Embed Size (px)
Citation preview

Mentor; prof, S. M. PRASAD MADE BY: DEEPAK YADAV
M.Sc-IV SEM.
RNA EDITING AND RNA TRANSPORT

RNA editing

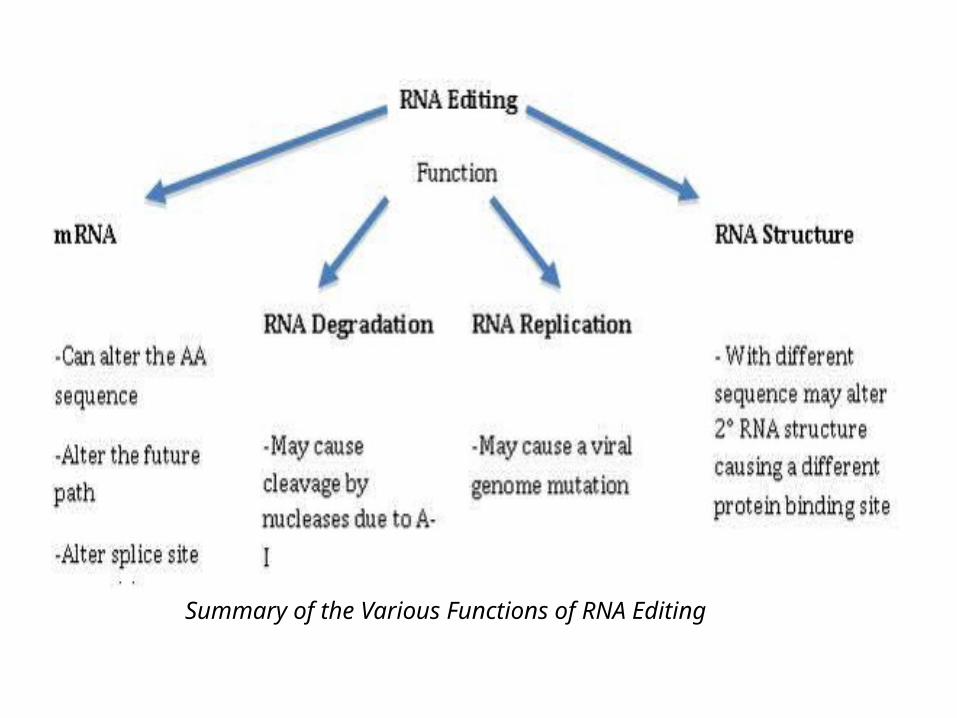
Summary of the Various Functions of RNA Editing

• RNA editing is a process in which information change at the level of mRNA. It is revealed by situations in which the coding sequence in an RNA differs from the sequence of DNA from which it was transcribed.
• Editing events may include the insertion, deletion, and base substitution of nucleotides within the edited RNA molecule.
• RNA editing has been observed in some tRNA, rRNA, mRNA molecules of eukaryotes and their viruses, archaea and prokaryotes.
• RNA editing occurs in the cell nucleus and cytosol, as well as within mitochondria and plastids.
RNA editing


• The diversity of RNA editing phenomena includes nucleobase modifications such as cytidine (C) to uridine (U) and adenosine (A) to inosine (I) deaminations, as well as non-templated nucleotide additions and insertions.
• RNA editing in mRNAs effectively alters the amino acid sequence of the encoded protein so that it differs from that predicted by the genomic DNA sequence.



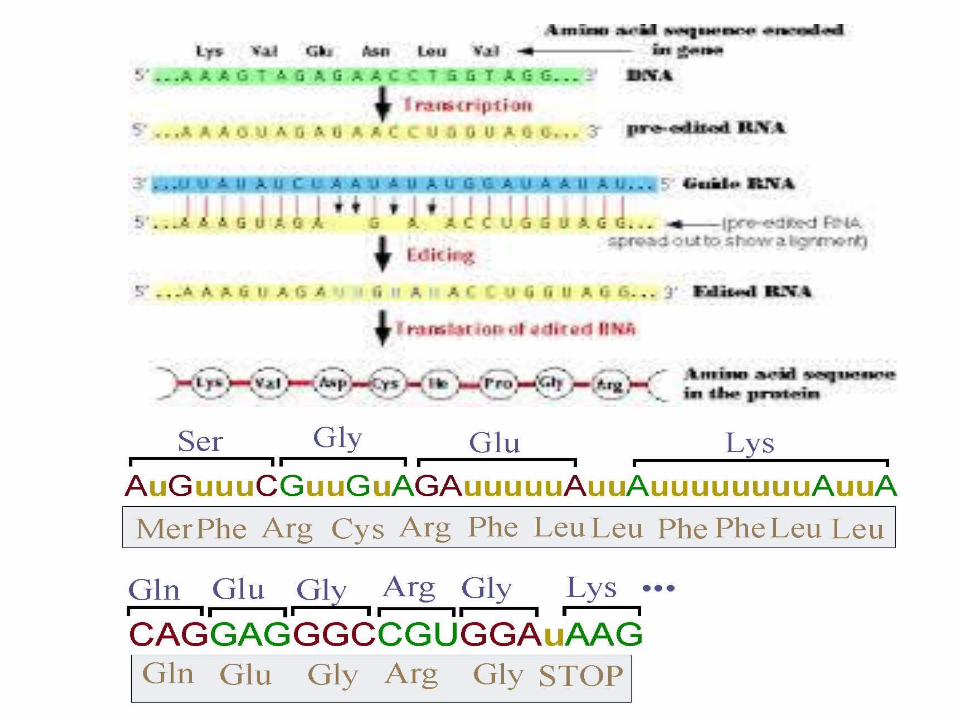
Editing by insertion or deletion• RNA editing through the addition and deletion
of uracil has been found in kinetoplasts from the mitochondria of Trypanosoma brucei Because this may involve a large fraction of the sites in a gene, it is sometimes called "pan-editing" to distinguish it from typical editing of one or a few sites.
• Pan-editing starts with the base-pairing of the unedited primary transcript with a guide RNA (gRNA), which contains complementary sequences to the regions around the insertion/deletion points.
• The newly formed double-stranded region is then enveloped by an editosome, a large multi-protein complex that catalyzes the editing.

• The editosome opens the transcript at the first mismatched nucleotide and starts inserting uridines.
• The inserted uridines will base-pair with the guide RNA, and insertion will continue as long as A or G is present in the guide RNA and will stop when a C or U is encountered.
• The inserted nucleotides cause a frameshift and result in a translated protein that differs from its gene.
• The mechanism of the editosome involves an endonucleolytic cut at the mismatch point between the guide RNA and the unedited transcript.
• The next step is catalyzed by one of the enzymes in the complex, a terminal U-transferase, which adds Us from UTP at the 3’ end of the mRNA.

• Another enzyme, a U-specific exoribonuclease, removes the unpaired Us.
• After editing has made mRNA complementary to gRNA, an RNA ligase rejoins the ends of the edited mRNA transcript.
• As a consequence, the editosome can edit only in a 3’ to 5’ direction along the primary RNA transcript.
• The complex can act on only a single guide RNA at a time. Therefore, a RNA transcript requiring extensive editing will need more than one guide RNA and editosome complex.



• C-U editing;-• The editing involves cytidine deaminase that deaminates a
cytidine base into a uridine base.
• An example of C-to-U editing is with the apolipoprotein B gene in humans.
• Apo B100 is expressed in the liver and apo B48 is expressed in the intestines.
• The B100 form has a CAA sequence that is edited to UAA, a stop codon, in the intestines. It is unedited in the liver.
Editing by deamination

The Effect of C-U RNA Editing on the Human ApoB gene


• A-I editing :- • A-to-I editing is the main form of RNA editing in mammals and
occurs in regions of double-stranded RNA (dsRNA).• Adenosine deaminases acting on RNA are the RNA-editing
enzymes involved in the hydrolytic deamination of Adenosine to Inosine (A-to-I editing).
• A-to-I editing can be specific (a single adenosine is edited within the stretch of dsRNA) or promiscuous (up to 50% of the adenosines are edited).
• Specific editing occurs within short duplexes, while promiscuous editing occurs within longer regions of duplex .


Systemic transport of RNA in plants

Plant vasculature was once thought to functionas a mere conduit for nutrients and hormones.
However, recent evidence indicatesthat this transport system can participate in the
dissemination of various signal moleculesthroughout the plant. Interestingly, these
signals include not only traditional signalingfactors, such as proteins and growth-regulating
small molecules, but also RNA.

Once RNA was thought to be an informationcarrier, functioning as a passive template for
protein synthesis. It is now also thoughtto act as an active signal molecule, regulatinggene expression and development in plants.Various types of RNA molecules travel long
distances from their site of synthesis to differentparts of the plant. This systemic movementoccurs through plant vasculature by as yet
unknown mechanisms. Here, three major typesof RNA systemic transport are summarized
(Fig. 1):

• Long distance movement of plant virusgenomic RNA.• Systemic transport of RNA elicitors ofpost-transcriptional gene silencing (PTGS).• Long distance transport of specific endogenous RNA molecules.


Following initial infection, usually by mechanical inoculation, plant RNA viruses spread from cell to cell through plasmodesmata until they reach the vascular system; the virus is then transported systemically throughthe vasculatureVirus-encoded nonstructural movement proteins mediate the cellto-cell spread of infection


Systemic transport of RNA signals forpost-transcriptional gene silencing
Because viruses often adapt existing cellularmachinery for their own needs, they probably
employ an endogenous pathway for the systemictransport of RNA. Indeed, recent evidence
indicates that numerous RNA speciestravel through plant vasculature, revealing anovel type of systemic signaling. Specifically,PTGS, an innate plant defense mechanism,is probably elicited by such systemic RNA
signals
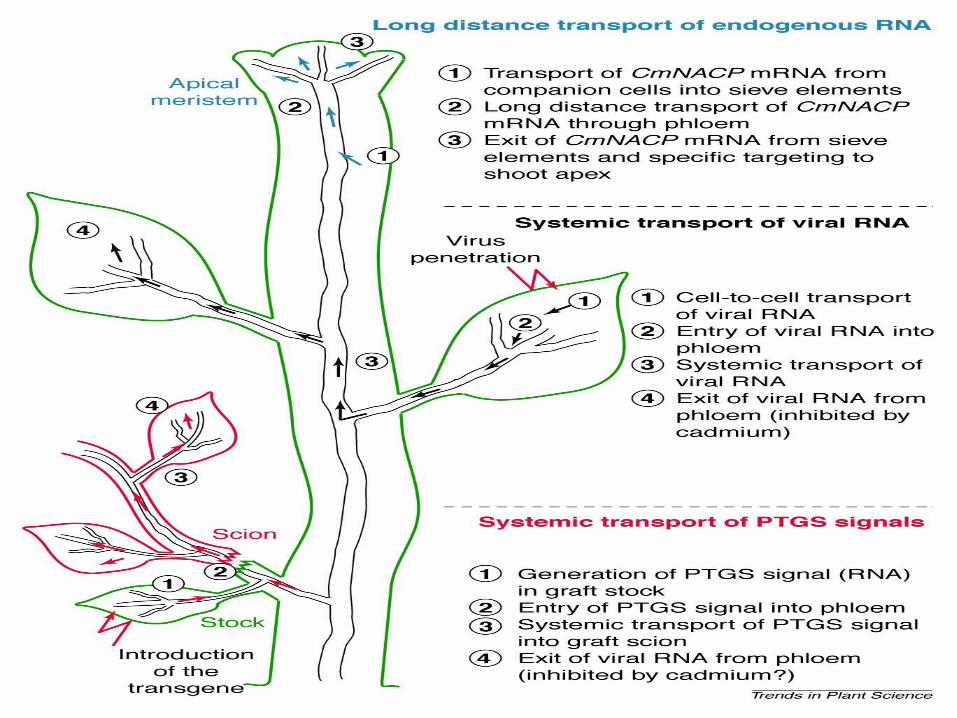

Thus, in plants, PTGS and virus-induced silencing provided the first indications of a regulatory role for the systemic transport of RNA.

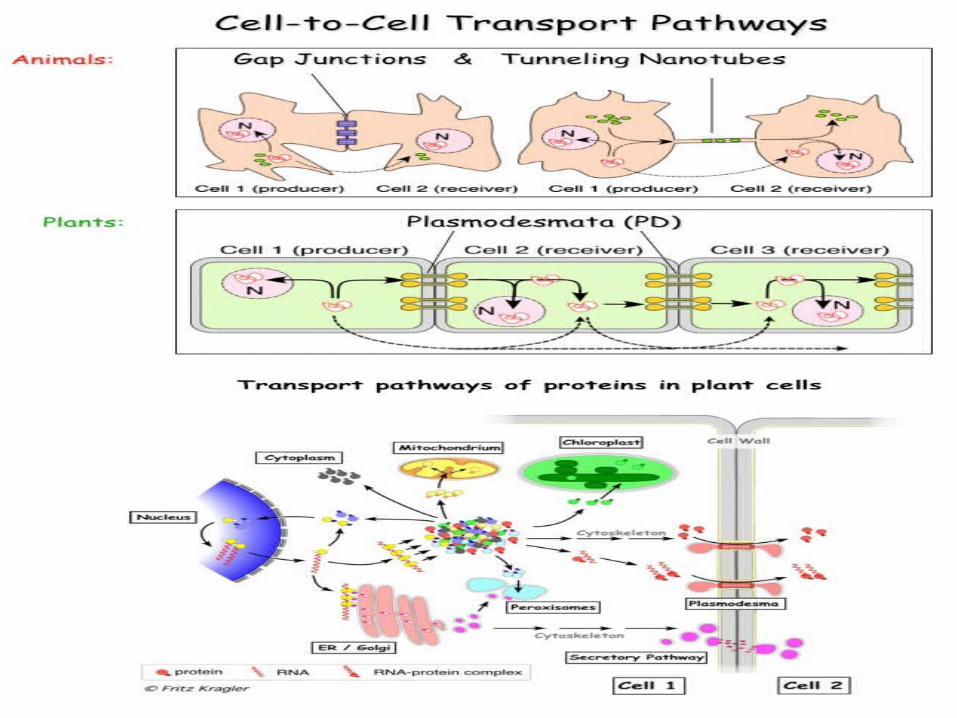
First, signal transduction between adjacent cell layers, as in Knotted transportation, is fast, whereas systemic signaling can occur at slower rates.
Second, cell-to-cell transport minimizes signal degradation or mistargeting, whereas movement through the entire phloem poses a higher risk of signal decay or delivery to the Wrong tissues.
Long distance transport of specific endogenous RNA molecules.


THANK YOU