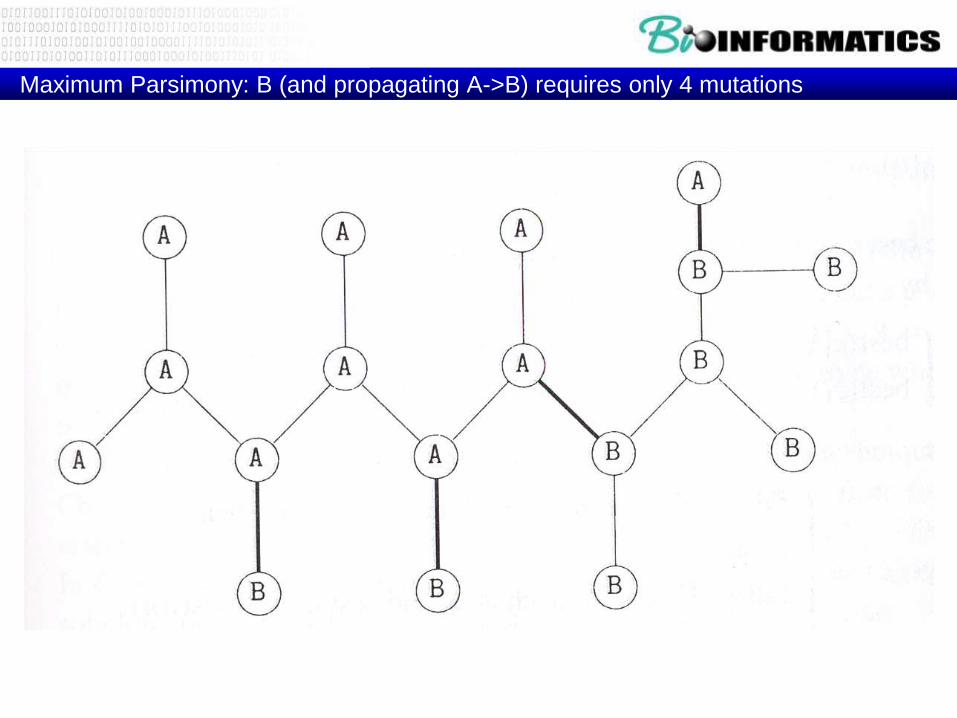
Maximum Parsimony: B (and propagating A->B) requires only 4 mutations

• The best tree is the one which needs the fewest changes.
• Problems :– If the evolutionary clock is not
constant, the procedure generates results which can be misleading ;
– within practical computational limits, this often leads in the generation of tens or more "equally most parsimonious trees" which make it difficult to justify the choice of a particular tree ;
– long computation time to construct a tree.
Maximum Parsimony

Phylogenetics
Introduction
Definitions
Species concept
Examples
The Tree-of-life
Phylogenetics Methodologies
AlgorithmsDistance Methods
Maximum Likelihood
Maximum Parsimony
Rooting
Statistical Validation
Conclusions
Orthologous genes
Horizontal Gene Transfer
Phylogenomics
Practical Approach: PHYLIP
Weblems

There is at present no statistical
methods which allow
comparisons of trees obtained
from different phylogenetic
methods, nevertheless many
studies have been made to
compare the relative consistency
of the existing methods.
Comparative evaluation of different methods

The consistency depends on many
factors, among these the topology
and branch lengths of the real tree,
the transition/transversion rate and
the variability of the substitution
rates.
One expects that if sequences have
strong phylogenetic relationship,
different methods will show the
same phylogenetic tree
Comparative evaluation of different methods

Comparison of methods
• Inconsistency
• Neighbour Joining (NJ) is very fast but depends on accurate estimates of distance. This is more difficult with very divergent data
• Parsimony suffers from Long Branch Attraction. This may be a particular problem for very divergent data
• NJ can suffer from Long Branch Attraction
• Parsimony is also computationally intensive
• Codon usage bias can be a problem for MP and NJ
• Maximum Likelihood is the most reliable but depends on the choice of model and is very slow
• Methods may be combined

Rooting the Tree
• In an unrooted tree the direction of evolution is unknown
• The root is the hypothesized ancestor of the sequences in the tree
• The root can either be placed on a branch or at a node
• You should start by viewing an unrooted tree

Automatic rooting
• Many software packages will root
trees automaticall (e.g. mid-point
rooting in NJPlot)
• Sometimes two trees may look very
different but, in fact, differ only in the
position of the root
• This normally involves assumptions…
BEWARE!

Rooting Using an Outgroup
1. The outgroup should be a sequence (or set
of sequences) known to be less closely
related to the rest of the sequences than they
are to each other
2. It should ideally be as closely related as
possible to the rest of the sequences while
still satisfying condition 1
The root must be somewhere between the
outgroup and the rest (either on the node or
in a branch)

How confident am I that my tree is correct?
Bootstrap values
Bootstrapping is a statistical
technique that can use random
resampling of data to determine
sampling error for tree topologies

Bootstrapping phylogenies
• Characters are resampled with replacement
to create many bootstrap replicate data sets
• Each bootstrap replicate data set is analysed
(e.g. with parsimony, distance, ML etc.)
• Agreement among the resulting trees is
summarized with a majority-rule consensus
tree
• Frequencies of occurrence of groups,
bootstrap proportions (BPs), are a measure
of support for those groups
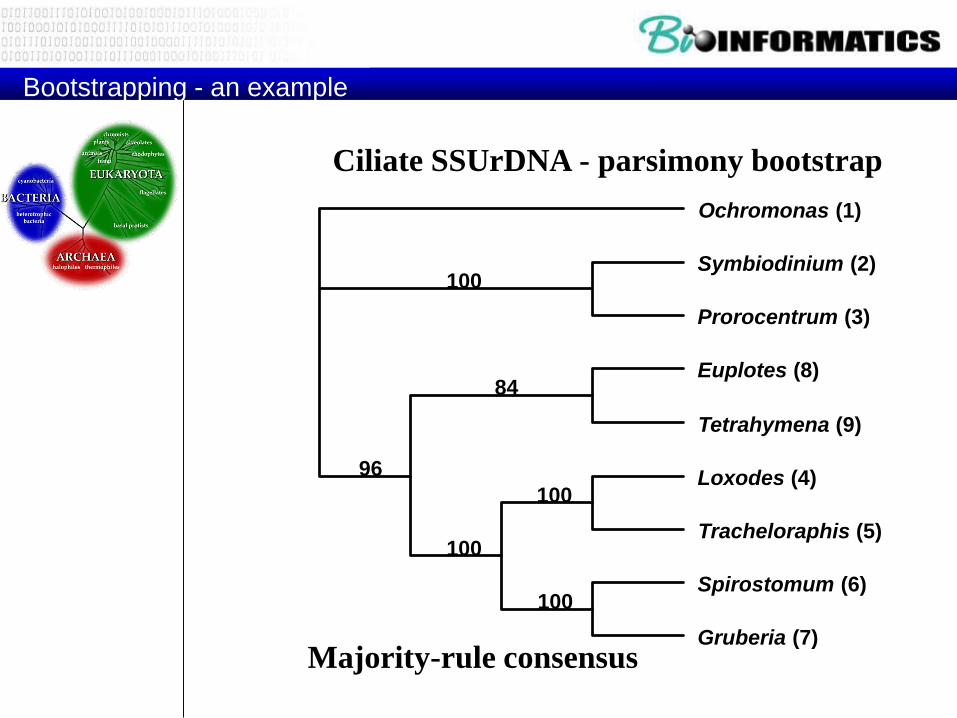
Bootstrapping - an example
Ciliate SSUrDNA - parsimony bootstrap
Majority-rule consensus
Ochromonas (1)
Symbiodinium (2)
Prorocentrum (3)
Euplotes (8)
Tetrahymena (9)
Loxodes (4)
Tracheloraphis (5)
Spirostomum (6)
Gruberia (7)
100
96
84
100
100
100

• Bootstrapping is a very valuable and widely used technique (it is demanded by some journals)
• BPs give an idea of how likely a given branch would be to be unaffected if additional data, with the same distribution, became available
• BPs are not the same as confidence intervals. There is no simple mapping between bootstrap values and confidence intervals. There is no agreement about what constitutes a ‘good’ bootstrap value (> 70%, > 80%, > 85% ????)
• Some theoretical work indicates that BPs can be a conservative estimate of confidence intervals
• If the estimated tree is inconsistent all the bootstraps in the world won’t help you…..
Bootstrap - interpretation

Jack-knifing
• Jack-knifing is very similar to
bootstrapping and differs only in the
character resampling strategy
• Jack-knifing is not as widely
available or widely used as
bootstrapping
• Tends to produce broadly similar
results

At present only sampling techniques allow testing the
topology of a phylogenetic tree
Bootstrapping
» It consists of drawing columns from a sample of
aligned sequences, with replacement, until one gets
a data set of the same size as the original one.
(usually some columns are sampled several times
others left out)
Half-Jacknife
» This technique resamples half of the sequence sites
considered and eliminates the rest. The final sample
has half the number of initial number of sites
without duplication.
Statistical evaluation of the obtained phylogenetic trees

Lineage.py
#Finding the lineage of an organism
#Staying with a plant example, let’s now find the lineage of the Cypripedioideae orchid family. First, we
search the Taxonomy database for Cypripedioideae, which yields exactly one NCBI taxonomy identifier:
from Bio import Entrez
Entrez.email = "[email protected]" # Always tell NCBI who you are
handle = Entrez.esearch(db="Taxonomy", term="Cypripedioideae")
record = Entrez.read(handle)
print (record["IdList"])
#Now, we use efetch to download this entry in the Taxonomy database, and then parse it:
handle = Entrez.efetch(db="Taxonomy", id="158330", retmode="xml")
records = Entrez.read(handle)
#Again, this record stores lots of information:
print (records[0].keys())
[u'Lineage', u'Division', u'ParentTaxId', u'PubDate', u'LineageEx',
u'CreateDate', u'TaxId', u'Rank', u'GeneticCode', u'ScientificName',
u'MitoGeneticCode', u'UpdateDate']
#We can get the lineage directly from this record:
print (records[0]["Lineage"])

WWW resources for molecular phylogeny (1)
Compilations A list of sites and resources:
http://www.ucmp.berkeley.edu/subway/phylogen.h
tml
An extensive list of phylogeny programshttp://evolution.genetics.washington.edu/
phylip/software.html
• Databases of rRNA sequences and
associated software The rRNA WWW Server - Antwerp, Belgium.
http://rrna.uia.ac.be
The Ribosomal Database Project - Michigan State University
http://rdp.cme.msu.edu/html/

Database similarity searches (Blast) :http://www.ncbi.nlm.nih.gov/BLAST/
http://www.infobiogen.fr/services/menuserv.html
http://bioweb.pasteur.fr/seqanal/blast/intro-fr.html
http://pbil.univ-lyon1.fr/BLAST/blast.html
Multiple sequence alignment
ClustalX : multiple sequence alignment with a graphical interface
(for all types of computers).http://www.ebi.ac.uk/FTP/index.html and go to ‘software’
Web interface to ClustalW algorithm for proteins:
http://pbil.univ-lyon1.fr/ and press “clustal”
WWW resources for molecular phylogeny (2)

• Sequence alignment editor
SEAVIEW : for windows and unixhttp://pbil.univ-lyon1.fr/software/seaview.html
• Programs for molecular phylogeny PHYLIP : an extensive package of programs for all platforms
http://evolution.genetics.washington.edu/phylip.html
CLUSTALX : beyond alignment, it also performs NJ
PAUP* : a very performing commercial packagehttp://paup.csit.fsu.edu/index.html
PHYLO_WIN : a graphical interface, for unix onlyhttp://pbil.univ-lyon1.fr/software/phylowin.html
MrBayes : Bayesian phylogenetic analysis http://morphbank.ebc.uu.se/mrbayes/
PHYML : fast maximum likelihood tree buildinghttp://www.lirmm.fr/~guindon/phyml.html
WWW-interface at Institut Pasteur, Parishttp://bioweb.pasteur.fr/seqanal/phylogeny
WWW resources for molecular phylogeny (3)

Weblems
W6.1: The growth hormones in most mammals have very similar ammo acid sequences. (The growth hormones of the Alpaca, Dog Cat Horse, Rabbit, and Elephant each differ from that of the Pig at no more than 3 positions out of 191.) Human growth hormone is very different, differing at 62 positions. The evolution of growth hormone accelerated sharply in the line leading to humans. By retrieving and aligning growth hormone sequences from species closely related to humans and our ancestors, determine where in the evolutionary tree leading to humans the accelerated evolution of growth hormone took place.
W6.2: Humans are primates, an order that we, apes and monkeys share with lemurs and tarsiers. On the basis of the Beta-globin gene cluster of human, a chimpanzee, an old-world monkey, a new-world monkey, a lemur, and a tarsier, derive a phylogenetic tree of these groups.
W6.3: Primates are mammals, a class we share with marsupials and monotremes; Extant marsupials live primarily in Australia, except for the opossum, found also in North and South America. Extant monotremes are limited to two animals from Australia: the platypus and echidna. Using the complete mitochondnal genome from human, horse (Equus caballus), wallaroo (Macropus robustus), American opossum (Didelphis mrgimana), and platypus (Ormthorhynchus anatmus), draw an evolutionary tree, indicating branch lengths. Are monotremes more closely related to placental mammals or to marsupials?
W6.4: Mammals are vertebrates, a subphylum that we share with fishes, sharks, birds and reptiles, amphibia, and primitive jawless fishes (example: lampreys). For the coelacanth (Latimeria chalumnae), the great white shark (Carcharodon carcharias), skipjack tuna (Katsuwonus pelamis), sea lamprey (Petromyzon marinus), frog (Rana Ripens), and Nile crocodile (Crocodylus niloticus), using sequences of cytochromes c and pancreatic ribonucleases, derive evolutionary trees of these species.