Embed Size (px)
DESCRIPTION
Todo lo referente a Hidratos de Carbonos desde el punto de vista Bioquímico por el Profesor Vicente Boniello
Citation preview


BUENO RECORDARBUENO RECORDAR
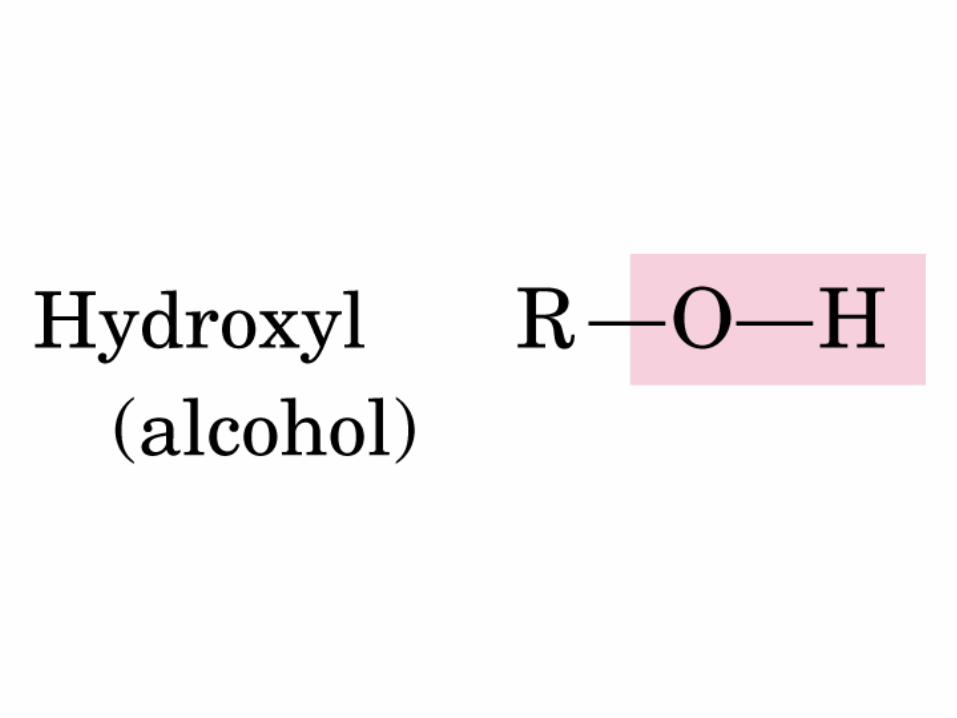
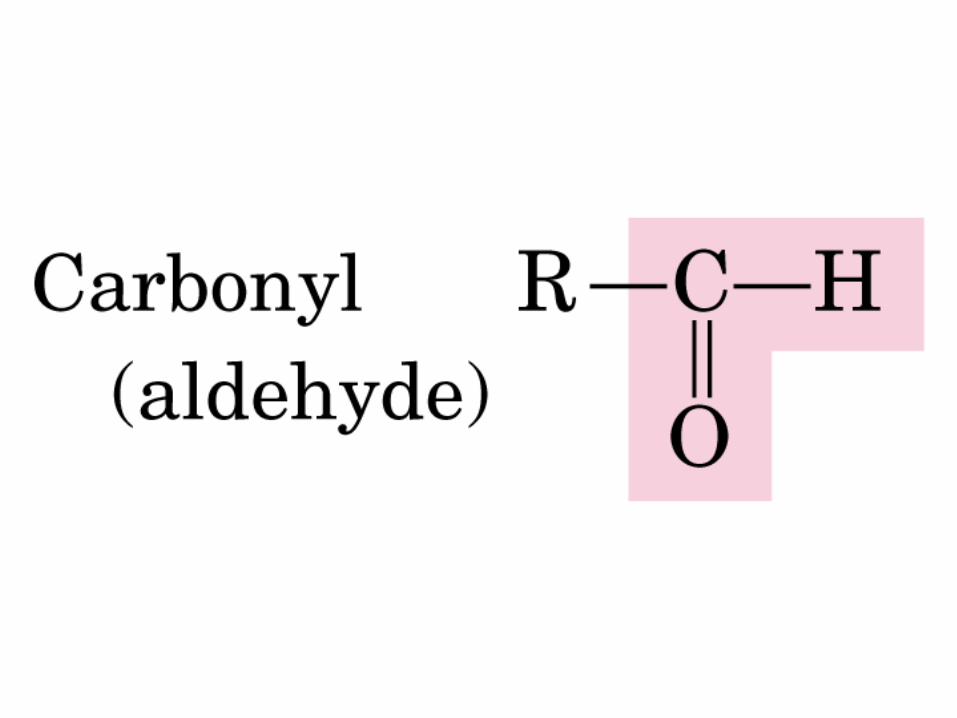
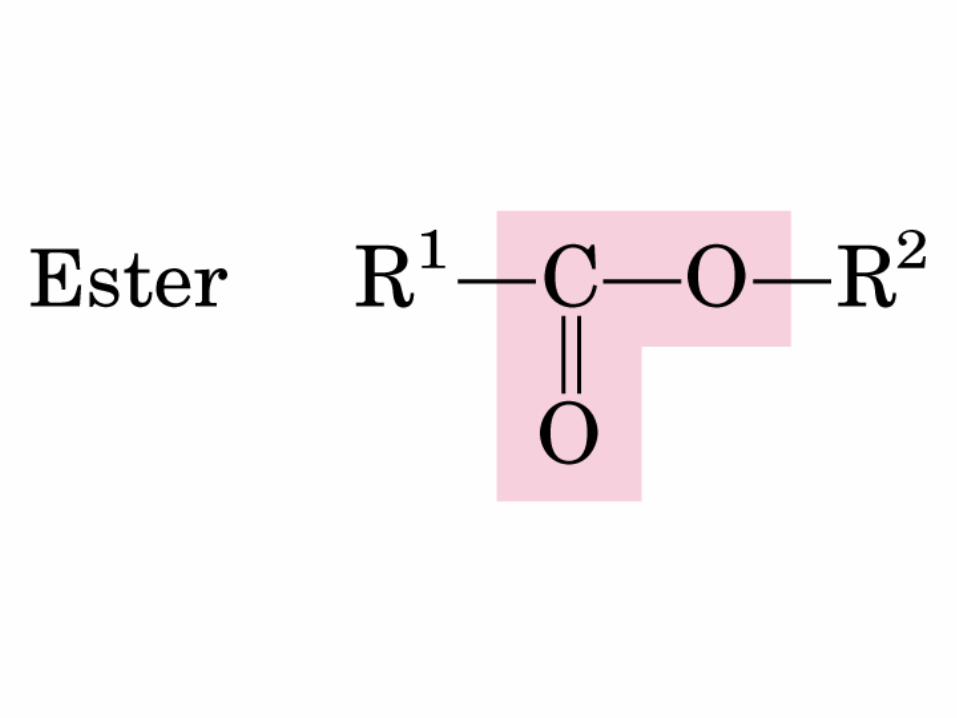
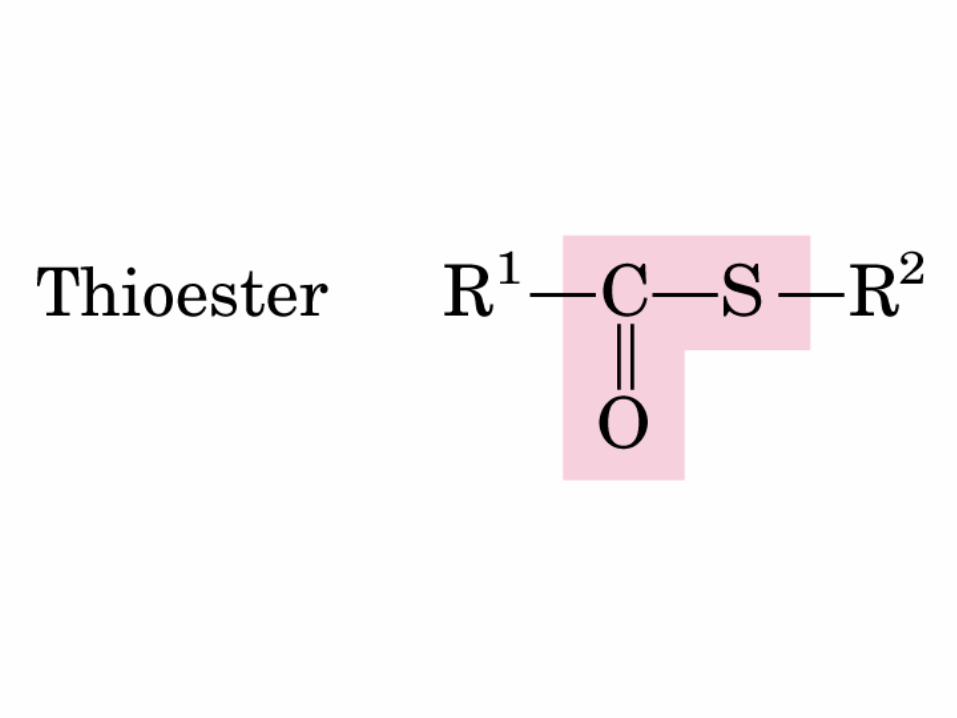
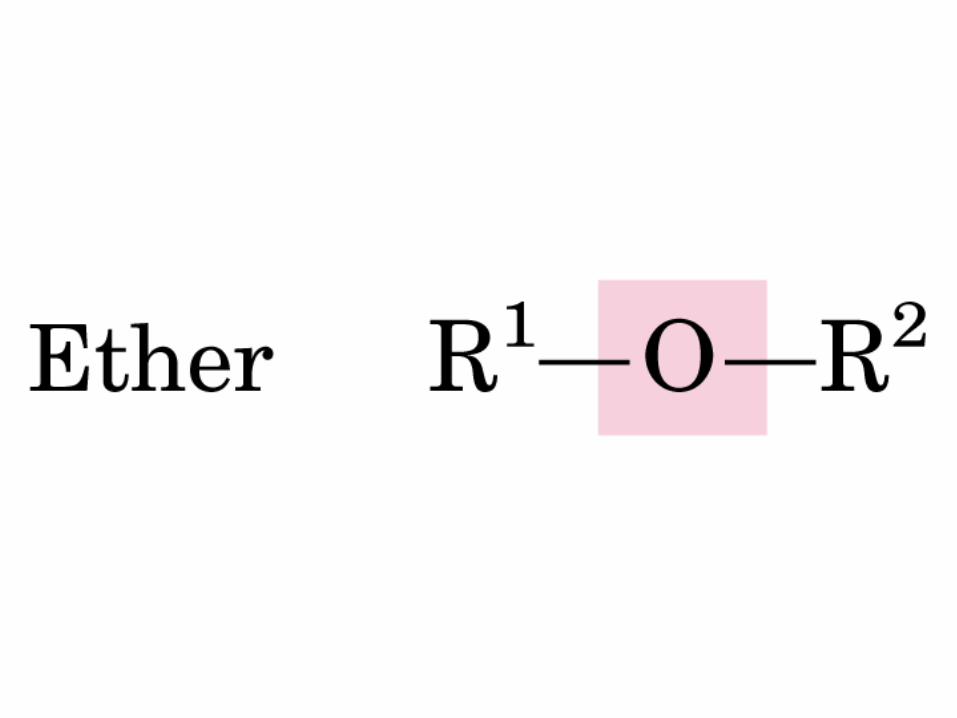
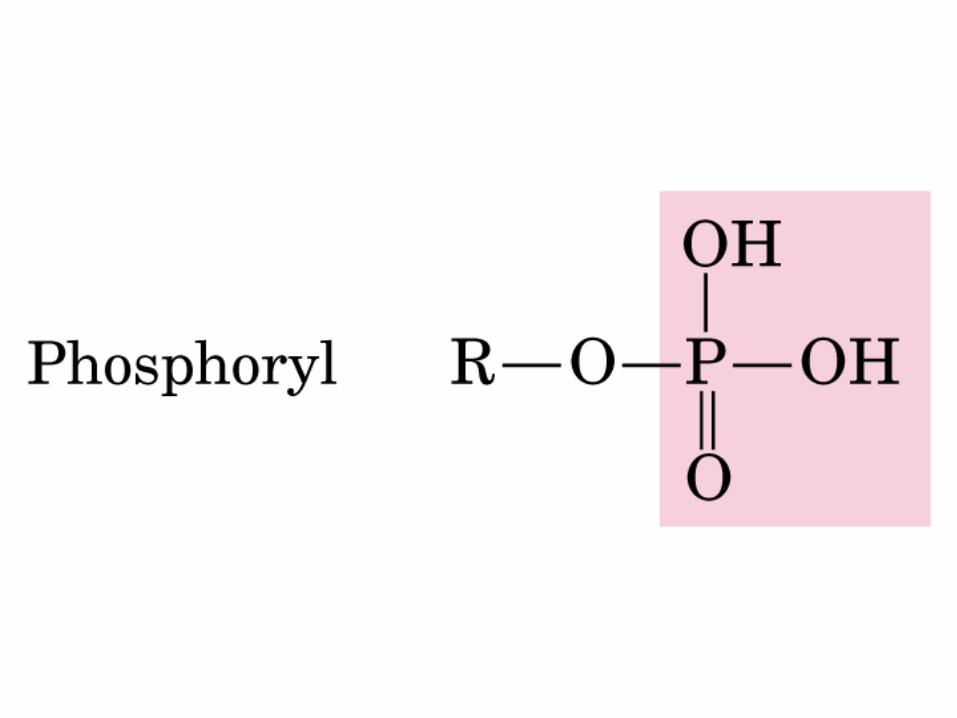
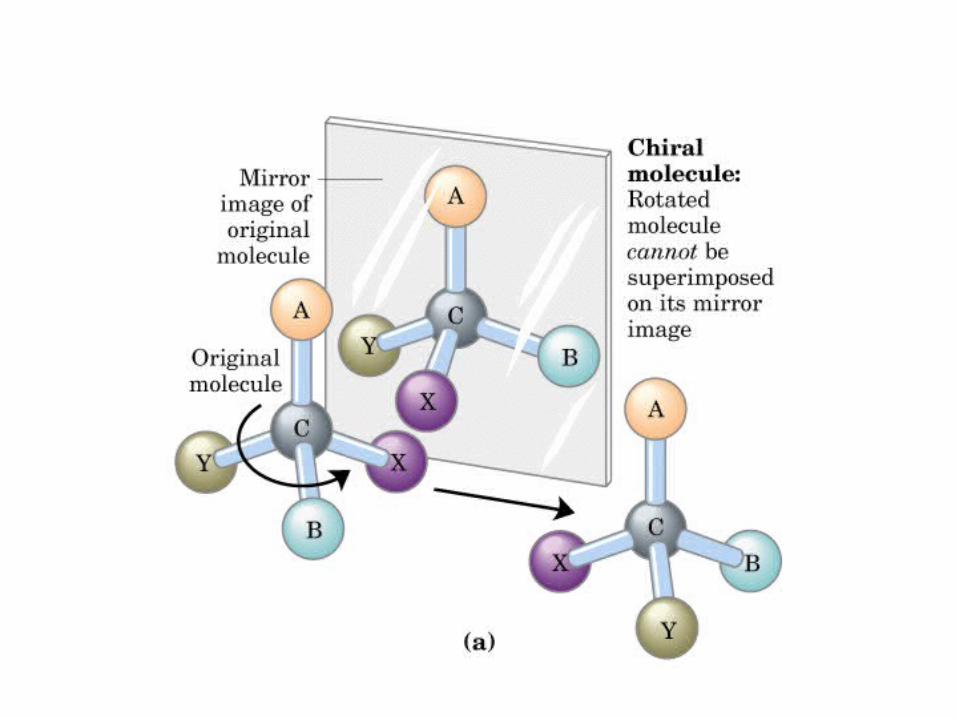
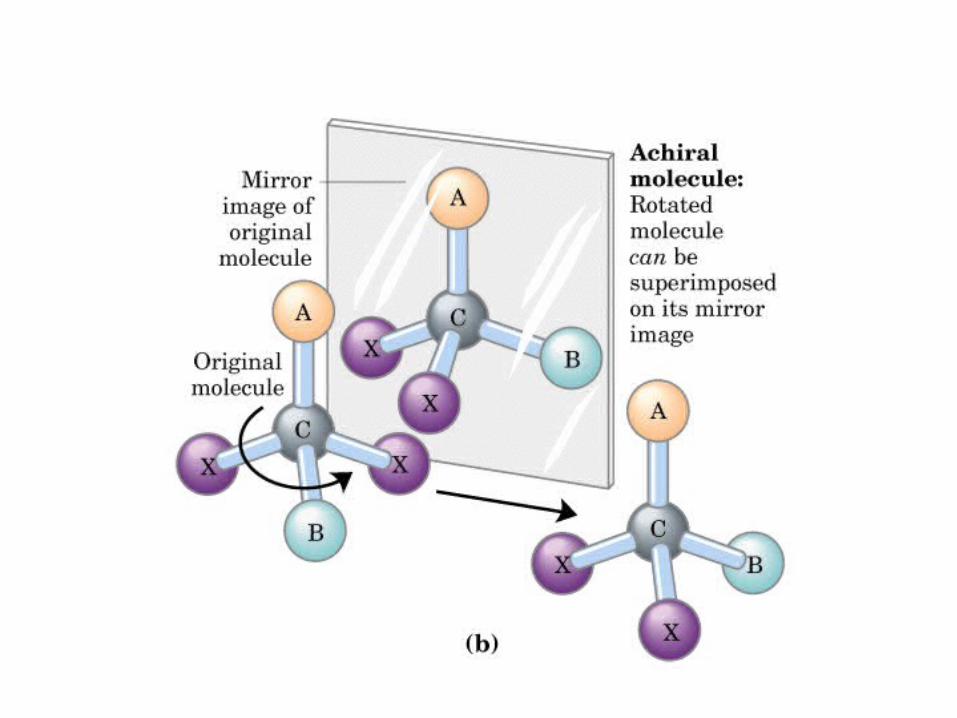
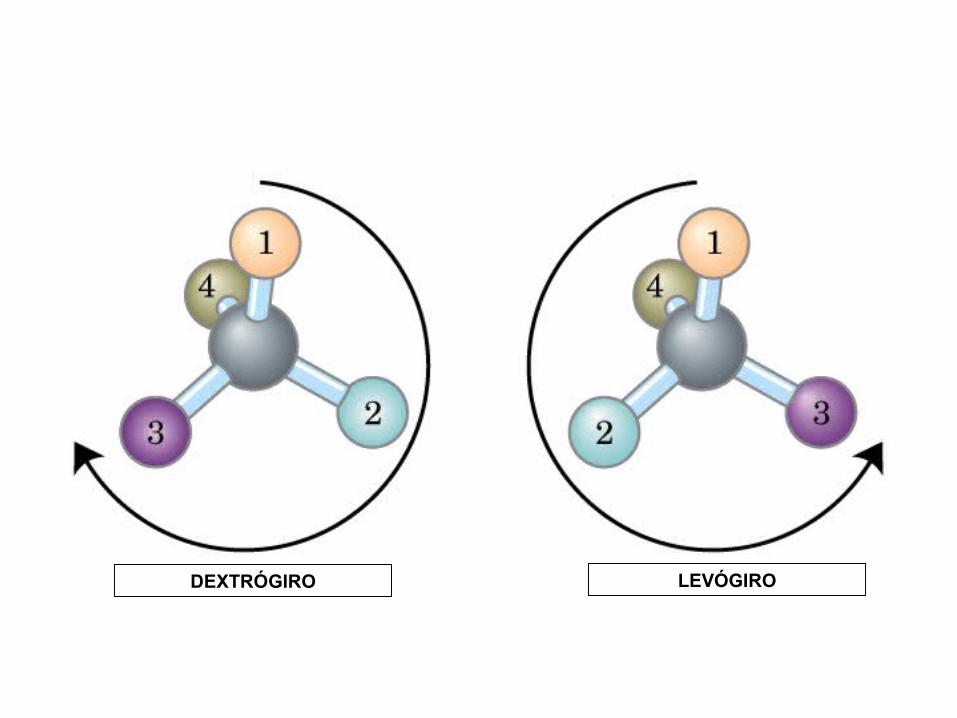
DEXTRÓGIRO LEVÓGIRO
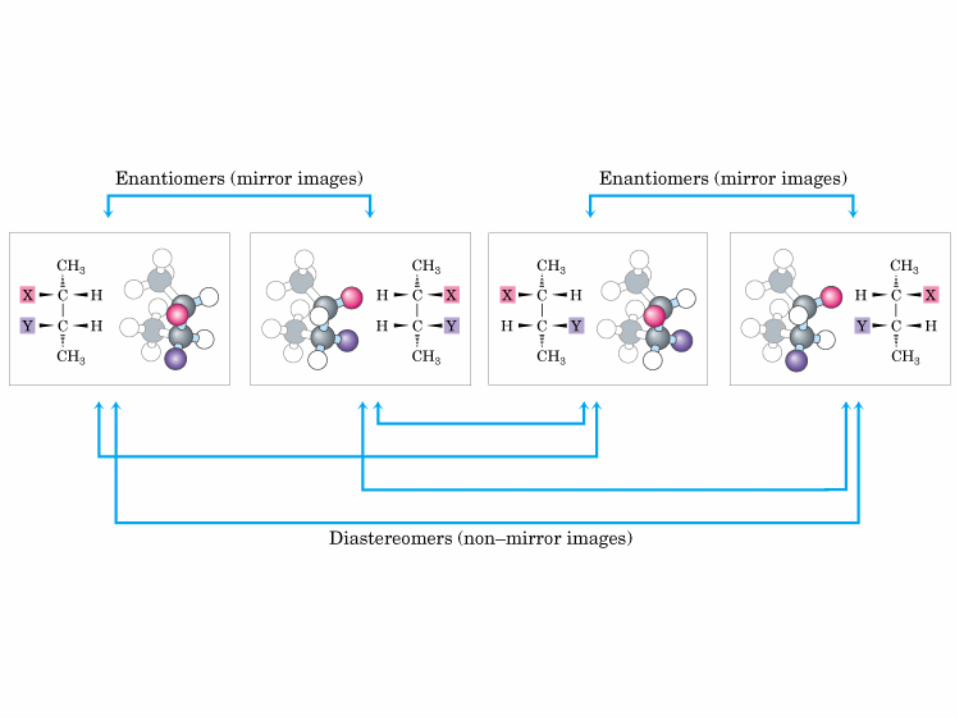
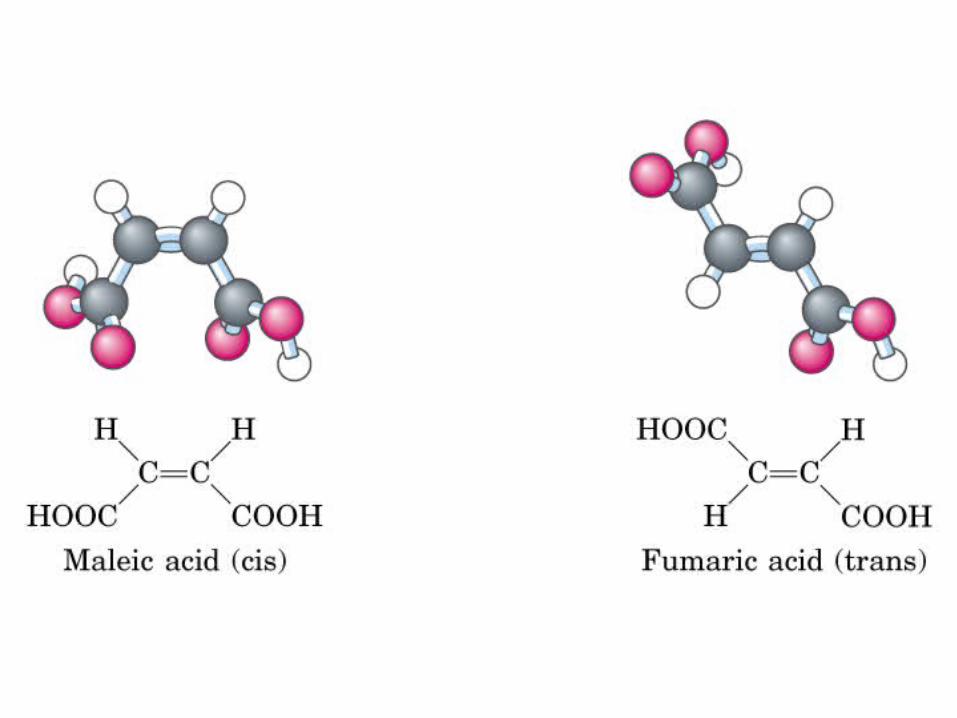

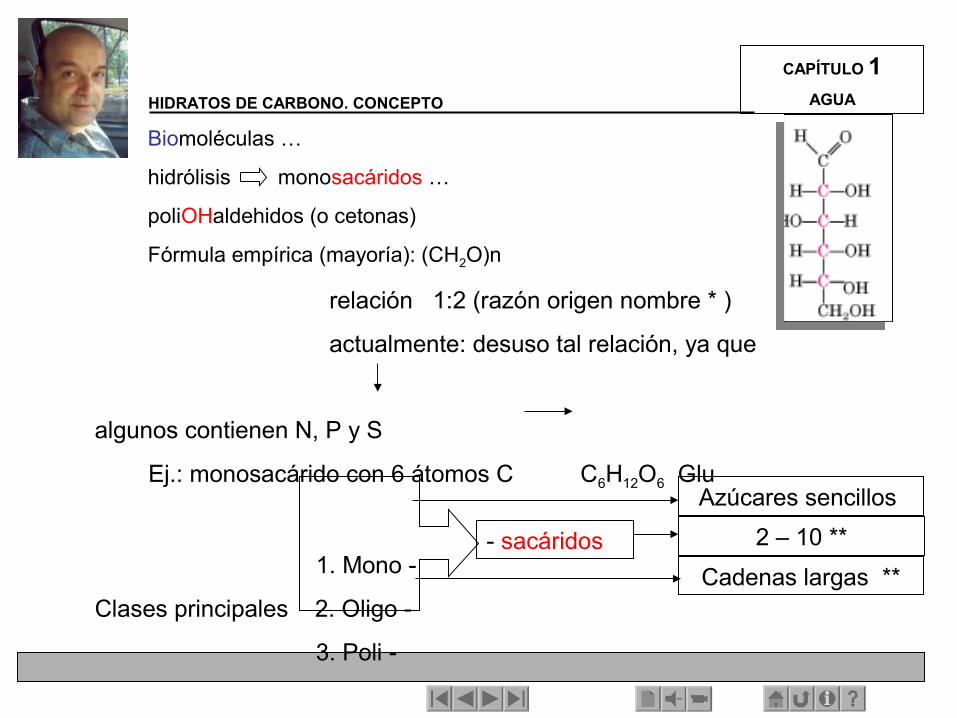
Biomoléculas …
hidrólisis monosacáridos …
poliOHaldehidos (o cetonas)
Fórmula empírica (mayoría): (CH2O)n
relación 1:2 (razón origen nombre * )
actualmente: desuso tal relación, ya que algunos contienen N, P y S
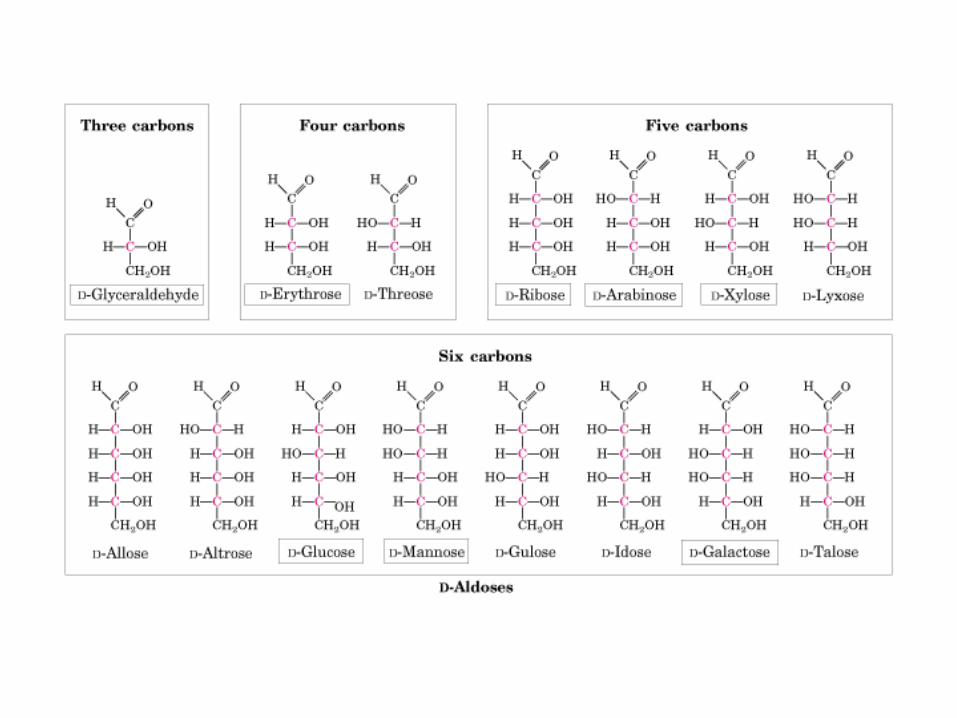
Ej.: monosacárido con 6 átomos C C6H12O6 Glu
1. Mono -
Clases principales 2. Oligo -
3. Poli -
- sacáridos
Azúcares sencillos
2 – 10 **
Cadenas largas **
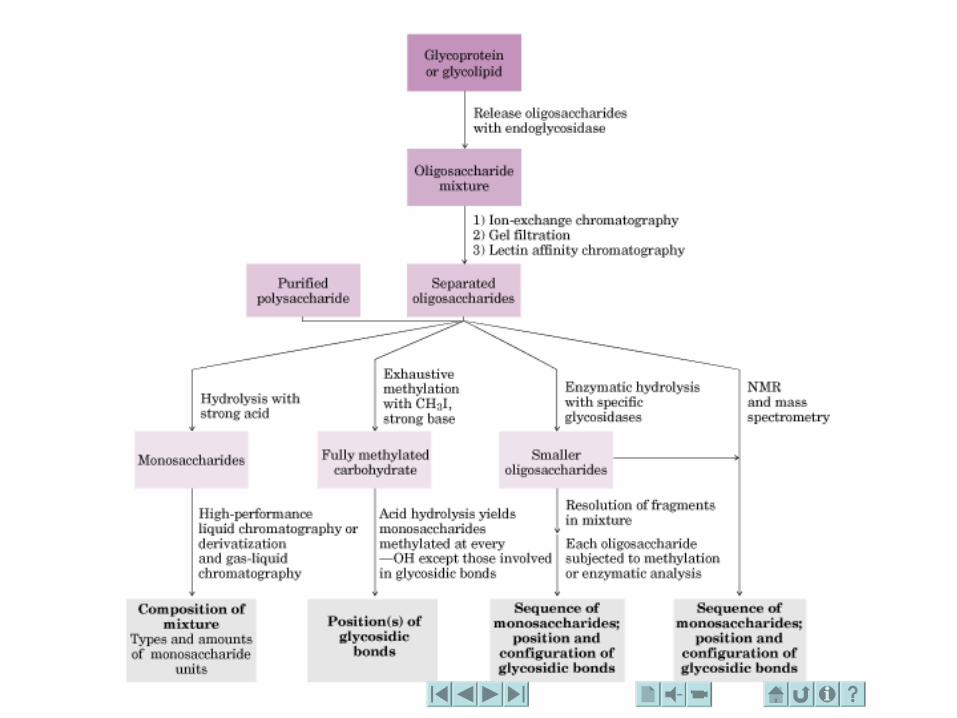
HIDRATOS DE CARBONO. CONCEPTO
CAPÍTULO 1AGUA
Med
. P
rof.
Bon
iello
, V
icen
te0101 0202GAD
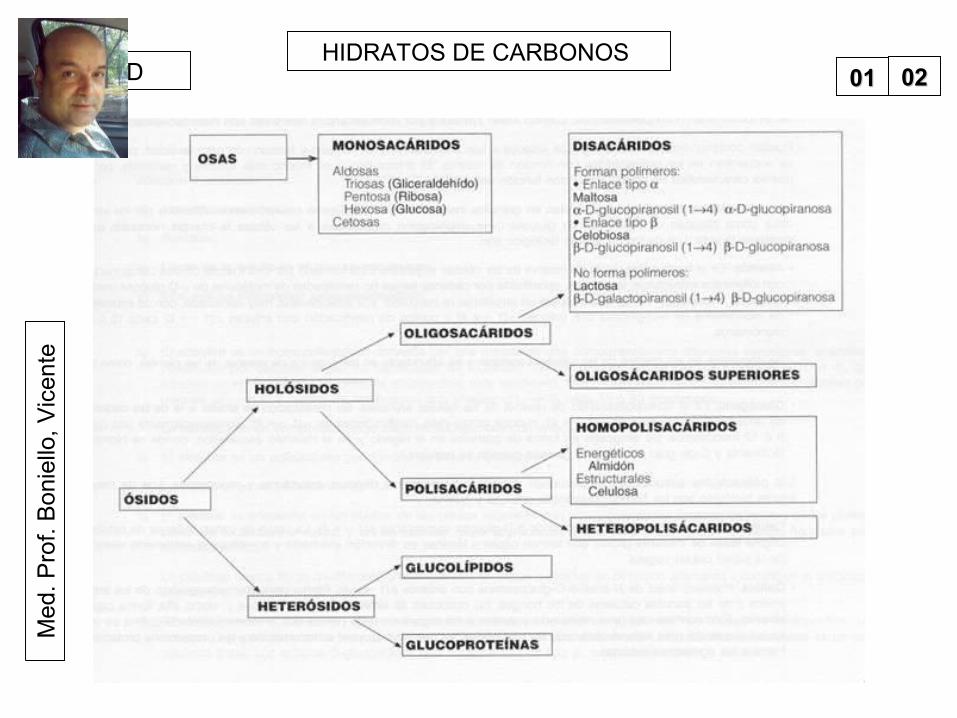
HIDRATOS DE CARBONOS
Med
. P
rof.
Bon
iello
, V
icen
te0101 0303
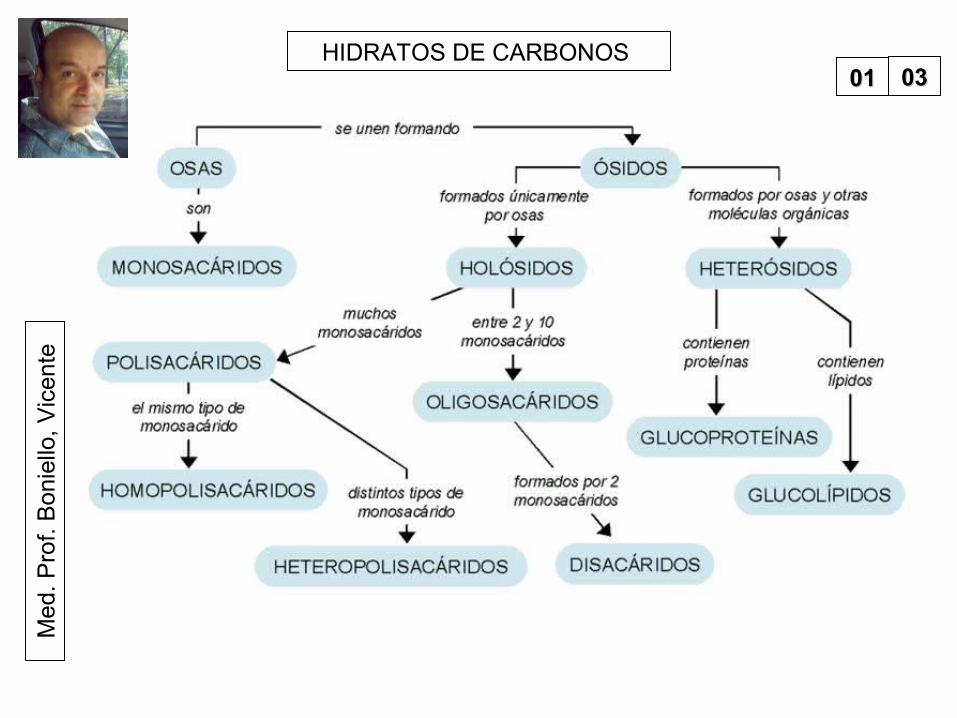
HIDRATOS DE CARBONOS
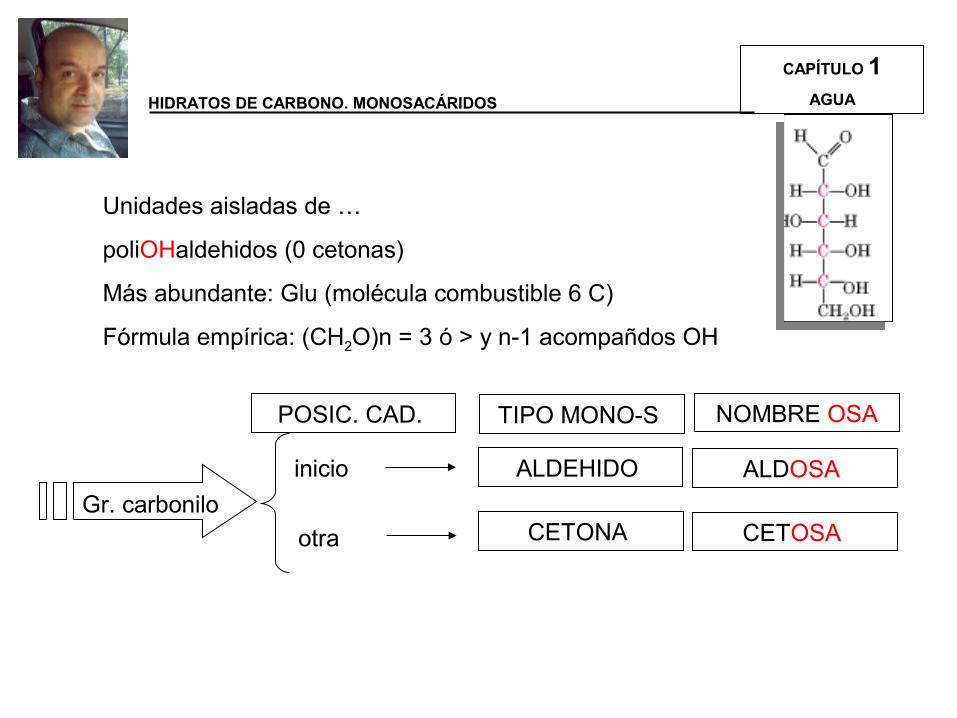
Unidades aisladas de …
poliOHaldehidos (0 cetonas)
Más abundante: Glu (molécula combustible 6 C)
Fórmula empírica: (CH2O)n = 3 ó > y n-1 acompañdos OH
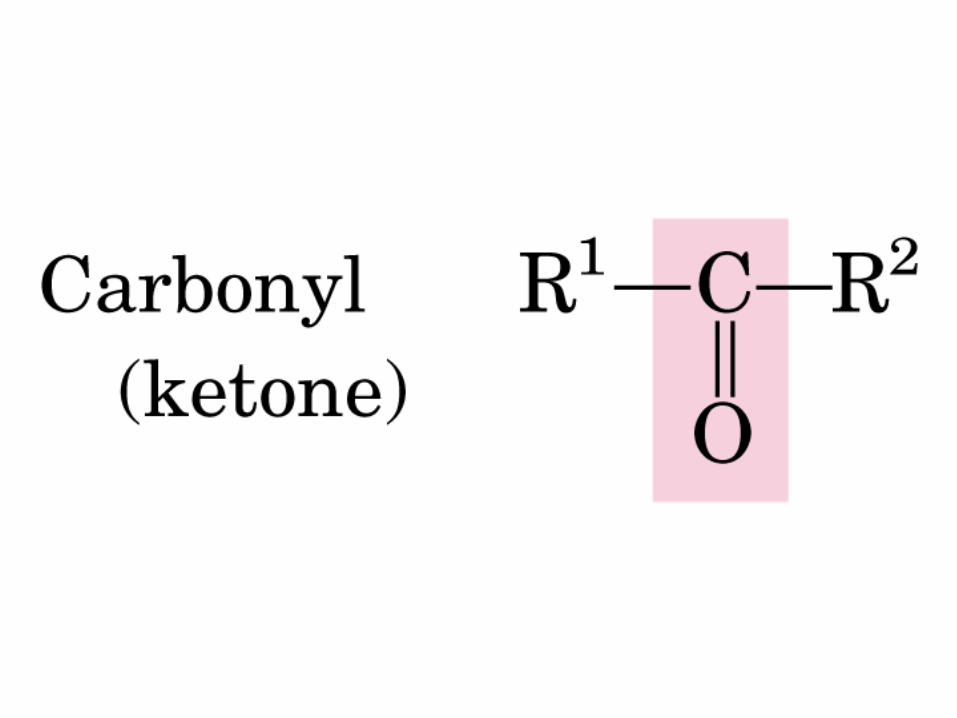
Gr. carbonilo
POSIC. CAD.
inicio
otra
ALDEHIDO
CETONA
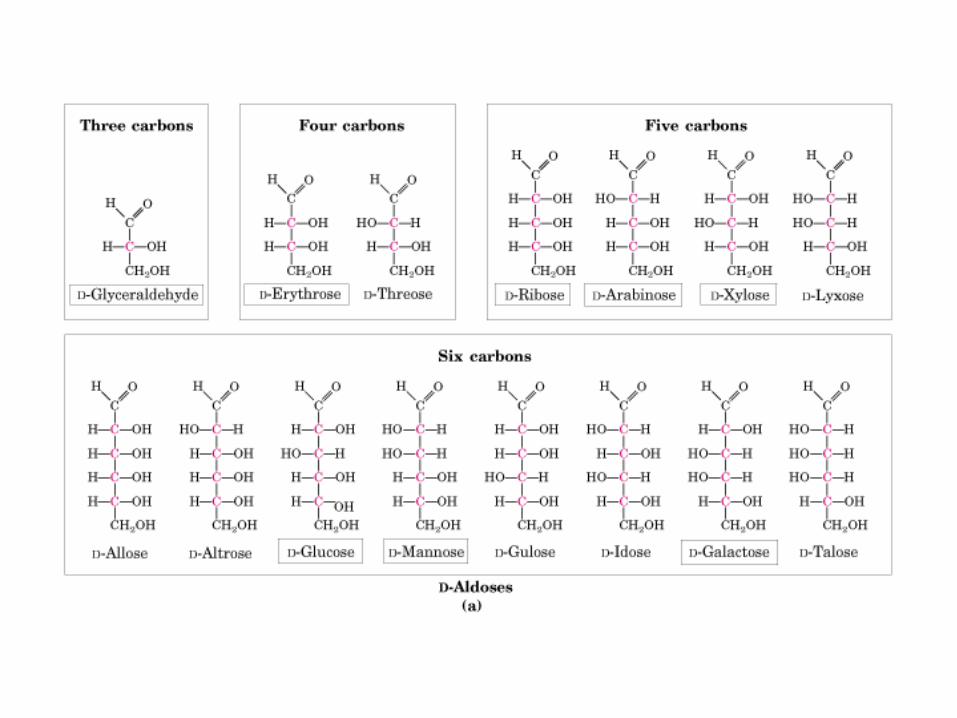
TIPO MONO-S NOMBRE OSA
ALDOSA
CETOSA
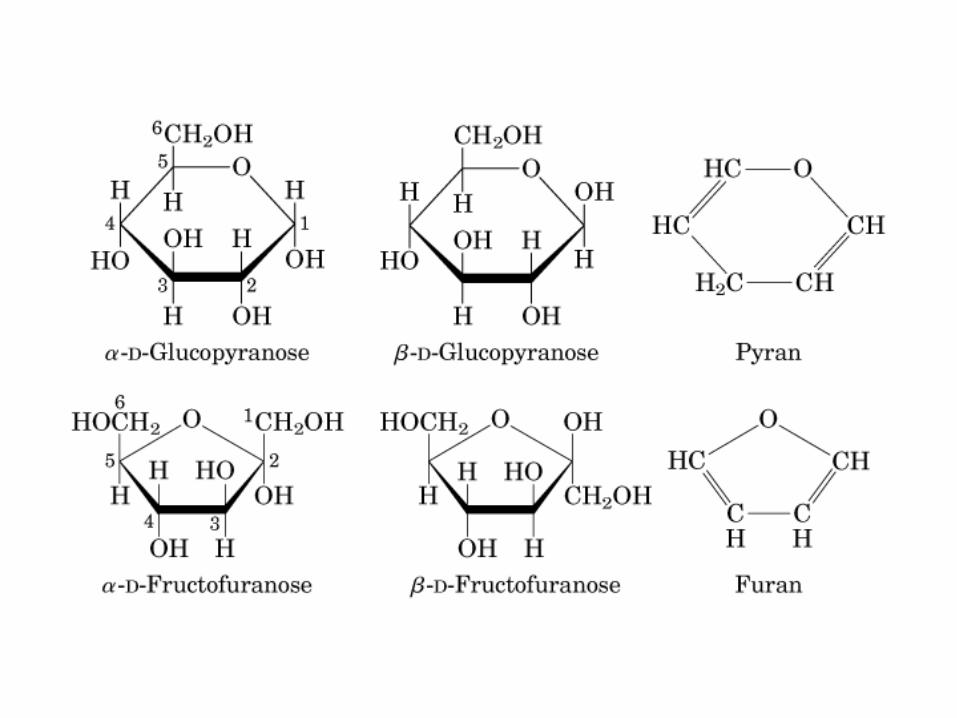
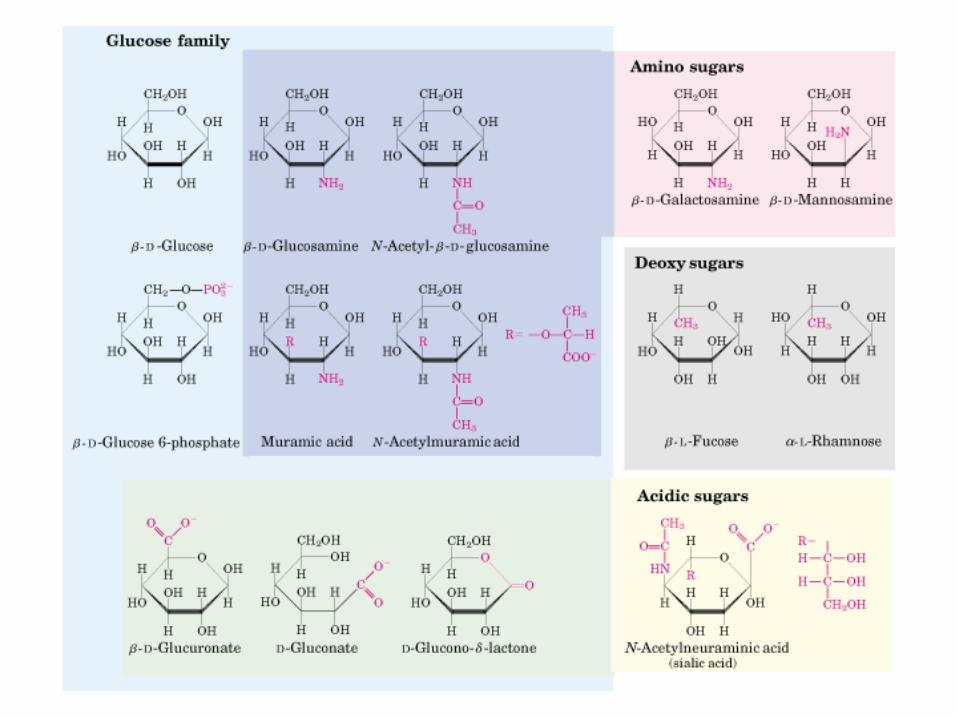
HIDRATOS DE CARBONO. MONOSACÁRIDOS
CAPÍTULO 1AGUA
Med
. P
rof.
Bon
iello
, V
icen
te0303
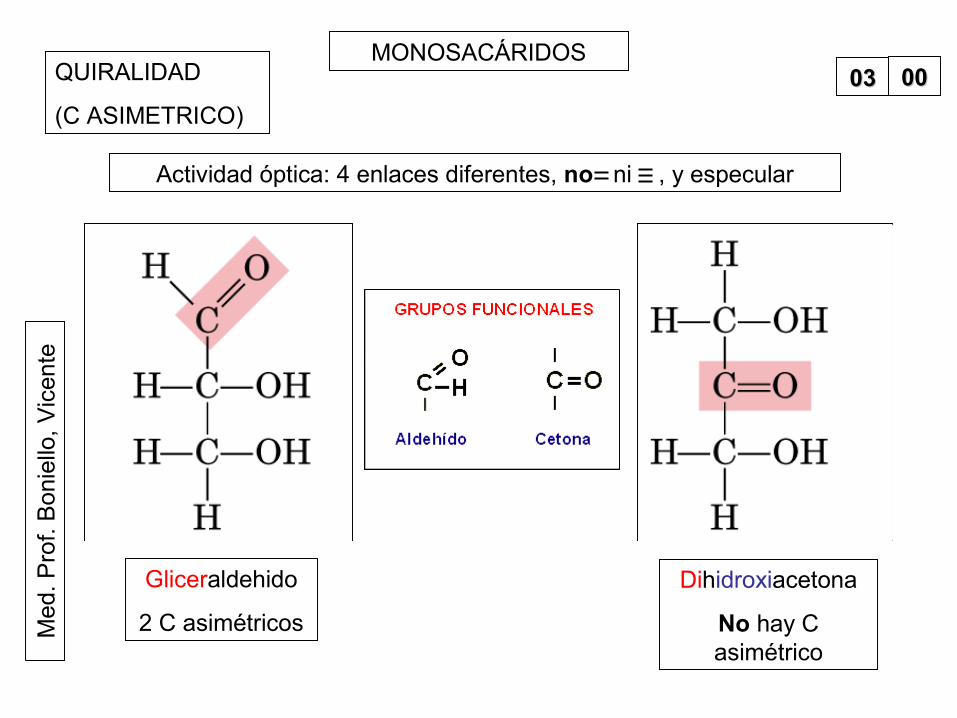
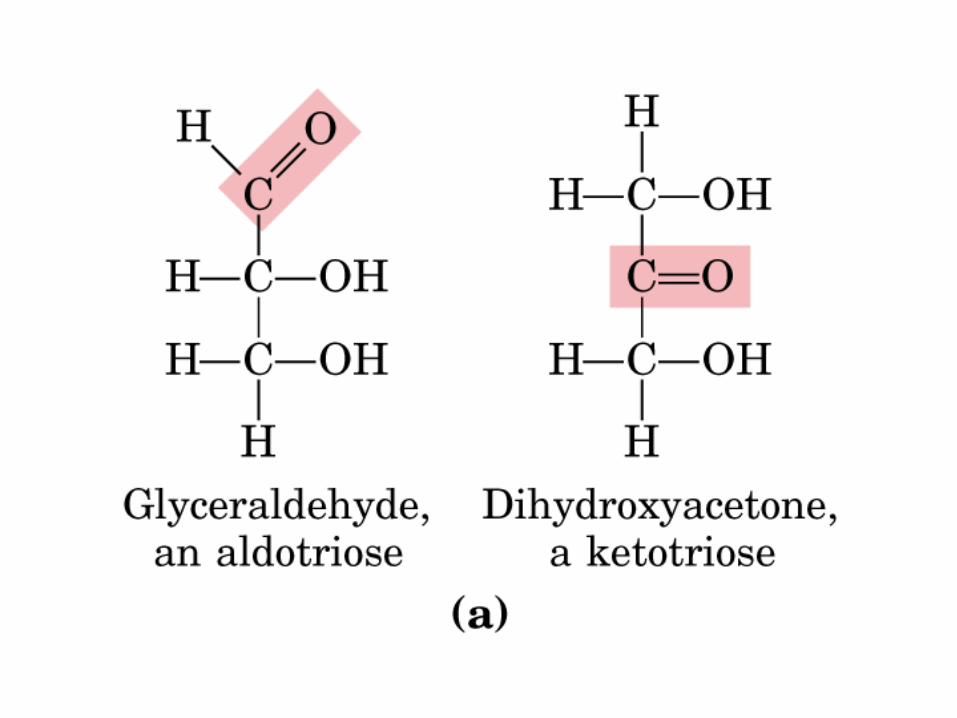
MONOSACÁRIDOS0000QUIRALIDAD
(C ASIMETRICO)
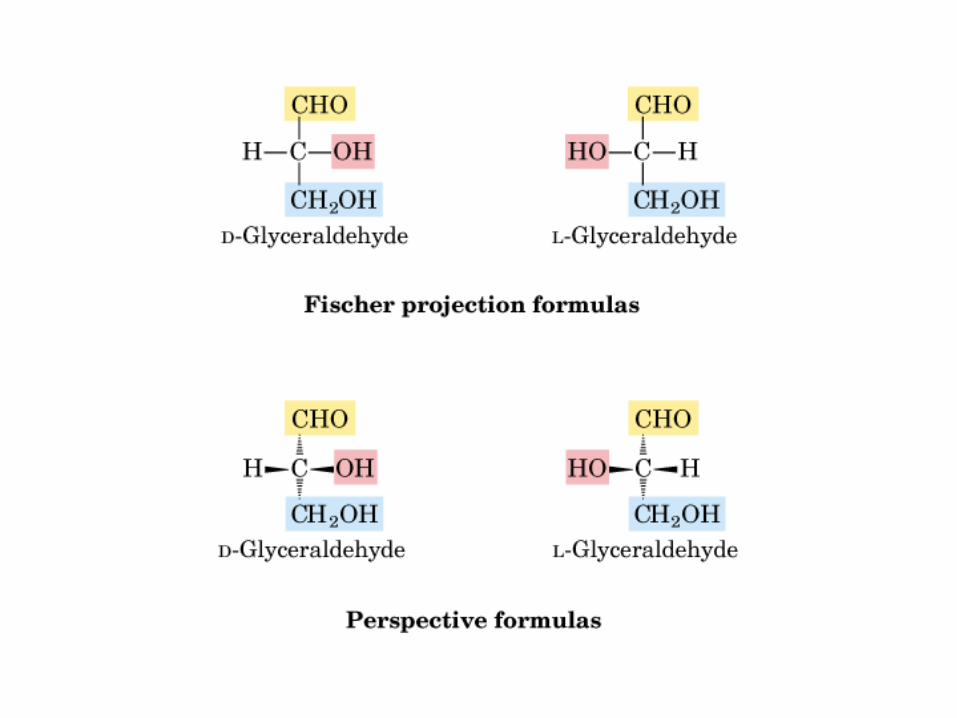
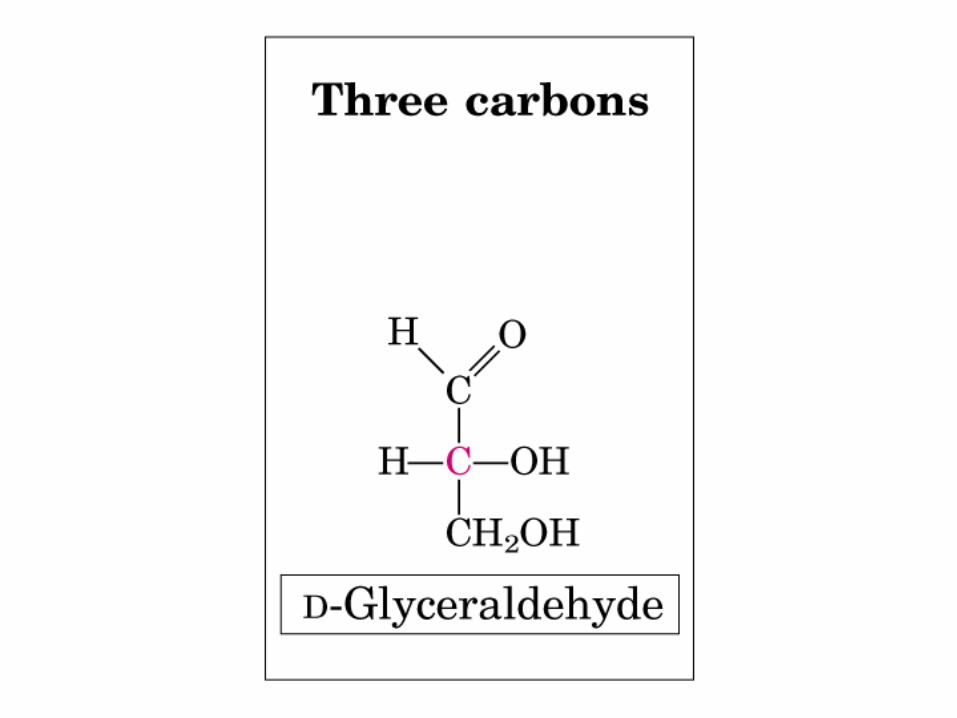
Gliceraldehido
2 C asimétricos
Dihidroxiacetona
No hay C asimétrico
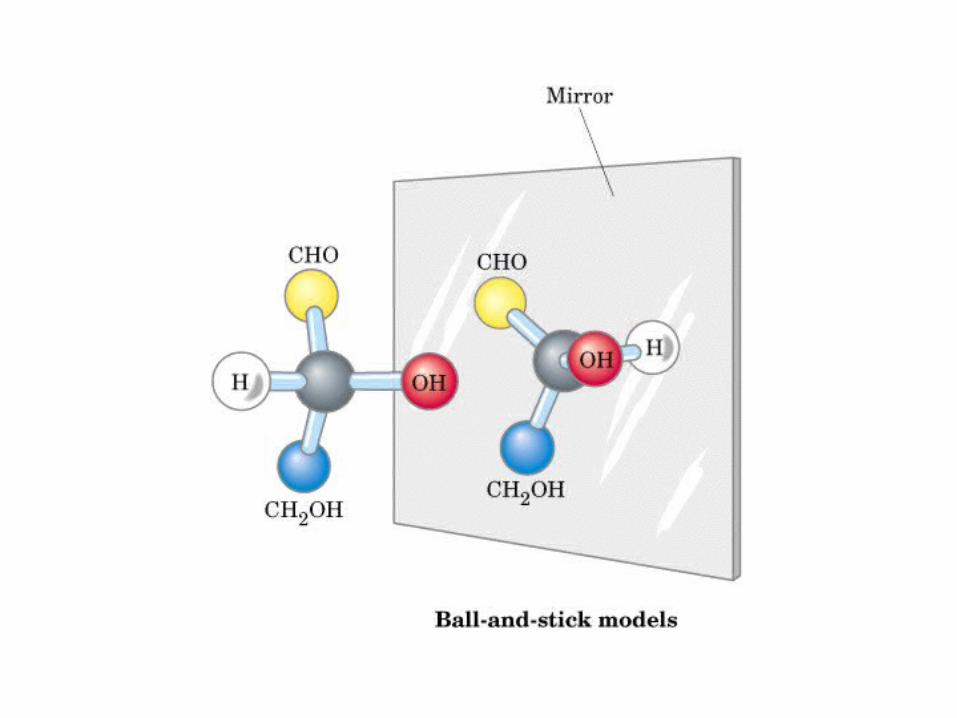
Actividad óptica: 4 enlaces diferentes, no ni , y especular
Med
. P
rof.
Bon
iello
, V
icen
te0404
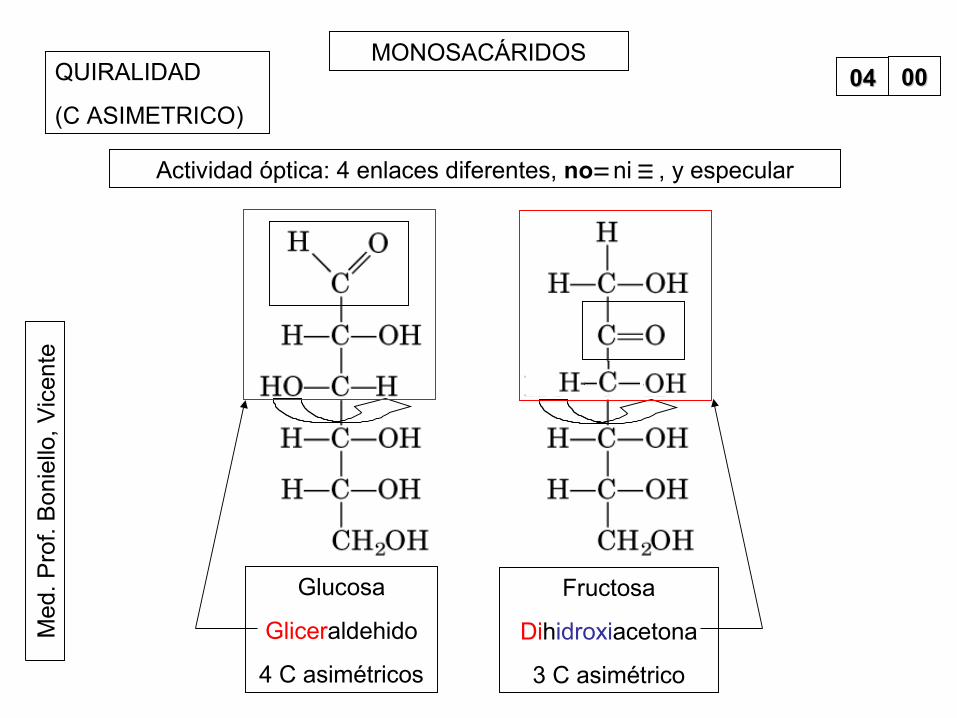
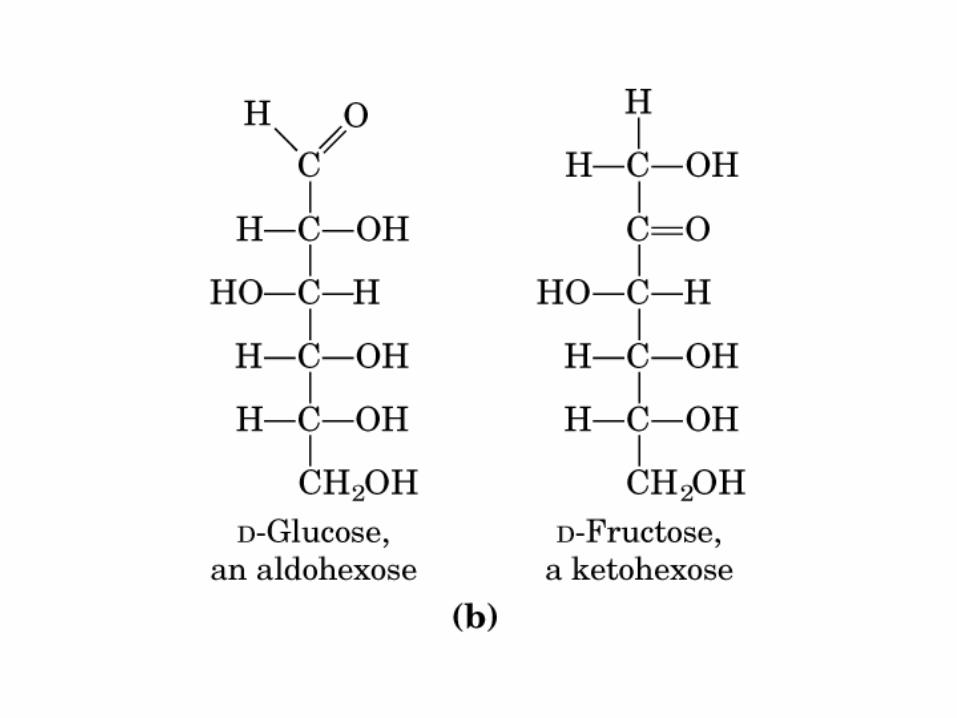
MONOSACÁRIDOS0000QUIRALIDAD
(C ASIMETRICO)
Glucosa
Gliceraldehido
4 C asimétricos
Fructosa
Dihidroxiacetona
3 C asimétrico
Actividad óptica: 4 enlaces diferentes, no ni , y especular
Med
. P
rof.
Bon
iello
, V
icen
teMONOSACÁRIDOS
0505 0000DD LL
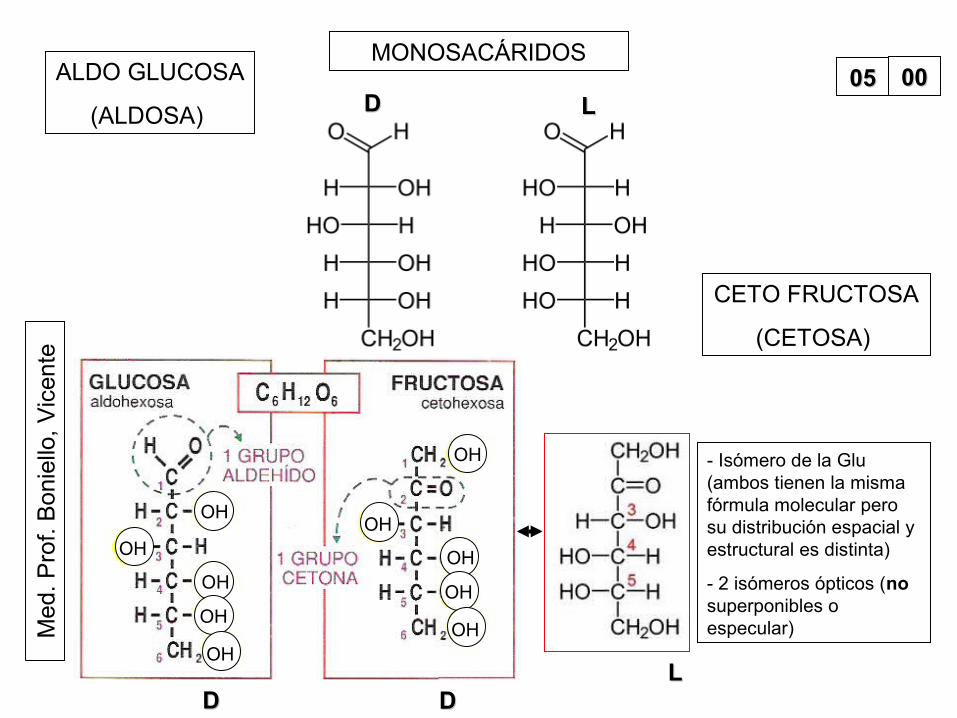
ALDO GLUCOSA
(ALDOSA)
OH
OH
OH
OH
OH
CETO FRUCTOSA
(CETOSA)
OH
OH
OH
OH
OH
DD DDLL
- Isómero de la Glu (ambos tienen la misma fórmula molecular pero su distribución espacial y estructural es distinta)
- 2 isómeros ópticos (no superponibles o especular)
Med
. P
rof.
Bon
iello
, V
icen
teMONOSACÁRIDOS
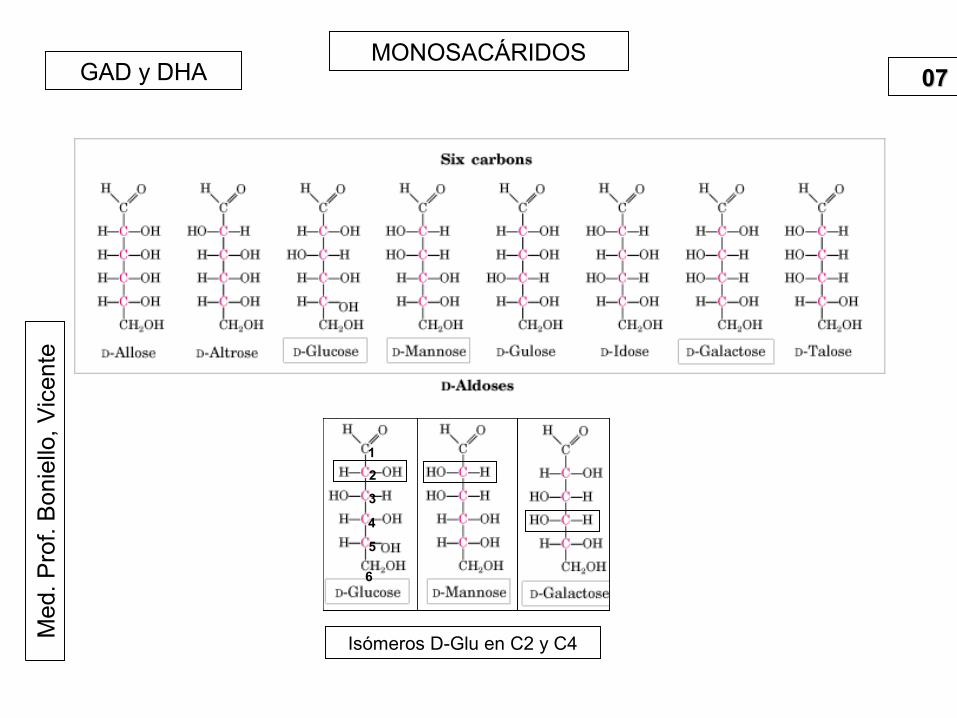
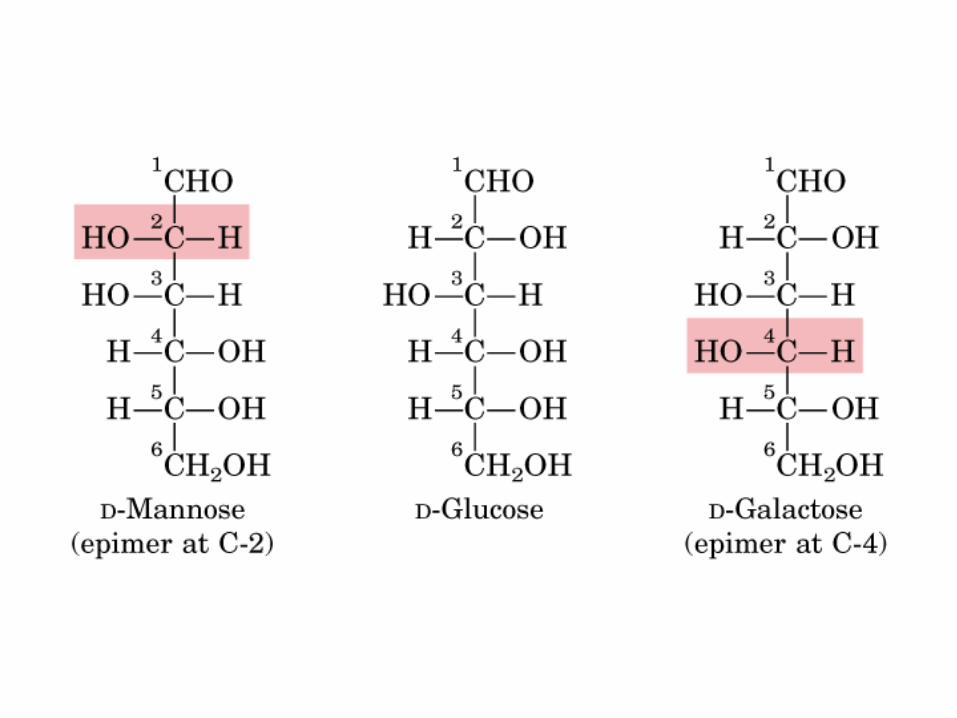
0707GAD y DHA
Isómeros D-Glu en C2 y C4
1
2
3
4
5
6
Med
. P
rof.
Bon
iello
, V
icen
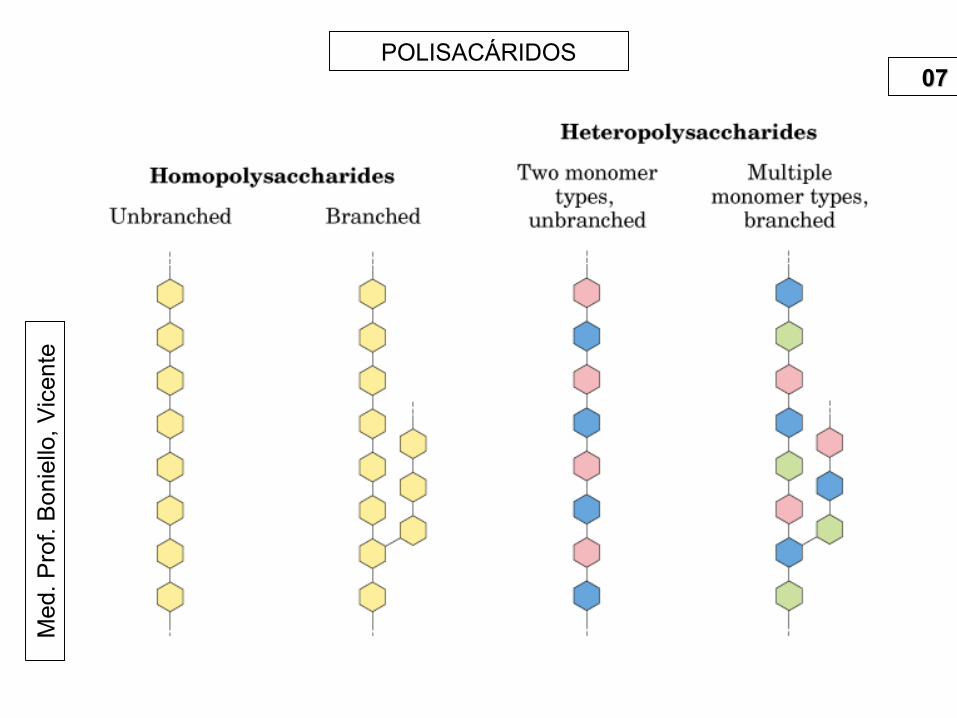
tePOLISACÁRIDOS
0707
Med
. P
rof.
Bon
iello
, V
icen
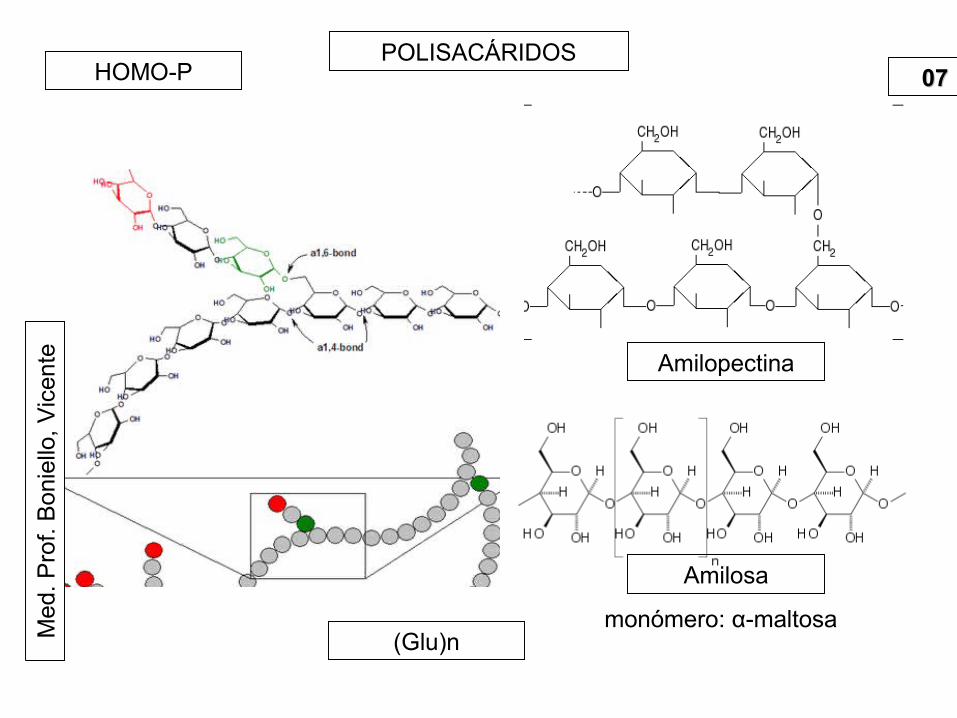
tePOLISACÁRIDOS
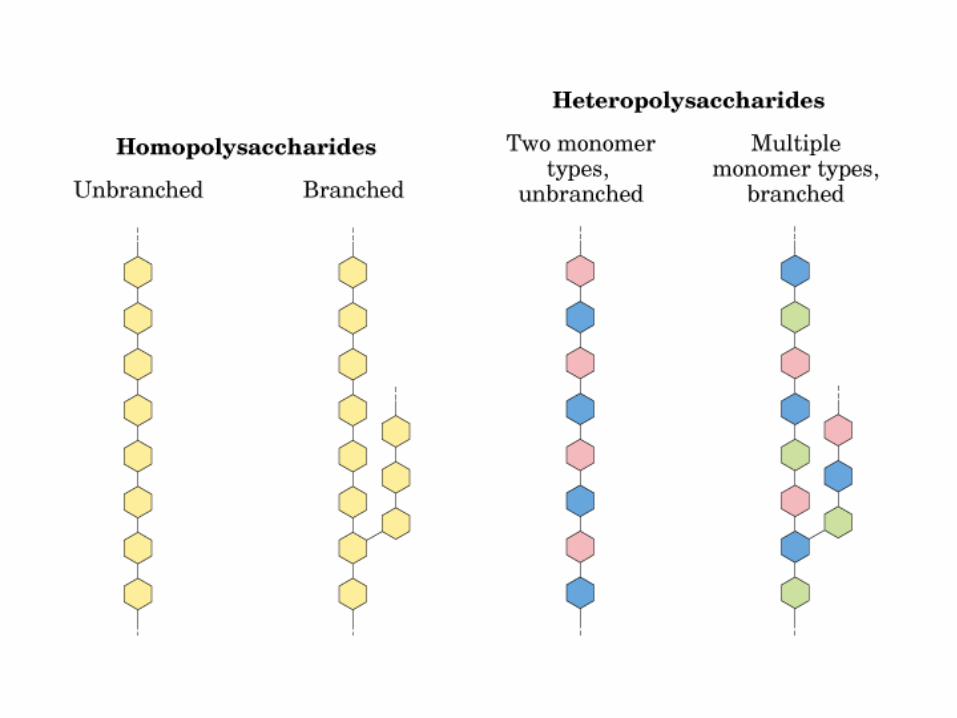
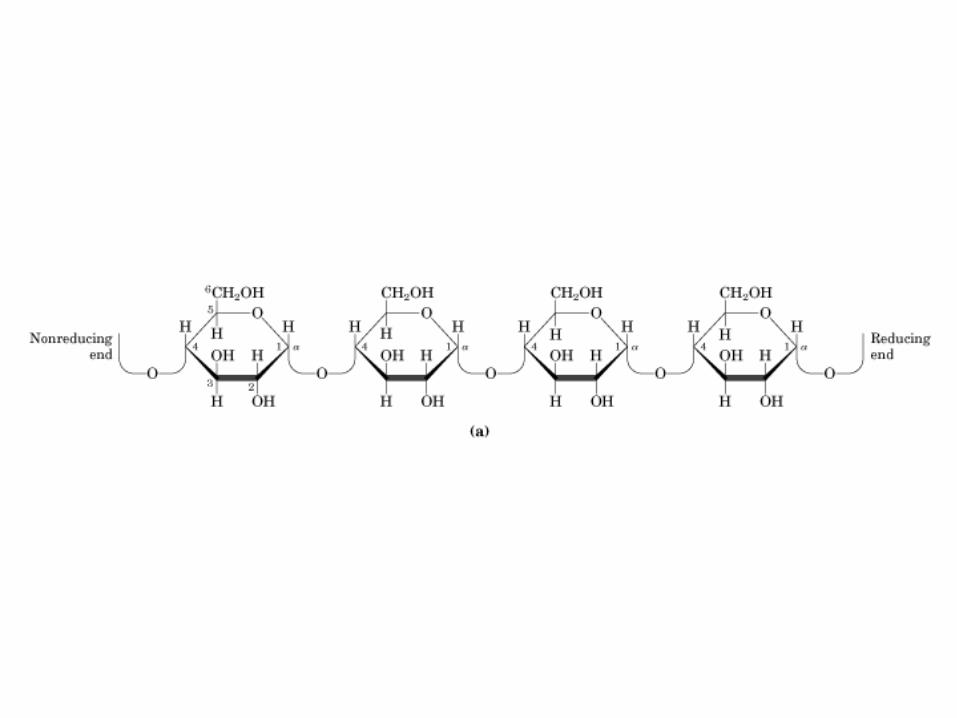
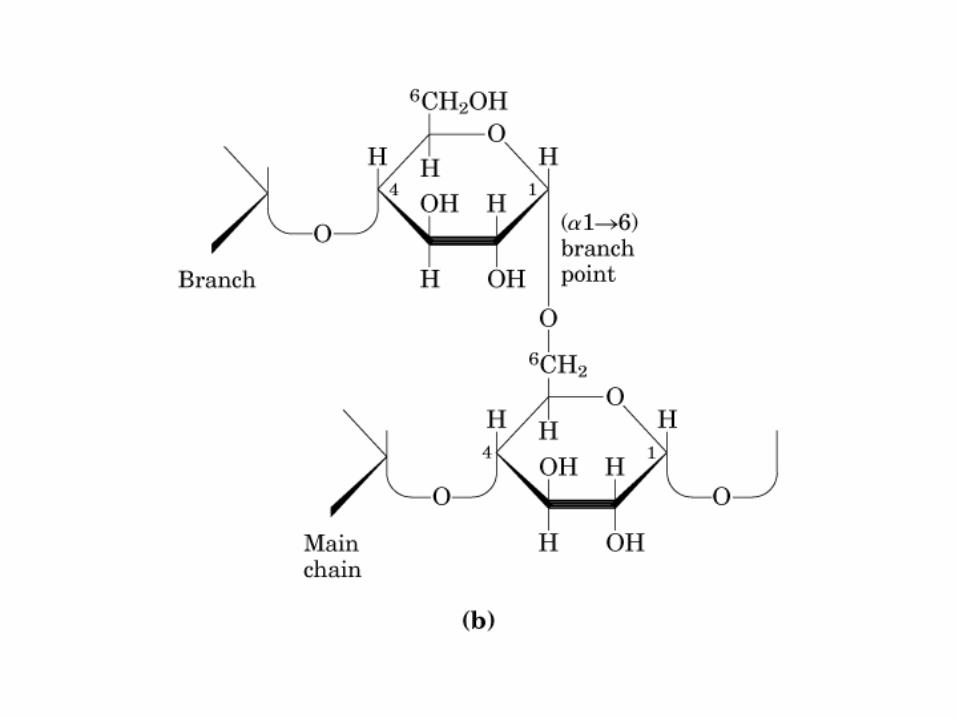
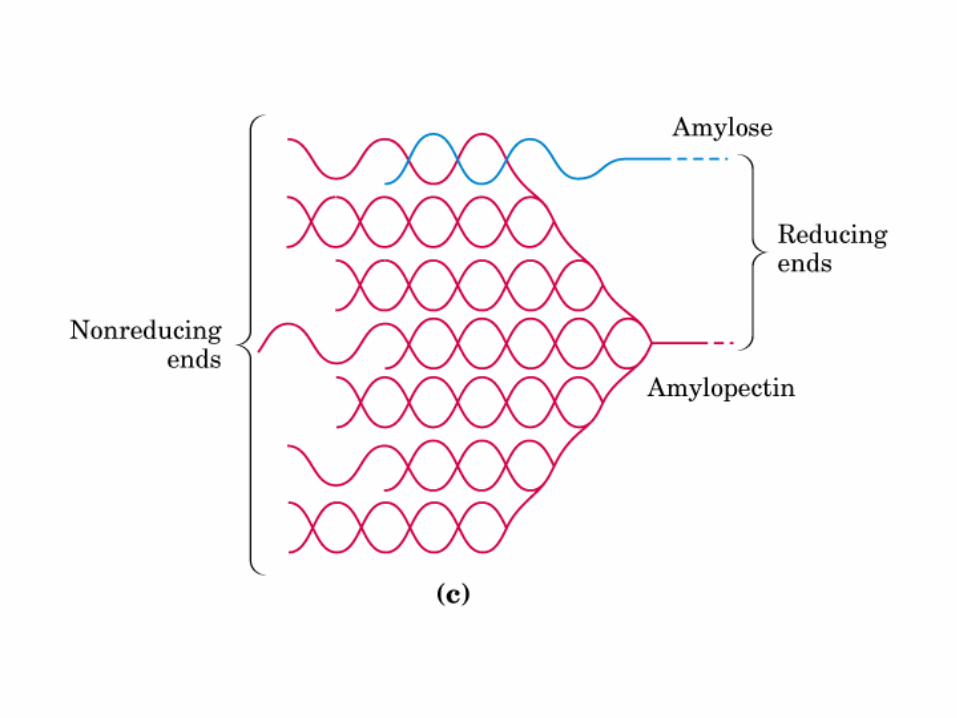
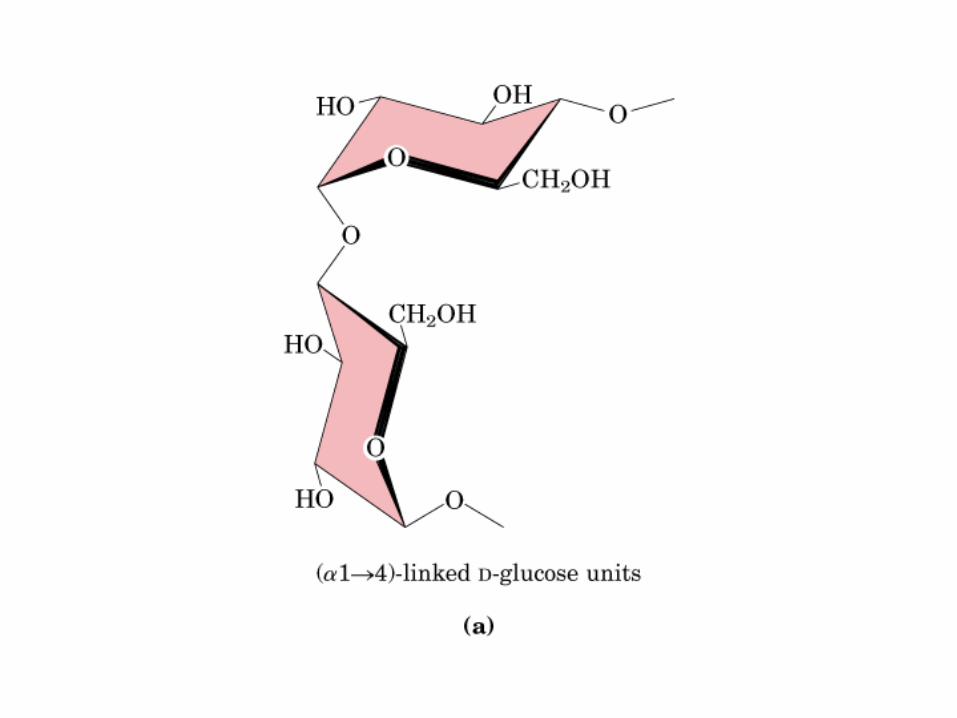
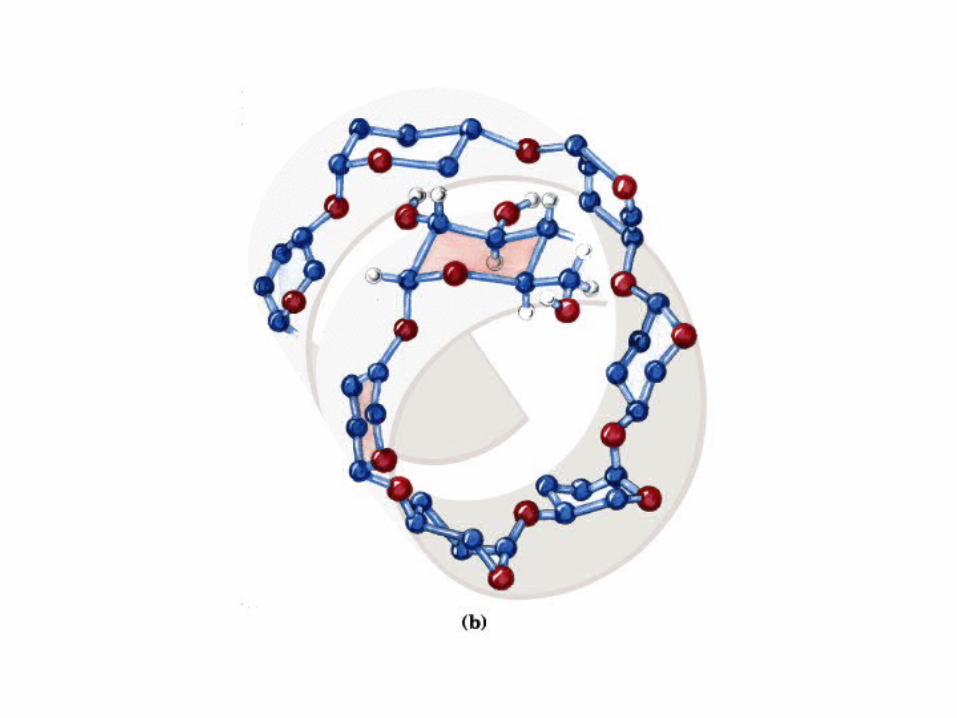
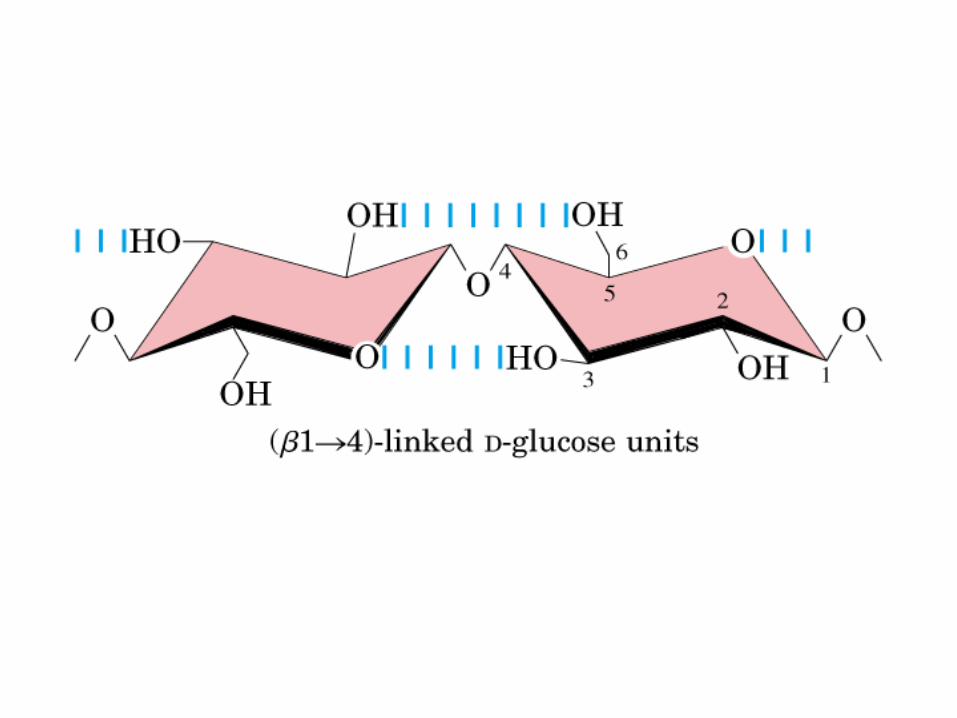
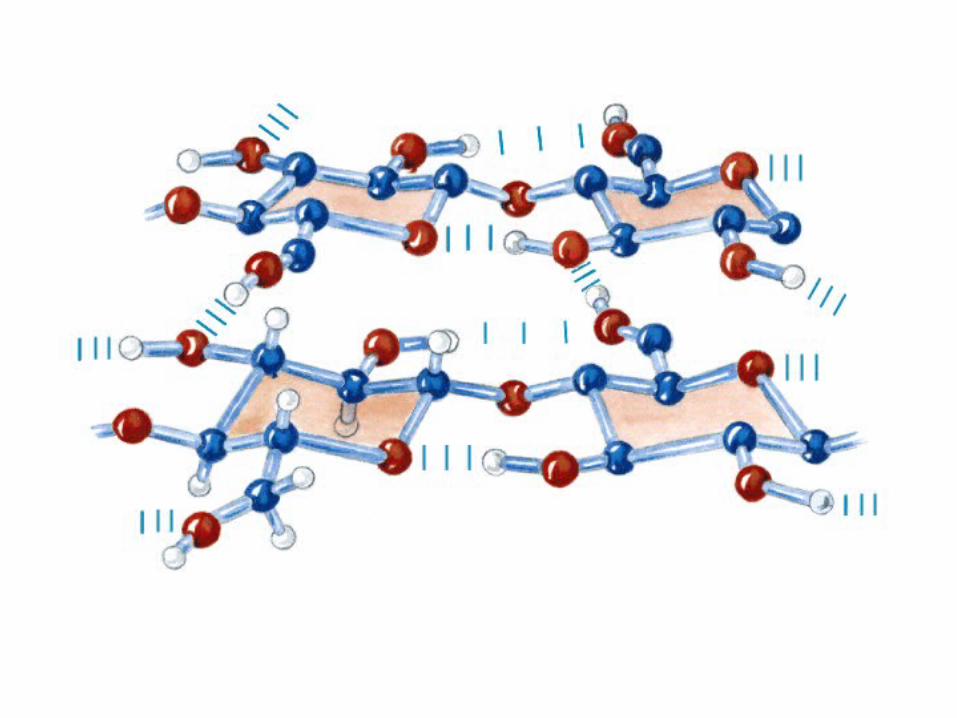
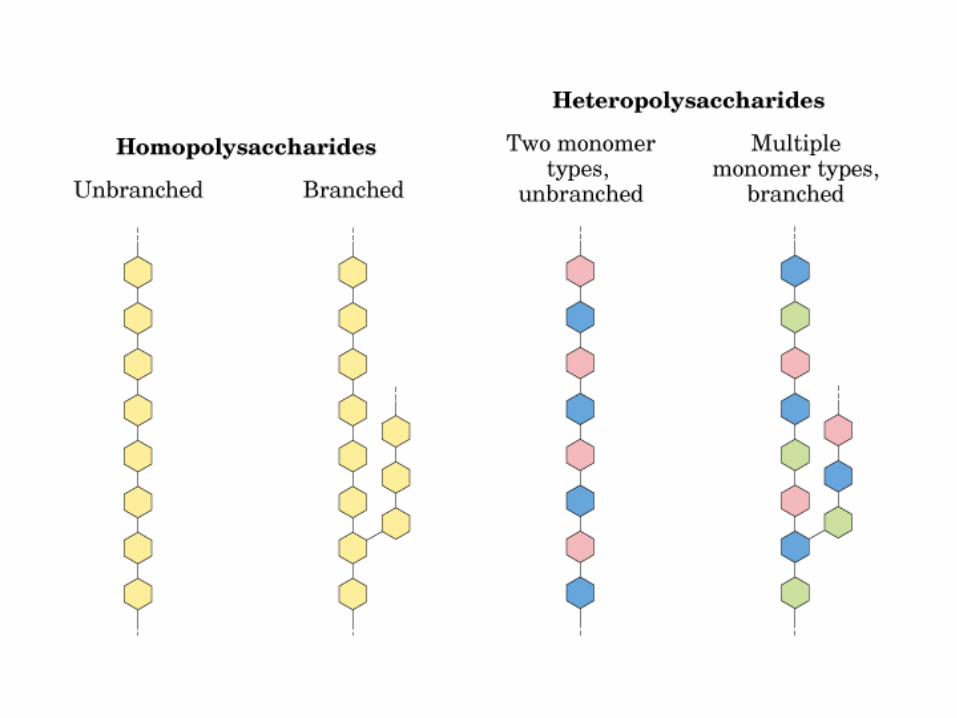
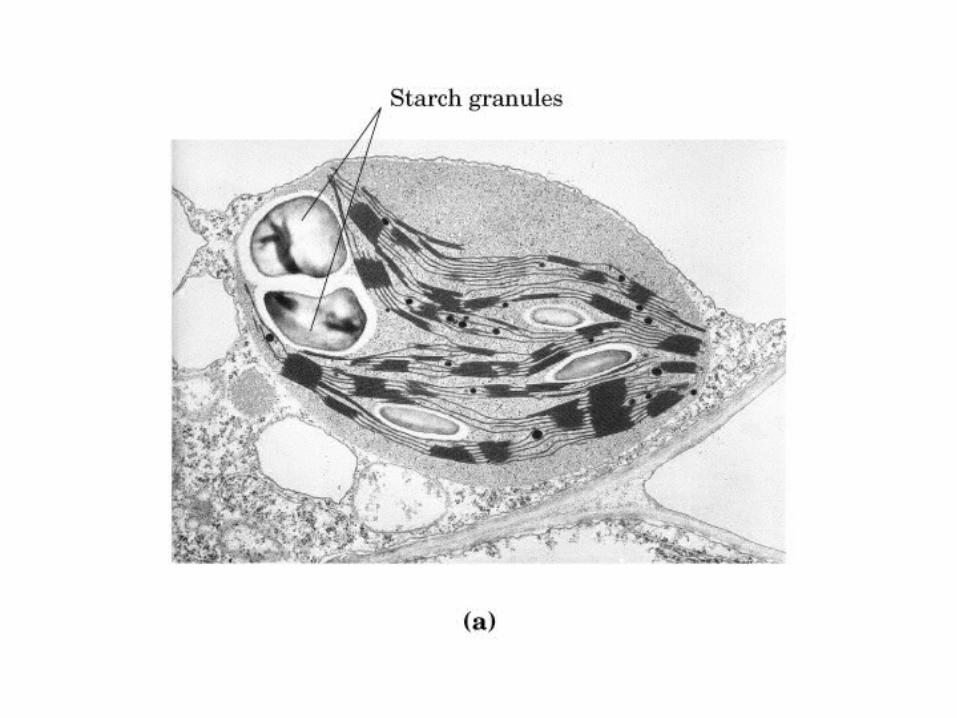
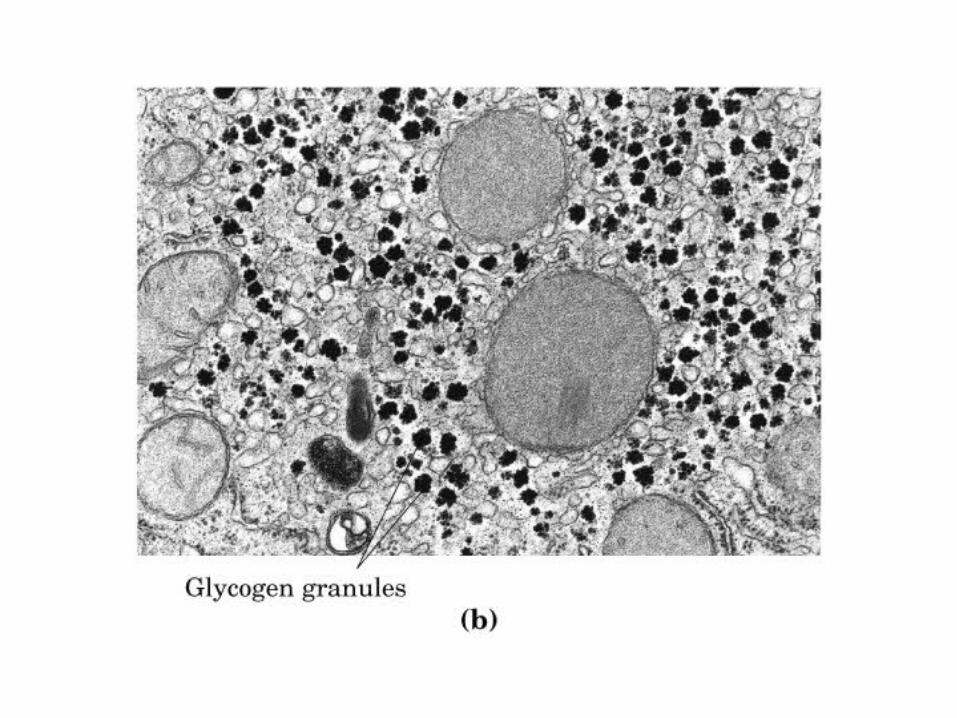
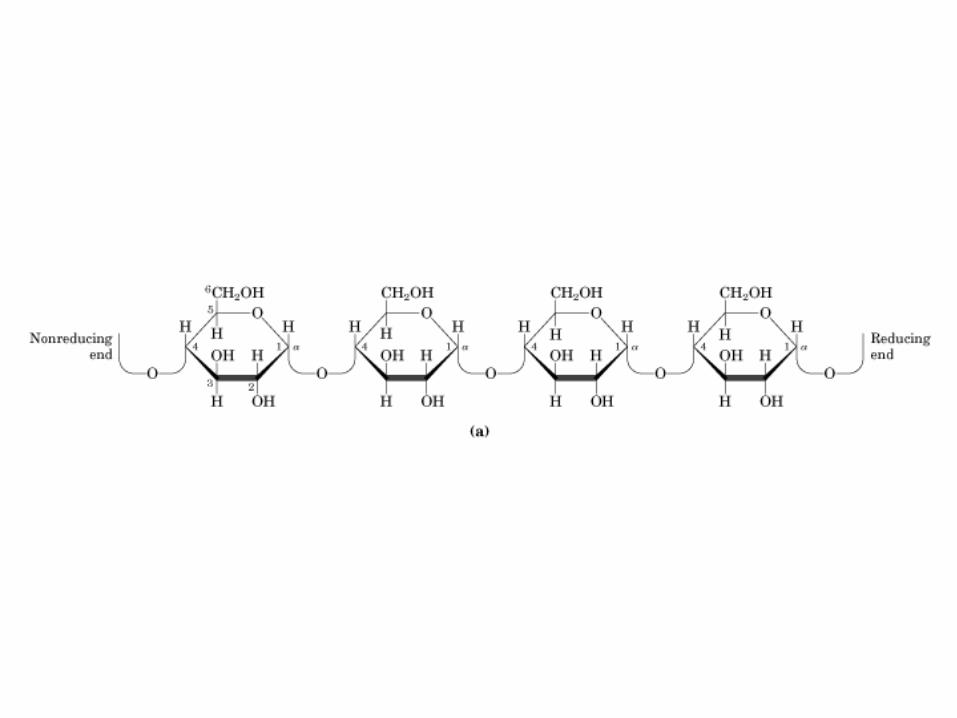
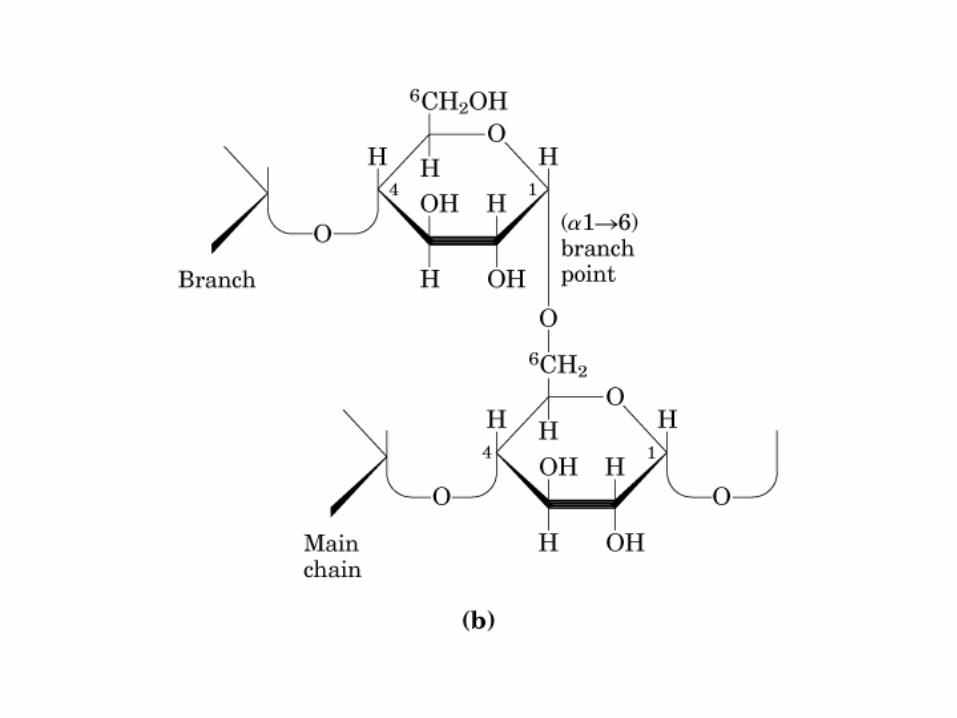
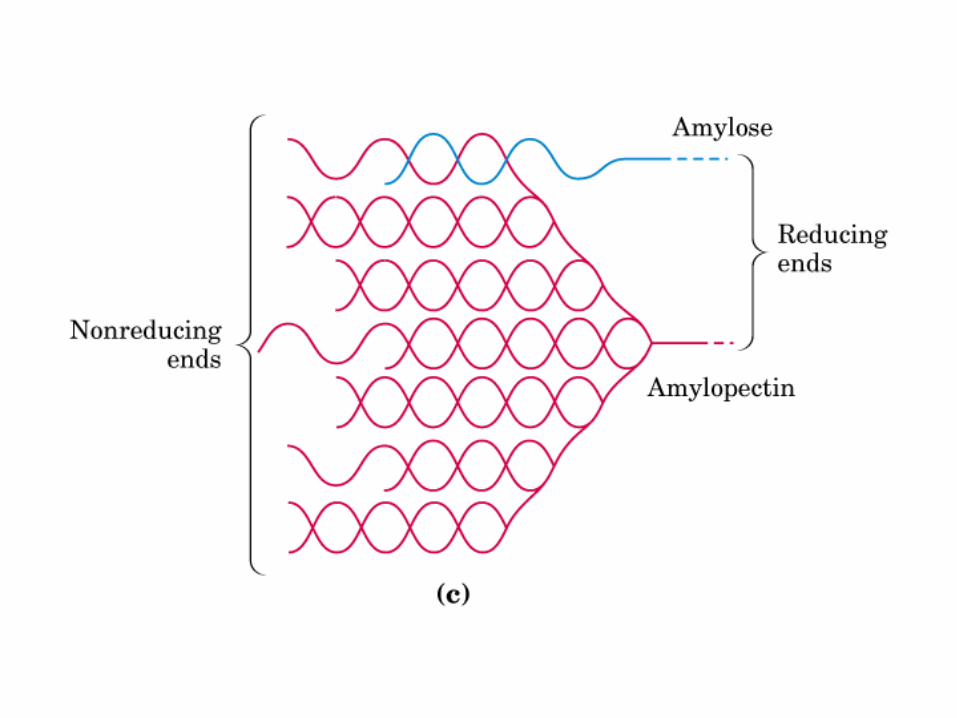
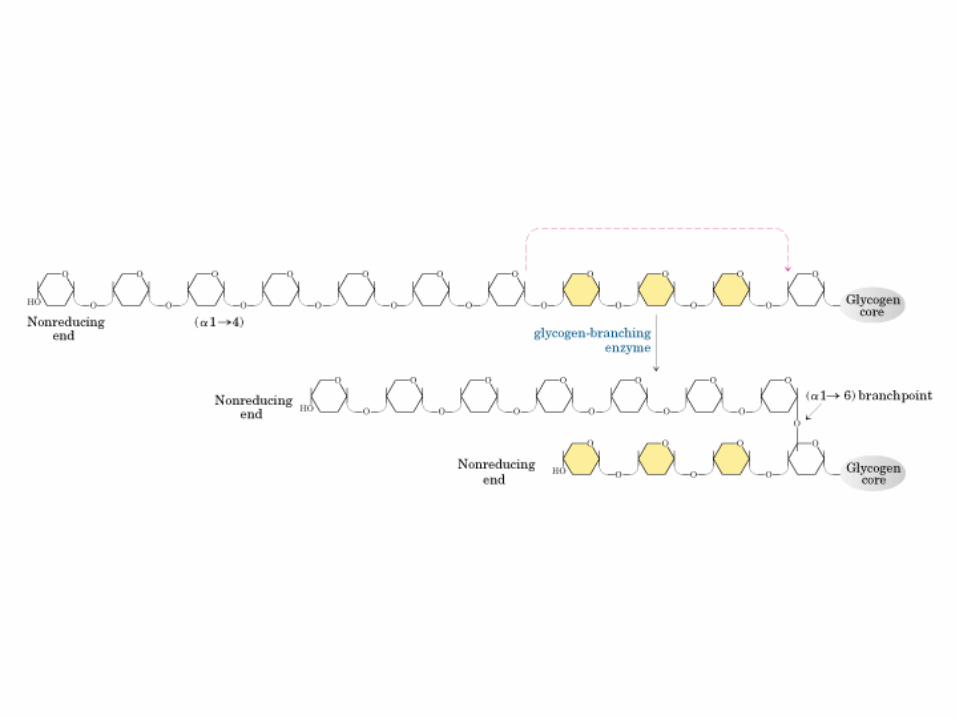
0707HOMO-P
(Glu)n
Amilopectina
monómero: α-maltosa
Amilosa
Med
. P
rof.
Bon
iello
, V
icen
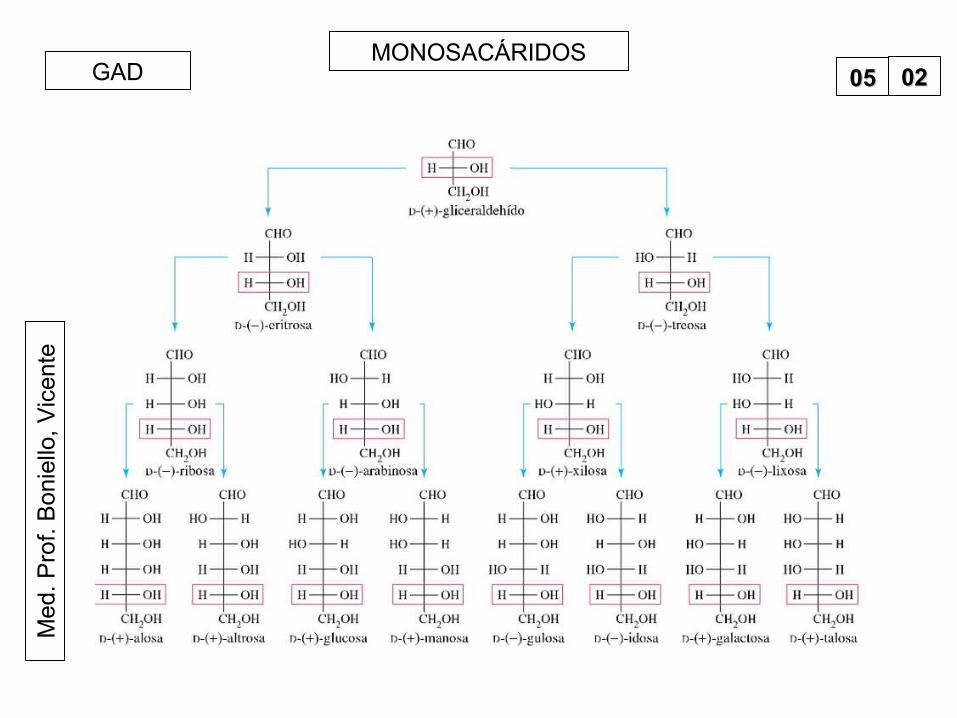
teMONOSACÁRIDOS
0505 0202GAD
Med
. P
rof.
Bon
iello
, V
icen
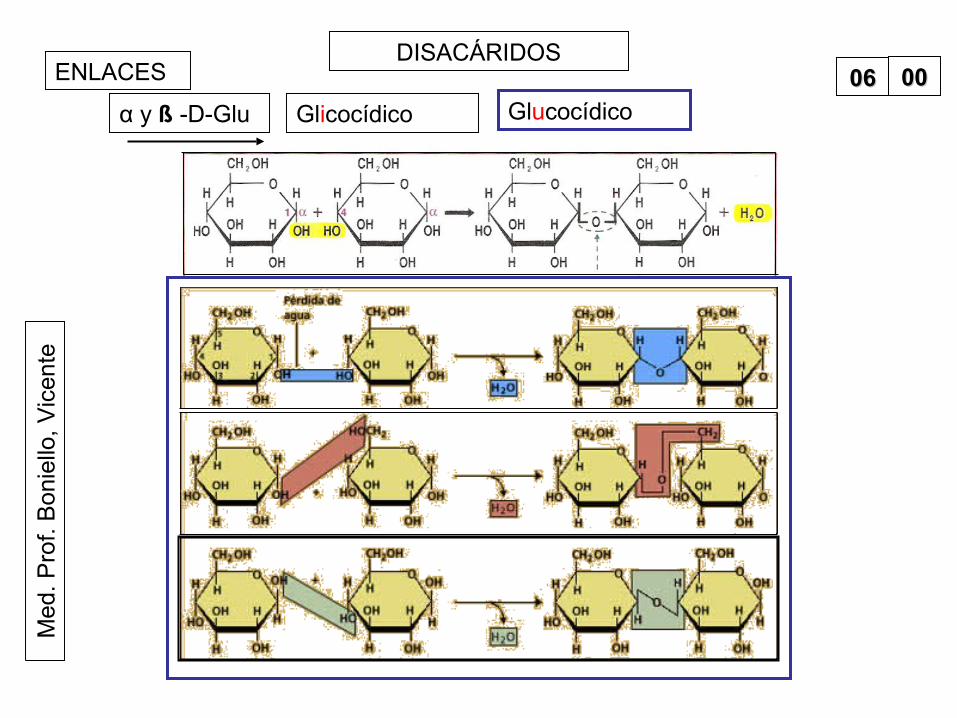
teDISACÁRIDOS
ENLACES
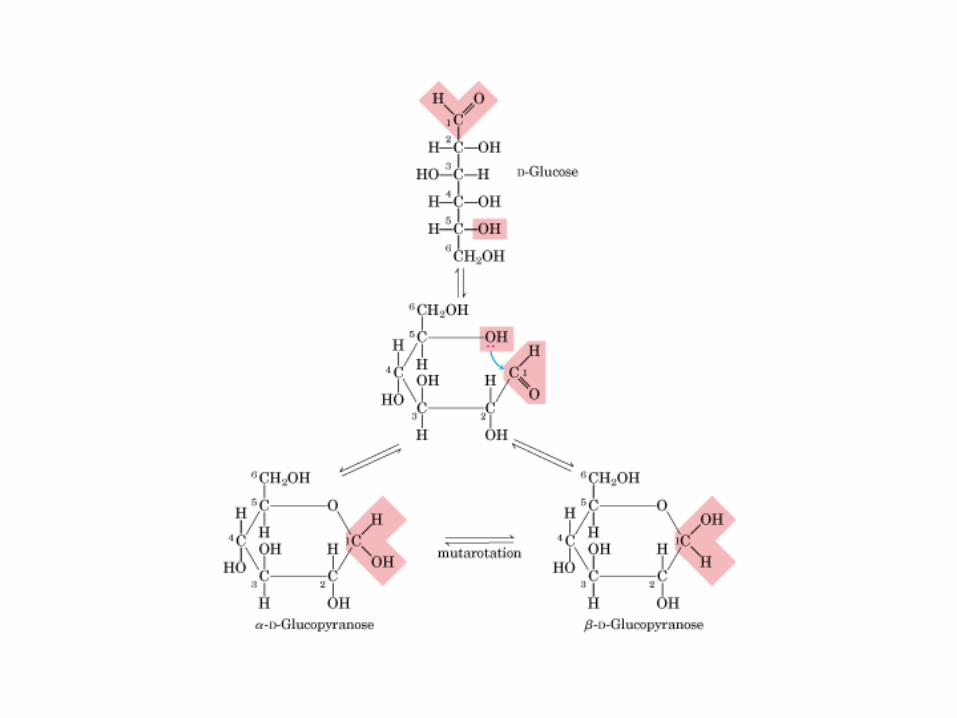
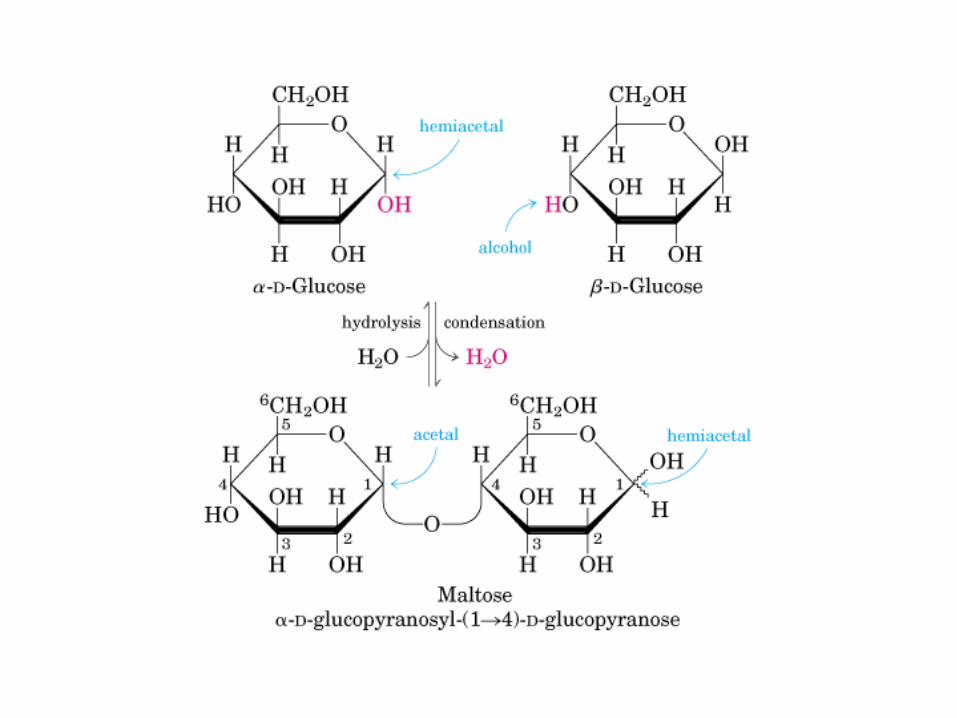
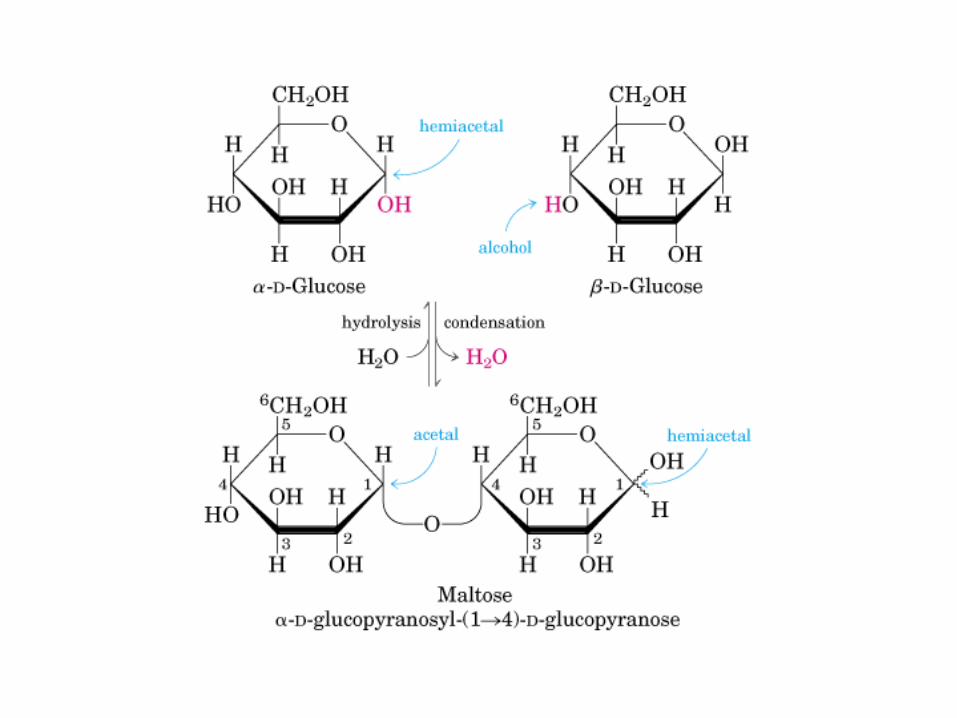
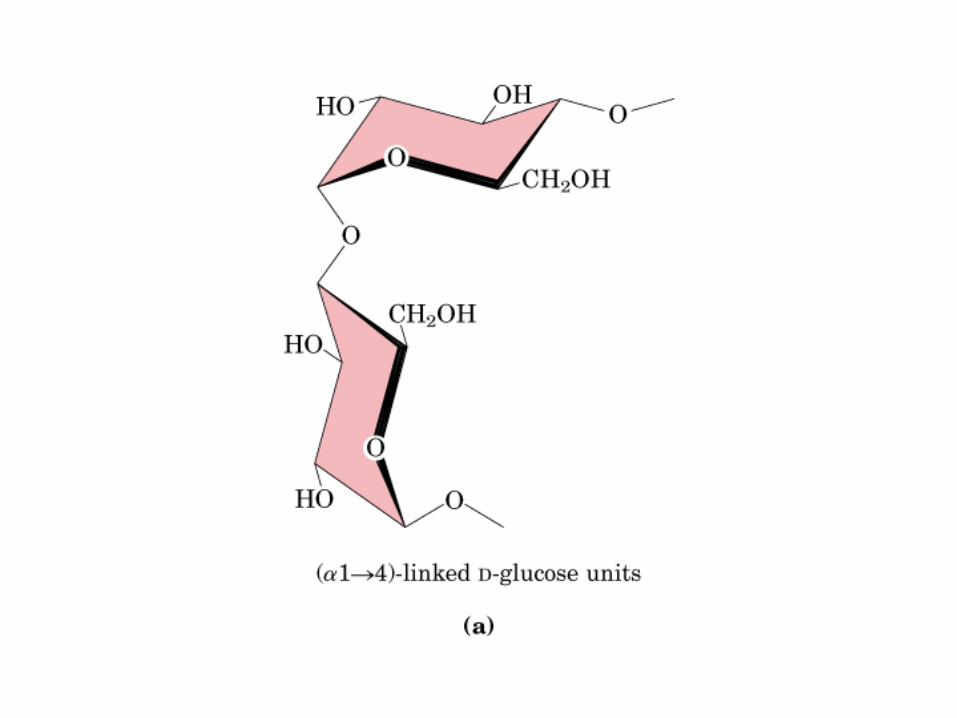
Glicocídicoα y ß -D-Glu Glucocídico0606 0000
Med
. P
rof.
Bon
iello
, V
icen
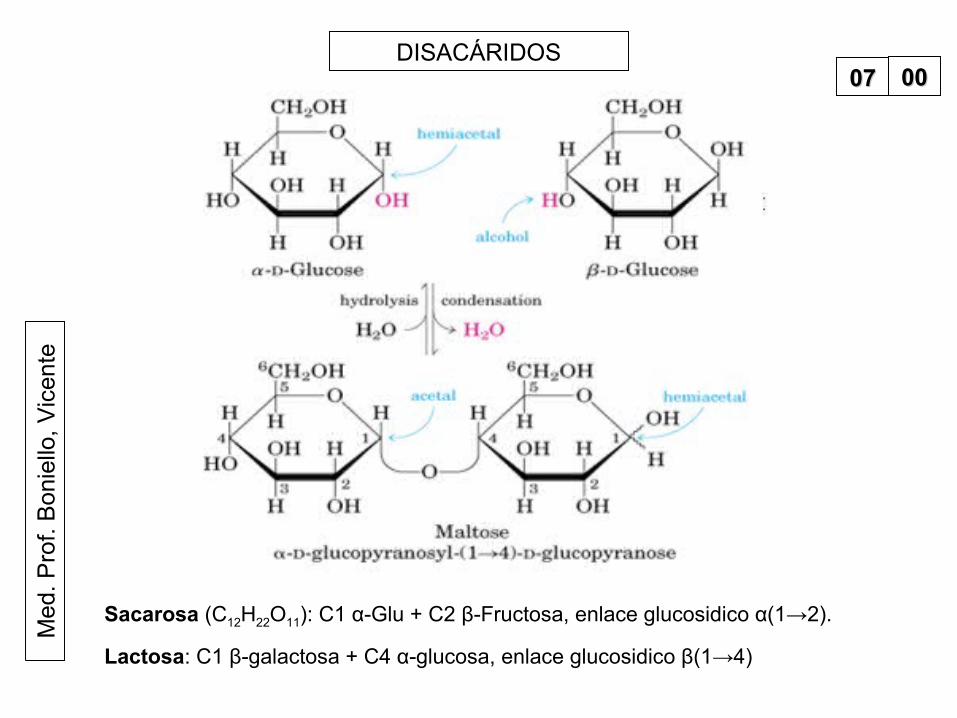
teDISACÁRIDOS
0707 0000
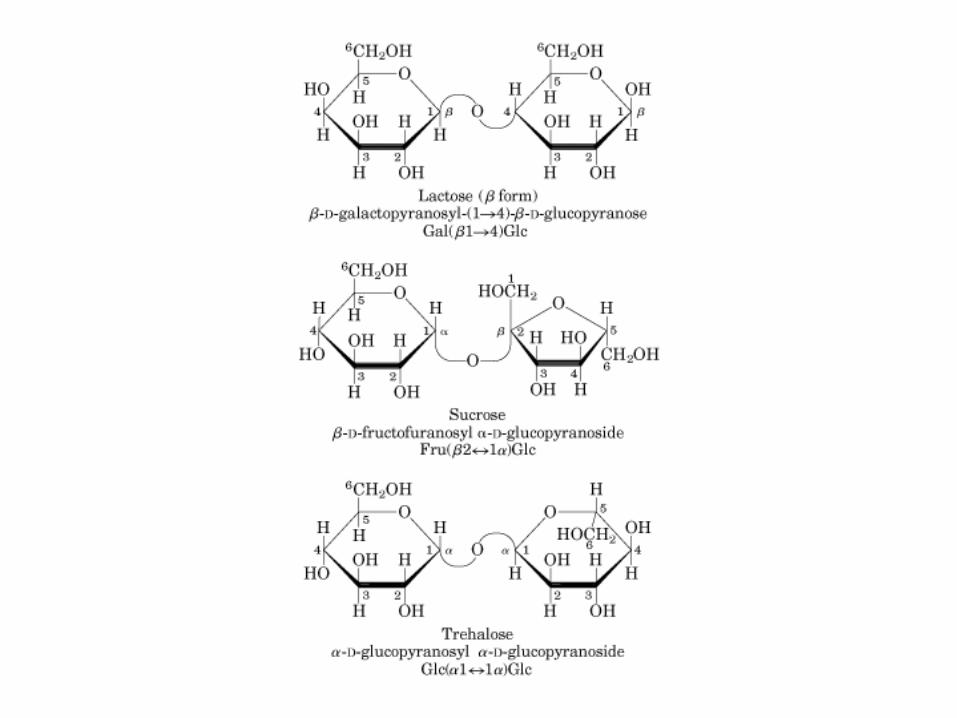
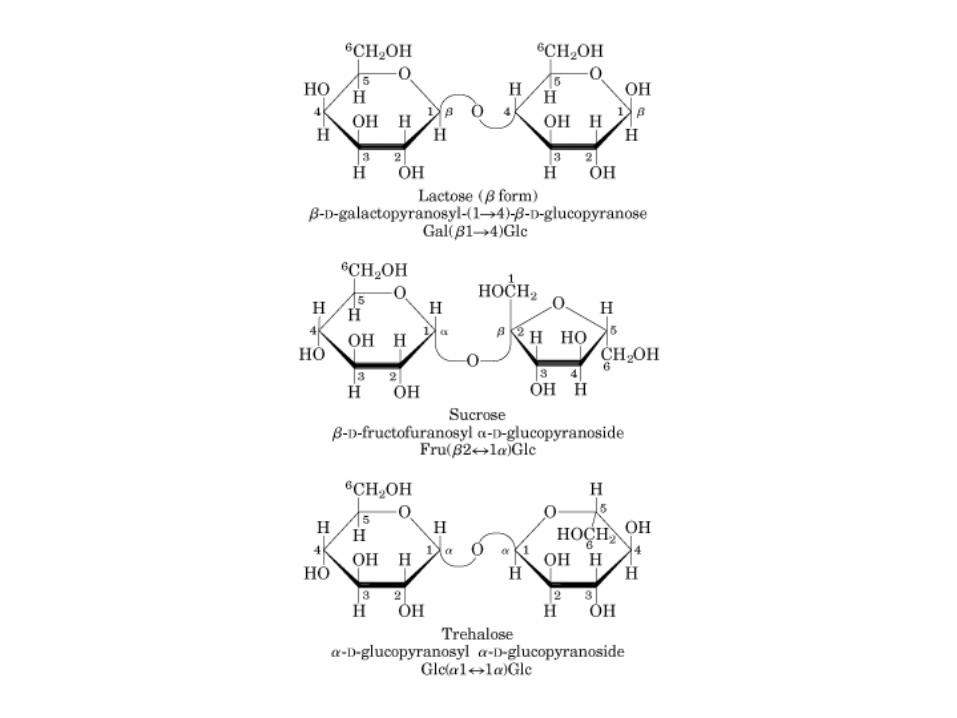
Sacarosa (C12H22O11): C1 α-Glu + C2 β-Fructosa, enlace glucosidico α(1→2).
Lactosa: C1 β-galactosa + C4 α-glucosa, enlace glucosidico β(1→4)
Med
. P
rof.
Bon
iello
, V
icen
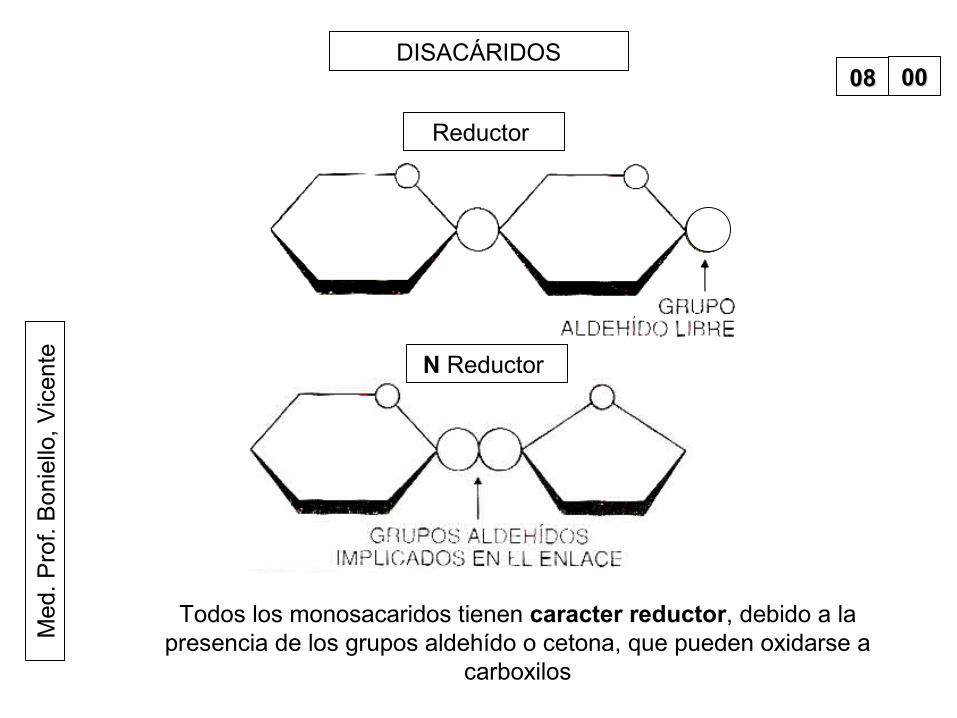
teDISACÁRIDOS
Reductor
N Reductor
0808 0000
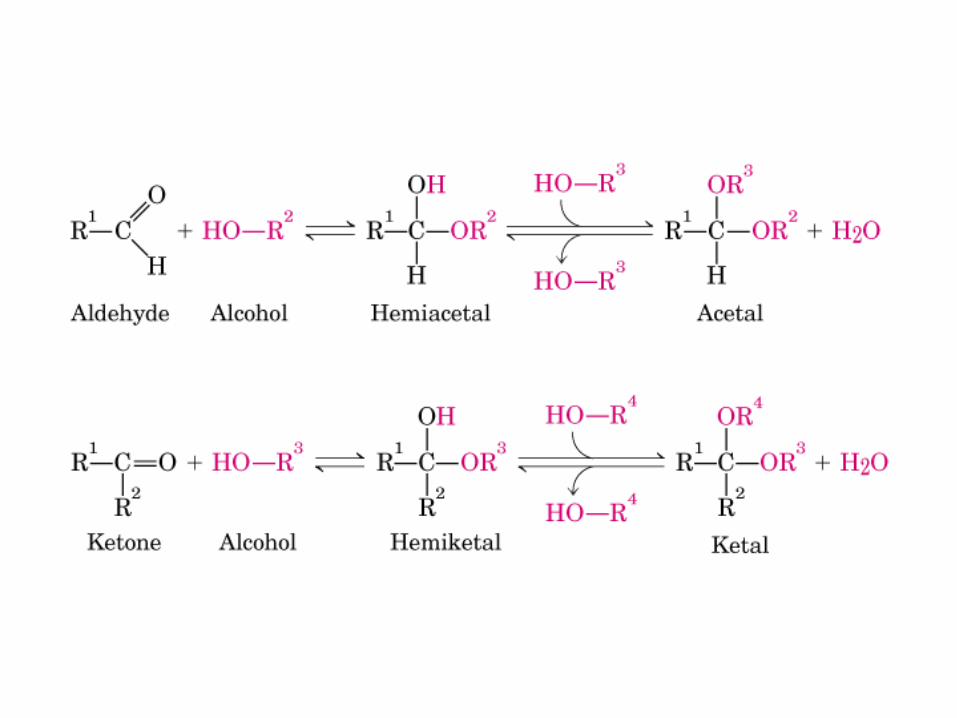
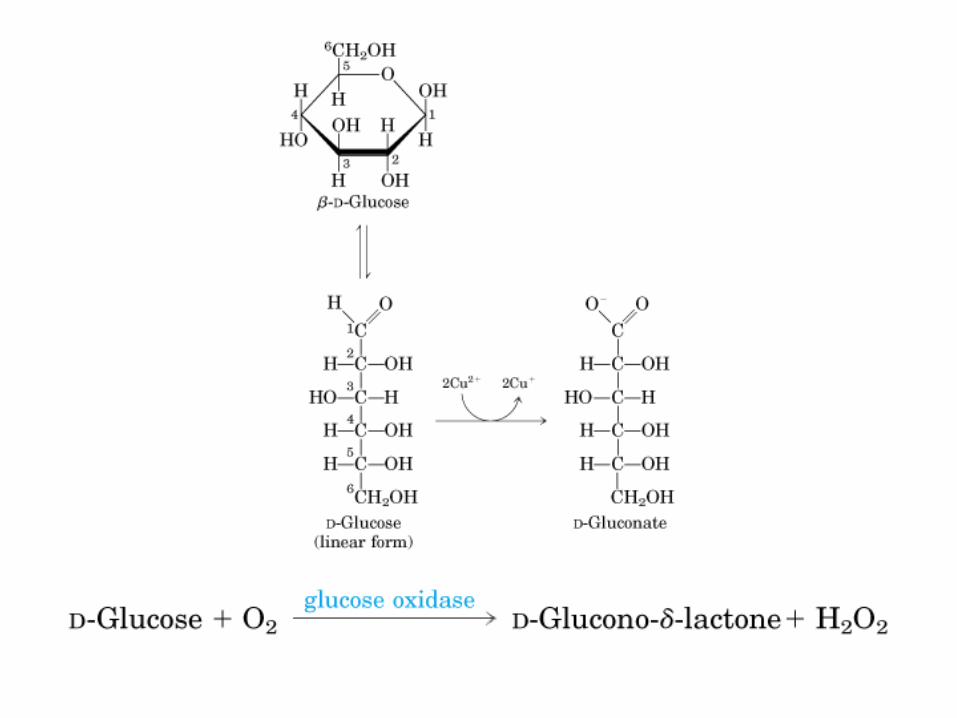
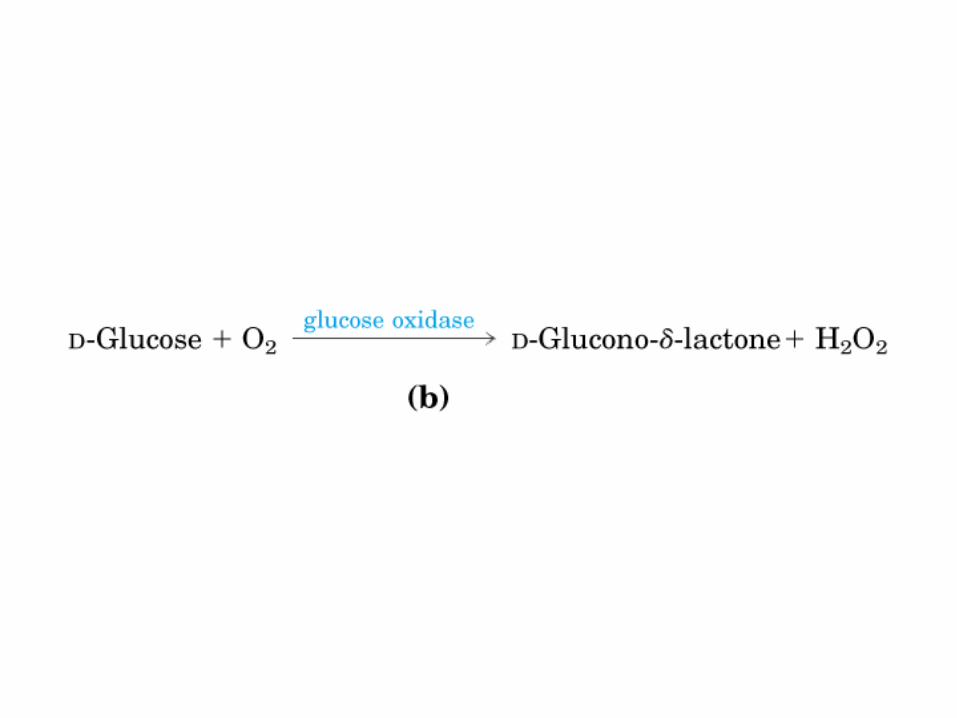
Todos los monosacaridos tienen caracter reductor, debido a la presencia de los grupos aldehído o cetona, que pueden oxidarse a
carboxilos
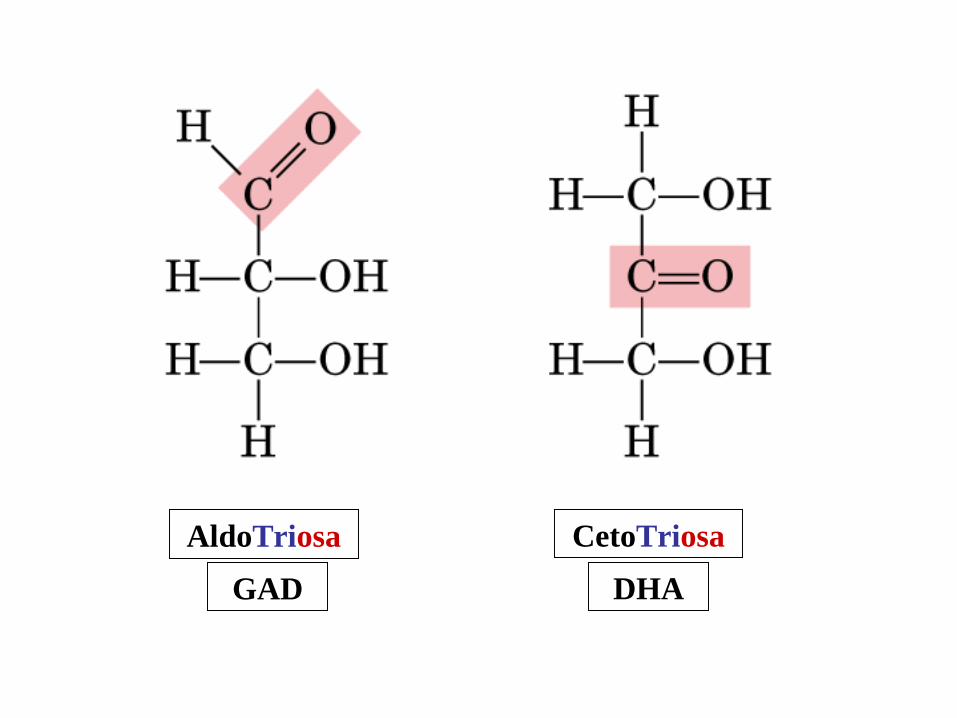
CetoTriosa
DHA
AldoTriosa
GAD
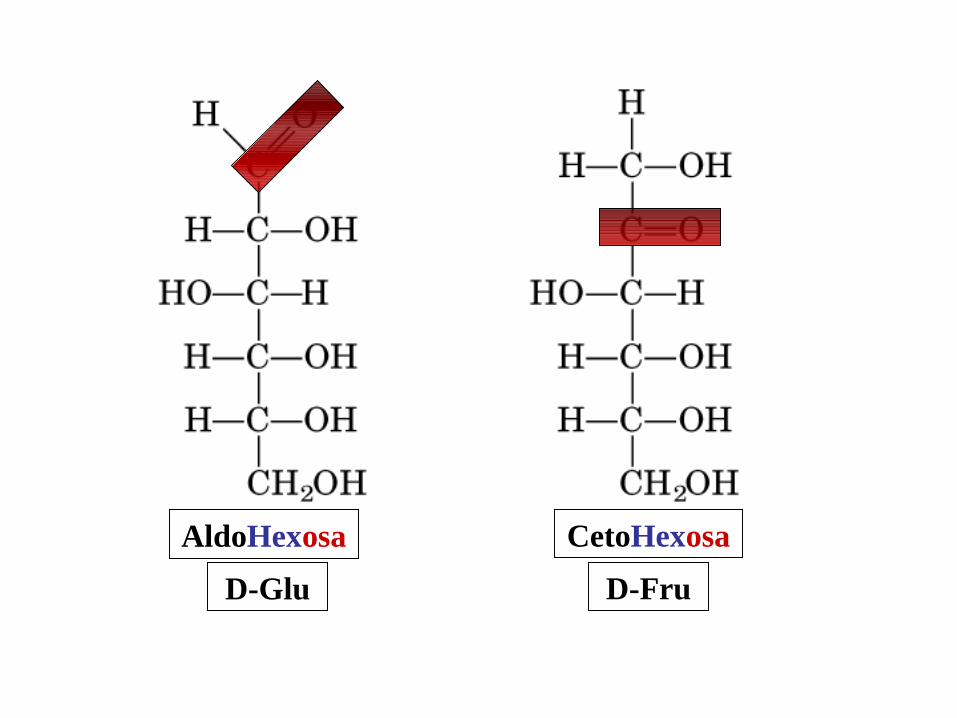
CetoHexosa
D-Fru
AldoHexosa
D-Glu
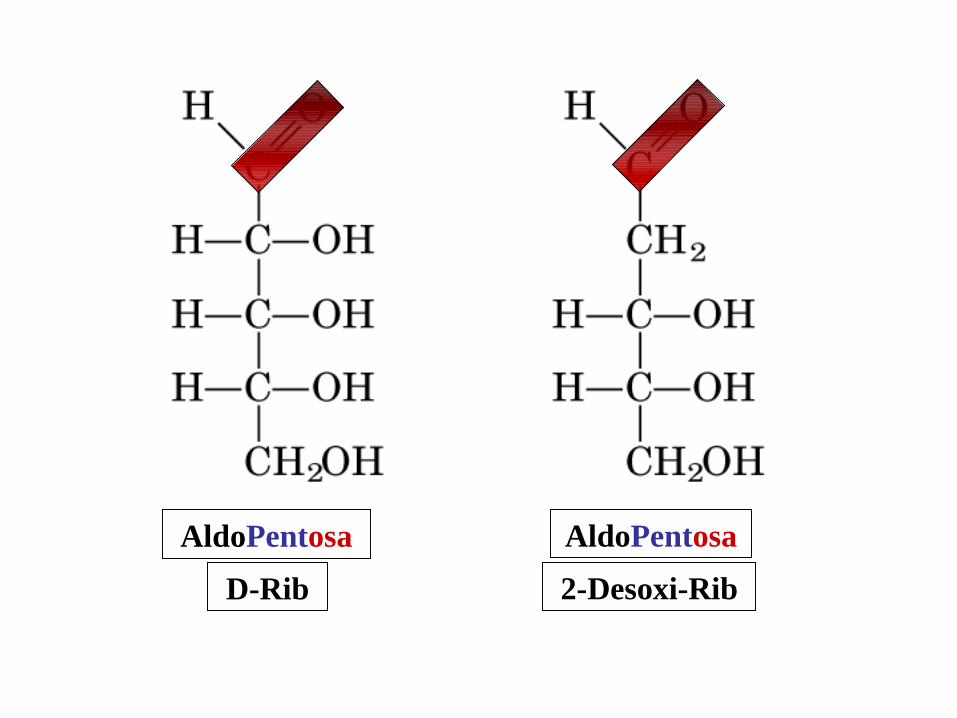
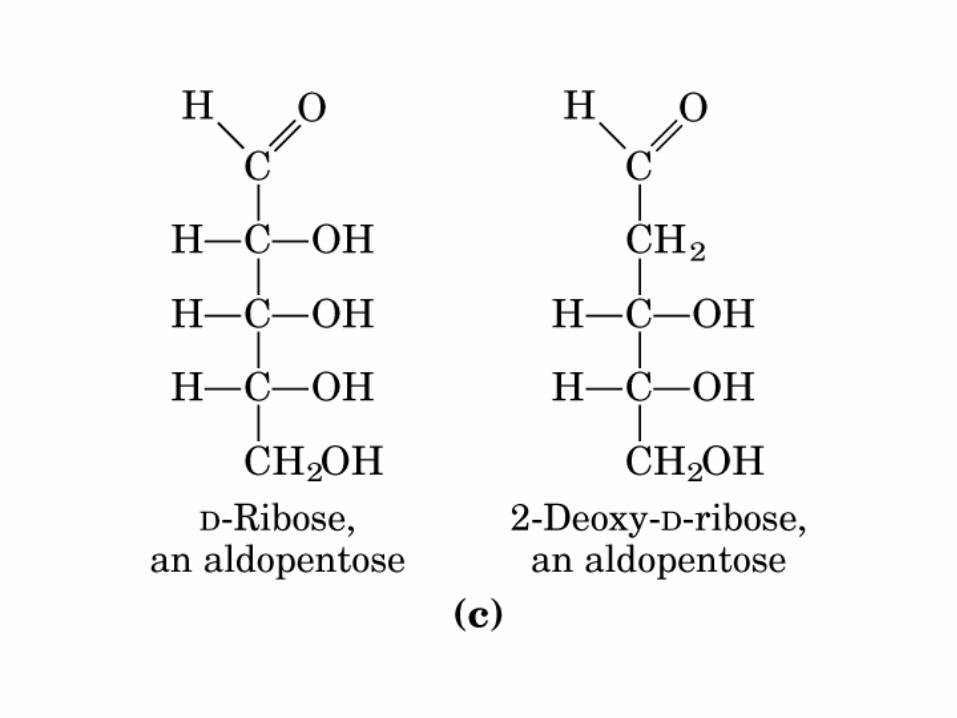
AldoPentosa
2-Desoxi-Rib
AldoPentosa
D-Rib
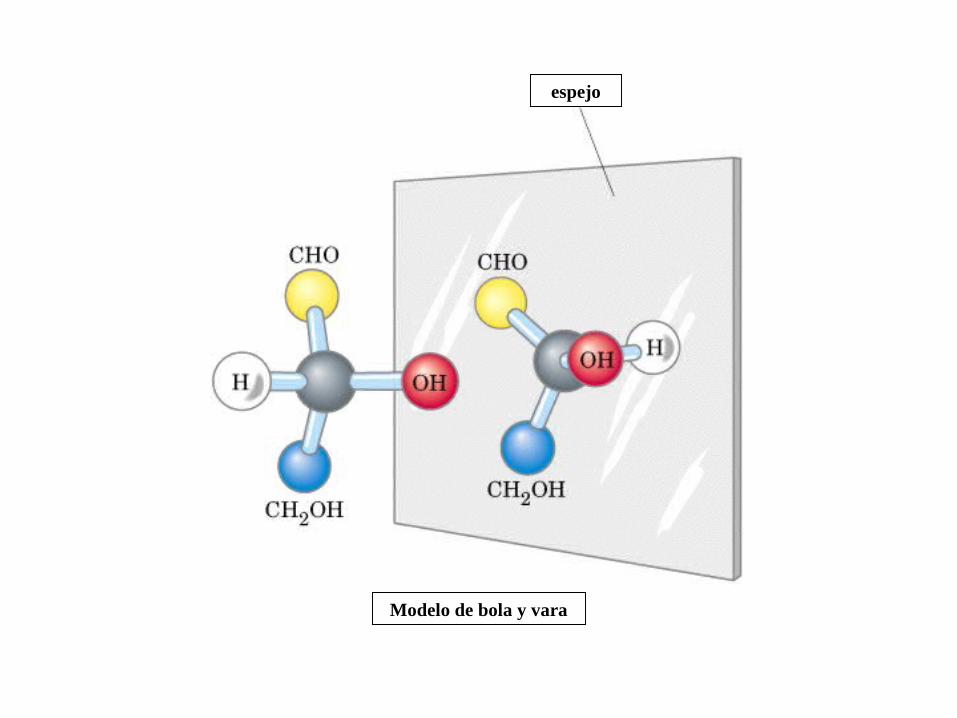
espejo
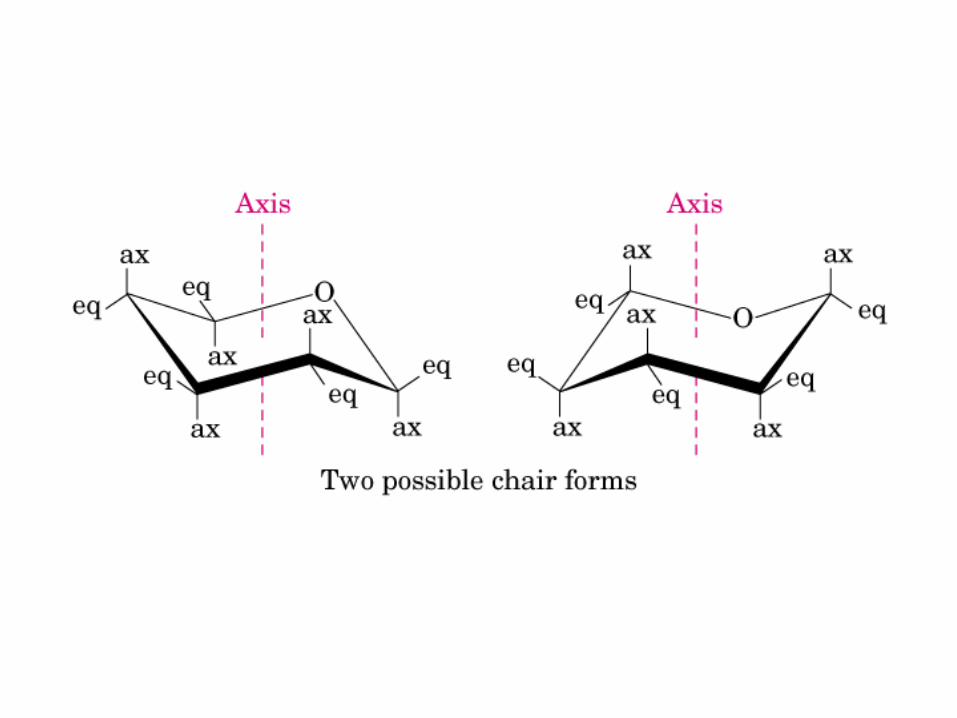
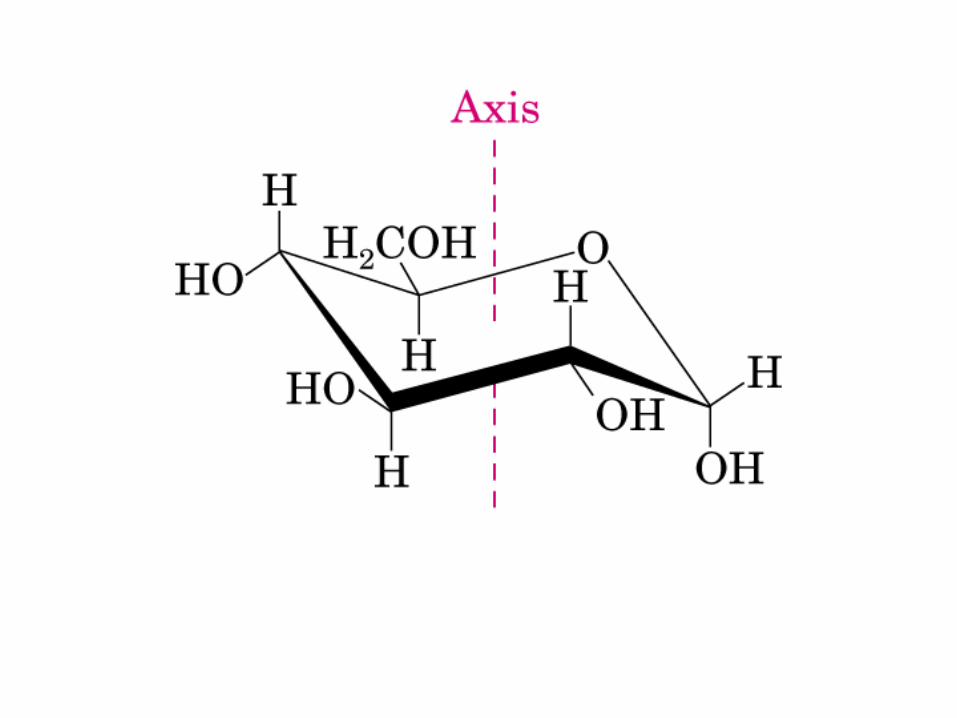
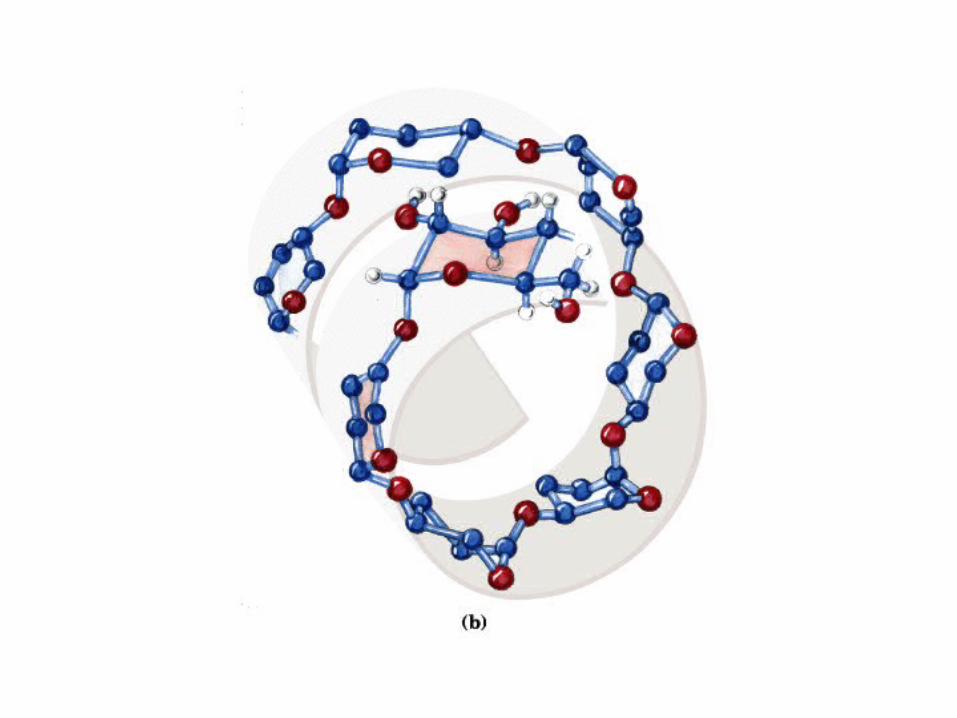
Modelo de bola y vara
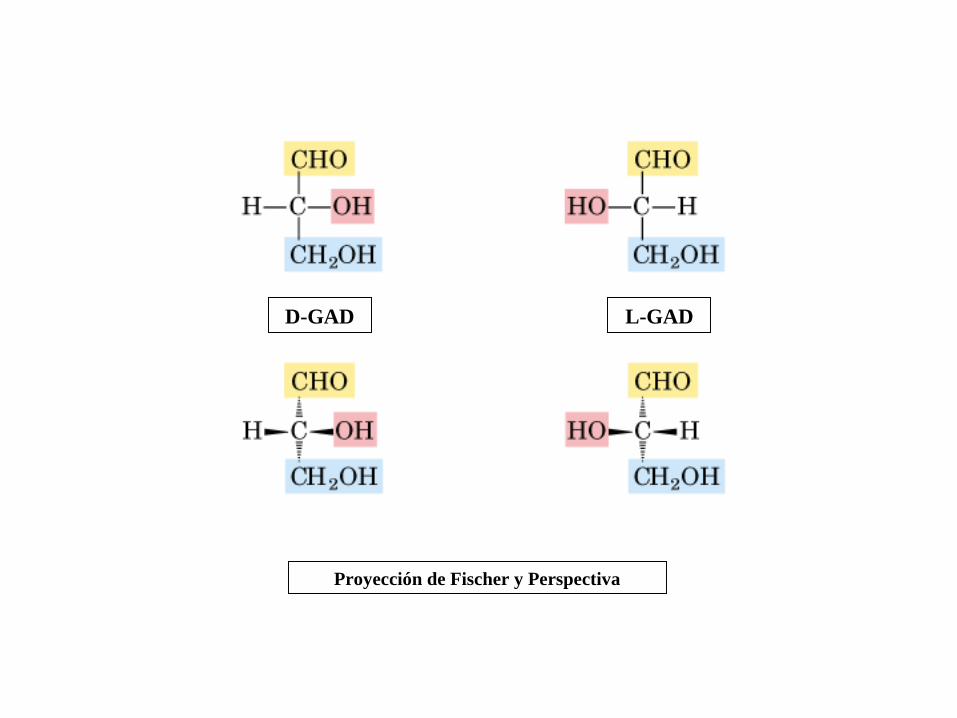
Proyección de Fischer y Perspectiva
D-GAD L-GAD

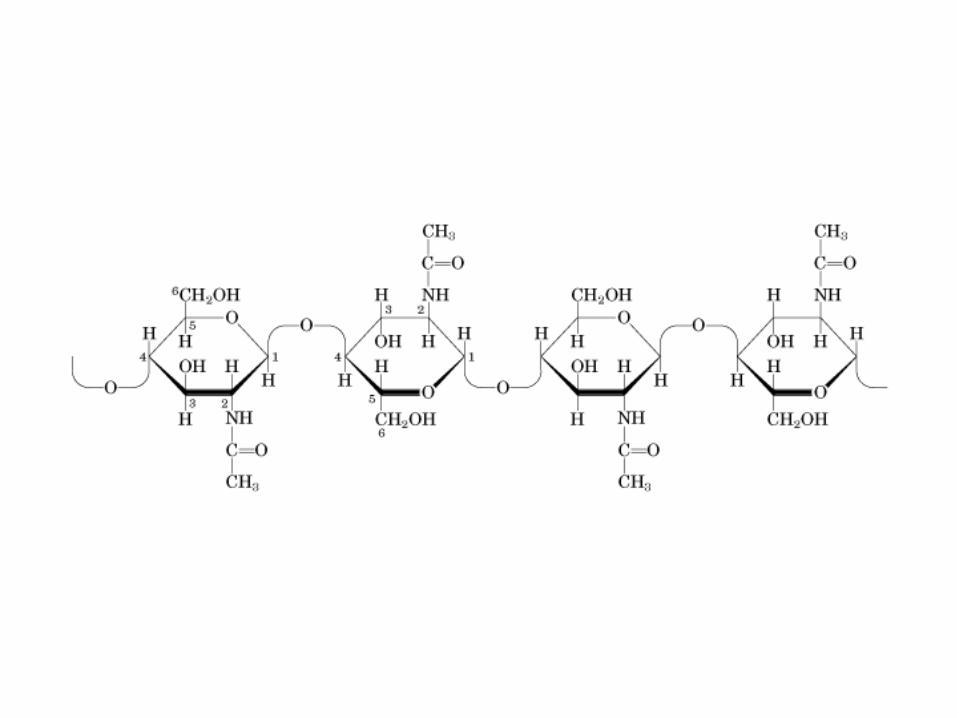
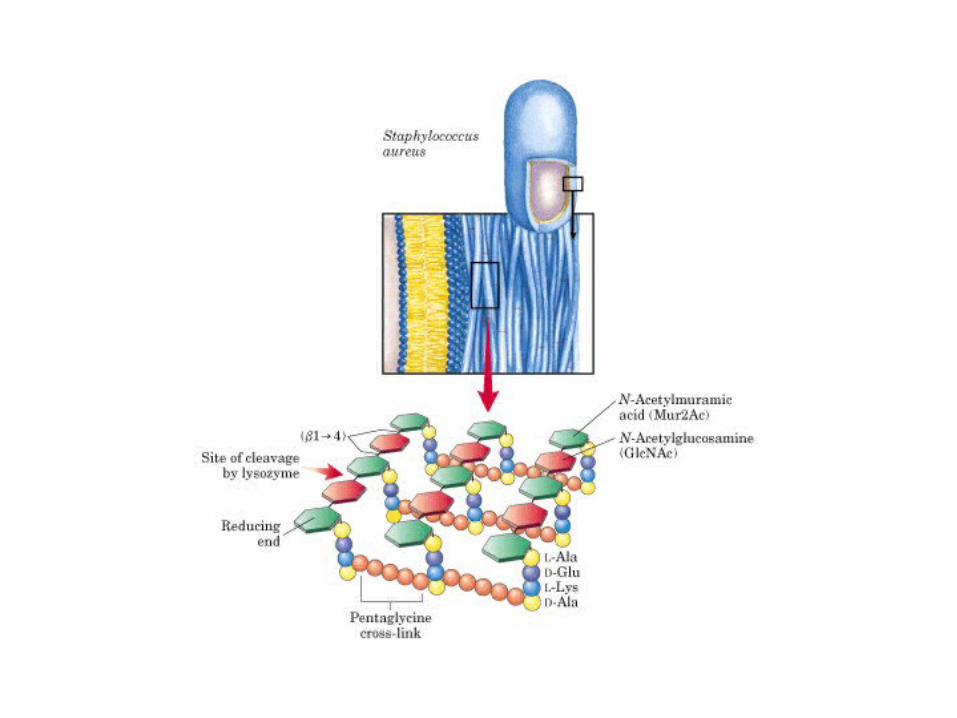
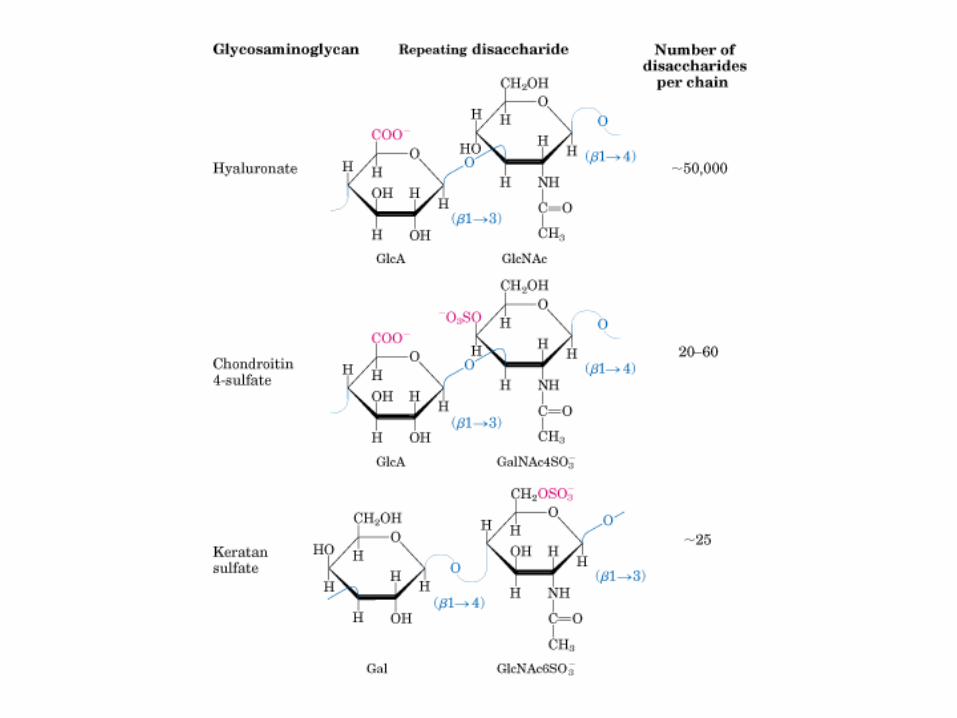
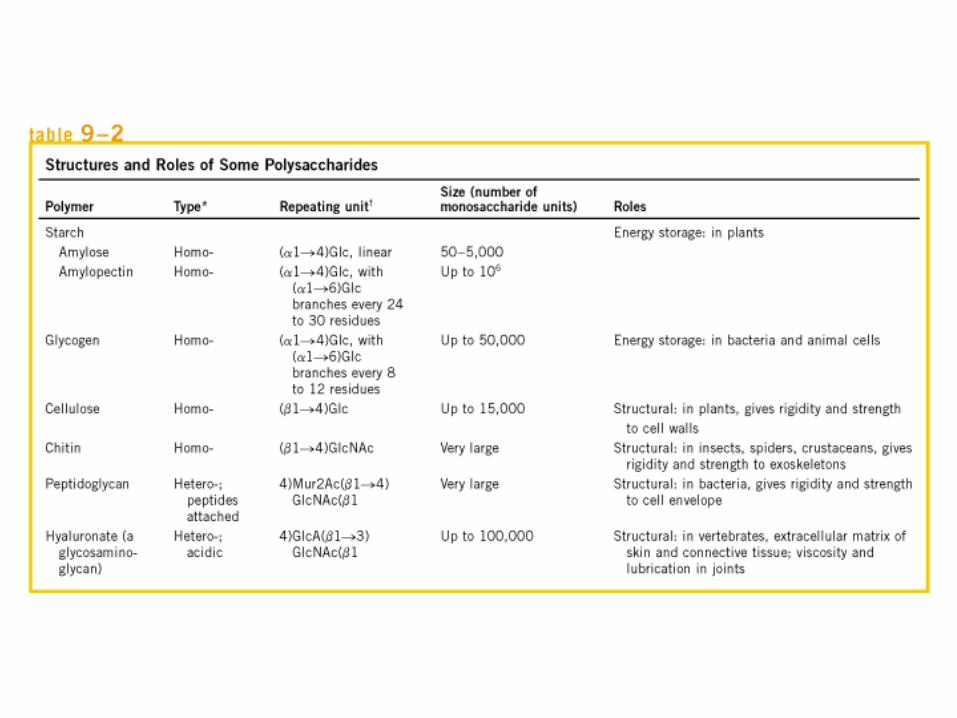

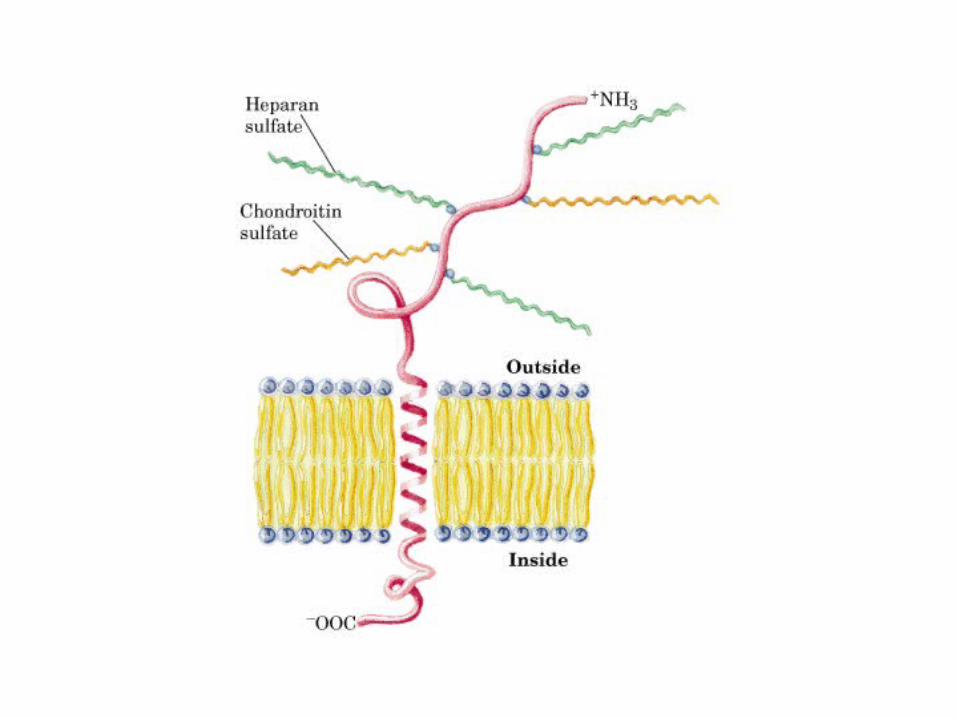
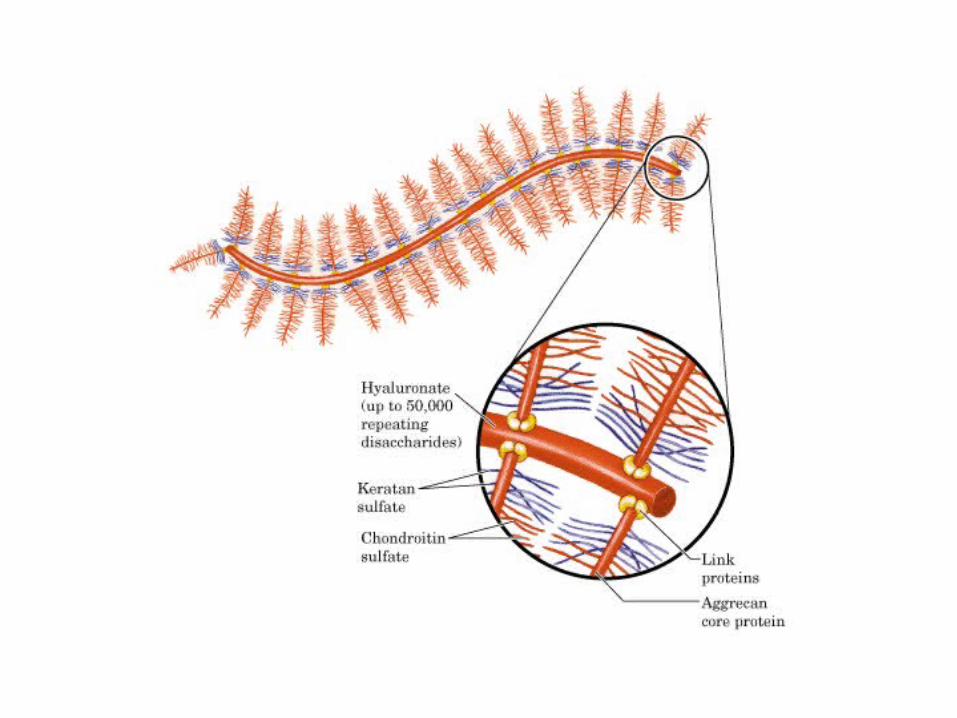
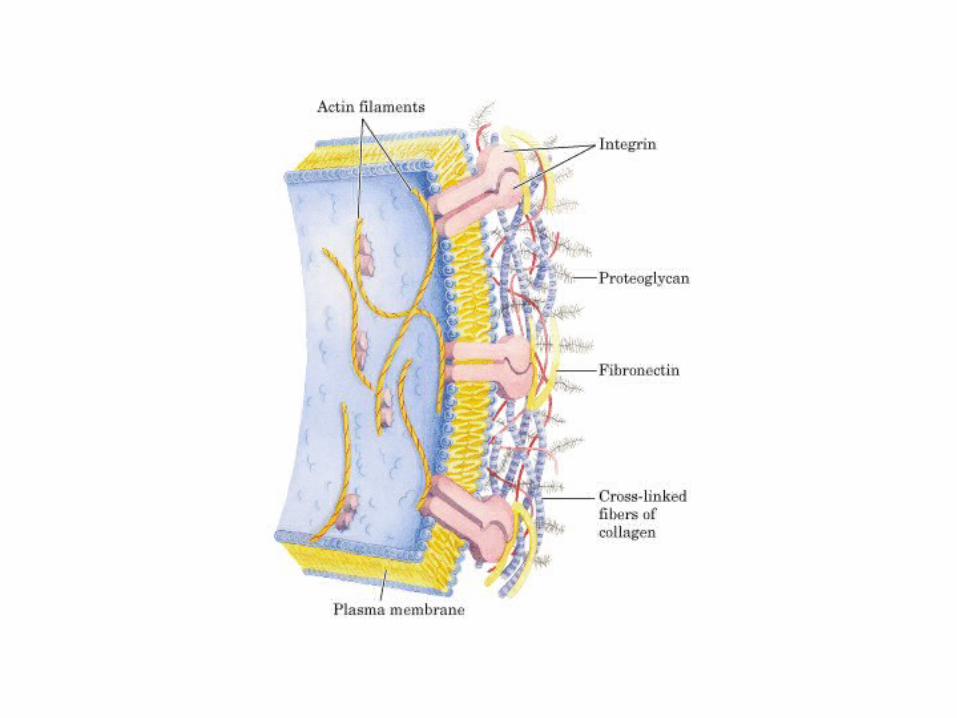
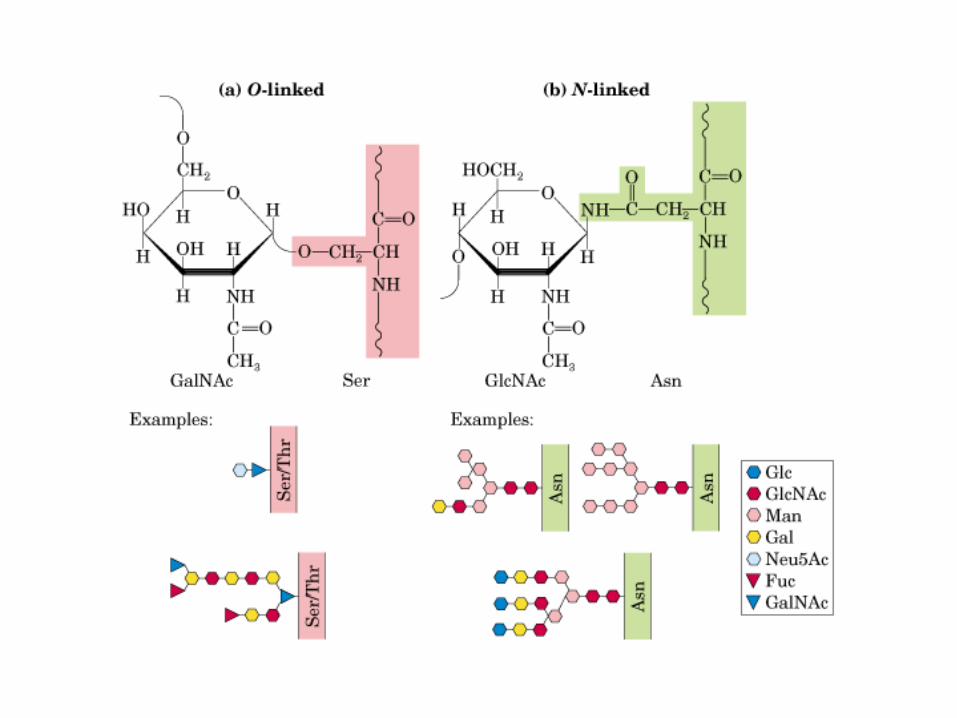
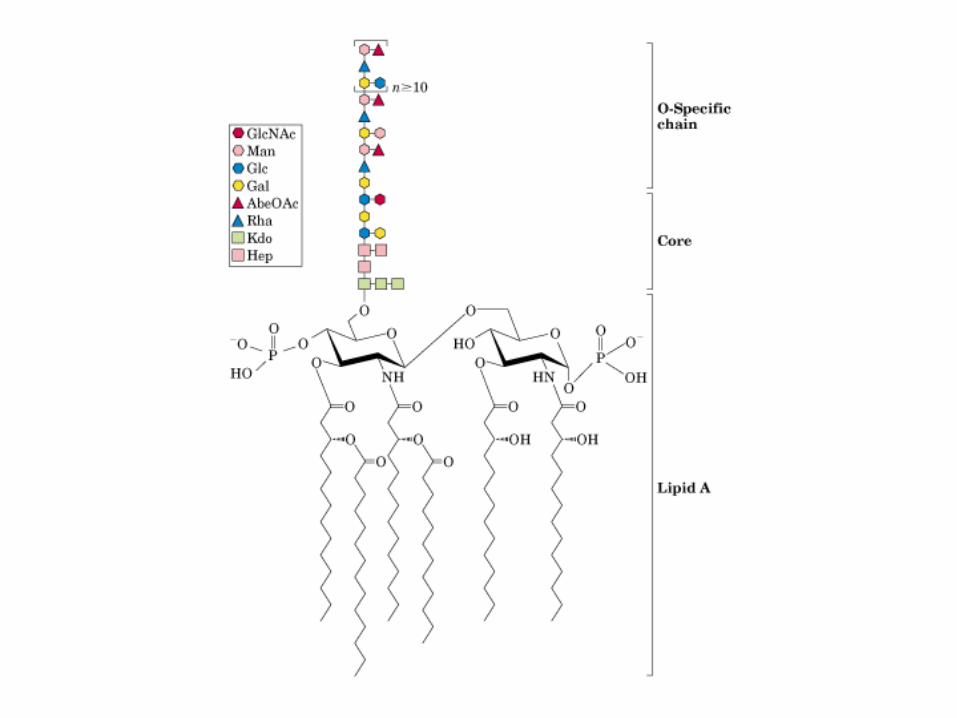
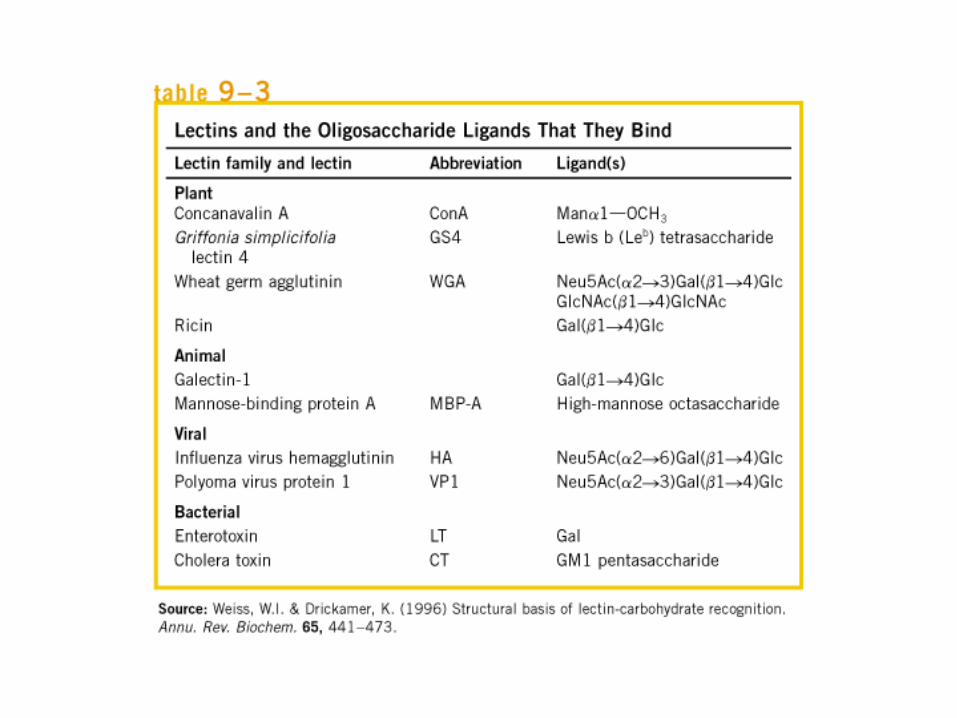
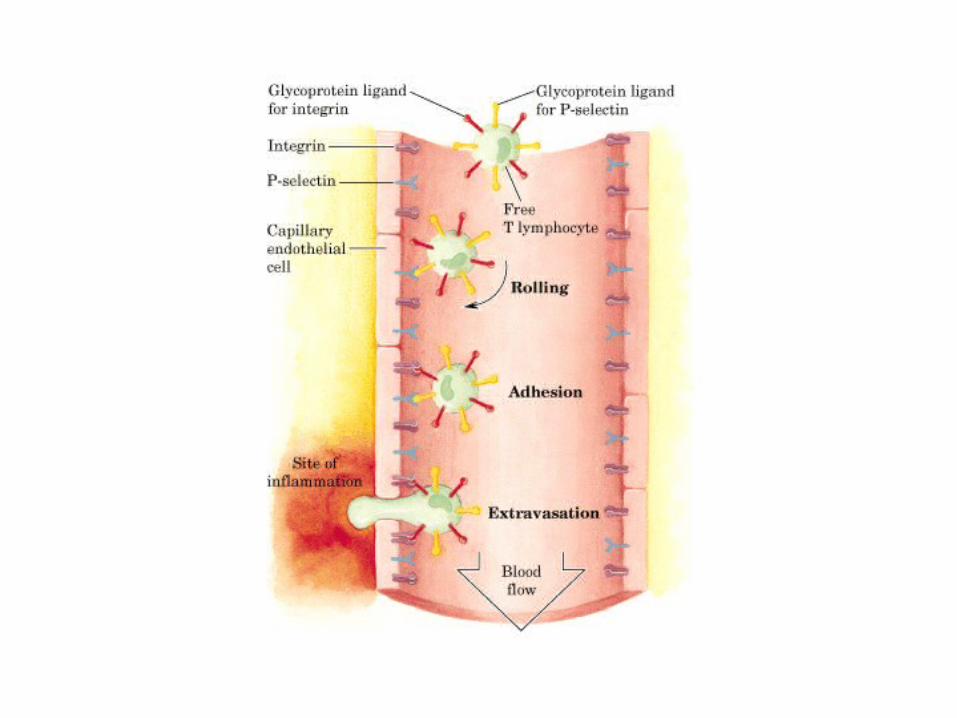
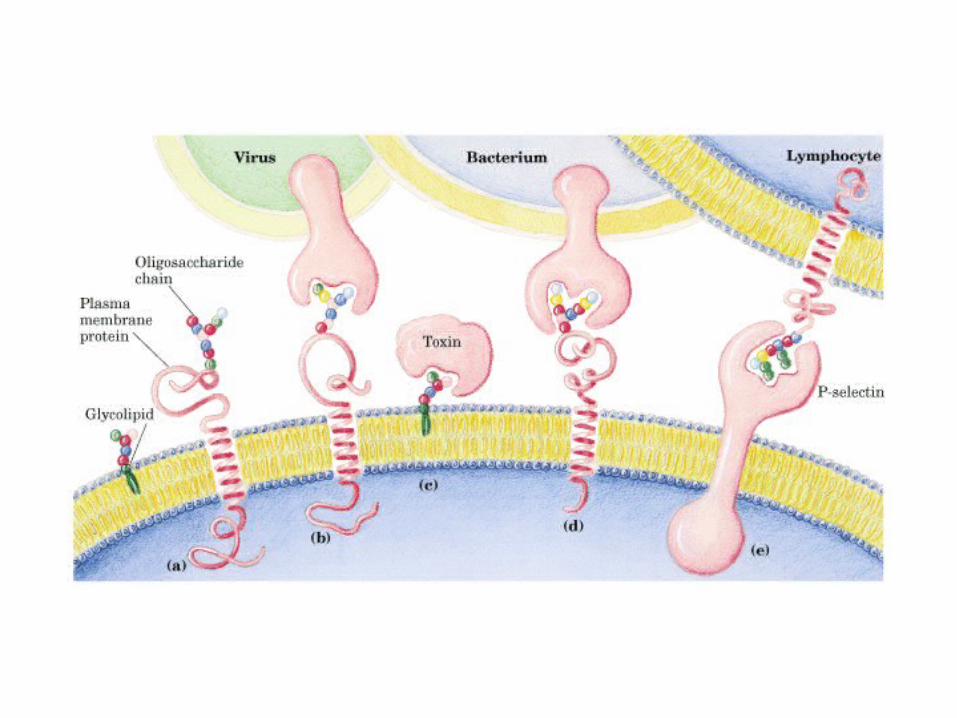
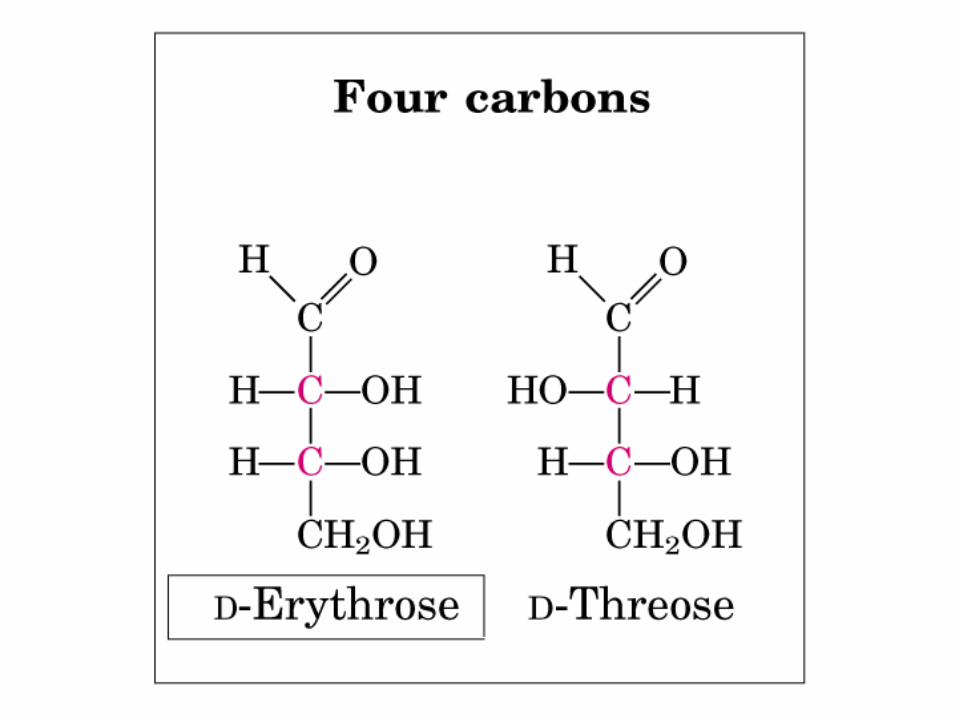
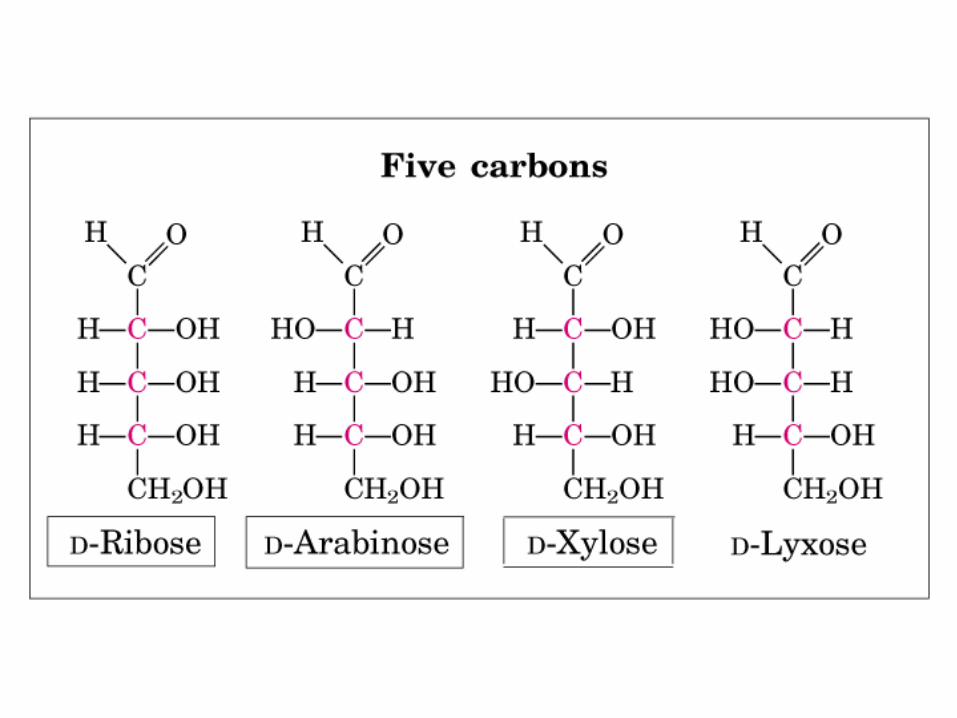

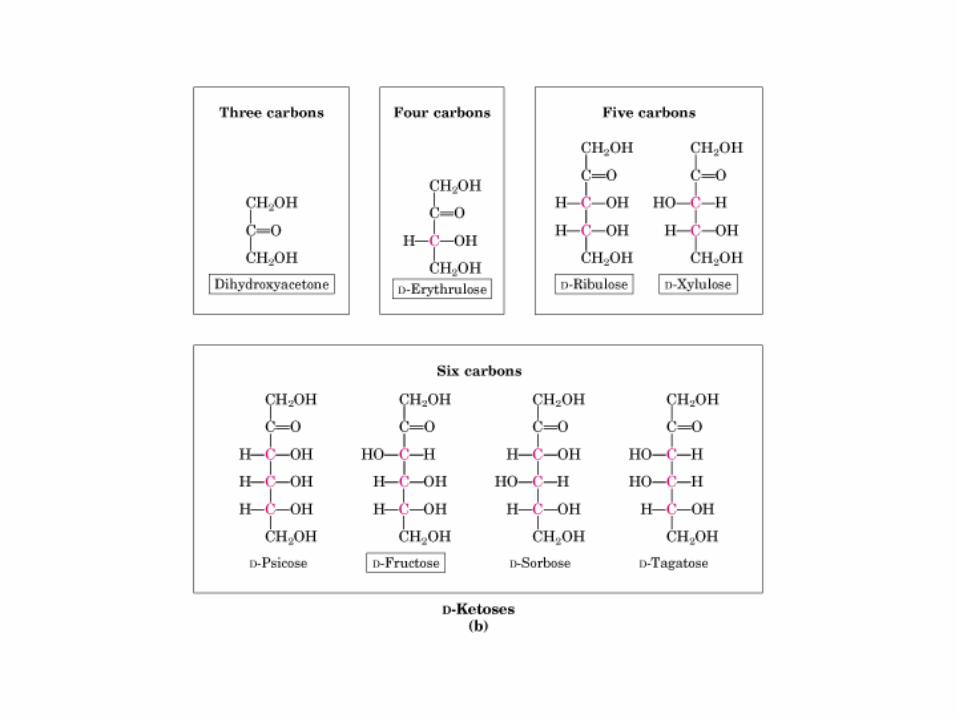
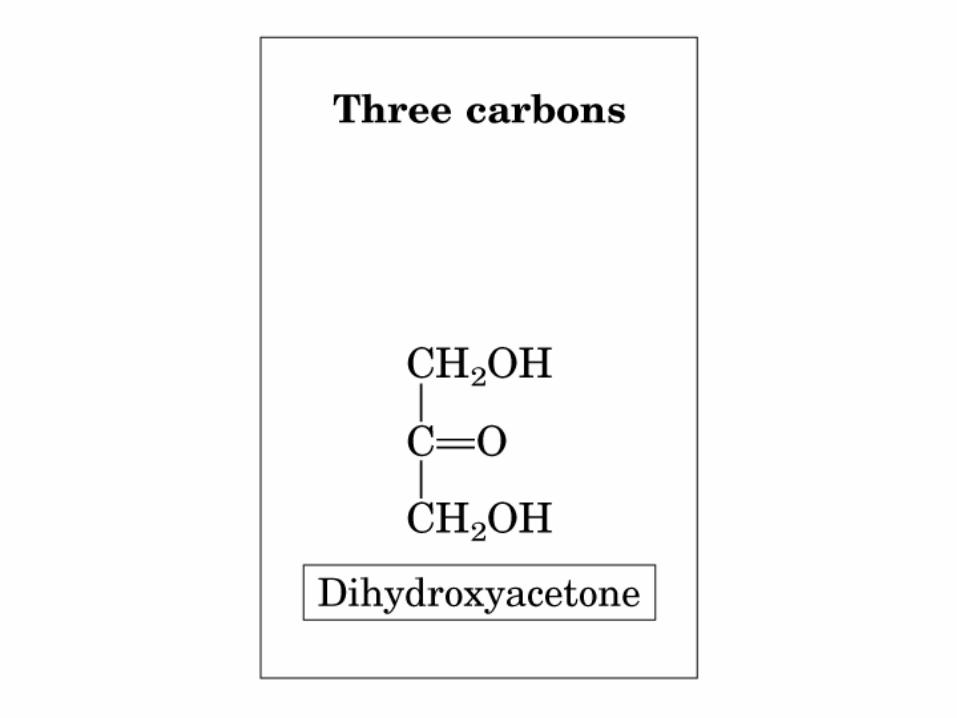
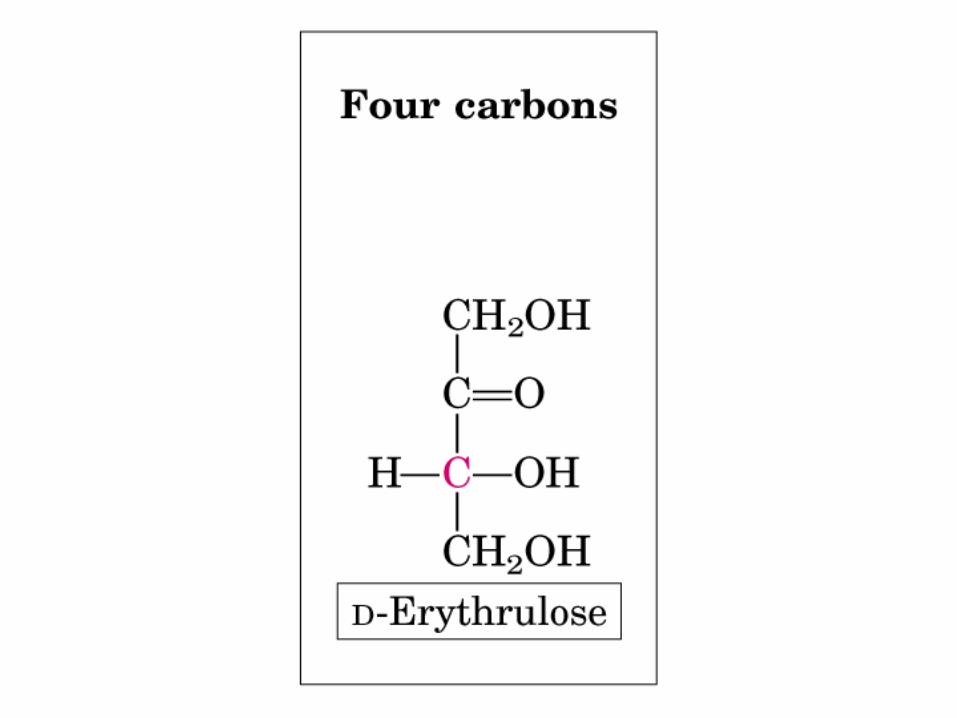
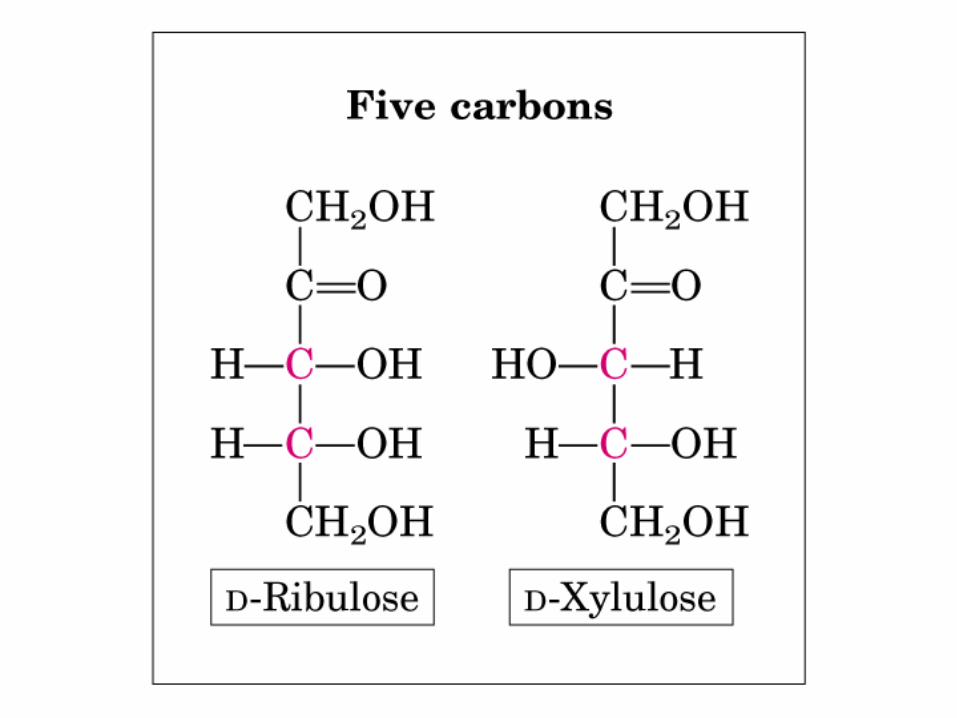
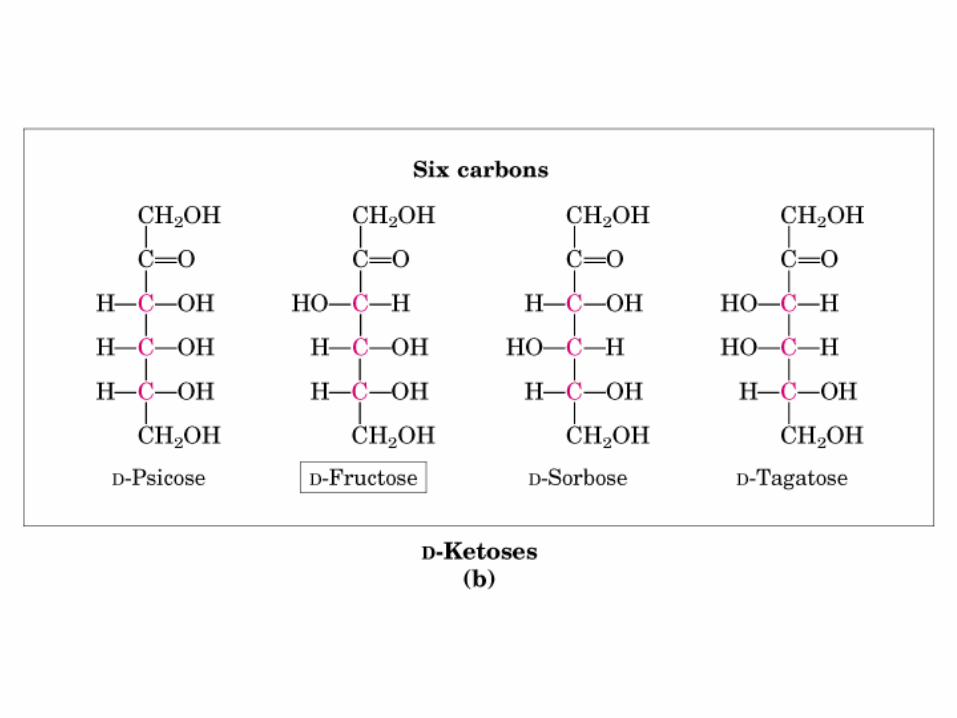
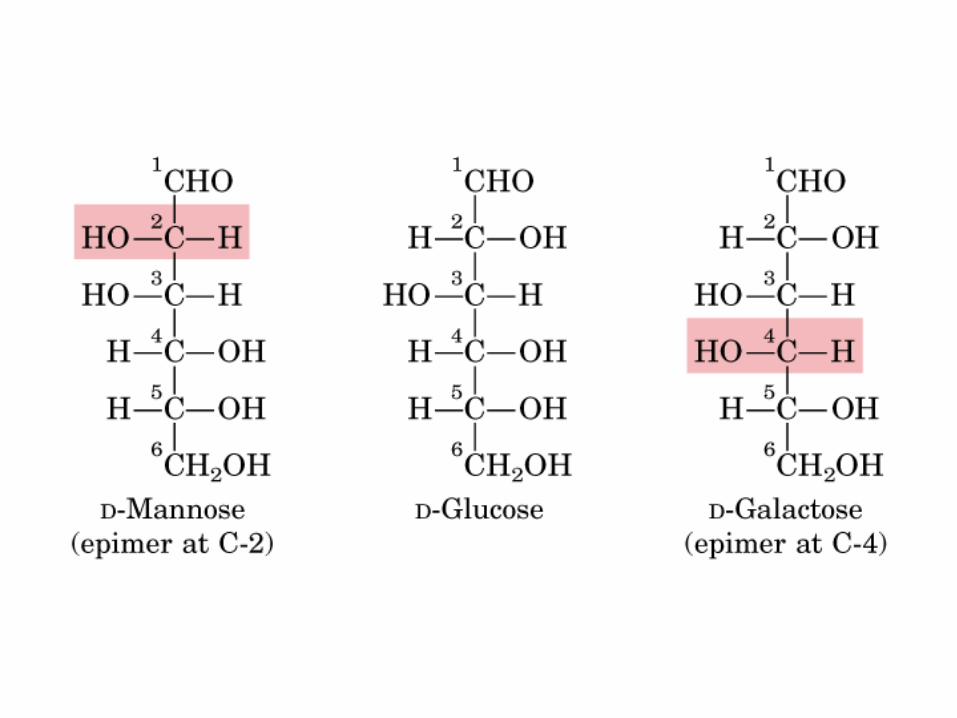
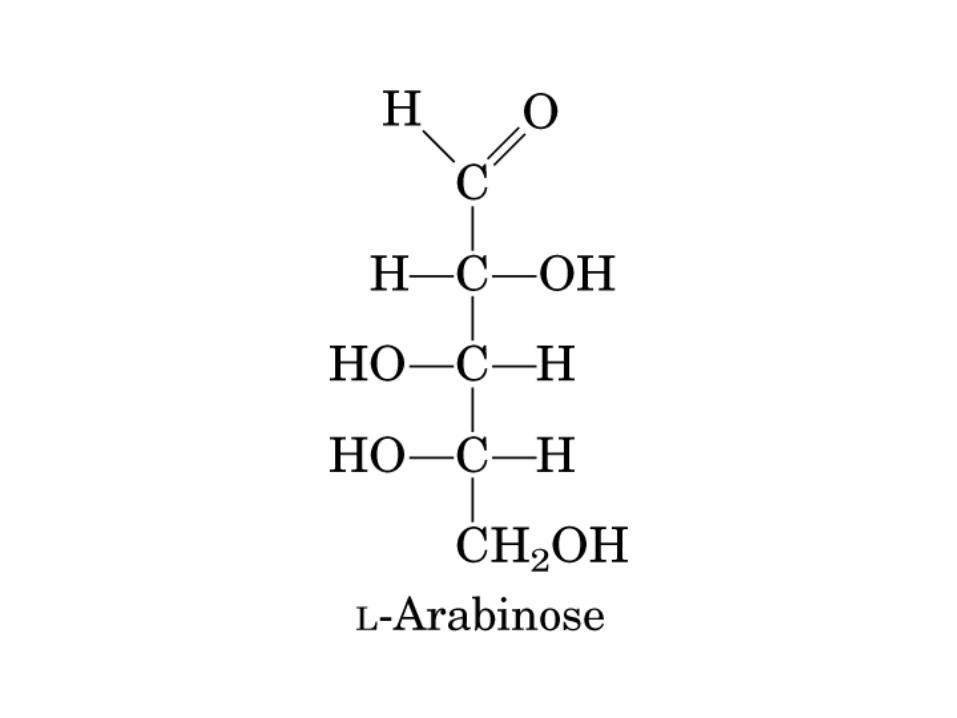
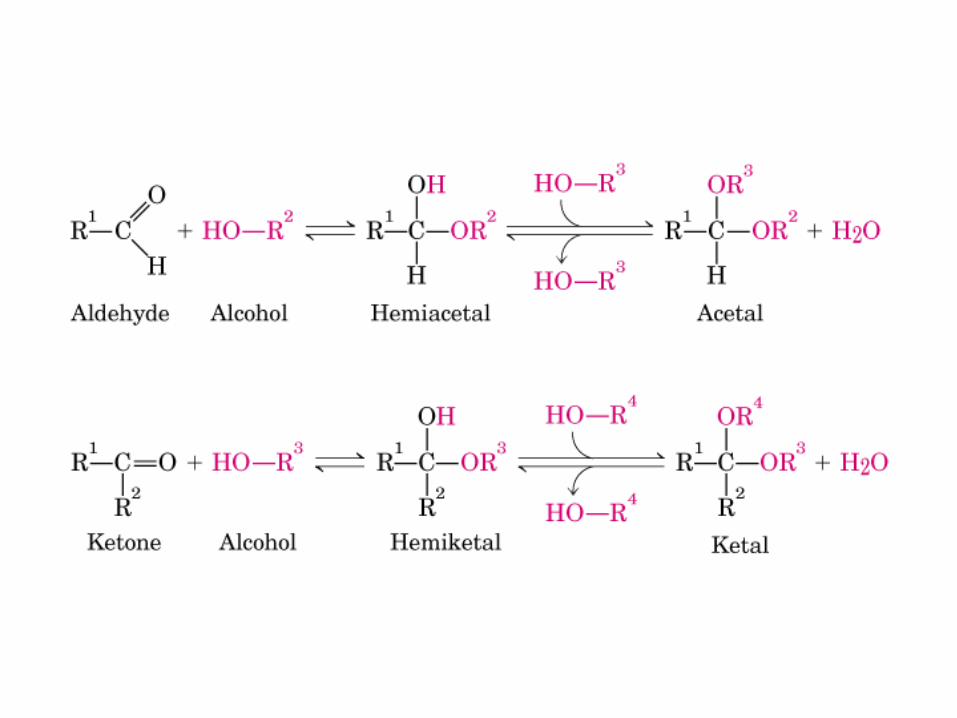
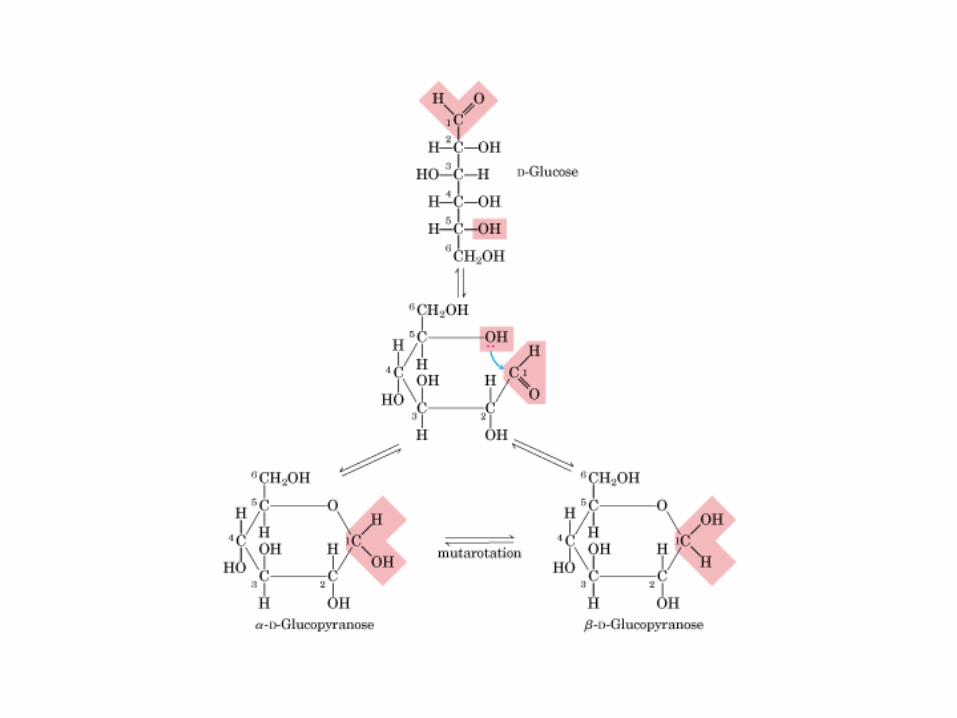
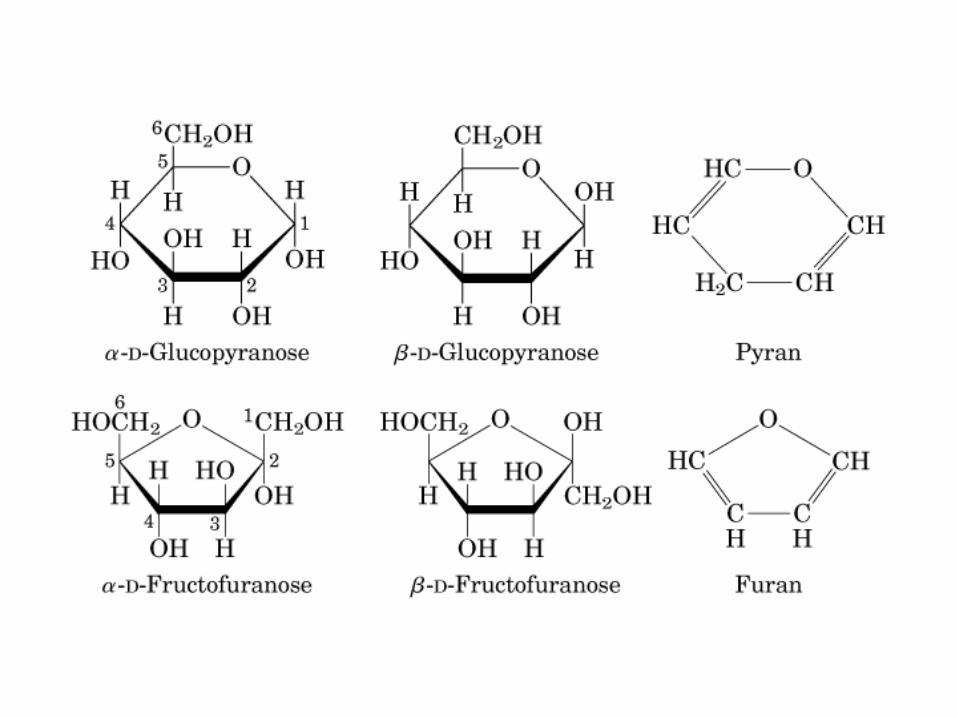
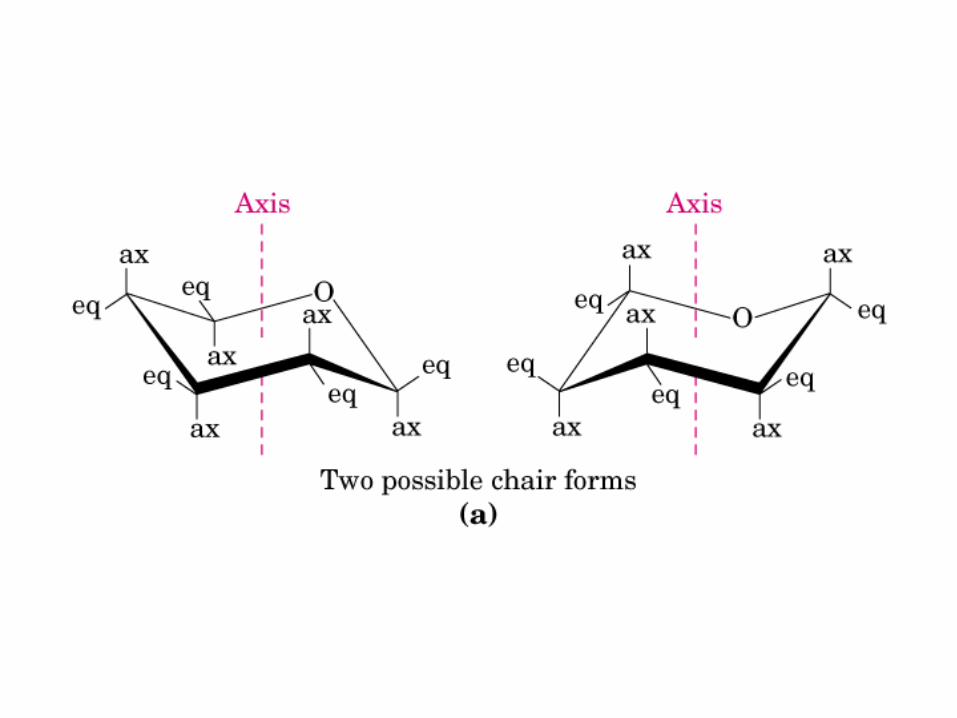
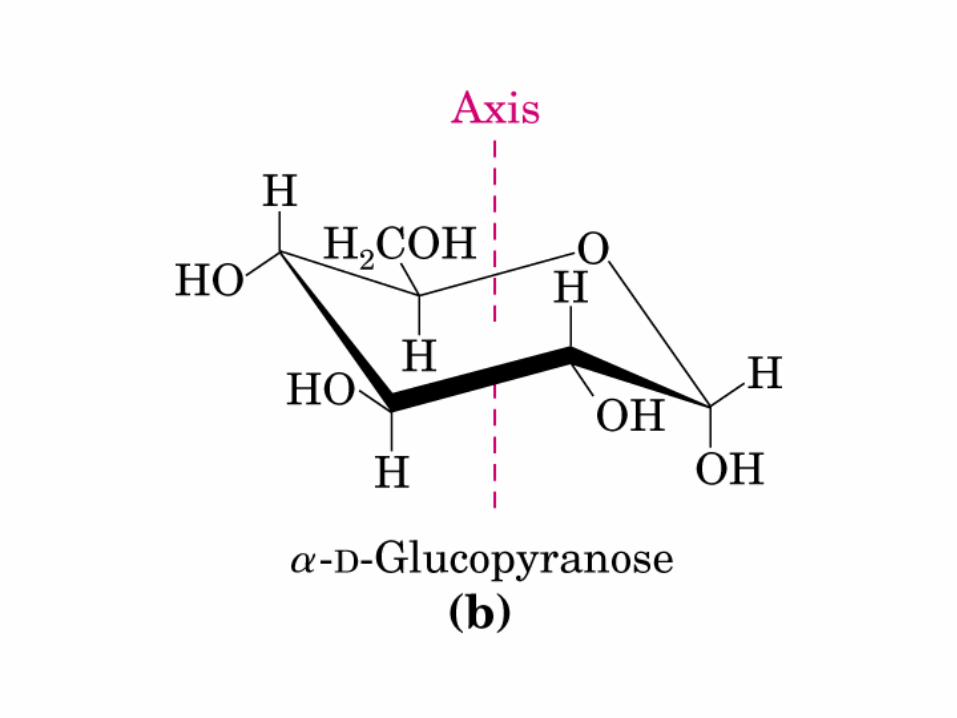
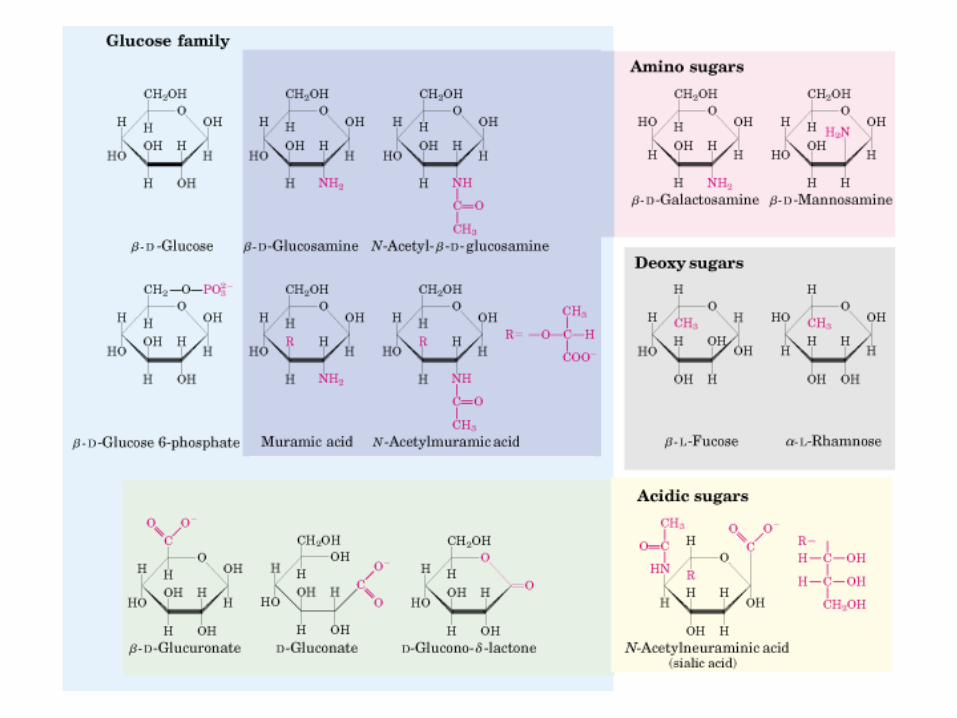
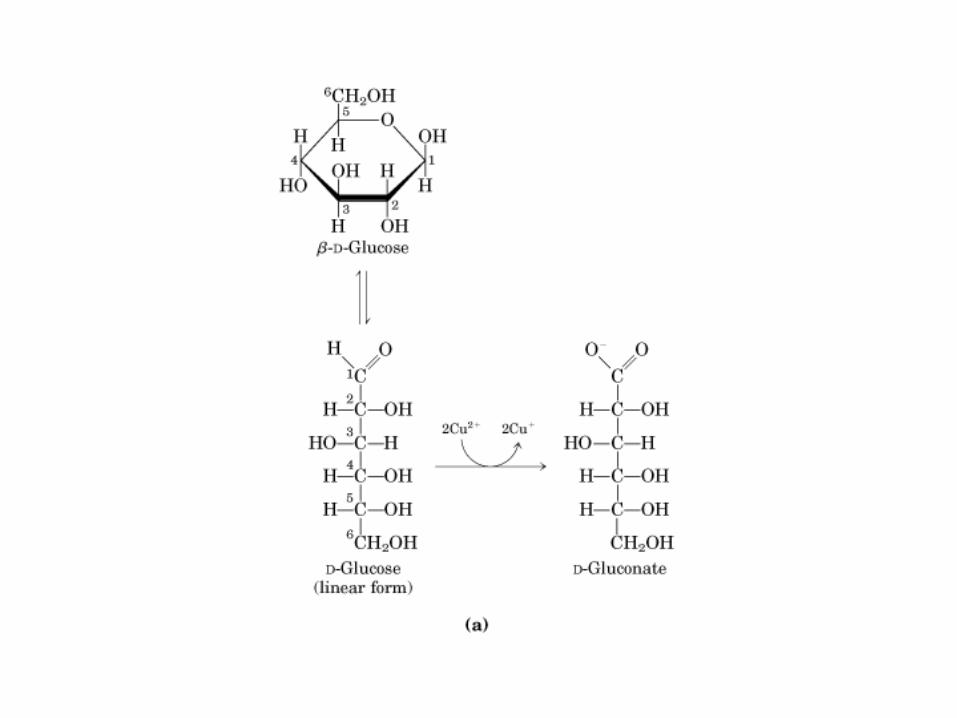
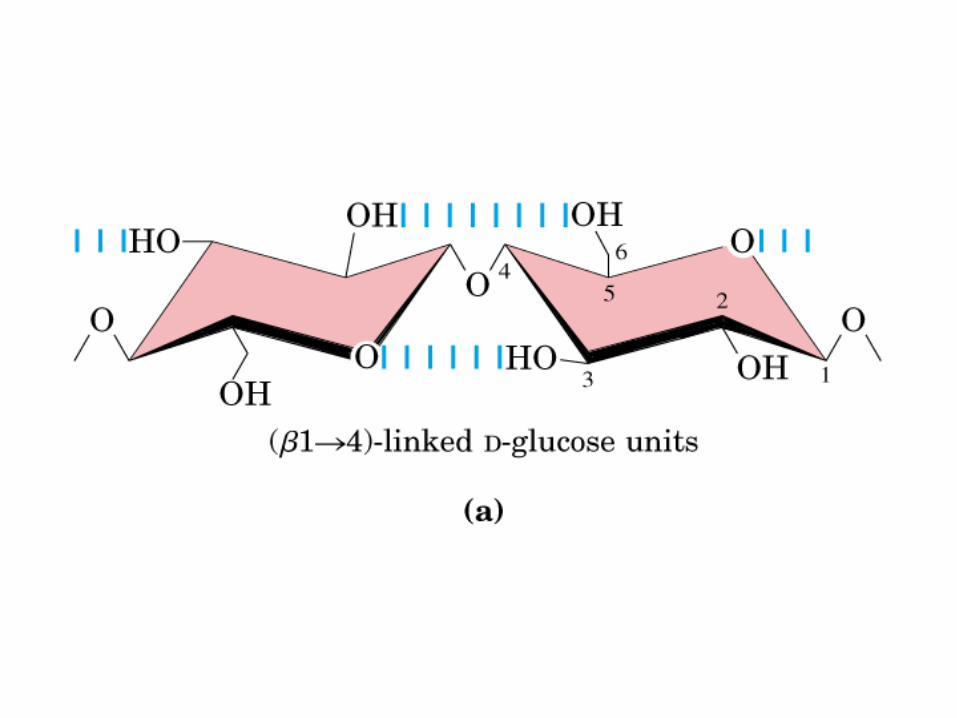
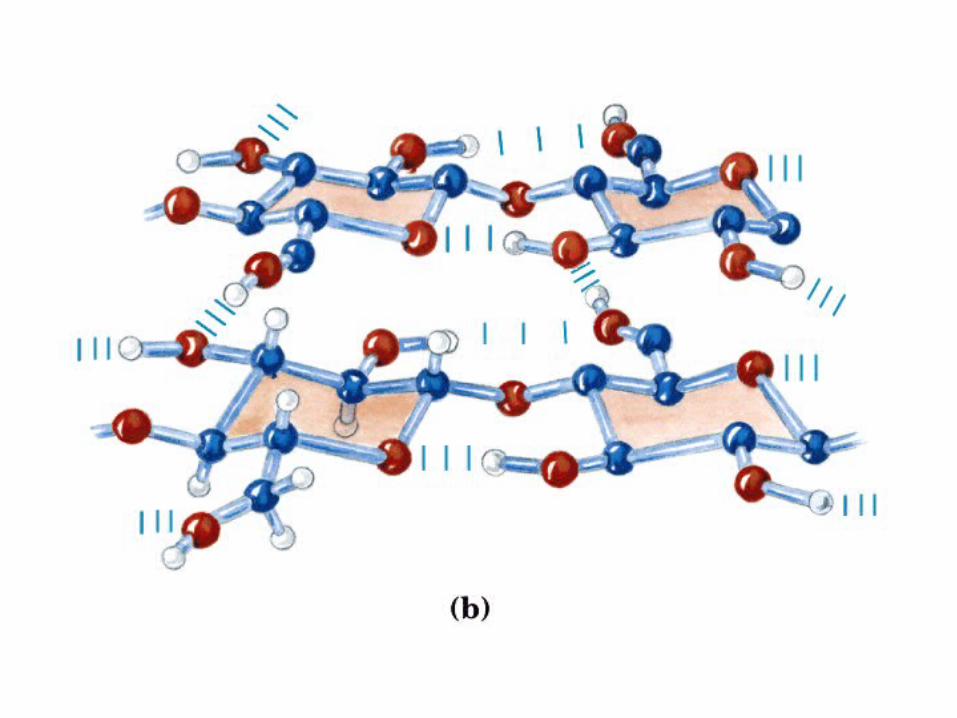
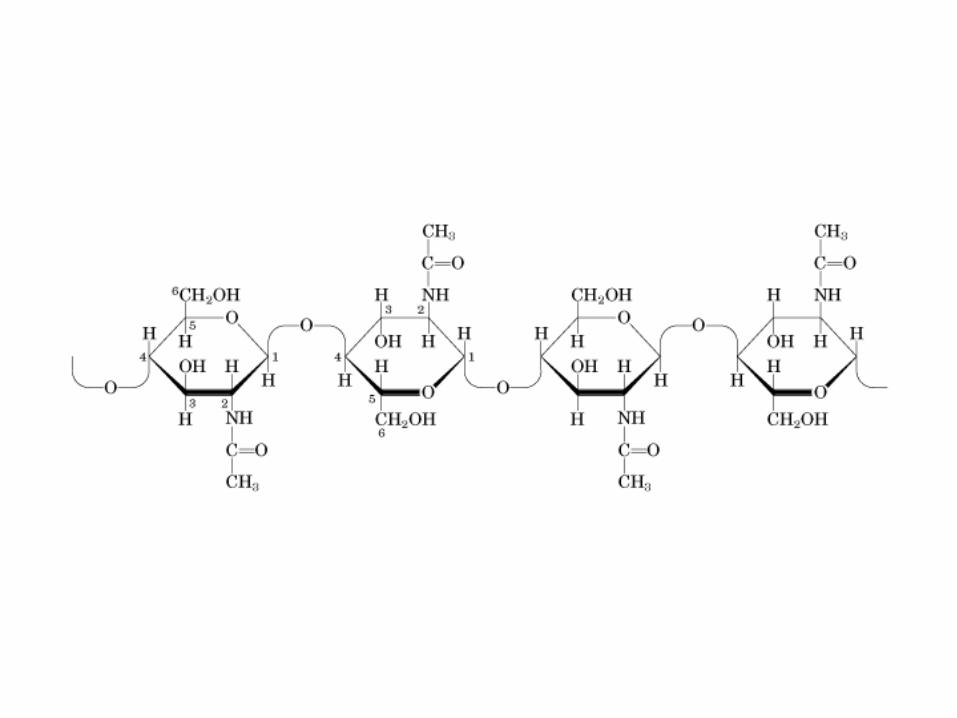
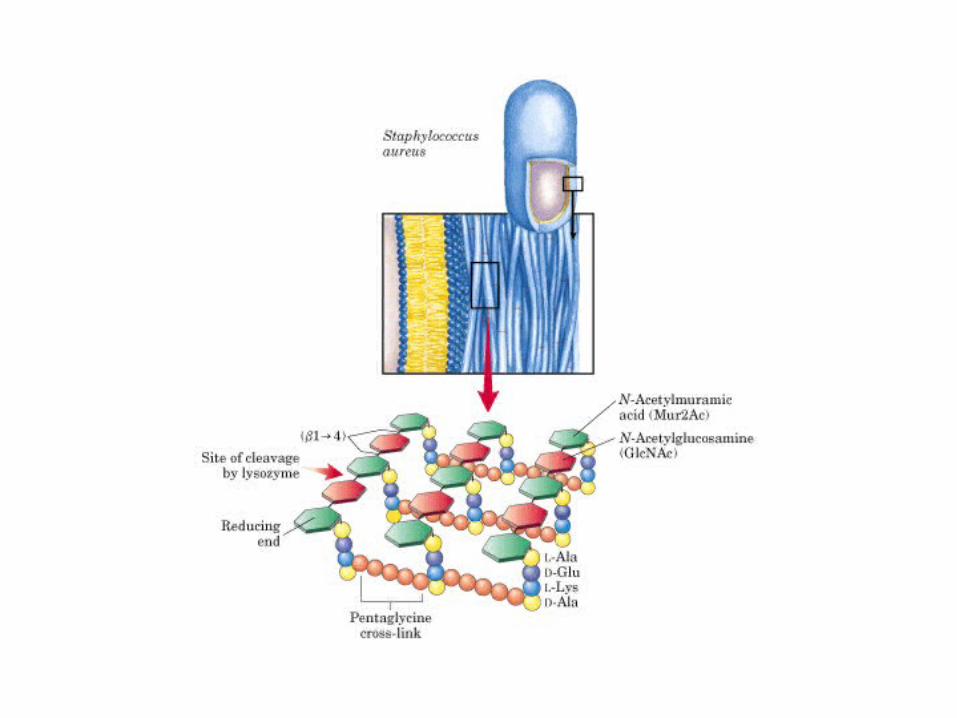
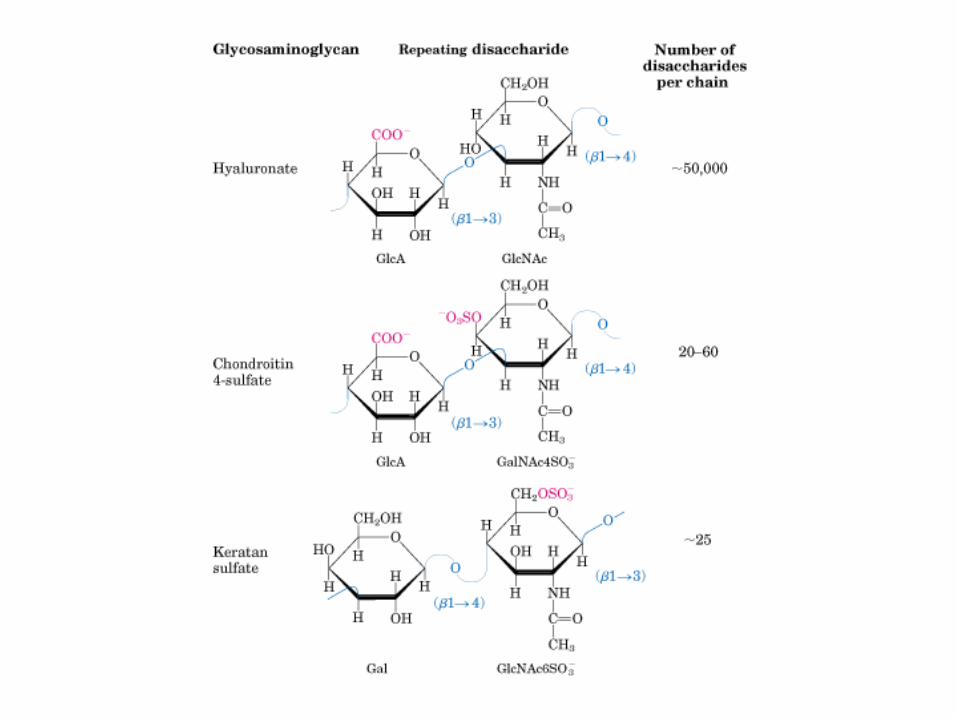
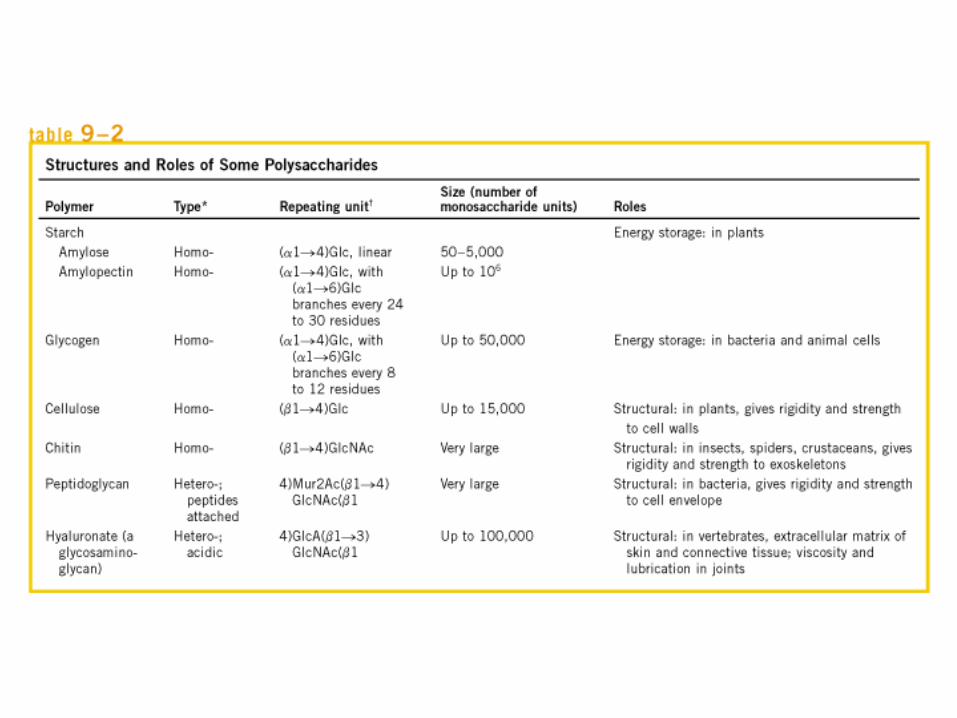

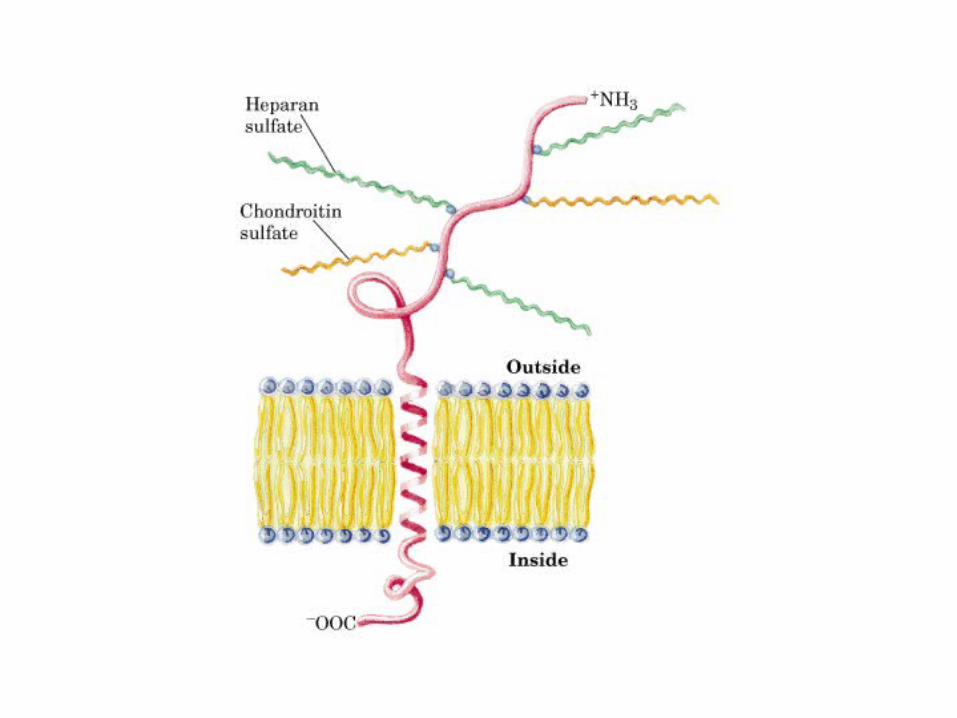
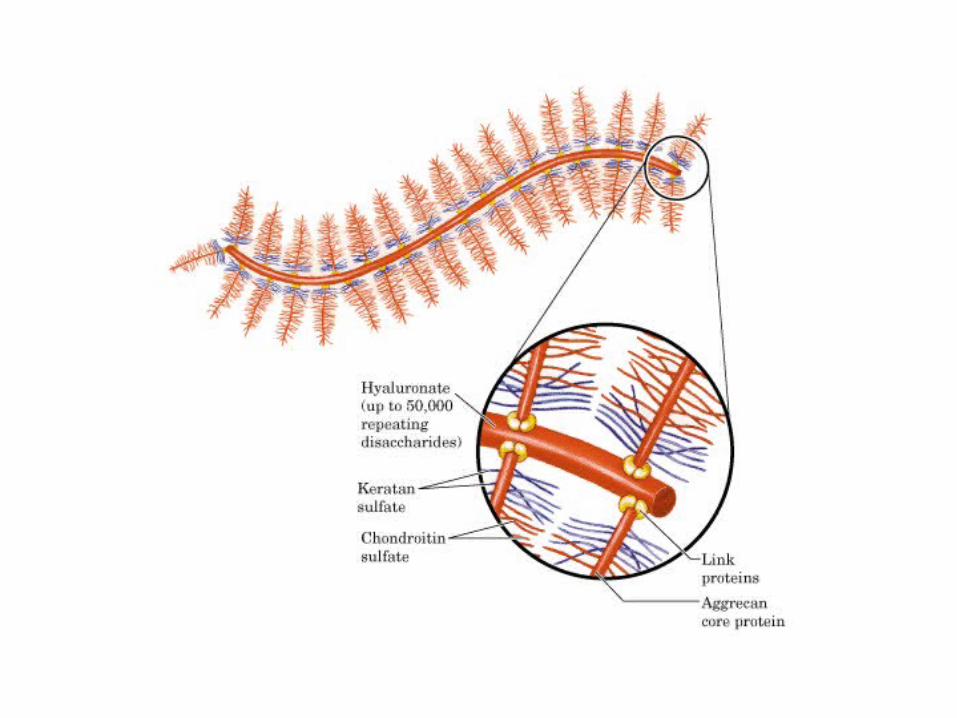
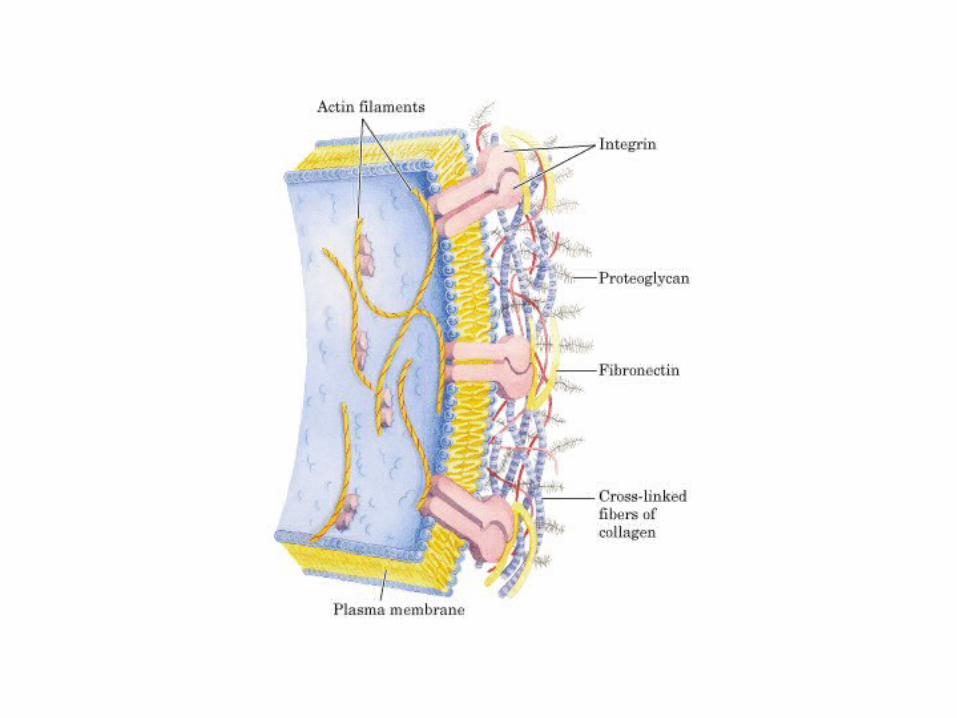
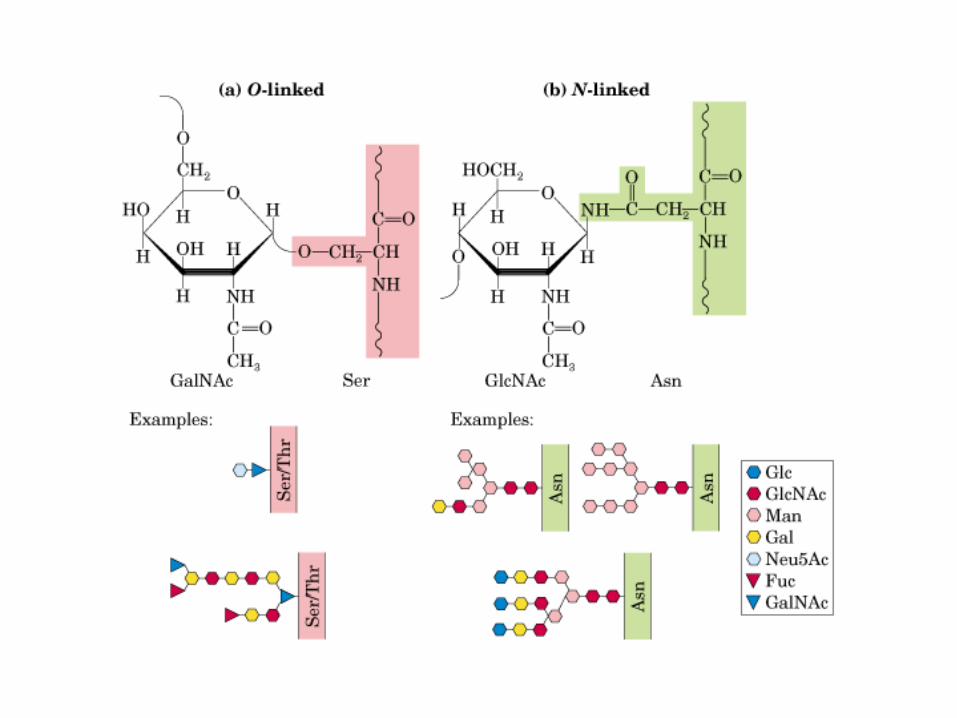
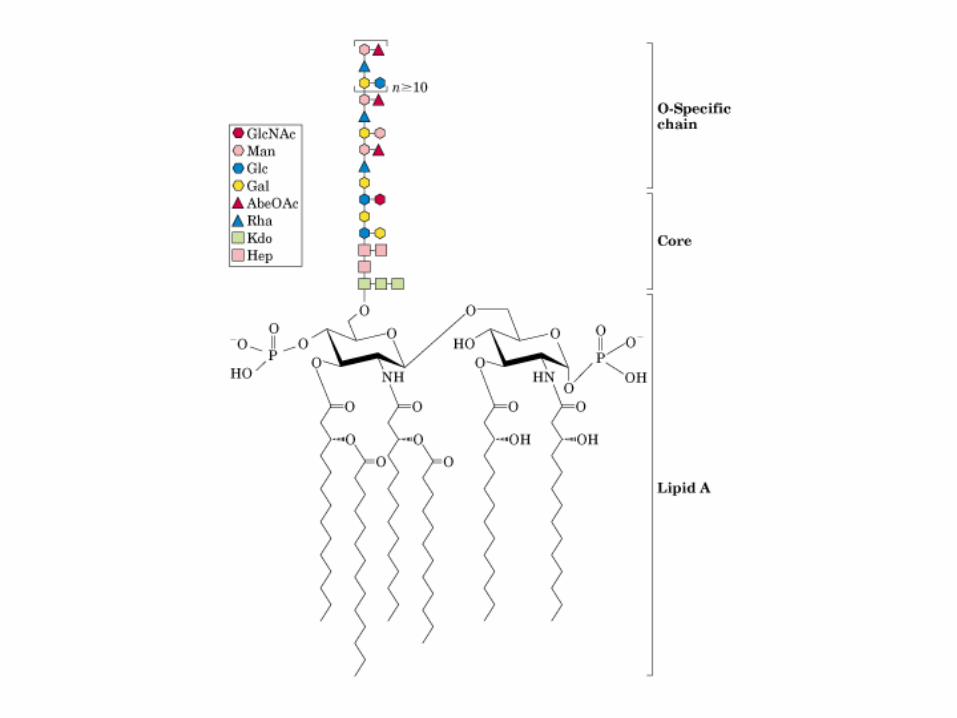
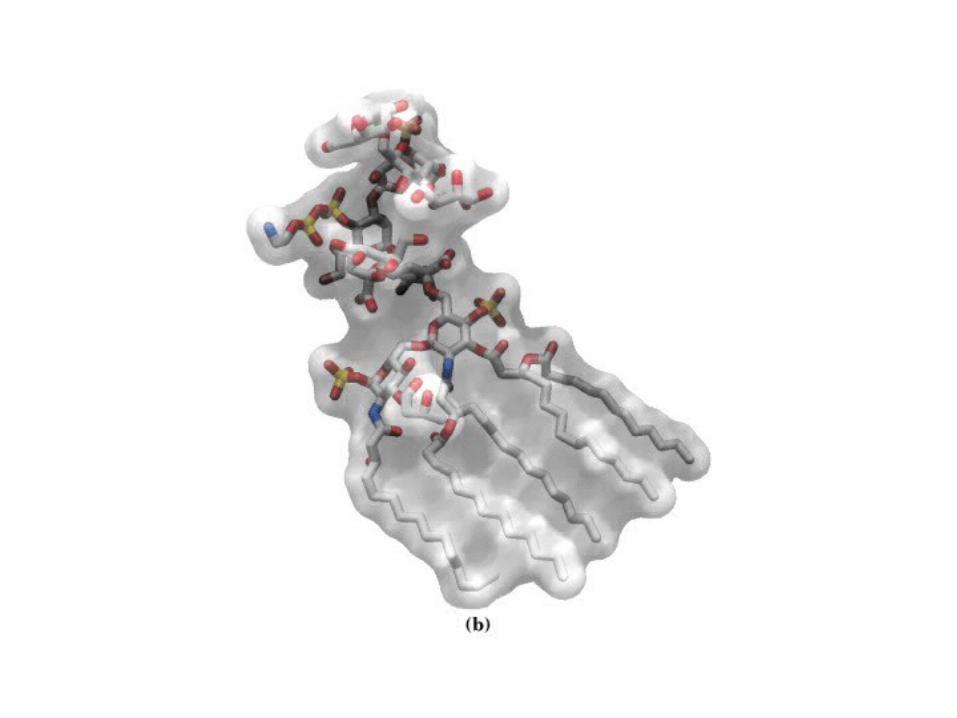
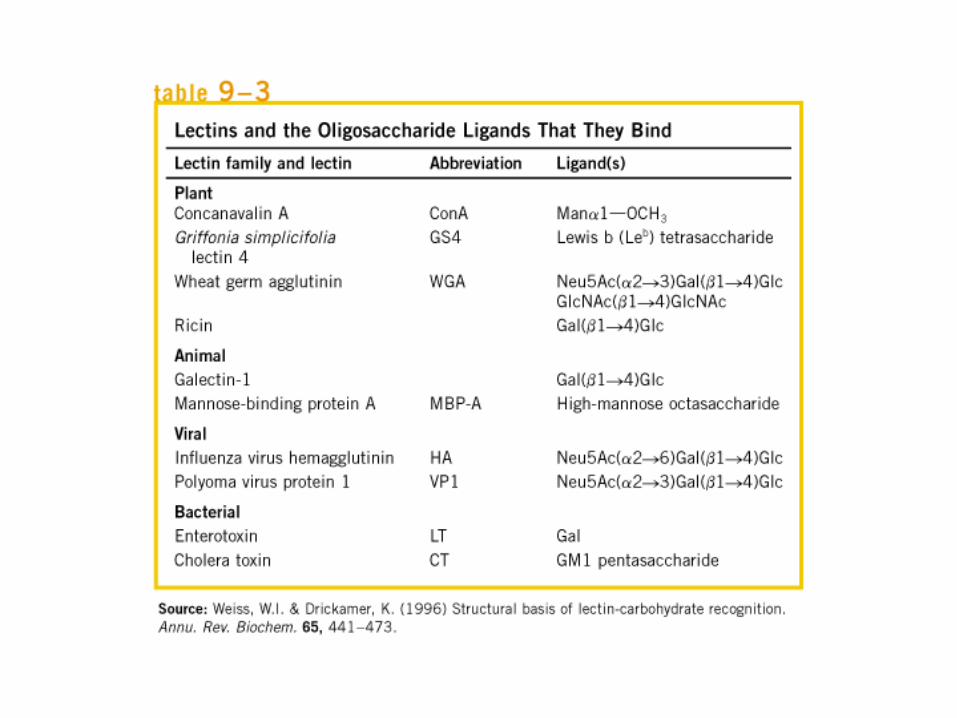
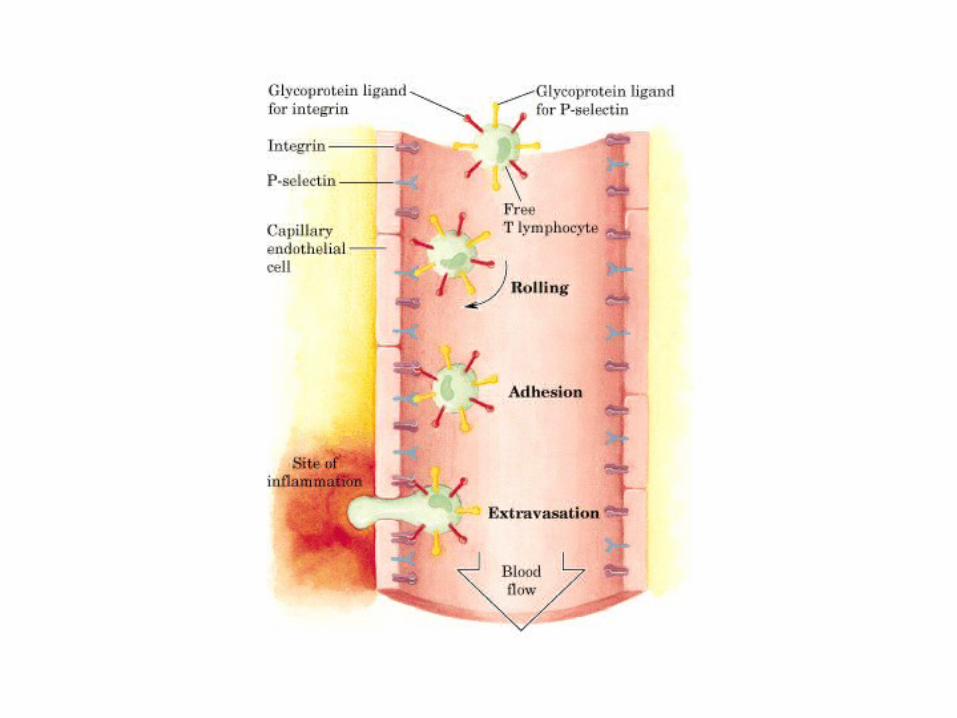
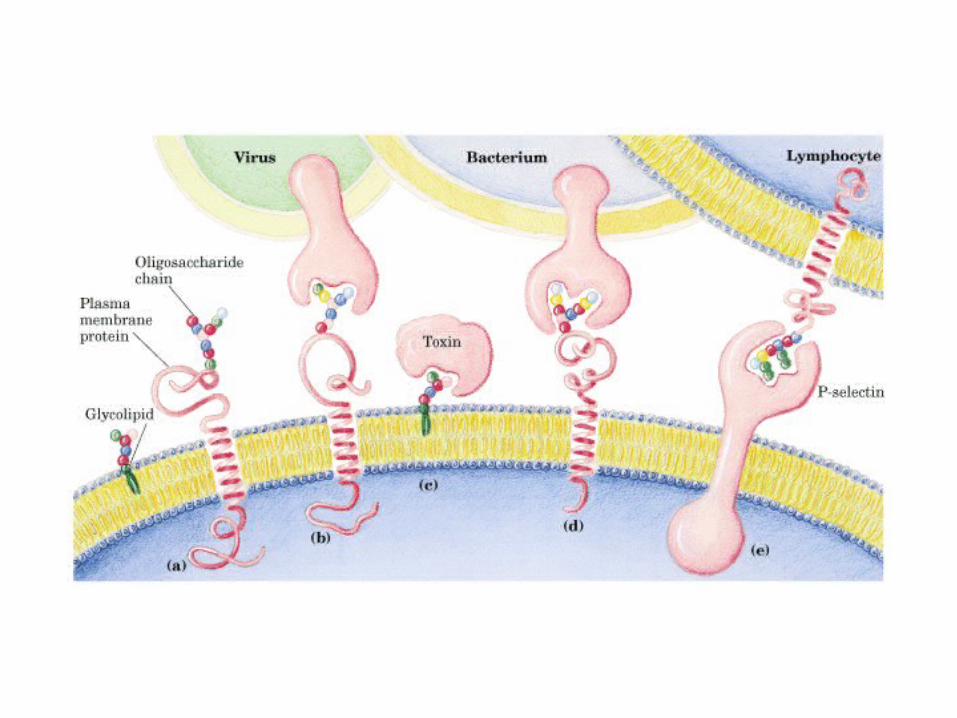


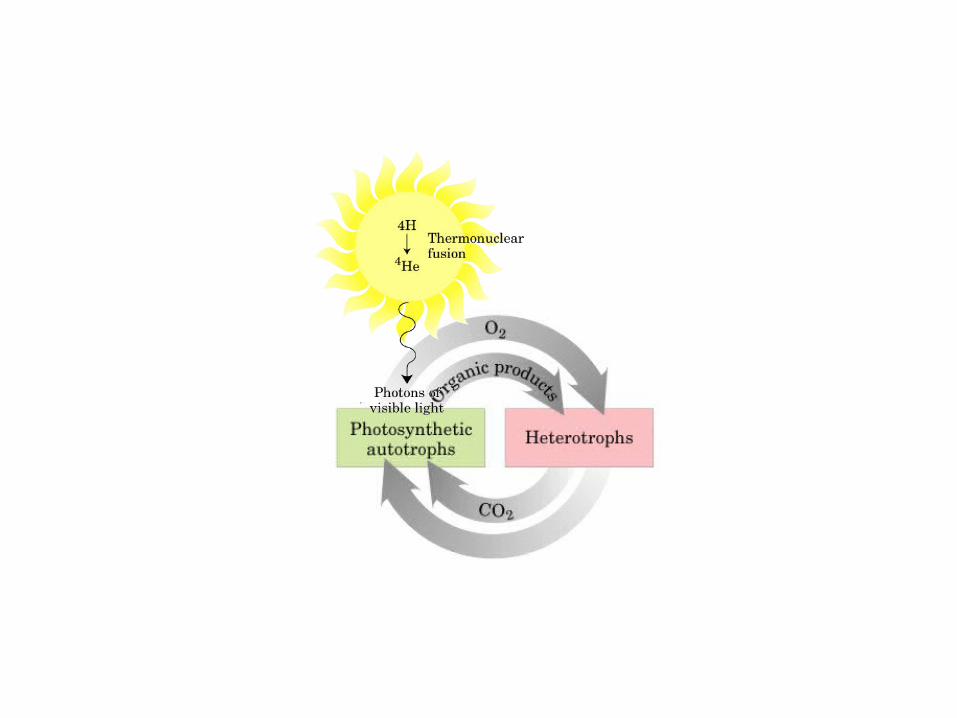
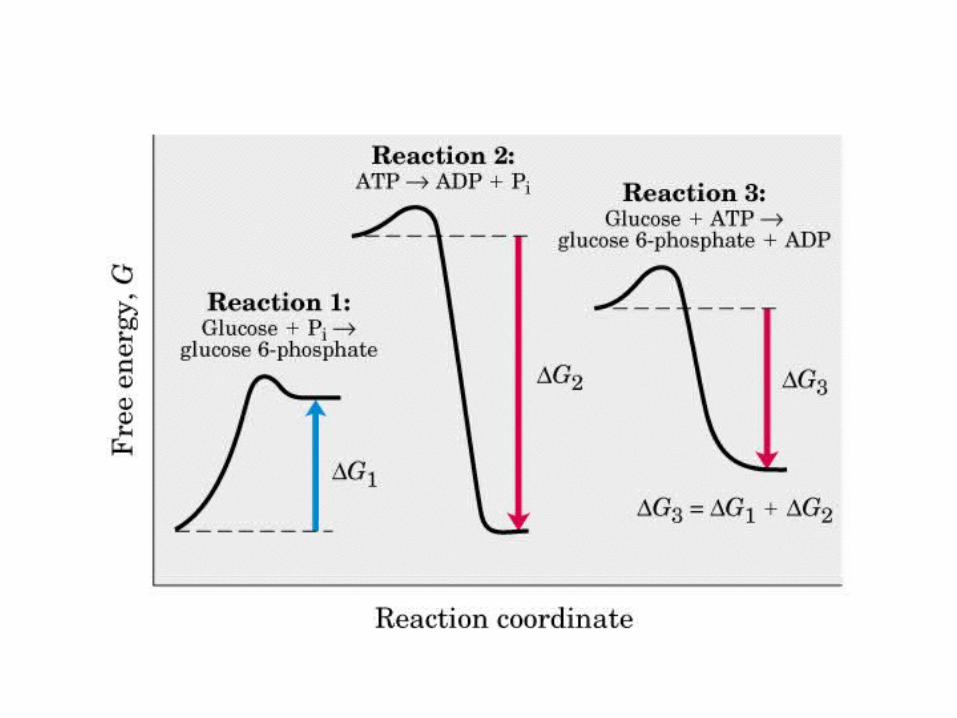
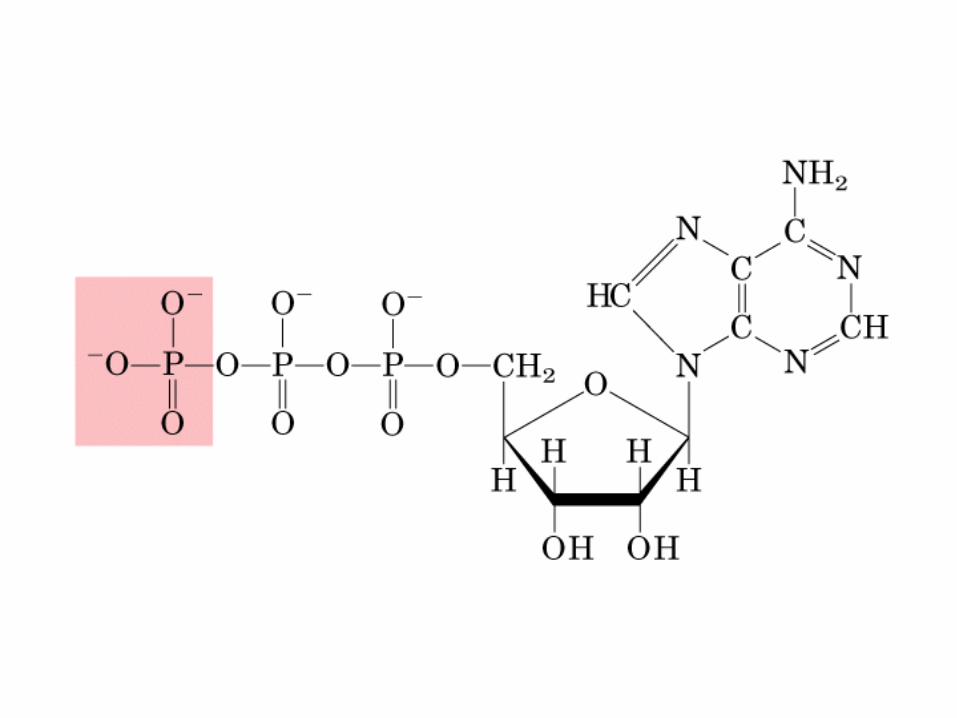
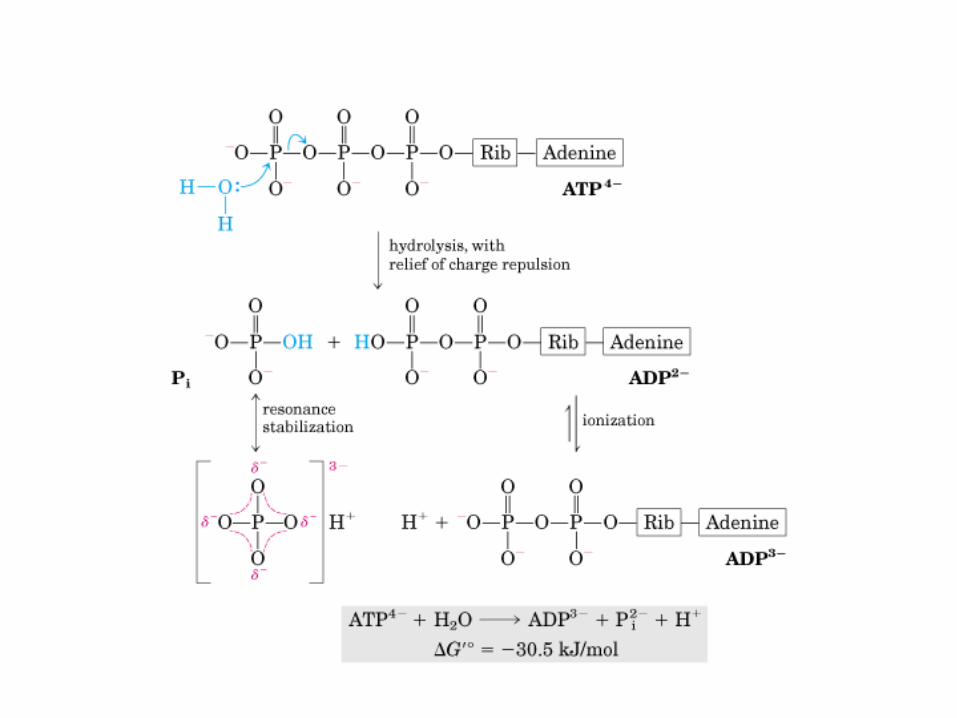
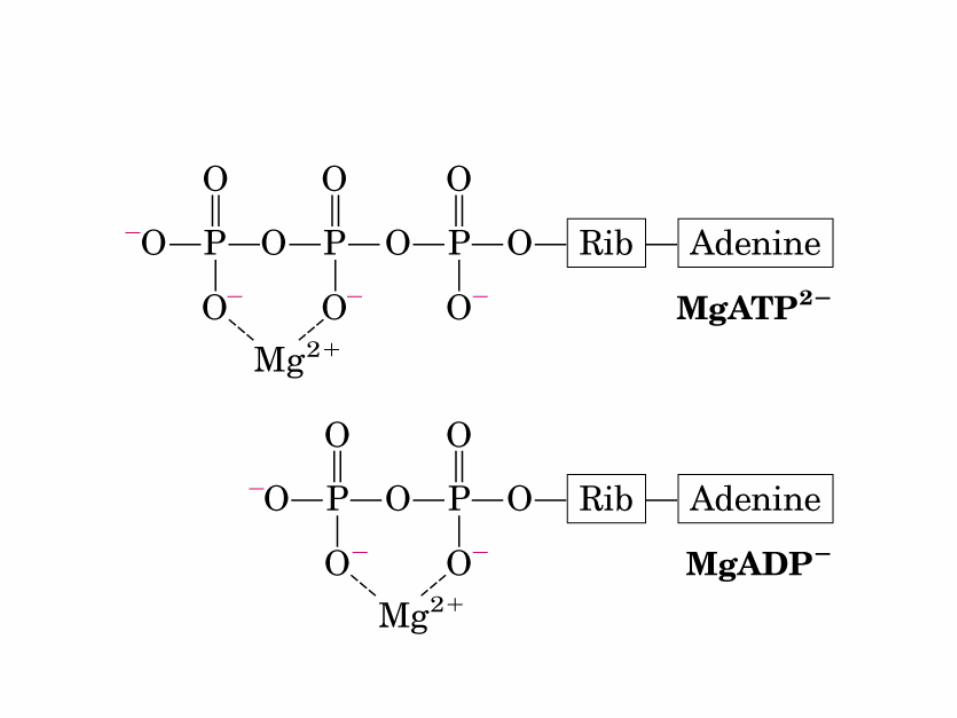
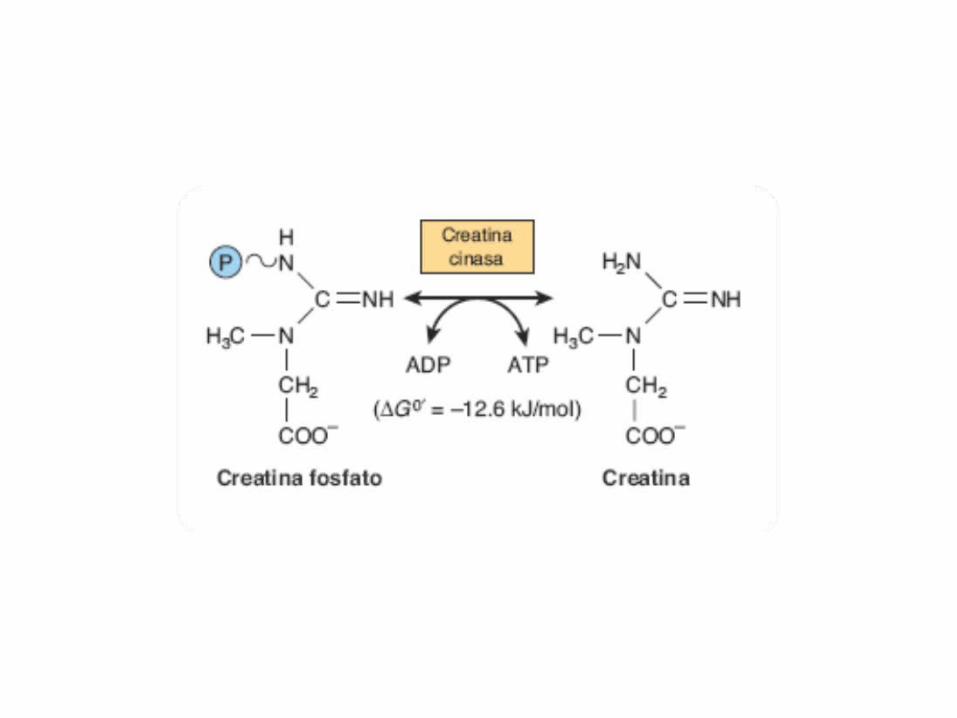
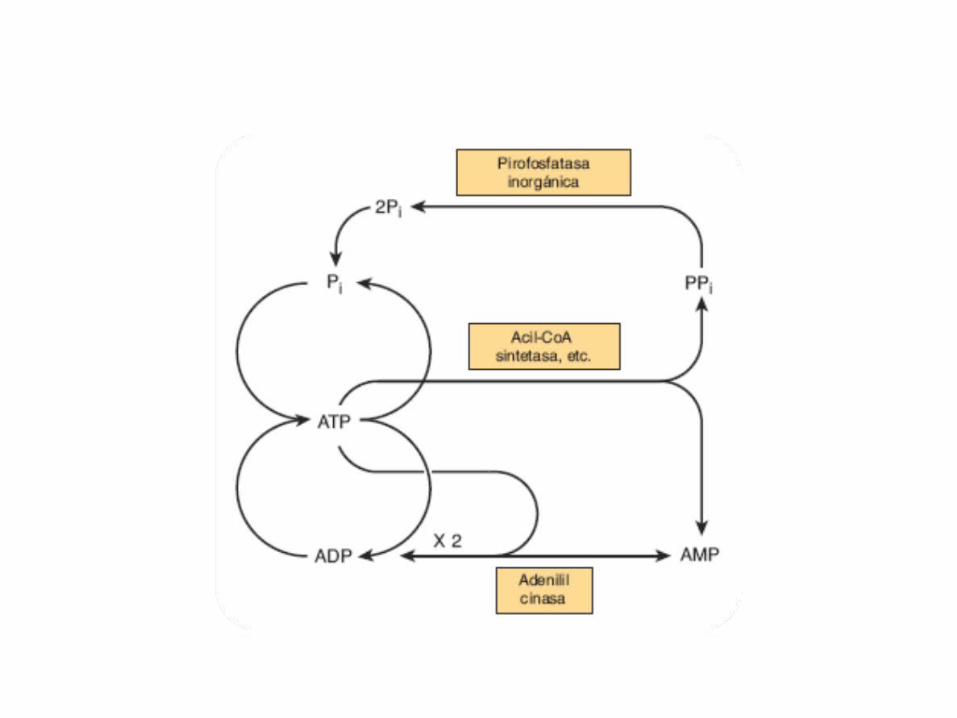
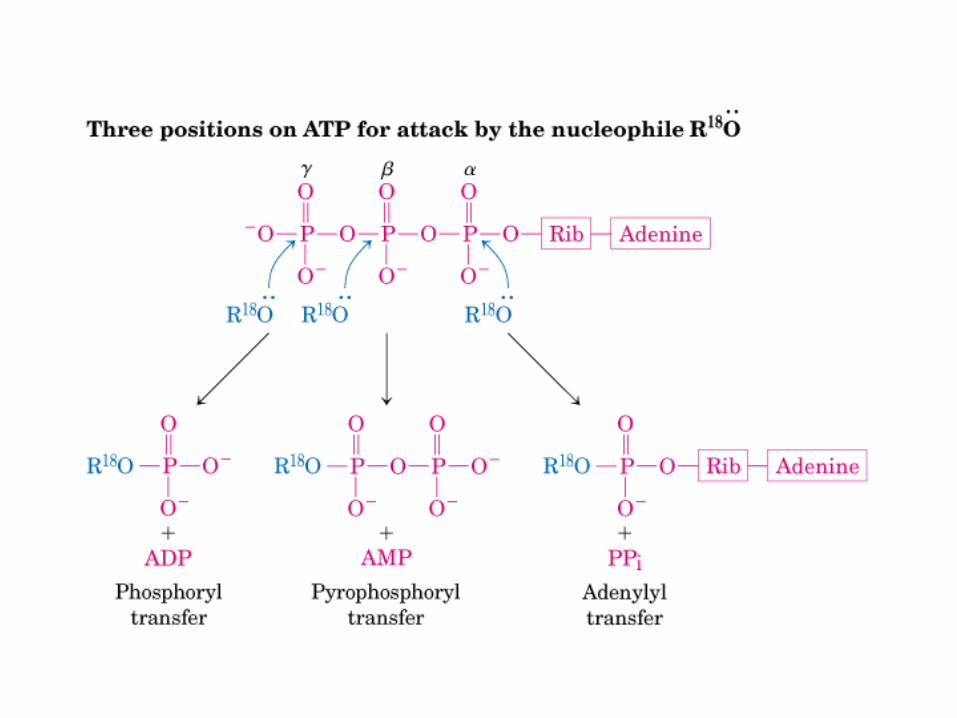
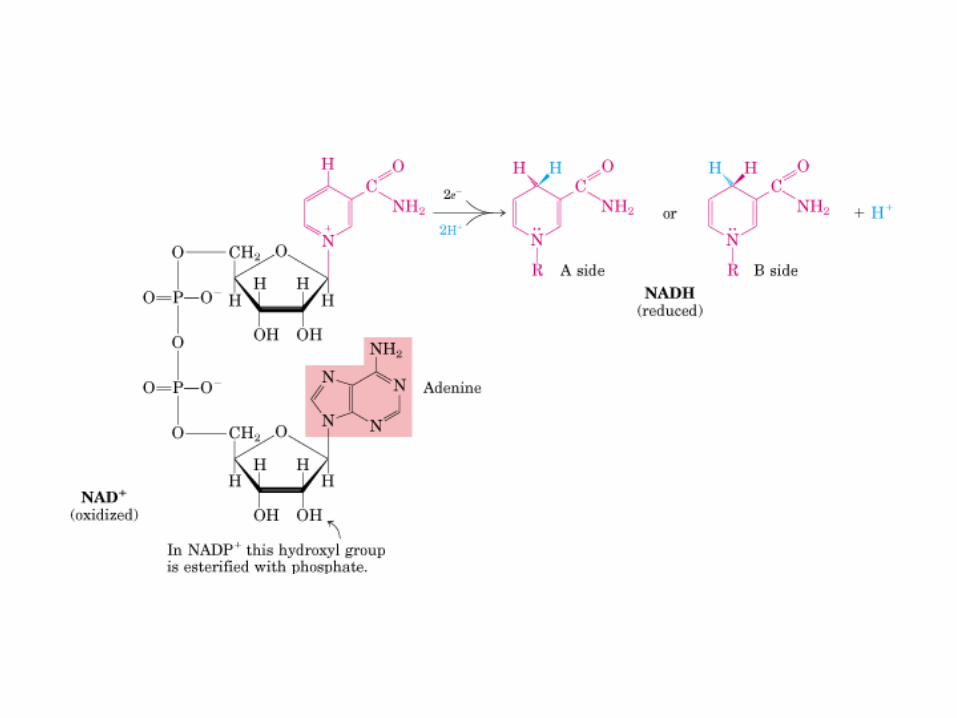
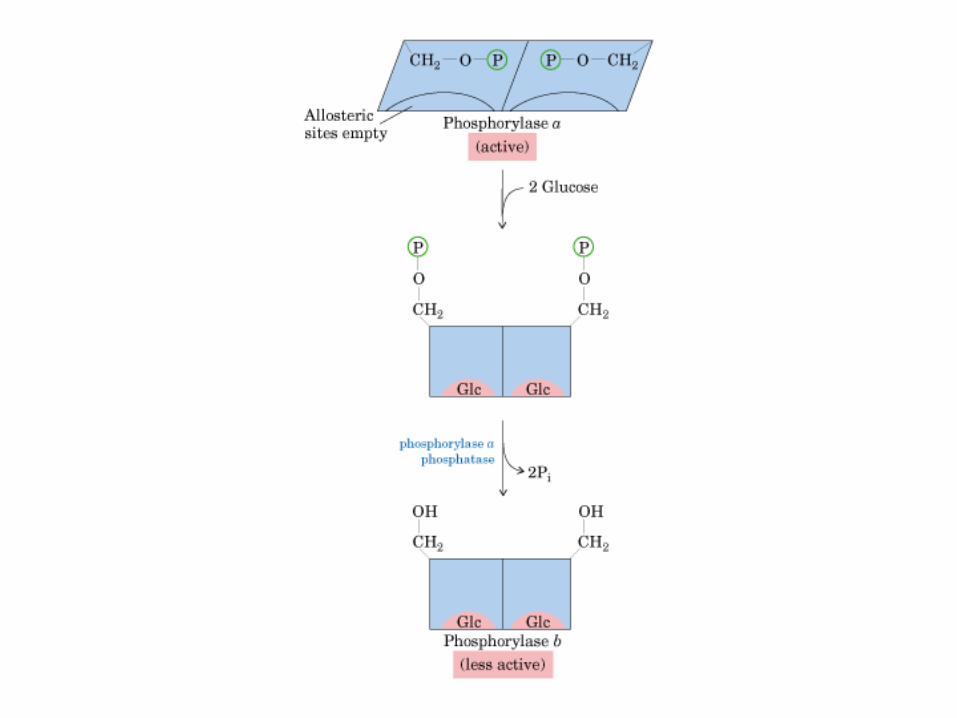
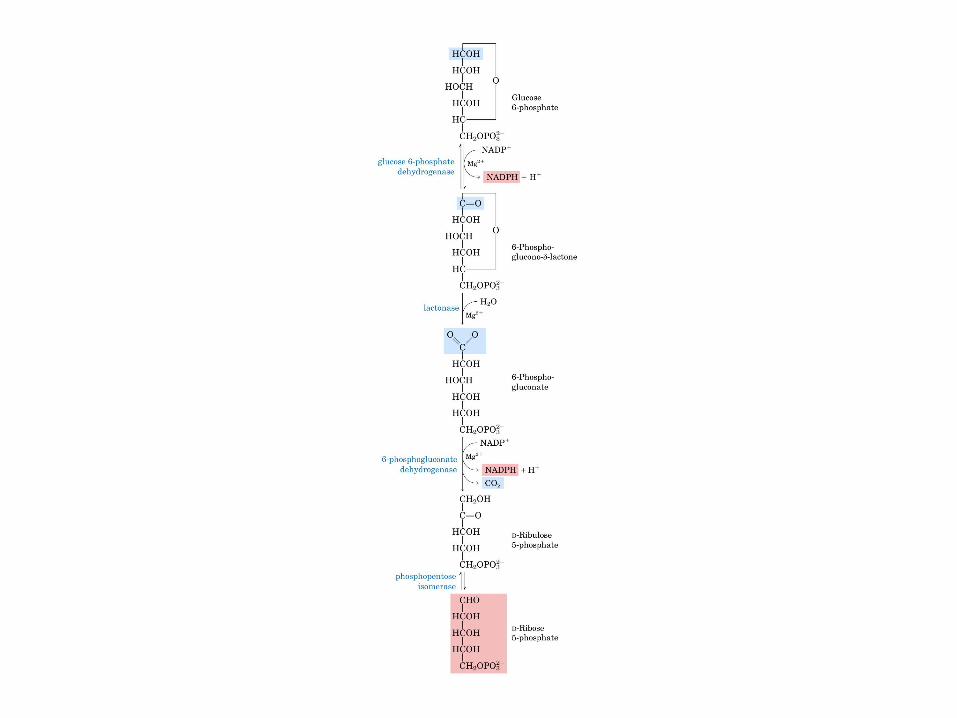
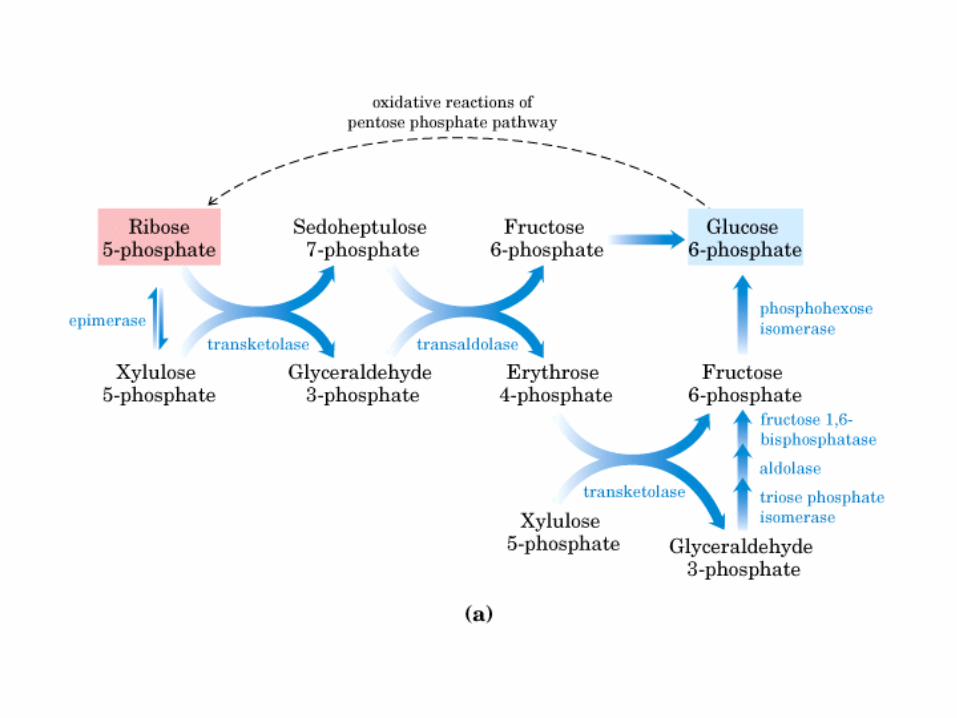
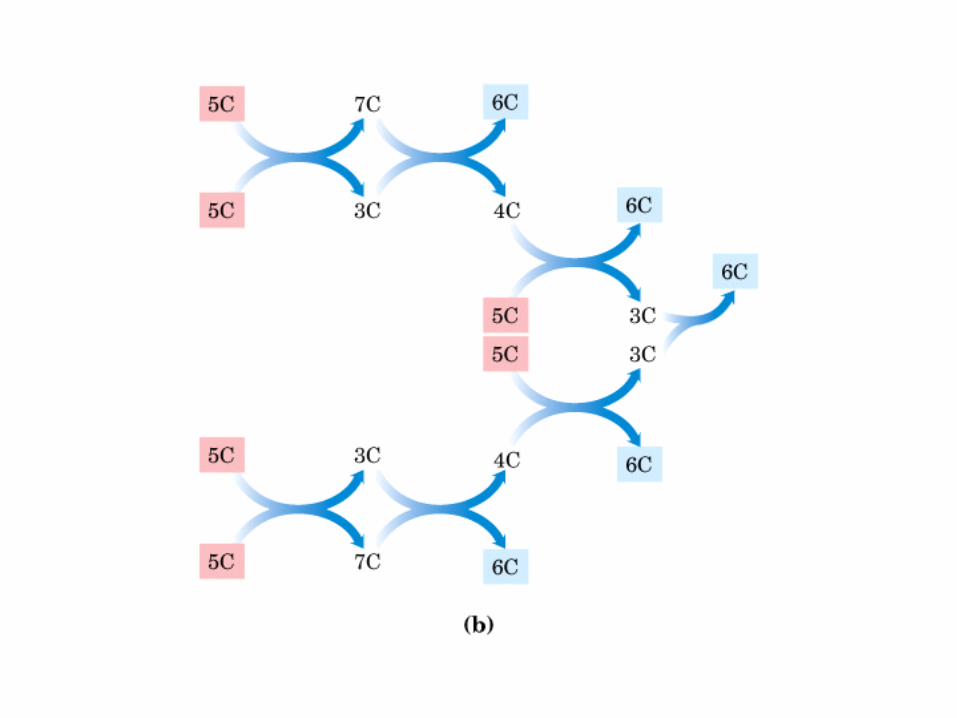
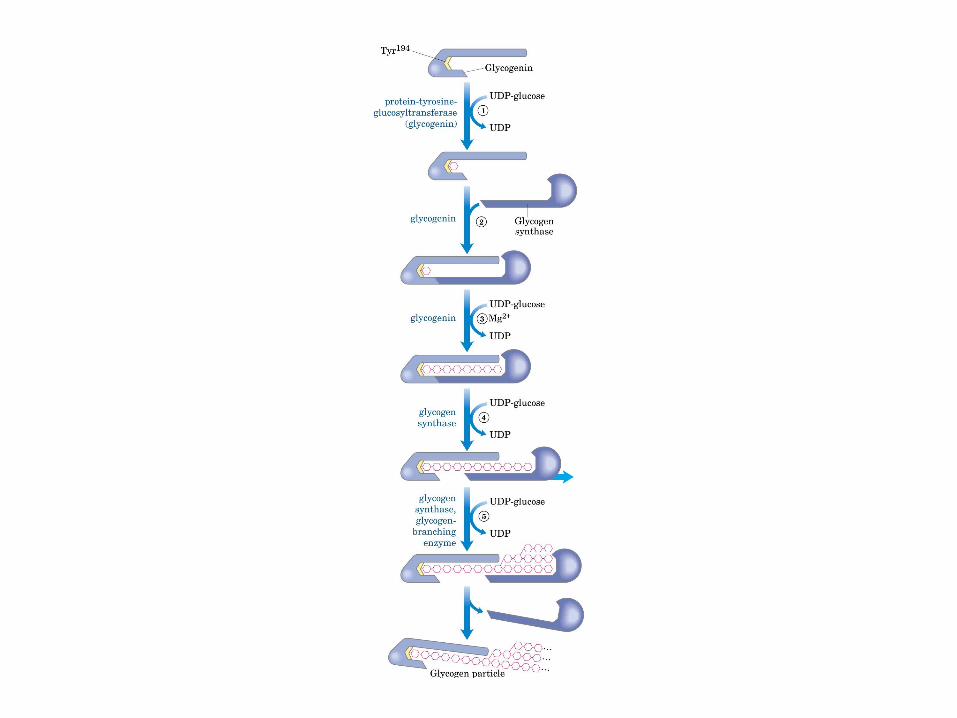
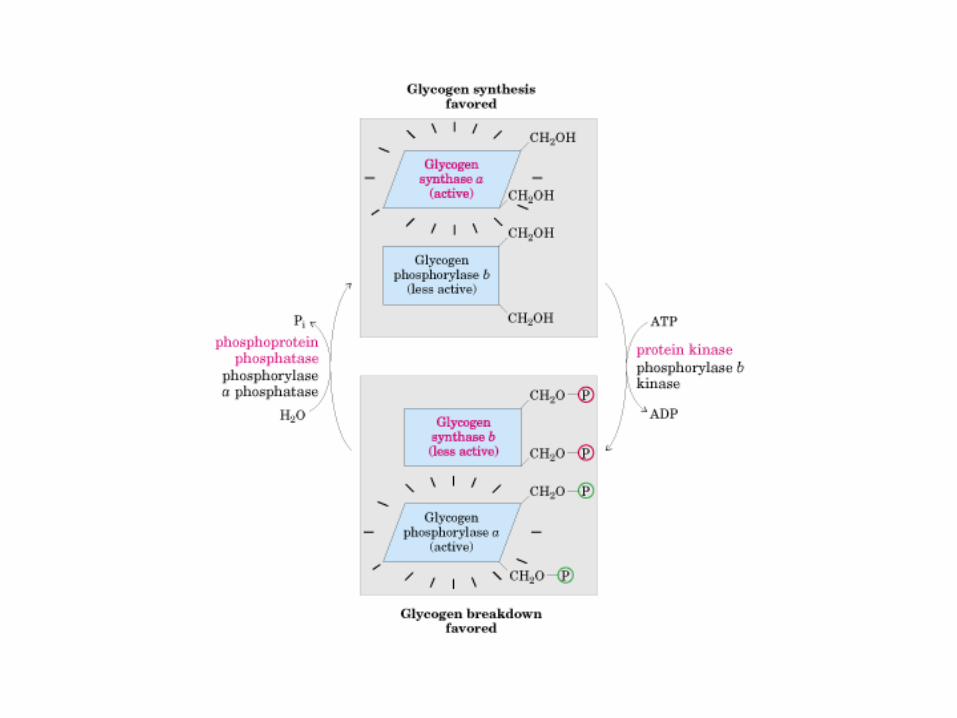
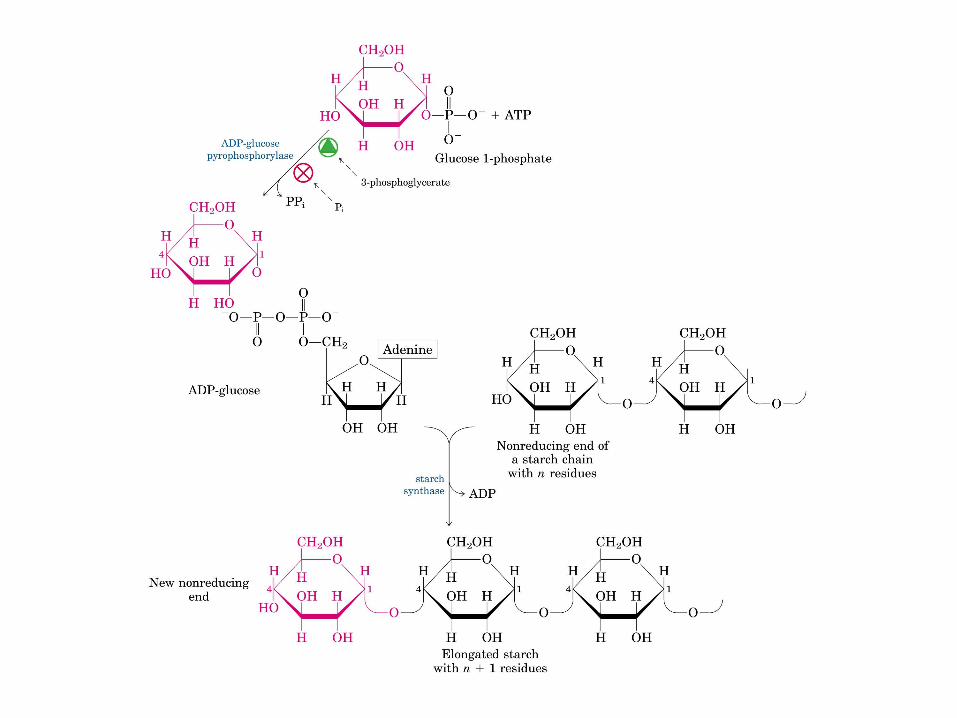
• GLUCOLISIS• GLUCONEOGENESIS• GLUCOGENOGENESIS• GLUCOGENOLISIS• VIA DE LAS PENTOSAS• VIA DE LOS POLIOLES• CICLO DE CORI• CICLO DE LA ALANINA
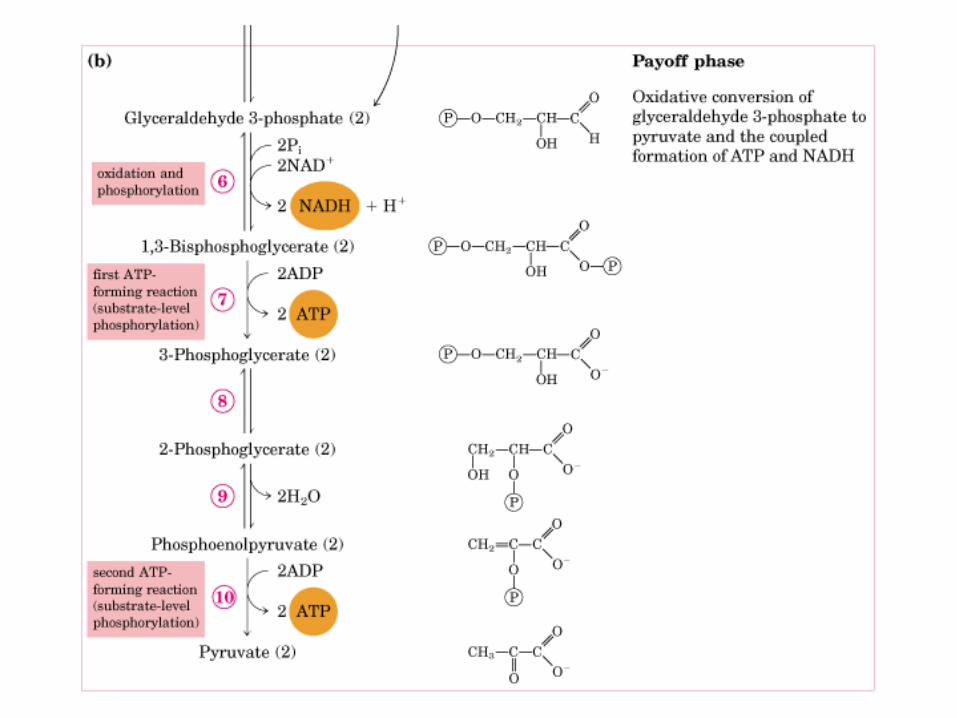
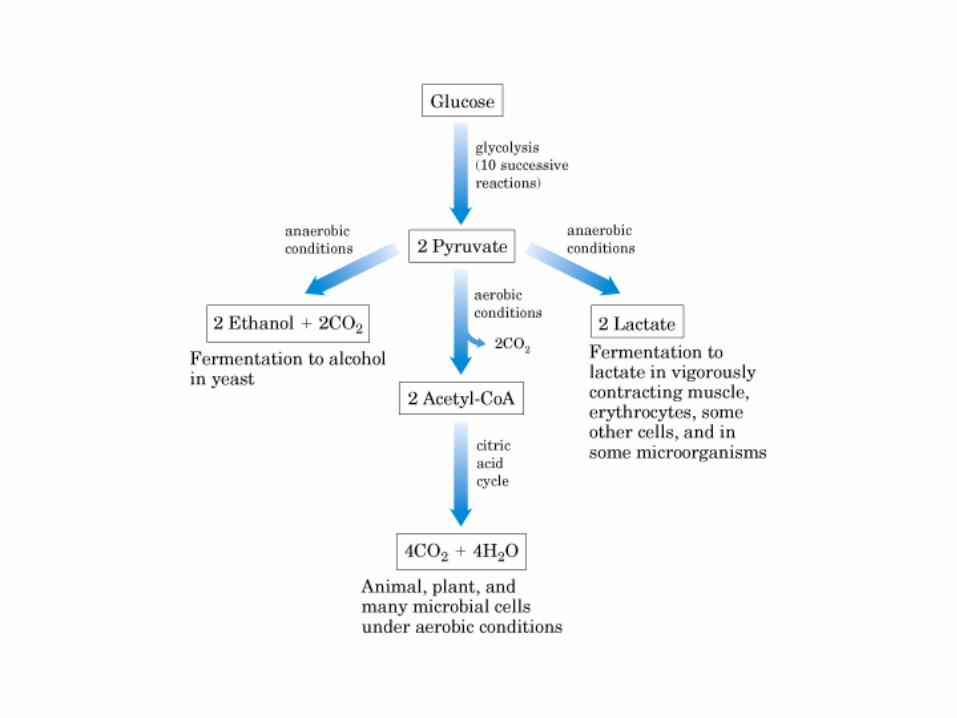
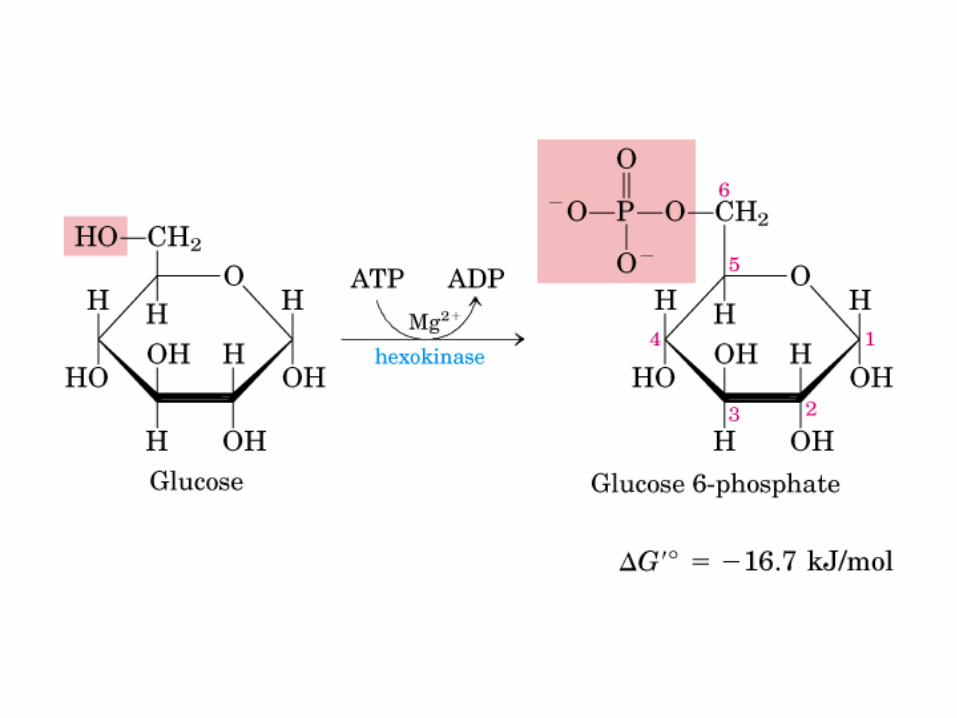
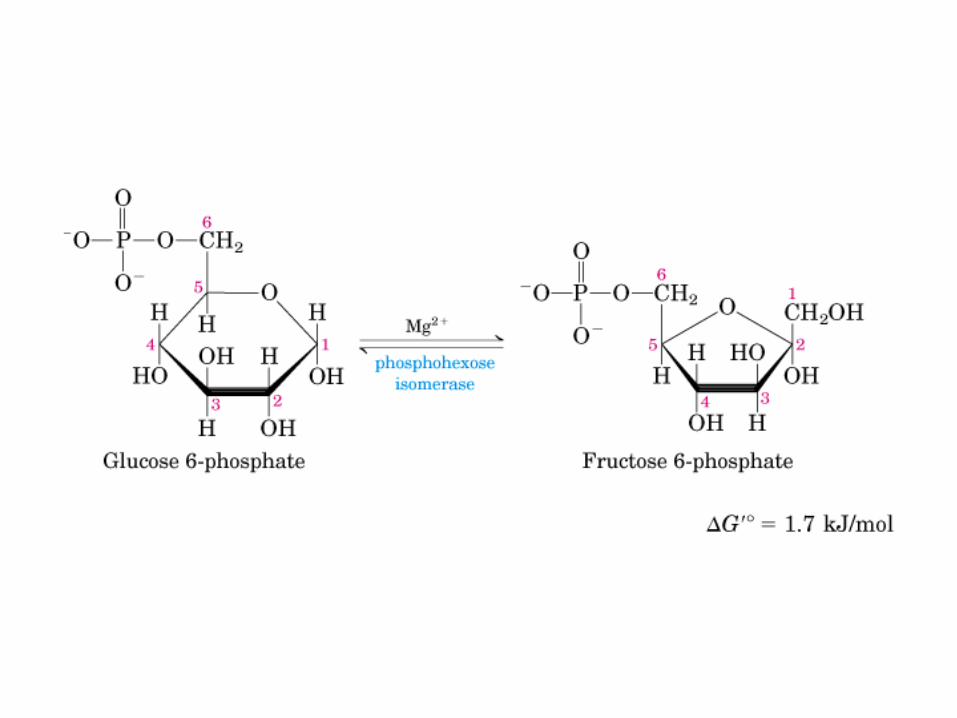
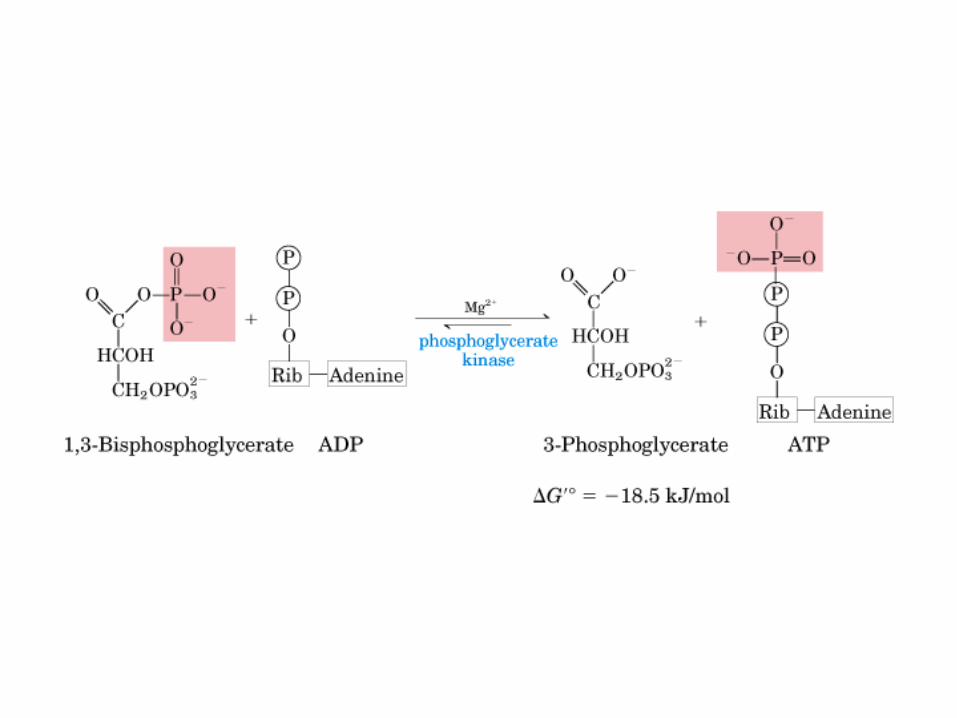
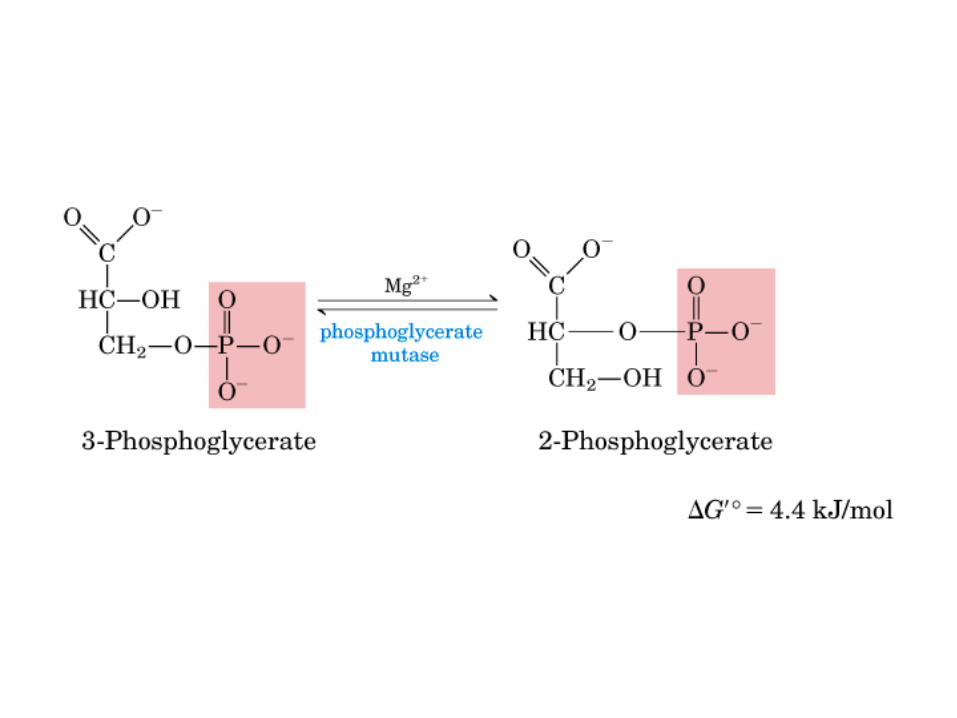
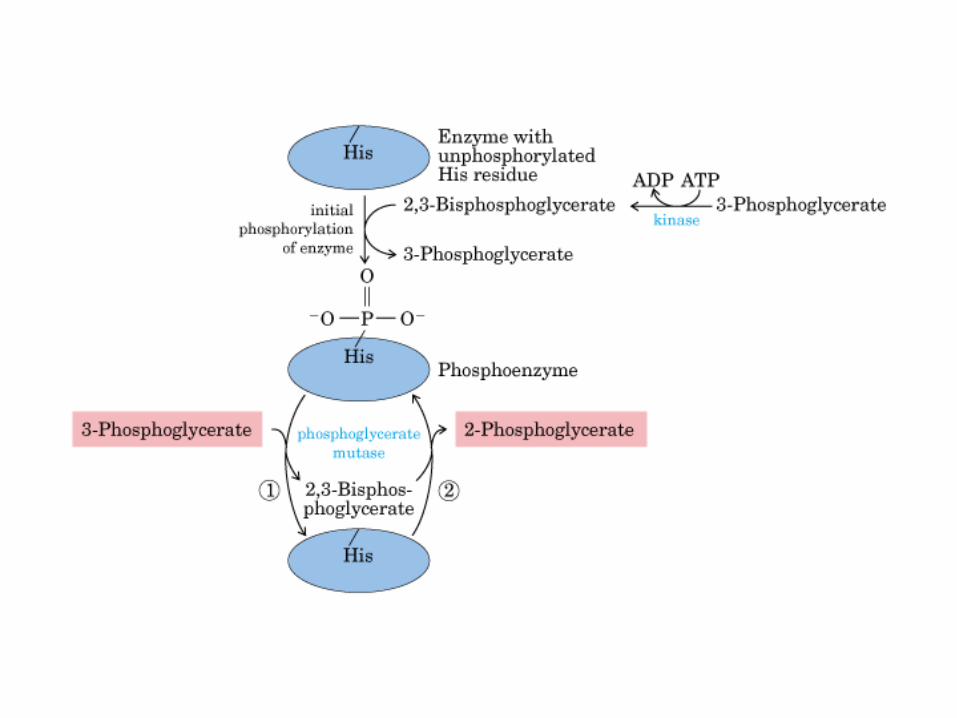
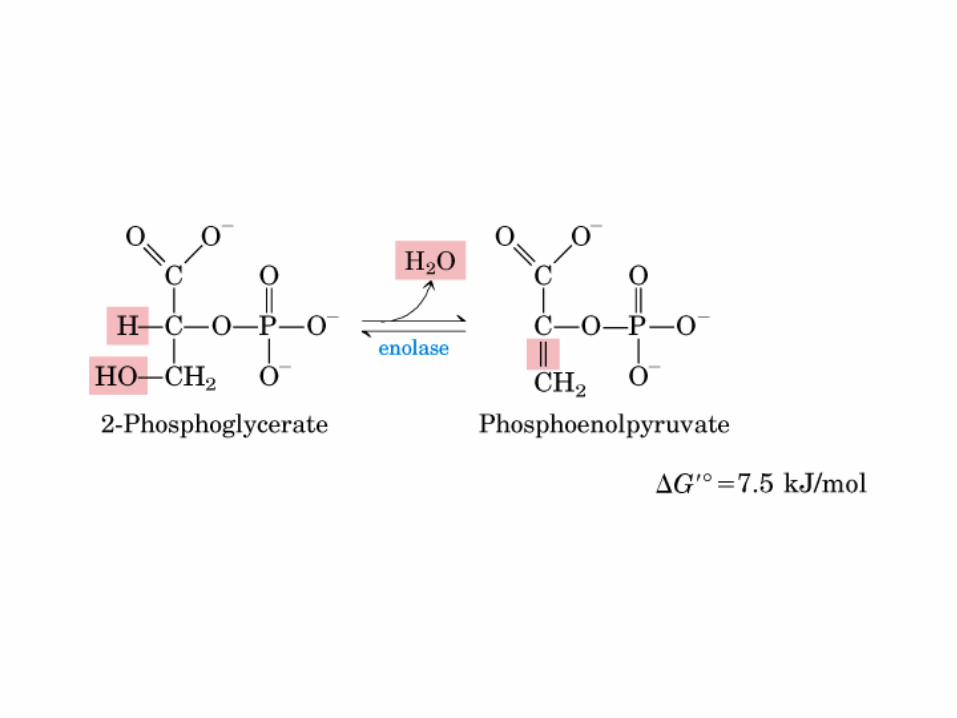
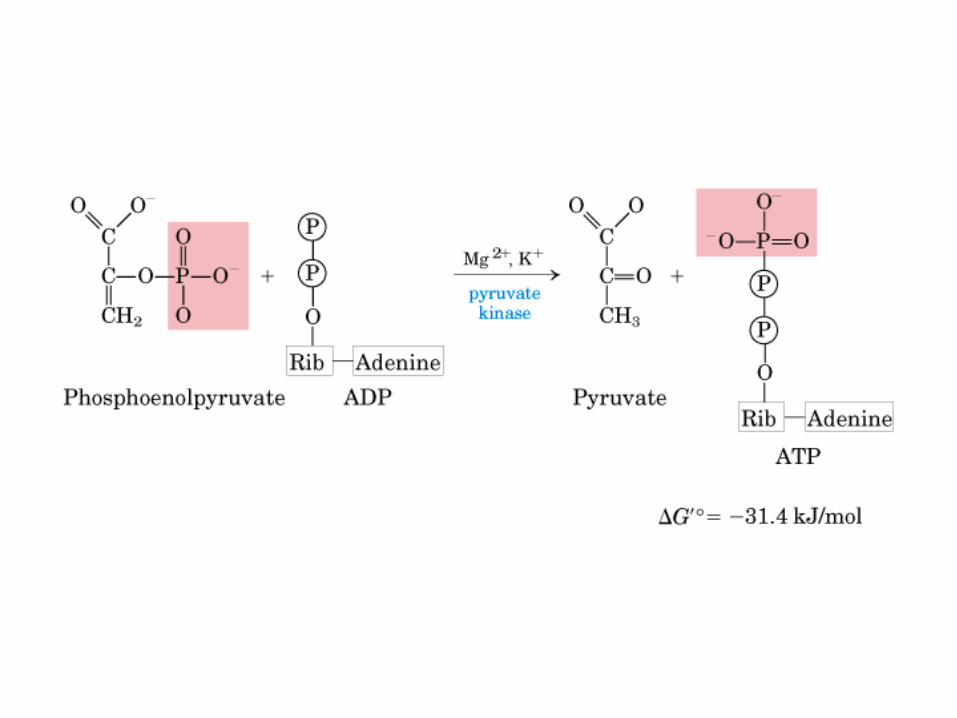
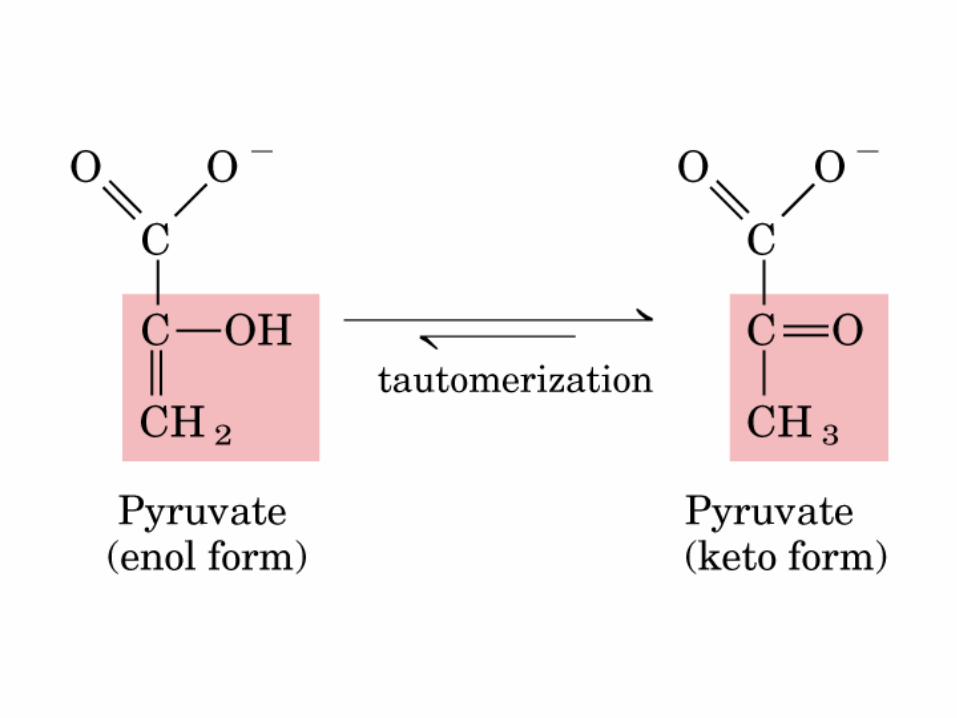
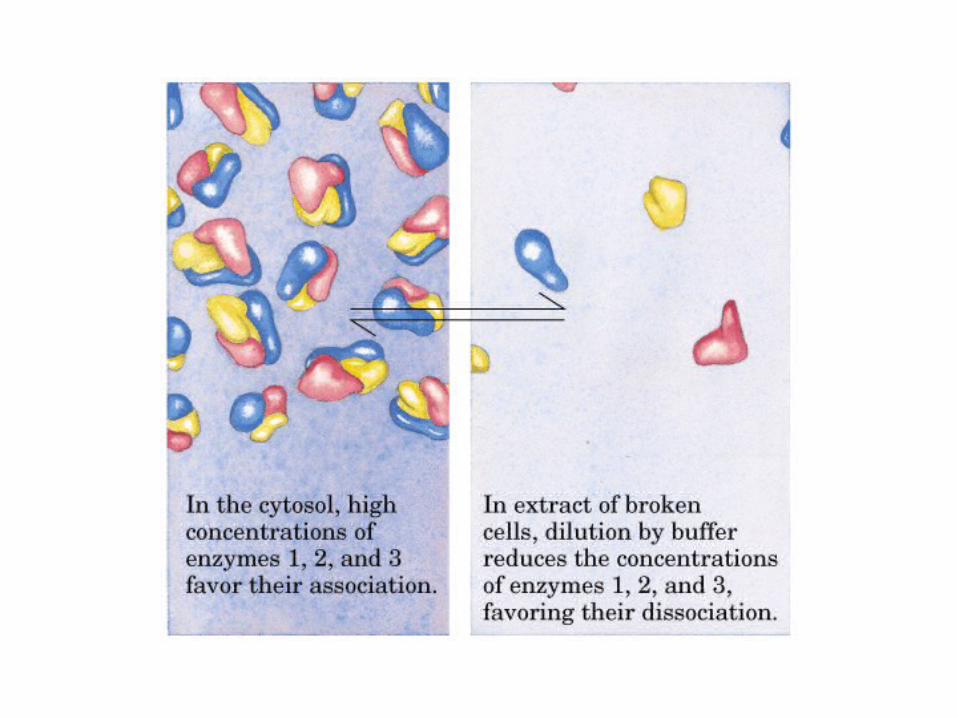
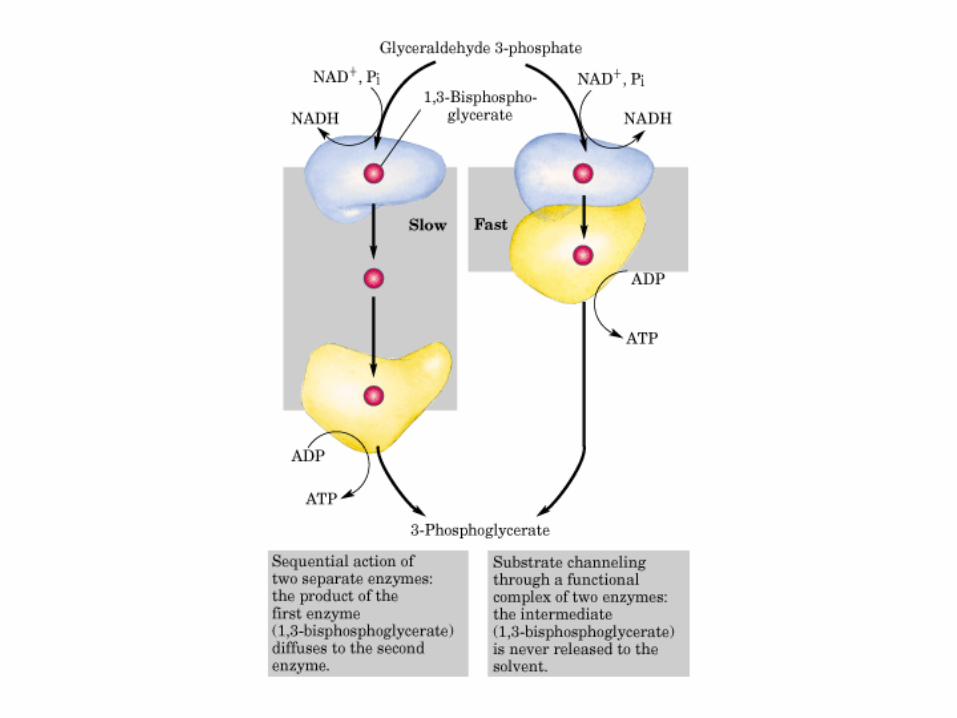
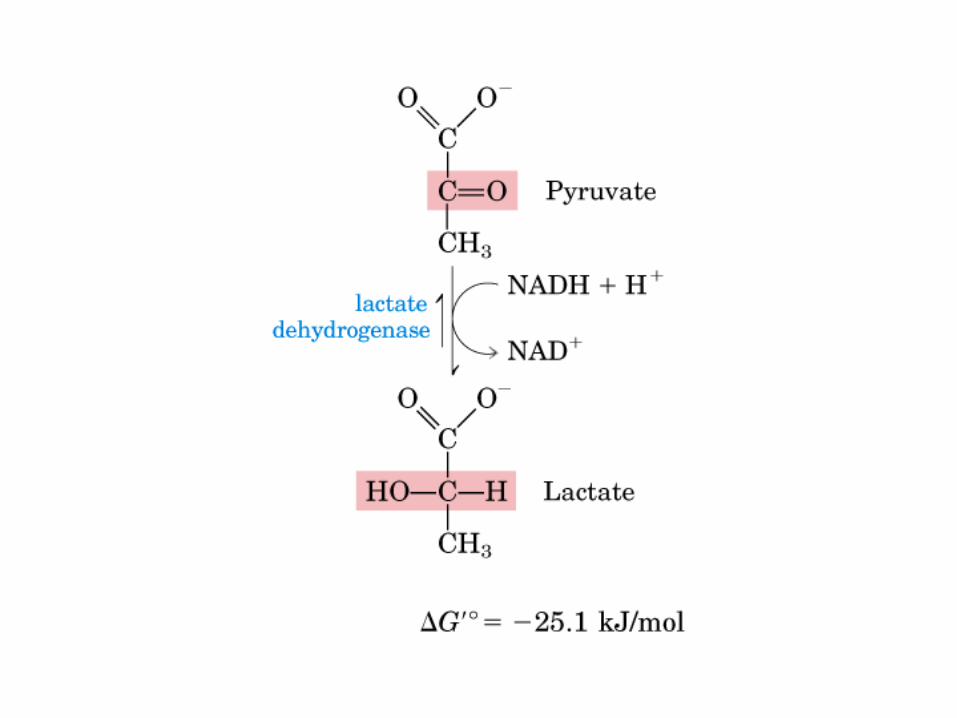

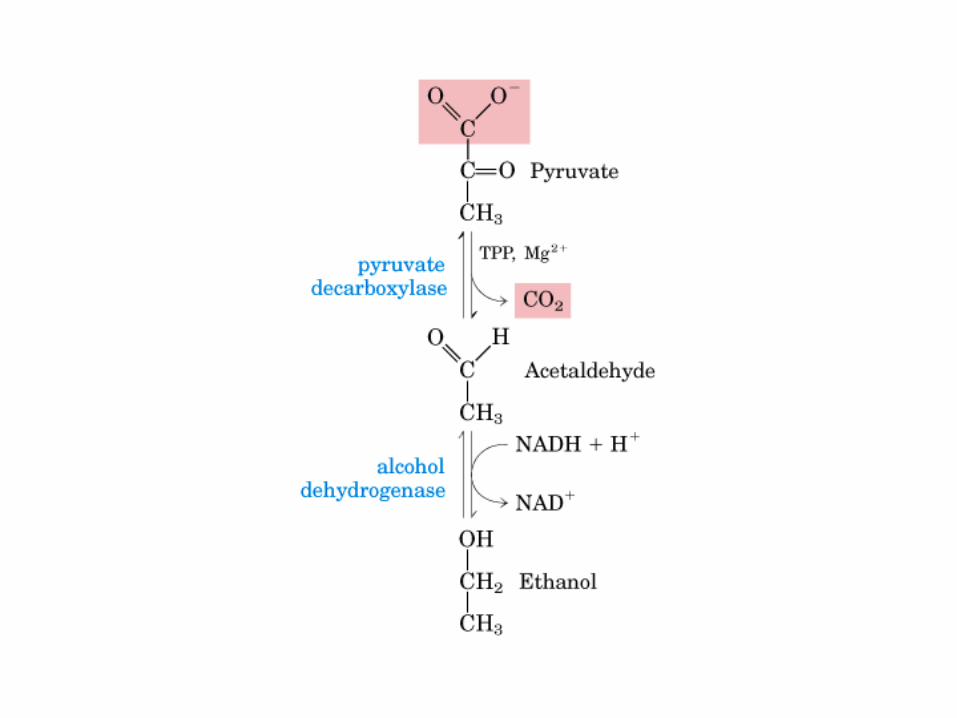
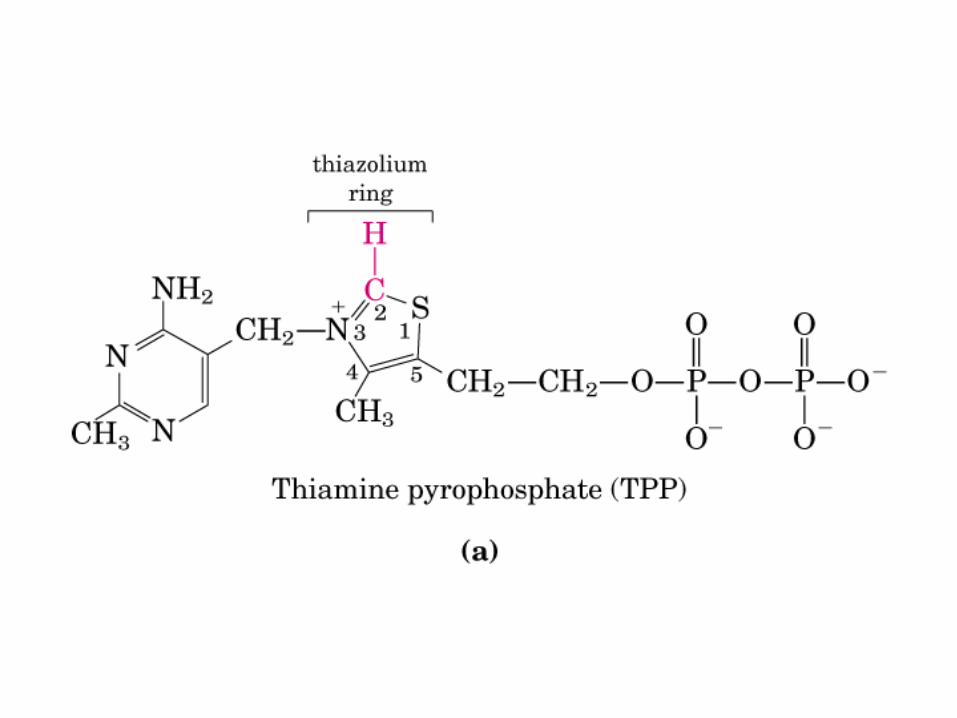
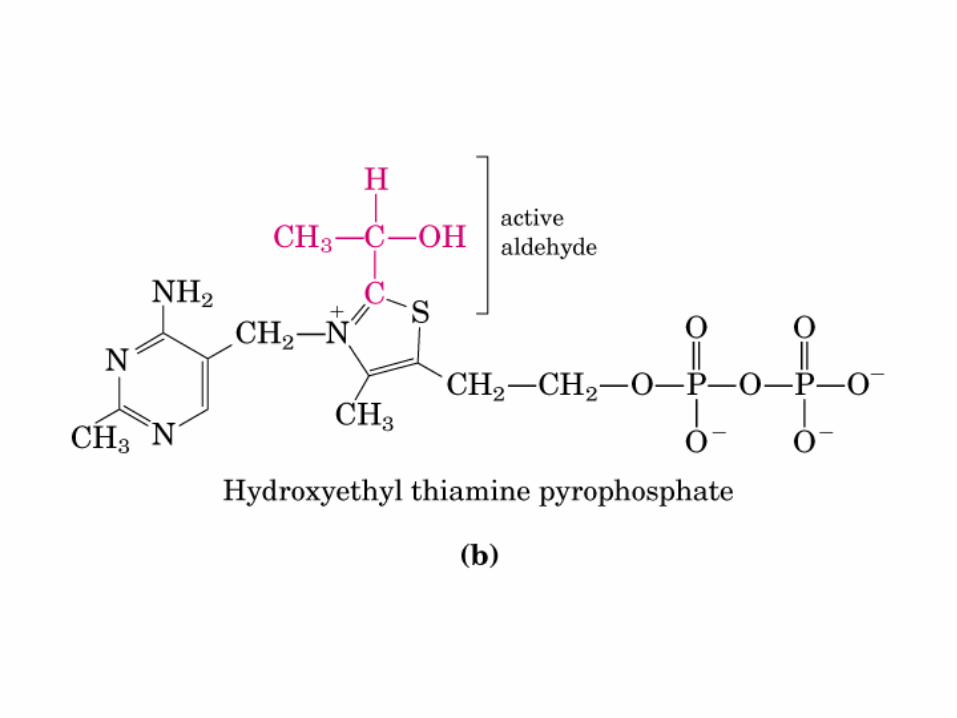
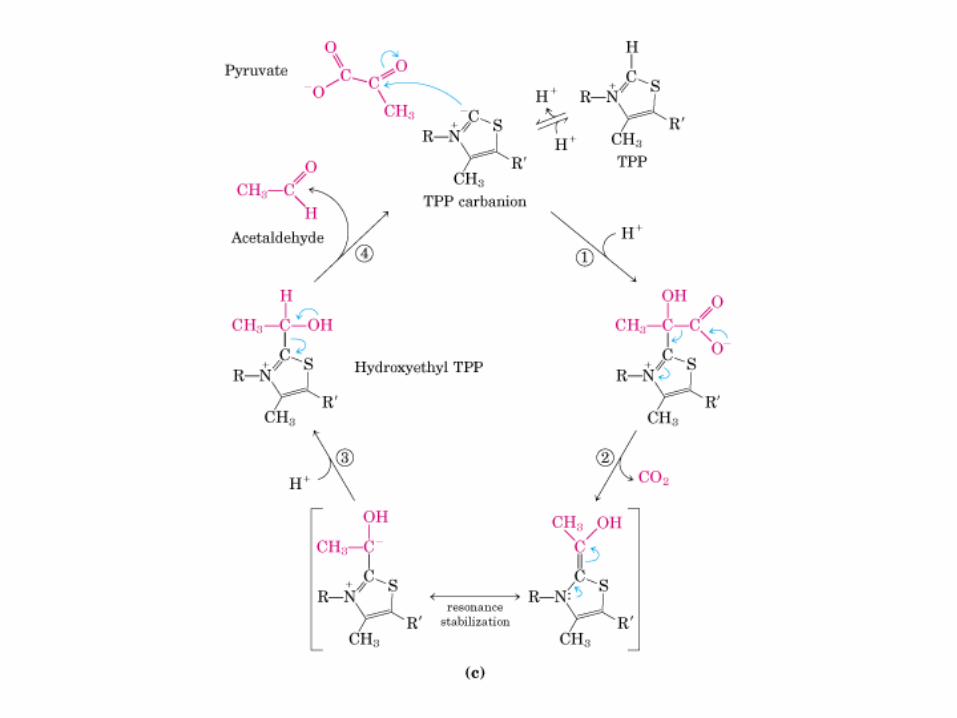
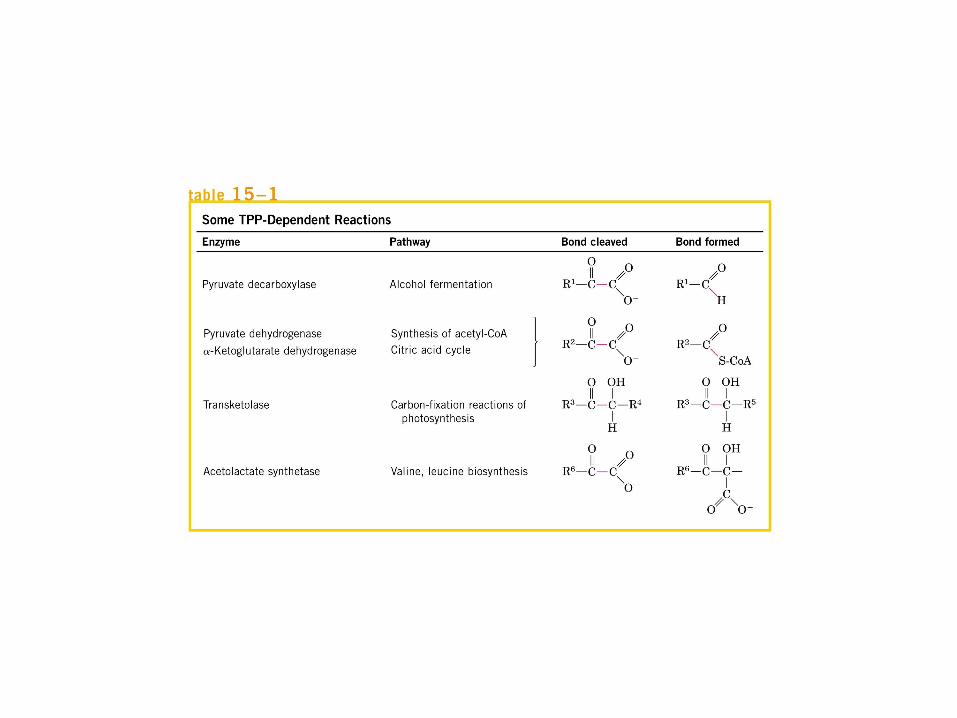

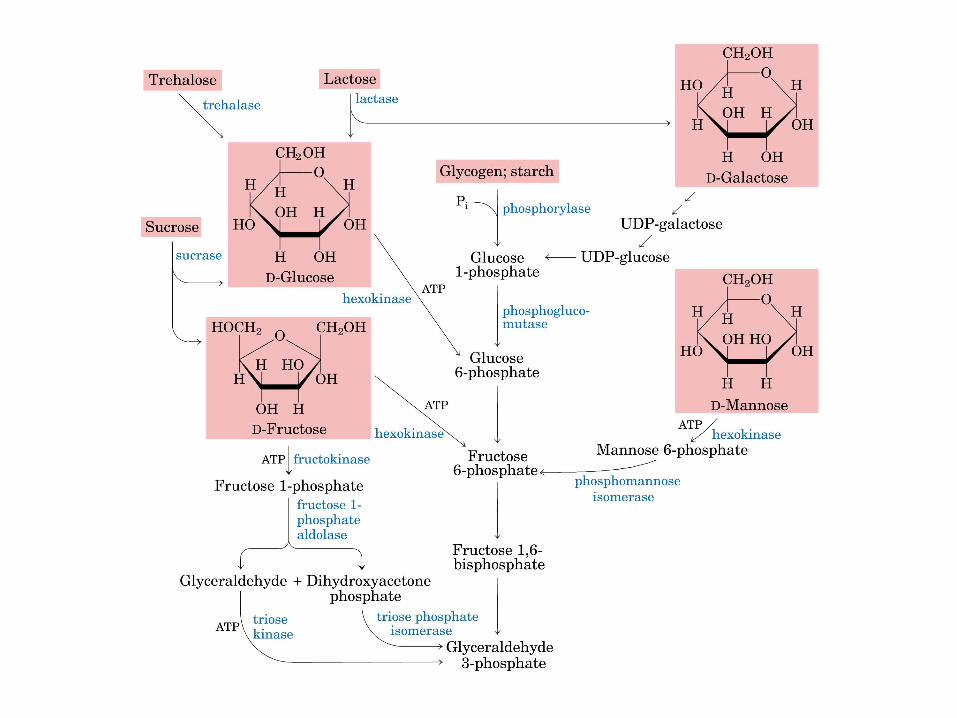
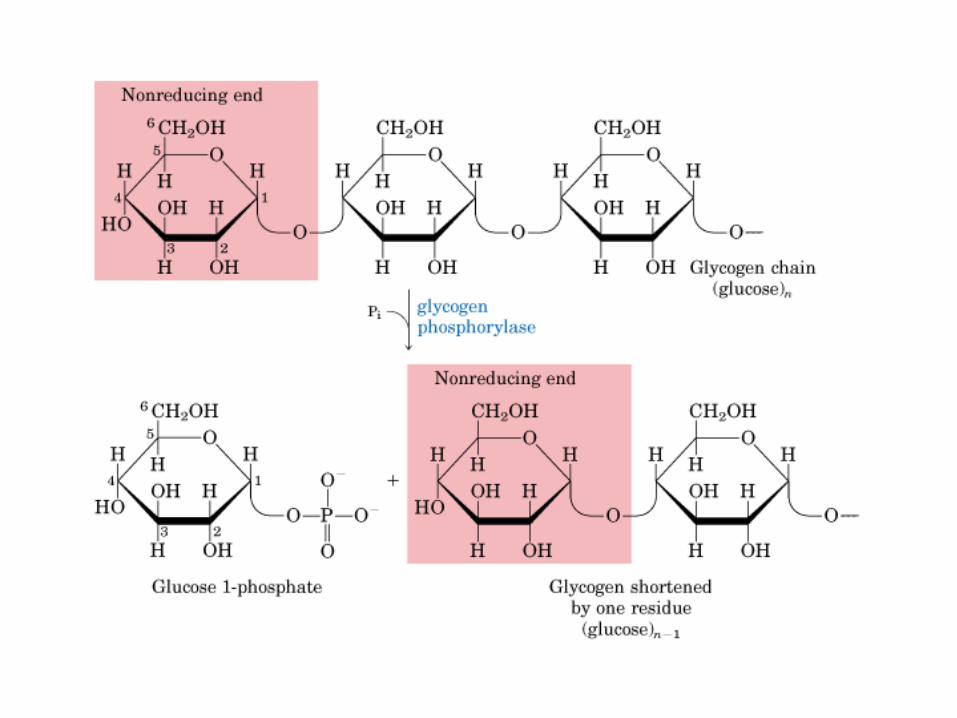

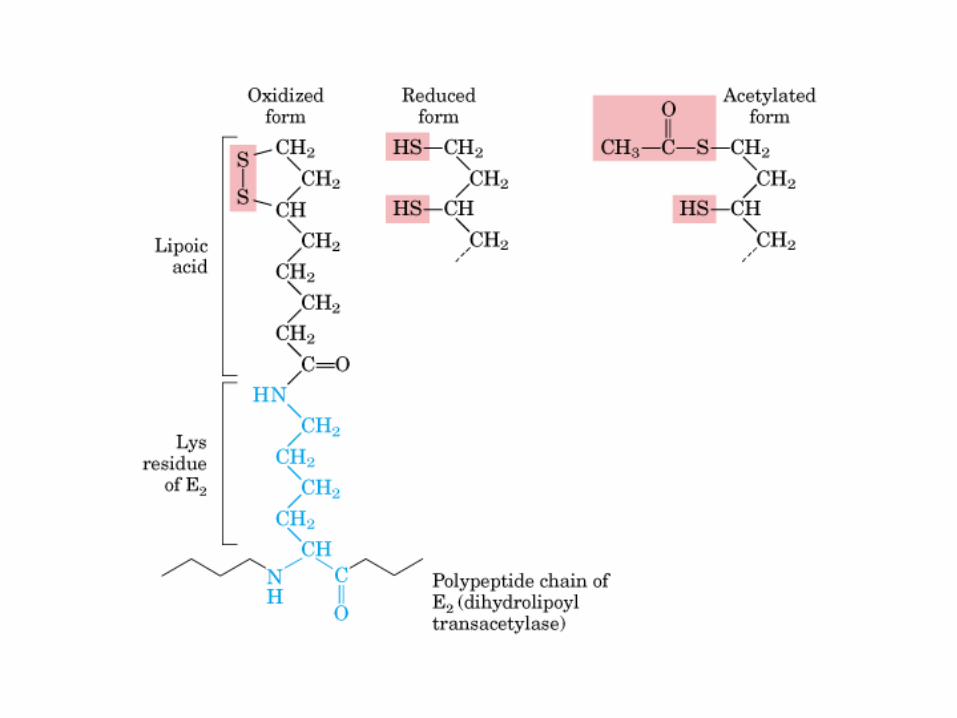
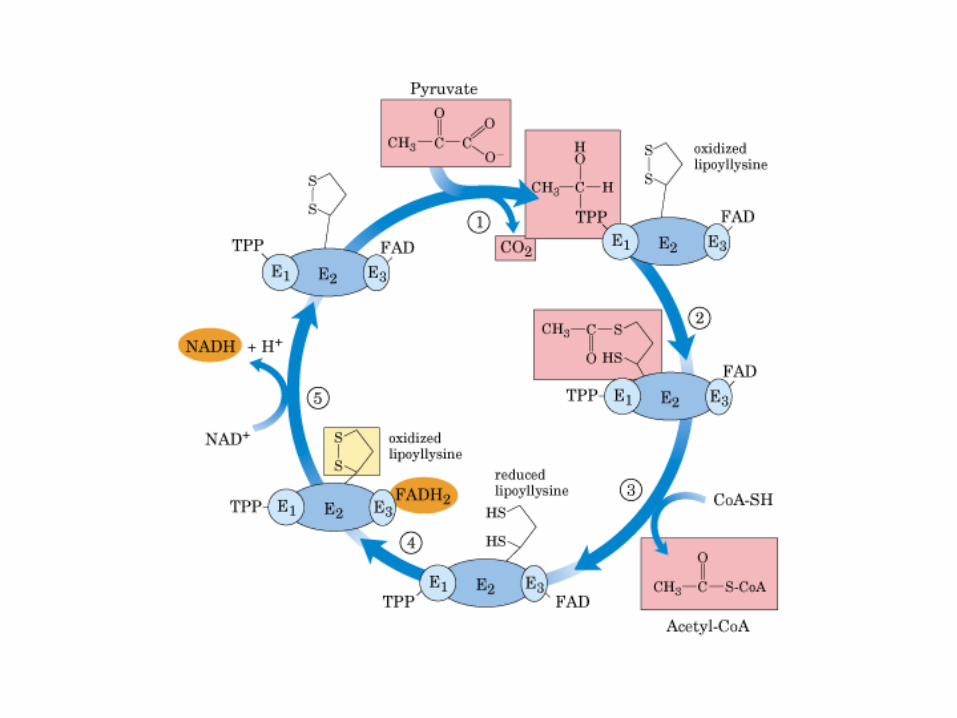
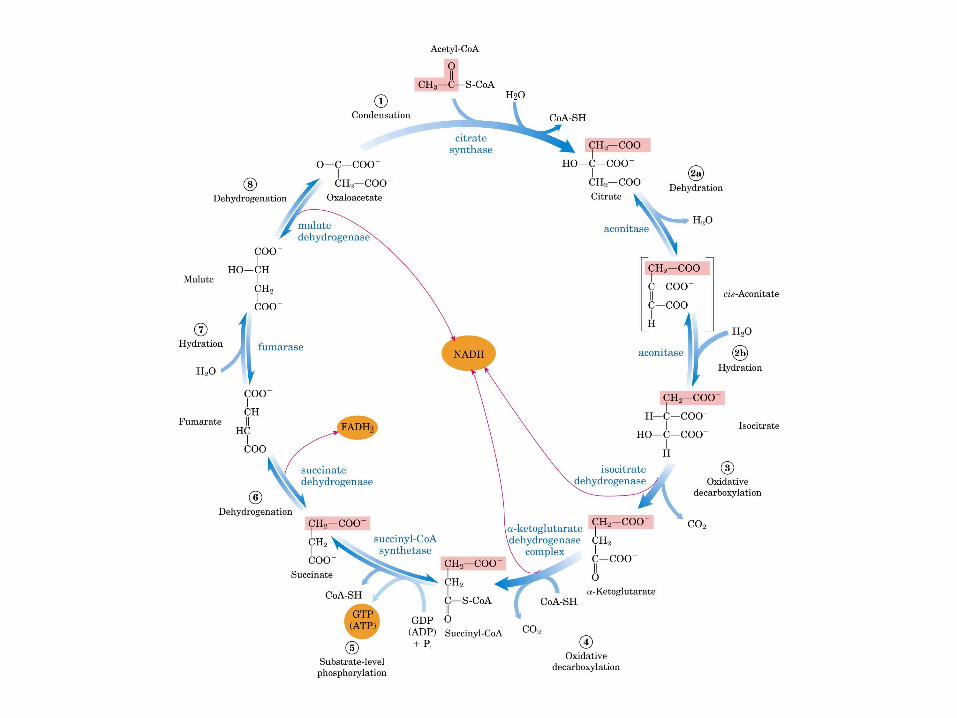
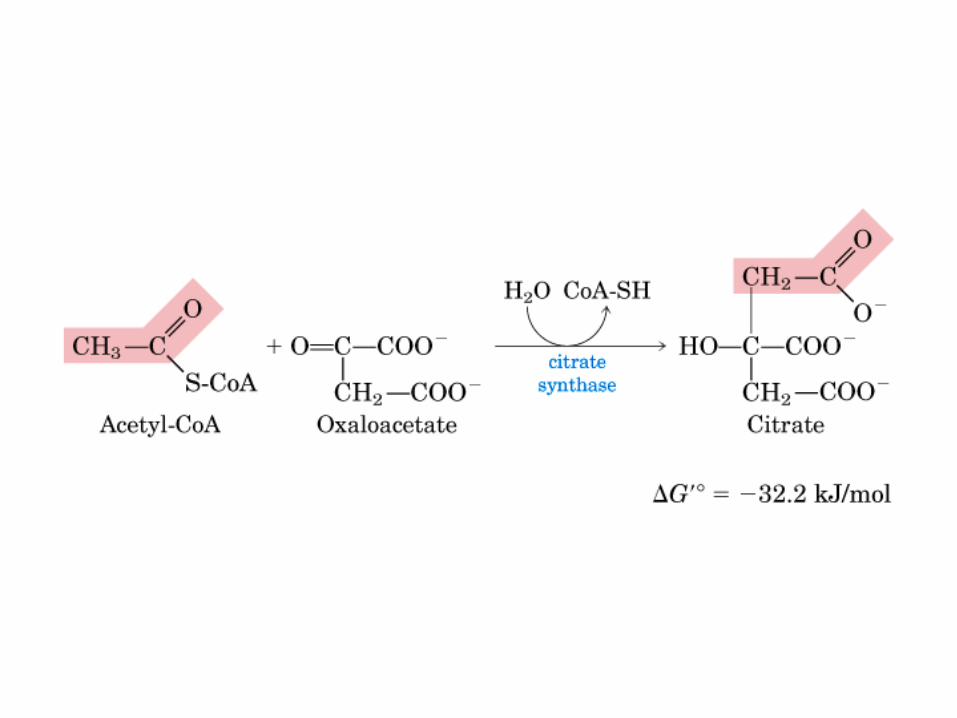
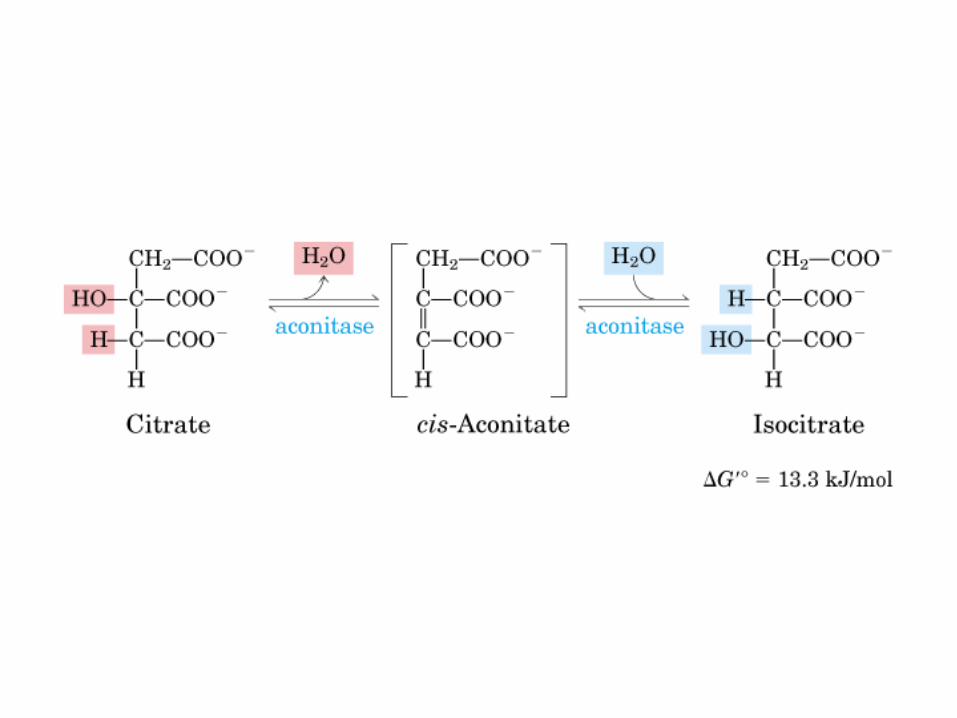
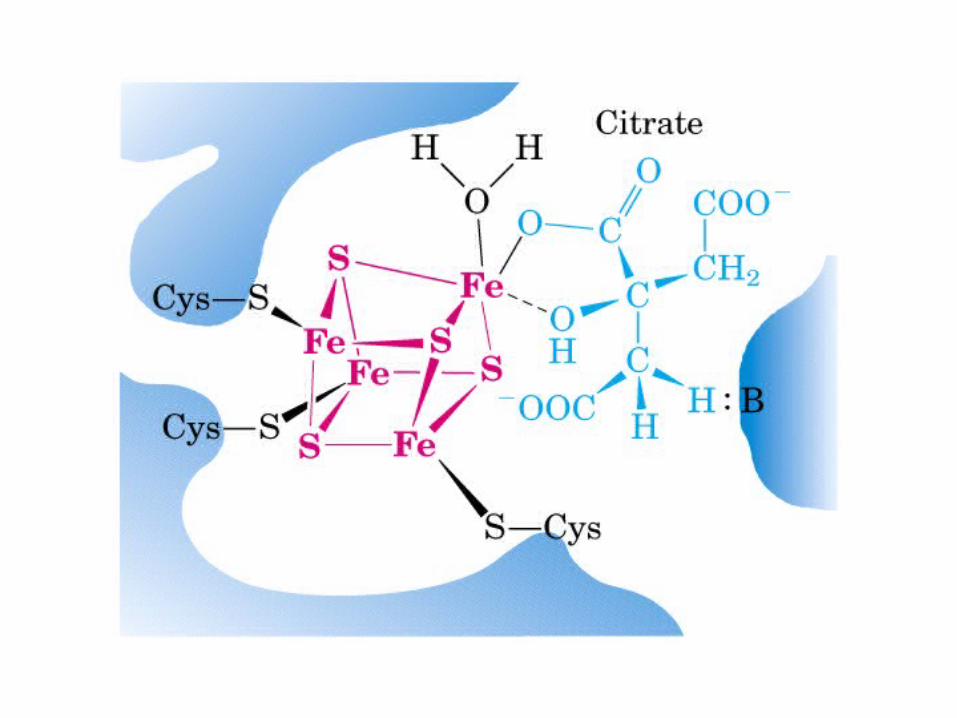
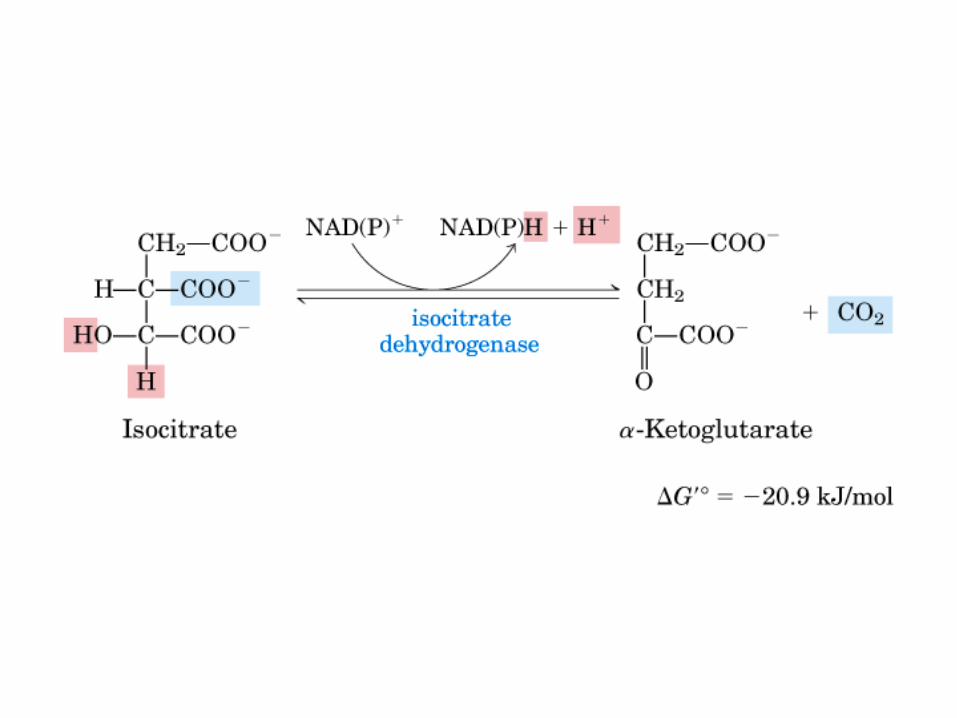
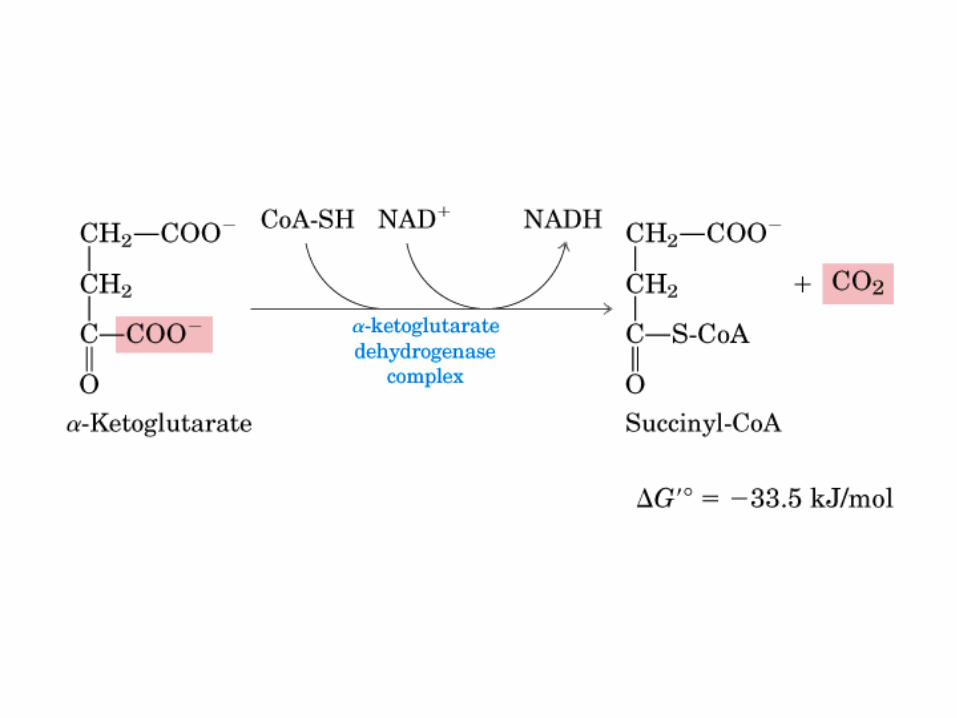
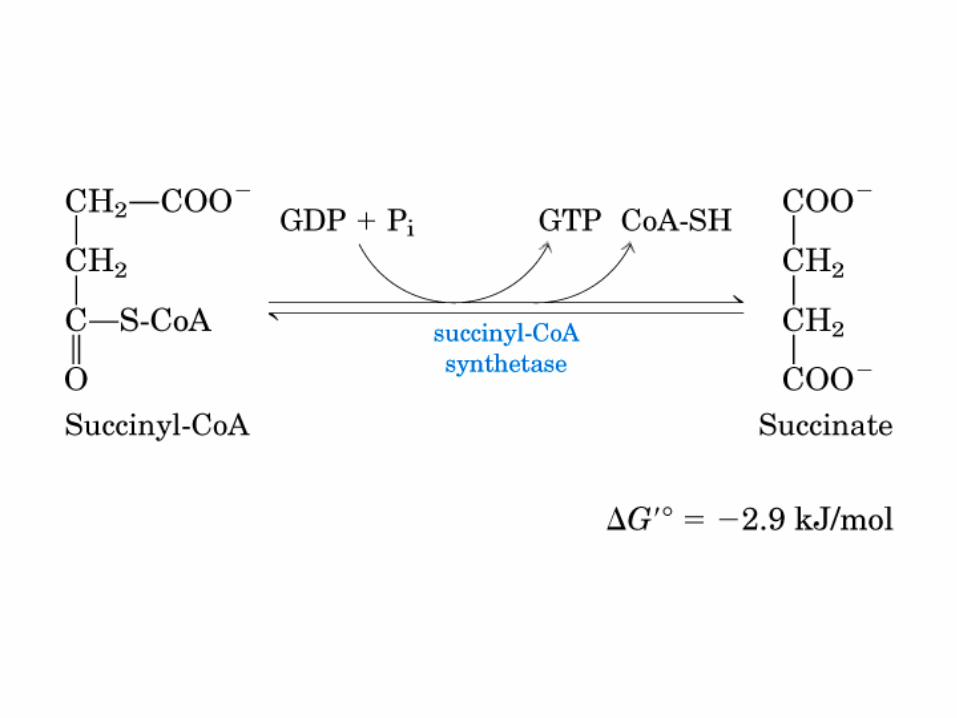
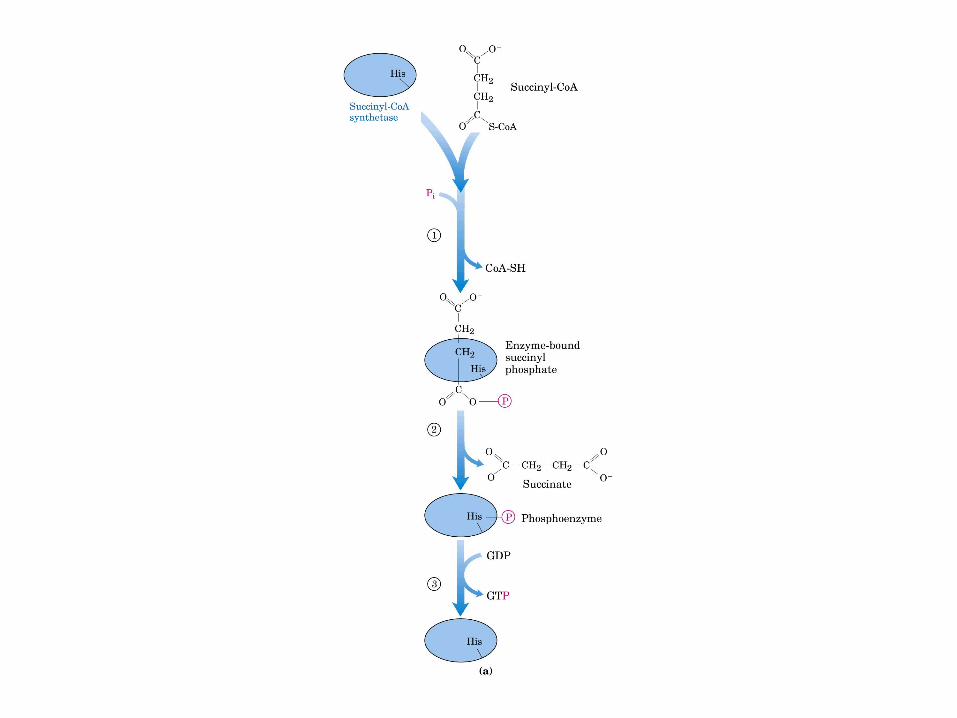
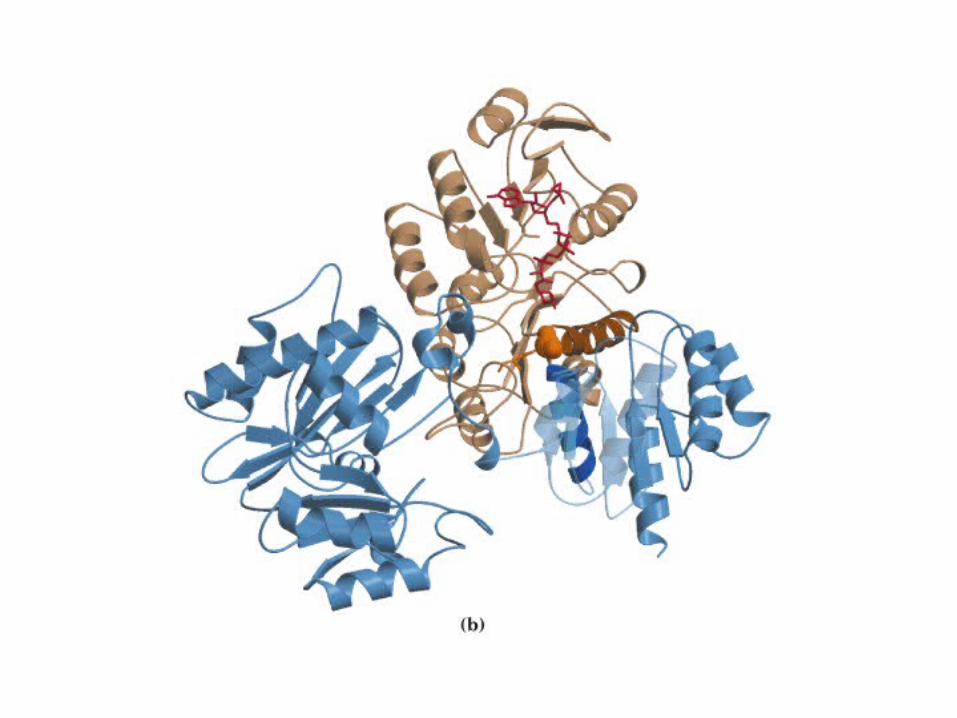
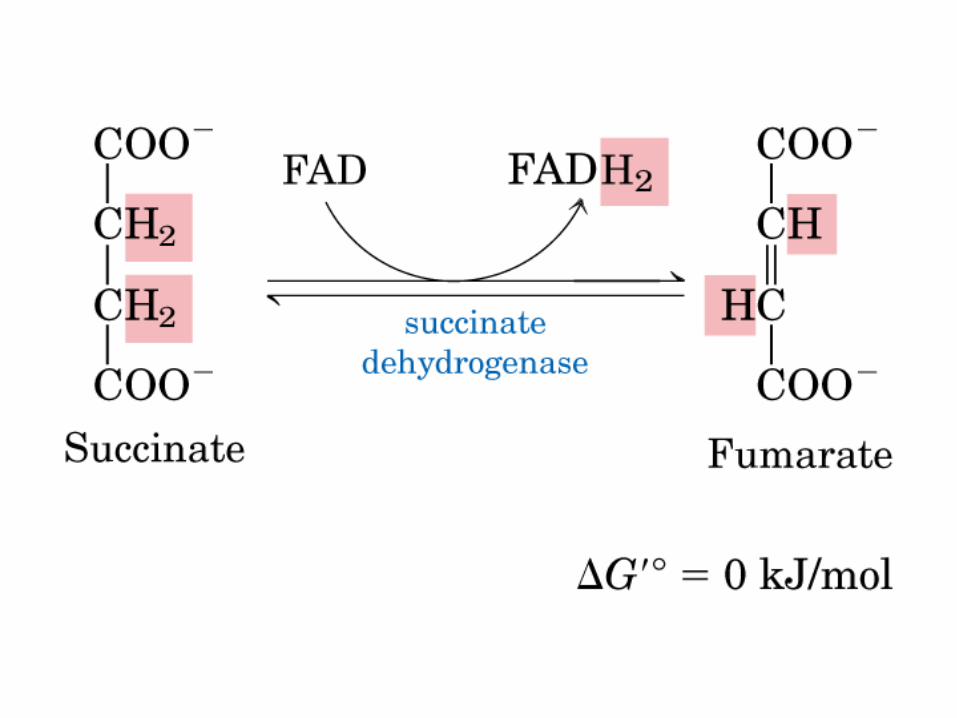
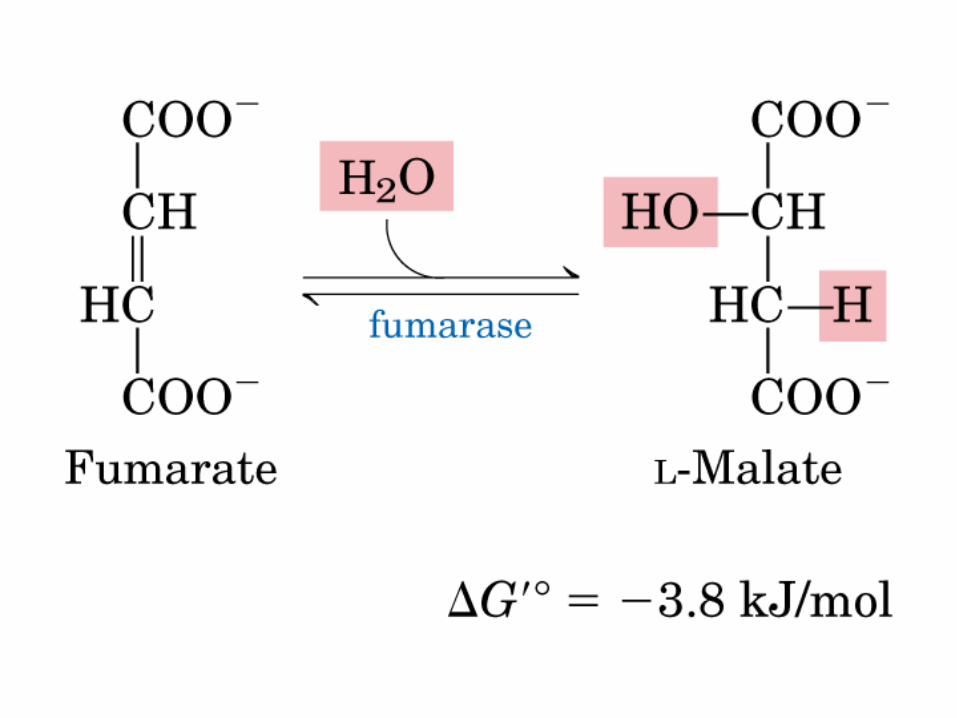
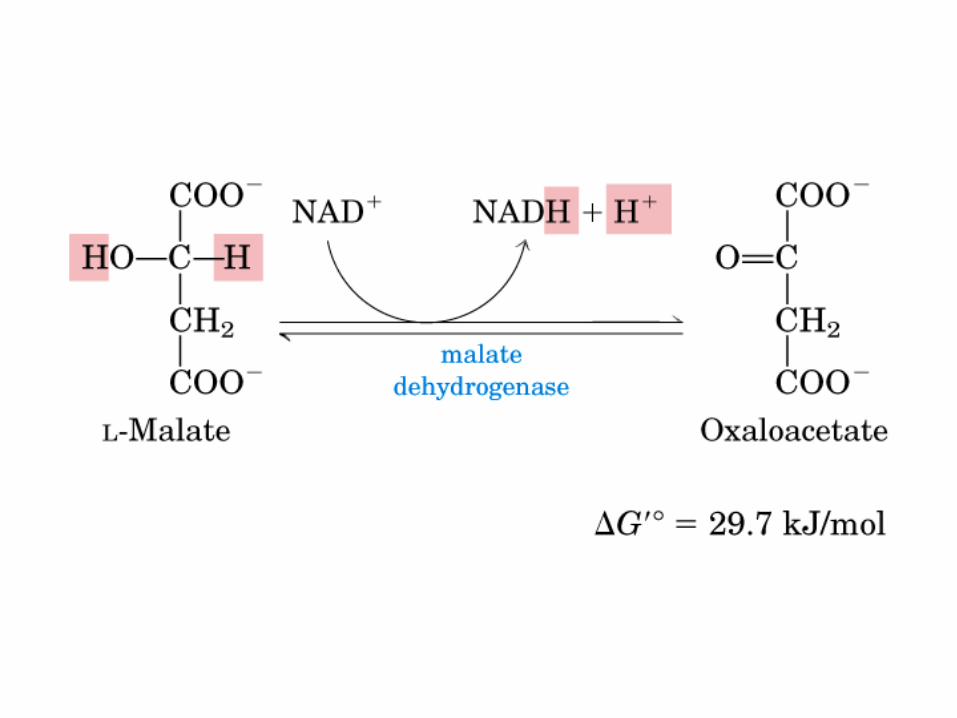
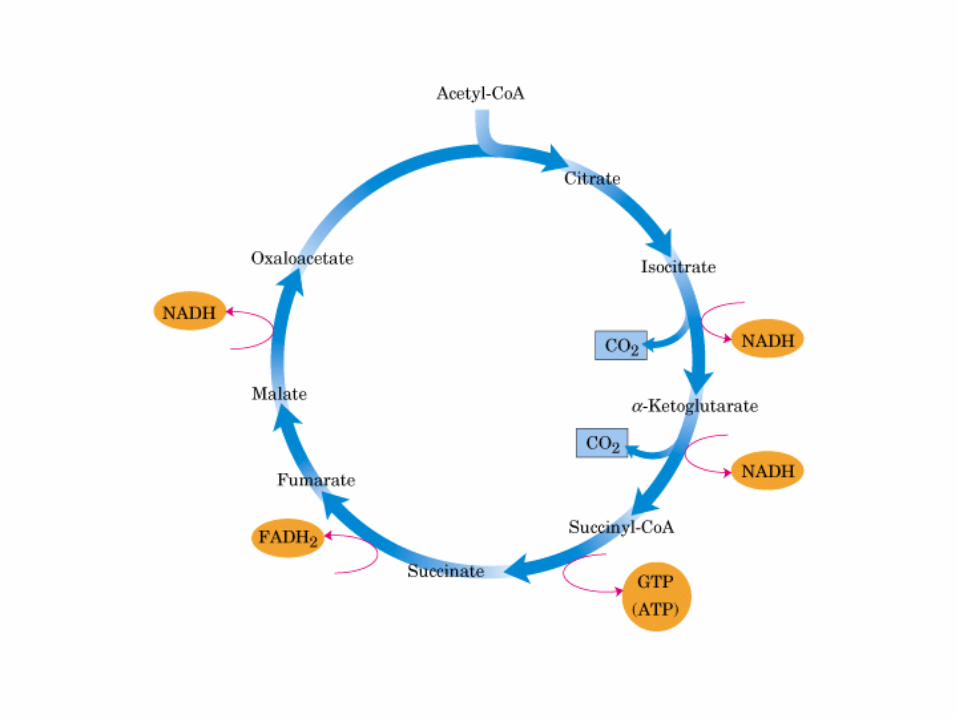
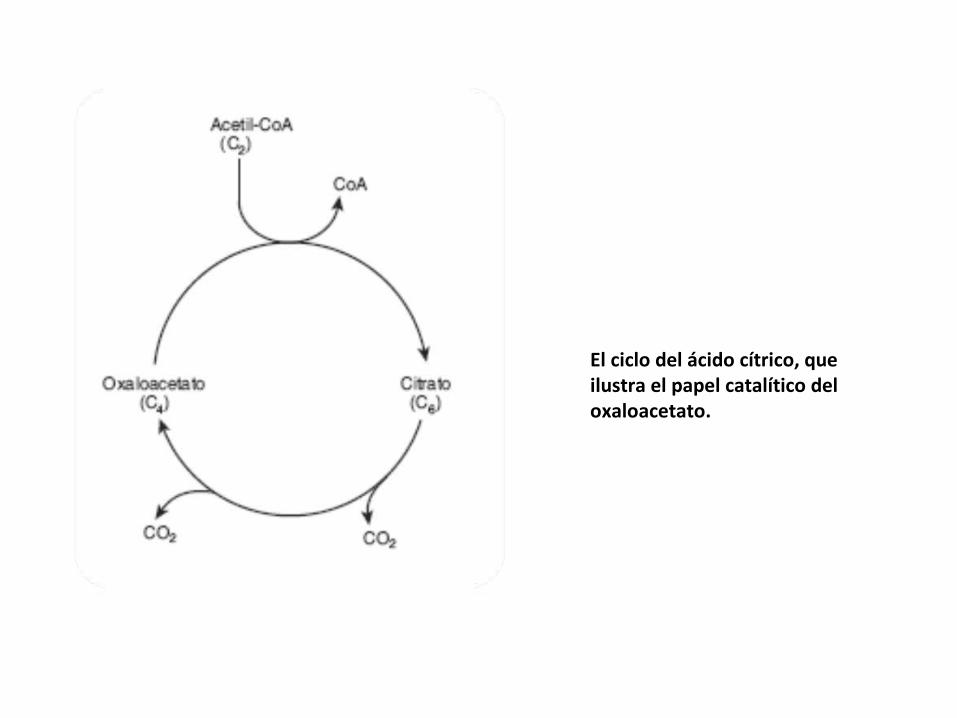
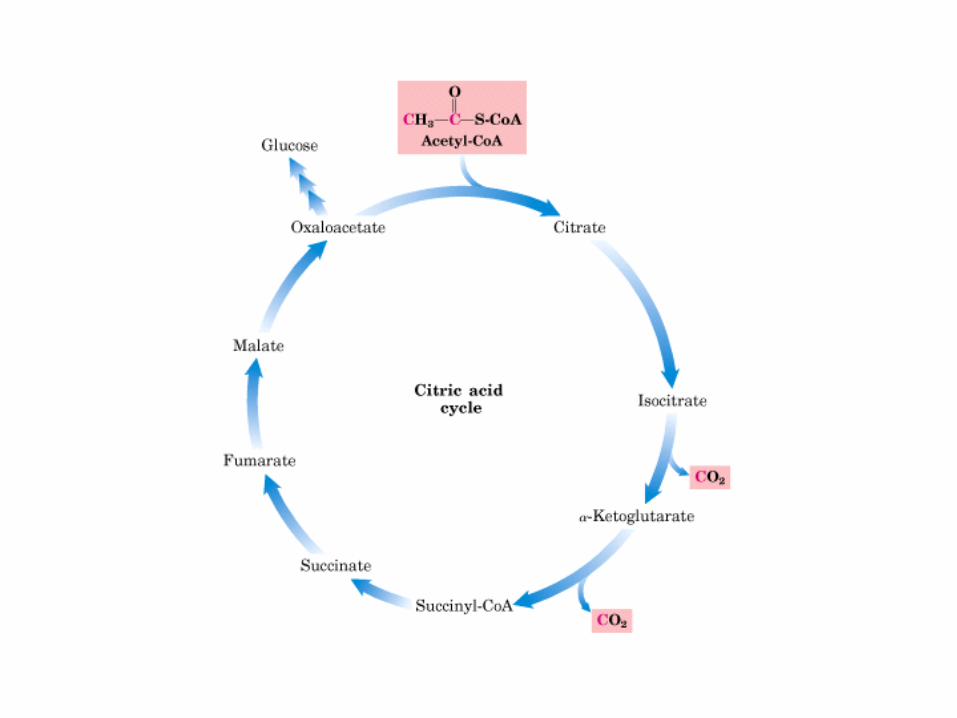
El ciclo del ácido cítrico, que ilustra el papel catalítico del oxaloacetato.
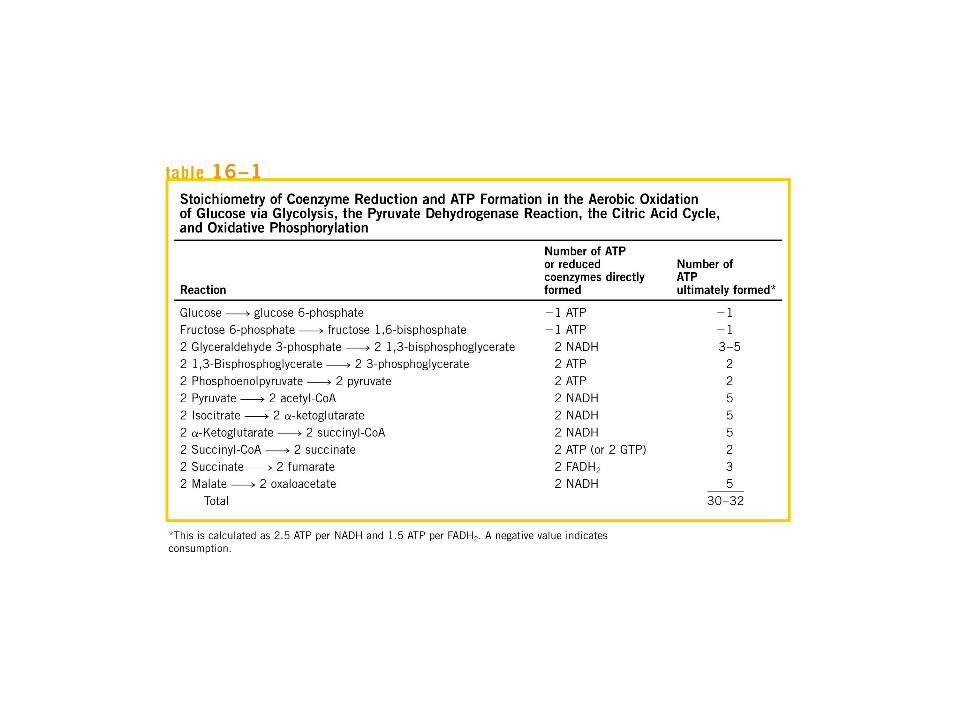
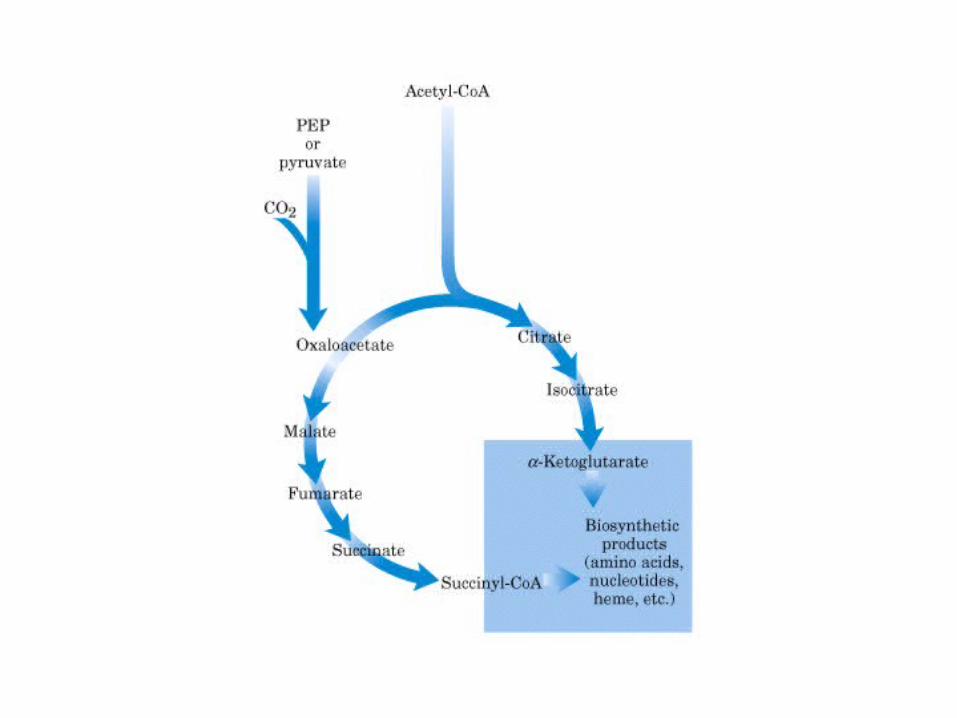

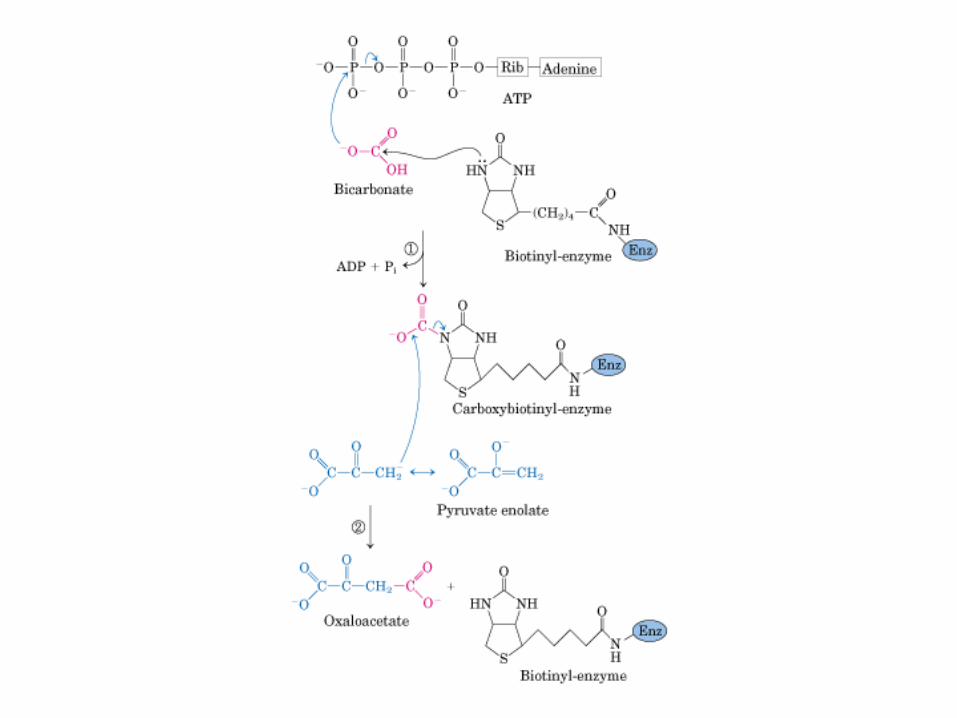
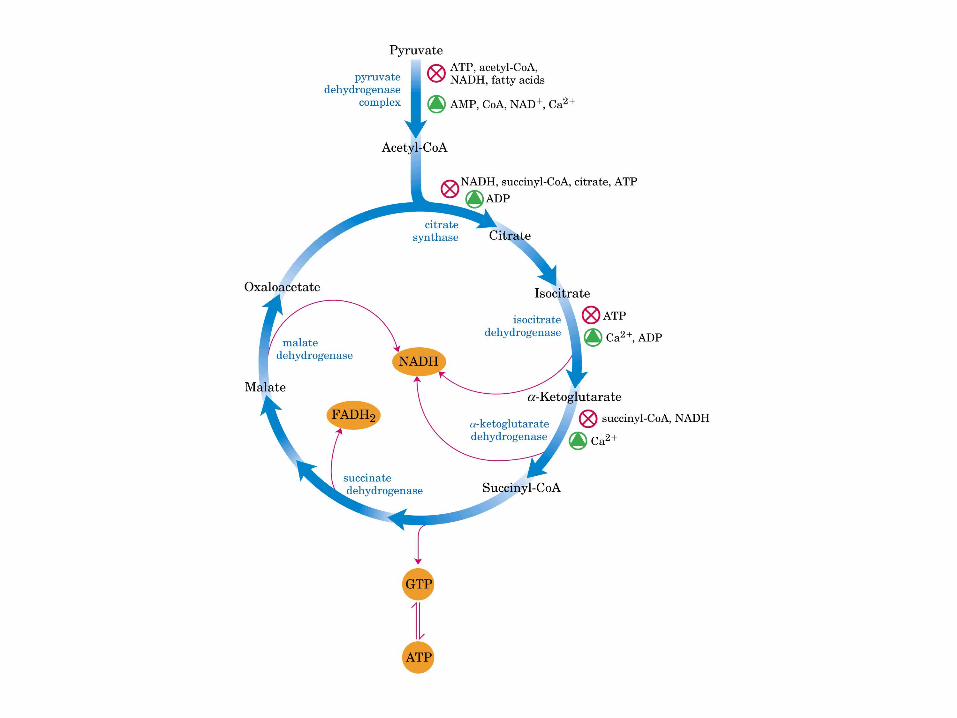
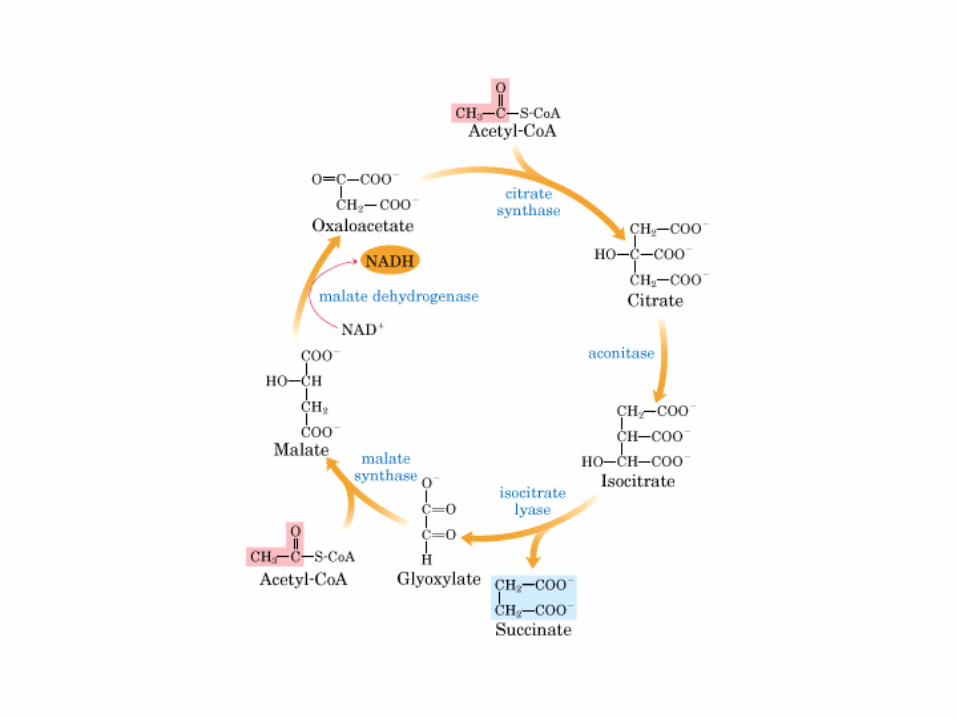
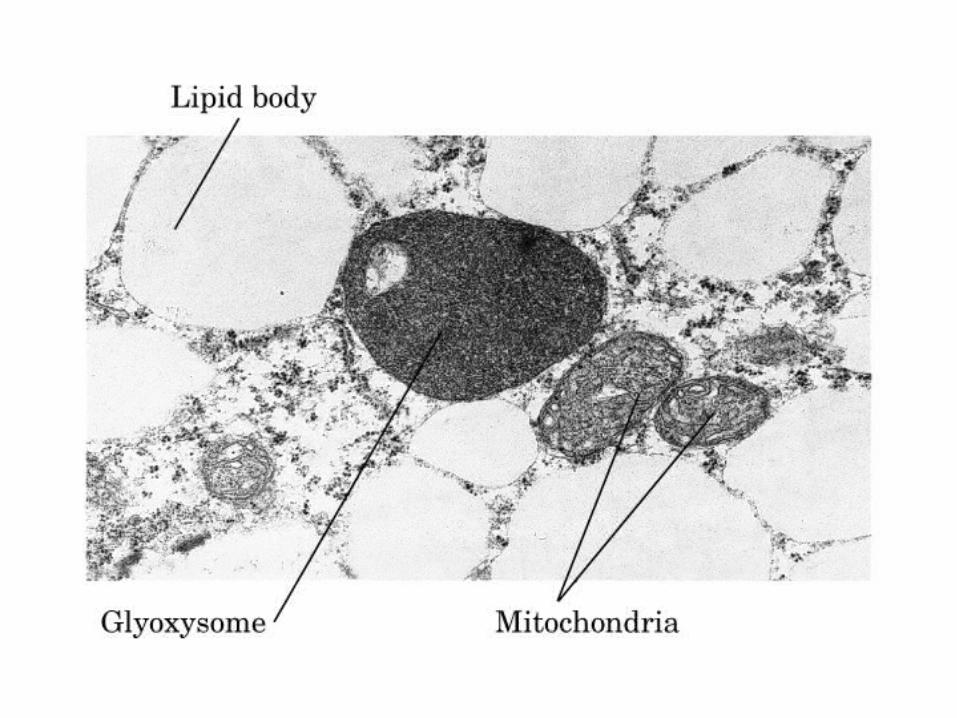
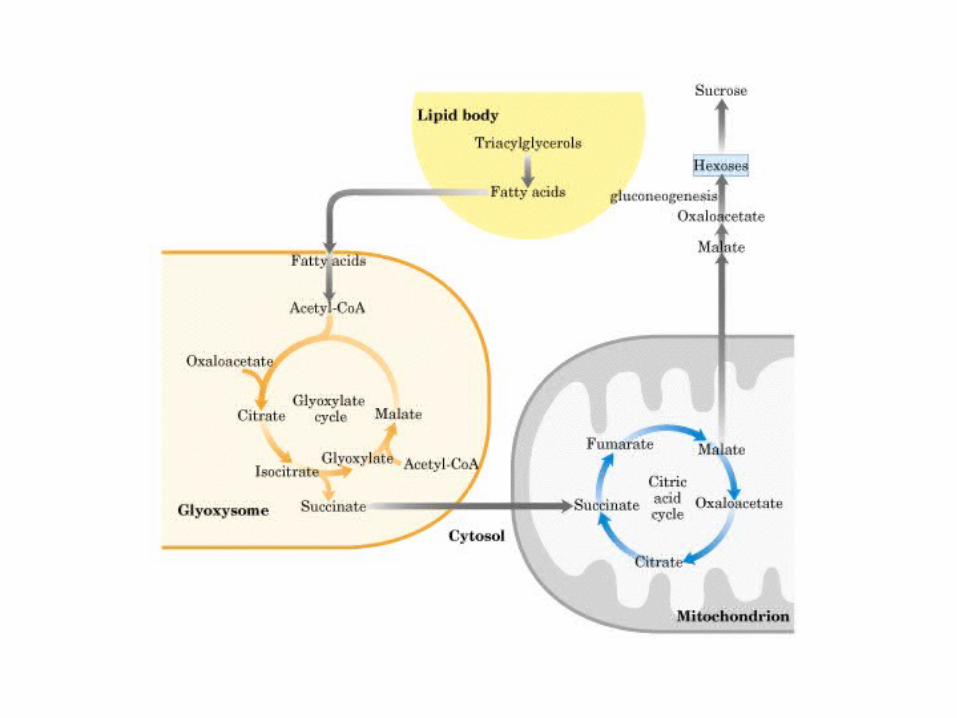
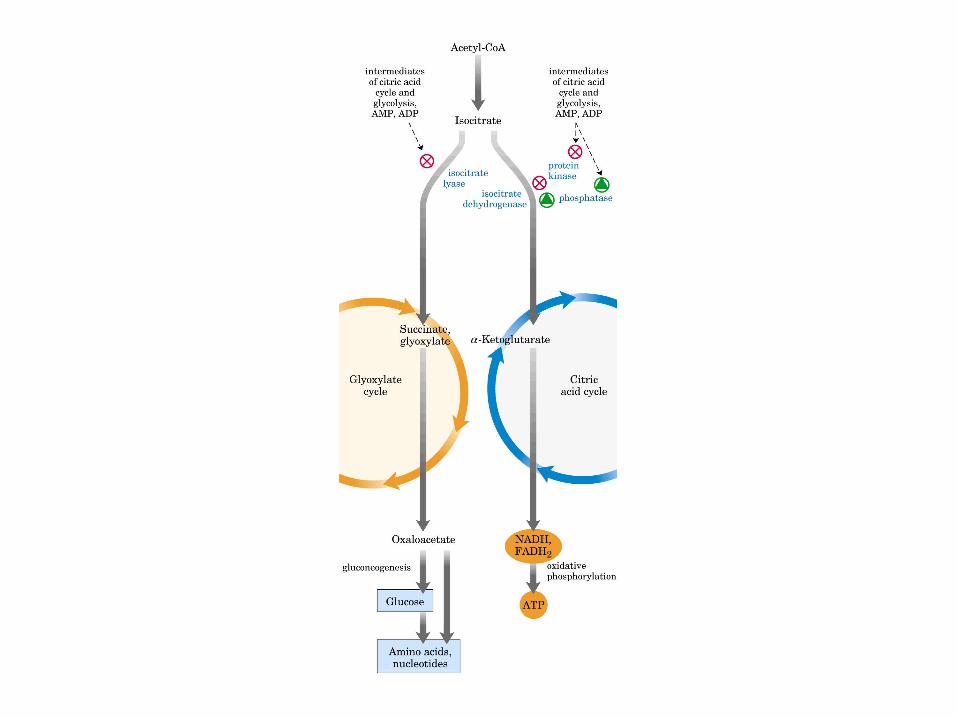
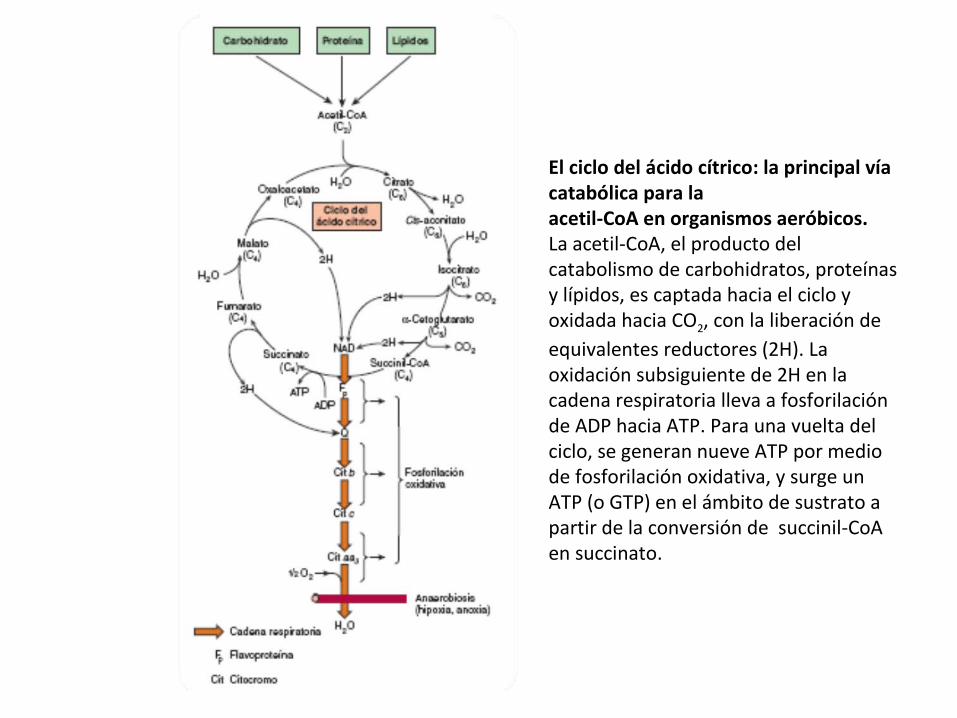
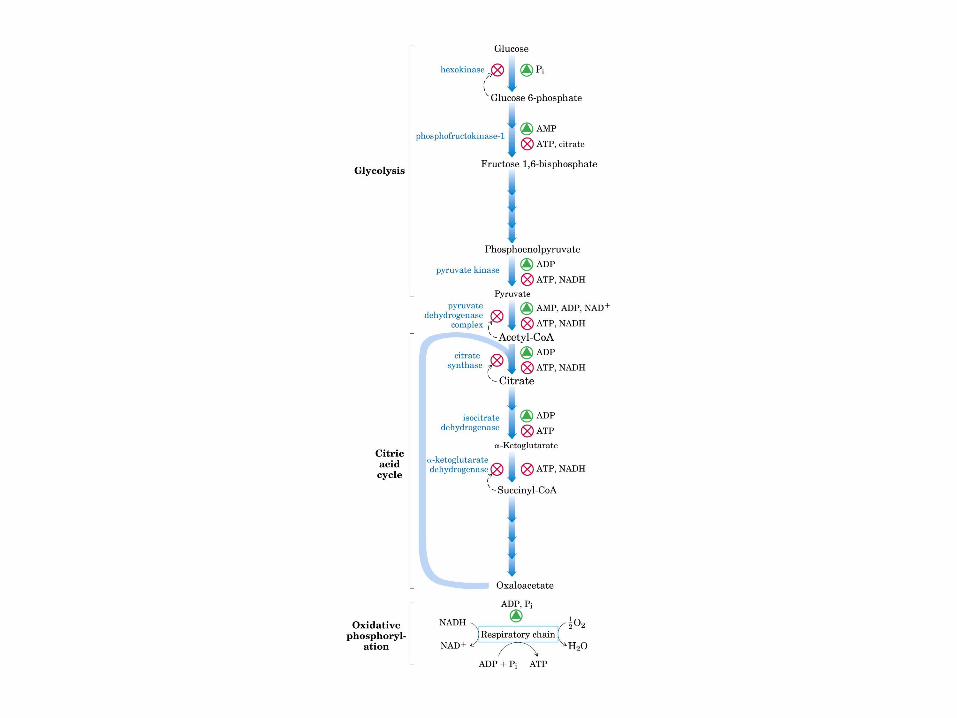
El ciclo del ácido cítrico: la principal vía catabólica para la acetil-CoA en organismos aeróbicos. La acetil-CoA, el producto del catabolismo de carbohidratos, proteínas y lípidos, es captada hacia el ciclo y oxidada hacia CO2, con la liberación de equivalentes reductores (2H). La oxidación subsiguiente de 2H en la cadena respiratoria lleva a fosforilación de ADP hacia ATP. Para una vuelta del ciclo, se generan nueve ATP por medio de fosforilación oxidativa, y surge un ATP (o GTP) en el ámbito de sustrato a partir de la conversión de succinil-CoA en succinato.
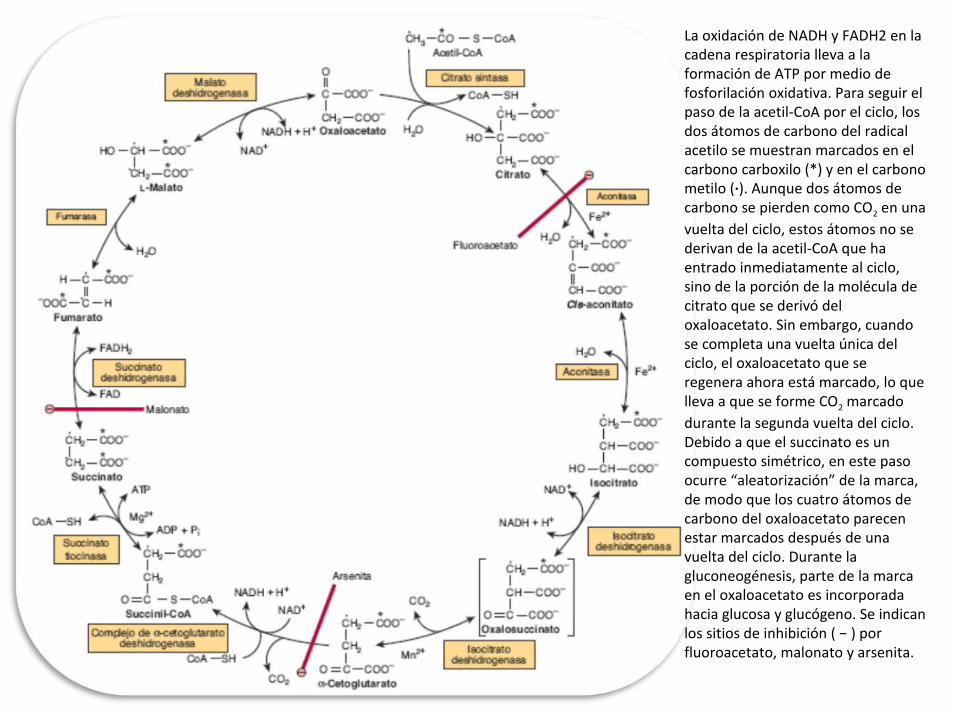
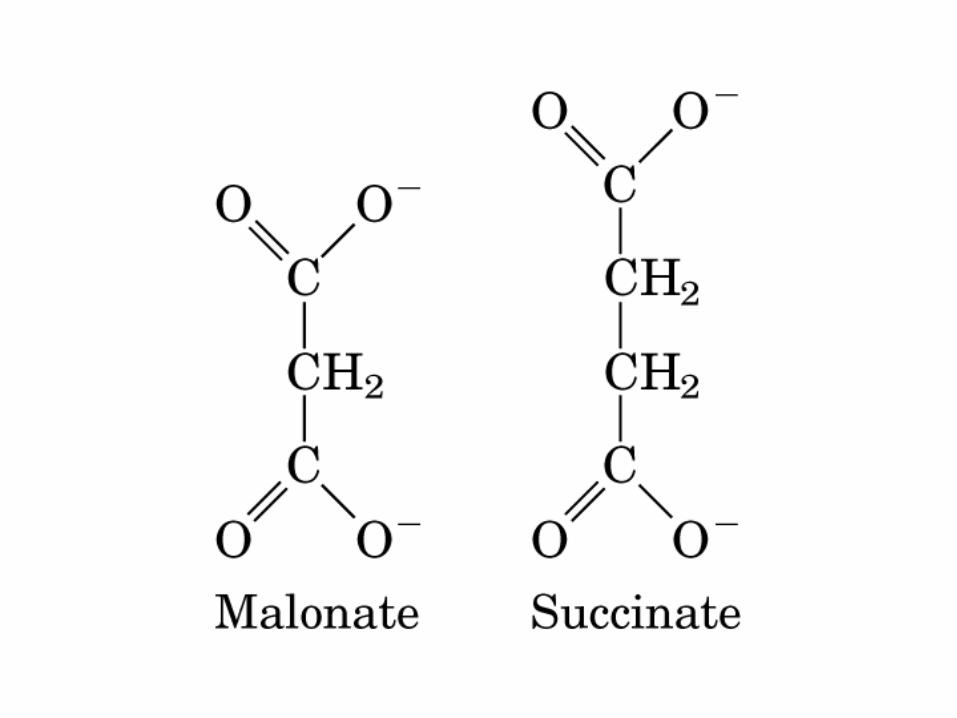
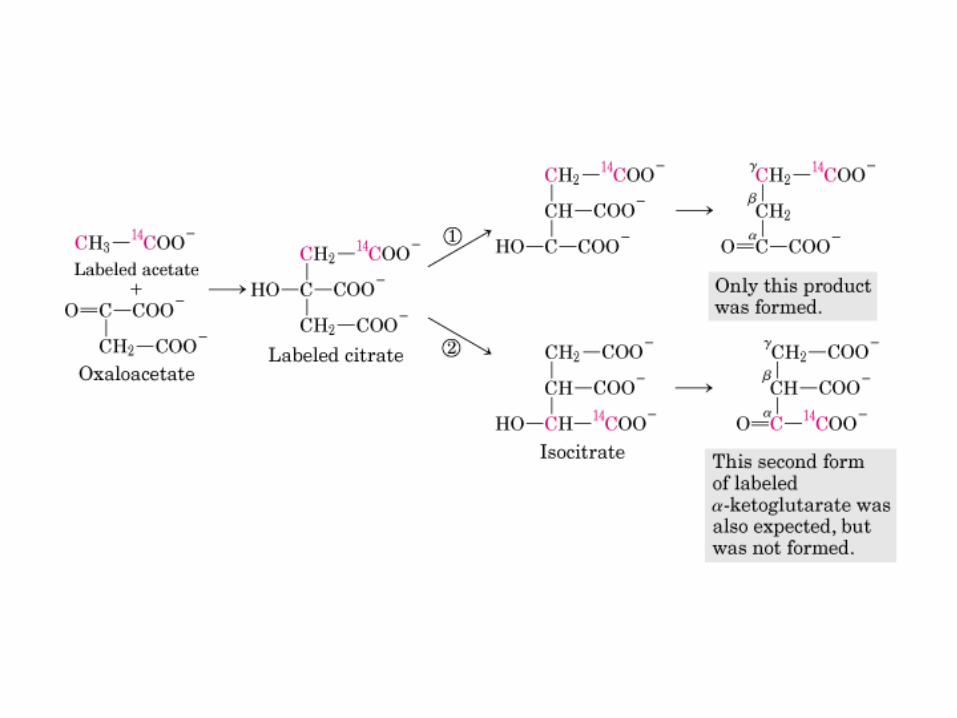
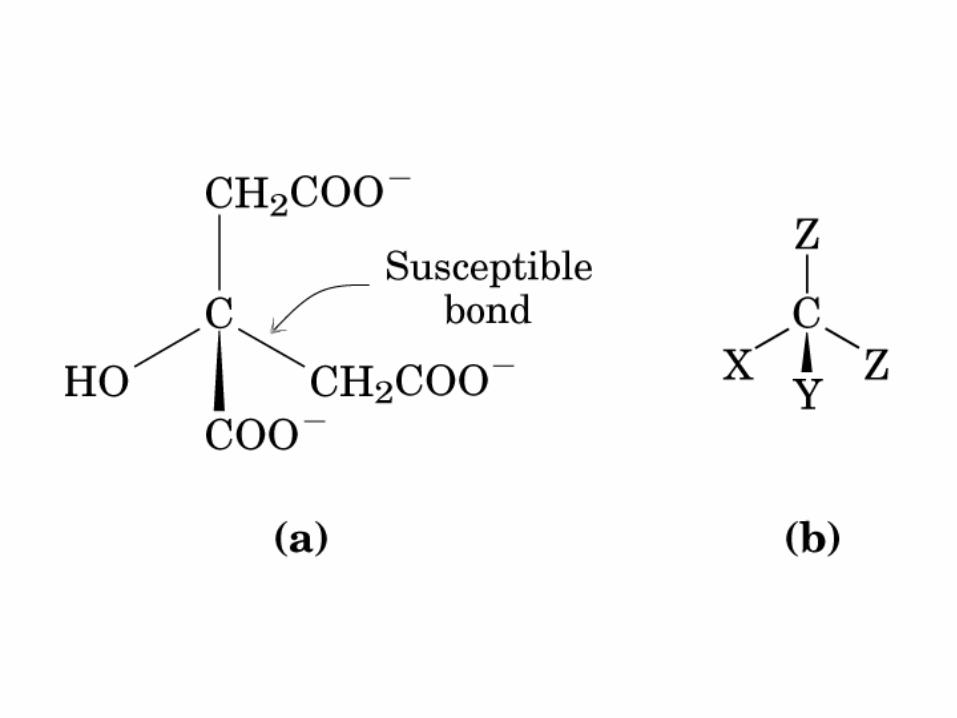
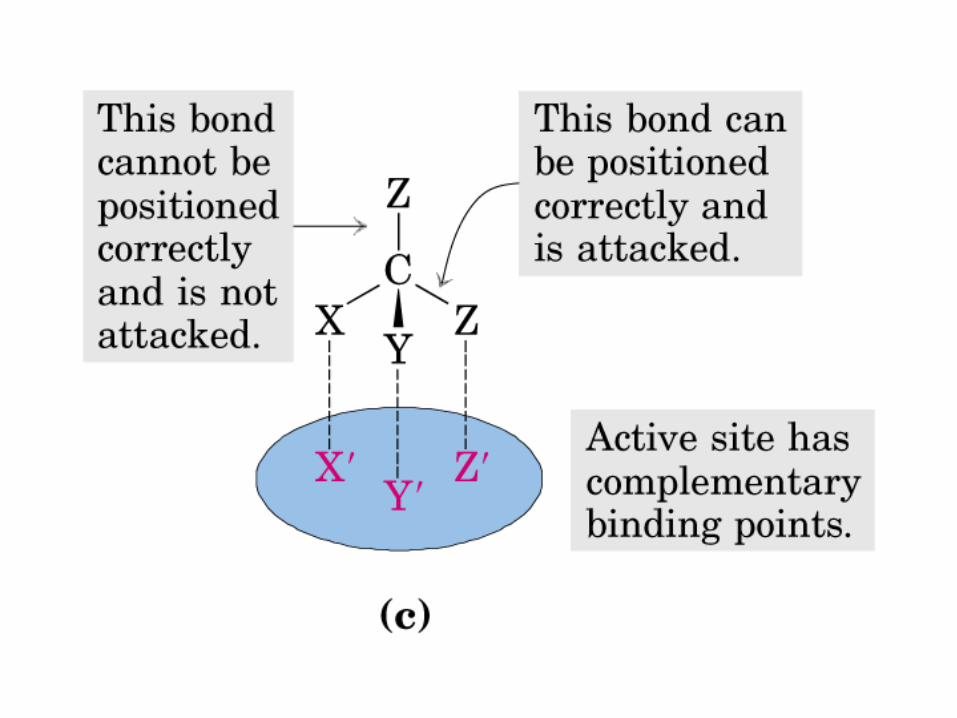
La oxidación de NADH y FADH2 en la cadena respiratoria lleva a la formación de ATP por medio de fosforilación oxidativa. Para seguir el paso de la acetil-CoA por el ciclo, los dos átomos de carbono del radical acetilo se muestran marcados en el carbono carboxilo (*) y en el carbono metilo (·). Aunque dos átomos de carbono se pierden como CO2 en una vuelta del ciclo, estos átomos no se derivan de la acetil-CoA que ha entrado inmediatamente al ciclo, sino de la porción de la molécula de citrato que se derivó del oxaloacetato. Sin embargo, cuando se completa una vuelta única del ciclo, el oxaloacetato que se regenera ahora está marcado, lo que lleva a que se forme CO2 marcado durante la segunda vuelta del ciclo. Debido a que el succinato es un compuesto simétrico, en este paso ocurre “aleatorización” de la marca, de modo que los cuatro átomos de carbono del oxaloacetato parecen estar marcados después de una vuelta del ciclo. Durante la gluconeogénesis, parte de la marca en el oxaloacetato es incorporada hacia glucosa y glucógeno. Se indican los sitios de inhibición ( − ) por fluoroacetato, malonato y arsenita.
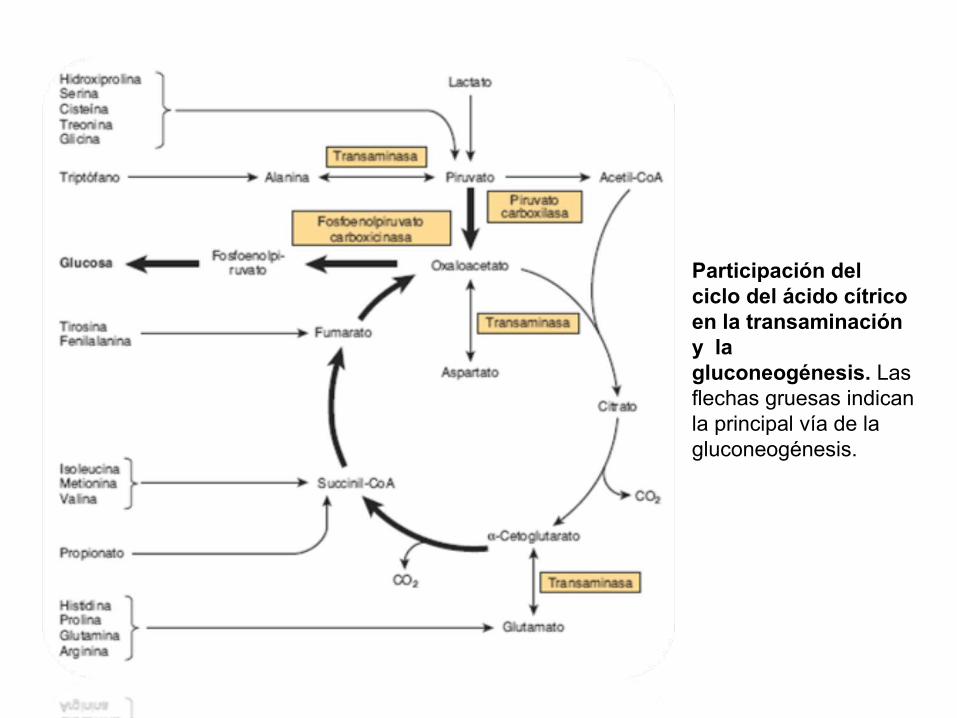
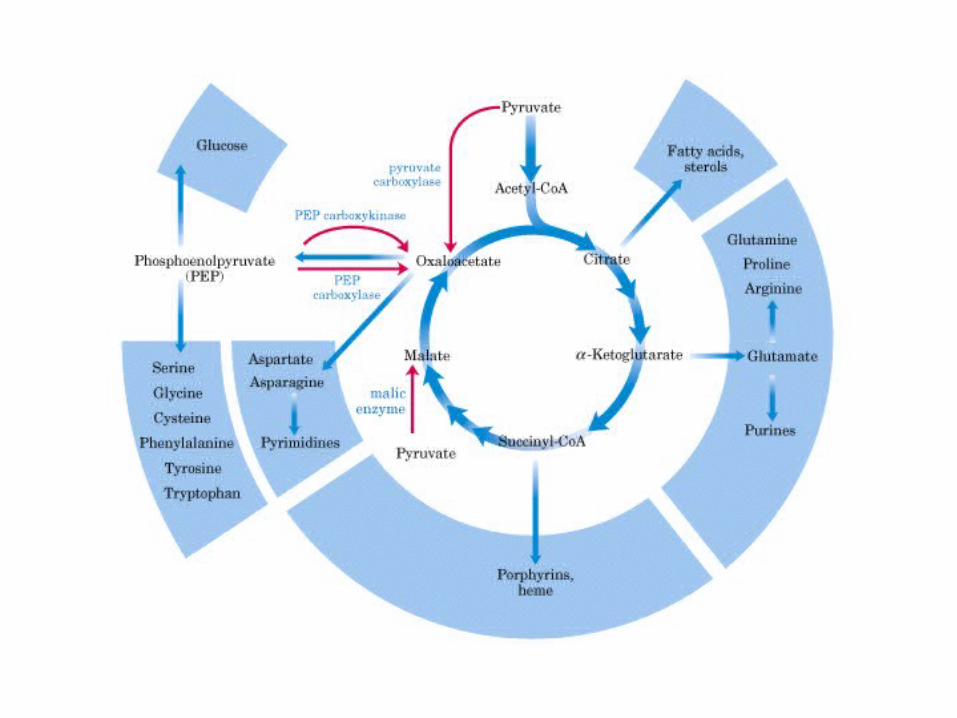
Participación del ciclo del ácido cítrico en la transaminación y la gluconeogénesis. Las flechas gruesas indican la principal vía de la gluconeogénesis.

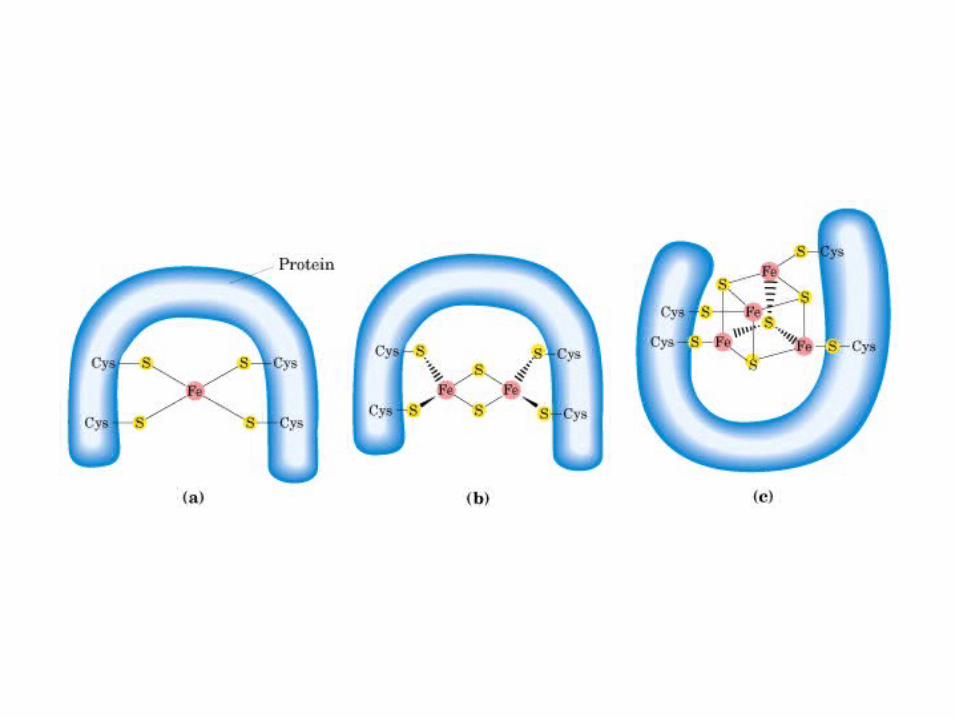
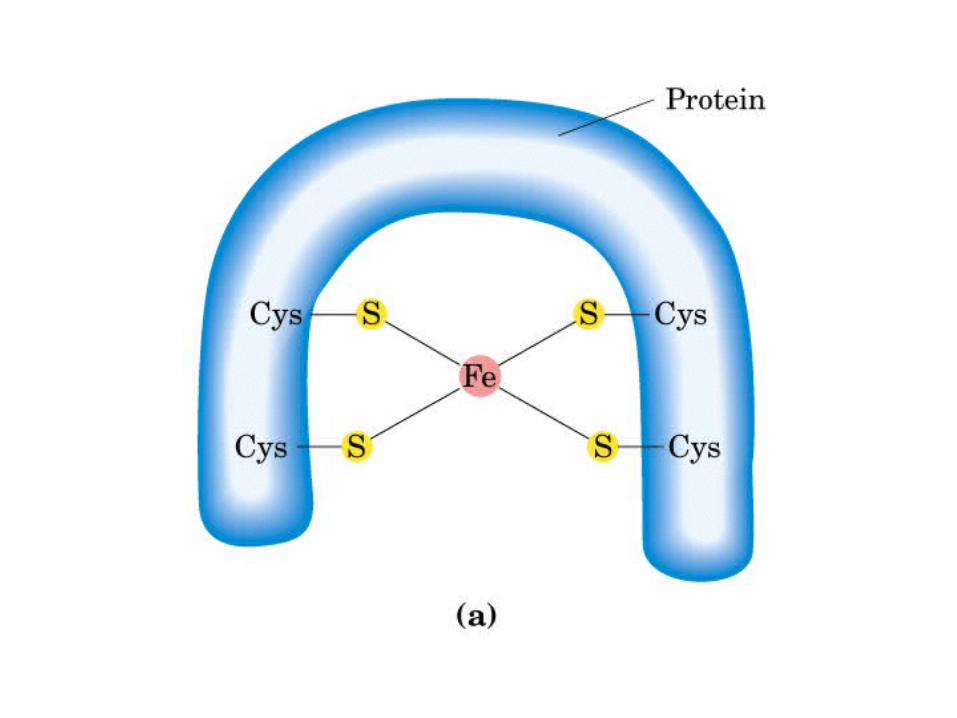
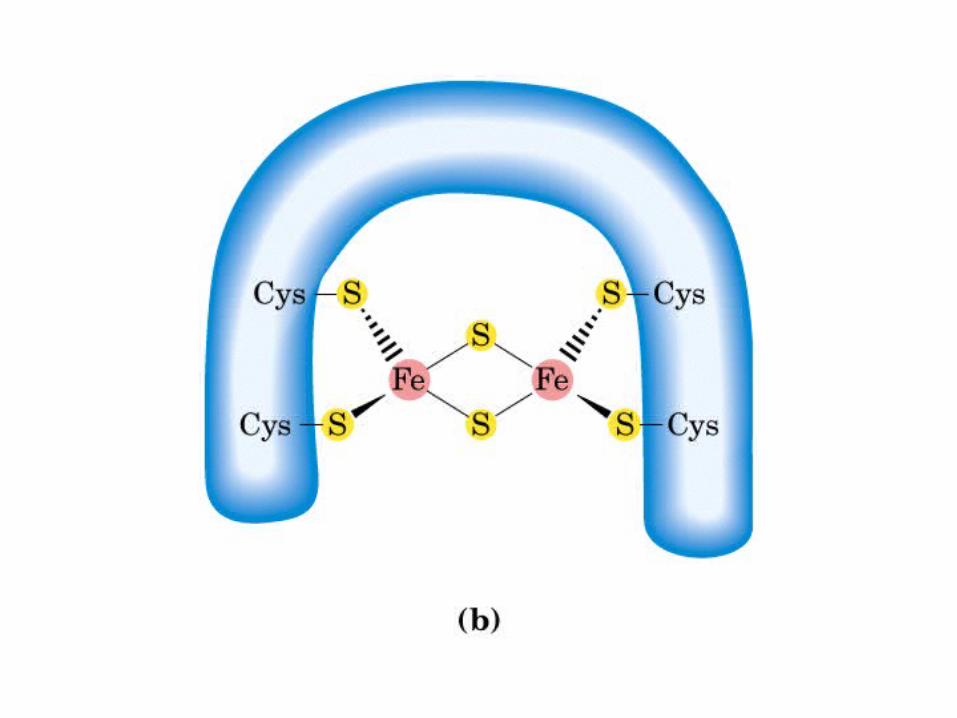
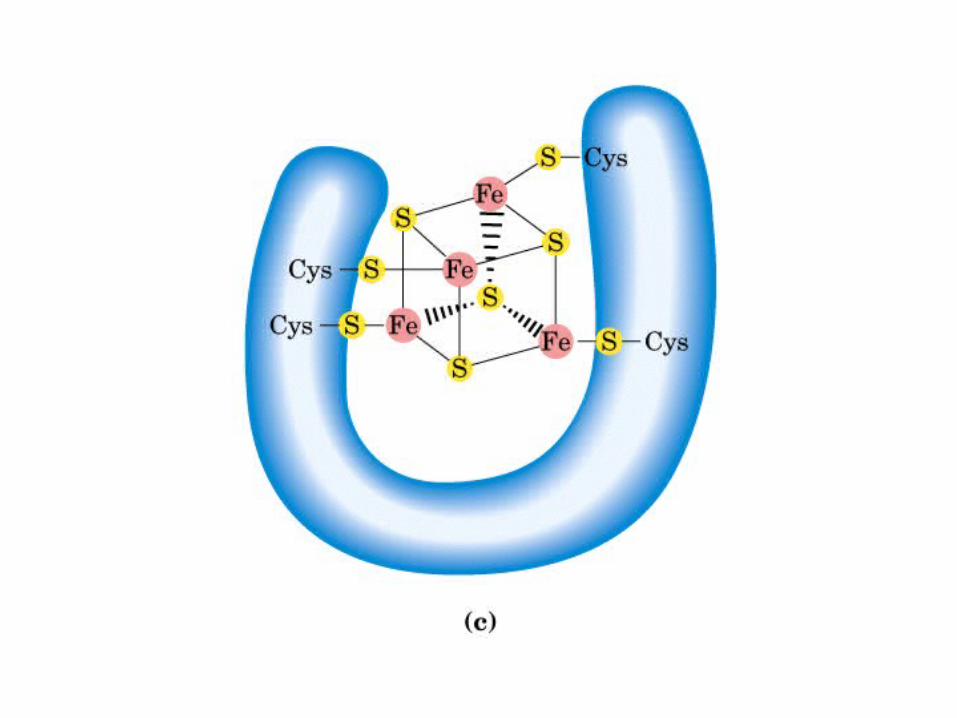

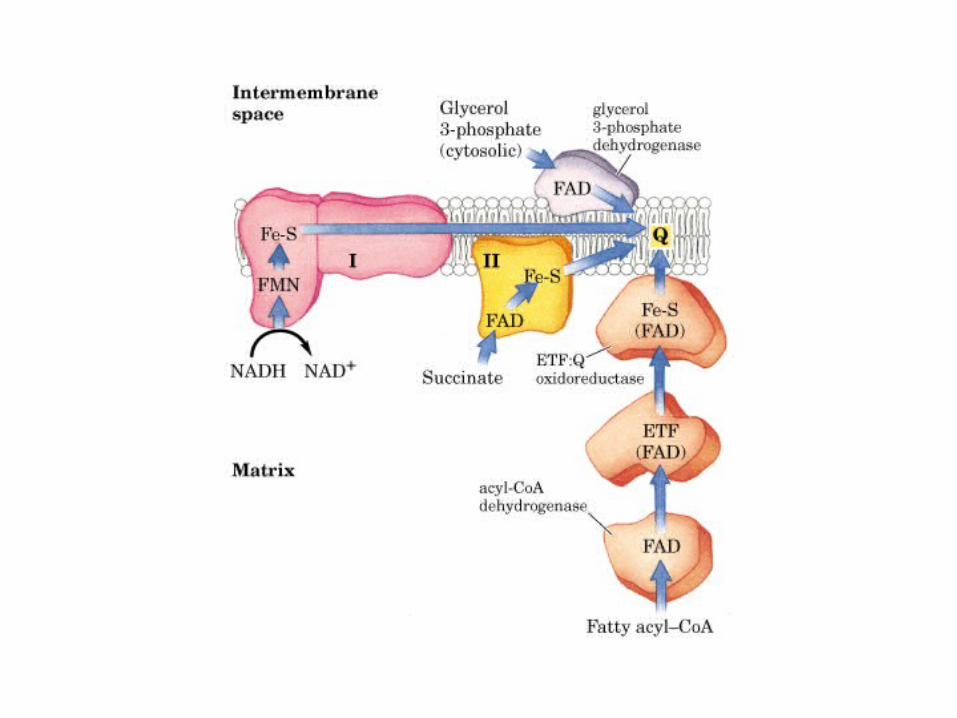
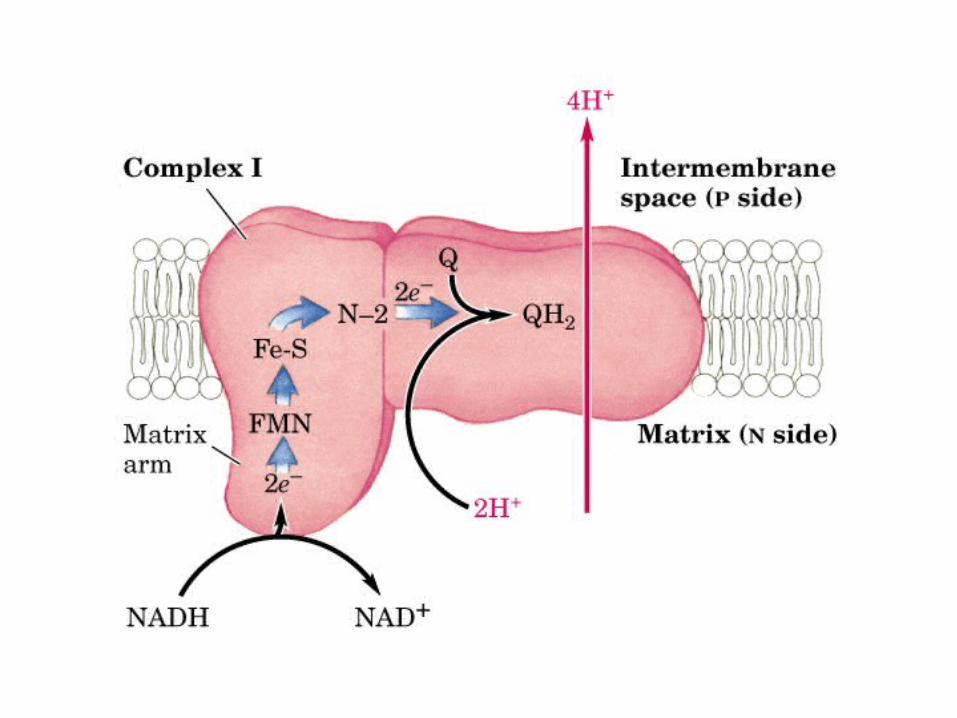
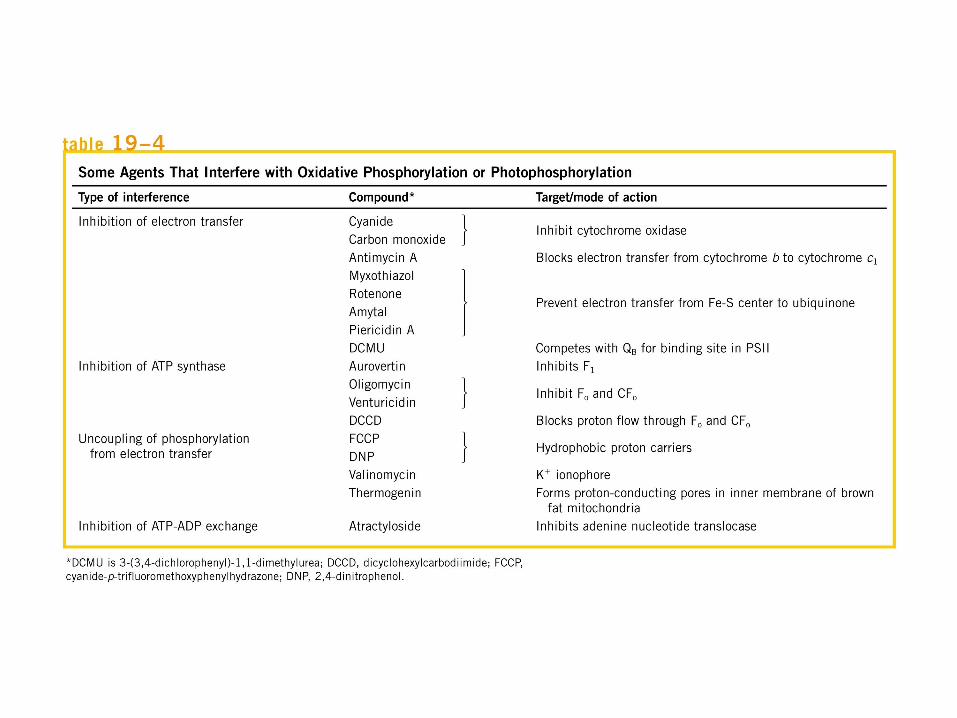
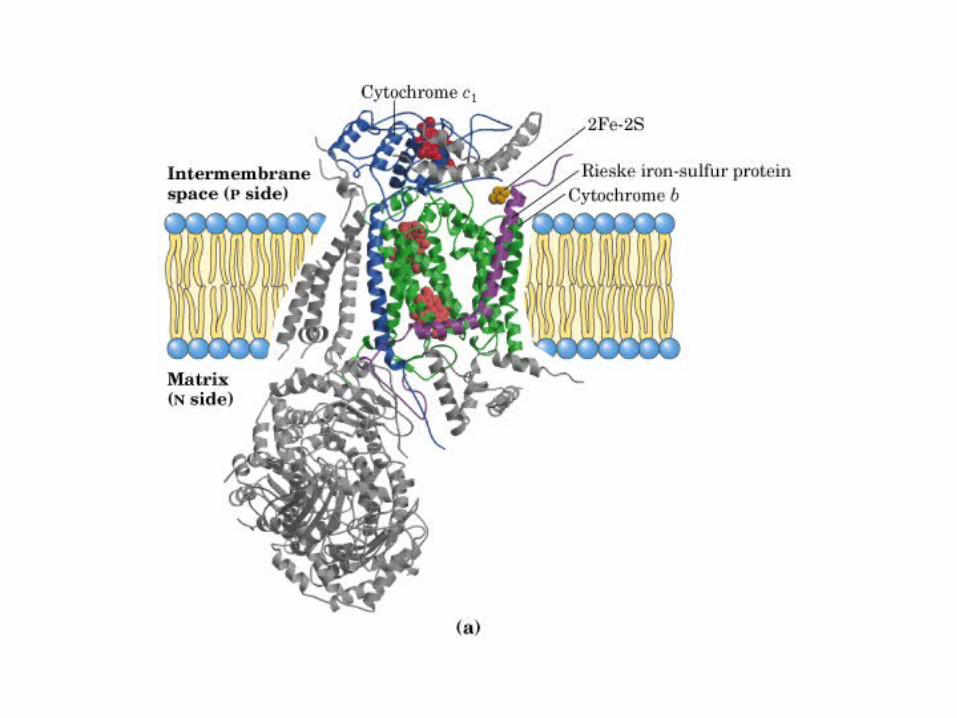
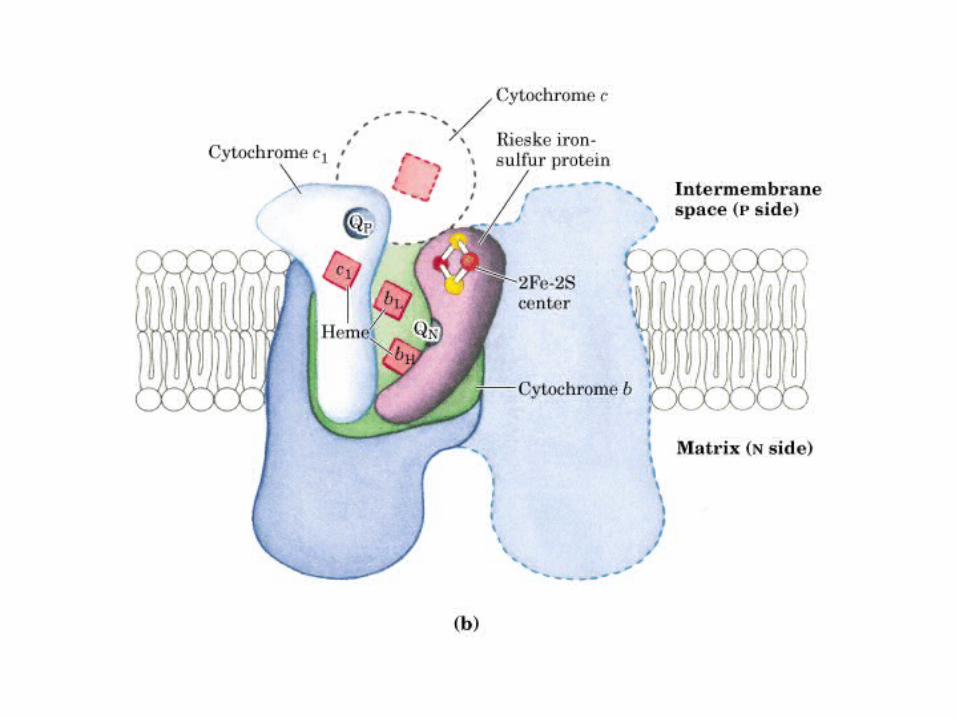
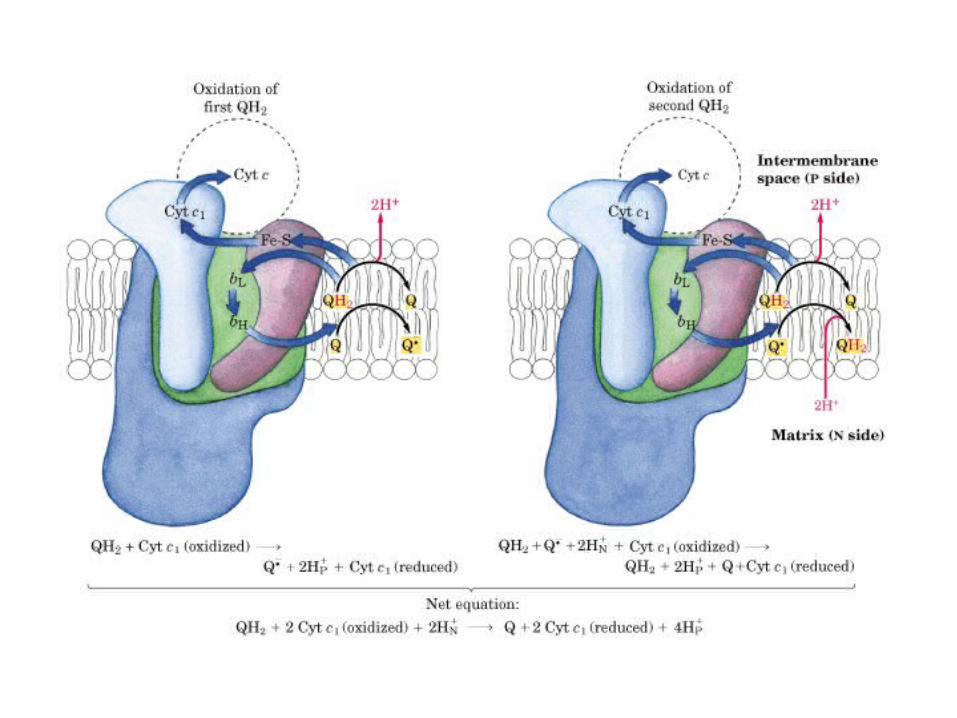
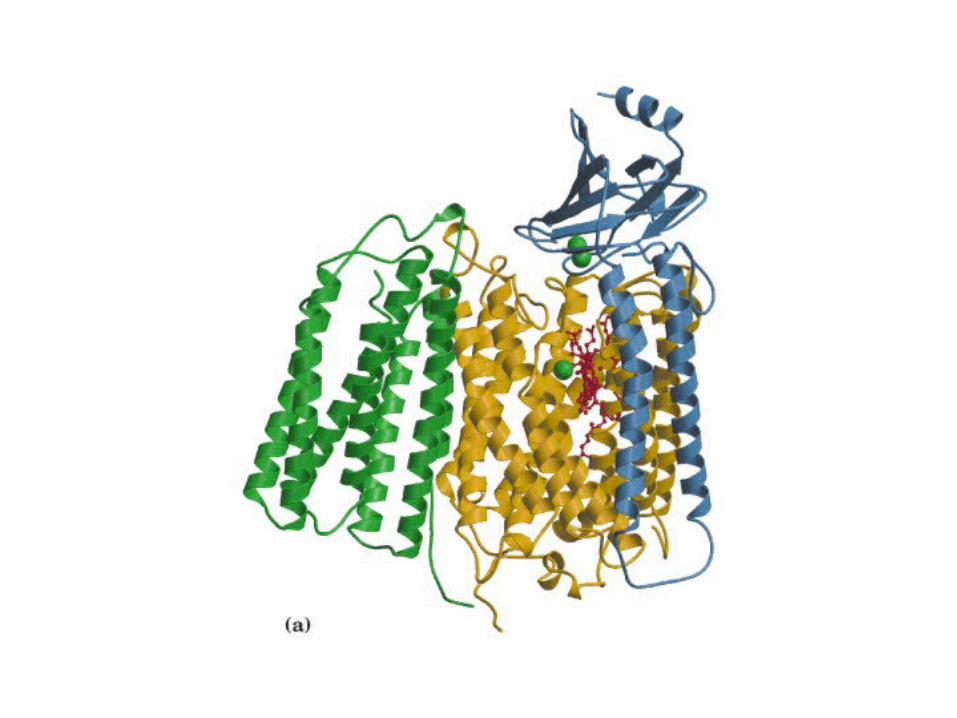
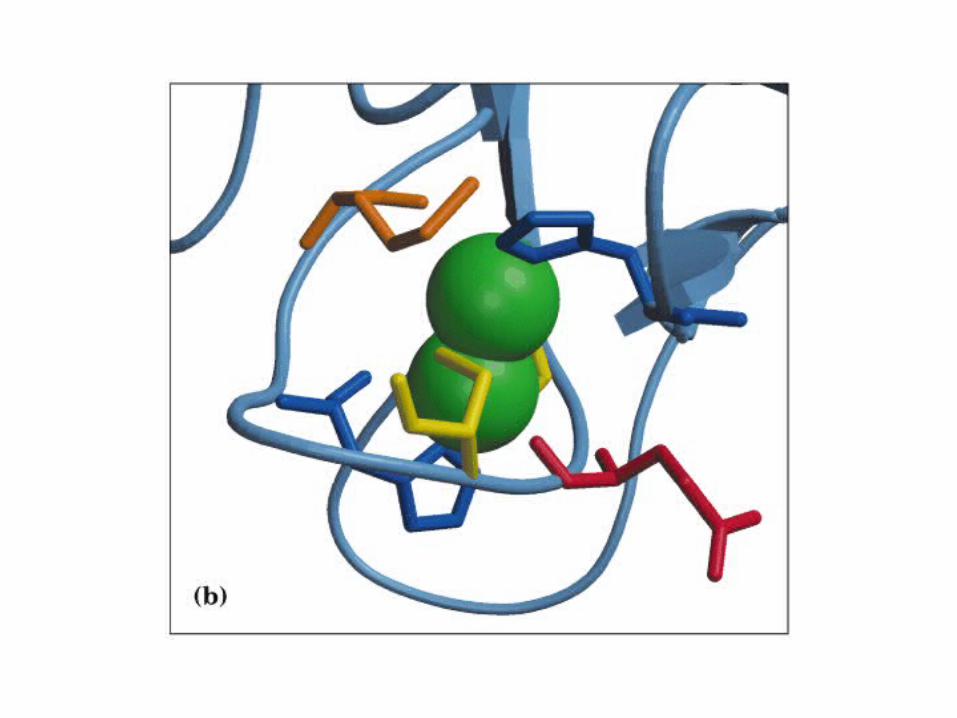
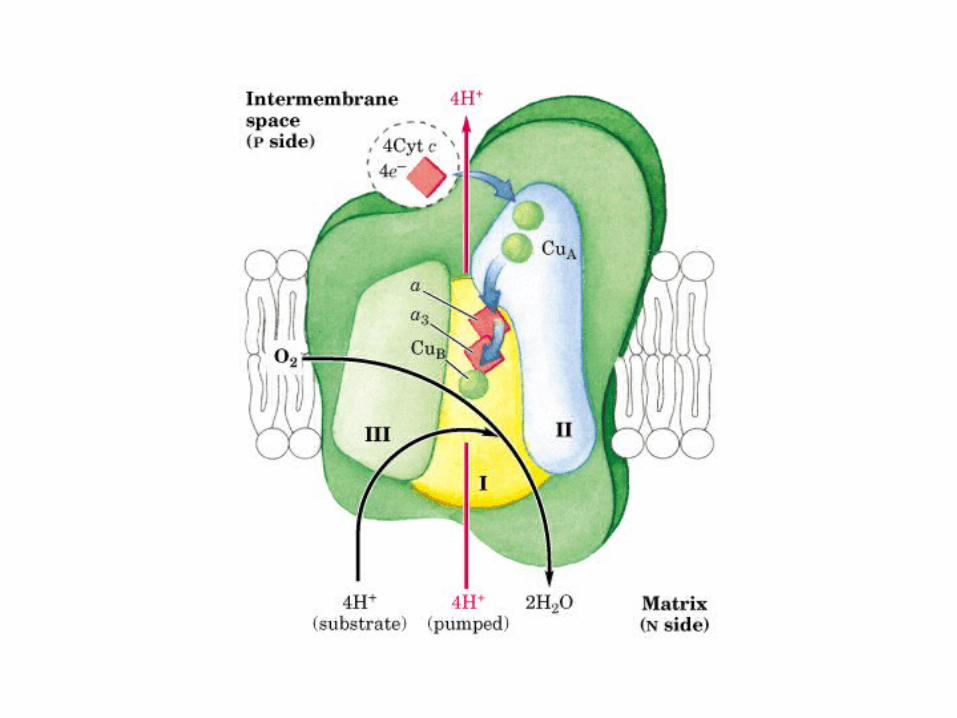
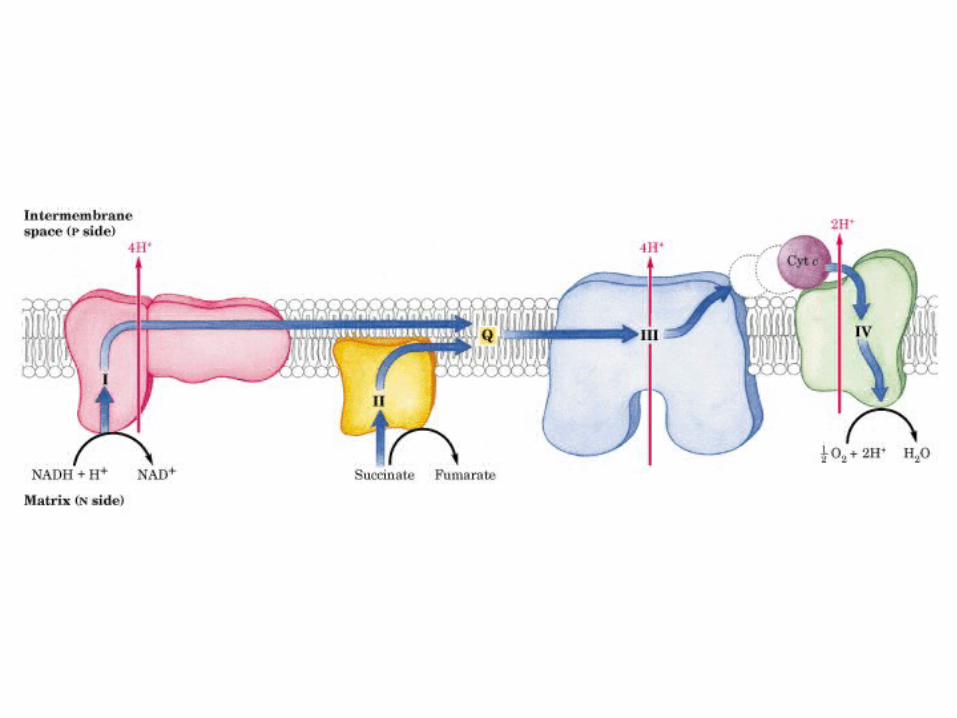
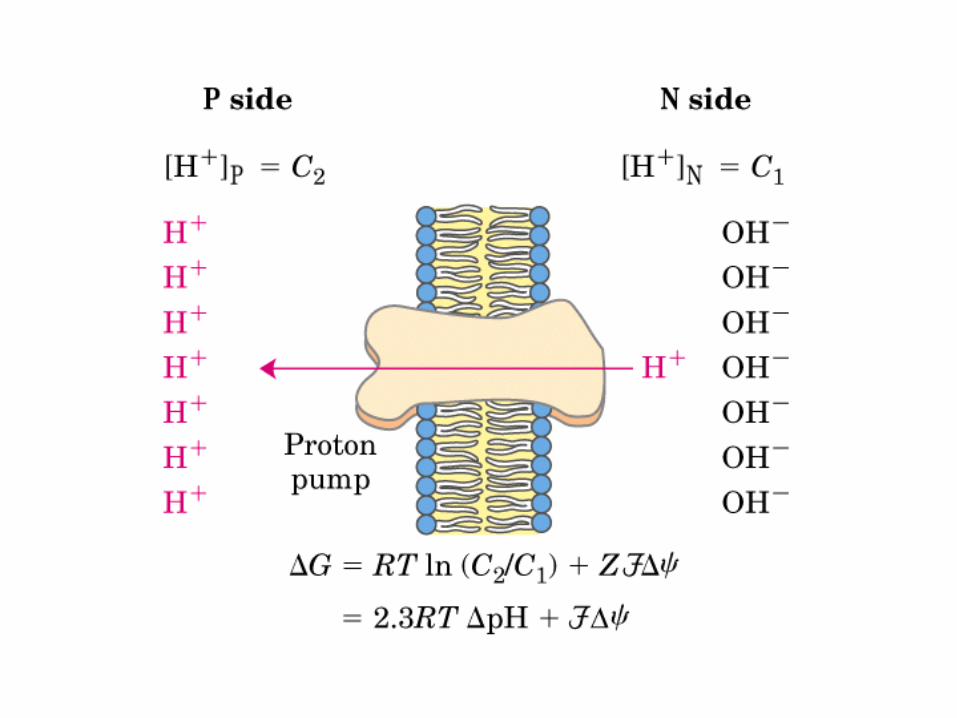

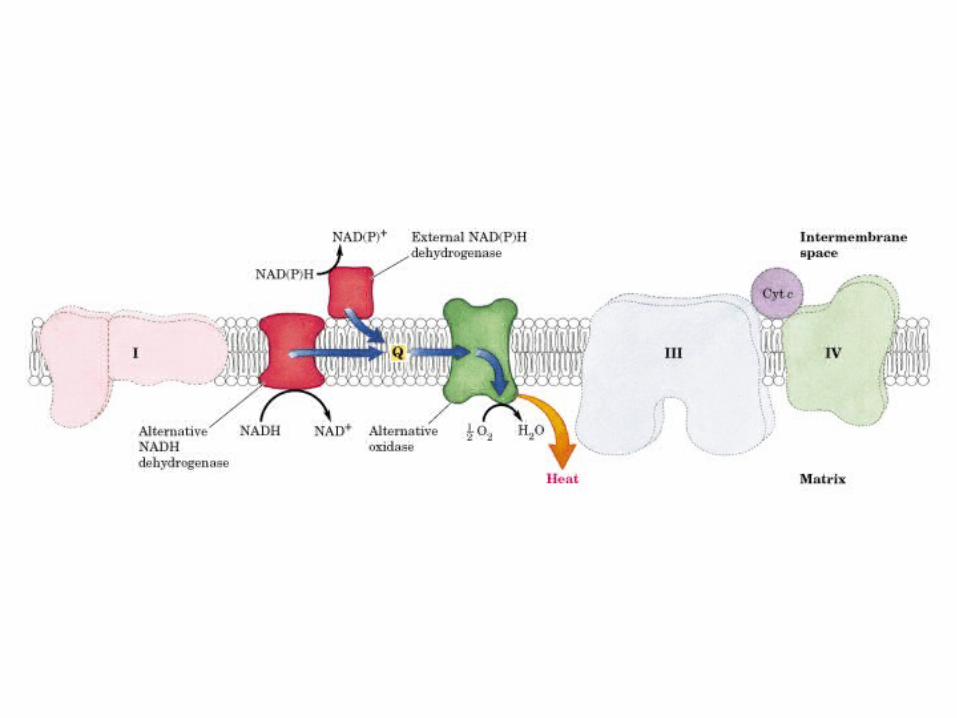
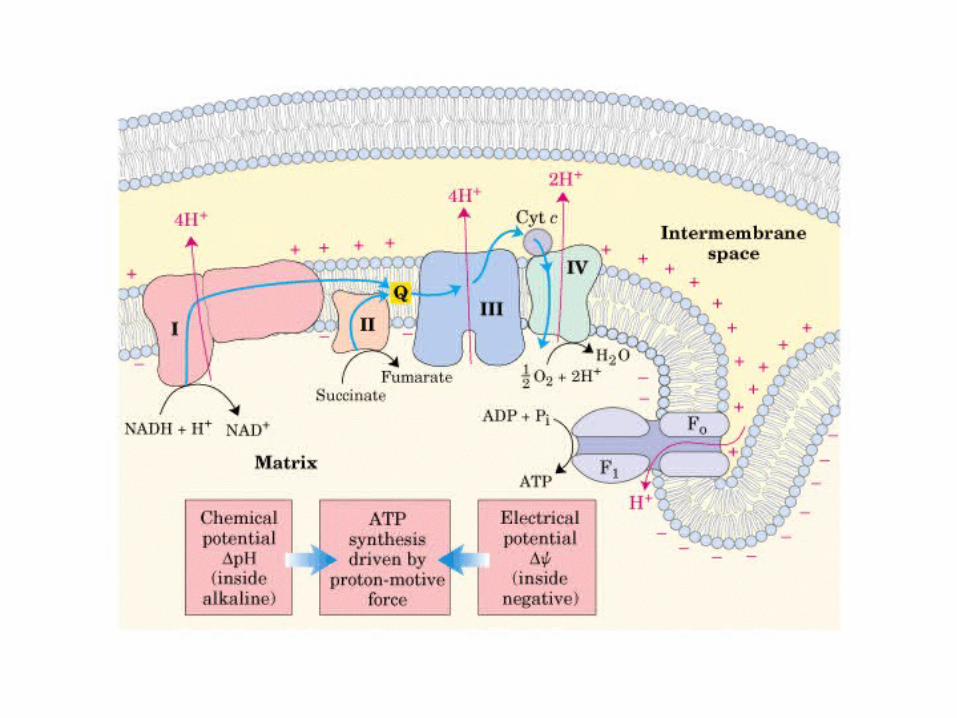
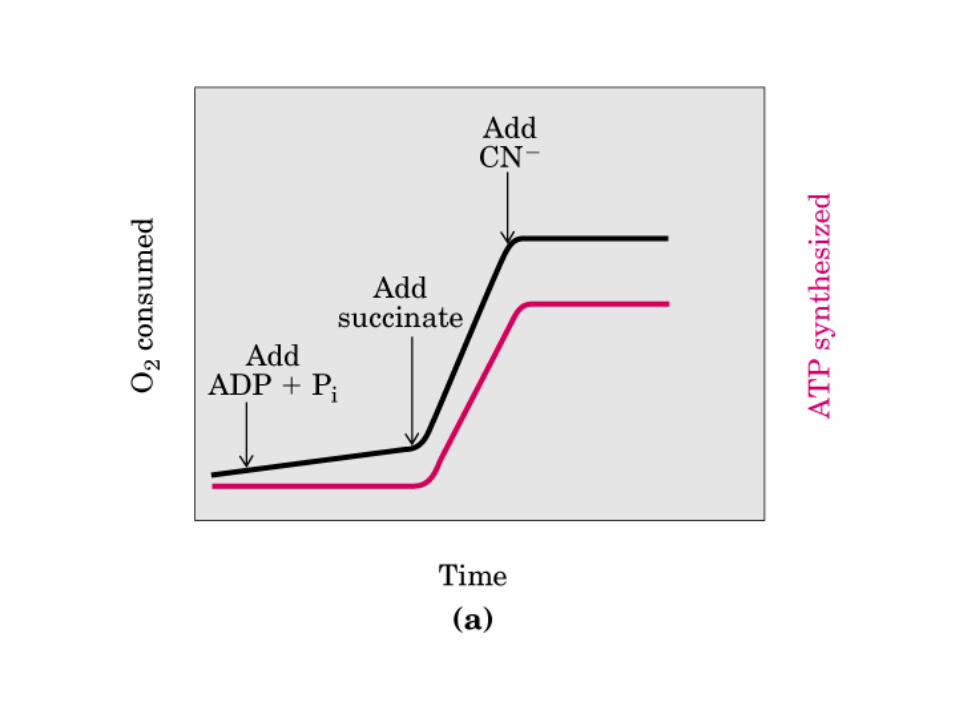
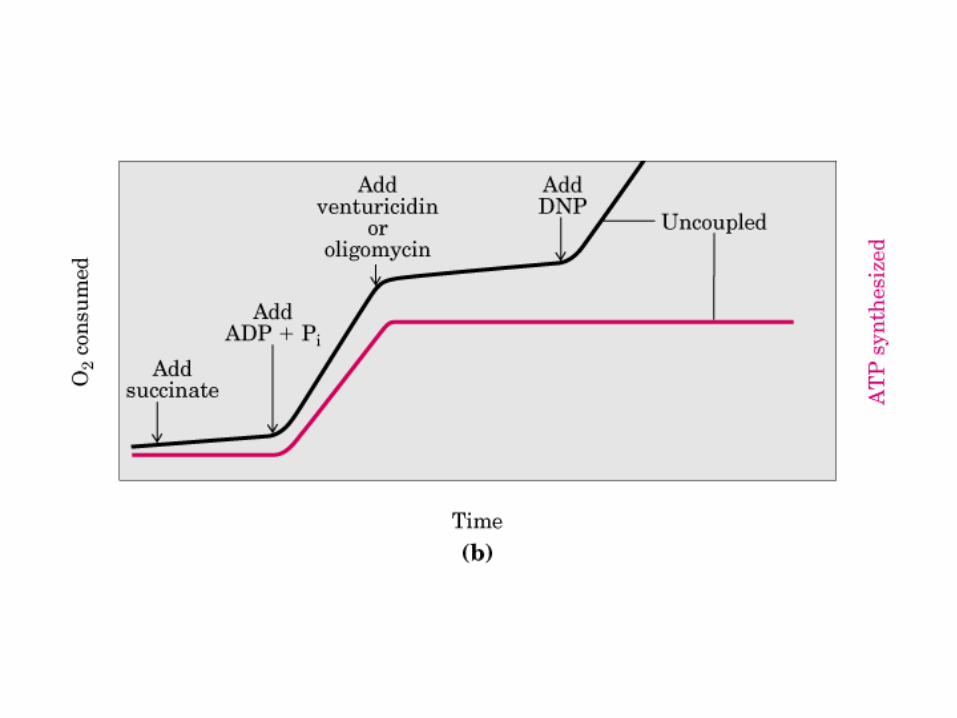
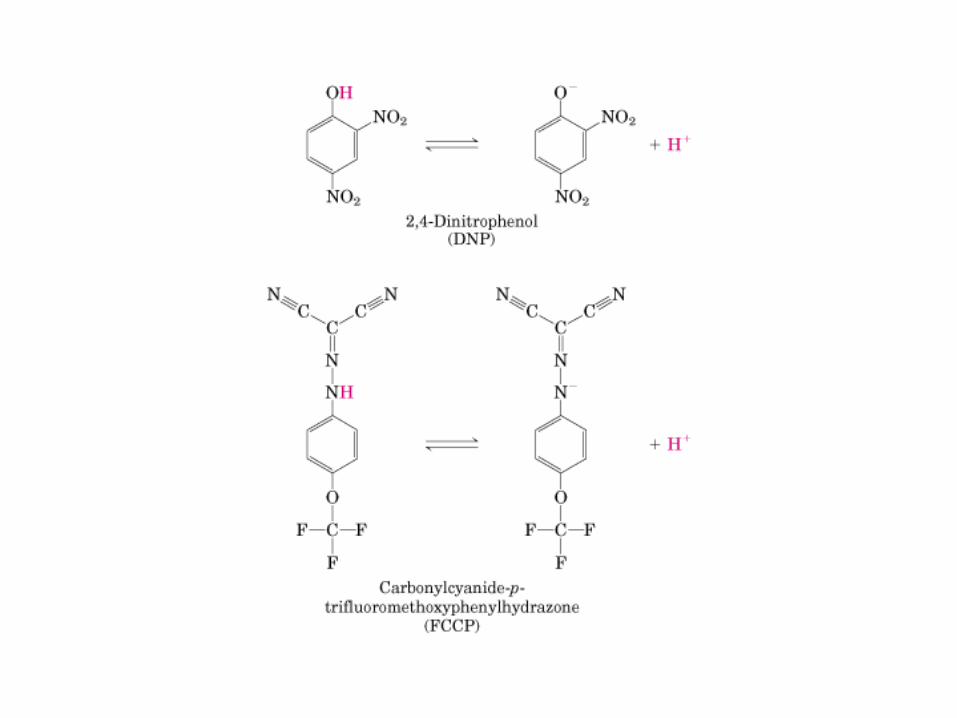
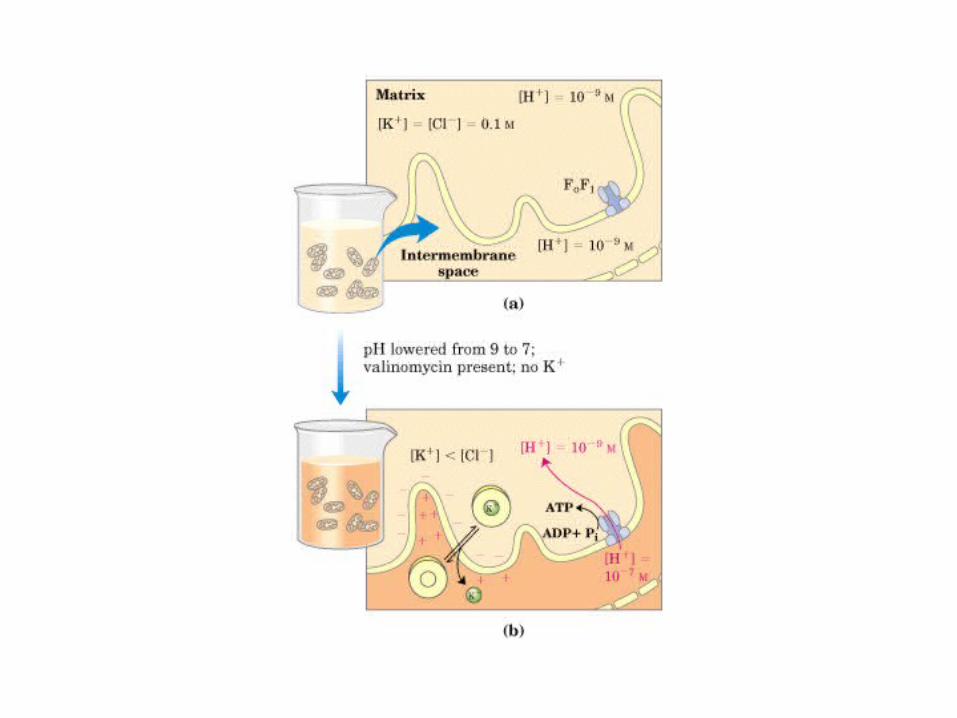
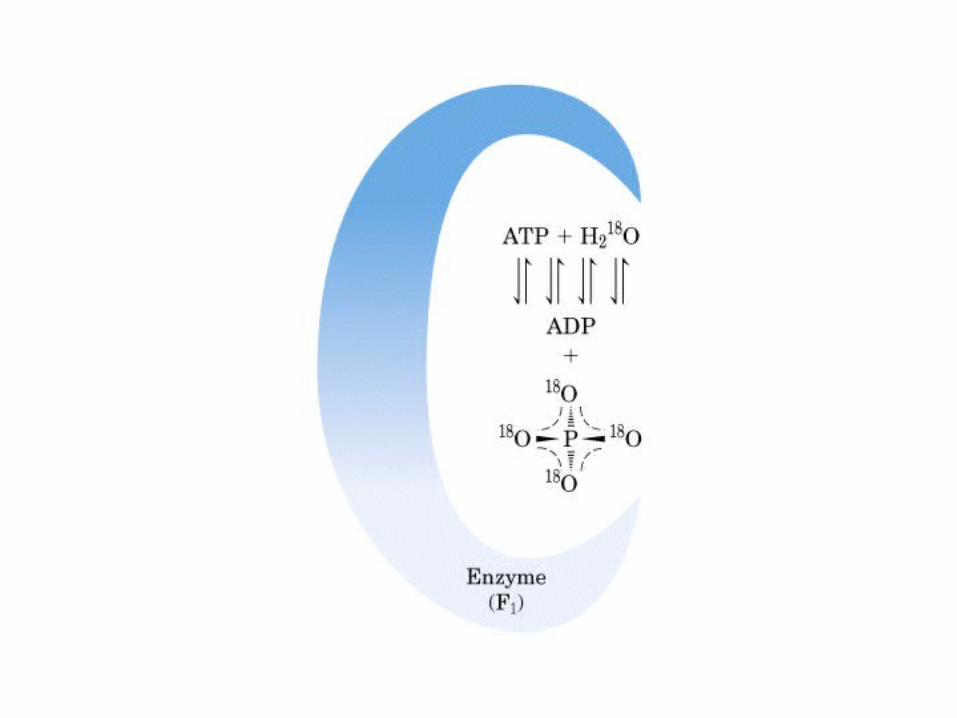
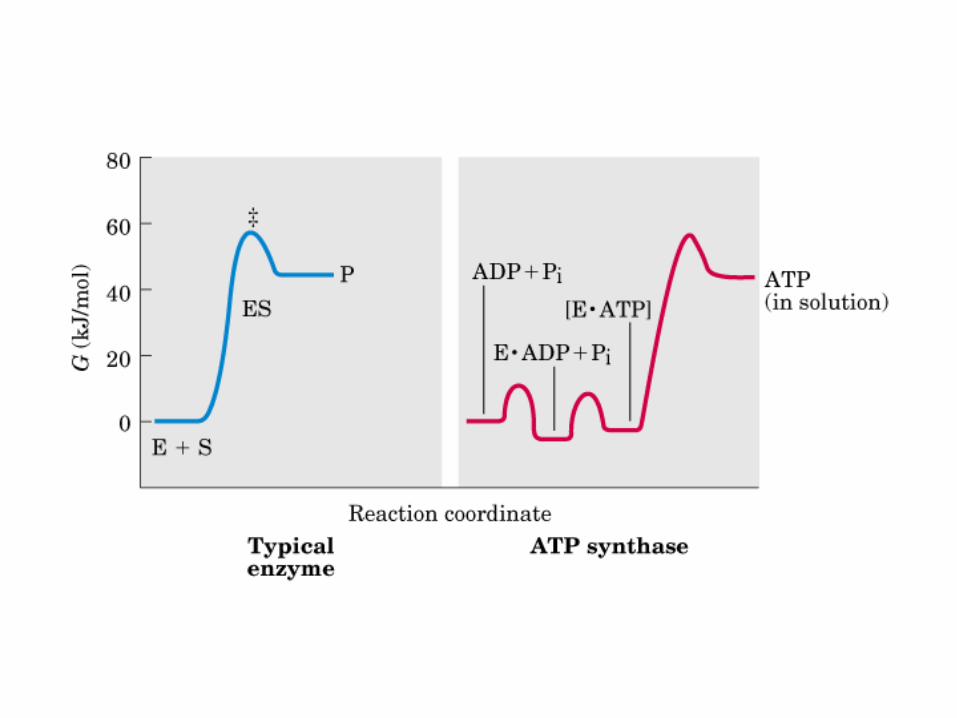
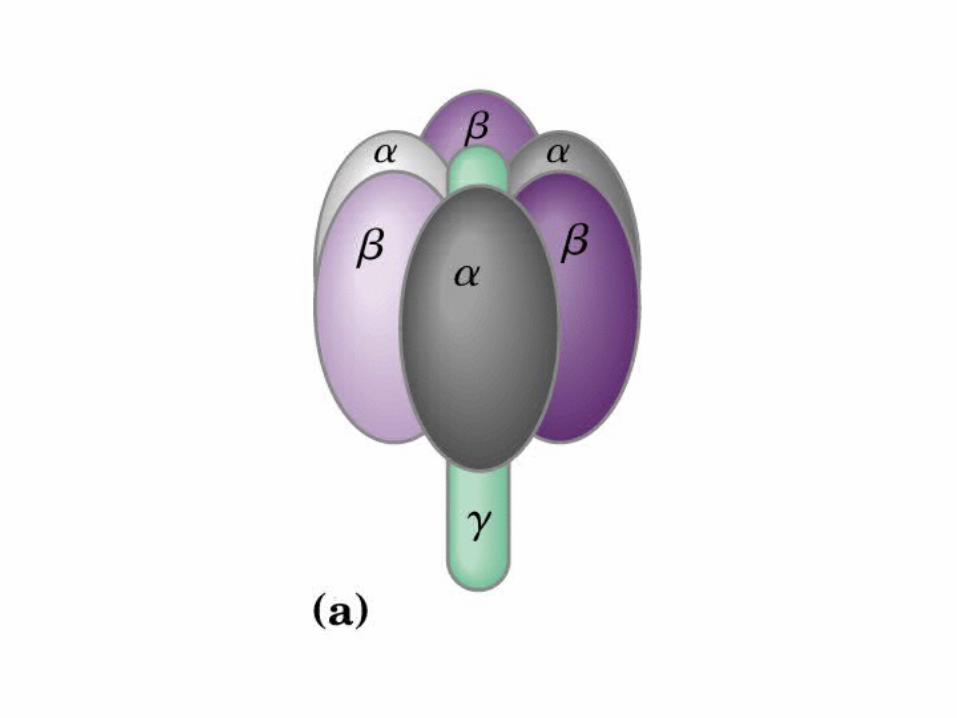
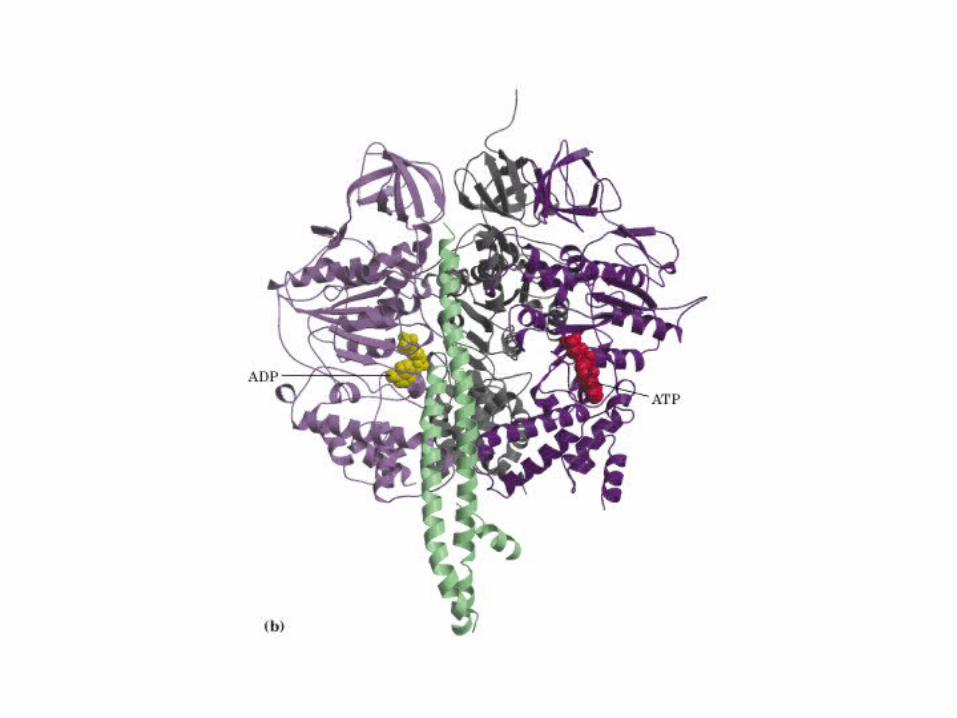
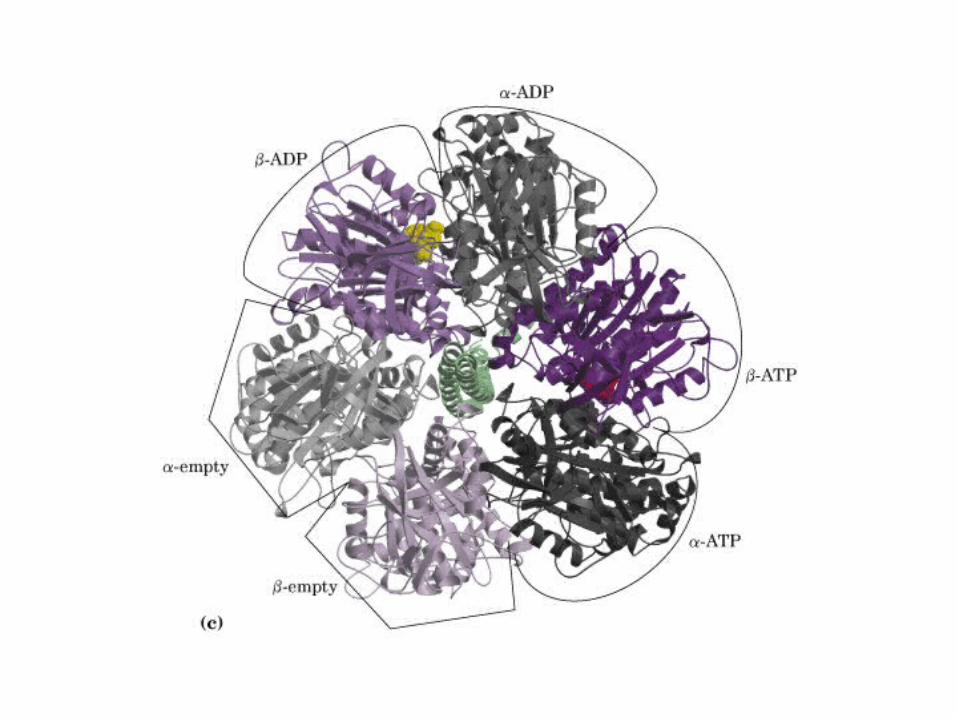
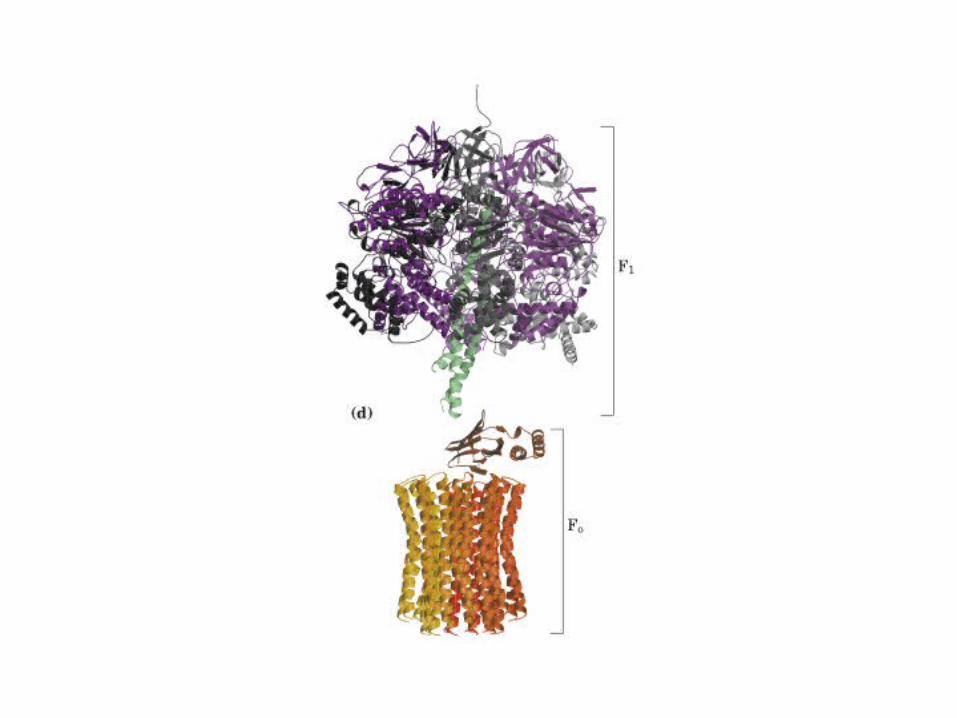
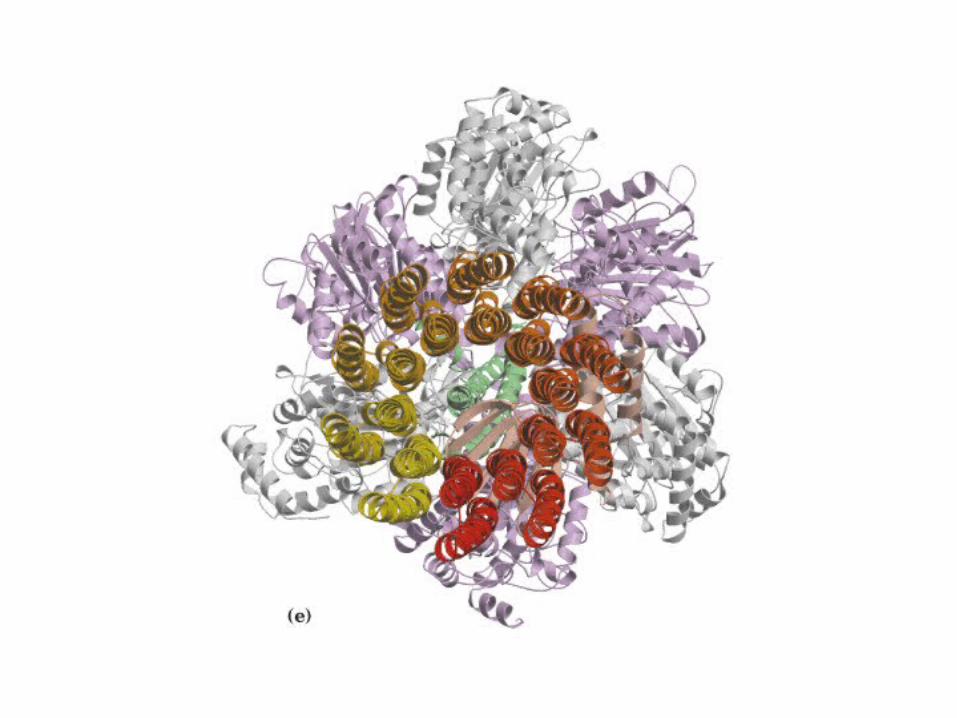
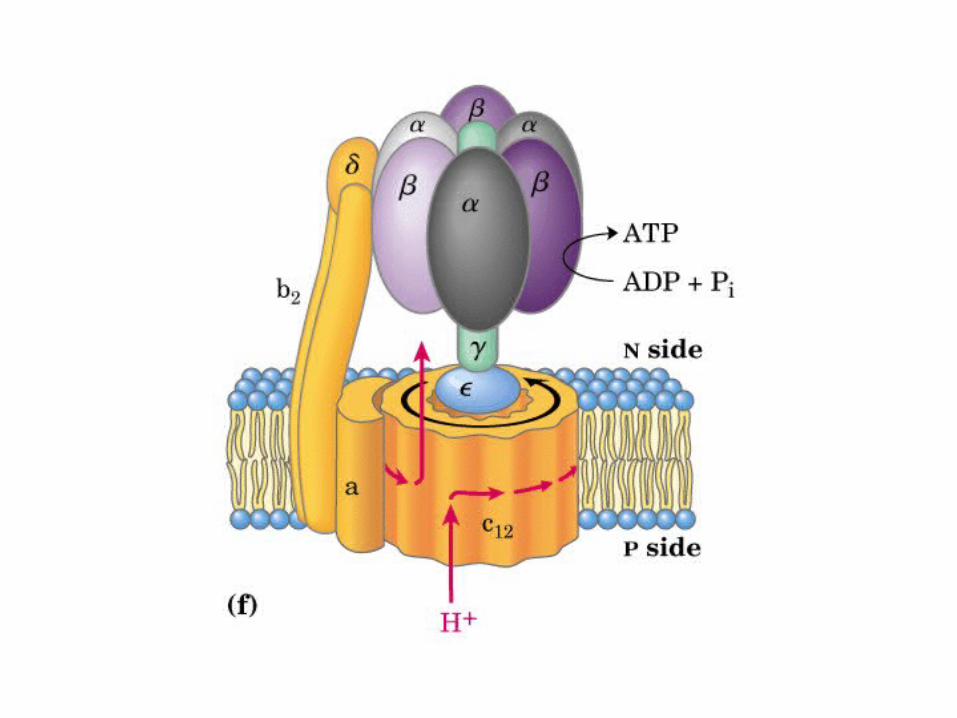
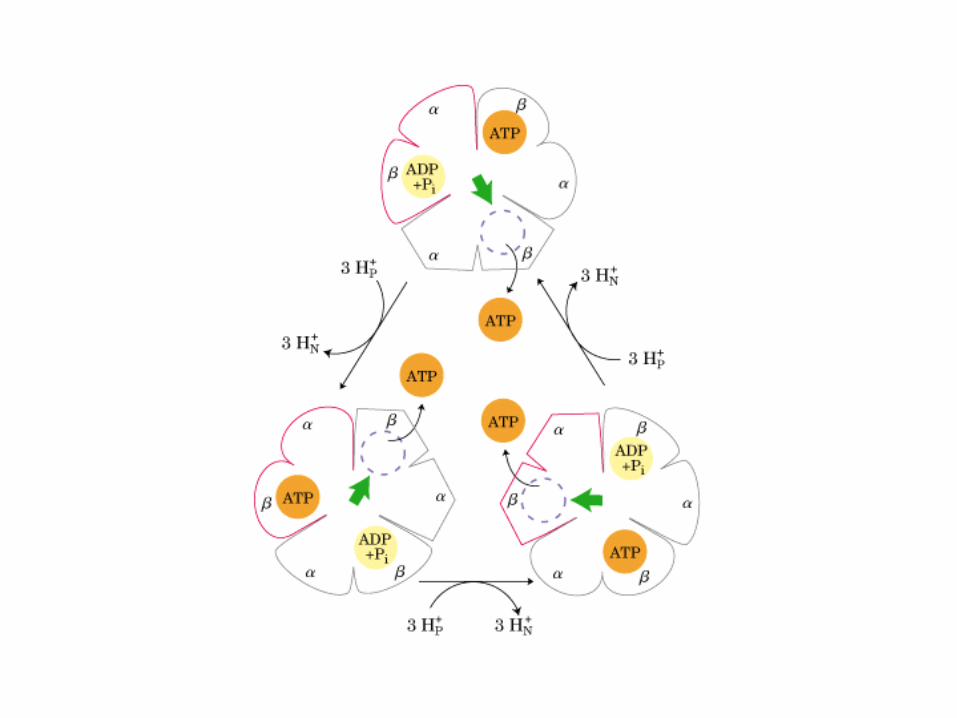
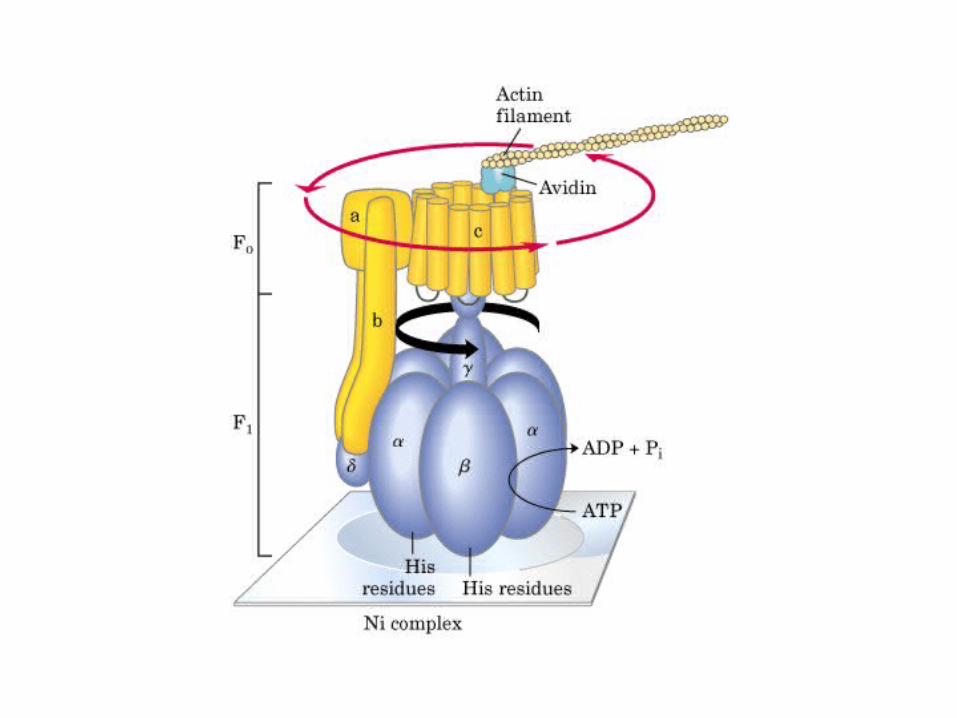

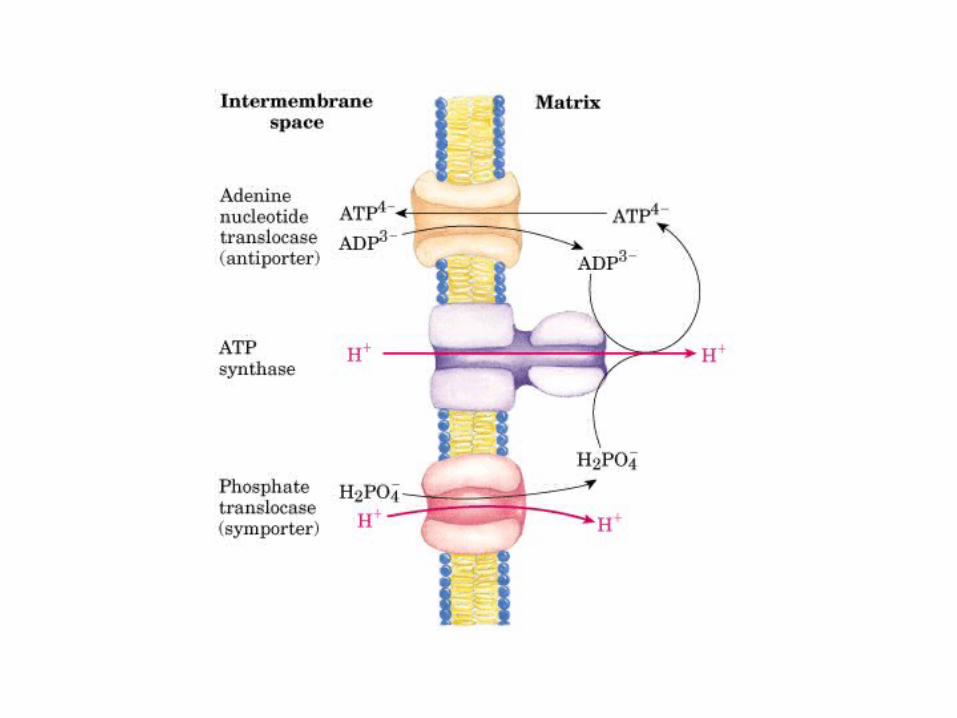
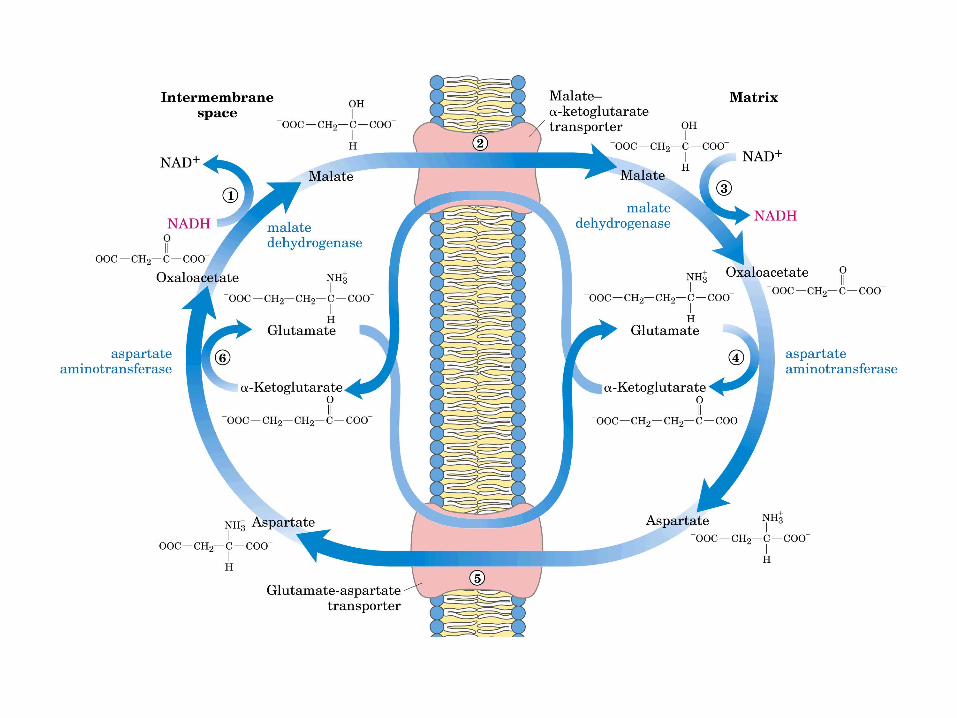
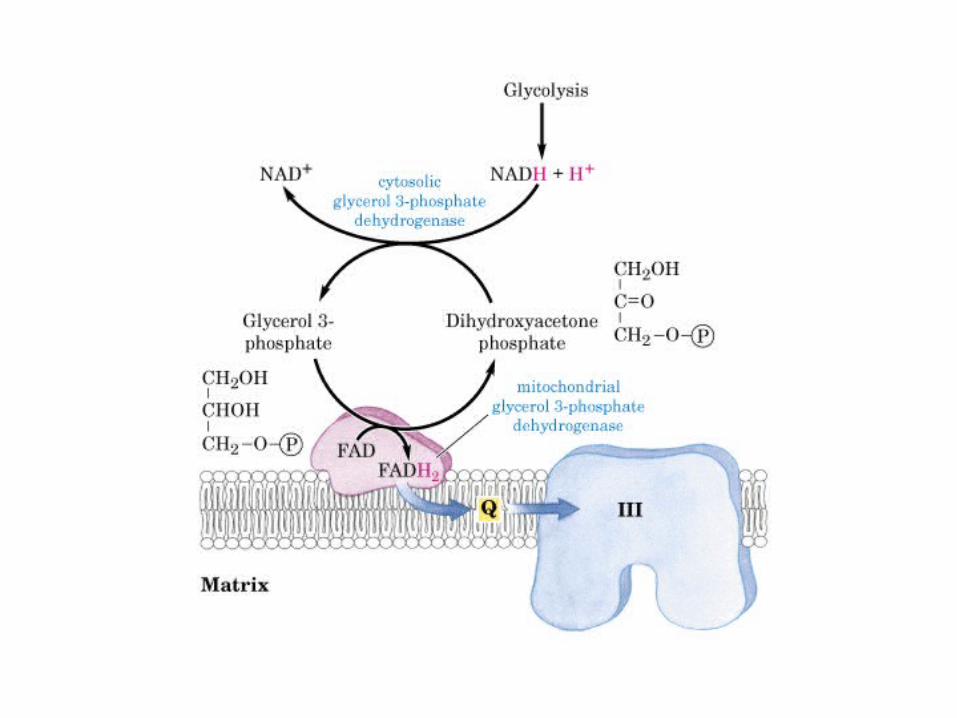
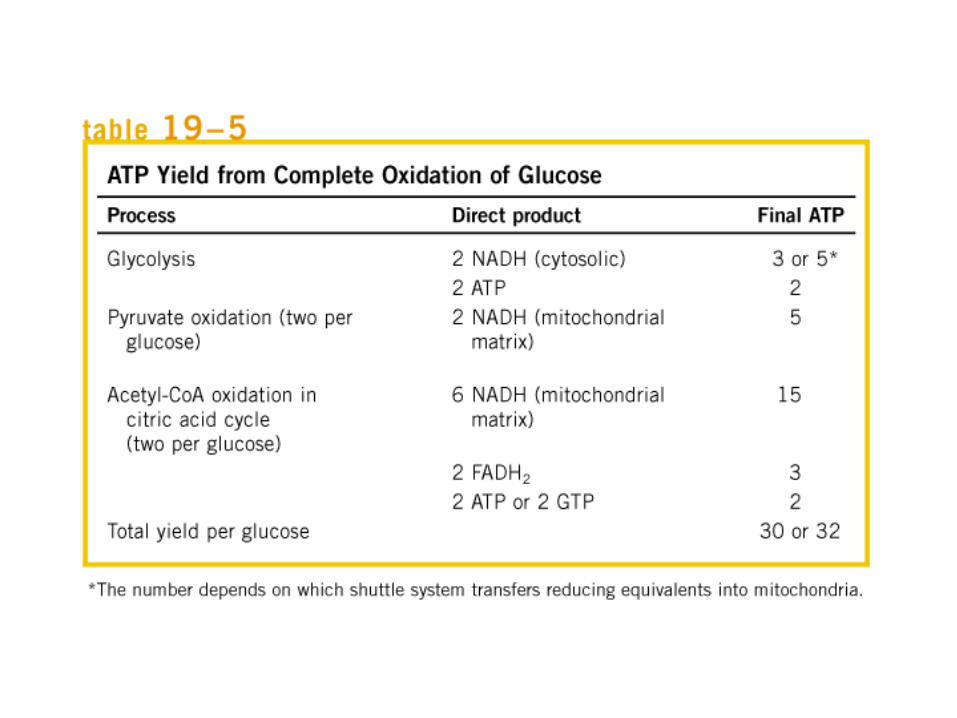
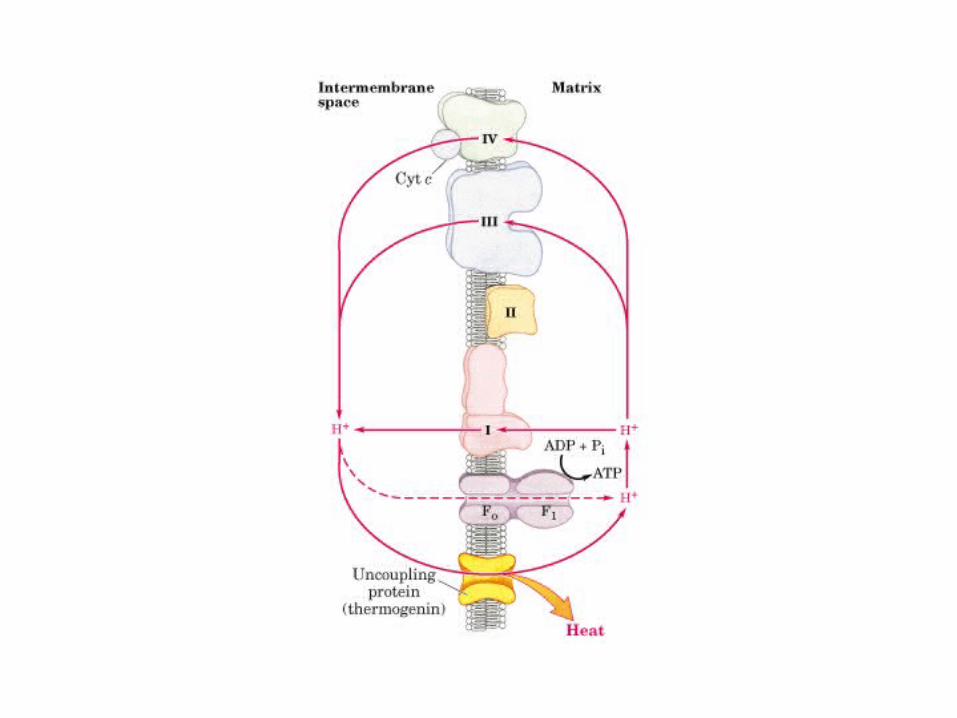
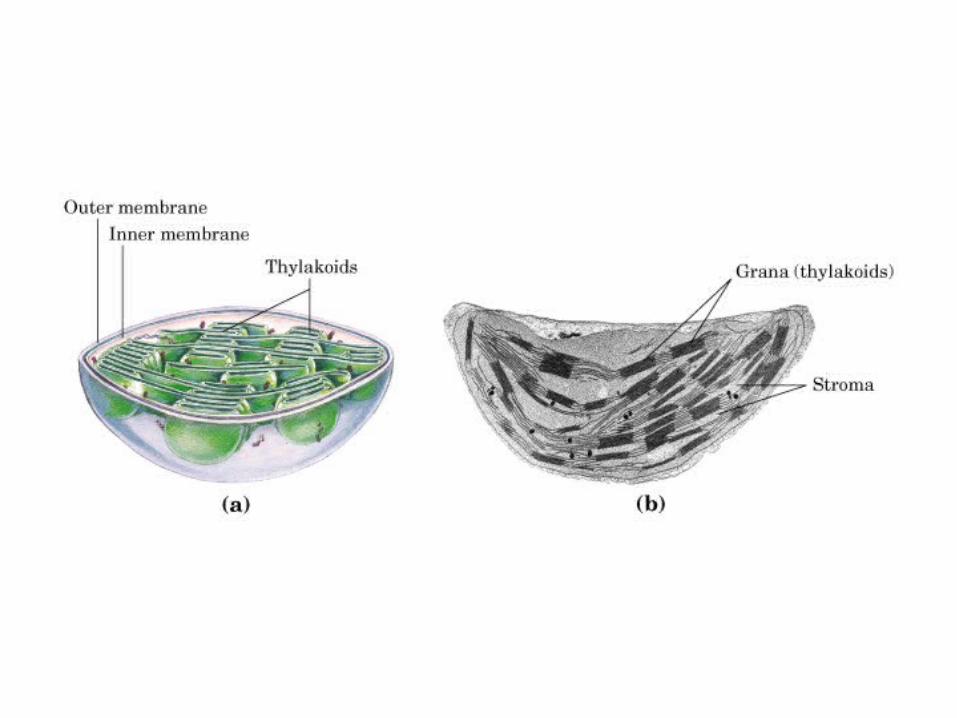
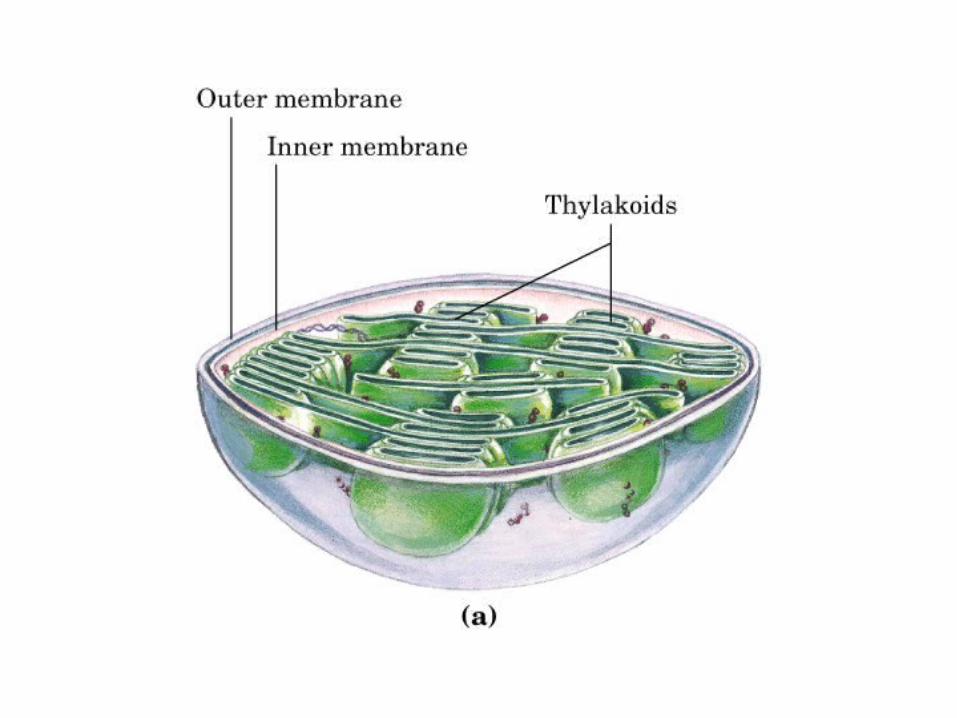
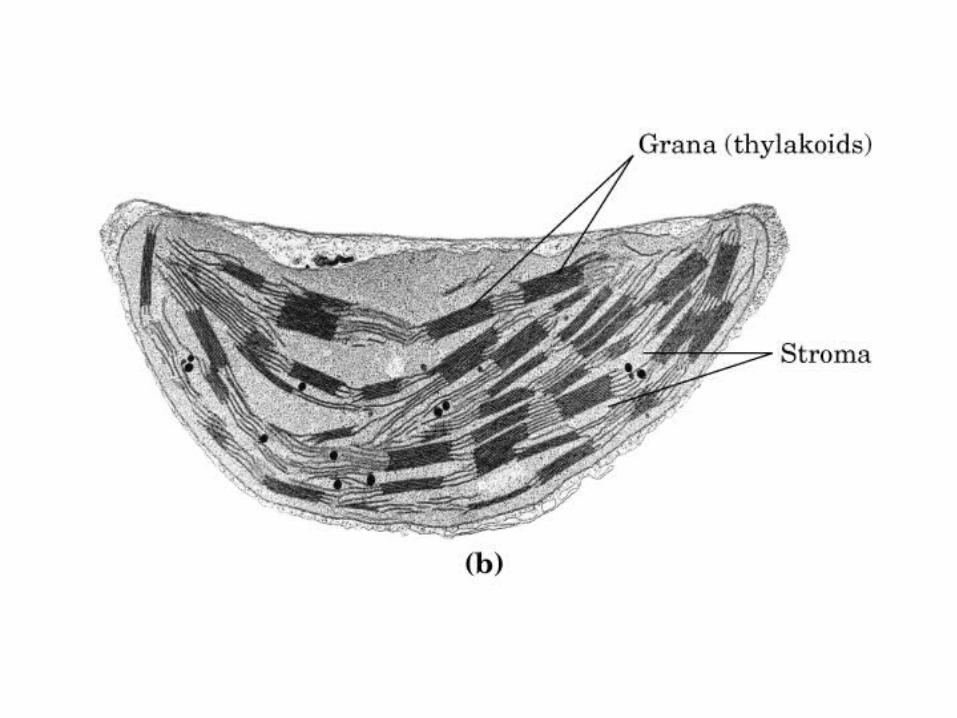
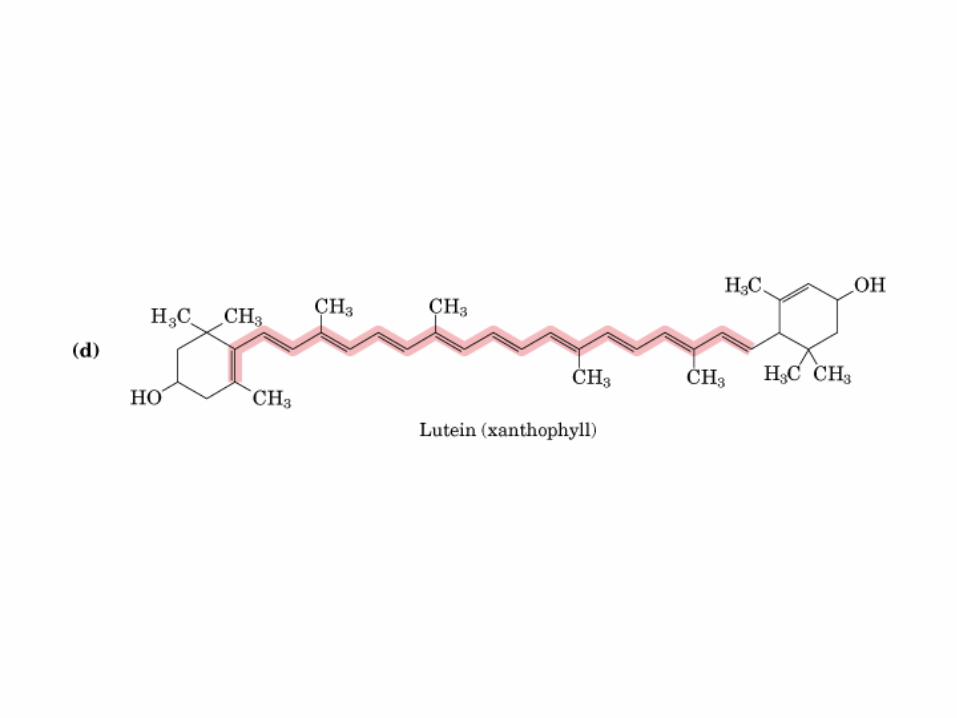
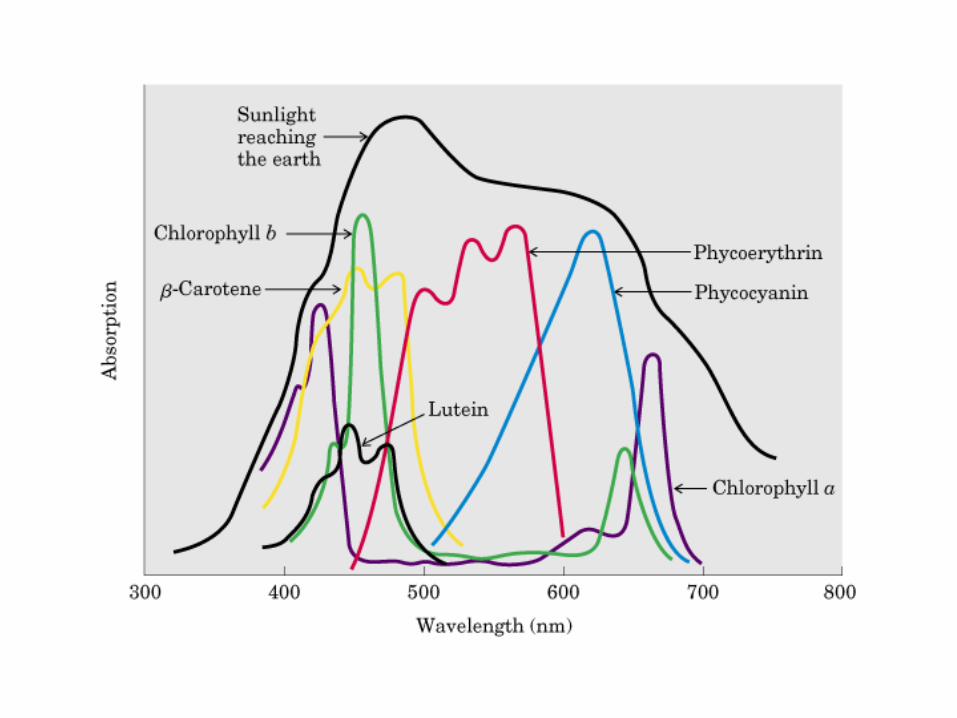
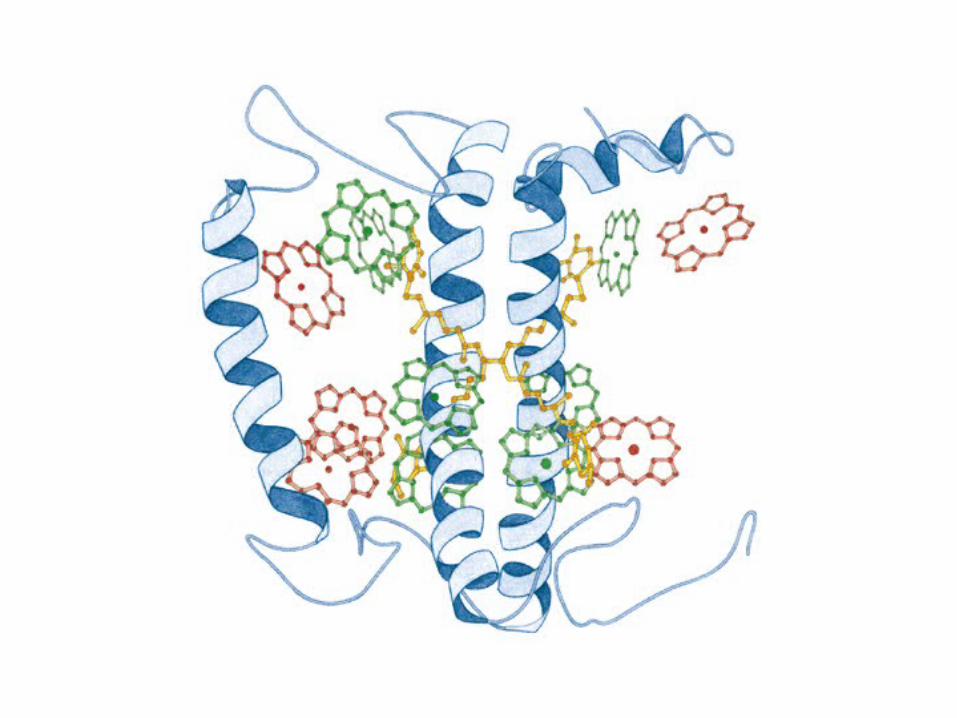
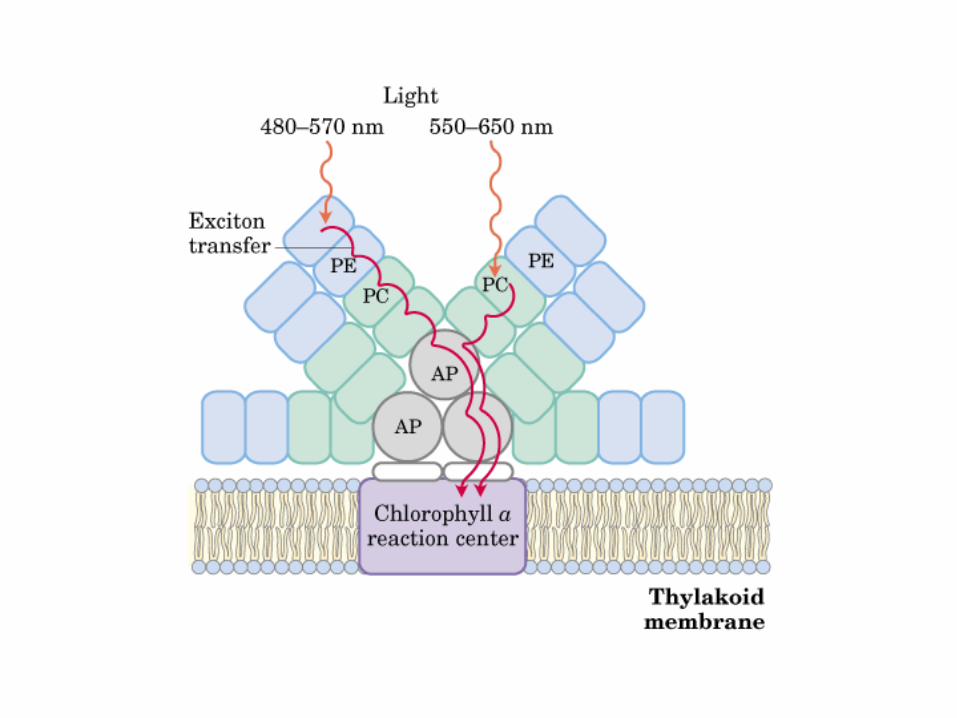
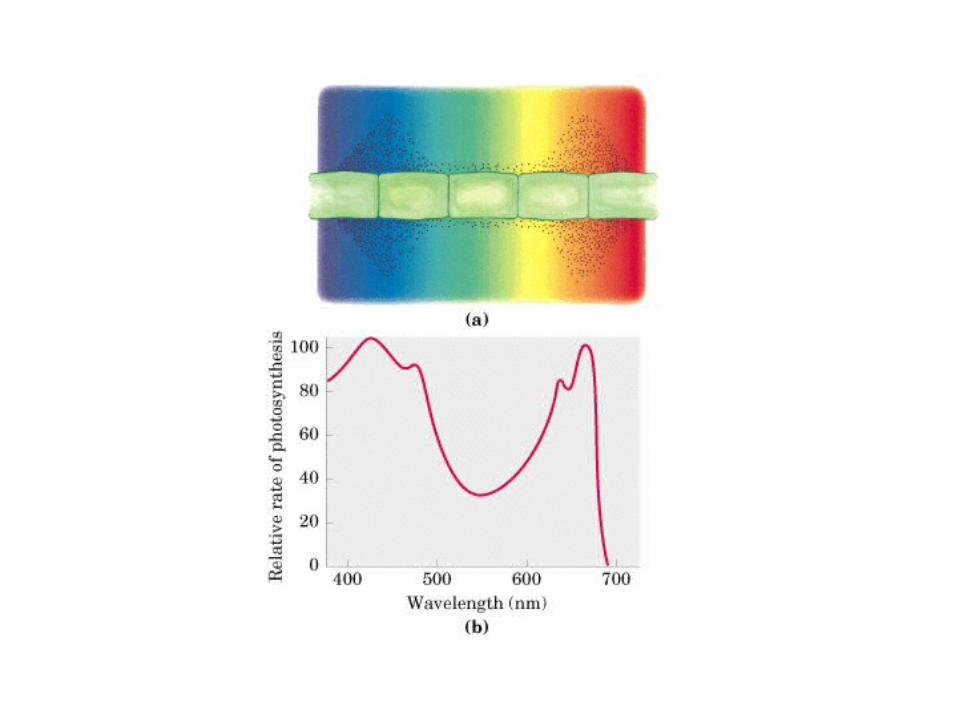
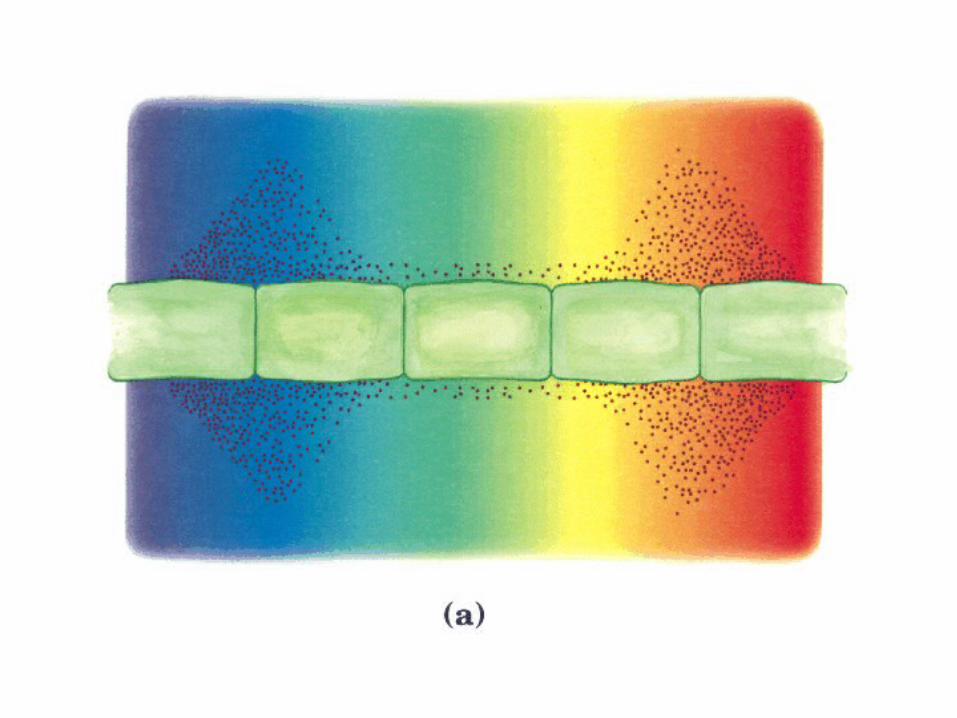
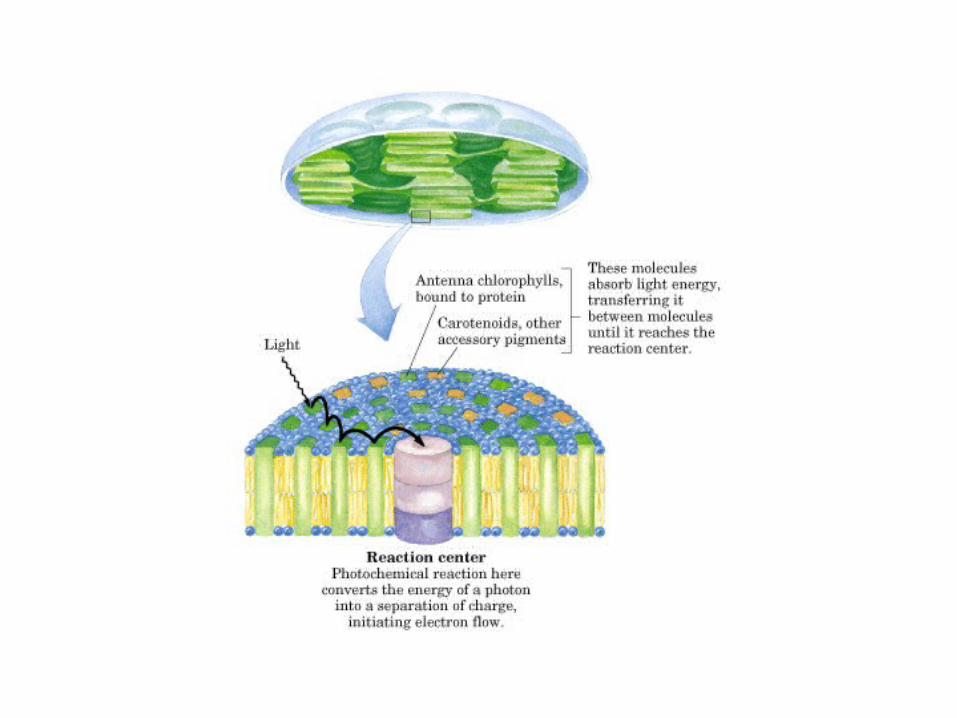
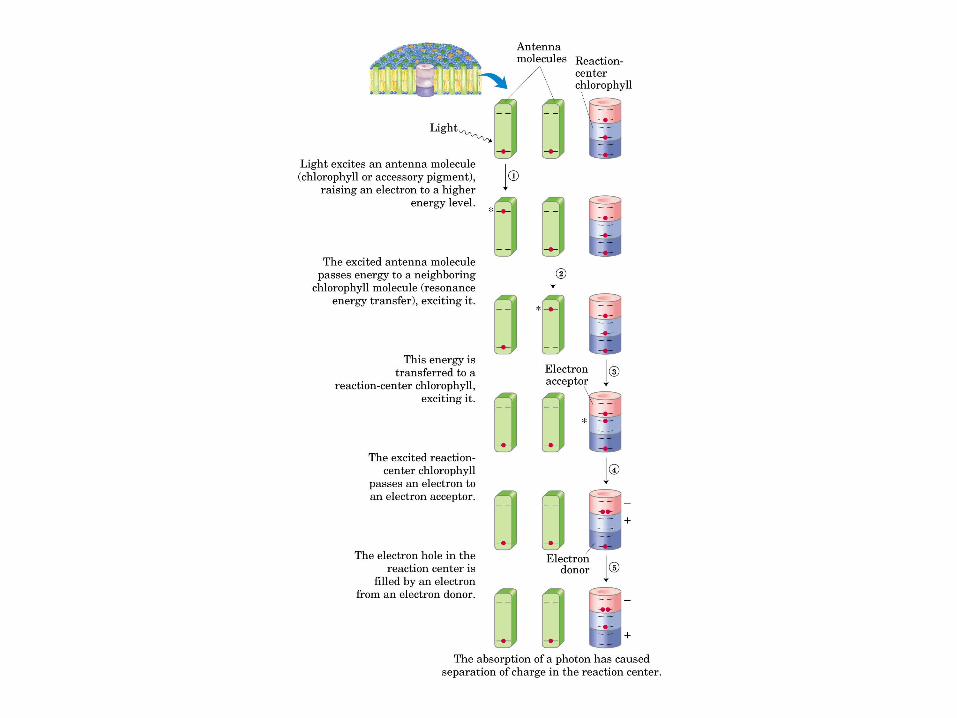
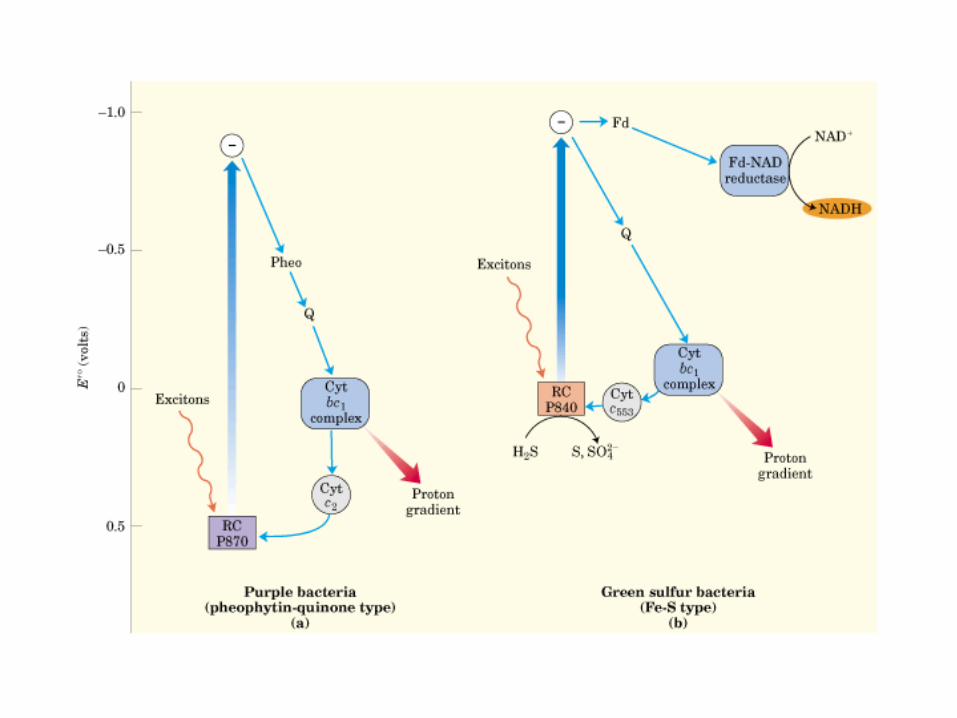
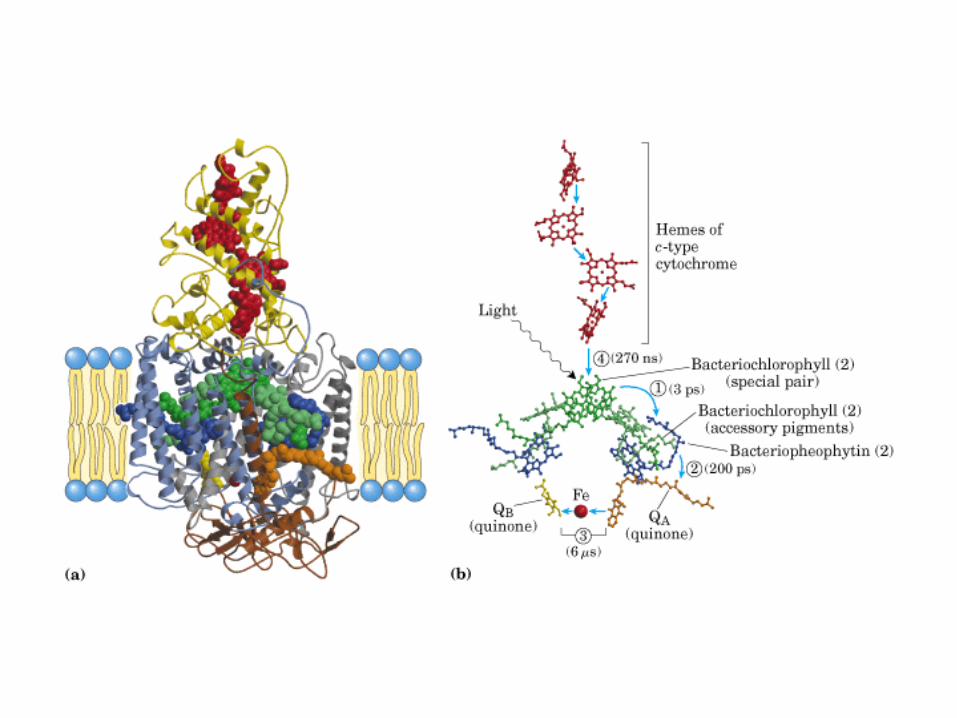
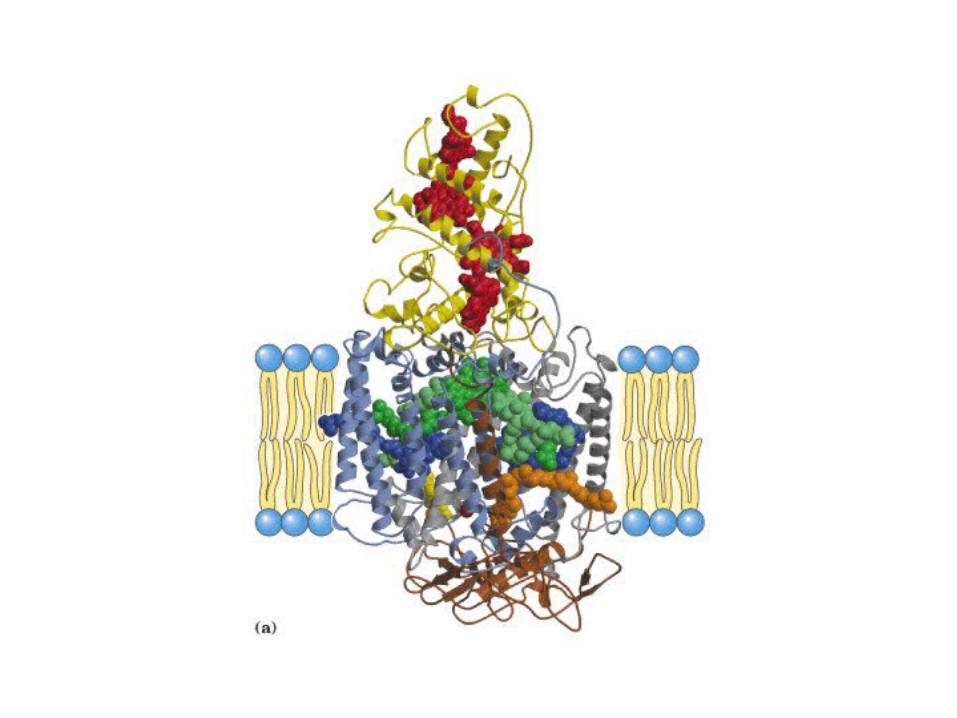
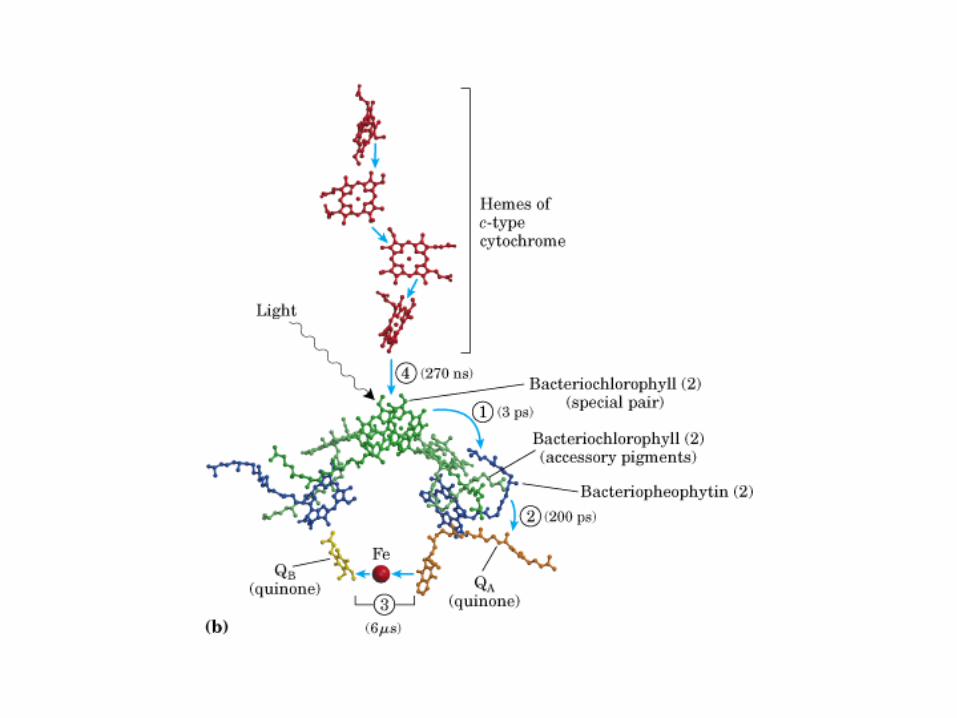
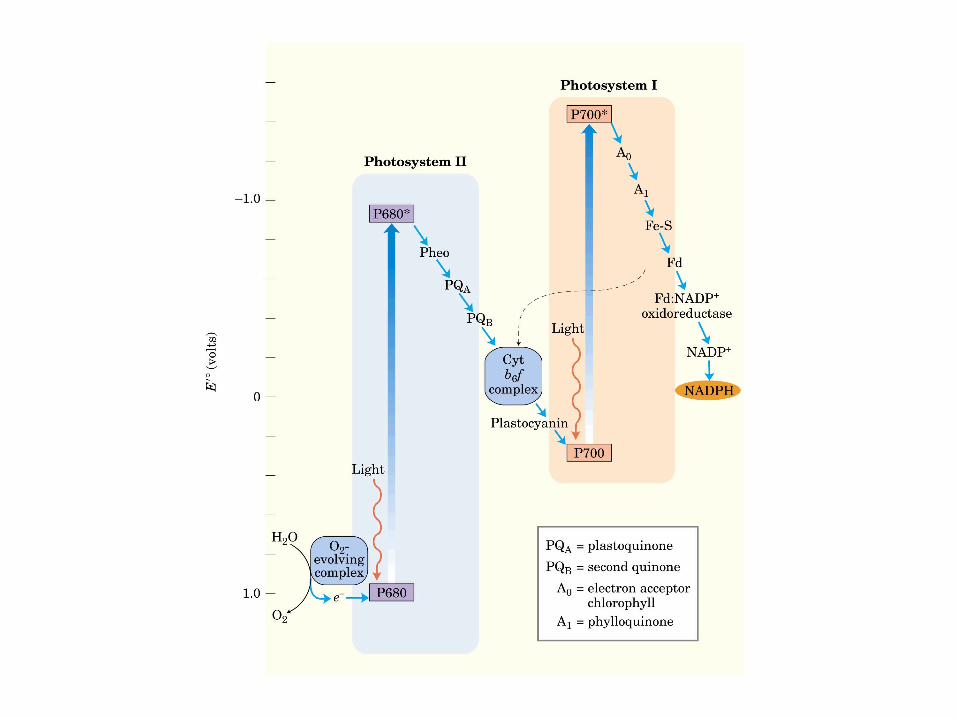
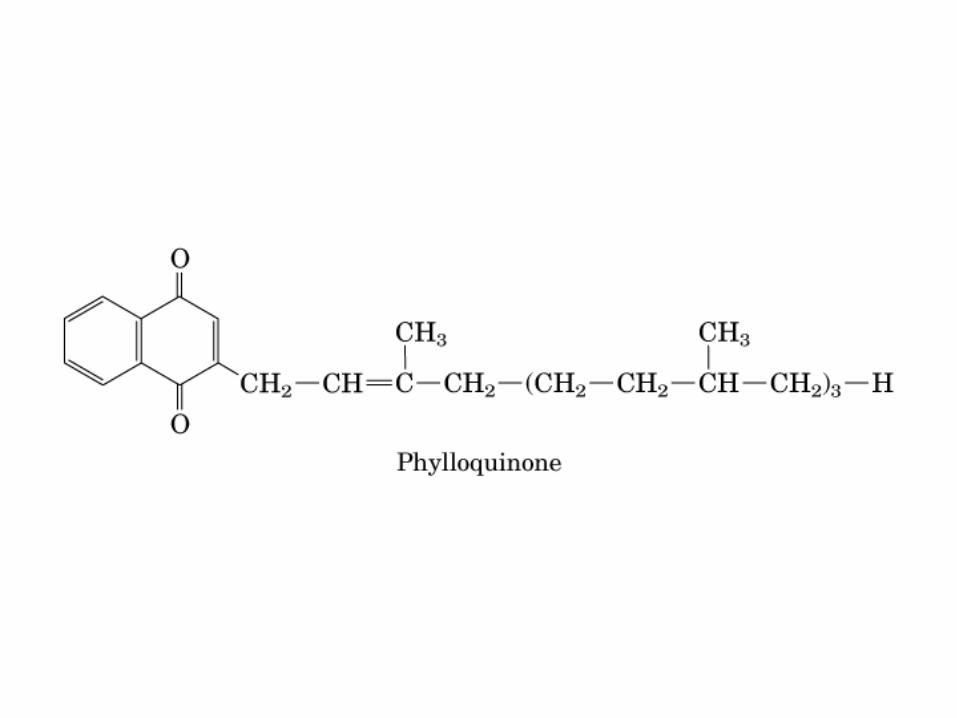
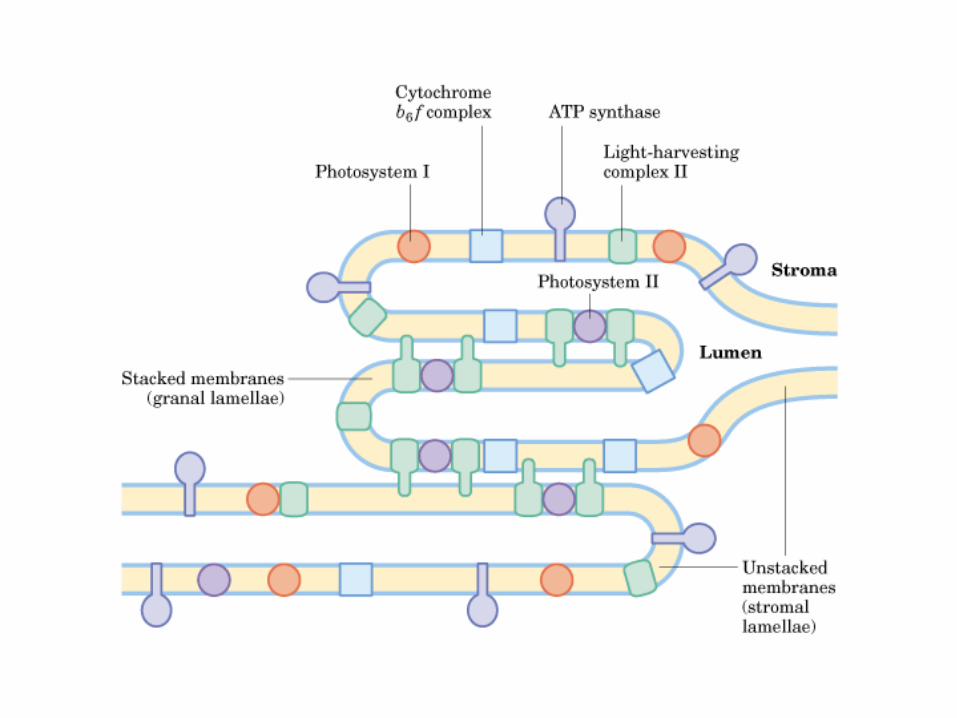
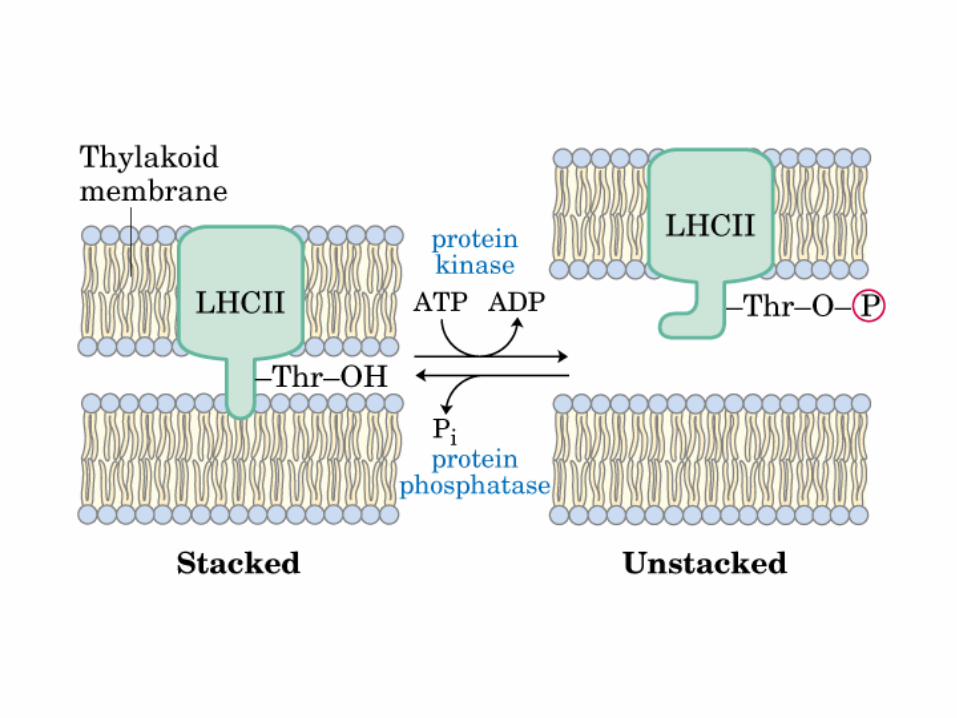
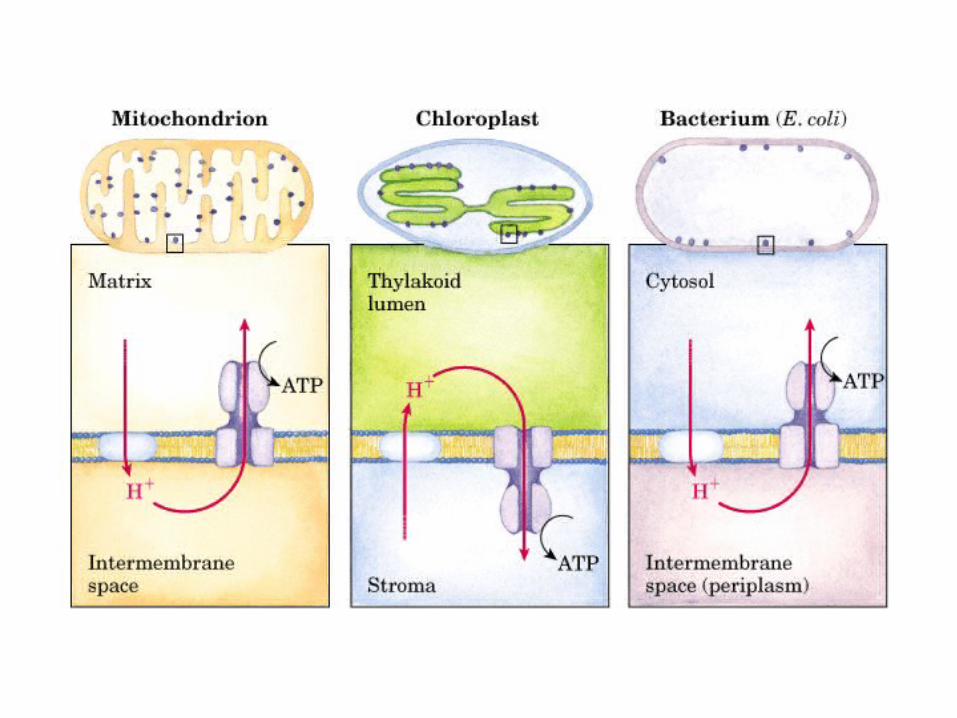
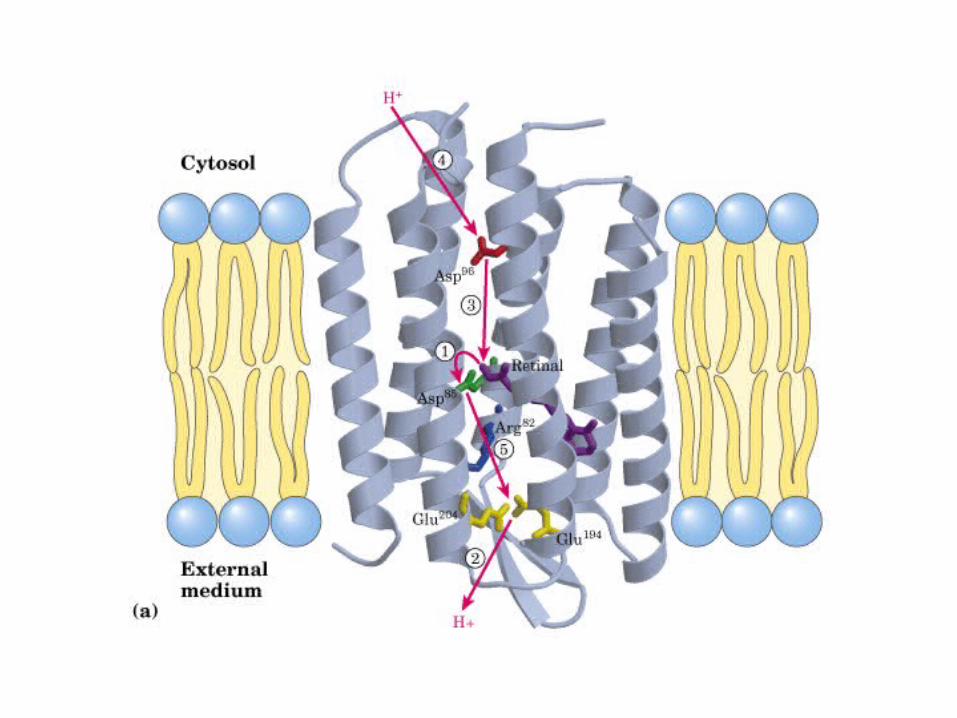
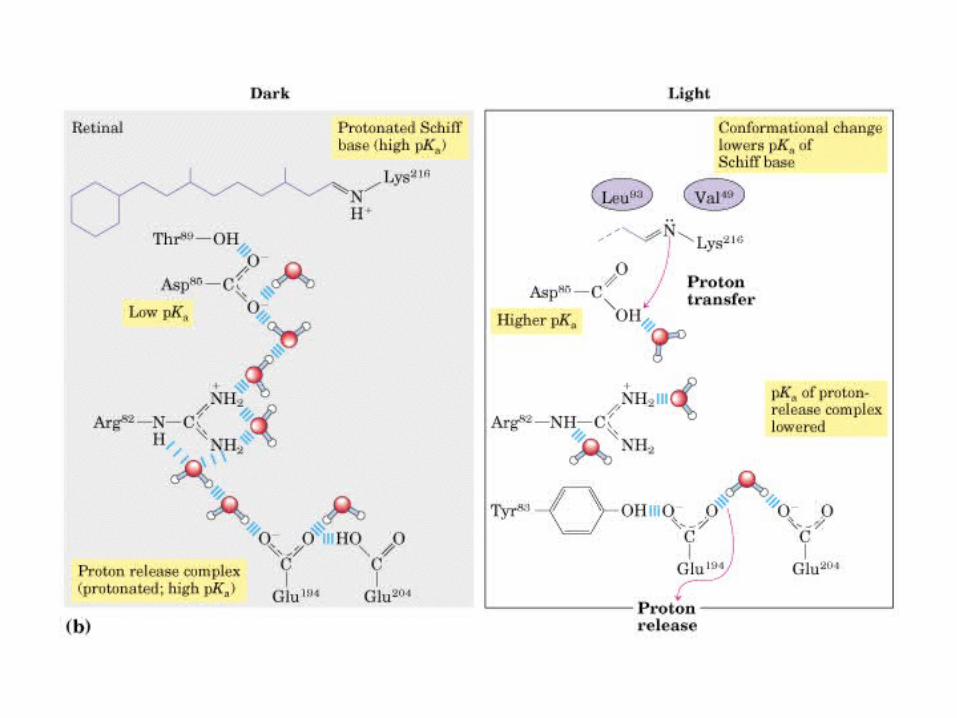


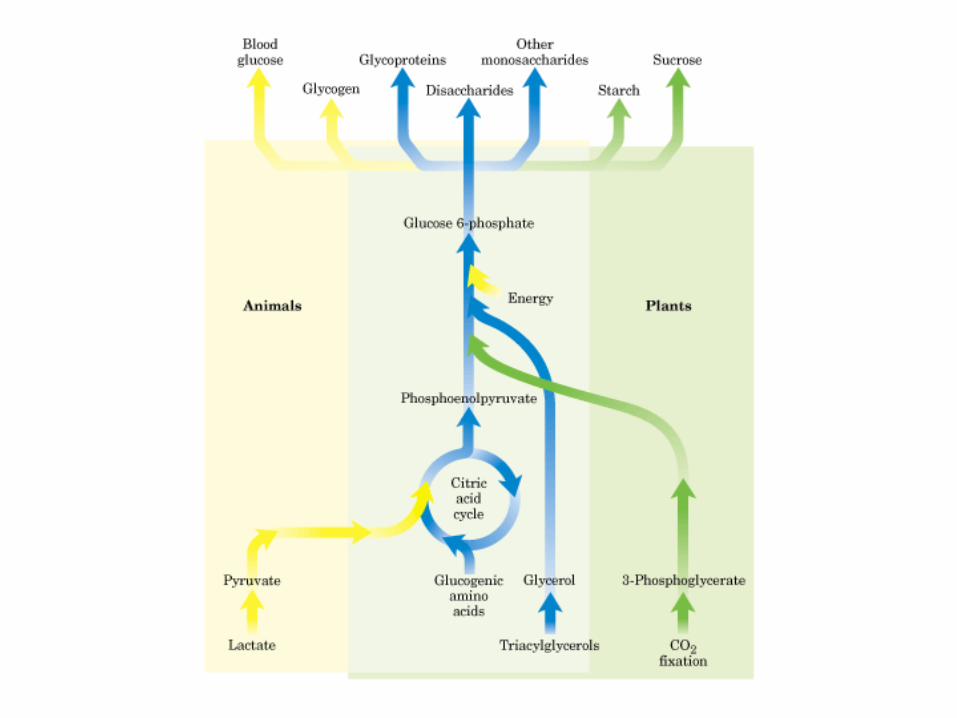
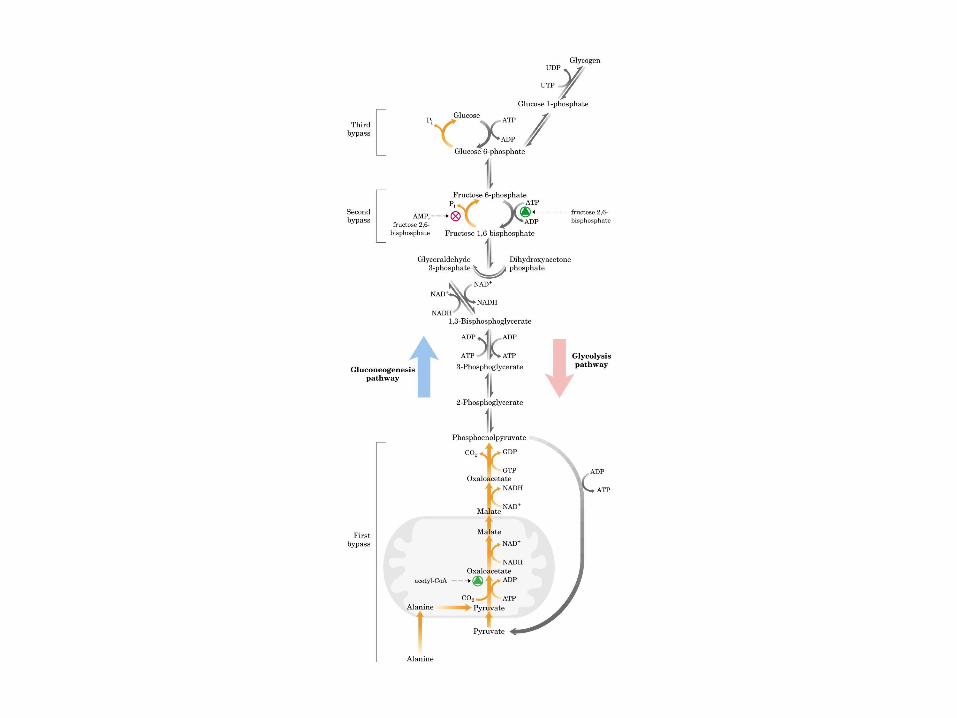
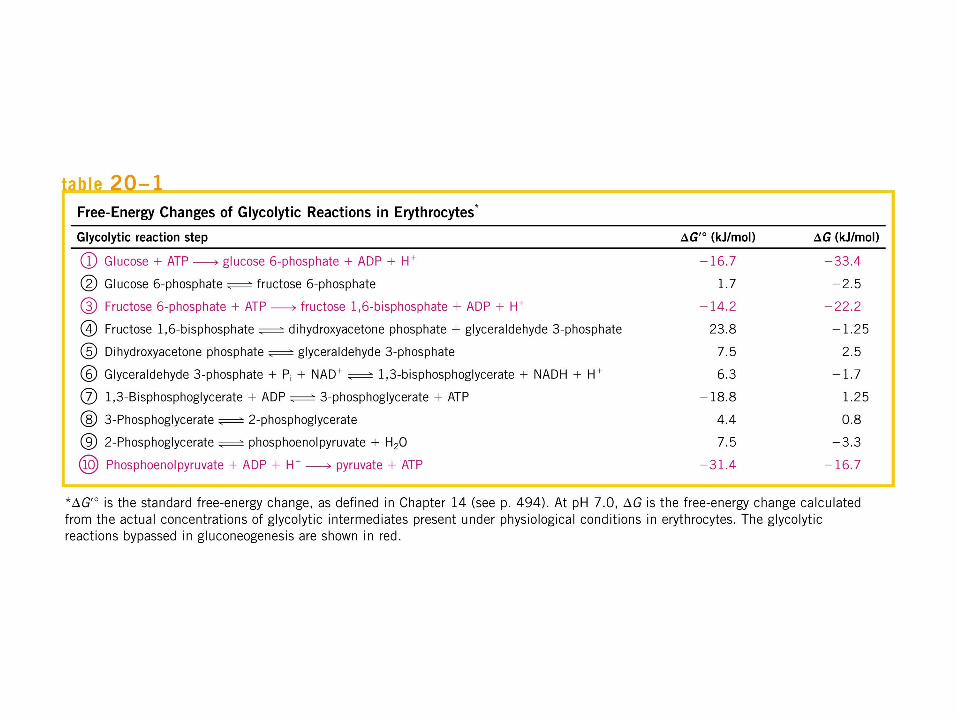
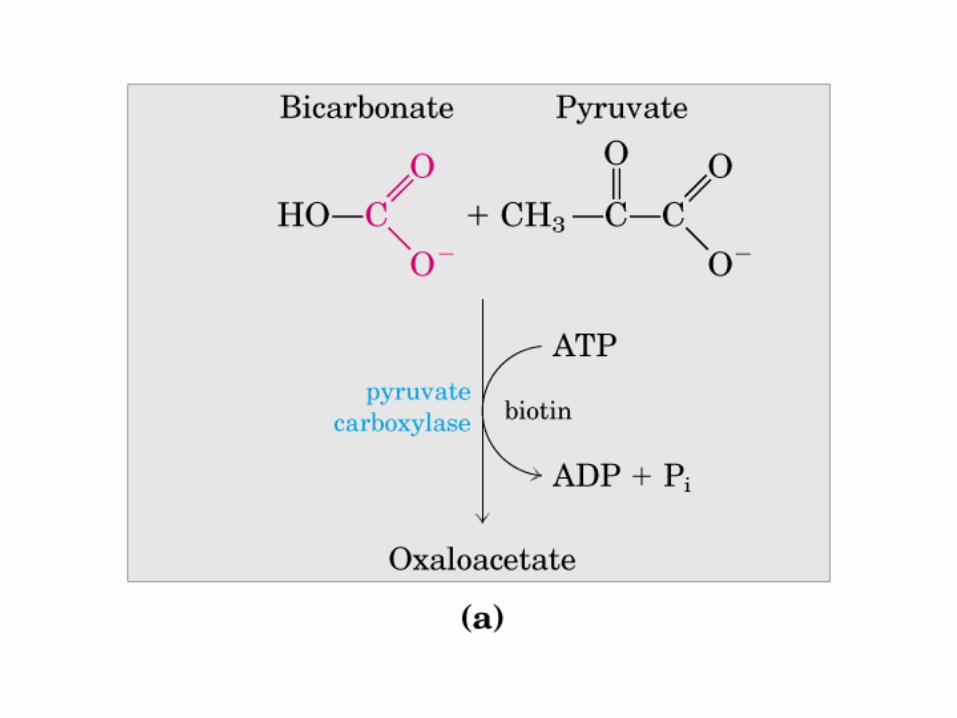
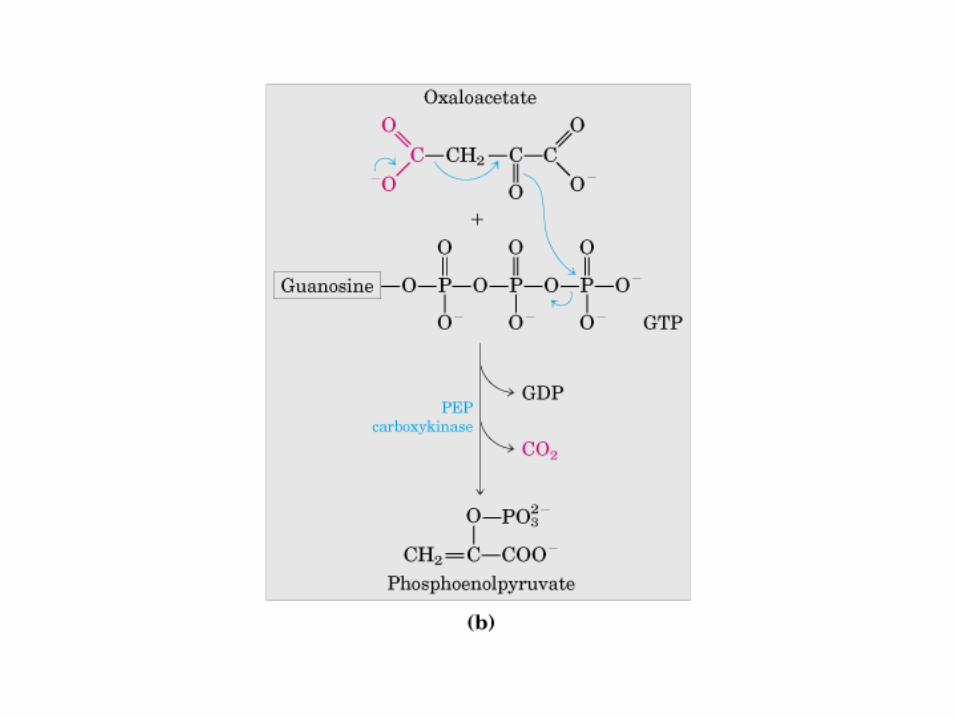
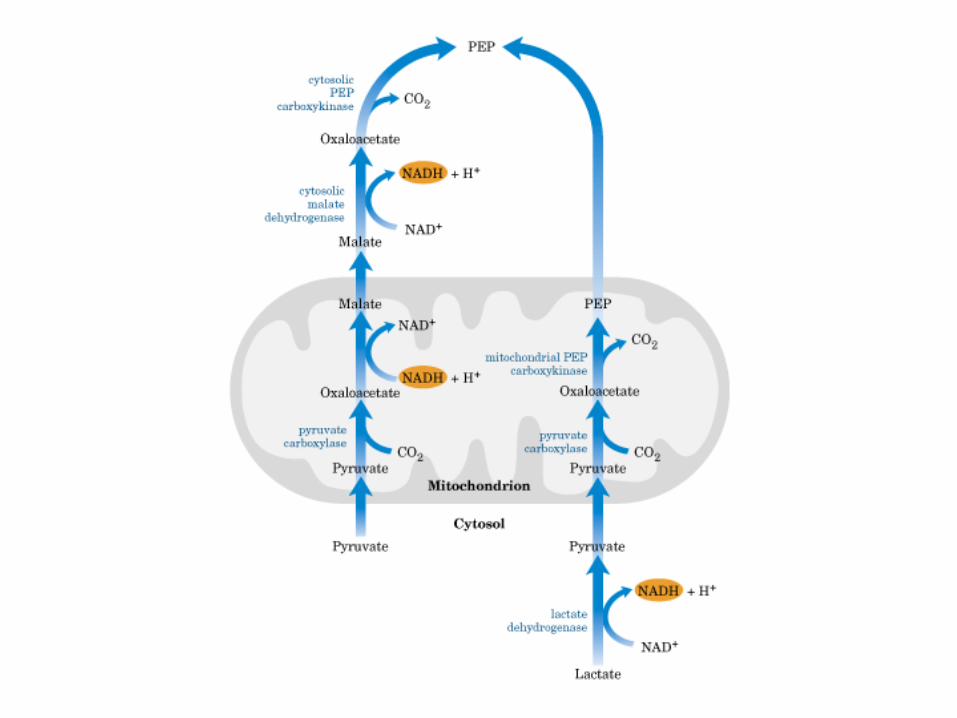


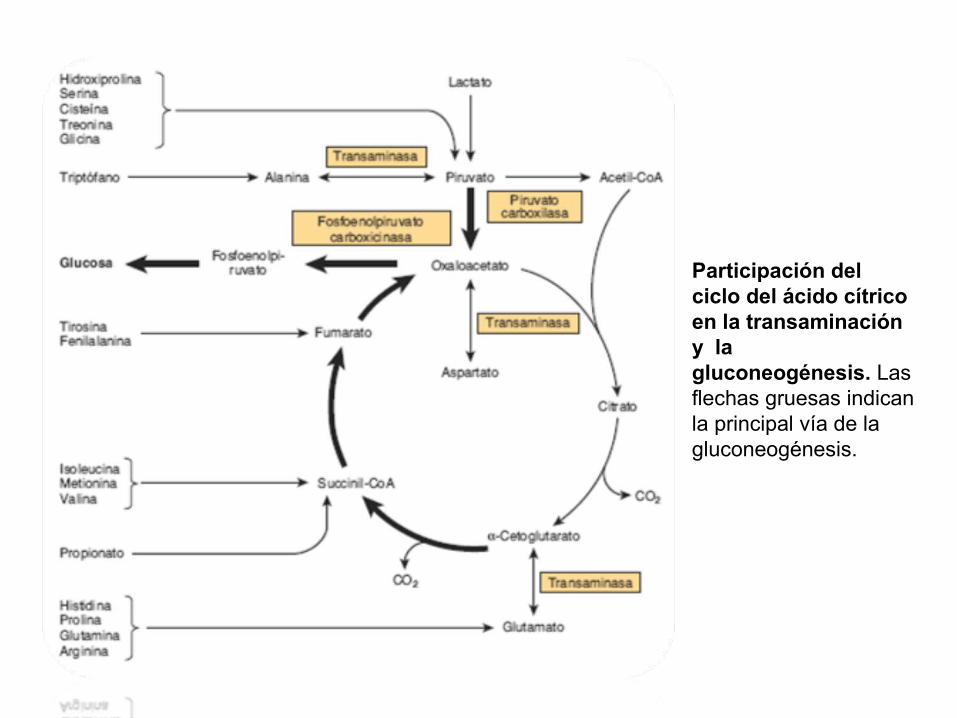
Participación del ciclo del ácido cítrico en la transaminación y la gluconeogénesis. Las flechas gruesas indican la principal vía de la gluconeogénesis.
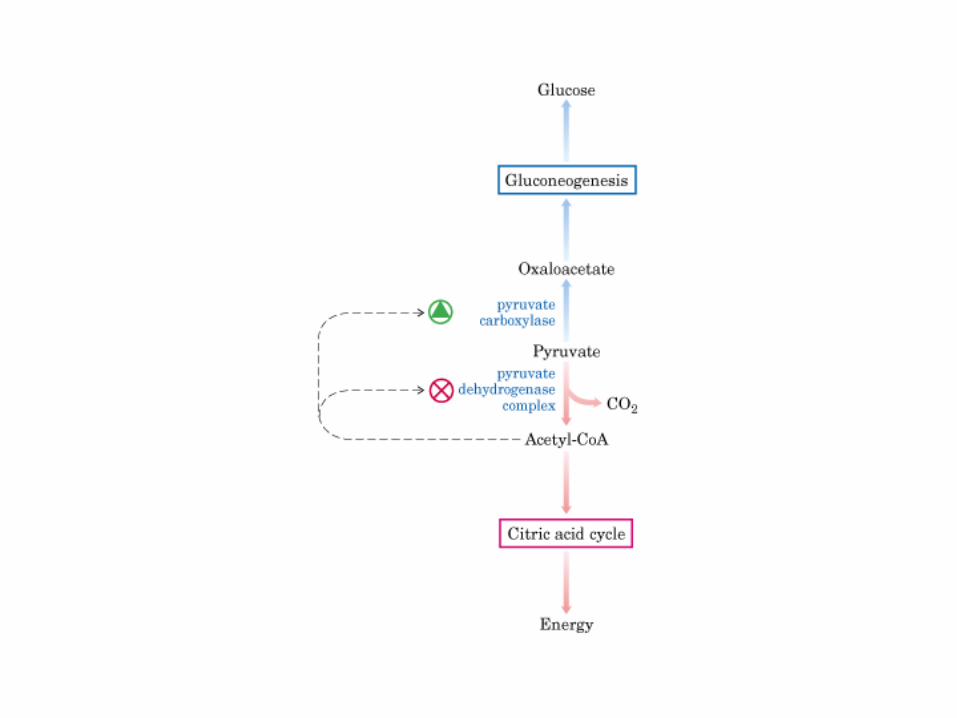
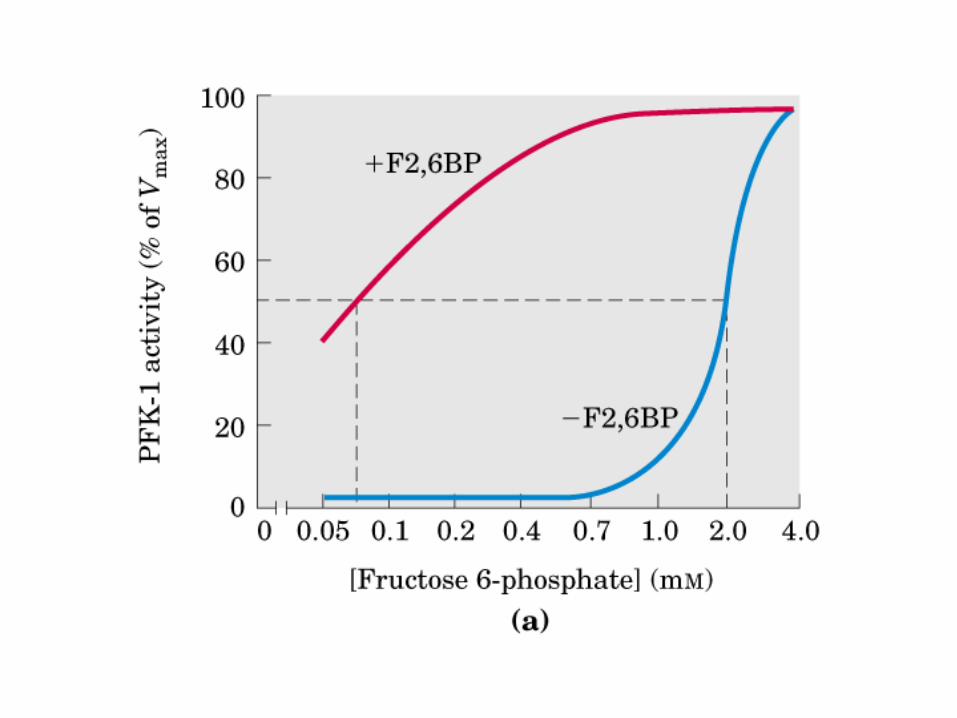
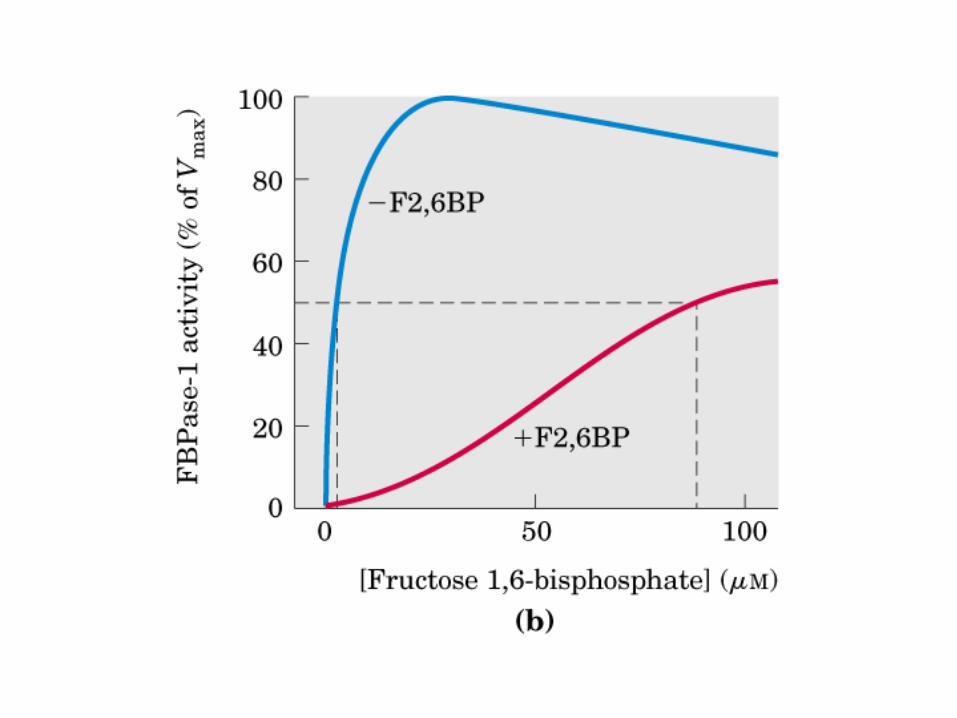
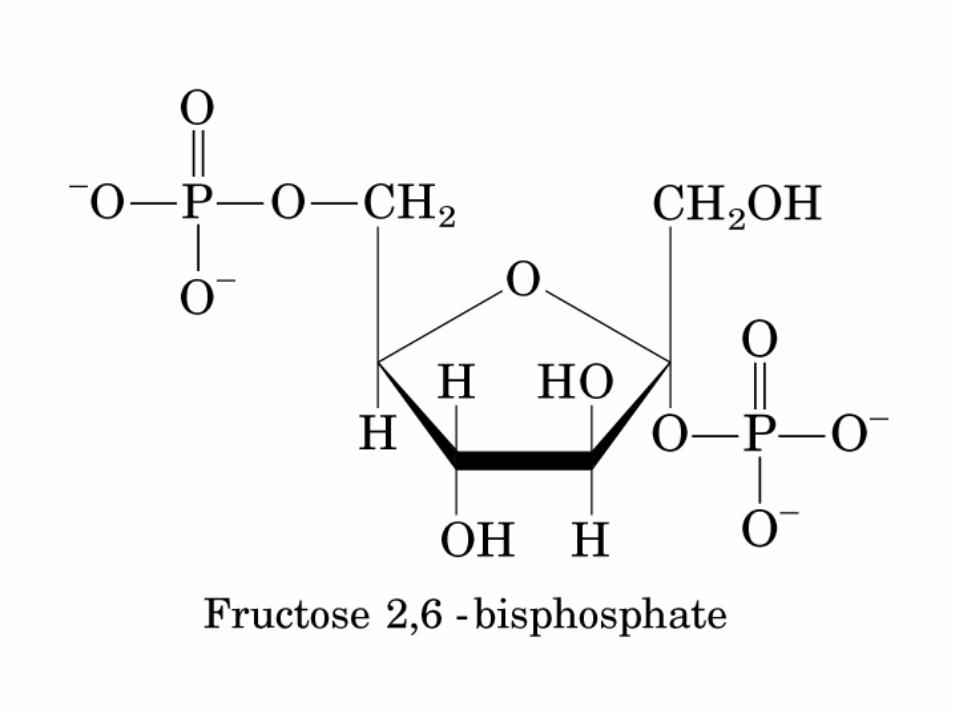
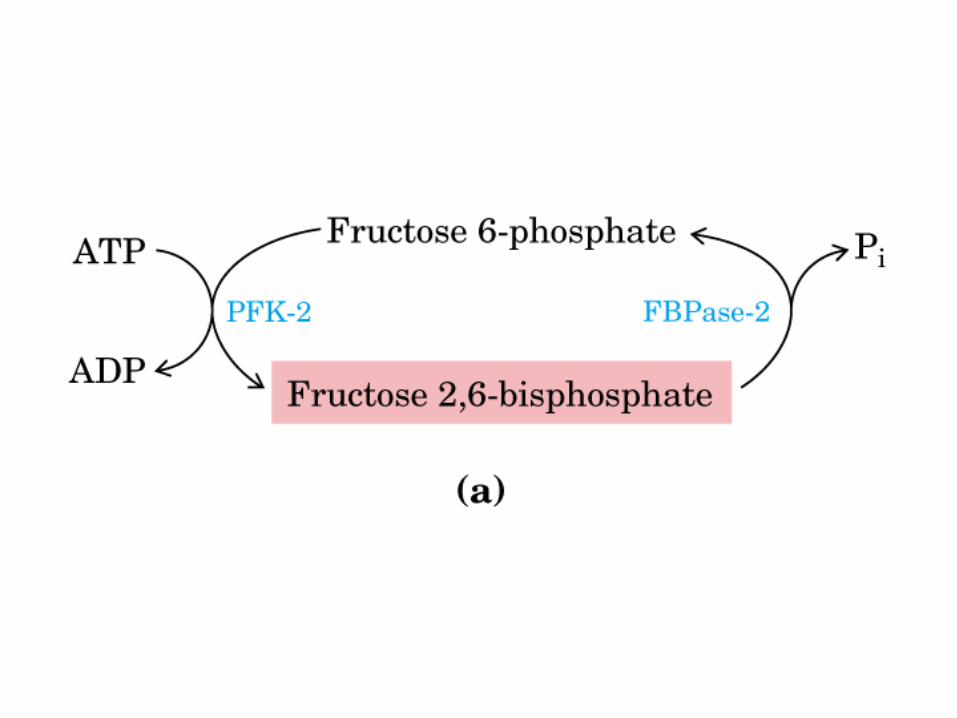
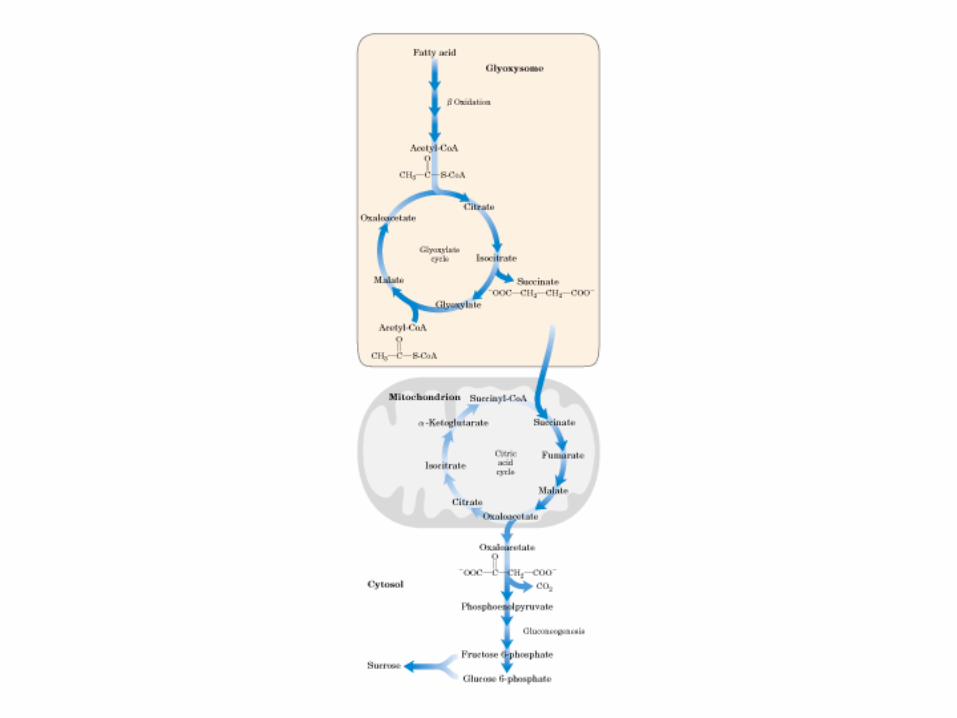
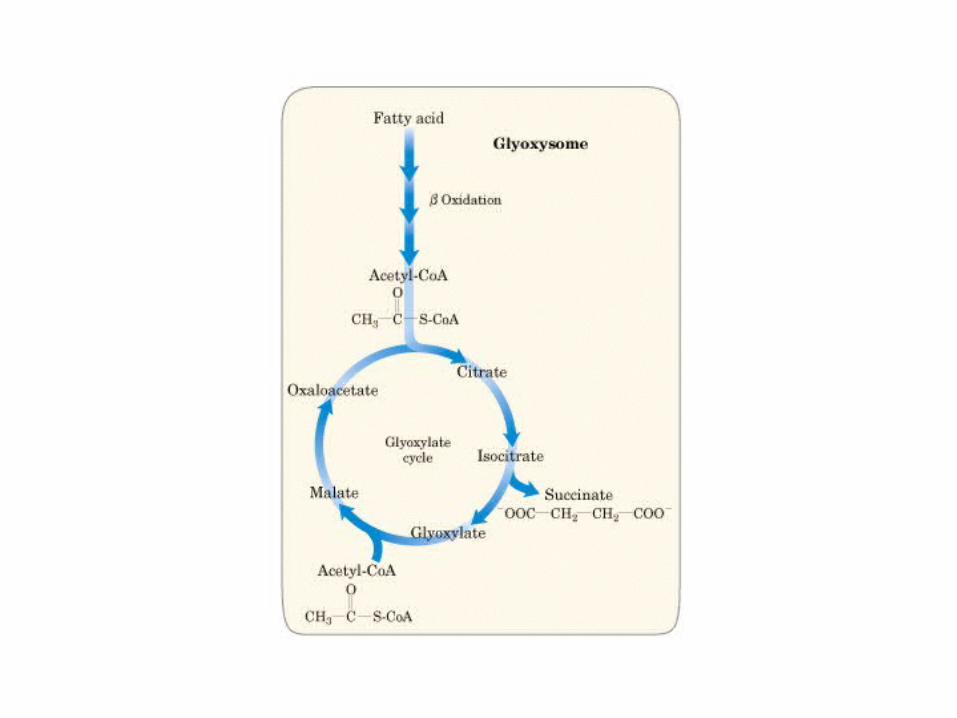
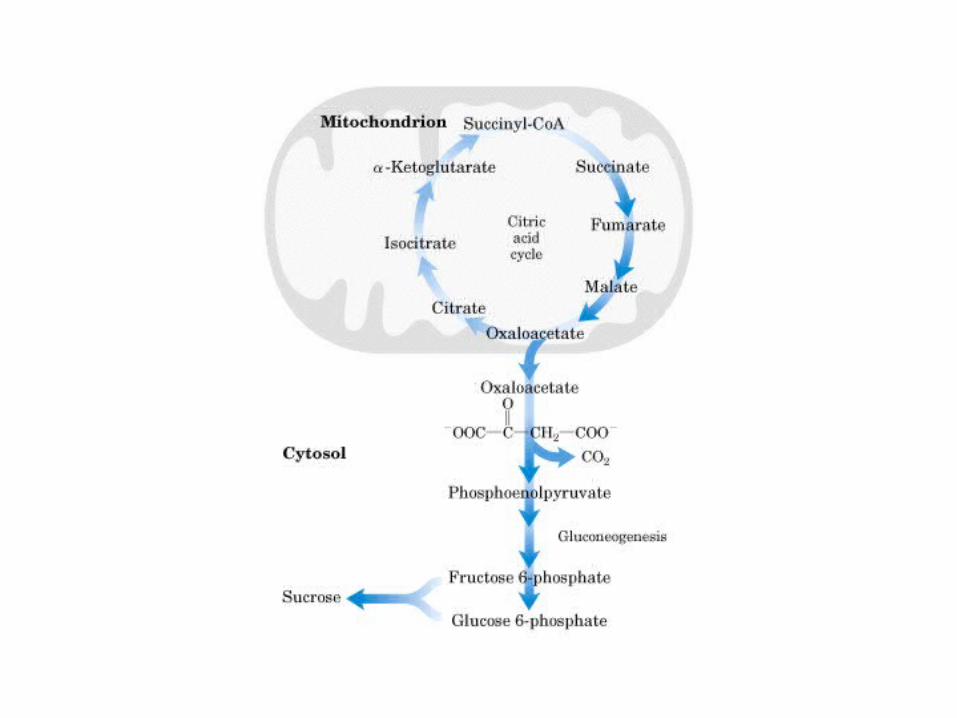
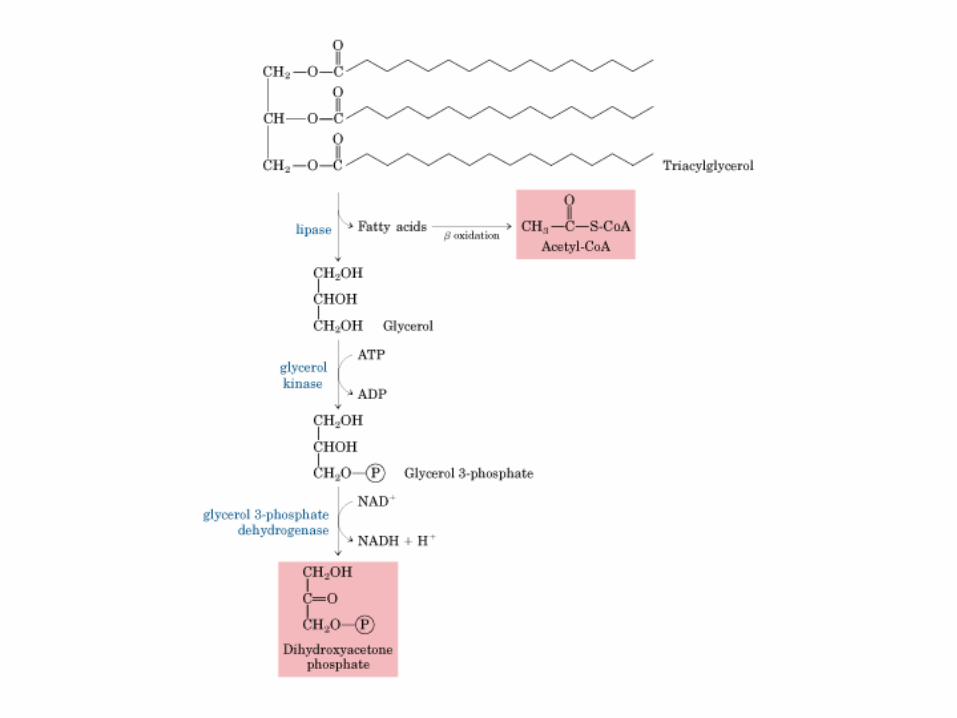
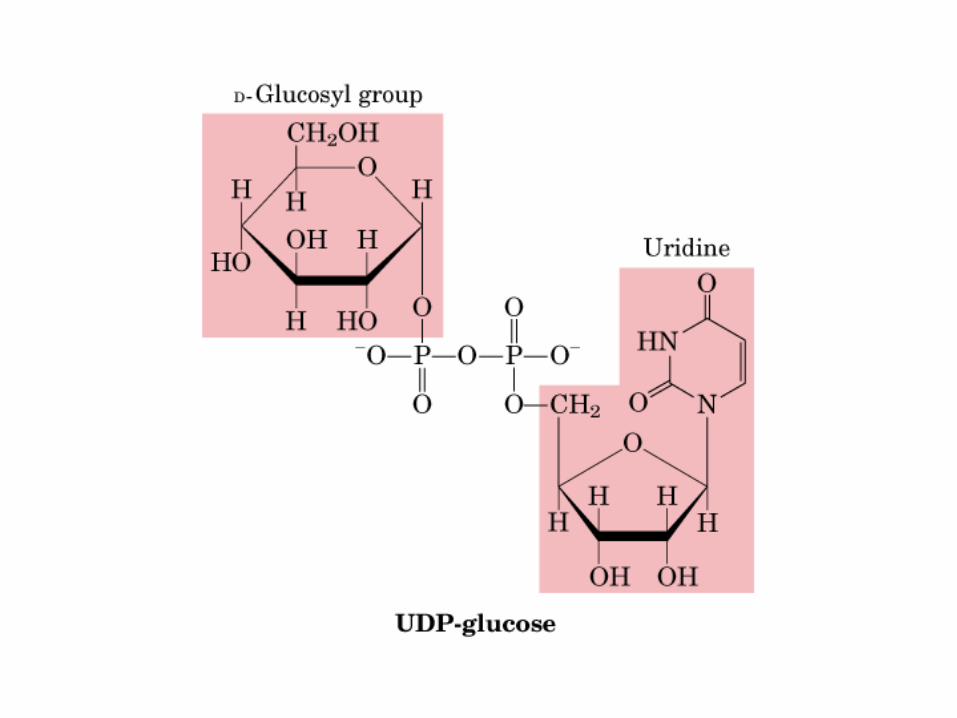
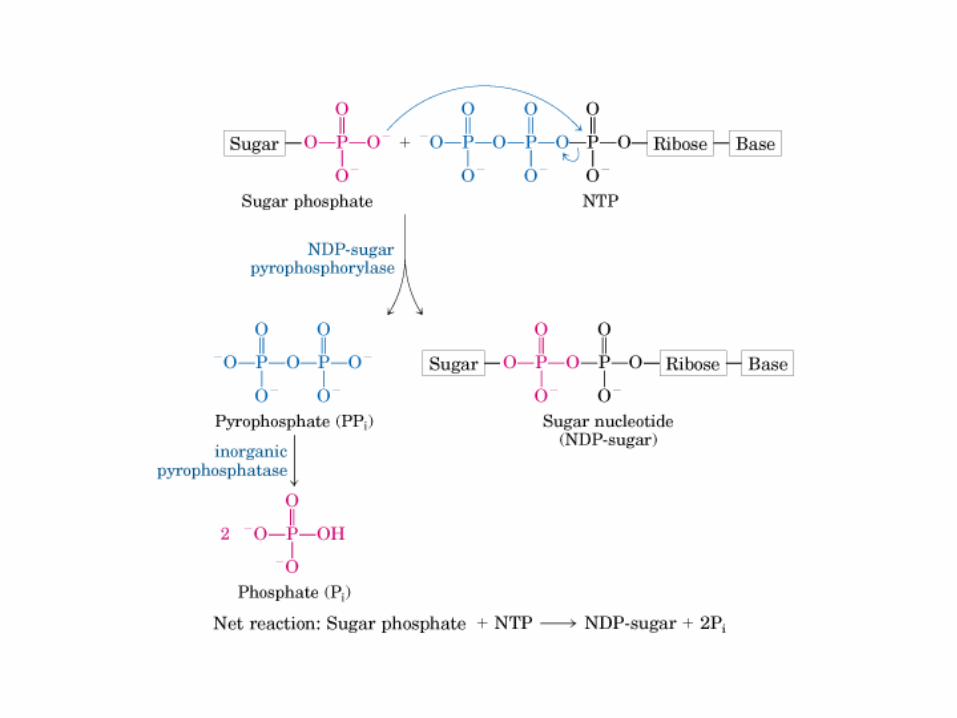
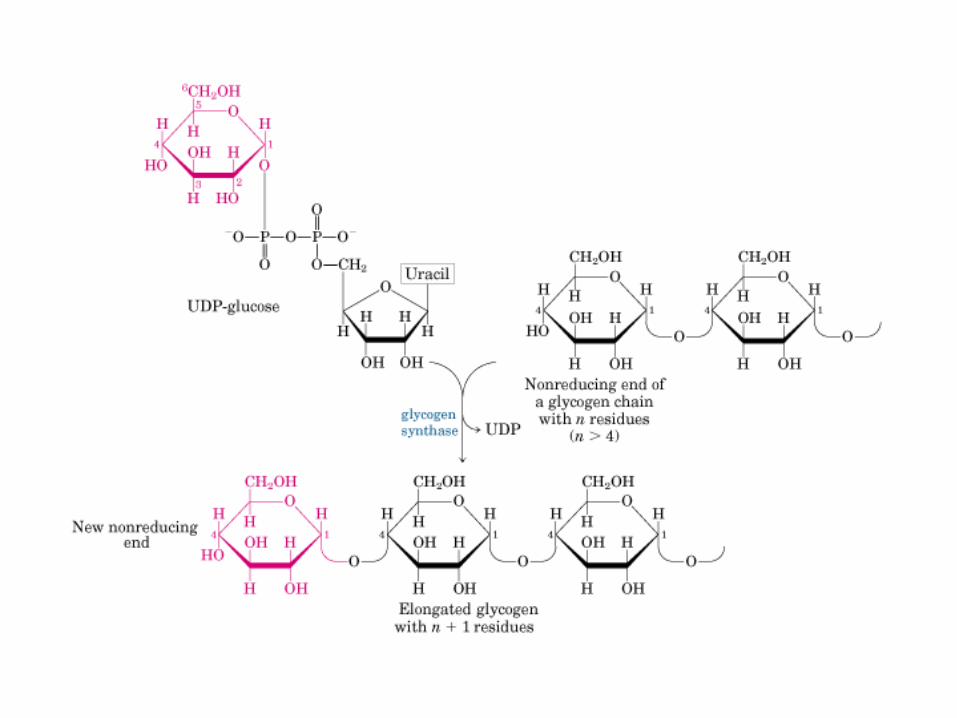
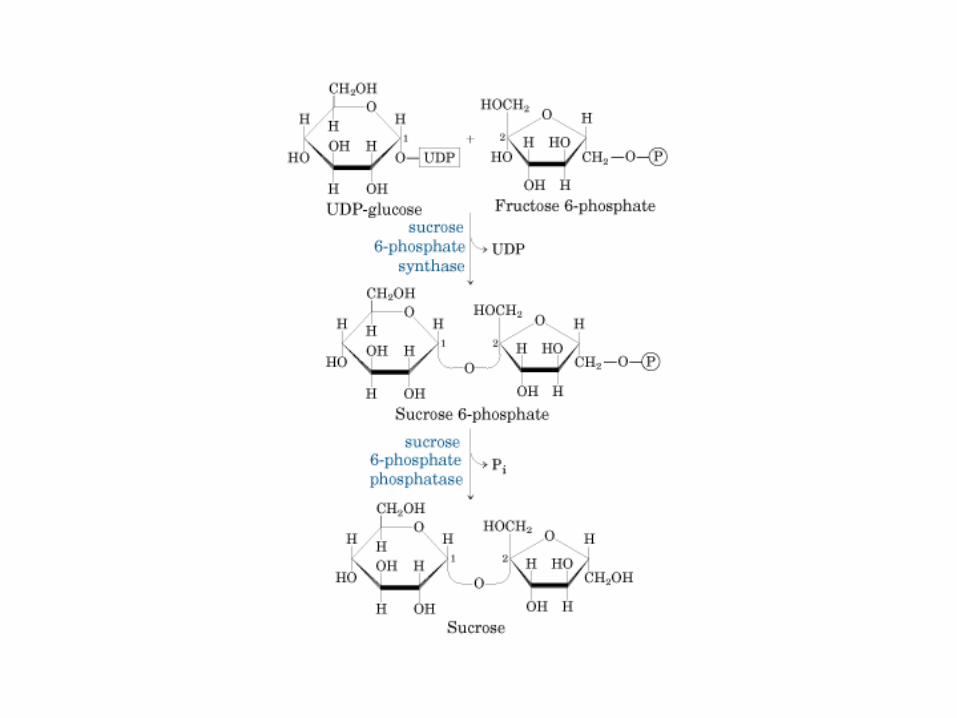
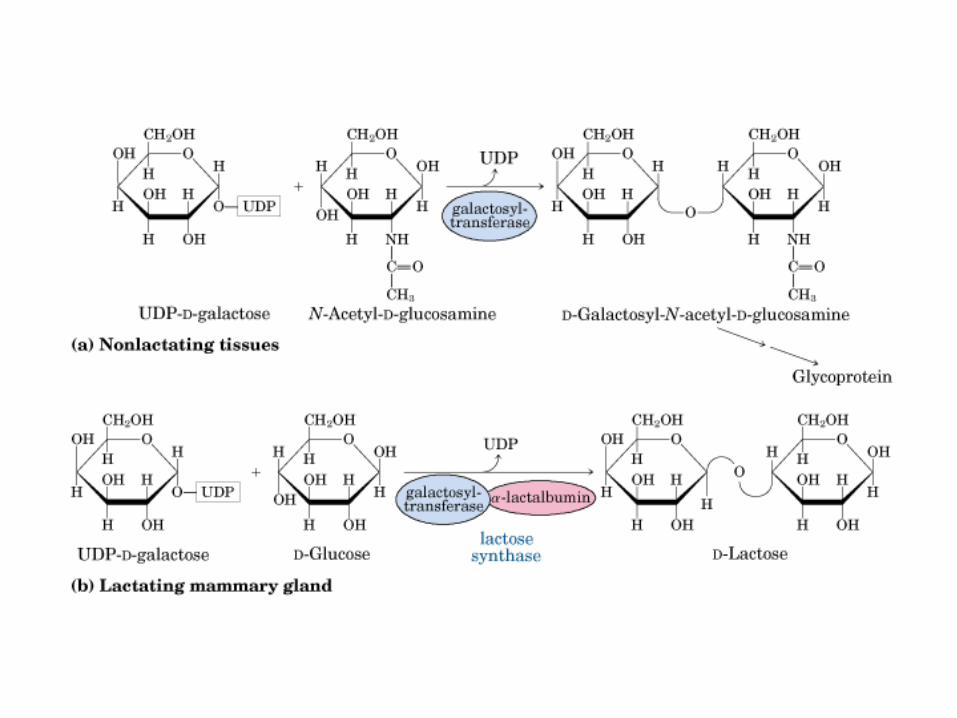
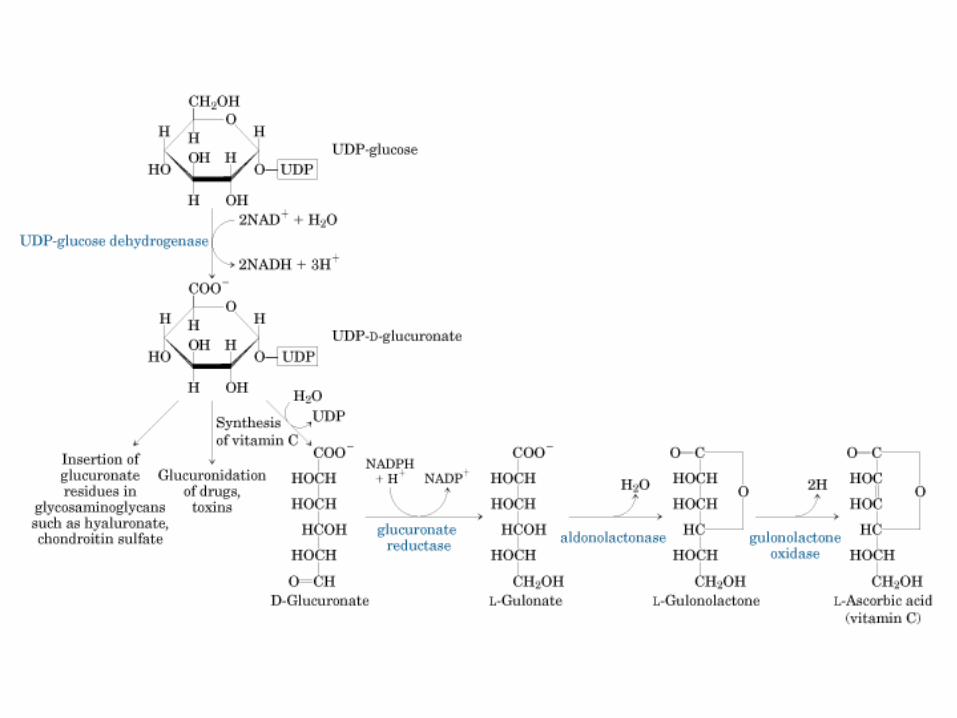
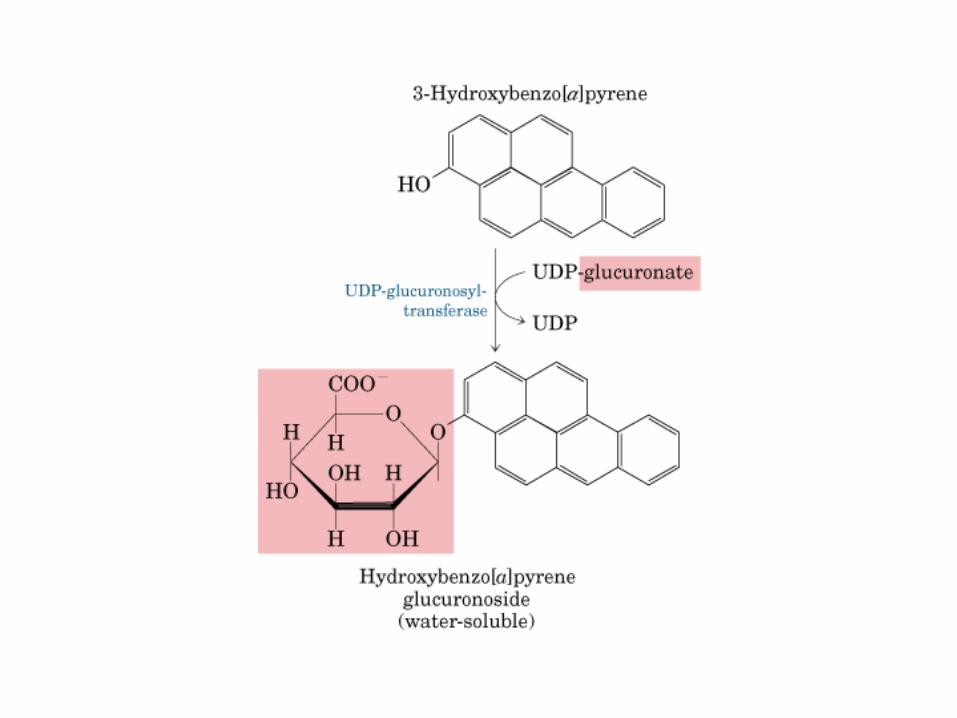
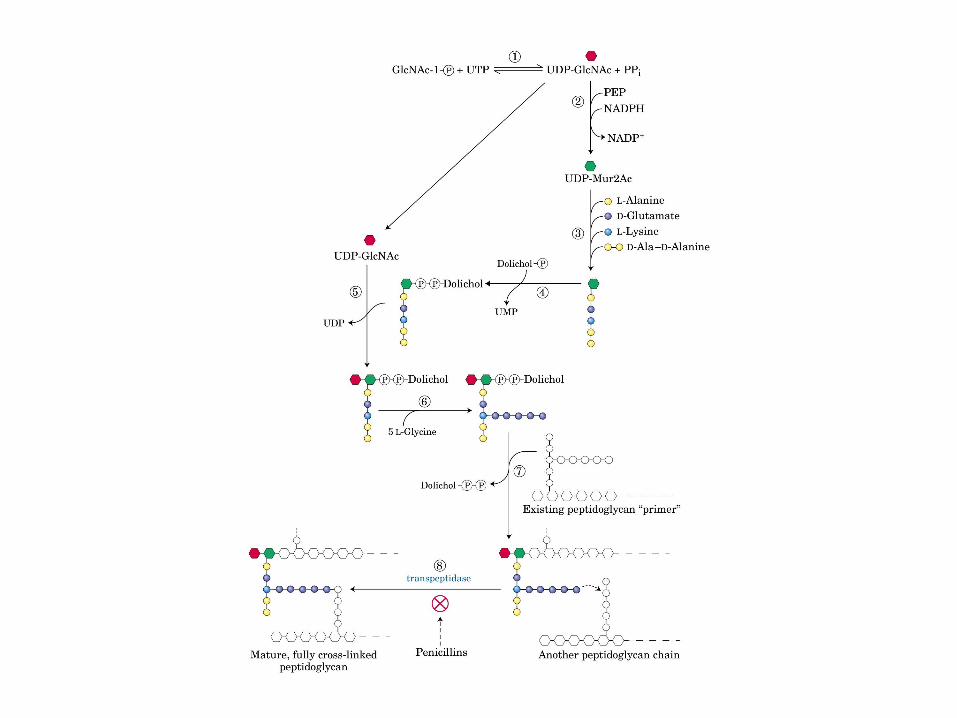
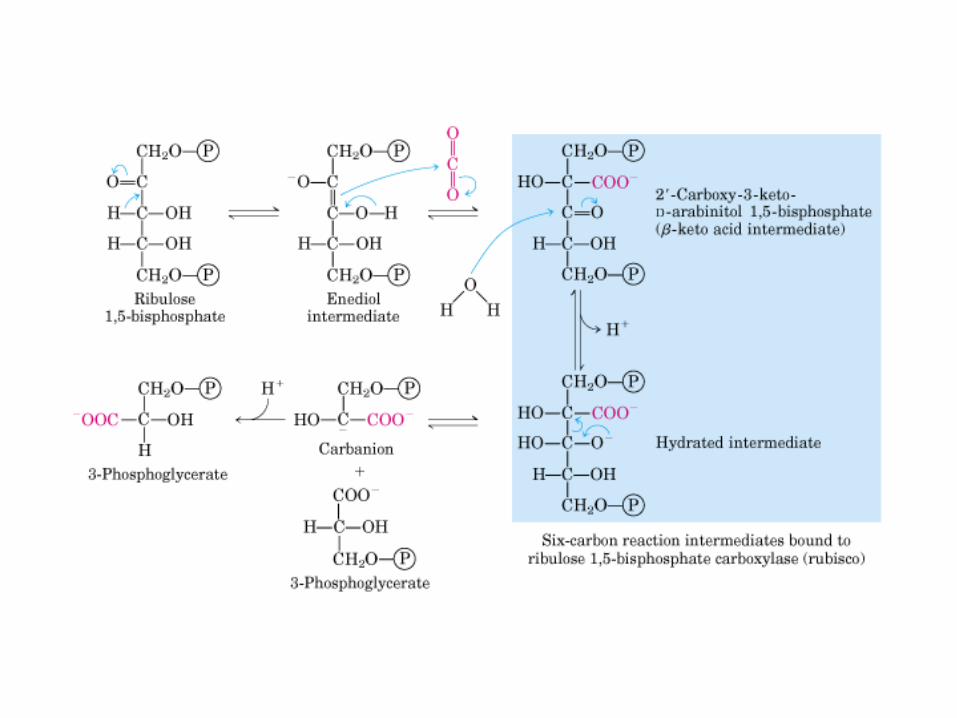
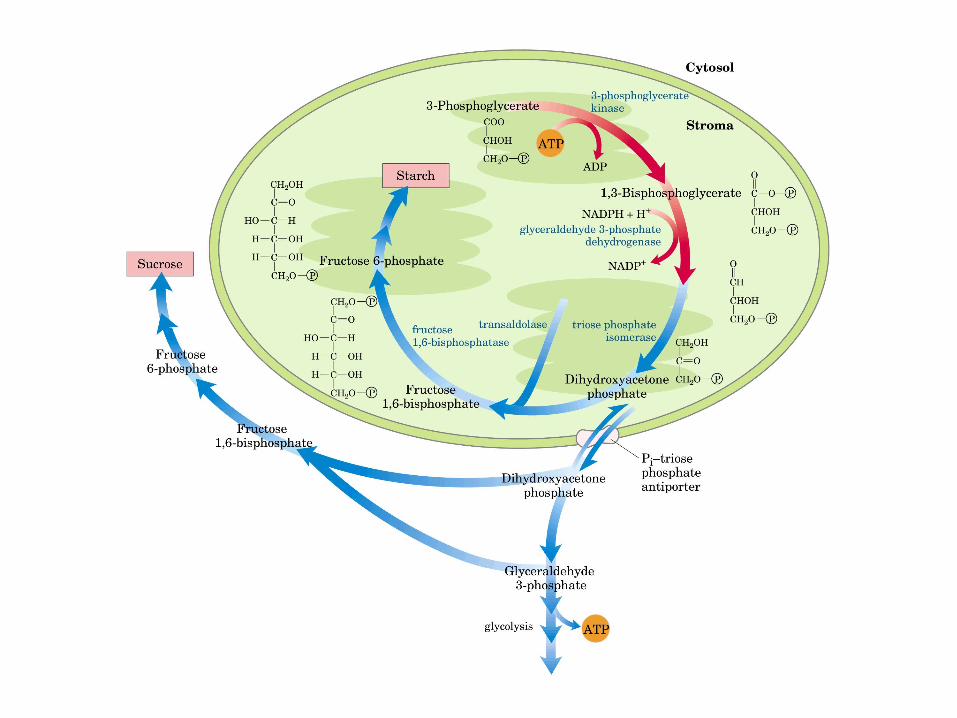
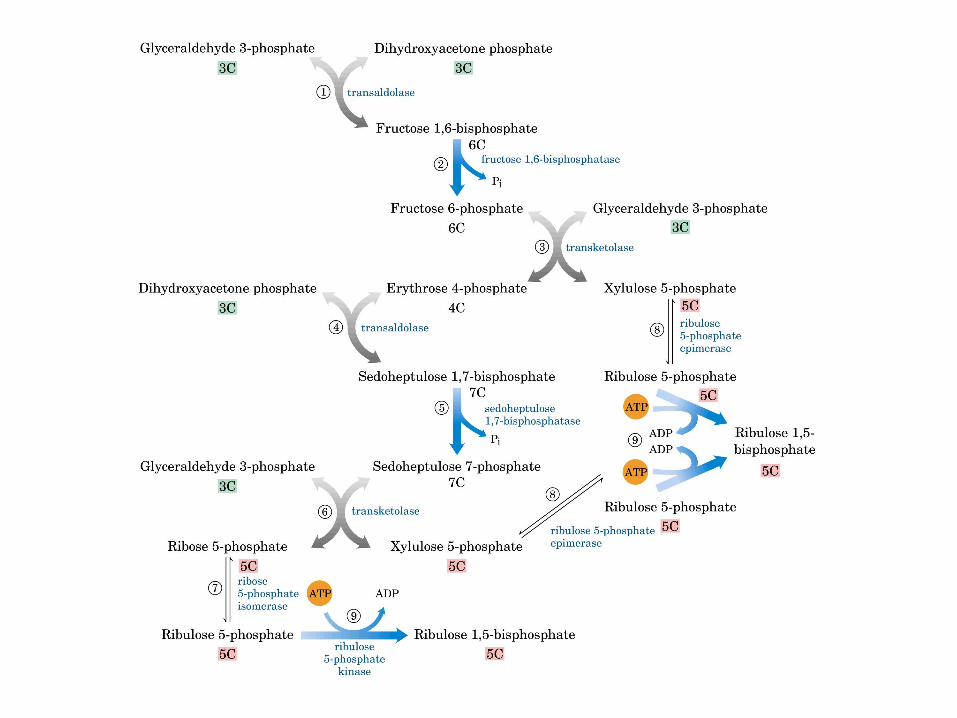
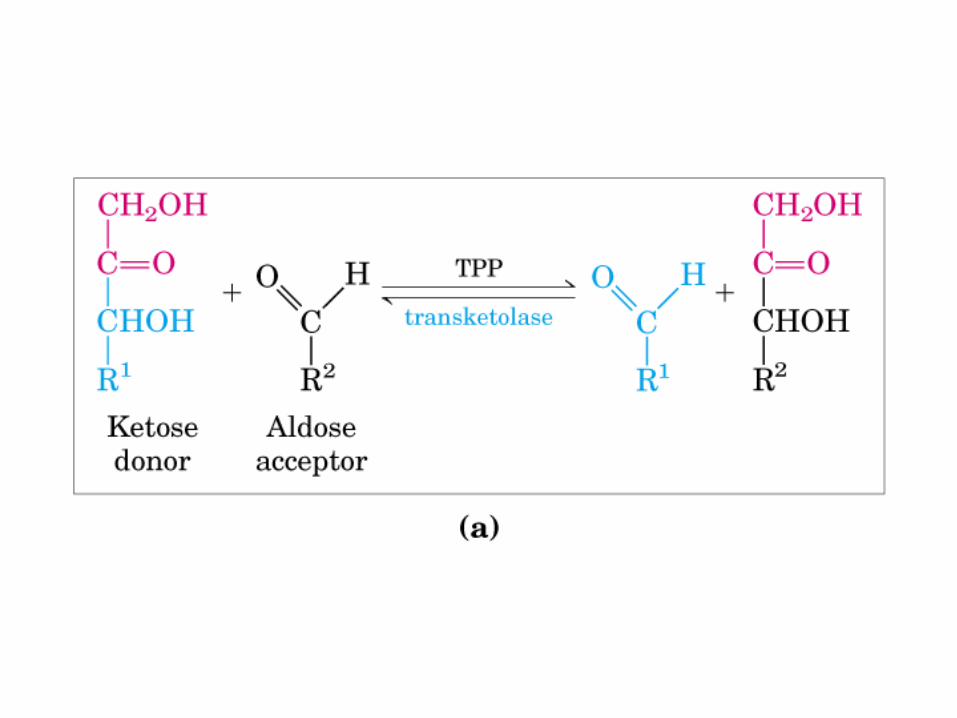
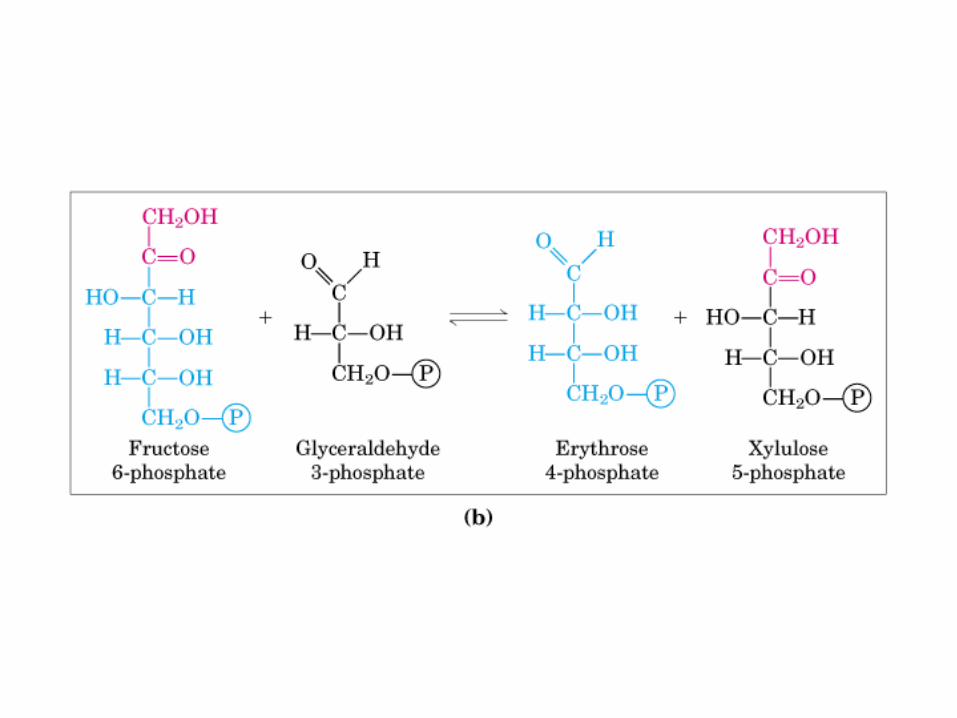
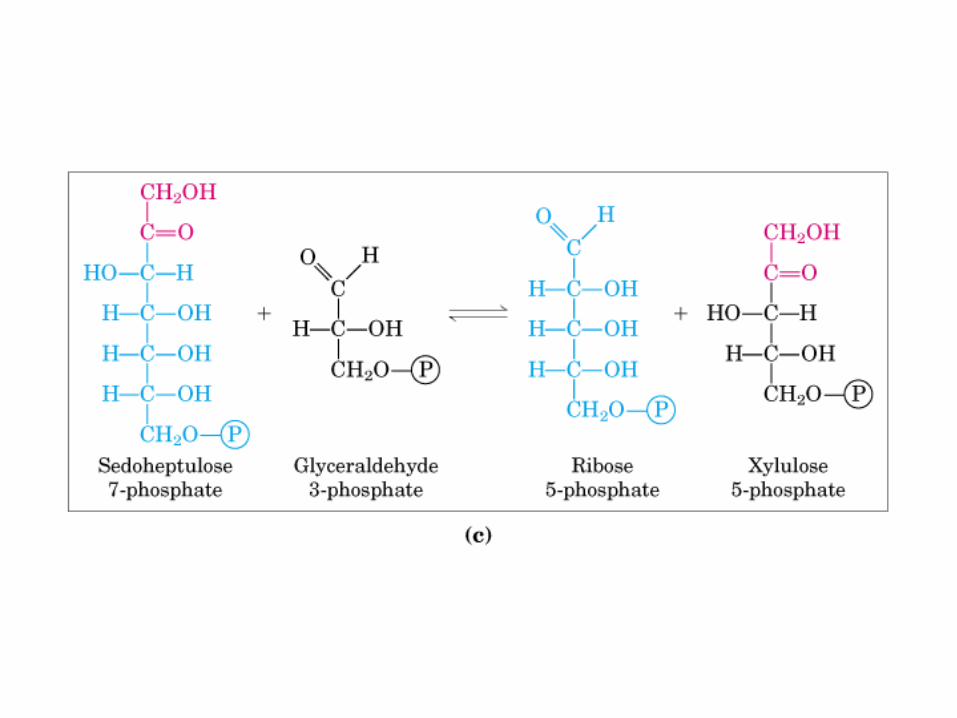
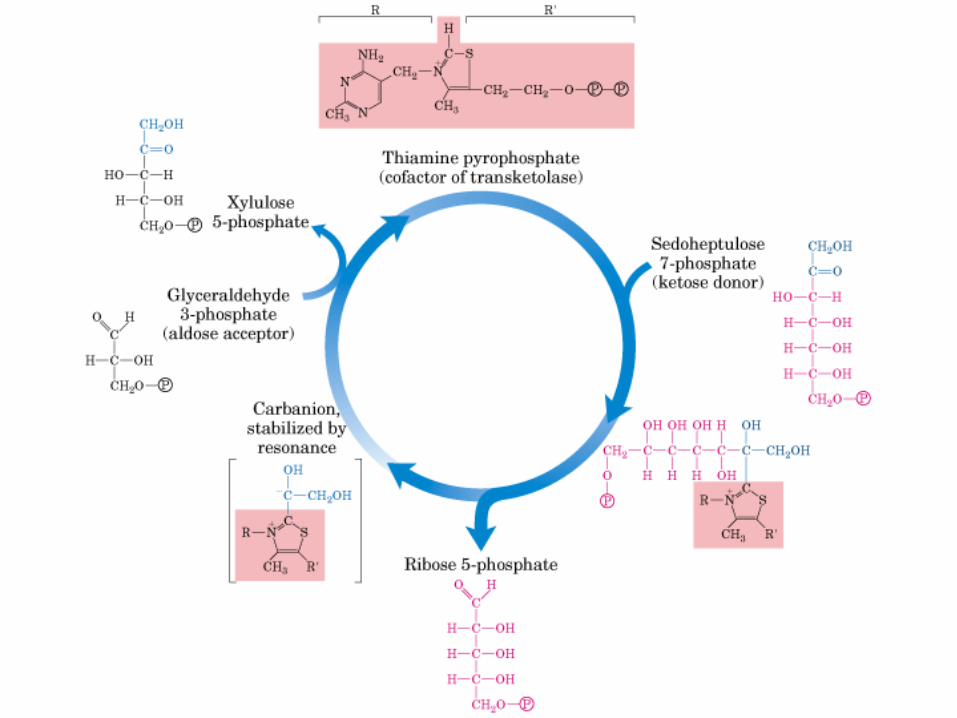
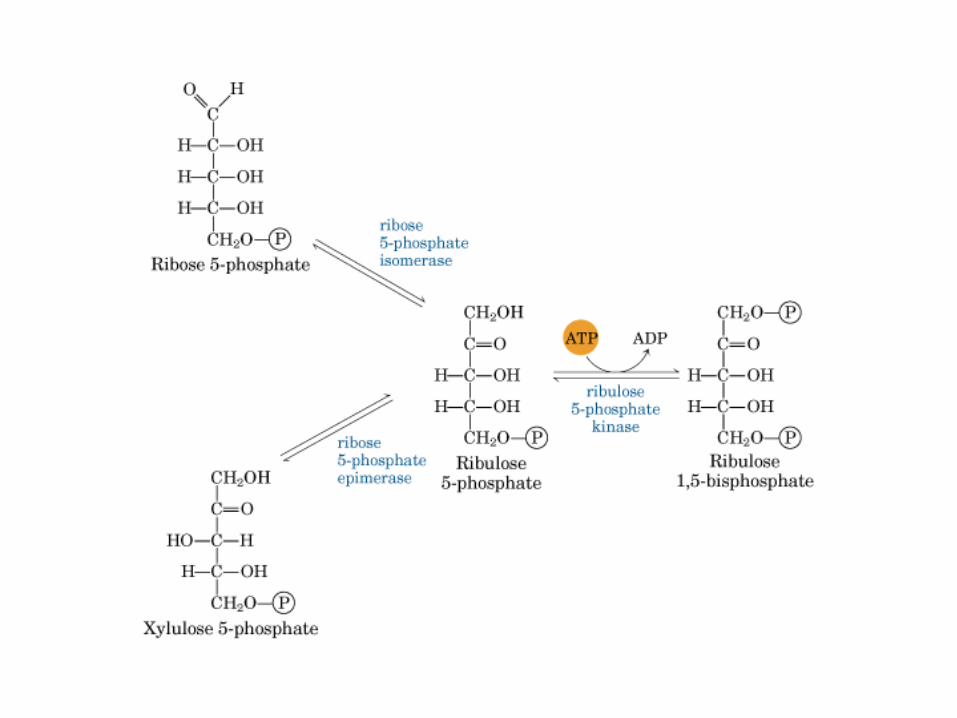
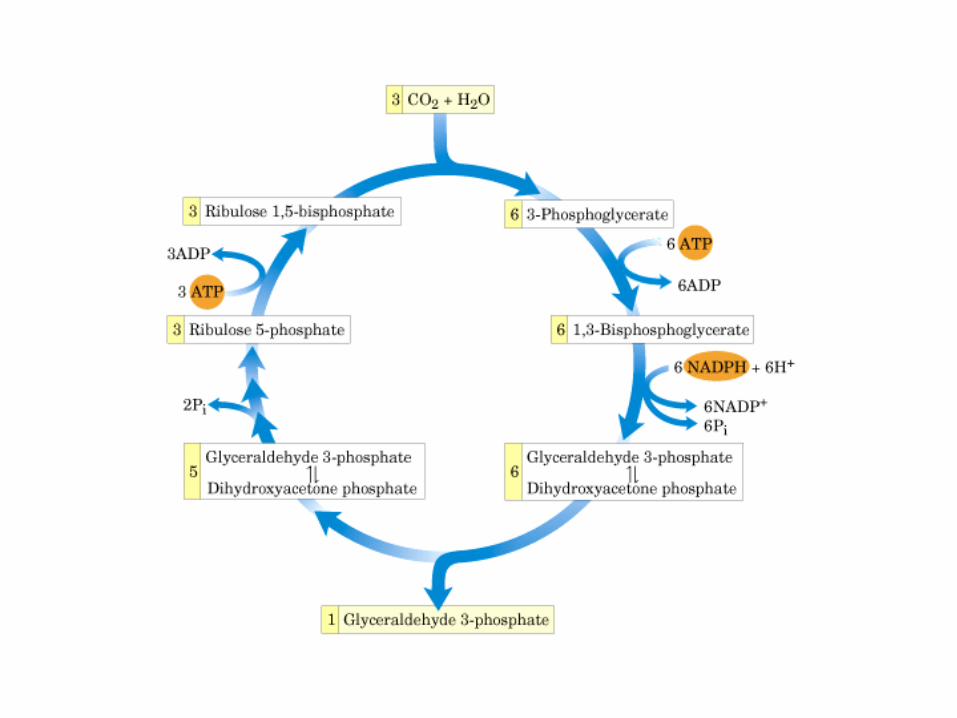
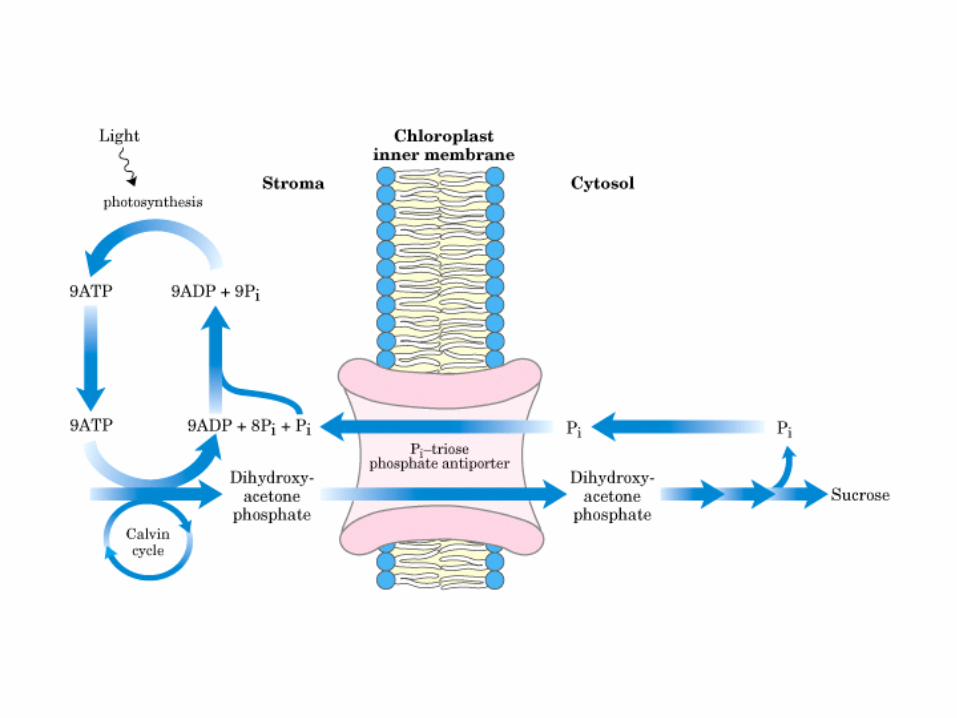
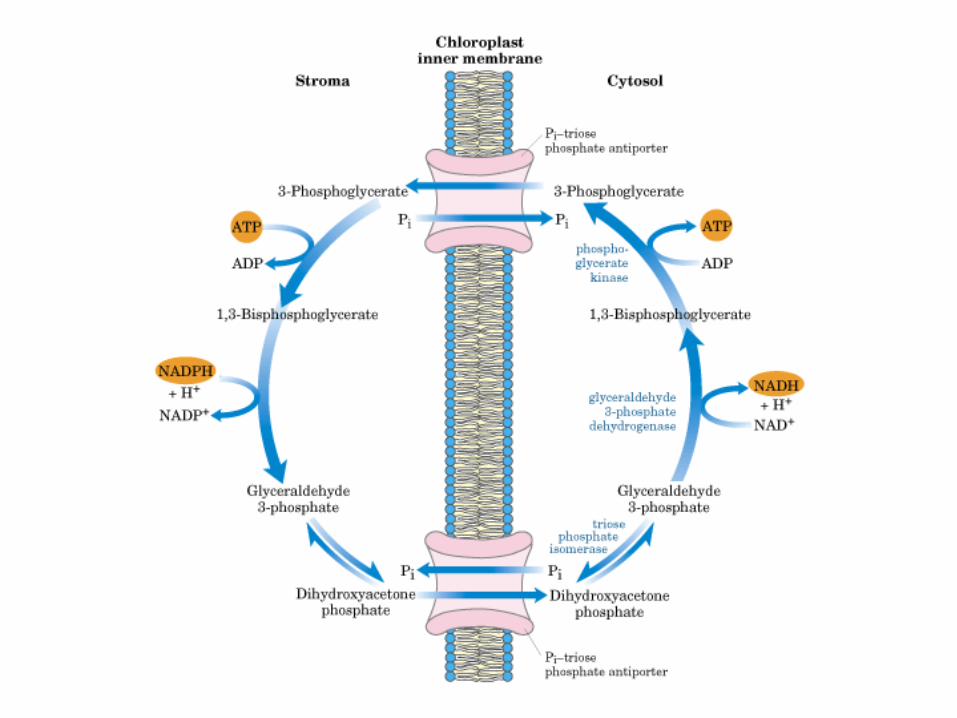
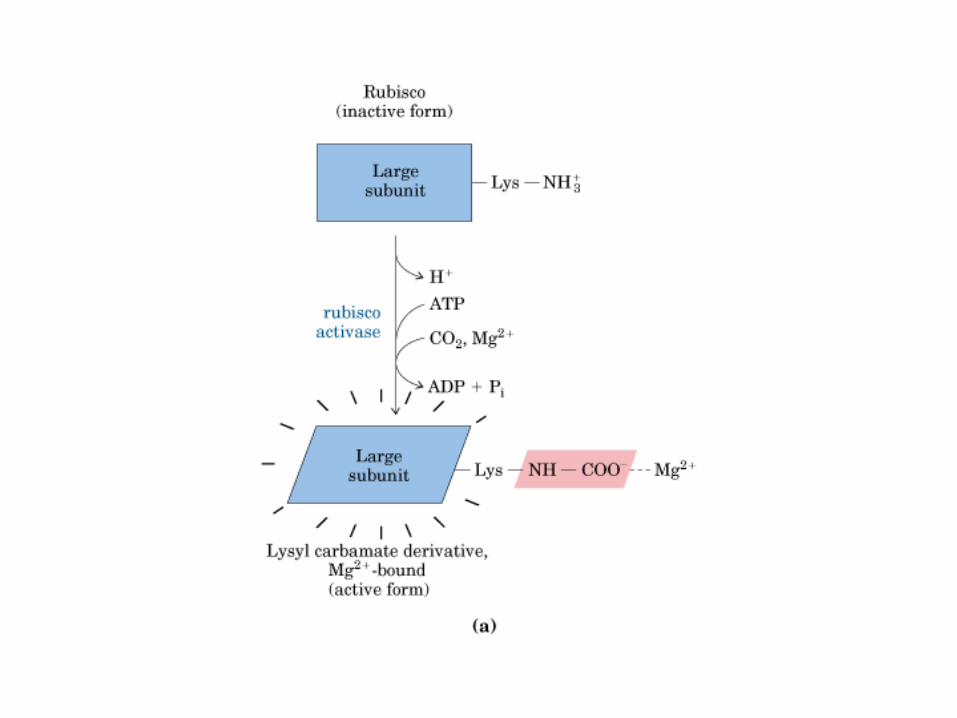
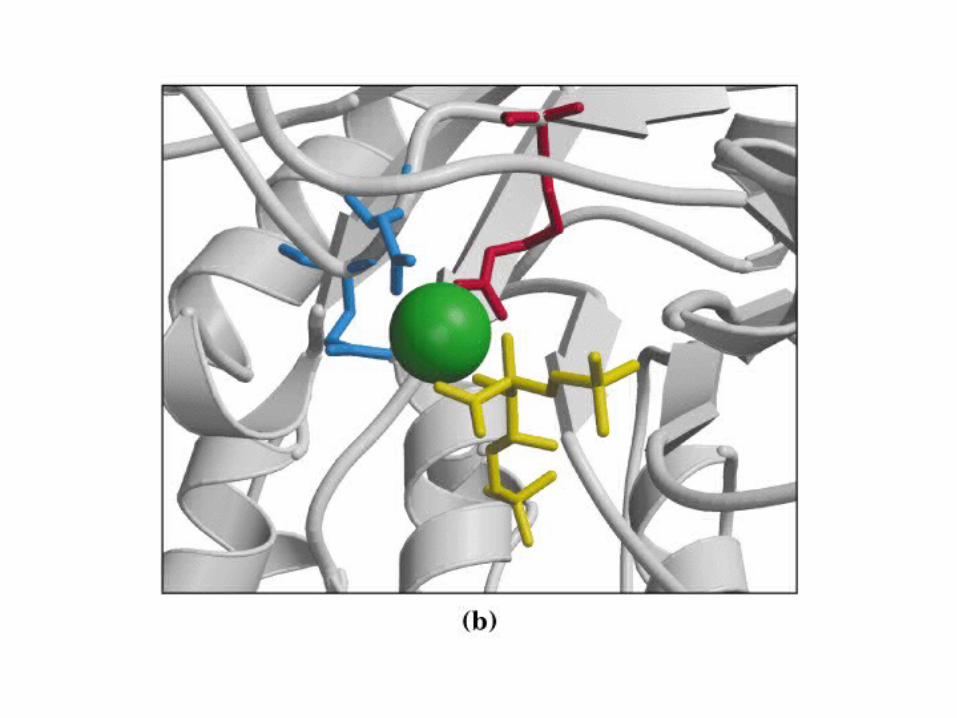
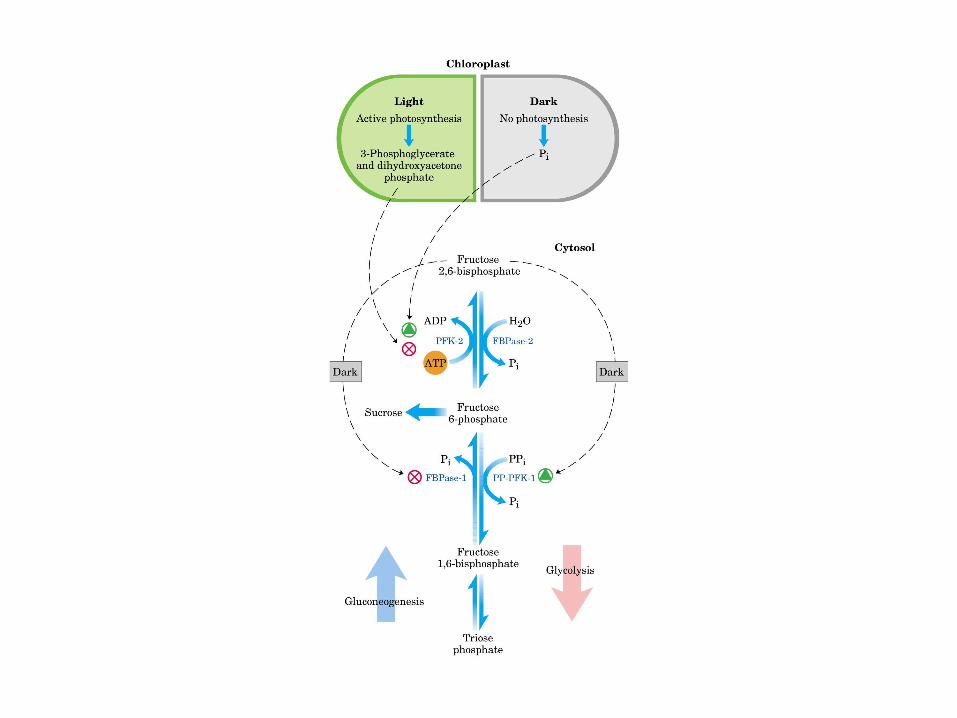
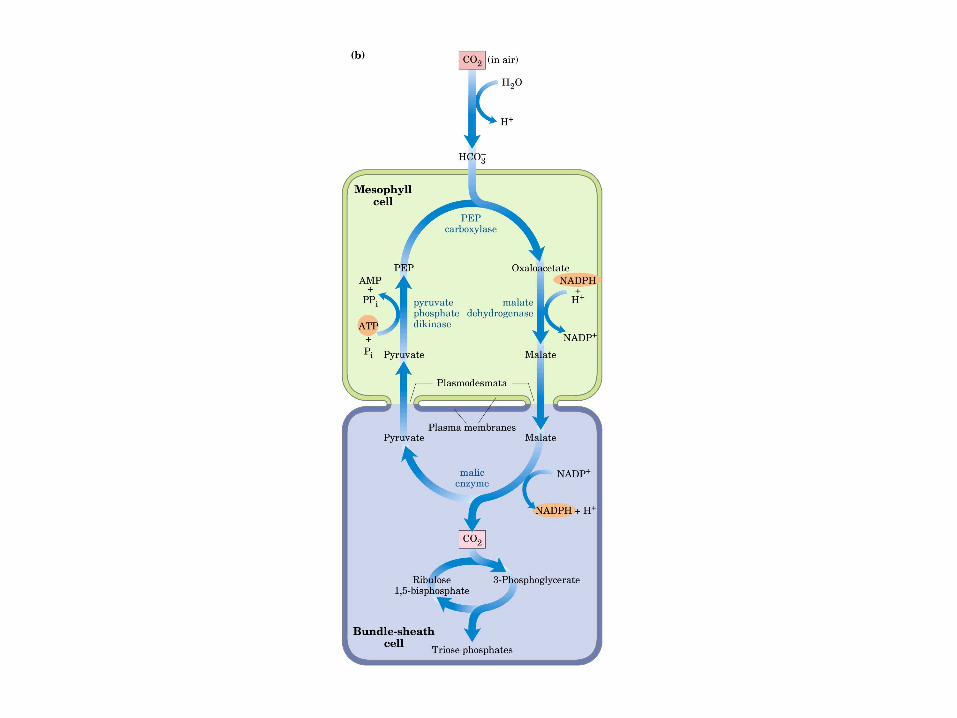

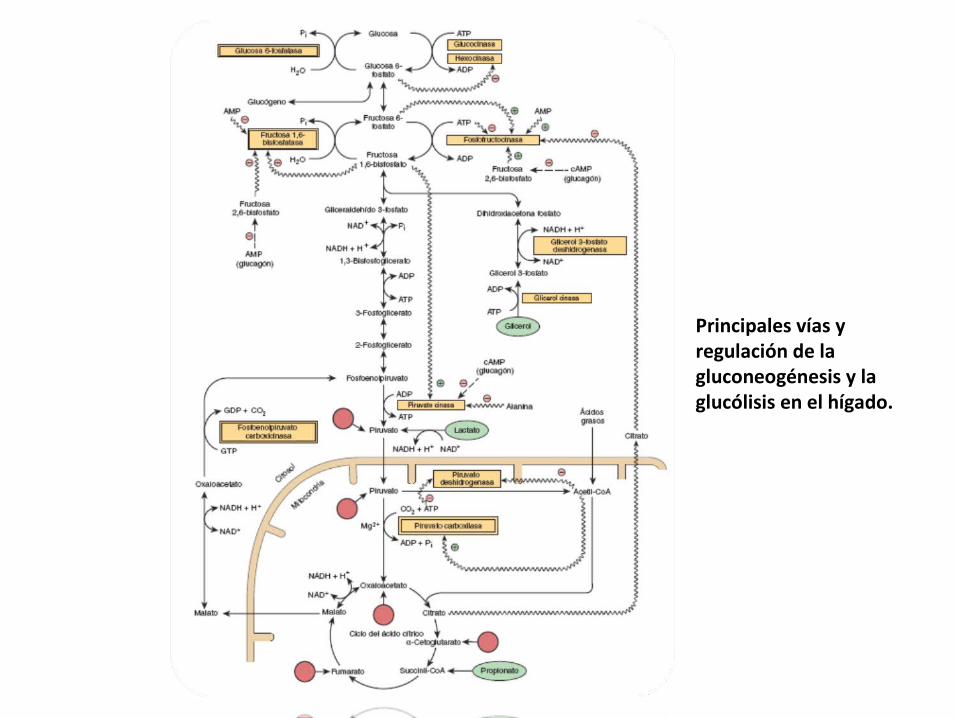
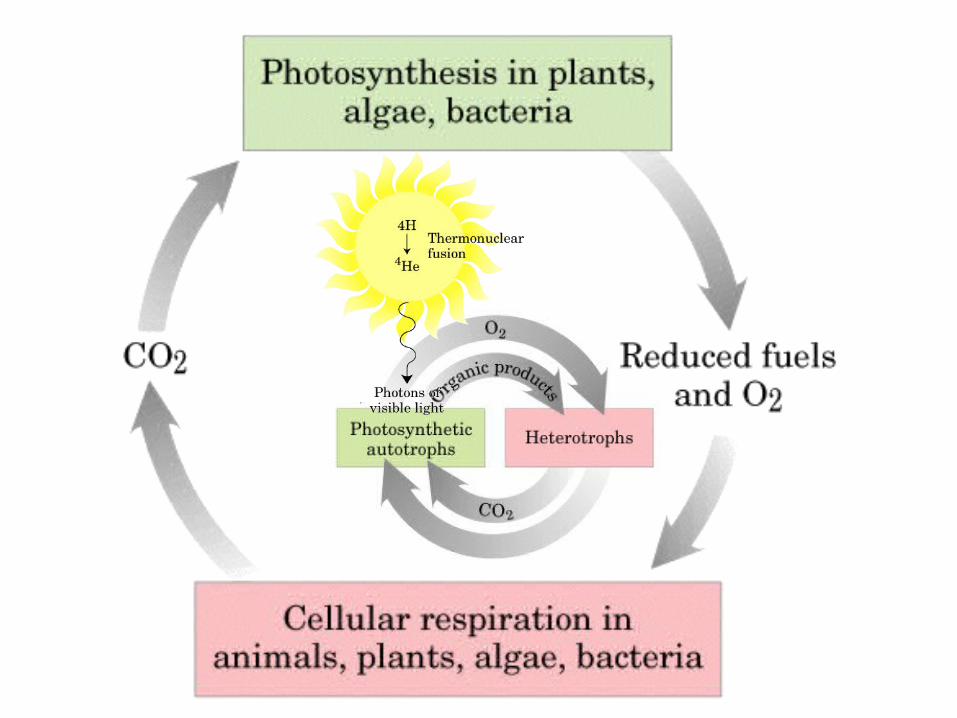
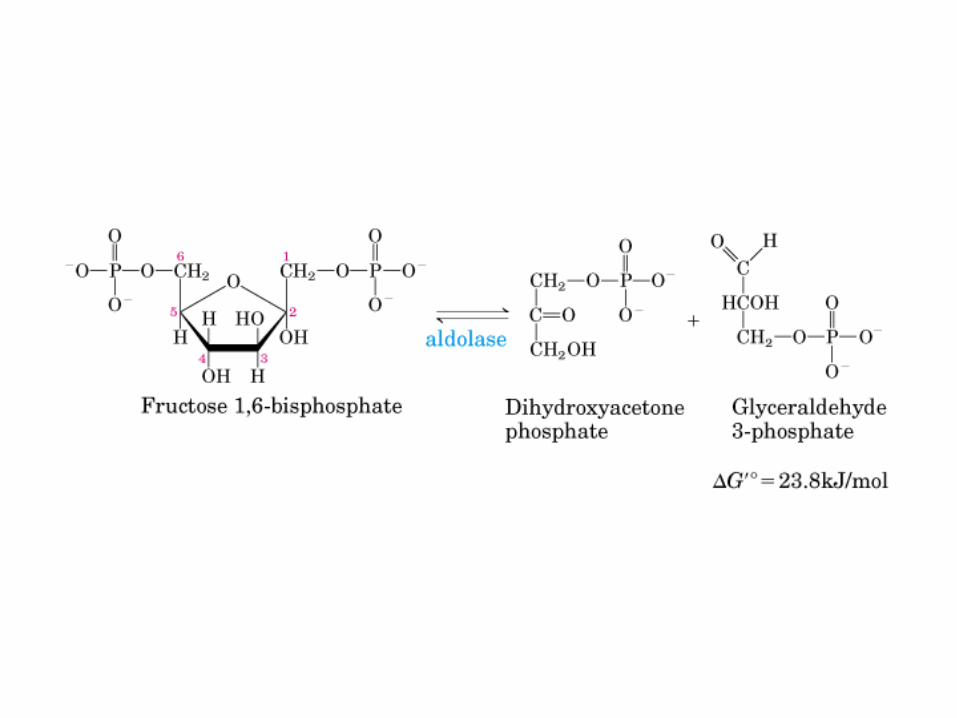
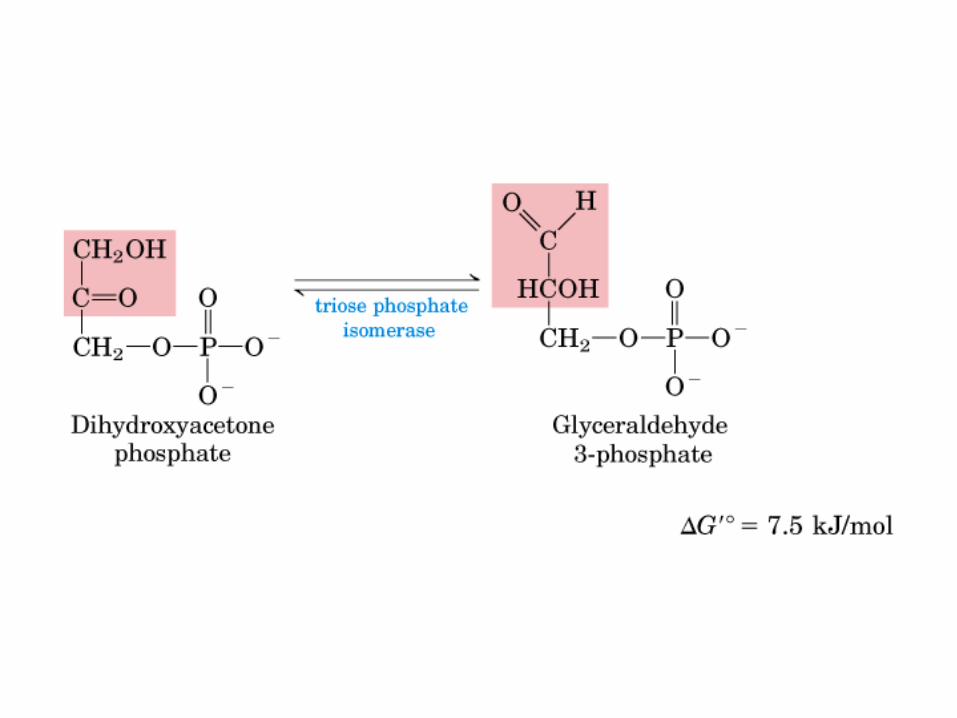
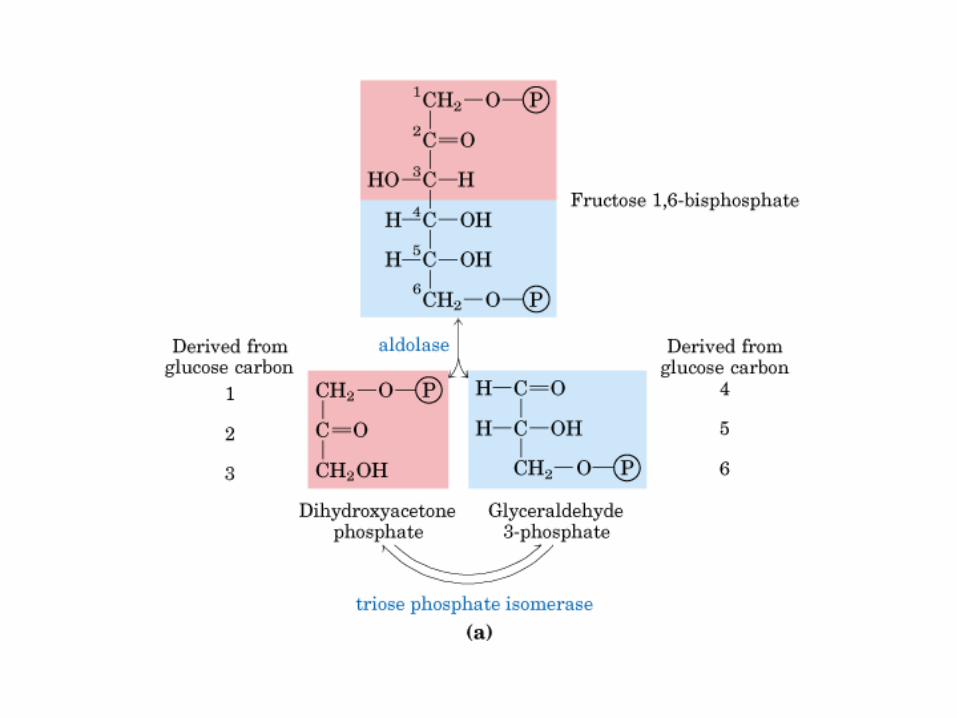
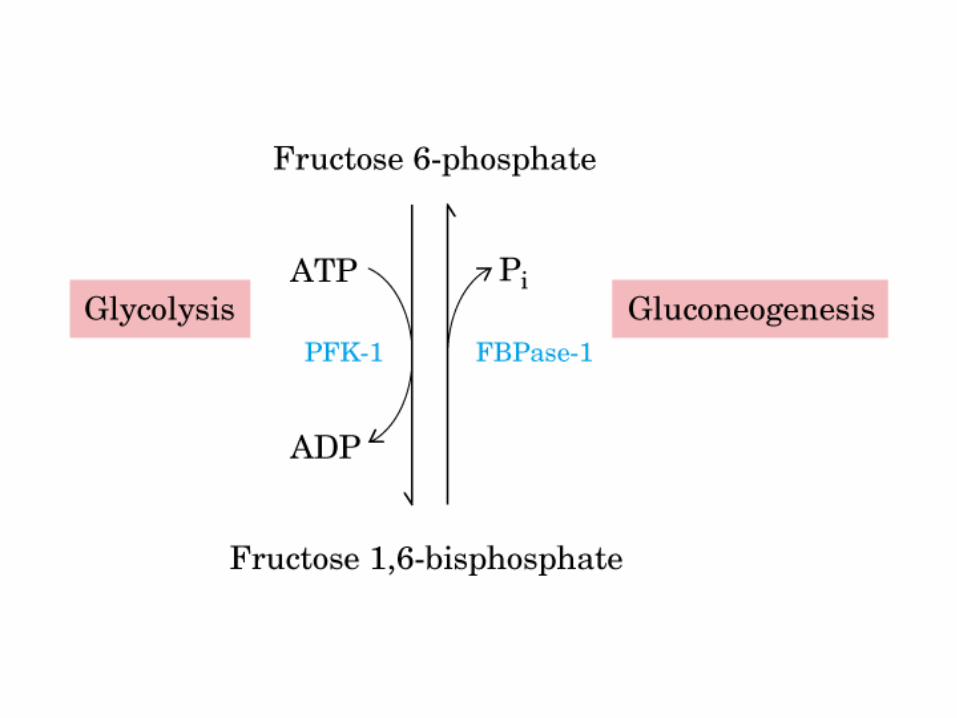
Principales vías y regulación de la gluconeogénesis y la glucólisis en el hígado.
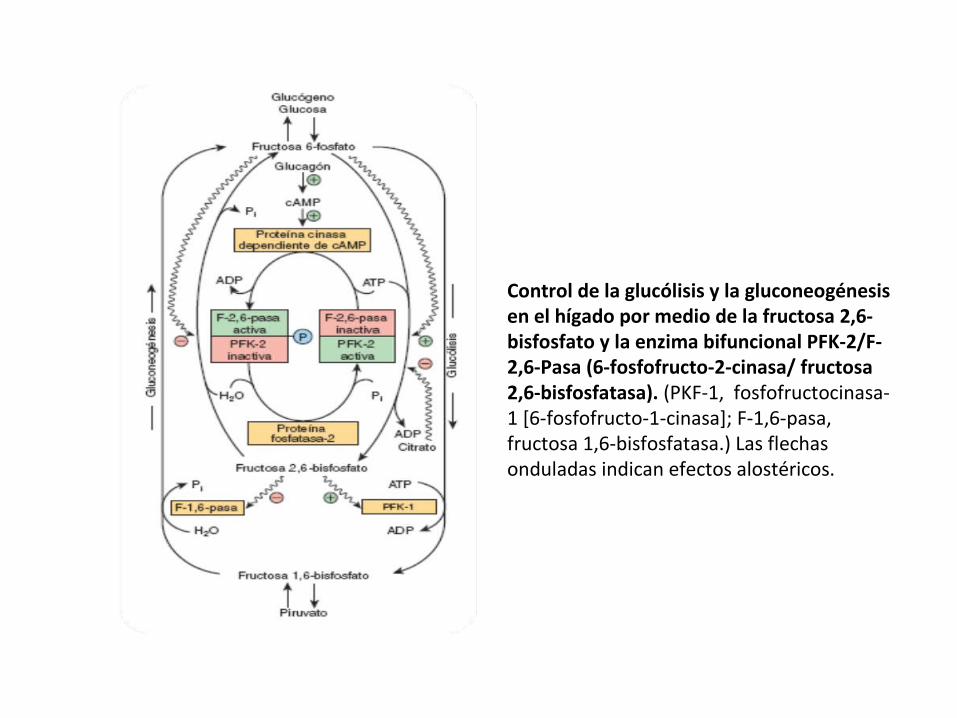
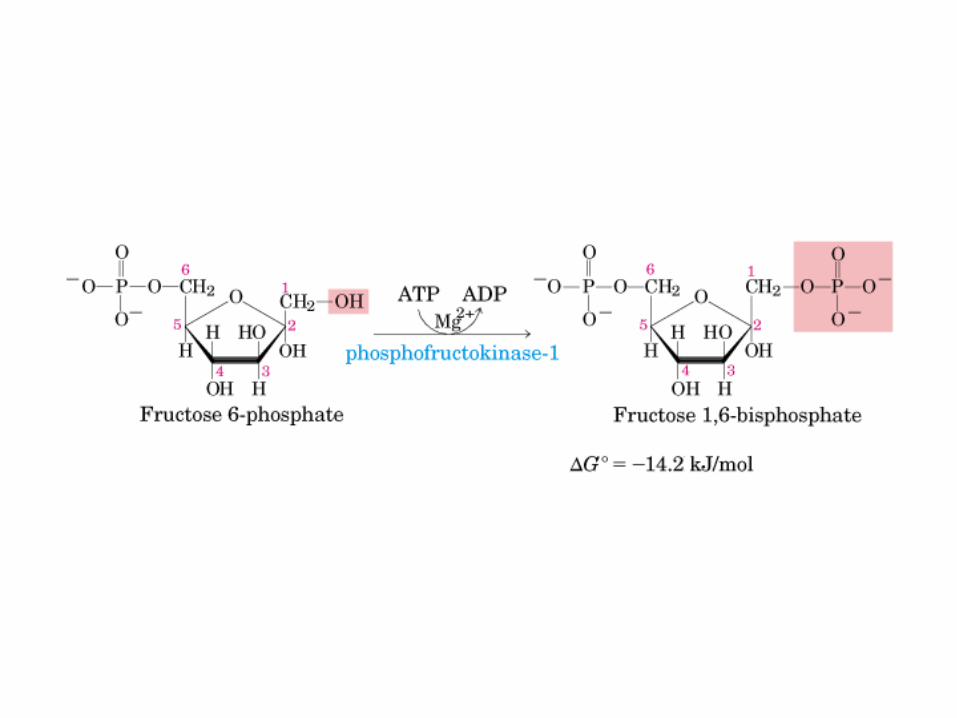
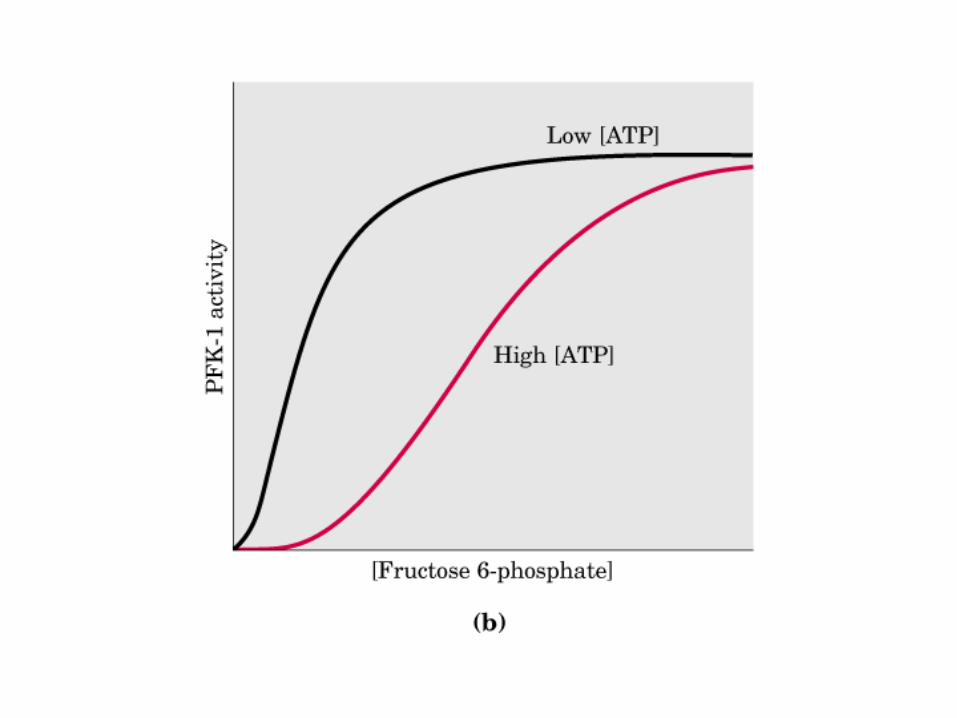
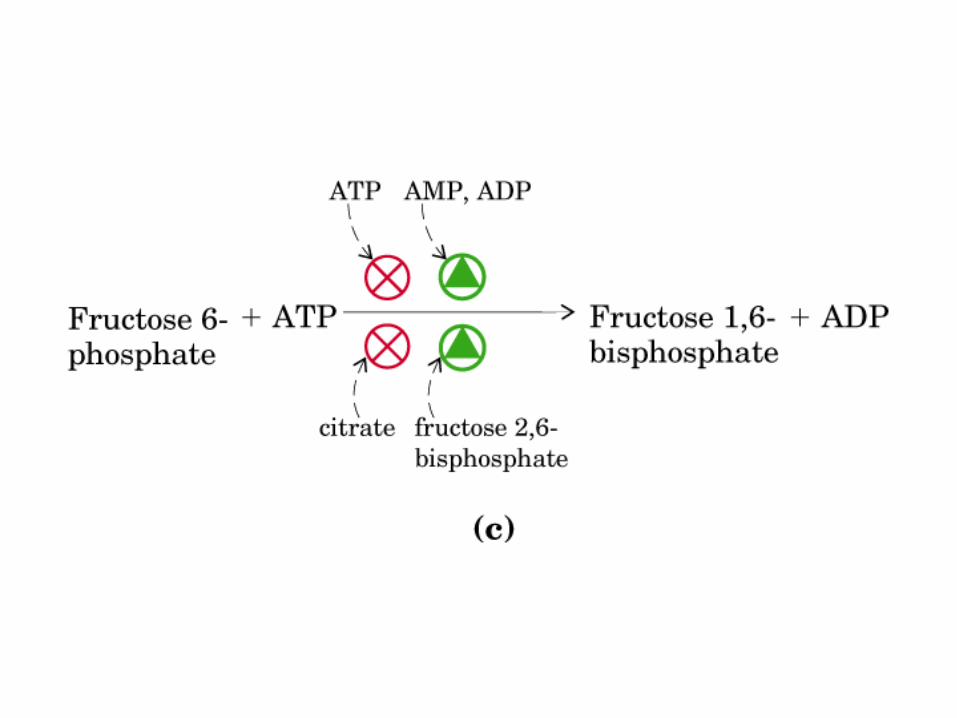
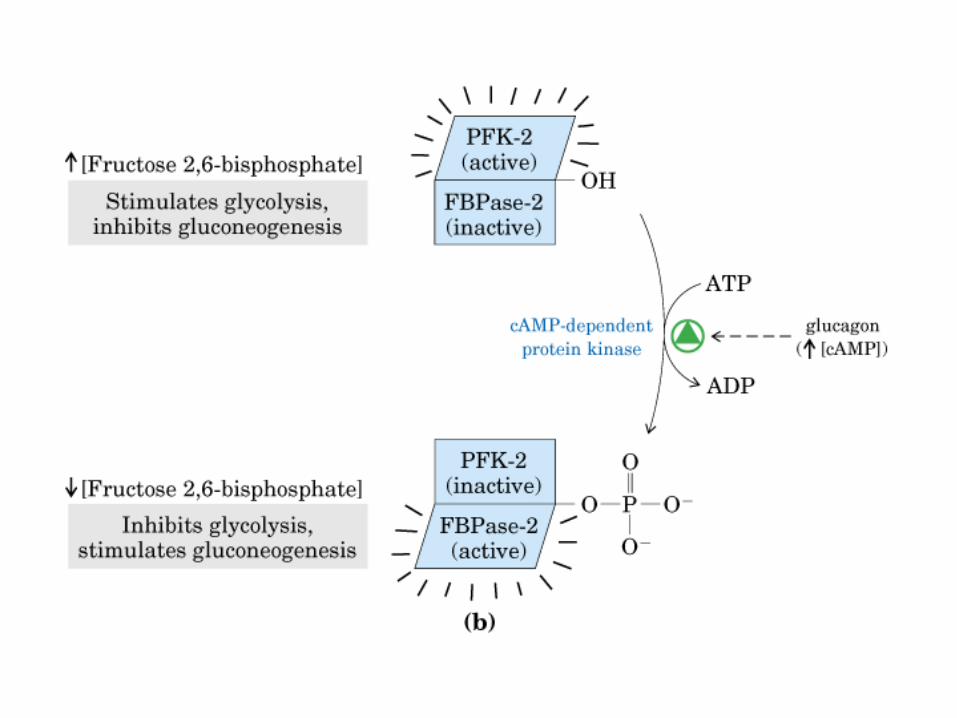
Control de la glucólisis y la gluconeogénesis en el hígado por medio de la fructosa 2,6-bisfosfato y la enzima bifuncional PFK-2/F-2,6-Pasa (6-fosfofructo-2-cinasa/ fructosa 2,6-bisfosfatasa). (PKF-1, fosfofructocinasa-1 [6-fosfofructo-1-cinasa]; F-1,6-pasa, fructosa 1,6-bisfosfatasa.) Las flechas onduladas indican efectos alostéricos.
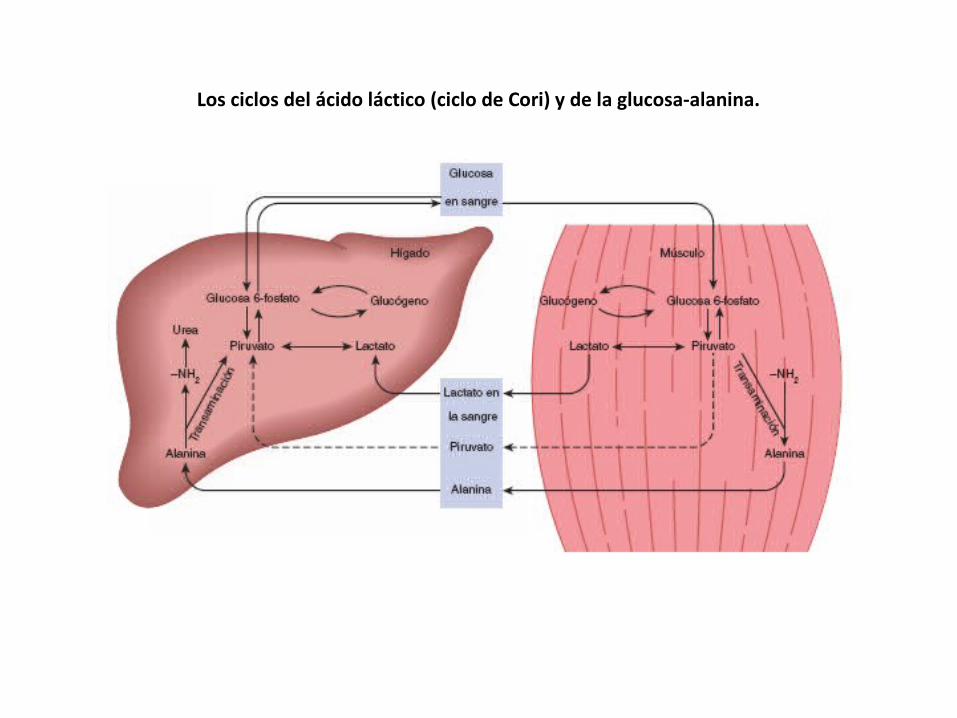
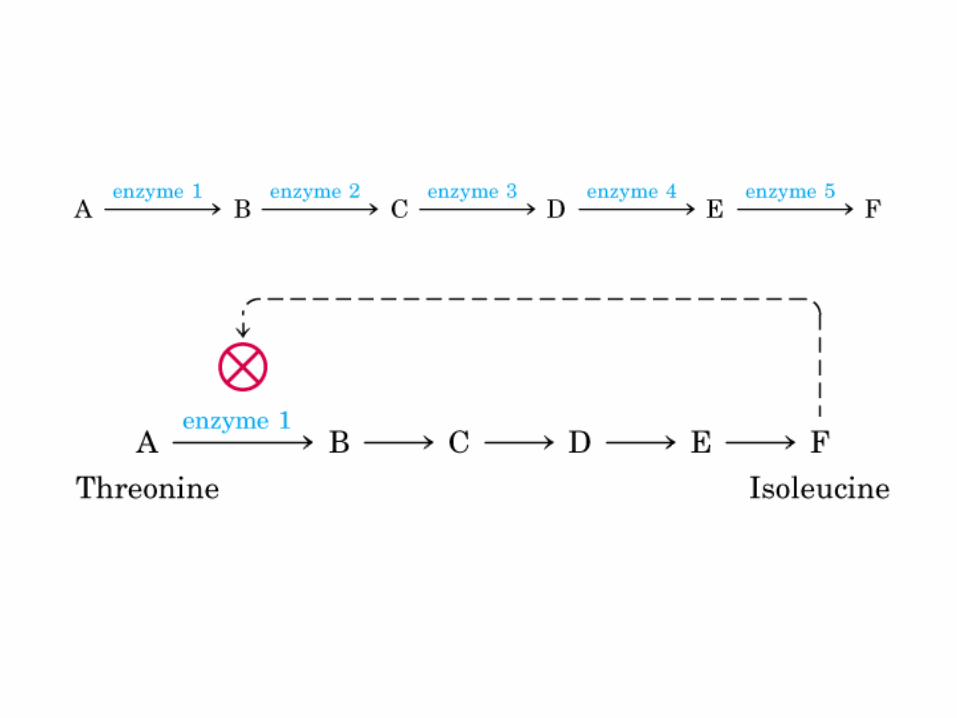
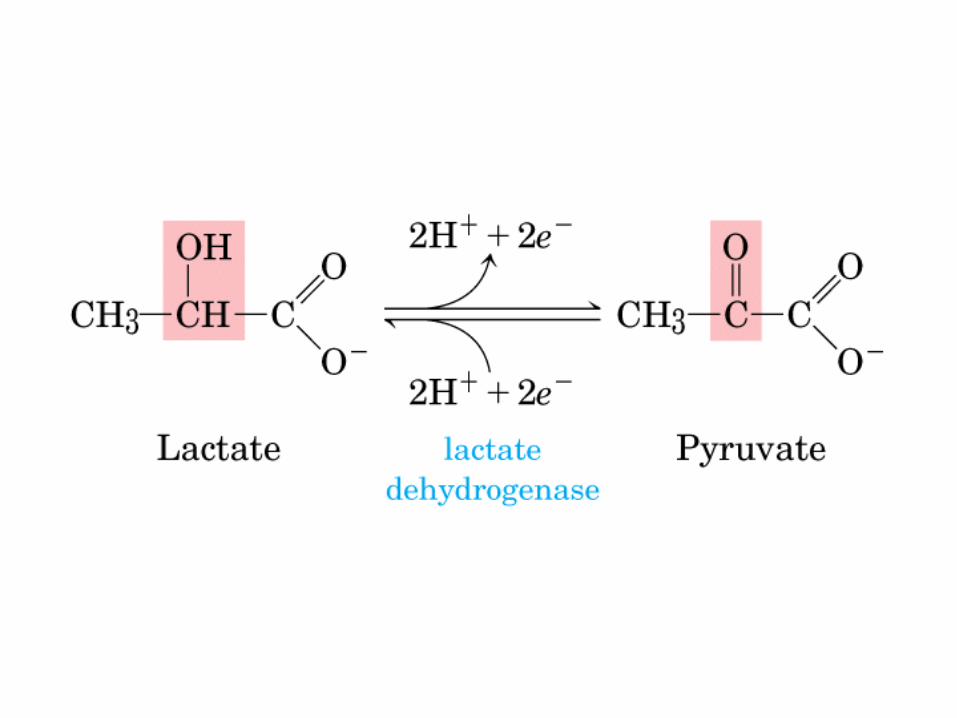
Los ciclos del ácido láctico (ciclo de Cori) y de la glucosa-alanina.




