Embed Size (px)
Citation preview

UNIVERSIDAD DE CUENCADepartamento de educación continua
Seminario – Taller herramientas informáticas para la docencia e investigación
CLASE VIRTUAL
MEMBRANA CELULAR
LCDO. MAURICIO BACULIMA

MEMBRANA CELULAR O PLASMÁTICA
Las células están separadas del ambiente externo por
una estructura denominada MEMBRANA PLASMÁTICA.
La membrana es sumamente delgada (5 a 10 nm de espesor)
Su estructura es posible estudiarla mediante el
microscopio electrónico (1950).
No es una pared rígida o cubierta inerte
Todas las membranas de la célula poseen una misma estructura, pero su composición química va a variar de
manera considerable.

Tiene una estructura de tres capas compuestas por dos capas de color obscuro orientadas hacia afuera y en medio una capa de color claro

Es la estructura que delimita a la célula
(constante).
En el inicio de la vida, permitió justamente el
delimitar un espacio interno y por tanto de los
seres vivos.
Permite mantener una composición diferente
entre la célula y el medio que le rodea.

FUNCIONES:
Límite.
Intercambio de substancias (barrera selectiva-permeable).
Comunicación entre las células.
Reconocimiento celular e interacción, (receptora de
sustancias)
Contacto entre las células, adherirse cuando es apropiado
e intercambiar materiales e información.
Recepción de mensajes (inmunoreacción)

ESTRUCTURA

“El método de congelamiento utilizado en microscopia electrónica hizo posible el estudio morfológico del
interior de la membrana”
El verdadero núcleo de la membrana consiste en una vaina de fosfolípidos, una delgada capa bimolecular (bicapa de lípidos).
Impide movimientos desordenados de materiales hidrosolubles hacía adentro y fuera de la célula.
Las proteínas , efectúan la mayor parte de las funciones específicas (selección previa).

COMPOSICION DE LA MEMBRANA
Composición química de la membrana:
Proteínas 50% Lípidos 40%, Glúcidos 10%.
Todas las membranas tienen está composición, las proporciones de cada una de ellos varia de un tipo celular a otro.
Singer y Nicholson (1972) proponen el llamado modelo del “mosaico fluido”

TEJIDO Y ESPECIE
PORCENTAJE DE
PROTEÍNAS
PORCENTAJE DE LÍPIDOS
SNC en el Humano 20 79
SNC en el Bovino
23 76
Eritrocito enHumano 60 40
Músculo de rata 70 27

En la membrana interna de las mitocondrias, la relación proteína/lípidos es muy alta en comparación con las membranas de la vaina de mielina que rodean una célula nerviosa.
La membrana interna de las mitocondrias contiene proteínas transportadoras de la cadena de transporte de los electrones
La mejor manera de describir a la vaina de mielina es como un aislante eléctrico realizada por una gruesa capa de lípidos


MOSAICO FLUIDO
Desde hace mas de 50 años se sabe que la membrana está compuesta
principalmente por lípidos y proteínas.
Las proteínas del mosaico fluido se presentan como un mosaico de
partículas discontinuas.
La membrana no es rígida (fluido), tanto los lípidos y proteínas tienen
notable libertad de movimientos laterales.
La membrana es asimétrica, las dos mitades no son iguales (cara
interna y externa).
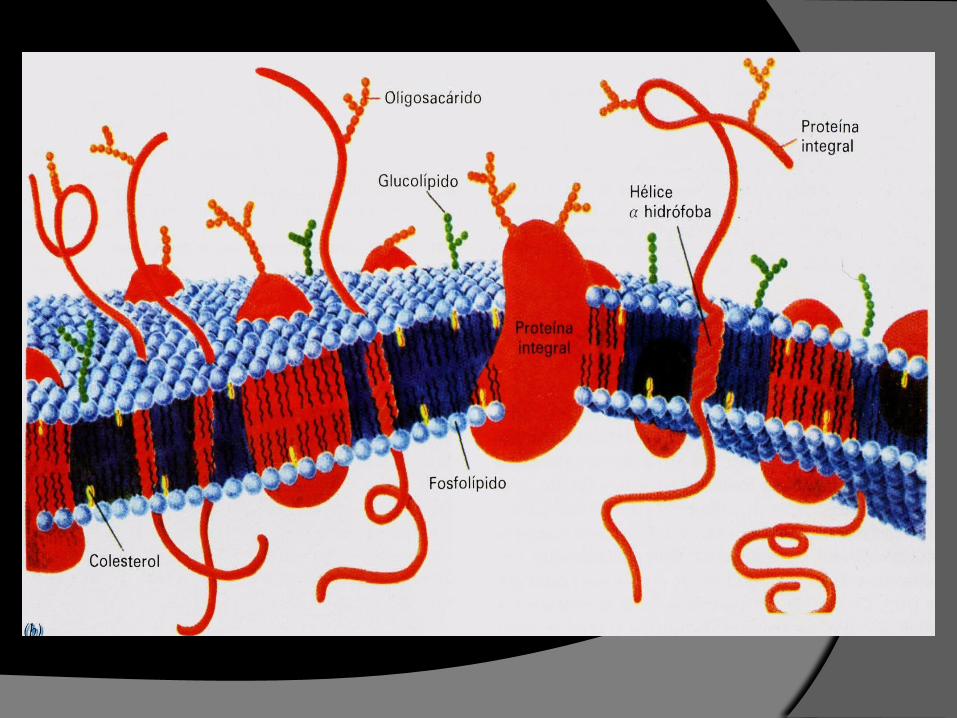

Lípidos de la membrana Contiene varios tipos de lípidos, todos anfipáticos.
La mayor parte de los lípidos contiene un grupo fosfato excepto colesterol y glucolípidos.
Los principales lípidos son :
1. Fosfolípidos1. Fosglicéridos
2. Esfingolípidos
2. Colesterol

FOSFOLÍPIDOS:
Poseen un esqueleto glicerol (fosfoglicéridos), los glicéridos de
la membrana son diglicéridos.
Los diglicéridos de la membrana contiene un grupo adicional
unido al fosfato :
Colina – fosfatidilcolina
Etanolamina – fosfatidiletanolamina
Serina – fosfatidilserina
Inositol – fosfatidilinositol
Constituyen mas de la mitad de los lípidos en la mayoría de las membranas


Un tipo menos abundante de lípidos son los esfingolípidos:
Derivan de esfingosina (Ác. Graso)Alcohol aminadoLarga cadena de hidrocarburo
CERAMIDO

Los diglicéridos de la membrana se une, Hidrato de carbono, (glucolípido):
Azúcar simple - cerebrósido Oligosacárido - gangliósido
También son anfipáticas y con estructura total básicamente similar a la de fosfoglicéridos.


Movimiento de los fosfolípidos:
FLIP-FLOP: pueden saltar de una monocapa a la otra, pero se produce poco porque requiere gran gasto de energía.
DIFUSIÓN LATERAL: cambian de
lugar con fosfolípidos vecinos, dentro de la misma monocapa. Esto da lugar a la difusión lateral.
ROTACIÓN: giran sobre su eje longitudinal con rapidez.
FLEXIÓN: Separación y
aproximación de los extremos de las colas, por flexión de las cadenas carbonadas de los ácidos grasos.

COLESTEROL Ciertas células animales pueden tener hasta 50% .
Es mas pequeño que otros lípidos y menos anfipático.
La mol. del colesterol se orienta con sus grupos OH (hidrófilos) hacia la superficie .
Su extremo hidrófobo integrado a la bicapa de lípidos.
La mayor parte de los vegetales y de todas las células bacterianas carecen de colesterol

COLESTEROL EN LA MEMBRANA
La colocación de las mol. De colesterol impide el apretado empacamiento de los fosfolípidos, que tiende a incrementar la fluidez de la bicapa.

El anillo esteroide interactúa con la cadena inicial de los ac. grasos, a los que inmoviliza parcialmente.
Produce dos efectos:○ Incrementa la impermeabilidad de la bicapa a mol. Hidrofílicas○ Decrece la flexibilidad y la fluidez de la membrana a 37°C.
El colesterol:
Aumenta la impermeabilidad de la capa lipídica al aumentar la temperatura. Esto hace a la membrana menos deformable y menos fluida, es decir, la estabiliza.
Mantiene la fluidez ante la disminución de la temperatura ,previene el compactamiento de las cadenas hidrocarbonadas, ya que evita que las colas se junten y se “cristalicen” (adopten una estructura muy compacta).

Tienen un importante papel en la regulación de las propiedades físico-químicas de la membrana biológicas regulando su resistencia y fluidez

PROTEÍNAS DE LA MEMBRANA
Son el componente funcional fundamental de las membranas biológicas.
No sólo son de estructura, sino también en su permeabilidad (canales o transportadores).
No se disponen al azar, sino cada una se localiza y orienta en una posición particular.
Se sitúan asimétricamente.

1. Proteínas integrales
o intrínsecas
2. Proteínas periféricas
o extrínsecas
3. Proteínas ancladas a
lípidos
Las proteínas de la membrana se pueden agrupar en tres tipos distintos según la intimidad con la bicapa lipídica:

Clases de proteínas:
Proteínas transmembrana (1 y 2) no transmembrana (3), membrana expuestas en la superficie celular externa (4), interacciones no covalentes con otras proteínas de membrana (5 y 6).

Proteínas integrales , intrínsecas o transmembrana
Son anfipáticas.
En su mayoría son
transmembranosas (glucoproteínas).
Las proteínas integrales (70%)de la membrana contienen residuos no polares en gran parte de su superficie expuesta.
a. una hélice alfa y b. una estructura globular terciaria

Las regiones no polares se integran al interior de la bicapa de lípidos
El resto de una proteína integral se compone de aa iónicos y no polares integrados a la bicapa.
Formando un canal
acuoso a través de la misma.
a. una hélice alfa y b. una estructura globular terciaria
Proteínas integrales , intrínsecas o transmembrana

Las proteínas integrales no están fijas,
más bien deben tener la capacidad para
desplazarse lateralmente.
Existen proteínas transmembranosas de
paso único y de paso múltiple

Las partes de estas proteínas que atraviesan la membrana
son regiones alfa-helicoidales (20 a 25 aa no polares)
Otra estructura proteica conocida que atraviesa bicapas
lipídicas es el barril-beta , formada por el plegamiento de
láminas-B

Proteínas periféricas o extrínsecas
Se unen a la membrana mediante débiles enlaces electroestáticos, (porciones hidrófilas, proteínas integrales).
Forman una red fibrilar que actúan como esqueleto flexible (cambios morfológicos, fijar proteínas integrales).
Otras funcionan como enzimas o factores transmisores de señales.


Proteínas de membrana ancladas a lípidos
Existen dos tipos :
1. Según los tipos de anclaje al lípido.
En la cara externa de la membrana se
enlazan mediante un oligosacárido corto
unido a una mol de glucofosfatidilinositol

2. La superficie de la membrana sobre la cual están expuestos.
Presentes en el lado citoplasmático ,
anclados a la membrana mediante
largas cadenas de hidrocarburos (hoja
interna de la bicapa)

Lípidos y proteínas de la membrana

Glúcidos de la membrana
Se representan en forma de oligosacáridos o menos
frecuente monosacáridos.
Unidos en forma covalente a lípidos-glucolípidos y
proteínas-glucoproteínas.
Otros componentes glucídicos de las membranas celulares
son los glucosaminoglucanos (GAGs) unidos a proteínas
(proteoglucanos).

Gal: galactosa Glc: glucosa GalNAc: N-acetil galactosamina
Los glúcidos se ubican casi en forma exclusiva en la hoja superficial de la membrana plasmática

Se piensa que los H de C ramificados participan
en la mediación de interacciones celulares.
Todavía no se comprende bien las funciones de los
glucolípidos de la membrana:
Los glucolípidos de la membrana plasmática de
eritrocitos determinan el grupo sanguíneo
Participan en reacciones inmunitarias
Actúan como receptores de membrana

Glucocaliz, glicocálix o cubierta celular
Los glucolípidos y glucoproteínas intrínsecos se proyectan a la superficie externa de la célula, conjuntamente con proteoglucanos integrales.
En la mayor parte de estas su grosor, de 10 a 20 nm, se halla por debajo del límite resolutivo del microscopio óptico.

GLUCOCALIZ

FUNCIONES.
Micro ambiente: “atrapar” ciertos iones que son importantes para la célula y agua del medido extracelular. Poseen muchas cargas negativas.
Protección celular: daños de origen químico o mecánico.
Reconocimiento celular: a partir de la composición diferencial que presentan los distintos tipos celulares en su Glucocáliz.


La adhesión de los GB a las células endoteliales que limitan los vasos sanguíneos, proceso que permite a los GB abandonar el sistema circulatorio e intervenir en la respuesta inflamatoria.
RECONOCIMIENTO CELULAR

PROPIEDADES DE LA MEMBRANA
FluidezAsimetríaElasticidadResistencia mecánicaFuerza de cohesiónCapacidad de regeneración Resistencia eléctrica Regenerabilidad

















