
This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Original article
A new interpretation of the Miocene rodent faunas fromComanesti 1 and Taut (W-Romania)§
Une nouvelle interprétation des faunes de rongeurs de Comanesti 1 et de Taut(Roumanie de l’ouest)
János Hír a,*, Jérôme Prieto b,c, Emanoil Stiuca d
a Municipal Museum of Pásztó, 3060 Pásztó, Pf. 15. Hungaryb Senckenberg Center for Human Evolution and Palaeoecology (HEP), Eberhard-Karls University Tübingen, Institute for Geoscience,
Sigwartstrasse 10, 72076 Tübingen, Germanyc Department for Earth and Environmental Sciences, Ludwig Maximilians University Munich & Bavarian State Collections for Palaeontology and Geology,
Richard Wagner Strasse 10, 80333 Munich, Germanyd Emil Racovita Institute of speleology, Romanian Academy, Str. Frumoasa nr. 11. C.P. 220-12, 78114, Bucharest, Romania
Received 30 April 2010; accepted 14 January 2011
Available online 24 February 2011
Abstract
The rodent faunas from the Comanesti 1 and Taut localities (Western Romania) are revised in light of the latest taxonomical, biostratigraphicaland palaeoenvironmental information. The main systematic results show that the two cricetid rodents Megacricetodon crisiensis andDemocricetodon iazygum are invalid, whereas Democricetodon zarandicus is retained. The original assigment of the cricetodontini remainsfrom Taut to Hispanomys is emended, as a relationship to Byzantinia is more likely. While the geological evidence suggests that the localities arelate Middle Miocene (Upper Volhynian-Bessarabian, late Sarmatian sensu stricto) in age, the association of Myoglis ucrainicus with Muscardinushispanicus rather argues for an MN9 correlation for Taut. Indeed, uncertainties and discontinuities in the Central and East European mammalianbiostratigraphy render any conclusion about the correlation of the localities to the MN ‘‘zonation’’ problematic. The high diversity of squirrels (fivegenera), as well as the presence of a pliopithecoid alongside, glirid and eomyid rodents, suggest a forest environment at the time of accumulation ofthe Taut fauna, which is in agreement with the indication of humid climate provided by ectothermic vertebrates.# 2011 Published by Elsevier Masson SAS.
Keywords: Biostratigraphy; Central Paratethys; Rodents; Miocene; paleoenvironment
Résumé
Une révision des faunes de rongeurs des localités Comanesti 1 et Taut (Roumanie de l’ouest) est réalisée à la lumière des dernièresconnaissances taxonomiques, biostratigraphiques et paléoenvironmentales. Les principaux résultats systématiques montrent que les deux rongeurscricetidés Megacricetodon crisiensis et Democricetodon iazygum ne peuvent être considérés comme valides, alors que Democricetodon zarandicusest conservé. L’attribution initiale des Cricetodontini de Taut au genre Hispanomys est corrigée, une relation avec Byzantinia étant plus probable.Bien que les informations géologiques suggèrent un âge Miocène moyen tardif (Volhynien supérieur-Bessarabien, Sarmatien tardif sensu stricto),l’association Myoglis ucrainicus + Muscardinus hispanicus est plus compatible avec une corrélation à MN9 pour Taut. Du fait des discontinuités etdes incertitudes liées à la biostratigraphie d’Europe centrale, toute conclusion est rendue fragile en ce qui concerne la correlation des sites avec la« zonation » MN. La grande diversité des sciuridés (cinq genres) ainsi que la présence d’un pliopithécoïde aux côtés de gliridés et d’éomyidés,peuvent être liées à une couverture végétale dense à l’époque de l’accumulation de la faune de Taut, une proposition en accord avec les donnéespluviométriques dérivées des vertébrés ectothermes.# 2011 Publié par Elsevier Masson SAS.
Mots clés : Biostratigraphie ; Parathéthys centrale ; Rongeurs ; Miocène ; Paléoenvironement
Geobios 44 (2011) 215–223
§ Invited editor: Jordi Agustí.* Corresponding author.
E-mail address: [email protected] (J. Hír).
0016-6995/$ – see front matter # 2011 Published by Elsevier Masson SAS.doi:10.1016/j.geobios.2011.01.003

Author's personal copy
1. Introduction
Late Middle Miocene to earliest Late Miocene smallmammal localities have rarely been reported from CentralEurope, and thus the faunal evolution during this period of timeremains poorly understood in this area. During the 5thSymposium of the Romanian Palaeontologists in Bucharest(2005), the first author and Márton Venczel therefore took theopportunity to re-study the original fossil material from Tautand Comanesti 1 that is kept in the Speleological Institute of theRomanian Scientific Academy (with the kind permission of thedirectorship). It became clear already after a first overview, thata detailed and thorough revision was necessary. At the sametime, an elaboration of the herpetofauna was completed(Venczel and Stiuca, 2008; excluding chelonians). In this paper,we present a taxonomical revision of the rodent faunas, anddiscuss the consequences of this revision on biostratigraphicaland palaeoenvironmental interpretations.
2. Historical and geological settings
2.1. Historical background
The faunas from Comanesti 1 and Taut have an importantposition in the history of Middle Miocene micromammalianresearch Europe. Indeed, the original descriptions of fossilsfrom these two localities represent a starting point for theHungarian and Romanian research in this region (Feru et al.,1979, 1980). During the following three decades, intensive fieldwork was conducted, which provided large collections ofMiocene micromammals and small vertebrates in general, thatform the basis for the present work (Hír and Kókay, 2004, 2009,2010; Hír and Venczel, 2005; Hír, 2006; Venczel and Stiuca,2008).
2.2. The localities
2.2.1. Comanesti 1This vertebrate locality is located at the eastern margin of the
village Comanesti, on the right side of the rivulet Hasmas. Thesediments of the eleven meter-high outcrop crop out in a privateyard (GPS: N 468 30.568’, E 228 02.807; section figured byIstocescu, 1971 and Feru et al., 1980), and are extremelyenriched in molluscs. From layer 21, which containscontinental vertebrates, Feru et al. (1980) listed Cardiumvindobonense vindobonense, C. latisulcum latisulcum, C.gracilicostatum, Ervilia dissita podolica and Trochus pictus.This layer belongs to the top of the lower lithological complexof the Sarmatian in the Crisul Alb Basin (sensu Istocescu, 1971;Istocescu and Istocescu, 1974) and correlates to the lateVolhynian/earliest Bessarabian (Feru et al., 1980) or to theentire Ervilia Zone of the Central Paratethys, late Volhynian(Harzhauser and Piller, 2004).
The continental vertebrate fossils collected from the lensesof tufa were initially described by Feru et al. (1980), andsubsequently followed by a study of the cricetid rodents withthe description of two new species (Radulescu and Samson,
1988). These authors correlate the fossils to the MN8 (latestMiddle Miocene). More recently, Grigorescu and Kazár (2006)described the odontocetes from this locality.
2.2.2. TautThis vertebrate locality is located south of the small village
Taut, in the Migiesului Valley (Arad District, WesternRomania). The tufaceous green clay containing Helix andPlanorbis shells was originally reported by Istocescu (1971)and Istocescu and Istocescu (1974). From the microvertebratematerial originally described by Feru et al. (1979), the cricetidrodents were emended by Radulescu and Samson (1988), withthe description of Democricetodon zarandicus nov. sp.McNulty et al. (1999) reported the lower deciduous premolarof a small-sized Pliopithecoid primate (Crouzelidae gen. andsp. indet.) from the Taut fauna. The herpetofauna (excludingchelonians) has been described by Venczel and Stiuca (2008).During the last decade, Márton Venczel and János Hír tried torediscover the locality in the field, but have been unsuccessfulso far.
Taut belongs to the middle lithological complex (sensuIstocescu, 1971; Istocescu and Istocescu, 1974). Feru et al.(1980) noticed that this complex corresponds to a distallacustrine facies that was accumulated, at least in its upper part,synchronously with the deposition of the lower lithologicalcomplex. Based on the evolutionary stage of the micromam-malian fauna, these authors suggest a lower Bessarabiancorrelation for this locality.
3. Material and methods
The nomenclature of the dental elements follows: Sciuridae:Cuenca-Bescos (1988); Gliridae: Daams (1981); Eomyidae:Engesser (1999); Cricetidae: Mein and Freudenthal (1971); andRummel (1998) for the special terminology of Cricetodontiniand Eumyarion teeth. Measurements were taken through ameasuring ocular mounted on a MBS 9 stereomicroscope andare given in mm. The micrographs were captured with a CanonEos 400D digital camera equipped with a Canon MP-E65 mmmacro-objective and a Canon MR-14EX macro ring light.Retouches were made by the first author using Photoshop 2 andPhotoshop 6.0. Drawings were made using a Zeiss drawingmicroscope.
4. Systematic palaeontology
Full paleontological descriptions of all identified taxa, aswell as the corresponding references, are given in Appendix A.A synoptical overview of the taxon occurrences resulting fromthe systematic analysis is given in Table 1. In addition, Figs. 1–
4 present rodent specimens from the localities Comanesti 1 andTaut.
The main results of our study show that:
� because of the small sample size from Comanesti 1, thecricetid rodent Democricetodon iazygum is considered asnomen dubium;
J. Hír et al. / Geobios 44 (2011) 215–223216

Author's personal copy
� the diagnosis of Democricetodon zarandicus is emended;� for similar reasons, we provisionaly assign the specimens
from Comanesti 1 to Megacricetodon cf. minutus, consider-ing Megacricetodon crisiensis as a junior synonym of theAustrian species;� the largest cricetid molars from Comanesti 1 are morpho-
logically similar to Byzantinia, but the lack of sufficientmaterial neither permits any confident assignment of thesample nor valuation of its taxonomical homogeneity;� the dormouse Miodyromys aff. grycivensis may have been
present in Comanesti 1;� Muscardinus hispanicus, Glirudinus sp., Myoglis ucrainicus
and Glirulus lissiensis as well as the eomyid Eomyopsoppligeri occur in Taut.
5. Discussion
5.1. Biostratigraphic implications
The taxonomic revision of the fossils from Comanesti 1 andTaut allows us to re-evaluate the biostratigraphic correlation ofthe localities.
5.1.1. Comanesti 1Because of the small number of specimens, the taxonomic
determinations, and thus the resulting biostratigraphic implica-tions are vague. Megacricetodon disappears during MN9 (e.g.,Kälin, 1999). The earliest occurrence of a form related toMegacricetodon minutus has been reported from Gratkorn ataround 12–12.5 My (Prieto et al., 2010; Gross et al., in press).The oldest Byzantinia specimens have been recorded for thelatest Middle Miocene of Anatolia (MN7+8; Ünay et al., 2003),
whereas a Cricetodon-Byzantinia transitional species also livedin SE-Europe at this time (localities not indicated; Rummel,1999). With regard to the mollusk fauna, the deposits of thelocality are assigned to the upper part of the Volhynian, thusclearly indicating a late Middle Miocene age for the fauna.
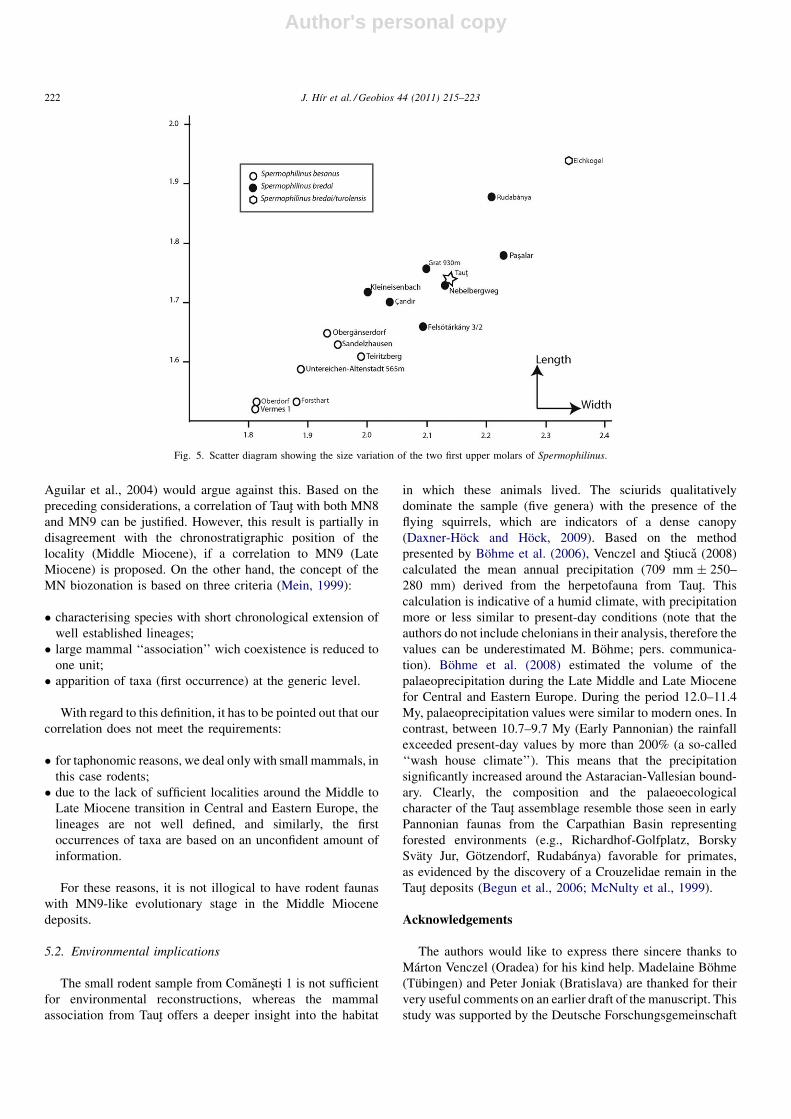
5.1.2. TautSquirrels are not normally useful for fine biostratigraphy, but
the relatively large-sized Spermophilinus molars from Taut arein agreement with data available for the late Middle Miocene to(?)earliest Late Miocene localities from Central Europe(Fig. 5). The cricetid rodents do not provide precise informationbecause of taxonomic uncertainty (Eumyarion), or limitedgeographic range (Democricetodon zarandicus). The presenceof Eomyops oppligeri might argue for an MN8 correlation, butthe species has also been reported from localities traditionallycorrelated to MN9 (early Late Miocene) such as Bełchatów A(Garapich, 2002) and Nebelbergweg (Kälin and Engesser,2001). Similarly, E. oppligeri co-occurs at the top of theFelsötárkány succession with Microtocricetus molassicus (Hírand Kókay, 2010). The importance of Microtocricetus as acharacteric taxon for the MN9 has been challenged by Prietoet al. (2010), and the presence of species related to the Collimyshiri-C. dobosi lineage in both Bełchatów A and Felsötárkány3/10 also probably argues for a latest Middle Miocenecorrelation of the two localities (Prieto, pers. opinion). Theglirid Muscardinus hispanicus first occurs in Spain, i.e. in theupper part of the zone G3 of the Calatayud-Daroca and Teruelbasins (MN8, Casanovas-Vilar et al., 2008). Myoglis ucrainicushas originally been described from Grytsiv, a locality thatbelongs to the MN9 because of the presence of hipparioninhorses, and has also been recorded for the Austrian LateMiocene (Daxner-Höck and Höck, 2009). On the other hand,
Table 1Overview of the taxon occurrences and synonymies resulting from the systematic analysis of the rodents from Taut and Comanesti 1.
Feru et al. (1979) Feru et al. (1980) Radulescu and Samson (1988) This paper
COMANESTI 1Peridyromys gr. hamadryas Miodyromys aff. grycivensisMyoglis meiniHispanomys cf. lavocati cf. Byzantinia sp. or div. sp.Hispanomys cf. bijugatus cf. Byzantinia sp. or div. sp.Democricetodon gr. gaillardi Democricetodon iazygum nov. sp. Democricetodon cf. freisingensis
vel. cf. gaillardiMegacricetodon gr. similis Megacricetodon crisiensis nov. sp. Megacricetodon cf. minutus
TAUvSpermophilinus bredai Spermophilinus bredaiMiopetaurista sp. Miopetaurista sp.Forsithya sp.Blackia sp. Blackia miocaenicaLeptodontomys gr. catalaunicus Eomyops oppligeriEomuscardinus gr. sansaniensis Muscardinus hispanicusGlirid gen. and sp. indet. Glirulus lissiensisMyoglis meini Myoglis ucrainicusDemocricetodon gr. gaillardi Democricetodon gr. gaillardi-brevis Democricetodon zarandicus nov. sp. Democricetodon zarandicusEumyarion cf. bifidus Eumyarion sp.
Glirudinus sp.?Albanensia sp.Neopetes hoeckarum
J. Hír et al. / Geobios 44 (2011) 215–223 217
Author's personal copy
[()TD$FIG]
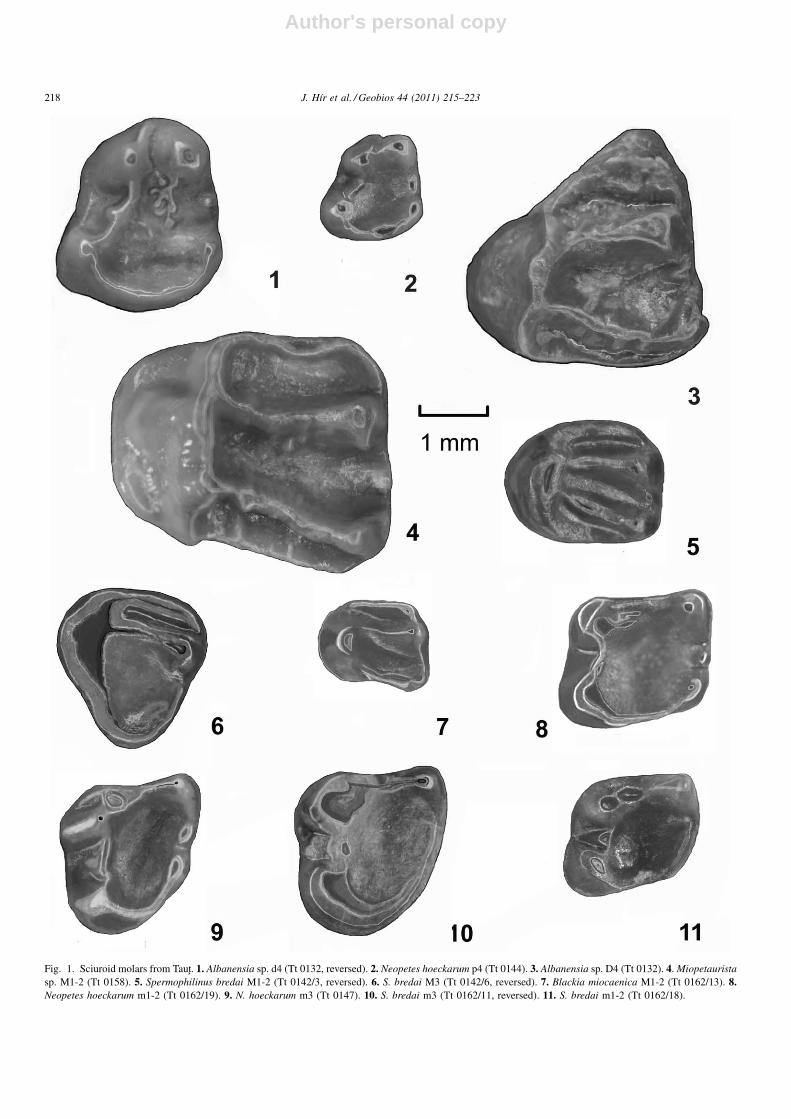
Fig. 1. Sciuroid molars from Taut. 1. Albanensia sp. d4 (Tt 0132, reversed). 2. Neopetes hoeckarum p4 (Tt 0144). 3. Albanensia sp. D4 (Tt 0132). 4. Miopetauristasp. M1-2 (Tt 0158). 5. Spermophilinus bredai M1-2 (Tt 0142/3, reversed). 6. S. bredai M3 (Tt 0142/6, reversed). 7. Blackia miocaenica M1-2 (Tt 0162/13). 8.Neopetes hoeckarum m1-2 (Tt 0162/19). 9. N. hoeckarum m3 (Tt 0147). 10. S. bredai m3 (Tt 0162/11, reversed). 11. S. bredai m1-2 (Tt 0162/18).
J. Hír et al. / Geobios 44 (2011) 215–223218

Author's personal copy
the fissure filling sediments from Grytsiv contain Sarmatimac-tra vitaliana (Topachevsky and Scorik, 1992; Topachevskiet al., 1996; Rzebik-Kowalska and Topachevsky, 1997;Vangegeim et al., 2006), a characteristic bivalve from theLate Sarmatian s. str. in the Central Parathethys (Piller et al.,
2007). The presence of M. ucrainicus in Taut indicates that itwas already present in the latest Middle Miocene. The presenceof Glirudinus, which is absent from the Late Miocene ofAustria (Daxner-Höck and Höck, 2009), could possibly be usedto confirm the correlation, but its presence in Jujurieux 1 (MN9;
[()TD$FIG]
Fig. 2. Eomyid and glirid molars from Taut and Comanesti 1. 1. Eomyops oppligeri m2 (Tt 0118, reversed). 2. Glirudinus sp. m2 (Tt 0200/1, reversed). 3. Gliruluslissiensis M1-2 (Tt 0201/1). 4. Miodyromys aff. grycivensis M1-2 (Com1-0115). 5. Muscardinus hispanicus M1 (Tt 0203/1). 6. M. hispanicus M1 (Tt 0203/2). 7.Myoglis ucrainicus p4 (Tt 0164). 8. M. ucrainicus M1 (Tt 0164, occlusal view). 9. M. ucrainicus M1 (Tt 0164, radical view). 10. M. ucrainicus m1 (Tt 0164). 11. M.ucrainicus m2 (Tt 0164).
J. Hír et al. / Geobios 44 (2011) 215–223 219
Author's personal copy
[()TD$FIG]
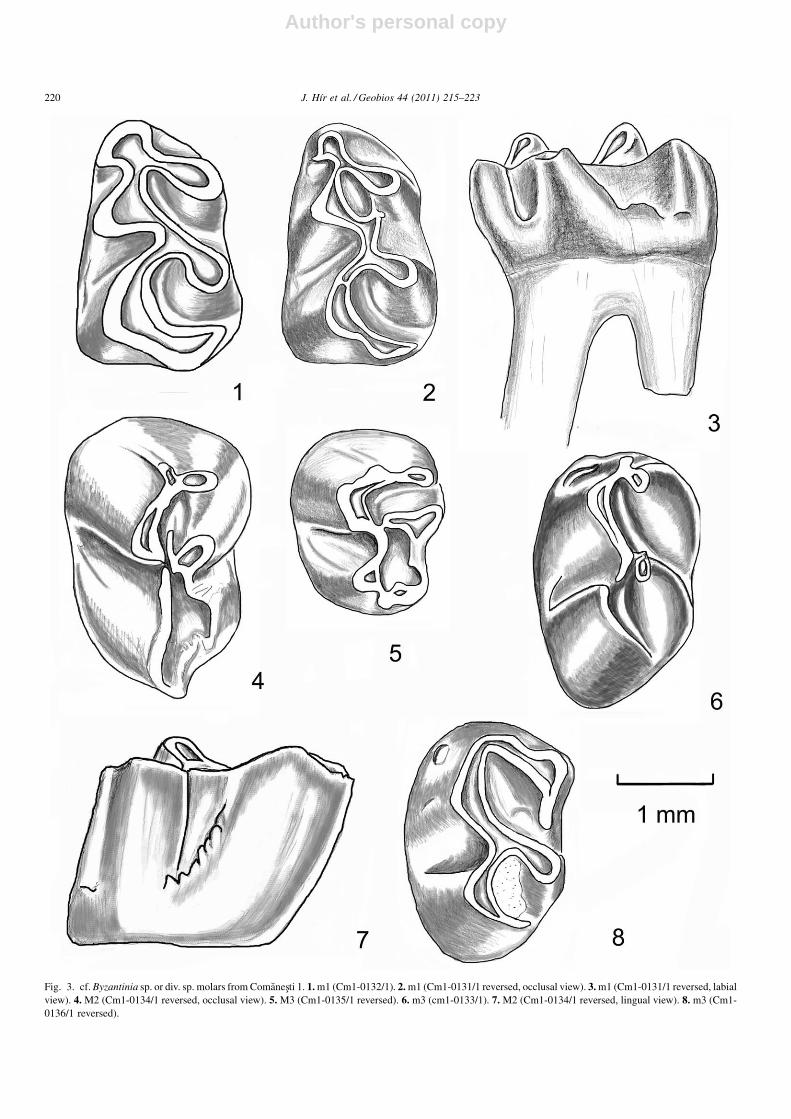
Fig. 3. cf. Byzantinia sp. or div. sp. molars from Comanesti 1. 1. m1 (Cm1-0132/1). 2. m1 (Cm1-0131/1 reversed, occlusal view). 3. m1 (Cm1-0131/1 reversed, labialview). 4. M2 (Cm1-0134/1 reversed, occlusal view). 5. M3 (Cm1-0135/1 reversed). 6. m3 (cm1-0133/1). 7. M2 (Cm1-0134/1 reversed, lingual view). 8. m3 (Cm1-0136/1 reversed).
J. Hír et al. / Geobios 44 (2011) 215–223220
Author's personal copy
[()TD$FIG]
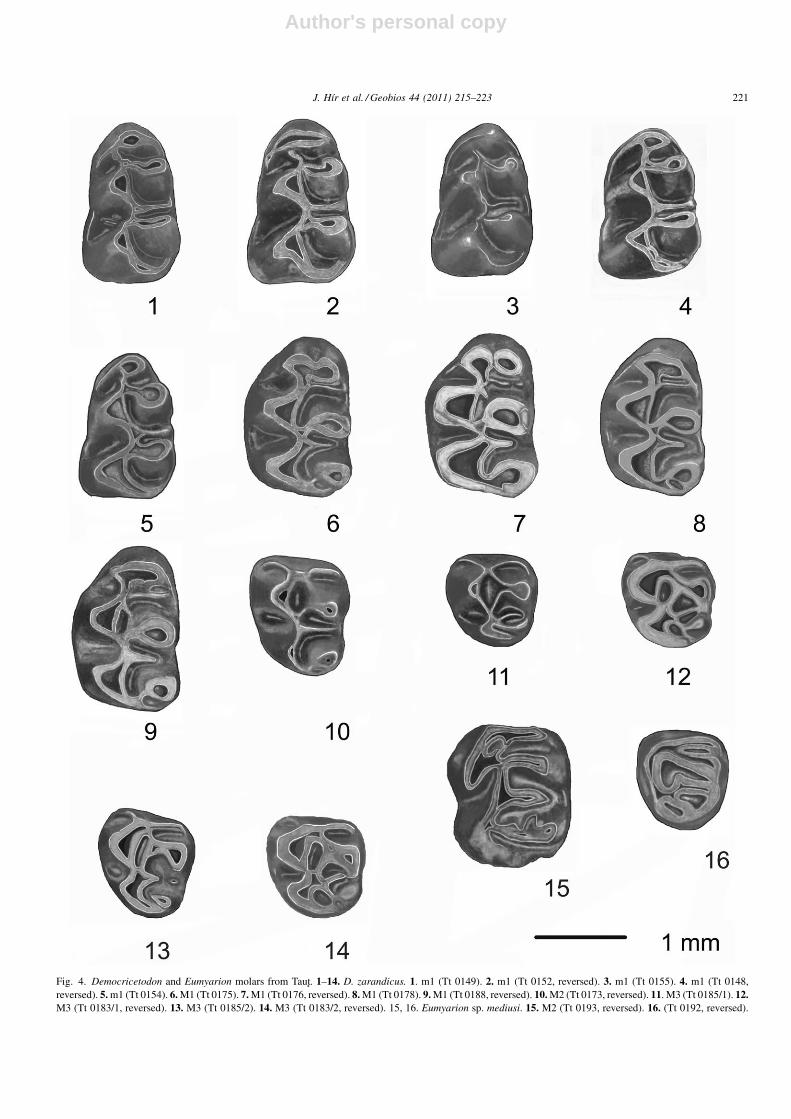
Fig. 4. Democricetodon and Eumyarion molars from Taut. 1–14. D. zarandicus. 1. m1 (Tt 0149). 2. m1 (Tt 0152, reversed). 3. m1 (Tt 0155). 4. m1 (Tt 0148,reversed). 5. m1 (Tt 0154). 6. M1 (Tt 0175). 7. M1 (Tt 0176, reversed). 8. M1 (Tt 0178). 9. M1 (Tt 0188, reversed). 10. M2 (Tt 0173, reversed). 11. M3 (Tt 0185/1). 12.M3 (Tt 0183/1, reversed). 13. M3 (Tt 0185/2). 14. M3 (Tt 0183/2, reversed). 15, 16. Eumyarion sp. mediusi. 15. M2 (Tt 0193, reversed). 16. (Tt 0192, reversed).
J. Hír et al. / Geobios 44 (2011) 215–223 221
Author's personal copy
Aguilar et al., 2004) would argue against this. Based on thepreceding considerations, a correlation of Taut with both MN8and MN9 can be justified. However, this result is partially indisagreement with the chronostratigraphic position of thelocality (Middle Miocene), if a correlation to MN9 (LateMiocene) is proposed. On the other hand, the concept of theMN biozonation is based on three criteria (Mein, 1999):
� characterising species with short chronological extension ofwell established lineages;� large mammal ‘‘association’’ wich coexistence is reduced to
one unit;� apparition of taxa (first occurrence) at the generic level.
With regard to this definition, it has to be pointed out that ourcorrelation does not meet the requirements:
� for taphonomic reasons, we deal only with small mammals, inthis case rodents;� due to the lack of sufficient localities around the Middle to
Late Miocene transition in Central and Eastern Europe, thelineages are not well defined, and similarly, the firstoccurrences of taxa are based on an unconfident amount ofinformation.
For these reasons, it is not illogical to have rodent faunaswith MN9-like evolutionary stage in the Middle Miocenedeposits.
5.2. Environmental implications
The small rodent sample from Comanesti 1 is not sufficientfor environmental reconstructions, whereas the mammalassociation from Taut offers a deeper insight into the habitat
in which these animals lived. The sciurids qualitativelydominate the sample (five genera) with the presence of theflying squirrels, which are indicators of a dense canopy(Daxner-Höck and Höck, 2009). Based on the methodpresented by Böhme et al. (2006), Venczel and Stiuca (2008)calculated the mean annual precipitation (709 mm � 250–
280 mm) derived from the herpetofauna from Taut. Thiscalculation is indicative of a humid climate, with precipitationmore or less similar to present-day conditions (note that theauthors do not include chelonians in their analysis, therefore thevalues can be underestimated M. Böhme; pers. communica-tion). Böhme et al. (2008) estimated the volume of thepalaeoprecipitation during the Late Middle and Late Miocenefor Central and Eastern Europe. During the period 12.0–11.4My, palaeoprecipitation values were similar to modern ones. Incontrast, between 10.7–9.7 My (Early Pannonian) the rainfallexceeded present-day values by more than 200% (a so-called‘‘wash house climate’’). This means that the precipitationsignificantly increased around the Astaracian-Vallesian bound-ary. Clearly, the composition and the palaeoecologicalcharacter of the Taut assemblage resemble those seen in earlyPannonian faunas from the Carpathian Basin representingforested environments (e.g., Richardhof-Golfplatz, BorskySväty Jur, Götzendorf, Rudabánya) favorable for primates,as evidenced by the discovery of a Crouzelidae remain in theTaut deposits (Begun et al., 2006; McNulty et al., 1999).
Acknowledgements
The authors would like to express there sincere thanks toMárton Venczel (Oradea) for his kind help. Madelaine Böhme(Tübingen) and Peter Joniak (Bratislava) are thanked for theirvery useful comments on an earlier draft of the manuscript. Thisstudy was supported by the Deutsche Forschungsgemeinschaft
[()TD$FIG]
Fig. 5. Scatter diagram showing the size variation of the two first upper molars of Spermophilinus.
J. Hír et al. / Geobios 44 (2011) 215–223222

Author's personal copy
grant BO1550/16-1 and the project no. T046719 of theHungarian Scientific Research Fund (OTKA).
Appendix A. Supplementary data
Supplementary text and data (Tables S1 to S13 and Figs. S1and S2) associated with this article can be found, in the onlineversion, at doi:10.1016/j.geobios.2011.01.003.
References
Begun, D.R., Nargolwalla, M.C., Hutchison, M.P., 2006. Primate diversity inthe Pannonian Basin: In situ evolution dispersal or both? Beiträge zurPaläontologie 30, 43–56.
Böhme, M., Ilg, A., Ossig, A., Küchenhoff, H., 2006. New method to estimatepaleoprecipitation using fossil amphibians and reptiles and the middle andlate Miocene precipitation gradients in Europe. Geology 34, 425–428.
Böhme, M., Ilg, A., Winklhofer, M., 2008. Late Miocene ‘‘washouse’’ climatein Europe. Earth and Planetary Science Letters 275, 393–401.
Casanovas-Vilar, I., Alba, D.M., Moya-Sola, S., Galindo, J., Cabrera, L.,Garces, M., Fùrio, M., Robles, J.M., Köhler, M., Angelone, M., 2008.Biochronological, taphonomical and paleoenvironmental background of thefossil great ape Pierolapithecus catalaunicus (Primate hominidae). Journalof Human Evolution 55, 589–603.
Cuenca-Bescos, G, 1988. Revisión de los Sciuridae del Aragoniense y delRambliense en la fosa de Calatayud – Montalban. Scripta Geologica 87, 2–
115.Daams, R., 1981. The dental pattern of the dormice Dryomys, Myomimus,
Microdyromys and Peridyromys. Utrecht Micropaleontological Bulletins.Special Publication 3, 1–115.
Daxner-Höck, G., Höck, E., 2009. New data on Eomyidae and Gliridae(Rodentia, Mammalia) from the Late Miocene of Austria. Annalen desNaturhistorisches Museums Wien 111A, 375–444.
Engesser, B., 1999. Family Eomyidae. In: Rössner, F.G., Heissig, F.K. (Eds.),The Miocene land Mammals of Europe. Verlag Dr. Friedrich Pfeil,München, pp. 319–335.
Feru, M., Radulescu, C., Samson, P., 1979. La faune de Micromammifères duMiocène de Taut (département d’Arad). Travaux de l’Institut de Spéologie« Emil Racovitza » 18, 185–190.
Feru, M., Radulescu, C., Samson, P., 1980. La faune de Micromammifères duMiocène de Comanesti (département d’Arad). Travaux de l’Institut deSpéologie « Emil Racovitza » 19, 171–190.
Garapich, A., 2002. An overview of Miocene rodents from Bełchatów (Poland).Folia Zoologica 51 (Suppl. 1), 59–66.
Grigorescu, D., Kazár, E., 2006. A new Middle Miocene odontocete (Mam-malia, Cetacea) locality and the Sarmatian Marine Mammal Event in theCentral Paratethys. Oryctos 6, 53–67.
Gross, M., Böhme, M., Prieto, J., in press. Gratkorn – a benchmark locality forthe continental Sarmatian s.str. of the Central Paratethys. InternationalJournal of Earth Sciences (doi:10.1007/s00531-010-0615-1).
Harzhauser, M., Piller, W., 2004. Integrated stratigraphy of the Sarmatian(Upper Middle Miocene) in the western Central Paratethys. Stratigraphy1, 1–22.
Hír, J., 2006. Late Astaracian (Late Sarmatian) Lagomorphs and Rodents fromFelsotárkány – Felnémet (Northern Hungary). Beiträge zur Paläontologie30, 155–173.
Hír, J., Kókay, J., 2004. Middle Miocene molluscs and rodents from Mátraszo-
los (Mátra Mountains, Hungary). Fragmenta Palaeontologica Hungarica 22,83–97.
Hír, J., Kókay, J., 2009. Middle Miocene Molluscs, Lagomorphs and Rodents ofFelsotárkány 1 and Felsotárkány 2. Fragmenta Palaeontologica Hungarica27, 81–89.
Hír, J., Venczel, M., 2005. New Middle Miocene vertebrate localities fromSubpiatra (Bihor County, Romania). Acta Palaeontologica Romaniae 5,211–221.
Hír, J., Kókay, J., 2010. A systematic study of the middle-late Miocene rodentsand lagomorphs (Mammalia) of Felsotárkány 3/8 and 3/10 (NorthernHungary). Geodiversitas 32, 307–329.
Istocescu, D., 1971. Geologisches Studium der West Zone des Crisul Alb-Beckens und ihre Umrahmung im Codru – und Highis – Gebirge. Studiitechnice si Economice, Institutul de Geologie si Geofizica, Bucuresti. SeriesJ8 1–201.
Istocescu, D., Istocescu, F., 1974. Geological considerations on the Neogenedeposits from Crisurilor Basin. Studii si Cercetari Geologice, Geofizice,Geografice, seria Geologie Bucuresti 19, 115–128.
Kälin, D., 1999. Tribe Cricetini. In: Rössner, G., Heissig, K. (Eds.), TheMiocene land Mammals of Europe. Verlag Dr. Friedrich Pfeil, München,pp. 373–387.
Kälin, D., Engesser, B., 2001. Die jungmiozäne Säugetierfauna vom Nebel-bergweg bei Nunningen (Kanton Solothurn, Schweiz). SchweizerischePaläontologische Abhandlungen 121, 1–61.
McNulty, K., Radulescu, C., Samson, P., Feru, M., Delson, E., 1999. Morphol-ogy and identification of a previously undescribed catarrhine tooth from themiddle Miocene of Romania. American Journal of Physical Anthropology28, 199.
Mein, P., 1999. European Miocene Mammal Biochronology. In: Rössner,G.E., Heissig, K. (Eds.), The Miocene land mammals of Europe. VerlagDr. Pfeil, Munich, pp. 25–38.
Mein, P., Freudenthal, M., 1971. Une nouvelle classification des Cricetidae(Mammalia, Rodentia) du Tertiaire de l’Europe. Scripta Geologica 2, 1–37.
Piller, W.E., Harzhauser, M., Mandic, O., 2007. The Middle and UpperOttnangian of the Simsee area (SE Germany): Miocene Central Para-tethys stratigraphy-current status and future directions. Stratigraphy 4,151–168.
Prieto, J., Böhme, M., Gross, M., 2010. The cricetid rodent fauna from Gratkorn(Austria, Styria): a benchmark locality for the continental Sarmatian s. str.(late Middle Miocene) in the central Paratehys. Geologica Carpathica 61,419–436.
Radulescu, C., Samson, P., 1988. Les Cricétides (Rodentia, Mammalia) duMiocène (Astarcien Supérieur) du Roumanie. Travaux de l’Institut deSpéologie « Emil Racovitza » 37, 67–78.
Rummel, M., 1998. Die Cricetiden aus dem Mittel- und Obermiozän der Türkeiunter besonderer Berückschtigung der Sickenberg/Tobien’schen Aufsamm-lung (1968–1970). Documenta naturae 123, 1–300.
Rummel, M., 1999. Tribe Cricetodontini. In: Rössner, G., Heissig, K. (Eds.),The Miocene land Mammals of Europe. Verlag Dr. Friedrich Pfeil,München, pp. 359–364.
Rzebik-Kowalska, B., Topachevsky, V.O., 1997. Insectivora (Mammalia) fromthe Miocene of Grytsiv in Ukraine. 1. Heterosoricidae Viret and Zapfe,1951. Acta Zoologica Cracoviensia 40, 237–247.
Topachevski, V.A., Nesin, V.A., Topachevsky, V., Semenov, Y.A., 1996. Theoldest locality of the Middle Sarmatian microtheriofauna (Insectivora,Lagomorpha, Rodentia) in Eastern Europe. Dopovidi NAN Ukrainy 2,107–110.
Topachevsky, V.A., Scorik, A.F., 1992. Neogene and Pleistocene primitivecricetoids of Southeastern Europe. Naukova Dumka Press, Kiev, pp. 1–242.
Ünay, E., Bruijn, H., de, Saraç, G., 2003. A preliminary zonation of thecontinental Neogene of Anatolia based on rodents. In: Reumer, J.W.F.,Wessels, W. (Eds.), Distribution and migration of Tertiary mammals inEurasia, 10. A volume in honour of Hans de Bruijn, DEINSEA, pp. 539–
547.Vangegeim, E.A., Lungu, A.N., Tesakov, A.S., 2006. Age of the Vallesian lower
boundary (Continental Miocene of Europe). Stratigraphy and GeologicalCorrelation 14, 655–667.
Venczel, M., Stiuca, E., 2008. Late middle Miocene amphibians and squamatereptiles from Taut, Romania. Geodiversitas 30, 731–763.
J. Hír et al. / Geobios 44 (2011) 215–223 223

Hír, Prieto, tiuca – Geobios Miocene rodent faunas from Com ne ti 1 and Tau
A new interpretation of the Miocene rodent faunas from Com ne ti 1 and
Tau (W-Romania)
Une nouvelle interprétation des faunes de rongeurs de Com ne ti 1 et de Tau (Roumanie de l’ouest)
János Hír, Jérôme Prieto, Emanoil tiuca
Auteur correspondant / Corresponding author : [email protected]
Article publié dans / Article published in : Geobios (http://www.sciencedirect.com/science/journal/00166995)
Please note: Geobios is not responsible for the content, quality or functionality of any supplementary materials supplied by the authors. Any queries (other than missing material) should be directed to the corresponding author for the article.
Matériel supplémentaire / Supplementary materials Supplementary text. Systematic paleontology: full paleontological description of all identified rodent taxa from Com ne ti 1 and Tau (W-Romania).
Table S1. Miodyromys aff. grycivensis Nesin and Kowalski, 1997: material and measurements (Com ne ti 1).
Table S2. Glirulus lissiensis Hugueney and Mein, 1965: material and measurements (Tau ).
Table S3. Glirudinus sp.: material and measurements (Tau ).
Table S4. Myoglis ucrainicus Nesin and Kowalski, 1997: material and measurements (Tau ).
Table S5. Muscardinus hispanicus de Bruijn, 1966: material and measurements (Tau ).
Table S6. cf. Byzantinia sp. or div. sp.: material and measurements (Com ne ti 1).
Table S7. Eumyarion sp.: material and measurements (Tau ).
Table S8. Albanensia sp.: material and measurements (Tau ).
Table S9. Miopetaurista sp.: material and measurements (Tau ).
Table S10. Blackia sp.: material and measurements (Tau ).
Table S11. Neopetes hoeckarum (de Bruijn, 1998): material and measurements (Tau ).
Table S12. Spermophilinus bredai (Von Meyer, 1848): material and measurements (Tau ).
Table S13. Eomyops oppligeri Engesser, 1990: material and measurements (Tau ).
Fig. S1. Size of the first upper and lower molars of some European Myoglis samples.
Fig. S2. Size of the first upper and lower molars of some European Muscardinus samples.
1

Hír, Prieto, tiuca – Geobios Miocene rodent faunas from Com ne ti 1 and Tau
Supplementary text. Systematic paleontology: full paleontological description of all identified rodent taxa from Com ne ti 1 and Tau (W-Romania).
NB: Figs. 1–5 refer to figures published in the printed part of this article; Figs. S1, S2 and Tables S1–S13 refer to supplementary figures and tables, respectively (this file).
Order RODENTIA Bowdich, 1821 Family GLIRIDAE Muirhead, 1819 Subfamily MYOMIMINAE Daams, 1981 Genus Miodyromys Kretzoi, 1943 Type species: Miodyromys hamadryas (F. Major, 1899). Other species included in Miodyromys: M. prosper (Thaler, 1966); M. aegercii (Baudelot, 1972); M. biradiculus Mayr, 1979; M. vagus Mayr, 1979; M. hugueneyae Agusti and Arbiol, 1989; M. praecox Wu, 1993; M. grycivensis Nesin and Kowalski, 1997. M. alter Kretzoi, 1984 is invalid. Diagnosis: See Daams and de Bruijn (1995: 44) and Daams (1999). Miodyromys aff. grycivensis Nesin and Kowalski (1997) Fig. 2(4) 1980. Peridyromys gr. hamadryas (F. Major) - Feru et al., pp. 174, 180. Type locality: Grytsiv (Ukraine). Age: Traditionally late Miocene, MN9; see section 5. Origin of the studied material: Com ne ti 1. Material and measurements: Table S1. Diagnosis: See Nesin and Kowalski (1997). Description: M1-2. Rectangular outline, three roots. The triangular anteroloph is isolated and strongly worn. The anterior centroloph is longer than the posterior one. An extra ridge is developed between protoloph and anterior centroloph. m3. The lingual and the posterior margins of the crown are damaged. Six main ridges are developed, but no extra ridge. Remarks: The validity of the genus Miodyromys is still uncertain and synonymies (with Prodryomys Mayr, 1979, Pseudodryomys de Bruin, 1966 or Peridyromys Stehlin and Schaub, 1951) are possible (Daams and de Bruijn, 1995; Freudenthal and Martín-Suárez, 2006: 17). Furthermore some species of Miodyromys, for instance M. hamadryas (Forsyth-Major, 1899) and M. aegercii Baudelot, 1972, are difficult to distinguish (e.g., Wu, 1990; Heissig, 2006). The M1-2 from Com ne ti has two distinctive features: its small size and the missing of the posterior extra ridge, making these molars close to the small-sized Miodyromys grycivensis from Ukrainian locality Grytsiv (Nesin and Kowalski, 1997). With only two specimens, the material from Com ne ti 1 is limited, and a confident determination cannot be proposed at the species level. Subfamily DRYOMYINAE de Bruijn, 1967 Genus Glirulus Thomas, 1906 Type species: Glirulus japonicus (Schinz, 1845). Other species included in Glirulus: G. pusillus (Heller, 1936), G. lissiensis (Hugueney and Mein, 1965), G. diremptus (Mayr, 1979), G. minor (Wu, 1993), G. ekremi Ünay, 1994. The synonymy Paraglirulus conjunctus Mayr, 1979 - Glirulus lissiensis is debated. Fejfar and Storch (1990) suggested that G. gemulla described by Kretzoi (1962, Hungarian locality Csarnota 2) is a junior synonym of G. pusillus. Emended diagnosis: See Van der Meulen and de Bruijn (1982). Glirulus lissiensis Hugueney and Mein, 1965 Fig. 2(3) 1979. Gliridé gen. et sp. indet. - Feru et al., p. 186. Type locality: Lissieu (France). Age: late Miocene, MN13 Origin of the studied material: Tau . Material and measurements: Table S2. Diagnosis: See Hugueney and Mein (1965). Description: M1-2. Square outline and continuous endoloph. All the five main ridges connect to the endoloph. The anterior centroloph is isolated from the paracone. Four extra ridges, one between each main ridge. m3. Subtriangular outline. Four main ridges and 4 extra ridges. Medium-sized centrolophid. The endolophid is interrupted between the centrolophid and the mesolophid.
2

Hír, Prieto, tiuca – Geobios Miocene rodent faunas from Com ne ti 1 and Tau
Remarks. The distinction between Paraglirulus and Glirulus is very problematic, and, as a result, it is sometime difficult to recognize species because of morphological stability through time (e.g., Daxner-Höck and de Bruijn, 1981; de Bruijn, 1998: 117; Daxner-Höck, 2005). This is especially true for Paraglirulus conjunctus and Glirulus lissiensis. The two species are synonym after Daxner-Höck and de Bruijn (1981), Daxner-Höck and Höck (2009) and Kowalski (1997), but this proposal is not followed by Daams and de Bruijn (1995) and Daams (1999). Mayr (1979: 287) distinguishes G. lissiensis from Paraglirulus conjunctus by the organisation of the fully molarised premolars with labially isolated anterior centroloph and the absence of a continuous endolophid in the lower molars. In the m3 from Tau , centrolophid and mesolophid are not linked. Thus the Glirulus remains belong to G. lissiensis. Occurs G. lissiensis, in the Late Astaracian - Early Vallesian of East and Central Europe: Rudabánya, MN9 (Daxner-Höck, 2005); Schernham, MN10 (Daxner-Höck, 2004a); Richardhof-Golfplatz, MN9 (Daxner-Höck, 2005); Richardhof-Wald, MN9 (Daxner-Höck, 2005); Borsky Sväty Jur, MN9 (Sabol et al., 2004); Be chatów A, MN9 (Kowalski, 1997; Garapich, 2002); Subpiatra 2/2, MN7/8 (Venczel et al., 2005; Hír and Venczel, 2005); Fels tárkány 2, MN7/8 (Hír and Kókay, 2009); Fels tárkány 3/8, MN9 (Hír and Kókay, 2010). Subfamily GLIRINAE Thomas, 1897 Genus Glirudinus de Bruijn, 1966 Type species: Glirudinus gracilis (Dehm, 1950). Other species included in Glirudinus: G. glirulus (Dehm, 1935); G. modestus (Dehm, 1950); G. undosus Mayr, 1979; G. euryodon Van der Meulen and de Bruijn, 1982; G. minutus Wu, 1993; G. engesseri Ünay, 1994; G. haramiensis Ünay, 1994; G. eggingensis Werner, 1994; G. antiquus Vianey-Liaud, 2003; G. intermedius Aguilar and Lazzari, 2006; G. magnus Aguilar and Lazzari, 2006. Diagnosis: See de Bruijn (1966). Glirudinus sp. Fig. 2(2) Origin of the studied material: Tau . Material and measurements: Table S3. Description: m2. The lingual part of the crown is slightly damaged. The tooth is almost rectangular in occlusal view. Thirteen thin enamel ridges are developed (main ridges: 1th, 4th, 6th, 8th, 10th, 12th and13th). Only the first ridge is transversal to the longitudinal axis of the molar, while the others are inclinated (~30° angle). The posterior four main ridges are lingually connected. The 2nd, 3rd, 5th, 7th and 9th ridges are ligual, while the tiny 11th extra ridge is central. Remarks: This m2 is inadequate for a species determination. Its size refers to a panel of species including G. minutus (Wu, 1993), G. antiquus (Vianey-Liaud, 2003) and G. intermedius (Aguilar and Lazzari, 2006). The long biostratigraphic range of Glirudinus starts in the Lower Oligocene MP23 (Vianey-Liaud, 2003) and extends traditionally to the end of the Middle Miocene (Daams, 1999). Aguilar et al. (2004) report G. undosus in the MN9 fauna of Jujurieux 1. Glirudinus was unknown from the Miocene of the Carpathian Basin. Genus Myoglis Baudelot, 1965 Type species: Myoglis meini (de Bruijn, 1966). Other species included in Myoglis: M. truyolsi (Daams, 1976); M. antecedens Mayr, 1979; M. ucrainicus Nesin and Kowalski, 1997; M. houlezi Aguilar et al., 1997. Diagnosis: See Baudelot (1965). Myoglis ucrainicus Nesin and Kowalski, 1997 Fig. 2(7–11) 1979. Myoglis meini (de Bruijn) - Feru et al., pp. 186, 188. Type locality: Grytsiv (Ukraine). Age: Traditionally late Miocene, MN9; see chapter 5. Discussion. Origin of the studied material: Tau . Material and measurements: Table S4. Diagnosis: See Nesin and Kowalski (1997). Description: M1. The antero-labial part of the tooth is damaged. Six main ridges and two short labial extra ridges (respectively between protoloph and anterior centroloph, and between meta- and posteroloph). Protoloph and metaloph do not connect lingually. Four roots. p4. Triangular outline with rounded angles and four transversal ridges. The anterolophid is a shortest crest. All ridges are isolated. One root. m1. Rectangular outline with narrower anterior part. Four transversal ridges (antero-, meta-, meso- and posterolophid) and four extra ridges. Two lingually-connected extra ridges are developed between antero- and
3

Hír, Prieto, tiuca – Geobios Miocene rodent faunas from Com ne ti 1 and Tau
metalophid. The long posterior extra ridge connects to the posterolophid on the lingual border of the m1. Long and centrally-placed extra ridges between meta- and mesolophid. m2. Rectangular outline. Four main ridges and five extra ridges. Three of the extra ridges are developed on the lingual part of the molar (between antero- and metalophid). The two last extra ridges are long and isolated, and developed in the two posterior synclines. Three roots. m3. Subrectangular outline with rounded posterior margin and three roots. The occlusal surface is worn. Remarks: The Myoglis teeth refer to M. ucrainicus because of the following characters: 1) four-rooted M1 without connection between proto- and metaloph, 2) m1 and m2 without accessory ridge behind the posterolophid, 3) the molar size fits very well with the Ukrainian species (Fig. S1). Beside the type locality Grytsiv, M. ucrainicus is also reported from Hungarian and Austrian deposits (Daxner-Höck, 2005; Daxner-Höck and Höck, 2009). M. cf. ucrainicus molars from Rudabánya are larger than the type sample, and inversely the molar from Austria are clearly smaller. Furthermore the molars from Schernham (youngest locality recording Myoglis) are often smaller, especially with regards to the teeth from Götzendorf (oldest locality). Size increase of this species through time is suspected, but this proposal is in contradiction to the large-sized molar from Rudabánya, a locality which is definitively older than Schernham. Up to date, M. ucrainicus is not recorded before the Late Miocene, but this statement depends on the dating of Grytsiv (see comments in the section 5). Subfamily LEITHIINAE Lydekker, 1896 Genus Muscardinus Kaup, 1829 Type species: Muscardinus avellanarius (Linneaus, 1758). Other species included in Muscardinus (subspecies are not taken in account): M. sansaniensis (Lartet, 1851); M. dacicus Kormos, 1930; M. pliocaenicus Kowalski, 1963; M. davidi Hugueney and Mein, 1965; M. vireti Hugueney and Mein, 1965; M. thaleri de Bruijn, 1966; M. hispanicus de Bruijn, 1966; M. vallesiensis Hartenberger, 1966; M. heinzi Aguilar, 1981-1982; M. cyclopeus Agusti, Moyá-Solá and Pons-Moyá, 1982; M. helleri Fejfar and Storch, 1990; M. malatestai Gliozzi, 1995; M. meridionalis García-Alix, Minwer-Barakat, Martín-Suárez and Freudenthal, 2008. M. topachevskii Nesin and Kowalski, 1997 is considered as junior synonym of M. hispanicus by Daxner-Höck and Höck (2009: 400). Diagnosis: See Ellerman (1941). Muscardinus hispanicus de Bruijn, 1966 Figs. 2(5, 6) and 7 1979. Eomuscardinus gr. sansaniensis (Lartet) - Feru et al., pp. 186, 188. Type locality: Pedregueras II (Spain). Age: late Miocene, MN9. Origin of the studied material: Tau . Material and measurements: Table S5. Diagnosis: See de Bruijn (1966). Description: M1. Longitudinally elongated and rectangular outline. The molars have seven enamel ridges. The first ridge (anteroloph) ends free on both sides of the teeth. The second, fourth, sixth, and seventh ridges are long and lingually connected. The third ridge is short and labial. The fifth ridge is variable (short or reaching the labial border of the molar). m1. Rectangular outline. The anterior margin is narrower than the posterior part of the m1. The molar has seven enamel ridges. The first and second ridges, as well as the sixth and seventh ridges, are lingually and labially connected. The fifth ridge is lingually connected to the sixth and seventh ridges. The fourth ridge is short and labial. Remarks: The molars from Tau are relatively small, and certainly not belong to the larger species M. vallesiensis. M. sansaniensis-related molar samples are also excluded because of their less elongated and more complex molars. M. thaleri M1/m1 are also less elongated. The size range of the teeth from Tau is included within the lower part of the size variation of M. pliocaenicus austriacus Bachmayer and Wilson, 1970 (Fig. S2), which is an intermediate form between M. hispanicus and M. pliocaenicus pliocaenicus (Daxner-Höck and Höck, 2009: 404). Thus the Romanian sample bears a close resemblance to M. hispanicus. Nesin and Kowalski (1997) described the species M. topachevskii from Grytsiv. They noticed: “From M. hispanicus, the species closest to [M. topachevskii] in morphology and dimensions, the Ukrainian form differs in a higher number of ridges on P4 and lower on m1 (five instead of six). Besides, M. hispanicus has three roots on m2, whereas M. topachevskii has two”. The large Austrian M. hispanicus record shows the morphologic variability of the species (Daxner-Höck and Höck, 2009): P4 have maximum five ridges, m1 have 6 main ridges. The m2 have two roots. In these characters the Ukrainian and Austrian samples are basically identical. M. topachevskii is different at least in the reduction of the 2nd ridge in the m1, and considered as valid species. As the m1 from Tau has a full 2nd ridge, and the M1 has a small anterior extra ridges and a long posterior centroloph, the Romanian molars are assigned to M. hispanicus.
4

Hír, Prieto, tiuca – Geobios Miocene rodent faunas from Com ne ti 1 and Tau
M. hispanicus record: The first evidence of the species is found in the Spanish latest Middle Miocene [Vallès-Penedès, Megacricetodon ibericus + Democricetodon crusafonti zone (Casanovas-Vilar, 2007, Casanovas-Vilar et al., 2008); upper part of the zone G3 of the Calatayud-Daroca and Teruel basins, MN8 sensu Mein and Ginsburg (2002)]. In Austria the species is not known before the Late Miocene, but it has to be stressed that the small mammal record is poor for the Sarmatian. The new late Middle Miocene locality of Gratkorn (Styria, 12-12.5 My; Gross et al., 2007a, 2007b, in press; Harzhauser et al., 2008; Prieto et al., 2010a, 2010b) contains M. aff. sansaniensis (Daxner-Höck, 2010). This record fit well with the occurrence of M. sansaniensis in Barranc de Can Vila 1 (Vallès-Penedès basin, 12-12,5 My; Casanovas-Vilar et al., 2008). In the North Alpine Foreland Basin, M. hispanicus is recorded from Marktl, Hammerschmiede 1 (as M. pliocaenicus hispanicus in Mayr, 1979, Daxner-Höck and Höck, 2009) and 3, Hillenloh (Jung and Mayr, 1980) and Nebelberg TGL II (as M. cf. crusafonti in Kälin and Kempf, 2009). These localities are traditionally correlated to MN9 because of Microtocricetus. On the other hand Prieto and Rummel (2009a) described the cricetid rodent lineage Collimys hiri-C. dobosi. The primitive Collimys hiri is suggestive of an older relative age of Hammerschmiede than Fels tárkány 3/2 and Hillenloh (both C. dobosi), and the German localities could be Middle Miocene. Kälin and Engesser (2001) report Muscardinus sp. (as Eomuscardinus aff. sansaniensis in Kälin and Kempf, 2009) in Nebelbergweg. Daxner-Höck (2010) assigns the M1 to M. aff. sansaniensis, and proposes a correlation to the Middle Miocene (MN7+8) for the locality. In our opinion the fragmentary tooth is less complicated than M. aff. sansaniensis from Anwil, and does not differ from the M. hispanicus German material. The lack of sufficient material in Nebelbergweg does not permit a confident determination at the species level. While the Badenian Muscardinus samples from Hungary (Mátrasz l s, Sámsonháza) can be easily related to M. sansaniensis, the younger samples – originally also related to the former species – deserve comments. A single tooth from T ad is tentatively assigned to M. aff. sansaniensis (Hír et al., 2001). Hír (2003) divides the Muscardinus sample from Felsötárkány 3/2 into two species, the larger one being assigned to M. aff. sansaniensis. The smaller and simpler species is not determinated. Actually the size of the larger molars fits better the sample from Grytsiv. The molars from Scherham illustrate how great can be the size variability. We cannot, because of lack of material, definitively conclude on the homogeneity of the sample from Felsötárkány 3/2. Similarly the two m1 from Fels tárkány-Felnémet (Hír, 2006) differs from M. aff. sansaniensis in their complete 2nd ridge and the isolated 3rd ridge. Family MURIDAE Illiger, 1811 Subfamily CRICETODONTINAE Schaub, 1925 Tribe COPEMYINI Jacobs and Lindsay, 1984 Genus Democricetodon Fahlbusch, 1964 Type species: Democricetodon crassus Freudenthal, 1969 [= D. minor (Lartet, 1851) sensu Fahlbusch, 1964]. Other species included in Democricetodon (here we limit the list of the species included in Democricetodon fide Freudenthal, 2006): Democricetodon affinis (Schaub, 1925); D. brevis (Schaub, 1925); D. gaillardi (Schaub, 1925); D. vindobonensis (Schaub and Zapfe, 1953); D. romieviensis (Freudenthal, 1963); D. sulcatus Freudenthal, 1964; D. freisingensis Fahlbusch, 1964 (considered by Theocharopoulos in de Bruijn et al., 2003 as junior synonym of D. gaillardi); D. gracilis Fahlbusch, 1964; D. mutilus Fahlbusch, 1964; D. hispanicus Freudenthal, 1964; D. franconicus Fahlbusch, 1966; D. cretensis de Bruijn and Meulenkamp, 1972; D. nemoralis Agustí, 1981; D. kohatensis Wessels et al., 1982; D. hasznosensis Kordos, 1986; D. walkeri Tong and Jaeger, 1993; D. lindsayi Qiu, 1996; D. tongi Qiu, 1996; D. hanniae Aguilar et al., 1999; D. sudrei Aguilar et al., 1999; D. moralesi van der Meulen et al., 2004; D. zarandicus Radulescu and Samson, 1988; D. fourensis Maridet et al., 2000; D. doukasi Theocharopoulos, 2000; D. anatolicus Theocharopoulos, 2000. The statu of D. iazygum R dulescu and Samson, 1988 is discussed below. Diagnosis: See Fahlbusch (1964). Democricetodon zarandicus R dulescu and Samson, 1988 Fig. 4(1–14) 1979. Democricetodon gr. gaillardi (Schaub) - Feru et al., pp. 186, 188. 1980. Democricetodon gr. gaillardi-brevis - Feru et al., p. 181. 1988. Democricetodon zarandicus nov. sp. - R dulescu and Samson, pp. 74–78, fig. 3. Type locality: Tau (Romania). Age: see section 5. Material and measurements: see R dulescu and Samson, 1988. Original diagnosis (R dulescu and Samson, 1988, translated from French): species of size intermediate between D. brevis and D. freisengensis. m1 with simple anteroconid; the long mesolophid reaches the labial border in m1-3; M1 with anterocone subdivided into two cuspids (apical and mesial furrow, this character is attenuated by wear process); very developed anteromesoloph [éperon labial] connecting the crest which closes the anterosinus; long mesoloph which does not extends to the labial border; M2 with double protolophule and mesoloph similar to M1.
5

Hír, Prieto, tiuca – Geobios Miocene rodent faunas from Com ne ti 1 and Tau
Emended diagnosis: medium-sized Democricetodon species; m1 elongated, almost as long as M1 in mean size, with simple anteroconid; lower molars with long mesolophid; anterocone of the M1 at least superficially subdivided; M1 with complete anteromesoloph, medium-sized mesoloph, protolophule posterior or double (anterior then less developed) and posterior metalophule; M2 and M3 with double protolophule. Description: M1. The anterocone is simple (8 out of 11 molars) or biparted (3). The labial and lingual branches of the anteroloph are equally developed. A double metalophule is present (8) or only the posterior metalophule is developed (3). The mesoloph is long but often does not reach the lingual margin (9). In two M1 the mesoloph is complete. M2. The labial anterolophule is slightly better developed than the lingual one. Double protolophule. Mesoloph long, but it does not extend to the labial margin. The metalophule is posterior. M3. The labial anterolophule is slightly better developed than the lingual one. The protolophule is double. The mesoloph can be either short (2 out of 8), long and does not reach the labial margin (1), long and extends to the posterior basis of the paracone (3), or it reaches the labial margin (2). Hypocone and metacone are reduced, but are distinct cusps. m1. The anteroconid is simple. Lingual and labial anterolophulids are equally developed (3 out of 5) or only the labial branch is observed. The anterolophulid connects either to the anteroconid (2 out of 5) or to the labial branch of the anterolophulid. In one molar the anterolophulid has two branches. A short ectomesolophid is developed in one specimen (Tt 0149). Remarks: In the differential diagnosis of D. zarandicus provided by R dulescu and Samson (1988), only the two species D. brevis and D. freisingensis are considered. Therefore a more in-depth analyze is necessary. In the following we restrict the comparison with the Democricetodon species having M1 with at least partial division/fissuration of the anterocone and well developed mesoloph(id)s. • Democricetodon freisingensis and D. gaillardi: The taxonomic status of the two species is still a subject of debate because they differ in small details only (e.g., Heissig, 1995; Maridet, 2003; de Bruijn et al., 2006). The following comparison considers the two species under a single paragraph, but we do not synonymize them. D. freisingensis was defined based on the German material from Giggenhausen (Fahlbusch, 1964), reported also from Kleineisenbach (Prieto, 2007) and Anwil (Switzerland, Engesser, 1972). The proposal of Maridet (2003), assigning the Anwil sample to D. gaillardi, is not followed because there is in our opinion no differences between this sample and the molars of the type locality. The molars of D. freisingensis are morphologically highly variable, but the sample from Tau cannot be assign to this species at least because: the molars are most often smaller; the ratio LM1/Lm1 is lower; the lingual anteroloph is less developed in the M1: in Tau this crest is a relatively straight, whereas in D.
freisingensis the crest is rounded and most often fused with the anterocone; in the m1, the metaconid is close to the anteroconid: the molars from Tau have a clear valley between the
two cuspids, and the metalophid is directed transversally to forwards. In D. freisingensis, the anterior wall of the metaconid often connects to the anteroconid, and closes the anterosinusid; the metalophid is often strongly angular.
• D. gaillardi: Heissig (1995: 90) concluded that the lectotype of the species must, because of the rule of priority, belongs to the German locality Steinheim, and not as proposed by Baudelot (1972) to the French locality Sansan [sample without certain specific assignment in Heissig (1995: 93), tentavely assigned to D. aff. freisingensis]. The sample from Tau cannot be assigned to the species from Steinheim at least because: the molars are smaller; the lingual anteroloph is less developed in the M1; the anterior protolophule is less developed in the M1; The sample from Tau cannot be assigned to the species from Sansan because: the molars are most often smaller; the ratio LM1/Lm1 is lower; the anteroconid of the m1 is simple. • Democricetodon affinis: The type sample from La Grive M was poorly described, and Heissig (1995: 91) related this species to the relative large molars with simple morphology from Steinheim. Recently, Maridet (2003) intensively studied the type material, and showed the great morphological variability of the species. The sample from Tau cannot be assigned to D. affinis at least because: the molars are most often smaller; the ratio LM1/Lm1 is lower; the anterocone is central, and at least superficially divided in the M1; the lingual cusp of the anterocone is always present and do not fuse with the anteroloph; the mesoloph(id)s are longer; the anteromesoloph is complete in the M1.
6

Hír, Prieto, tiuca – Geobios Miocene rodent faunas from Com ne ti 1 and Tau
• Democricetodon brevis: Maridet (2003) provided an excellent description of the material from the type locality La Grive M. The sample from Tau cannot be assigned to D. brevis at least because: the ratio LM1/Lm1 is lower; the anteromesoloph is better developed; a well developed ectomesolophid is missing in m1-2 (but the material from Tau is limited); the anterolophulid often connects to the labial anterolophid in the m1; this crest is most often straight or
connects lingually in La Grive M. • Democricetodon nemoralis: This Spanish species is close to D. brevis, and some authors consider it as a subspecies (e.g., Casanovas-Vilar, 2007). It is characterized by a well developed mesoloph(id) and the subdivion of the anteroconid in some m1. The last character is unknown in Tau , and the m1 are relatively longer. • Democriceton vindobonensis: The species from Neudorf Fissures 1 and 2 (Fejfar, 1974) differs from D. zarandicus in the narrower M1 and the somewhat smaller m1. The mesoloph of the M1 is complete in all 6 M1 from Neudorf, the anterocone less fissured, and the anteromesoloph may be absent. The anteroconid of the m1 is shorter, most often close to the metaconid, and the mesolophid may not reach the border of the tooth. • Democricetodon sulcatus: The species from Pedregueras IIC is clearly smaller than D. zarandicus, and the mesolophid is clearly shorter. Furthermore a double anterolophule may be developed in D. sulcatus (Freudenthal, 1967: pl. 1, fig. 9 and 10). • Democricetodon lindsayi: The species from Tunggur (China) has less elongated m1, superficial fissuration of the anterocone and shorter anteromesoloph in the M1. • Democricetodon kohatensis: The Pakistanis species is larger (m1 excepted) and lacks complete anteromesoloph in the M1. The species from Tau is clearly less variable. • D. cretensis is defined on a small molar sample without m1, m2 and M3 (de Bruijn and Meulenkamp, 1972). A comparison with Tau is tentative. The notable difference is the absence of an anteromesoloph in D. cretensis. • Democricetodon walkeri from Kenya is out of consideration because it is extremely large (Tong and Jaeger, 1993). In view of these differences, it is reasonable to retain the species Democricetodon zarandicus. Beside the type samples considered above, some Democricetodon records deserve comments. We restrict the analysis to samples from Central Europe to Anatolia. • Democriceton samples from the North Alpine Foreland basin (NAFB): Democricetodon cf. brevis: This species is found e.g. in Anwil (Engesser, 1972), Giggenhausen and
Kleineisenbach (Fahlbusch, 1964; Prieto, 2007). D. cf. brevis differs from D. zarandicus in the slightly smaller upper molars, and clearly smaller lower molars. The labial cusp of the anterocone in the M1 from the NAFB (comparison: Kleineisenbach, Giggenhausen) is most often high, whereas the lingual one is fused in the long and rounded anteroloph;
Democricetodon nov. sp. from Nebelbergweg: After Kälin and Engesser (2001: 30) this Democricetodon differs from the former D. cf. brevis from Anwil in the less often developed of the anteromesoloph (M1) and the missing mesolophid in some m2. We add that the anterocone is much divided in the M1, and the lingual anteroloph is shorter;
Democricetodon zarandicus differs from the species from Nebelbergweg in the smaller upper molars, in the complete anteromesoloph in M1, and in the complete mesolophid.
• Austrian Democricetodon: Samples related to the species from Nebelbergweg are also known from Götzendorf (Pannonian; Bachmayer and Wilson, 1984) and Gratkorn (Sarmatian; Prieto et al., 2010a, Gross et al., in press). The Democricetodon molars from Vösendorf (Pannonian; Daxner-Höck 1972, as Kowalskia sp.) are slightly larger than the Swiss molars. • Democricetodon-record from Romania: The small molar sample of D. brevis from T ad (Sarmatian; Hír et al., 2001) is very similar to D. zarandicus. The figured M1 (Hír et al., 2001: pl. III, fig. 5) differs from the molars from Tau in the double anterolophule and the longer mesoloph. Megacricetodon aff. germanicus is also described in the Romanian locality on the basis of a single M1 (Hír et al., 2001: pl. IV, fig. 3). This molar belongs in fact to Democricetodon, and shows that the length of the anteromesoloph is variable in T ad. Different Democricetodon species are described from two layers of Subpiatr (Hír and Venczel, 2005; Venczel et al., 2005). The older layer of the locality contains D. brevis. The two M1 have short to complete anteromesoloph. Based on the limited material the taxonomy is unclear. In the younger layer 2/2, D. freisingensis is represented by two M1 and one m1. Hír and Venczel (2005) exclude D. zarandicus because of, in this species, the division of the anterocone in the M1, the anteroconid-protoconid connection, as well as the lack of an ectomesolophid in the m1. On the view of the new description of the material from Tau , there is no clear argumentation not to assign the molars from Subpiatr 2/2 to a species morphologically close to D. zarandicus. • Czech Republic and Slovak Republic: Sabol et al. (2004) assign the Badenian Democricetodon from Bonanza (Slovak Republic) to D. vindobonensis. D. cf. freisingensis is reported from Suchomasti (Czech Republic, Fejfar, 1990; Kretzoi and Fejfar, 2005). The species is equivalent to the Democricetodon from Rudabánya. The Democricetodon from Borský Svätý Jur (Slovak Republic; Sabol et al., 2004; Joniak, 2005) is not discussed here
7

Hír, Prieto, tiuca – Geobios Miocene rodent faunas from Com ne ti 1 and Tau
because the possibility that the sample represent two species cannot be ignored. The association Kowalskia + Democricetodon is known from Suchomasti and Rudabánya. • Hungarian Democricetodon record: Kretzoi and Fejfar (2005) reports D. cf. freisingensis in Rudabánya. A comparison with Tau is difficult because Kowalskia cf. schaubi also occurs in the locality, and is similar in size to Democricetodon. The authors conclude that D. cf. freisingensis is similar to Democricetodon nov. sp. from Nebelbergweg, but we refrain at present to accept this interpretation. Anyway the ratio length LM1/Lm1 is higher in Rudabánya and the lower molars are larger. Furthermore the anteromesoloph is rarely complete (18%) and may be absent (50%) in Rudabánya. Hír (2006) describes D. brevis and Megacriceton germanicus in Felsötárkány-Felnémet (Sarmatian). These samples are assigned to Democriceton nov. sp. from Nebelbergweg by Prieto and Rummel (2009a), and fit well in size with D. zarandicus. The anteromesoloph is less developed than in the M1 from Tau . D. cf. freisingensis from the late Badenian of Mátrasz l s 2 (Gál et al., 2000; Hír and Kókay, 2004) is very large and thus cannot be confused with D. zarandicus. The second Democricetodon species present in the locality is smaller and assigned to D. mutilus, but the molar sample is limited (5 molars, no m1). The M1 is narrower than in the molars from Tau and a complete anteromesoloph is missing. The Hungarian species cannot be assigned to D. vindobonensis because of the medium-sized mesoloph in the M1. D. hasznosensis from Hasznos and Szentendre (Badenian; Kordos, 1986) cannot be confused either with D. zarandicus nor D. vinobonensis. • Poland: Kowalski (1967) listed D. gaillardi in Opole 1 and 2, Democricetodon cf. vindobonensis and D. gracilis in Be chatów A. Based on the figures provided by Kowalski (1967), Prieto and Rummel (2009a: 245) recognize Collimys rather than Democricetodon. The molars from Be chatów A are unfortunately insufficiently described or figured, but Garapich (2002) listed a large D. cf. brevis and Democricetodon nov. sp. • Anatolian Democricetodon: Acording to Ünay et al. (2003) the genus disappears from the Antolian fossil record during the late Middle Miocene zone H. The youngest Democricetodon samples are published in detail and discussed here: D. aff. gaillardi is well represented in the sample from Çandir 2 (zone F; de Bruijn et al., 2003). The molars are slightly larger than those of D. zarandicus, with higher M1/m1 ratio. In other morphological characteristics the species from Çandir 2 does not differ basically from the Romanian species. D. brevis from Pa alar (zone F; Peláez-Campomanes and Daams, 2002) differs from D. zarandicus in the slightly larger M1-2, and the smaller m1. In the description of the M1 it is stated that the “anterocone is simple and kidney-shaped”. In fact one figured molar (fig. 1(A), 1 out of three figured M1) has the anterocone with two distinct cusps. The anteromesoloph, although most often present in the molars from Pa alar, is shorter. Similarly the mesolophid is long but does not reach the lingual border of the m1. D. freisingensis von Yukari-Kiliza is poorly documented (probably zone H, because Byzantinia cariensis is present in this locality; Rummel, 1998). The fragmentary M1 has a medium-sized anteromesoloph. The fragmentary m1 has a complete mesolophid. Democricetodon sp. from Sofça (zone H; Rummel, 1998) is clearly larger than D. zarandicus. Furthermore the anteromesoloph of the M1 is less developed, the anteroconid of the m1 may be bifid, and the mesolophid is very short. As a conclusion it can be shown that D. zarandicus cannot be confidently recognized outside the type locality, but a panel of molar samples differ from the Romanian species in just slight morphologic aspects. A phylogenetic interpretation is at present hazardous because shape and morphology of the rodent molars might change along geographic and climatic clines (McGuire, 2010). Democricetodon cf. freisingensis Fahlbusch, 1964 vel. cf. gaillardi (Schaub, 1925) 1980. Democricetodon gr. gaillardi - R dulescu and Samson, pp. 71–74, fig. 2. 1988. Democricetodon iazygum nov. sp. - R dulescu and Samson, pp. 71–74, fig. 2. Origin of the materiel: Com ne ti 1. Remarks: R dulescu and Samson (1988) described Democricetodon iazygum nov. sp., although the type material is restricted to only three intact molars. The M3, m2, m3 are completely unknown, and there is a posterior fragment of a m1. The most important morphological characters of the differential diagnosis are the divided anterocone, the short anteromesoloph, the absence of an anterior protolophule, and the long but not complete mesoloph. The absence of the ectomesolophid is also noticed in the m1. This morphology is found in Democricetodon freisingensis-related species from the NAFB (Fahlbusch, 1964; Engesser, 1972; Kälin et al., 2001; Prieto, 2007). The division of the anterocone in the M1 figured by R dulescu and Samson (1988) can be observed in some molars from Anwil (Maridet, 2003), but is not evident in Germany (Kleineisenbach, Giggenhausen). The species from Com ne ti cannot be confused with D. fourensis because the positioning of the metacone of the M1 (Maridet et al., 2000). de Bruijn et al. (2006) report D. aff. gaillardi (with D. gaillardi = D. freisingensis) in the Turkish locality Çandir. The molars are in size and morphology close to the Com ne ti 1 sample: the anterocone is often more or less bifid (de Bruijn et al., 2006: pl. 6, fig. 1), the labial spur of the anterolophule is developed, and the mesoloph medium-sized. We consider that the material from Com ne ti 1 is too limited, and D. iazygum is a nomen dubium. Genus Megacricetodon Fahlbusch, 1964 Type species: Megacricetodon gregarius (Schaub, 1925).
8

Hír, Prieto, tiuca – Geobios Miocene rodent faunas from Com ne ti 1 and Tau
Other species included in Megacricetodon: M. minor (Lartet, 1851); M. bourgeoisi (Schaub, 1925); M. ibericus (Schaub, 1944); M. collongensis (Mein, 1958); M. lappi (Mein, 1958); M. crusafonti (Freudenthal, 1963); M. primitivus (Freudenthal, 1963); M. bavaricus Fahlbusch, 1964; M. similis Fahlbusch, 1964; M. minutus Daxner, 1967; M. bezianensis Bulot, 1980; M. germanicus Aguilar, 1980; M. gersii Aguilar, 1980; M. sinensis Qiu et al., 1981; M. lopezae Garcia Moreno, 1986; M. roussilonensis Aguilar et al., 1986; M. rafaeli Daams and Freudenthal, 1988; M. fournasi Aguilar, 1995; M. lemartinelli Aguilar, 1995; M. pusillus Qiu, 1996; M. fahlbuschi Aguilar et al., 1999; M. wuae Aguilar et al., 1999; M. lalai Aguilar et al., 1999; M. andrewsi Pelaez-Campomanez and Daams, 2002; M. tautavelensis Lazzari and Aguilar, 2007; M. aunayi Lazzari and Aguilar, 2007; M. yei Bi et al., 2008. M. aguilari Lindsay, 1988; M. sivalensis Lindsay, 1988; M. daamsi Lindsay, 1988 and M. mythikos Lindsay, 1988 are excluded from Megacricetodon by Wessels (1996). M. debruijni Freudenthal, 1968 is considered by Joniak (2005) as junior synonym of M. minutus. The validity of M. crisiensis R dulescu and Samson, 1988 is discussed in the following. Diagnosis: See Fahlbusch (1964). Megacricetodon cf. minutus (Daxner-Höck, 1967) 1980. Megacricetodon gr. similis (Fahlbusch) - Feru et al., p. 174. 1988. Megacricetodon crisiensis nov. sp. - R dulescu and Samson, pp. 67–71, fig. 1. Type locality: Brunn-Vösendorf (Austria). Age: late Miocene, MN9. Origin of the studied material: Com ne ti 1. Diagnosis: See Daxner-Höck (1967). Description: See R dulescu and Samson (1988). Remarks: The main characters defining M. crisiensis are the small size, the bifid anterocon(id) of the first molars, and the reduction of the posterior arm of the protocone in the M2 (R dulescu and Samson, 1988). We can add that the anterolophule of the M1 is double. Small-sized Megacricetodon species are reported from a wide range of localities. Their affiliation is often controversial, and the scope of this paper is not a revision of these forms. Thus we restrict our analysis to Central/East European and Anatolian species. M. similis and M. cf. minor from the NAFB (Fahlbusch, 1964; Engesser, 1972) and M. aff. similis from the Frankian Alb (Prieto, 2007; Prieto and Rummel, 2009b) can be easily excluded. The indeterminate species from Mörgen (Germany, MN8; Seehuber, 2009) is of interest because the anteroconid of the m1 is clearly divided. The species cannot be discussed under these lines because the taxonomic homogenity of this Megacricetodon sample is questionable. The M1 from Hasznos do not have a double anterolophule and the anteroconid of the m1 is rarely bifid (Kordos, 1986). M. andrewsi from Pa alar do not have a double anterolophule in the M1, and the anteroconid of the m1 is not well divided (Peláez-Campomanes and Daams, 2002). Similarly M. collongensis from Çandir has a simple anterolophule in the M1 (de Bruijn et al., 2003). The morphology of the M. crisiensis M2 is observed in M. minor from Nebelbergweg (Kälin and Engesser, 2001), but the m1 do not have a bifid anteroconid in the Swiss locality. In Hammerschmiede 1 (Mayr and Fahlbusch, 1975; unpublished material) some m1 have this structure, and the anterolophule of the M1 is Y-shaped to double. The German, Swiss and Romanian samples belong to a similar species. Prieto et al. (2010a) assign the species from the NAFB to M. minutus. Recently Joniak (2005) discussed intensively taxonomy of the Megacricetodon from Borský Sv. Jur and reevaluated M. minutus (= M. debruijni). He underlined the poorness of the sample in the M. minutus type locality. Therefore the comparison with M. crisiensis – which is also poorly documented – is extremely tentative. The two species might be separated at the species level, but the observation made on larger samples show that the differences can also be interpreted as intraspecific variations. A deeper analyzis of the Central and East European remains is necessary to conclude on the validity of M. minutus (nomen dubium?), and at present we choose to assign the species from Com nesti 1 to M. cf. minutus, and M. crisiensis is a junior synonym. In Central/East Europe, M. minutus is recorded in diverse MN8-MN9 localities from Austria (Daxner-Höck, 2004b, Prieto et al., 2010a), Germany (Mayr and Fahlbusch, 1975; Jung and Mayr, 1980), Switzerland (Kälin and Engesser, 2001) and Hungary (Hír and Kókay, 2009). Tribe CRICETODONTINI Schaub, 1925 Genus Byzantinia de Bruijn, 1976 Type species: Byzantinia pikermiensis de Bruijn, 1976. Other species included in Byzantinia: B. hellenicus (Freudenthal, 1970); B. nikosi de Bruijn, 1976; B. eskihisarensis (Tobien, 1978); B. sofcaensis (Tobien, 1978); B. dardanellensis Ünay, 1980; B. bayraktepensis Ünay, 1980; B. ozansyi Ünay, 1980; «B.» orientalis (Lungu, 1981); B. uenayae Rummel, 1998. B. menderesensis has been described by Sarica-Filoreau (2002) but is still unpublished. García-Alix et al. (2008) notice that Ruscinomys africanus Geraads, 1998 resembles rather Byzantinia. Diagnosis (emended): See Ünay, 1980. cf. Byzantinia sp. or div. sp.
9

Hír, Prieto, tiuca – Geobios Miocene rodent faunas from Com ne ti 1 and Tau
Fig. 3 1980. Hispanomys cf. lavocati (Freudenthal) - Feru et al., pp. 174, 179 and 184,185. 1980. Hispanomys cf. bijugatus (Mein and Freudenthal) - Feru et al., pp. 174, 179 and 184, 185. Origin of the studied material: Com ne ti 1. Material and measurements: Table S6. Description: M2. Juvenile, slightly worn specimen. The molar is elongated and rectangular; the anterior width (across the protocone-paracone axis) is wider than the posterior width (across the hypocone-metacone axis). The anterocone is anteriorly connected to an incipient lingual anteroloph, and posteriorly connected to the protocone via a strong anterolophule. The labial anteroloph arm is a well developed cusp. The protocone is disconnected from the endoloph. The protolophule is directed forwards, and does not connect to the entoloph; it ends free in the anterosinus. The anterosinus is labially closed. The posterior ectoloph is complete but angular. The mesoloph is not developed. M3. Juvenile, sligtly worn specimen. The anterior width is wider than the posterior width. The anterocone fused with the protocone. The lingual anteroloph arm is incipient (less developed than in M2). The labial anteroloph arm is well developed and its terminal part forms a small cusp. The anterosinus is labially open. The protolophule connects transversally to the entoloph. The posterior ectoloph is complete. The mesoloph is not developed. The metacone and hypocone are reduced. A posteroloph is not distinctly developed. m1. See description in Feru et al. (1980). m3. Two specimens: unworn and slightly worn. The outlines of the molars are rounded elliptic. The lingual anterolophulid is missing; the labial one is developed or incipient. The mesolophid is not developed. In one specimen the metaconid has a posterior spur which does not reach the base of the entoconid. Remarks: Feru et al. (1980) assigned the molars – actually the m1 – to two taxa: Hispanomys cf. lavocati and Hispanomys cf. bijugatus. In our opinion this determination needs correction, because of the following observations: • Regarding the morphologic and metric variability of the Cricetodontini m1 in general (e.g., Ünay, 1990), the presence of two species in this limited material (2 m1) is very doubtful; • Hispanomys bijugatus Mein and Freudenthal, 1971 is out of consideration because of the presence of a double metalophulid in the French m1 (Lopez-Antoñanzas and Mein, 2009); • Hispanomys lavocati (Freudenthal, 1966) is out of consideration because the m1 from Com ne ti 1 are somewhat narrower (comparison with m1 from Hostalets de Pierola in Agustí, 1980 and Nombrevilla 2 in López-Guerrero et al., 2008), and the posterior part of the teeth is not enlarged; the cuspids are more bulbous, and a small basin is present posteriorly to the posterolophid in the Spanish species. The small teeth sample from Com ne ti 1 is even inadequate for confident generic assignment. The limit between the genera Cricetodon, Hispanomys and Byzantinia is vague for the late Middle Miocene and earliest Late Miocene species. Although all teeth from Com ne ti 1 are large it is unfortunately not possible to demonstrate the taxonomic homogeneity. For instance the M2 might be difficult to assign in two co-existing equally-sized Byzantinia species (Ünay, 1980). Indeed, the simultaneous co-occurrence of two Cricetodontini species is widespread in the fossil record, as shown by Cricetodon-Deperetomys associations (e.g., Anwil; Engesser, 1972), Hispanomys-Cricetodon (e.g., La Grive L3; Mein and Ginsburg, 2002), Hispanomys-Hispanomys (e.g., La Grive L5; Mein and Ginsburg, 2002), Byzantinia-Byzantinia (e.g., Bayraktepe I; Ünay, 1980). In the following we consider the sample as mono-specific in order to simplify the analysis. The morphology of the molars from Com ne ti 1 does not fit with any Cricetodontini reported from late middle Miocene/earliest late Miocene localities of Central and Eastern Europe: • Cricetodon sp. from T ad is insufficiently documented (Hír et al., 2001), but the M2 is not elongated (“streched” sensu de Bruijn and Ünay, 1996). In the M3 the posterior ectoloph is interrupted. The labial anterolophulid is completely missing in the m3; • “Cricetodon” fandli from Gratkorn, Austria (Prieto et al., 2010a) is smaller. In the m1 the connection anteroconid-metaconid is interrupted or absent. In the m3 a short mesolophid is developed. The M2 is not “stretched”, and the lingual anteroloph is stronger; • Cricetodon klariankae from Fels tárkány-Felnémet, Hungary (Hír, 2007). In the upper molars the spur of the protocone is present, and the anterior and posterior widths are not different. In m1 the two metalophulid are equally developed. The m3 has a short mesolophid; • The newly discovered and unpublished Cricetodon sp. sample from V rciorog (Romania, Bihor District) has M2 with well developed lingual anteroloph, the spur of the protocone, and the molars are almost rectangular. The labial anterolophid of the lower molars is well developed, and closes the deep protosinusid. In the m1 the anteroconid has an independant lingual branch which connects to the metaconid; this connection is achieved close to the anterolophulid in Com ne ti. A short mesolophid is present in the m3; • Markovi (2003) listed Deperetomys hagni and Cricetodon sp. from Vra evi i (Serbia), but the material is currently under revision by the same author, and thus cannot be considered here; • Hispanomys sp. from Nebelbergweg (Kälin and Kempf, 2009: fig. 13) has m1 with completely different outline and morphology, especially the transversal mesolophid; • Cricetodon sp. from Bullendorf is insufficiently documented (Daxner-Höck, 1996).
10

Hír, Prieto, tiuca – Geobios Miocene rodent faunas from Com ne ti 1 and Tau
Some Anatolian Cricetodontini also deserve to be discussed: • B. cariensis m1 lack the anterior metalophid and the M2 is not “stretched” (Sen and Ünay, 1979; Rummel, 1998); • B. sofcaensis differs from the species from Com ne ti in having M3 with a well developed lingual anterolophule, M2 with a mesoloph, and m3 with a mesolophid (Tobien, 1978); • In B. bayraktepensis the protolophule is directed backwards, whereas this crest is transversal in Com ne ti. The Anatolian m3 has a mesolophid; • B. eskhisarensis, B. ozansoyi, B. dardanellensis and B. nikosi are clearly smaller. The molars from Com ne ti 1 share similarities with Byzantinia, but the lack of sufficient material does not allow a confident determination. Subfamily EUMYARIONINAE Ünay, 1989 Genus Eumyarion Thaler, 1966 Type species: Eumyarion medium (Lartet, 1851) (= Cricetodon helveticum Schaub, 1925). Other species included in Eumyarion: E. latior (Schaub and Zapfe, 1953); E. weinfurteri (Schaub and Zapfe, 1953); E. bifidus (Fahlbusch, 1964); E. leemanni (Hartenberger, 1965); E. valencianum Daams and Freudenthal, 1974 [nomen dubium in de Bruijn and Saraç (1991: 14) and de Bruijn (2009: 78)]; Eumyarion montanus de Bruijn and Saraç, 1991; E. intercentralis de Bruijn and Saraç, 1991; E. microps de Bruijn and Saraç, 1991; E. carbonicus de Bruijn and Saraç, 1991; E. orhani de Bruijn et al., 2006. Diagnosis (emended): See Mein and Freudenthal (1971). Differencial diagnosis: See Ünay (1989). Eumyarion sp. Fig. 4(15, 16) 1979. Eumyarion cf. bifidus (Fahlbusch) - Feru et al., p. 186, 187. Origin of the studied material: Tau . Material and measurements: Table S7. Description: M2. Rectangular outline and flat occlusal surface. The labial arm of the anteroloph is developed. A small interrupted crest runs parallel to the anterolophule. Paracone and metacone are incorporated respectively into the transversal protolophule and the metalophule. The mesoloph is broad and long, but does not reach the labial margin. The lingual sinus is directed forwards. The hypocone and the posteroloph are damaged. M3. The labial arm of the anteroloph is developed. Paracone, protocone and protolophule are fused. The posterior arm of the protocone extends to the hypocone. The metacone is incorporated into the posteroloph. The broad mesoloph connects to the labial border of the tooth. The metalophule is directed backwards. Remarks: The species of the genus Eumyarion are highly variable (e.g., de Bruijn, 2009). The taxonomic attribution of the small molar sample from Tau is thus difficult. In Romania and in Hungary only few Eumyarion finds are reported. The M2 does not have a double protolophule, and E. bifidus can be excluded. E. leemanni is characterized by the cusps which are integrated in the rigdes that join them. A crest is developed parallel to the anterolophule in some M2 from Neudorf (E. latior; Fejfar, 1974: fig. 22, molar 733920). Because of these evidences, it is not possible to assign the specimens to any species. Family SCIURIDAE Fischer von Waldheim, 1817 Subfamily PTEROMYINAE Brandt, 1855 Genus Albanensia Daxner-Höck and Mein, 1975 Type species: Albanensia albanensis (Major, 1893). Other species included in Albanensia: A. sansaniensis (Lartet, 1851), A. albanensis quiricensis (Villalta, 1950), A. grimmi (Black, 1966). Diagnosis (emended): See Daxner-Höck and Mein (1975). Albanensia sp. Fig. 1(1, 3) Origin of the studied material: Tau . Material and measurements: Table S8. Description: D4. Triangular outline, protuberant anterocone. The transversal ridges (anteroloph, protoloph, metaloph, posteroloph) are all connected to the protocone. Paracone and metacone are incorporated into the protoloph and the metaloph. The two ridges slightly converge on the protocone, and are ornamented. d3. Trapezoidal outline. The metaconid is the strongest cusp. Protoconid and metaconid are separated by a narrow trench. A mesial ridge merges from the protoconid. The posterior side of the metaconid is sculptured. The mesoconid is low and has a strong connections to the protoconid and to the hypoconid. The mesolophid is developed as a rounded cusp.
11

Hír, Prieto, tiuca – Geobios Miocene rodent faunas from Com ne ti 1 and Tau
Remarks: Mein (1970: figs. 5, 7, 10 and p. 15) figured and measured A. albanensis D4/d4. The morphology of these specimens corresponds to our teeth; the size of the molars from Tau exceeds the measurements of the D4/d4 provided by Mein. One A. grimmi d4 was described and figured from Rudabánya (Kretzoi and Fejfar, 2005: figs. 10/11) but not measured. In this specimen protoconid and metaconid are connected. In the same locality the d4 of Miopetaurista sp. might also belong to Albanensia (Kretzoi and Fejfar, 2005: 118). The measurements of the D4 from Tau are too small for Miopetaurista. Because the differences between A. albanensis and A. grimmi are based on P4-M2 (Daxner-Höck, 2004a) we cannot recognize the species without these molars. Genus Miopetaurista Kretzoi, 1962 Type species: Miopetaurista göriachensis (Hoffman, 1893) see comments in Daxner-Höck, 2004a: 392). Other species included in Miopetaurista: M. lappi (Mein, 1958); M. gaillardi (Mein, 1970); M. neogrivensis (Mein, 1970); M. crusafonti (Mein, 1970); M. thaleri (Mein, 1970); M. dehmi de Bruijn et al., 1980; M. asiatica Qiu, 2002. According to de Bruijn et al. (1980: 251), M. tobieni and M. thaleri are not different. M. diescalicus Daams, 1975 is considered as nomen dubium by de Bruijn (1999: 276). Engesser (1979) included a panel of North American species in Petauristodon, a proposal rejected by de Bruijn et al. (1980: 251), but followed by Qiu (2002: 188). Diagnosis (emended): See Daxner-Höck and Mein (1975). Miopetaurista sp. Fig. 1(4) 1979. Miopetaurista sp. - Feru et al., pp. 186, 188. Origin of the studied material: Tau . Material and measurements: Table S9. Description: Almost rectangular outline. The protocone is antero-posteriorly elongated. There are four thin transversal ridges: anteroloph, protoloph, metaloph, and posteroloph; all connect to the protocone. The paracone and the metacone are incorporated into the ridges. The three basins are labially open. A weak mesostyle is developed in the labial margin of the central basin. The lingual side of the protocone has two shallow imprints. The enamel surface is moderately granulated. Remarks: This M2 is assigned to Miopetaurista because of its size, the approximately parallel-running protoloph and metaloph, and the weak mesostyl. The species identification is not confident. This large-sized flying squirrel is a rare element of the late Middle Miocene and early Late Miocene vertebrate faunas of Eastern and Central Europe. Species determination is not given for the finds from Rudabánya (Kretzoi and Fejfar, 2005), Götzendorf and Schernham (Daxner-Höck, 2004a). Fahlbusch (1979) reports M. crusafonti in Germany (Gumpersdorf). Genus Blackia Mein, 1970 Type species: Blackia miocaenica Mein, 1970. Other species included in Blackia: The validity of the included species is still a controversial subject. De Bruijn (1999) synonymised B. parvula Baudelot, 1972 and B. ulmensis Werner, 1994 with B. miocaenica. B. woelfersheimensis Mein, 1970 (= B. polonica Black and Kowalski, 1974 in de Bruijn, 1999) is not different from B. miocaenica according to Dahlmann (2001). Diagnosis (emended): See Daxner-Höck and Mein (1975). Blackia miocaenica Mein, 1970 Fig. 1(7) 1979. Blackia sp. - Feru et al., p. 186. Type locality: La Grive L7 (France). Age: latest middle Miocene (MN7 sensu Mein and Ginsburg, 2002). Origin of the material: Tau . Material and measurements: Table S10. Diagnosis: See Mein 1970. Description: M1-2. The teeth are subrectangular. The lingual margin is rounded, and the posterior margin convex. The largest cusp is the protocone, then the paracone. The metacone is low. Three parallel ridges (anteroloph, protoloph and posteroloph) are connected to the protocone. An incipient anteroconule is incorporated into the anteroloph. The shallow basins are closed. The enamel surface is granulated. m1-2. The teeth are rhomboidal. The central basin is large and surrounded by three cusps: protoconid, metaconid, hypoconid. The entoconid is incipient and incorporated into the posterolophid. The mesoconid is not developed, but there is an enamel ridge between the base of the protoconid and the hypoconid. On the labial side a similar ridge is developed between the metaconid and the entoconid. Small groove between the protoconid and the anterolophid. The metalophid is incipient. The central basin is completely closed. The enamel surface is granulated or alveolated in the worn and senile tooth.
12

Hír, Prieto, tiuca – Geobios Miocene rodent faunas from Com ne ti 1 and Tau
Comments: The dimensional and morphological changes are minor in Blackia miocaenica from the Lower Miocene to the Pliocene (MN2-MN15). This small sized flying squirrel is a frequent element of the Mio-Pliocene vertebrate faunas of Eastern and Central Europe. Its latest occurrence is in Csarnóta 2, although it is not mentioned in the original faunal list of Kretzoi (1959, 1962). See the problem on the holotype of Pliopetes hungaricus (van de Weerd, 1979). Genus Neopetes Daxner-Höck, 2004 Type species: Neopetes hoeckarum (de Bruijn, 1998). Other species included in Neopetes: N. macedoniensis (Bouwens and de Bruijn, 1986), N. debruijni (Reumer and van den Hoek Ostende, 2003). Diagnosis: See Daxner-Höck (2004a). Neopetes hoeckarum (de Bruijn, 1998) Fig. 1(2, 8, 9) Type locality: Oberdorf (Austria). Age: Early Miocene, MN4. Origin of the studied material: Tau . Material and measurements: Table S11. Diagnosis: See de Bruijn (1998). Description: p4. Trapezoidal outline, the posterior margin being wider than the anterior one. The largest cusp is the metaconid. A small crest merges from the anterior wall of the protoconid. The mesoconid is ridge-shaped. Entoconid and mesolophid are developed. m1-2. Rectangular outline. The well developed entoconid and mesolophid are divided by a groove. The mesoconid is weak and elongated. The anterolophid and metalophid are distinct. m3. Subtriangular outline, elongated mesoconid. The development of the entoconid and the mesolophid is similar to the m1-2. Remarks: The lack of clear ornamentation, the presence of a mesostylid, and the weak mesoconid are characteristic for Neopetes (Daxner-Höck, 2004a). The identification of the finds from Tau as N. macedoniensis is not founded because of size differences. N. debruijni is out of consideration because its larger size. The stratigraphical range of Neopetes extends from the Early Miocene (Oberdorf, Austria: de Bruijn, 1998) to the Early Pleistocene (Tegelen, The Netherlands: Reumer and van den Hoek Ostende, 2003). In the Hungarian Miocene, Neopetes was reported from Fels tárkány 3/2 (Hír, 2003), Fels tárkány 3/10 (Hír and Kókay, 2010), and Rudabánya (Kretzoi and Fejfar, 2005). Subfamily SCIURINAE Fischer von Waldheim, 1817 Genus Spermophilinus de Bruijn and Mein, 1968 Type species: Spermophilinus turolensis de Bruijn and Mein, 1968. Other species included in Spermophilinus: S. bredai (Meyer von, 1848), S. besanus Cuenca Bescós, 1988 ; S. giganteus de Bruijn et al., 1970; S. minutus Zheng and Li, 1982. Diagnosis: See de Bruijn and Mein (1968). Remarks: Kretzoi and Fejfar (2005: 117) discussed intensively the synonymy Spermophilinus-Csakvaromys Kretzoi, 1951. This proposal is not followed here because the authors did not see the original material of Csakvaromys. Spermophilinus bredai (Von Meyer, 1848) Fig. 1(5, 6, 10, 11) 1979. Spermophilinus bredai (H. von Meyer) - Feru et al., p. 186. Type locality: Oehningen (Germany). Age: Middle Miocene, ?MN7 (sensu Kälin and Engesser, 2001). Origin of the material: Tau . Material and measurements: Table S12. Diagnosis: See de Bruijn and Mein (1968). Description: The species is well known: We avoid a detailed description, and refer to. e.g., Cuenca -Bescos (1988), Aldana-Carrasco (1992) and Kretzoi and Fejfar (2005). Remarks: Spermophilinus is a frequent element of the Miocene faunas from Central Europe, and especially from the Carpathian Basin. In Fig. 5 the mean size of a panel of Central European to Anatolian M1-2 are presented. The molars from Tau are intermediate in size between Fels tárkány 3/2 and Rudabánya, and close to Nebelbergweg. A size increase through time in Spermophilinus is observed by De Bruijn (1995). Because of the wide geographical
13

Hír, Prieto, tiuca – Geobios Miocene rodent faunas from Com ne ti 1 and Tau
repartition of the finds, and the limited number of localities in each region, this size increase should not be used, at present, in fine biostratigraphical context. Family EOMYIDAE Déperet and Douxami, 1902 Genus Eomyops Engesser, 1979 Type species: Eomyops catalaunicus (Hartenberger, 1966). Other species included in Eomyops: E. bodvanus (Janossy, 1972), E. oppligeri Engesser, 1990, E. hebeiseni Kälin, 1997, E. noeliae Ruiz-Sánchez, Calatayud and Freudenthal, 2009. Diagnosis: See Engesser (1979). Remarks: Engesser (1979, 1990) precisely developed the argumentation in favour of the separation of Leptodontomys and Eomyops. This proposal was, and is still not followed by a part of the scholars. For instance the Chinese material is reported to the North American genus Leptodontomys (e.g., Qiu, 1996: 183–184). It is not the purpose of this paper to deal with this synonymy, and we restrict arbitrary our species list to the “official” Eomyops species. Eomyops oppligeri Engesser, 1990 Fig. 2(1) 1979. Leptodontomys gr. catalaunicus Hartenberger - Feru et al., p. 186. Type locality: Anwil (Switzerland). Age: Middle Miocene, MN8 (sensu Kälin and Engesser, 2001). Origin of the studied material: Tau . Material and measurements: Table S13. Diagnosis: See Engesser (1990). Description: m1-2. Rectangular outline with rounded corners. Metalophid, hypolophid and anterolophids are transversal. The mesolophid is short and transversal. Remarks: There are two Eomyops species in the latest middle Miocene/early late Miocene faunas of Eastern and Central Europe (with exception of the large-sized Eomyops nov. sp. from Marktl and the unpublished, also large Eomyops nov. sp. from Be chatów A): E. catalaunicus (traditionally late Miocene) and the smaller E. oppligeri (middle Miocene-earliest late Miocene?). The size of the m1-2 from Tau clearly refers to E. oppligeri. The species is reported in Hungary and Romania in Mátrasz l s 2, Fels tárkány-Felnémet 2/3, Fels tárkány 1, Fels tárkány 3/2, Fels tárkány 3/8, and Fels tárkány 3/10.
References (in addition to the printed version)
Aguilar, J., Berggren, W., Aubry, M., Kent, D., Clauzon, G., Bennam, M., Michaux, J. 2004. Mid-Neogen Mediterranean marine – continental correlations: an alternative interpretation. Palaeogeography, Palaeoclimatology, Palaeoecology 204, 165–186.
Aguilar, J.-P., Lazzari, V., 2006. Nouvelles espèces de gliridés du gisement karstique de Blanquatère 1 (Miocène moyen, sud de la France). Geodiversitas 28, 277–295.
Agustí, J., 1980. Tendencias evolutivas de la línea Cricetodon-Ruscinomys (Rodentia, Mammalia) en la Península Ibérica. Acta Geològica Hispànica 17, 103–111.
Aldana-Carrasco, E., 1992. Los Sciurinae (Rodentia, Mammalia) del Miocene de la Cuenca del Vallés-Penedés (Cataluña, España). Treballs del Museu de Geologia de Barcelona 2, 69–97.
Bachmayer, F., Wilson, R., 1984. Die Kleinsaugerfauna von Götzendorf, Niederösterreich. Sitzungberichte der Österreichischen Akademie der Wissenschaften, matematisch-naturwissenschaftliche Klasse 193, 303–319.
Baudelot, S., 1965. Complément à l’étude de la faune de rongeurs de Sansan : les Gliridés. Bulletin de la Société géologique de France 7, 758–764.
Baudelot, S., 1972. Étude des chiroptéres insectivores et rongeurs du miocene de Sansan (Gers.). Ph.D. thesis, Université Paul Sabatier de Toulouse (unpublished).
Casanovas-Vilar, I., 2007. The rodent assemblages from the Late Aragonian and the Vallesian (Middle to Late Miocene) of the Vallès-Penedès Basin (Catalonia, Spain). Ph.D. thesis, Universitat autònoma de Barcelona (unpublished).
Casanovas-Vilar, I., Alba, D.M., Moya-Sola, S., Galindo, J., Cabrera, L., Garces, M., Fùrio, M., Robles, J.M., Köhler, M., Angelone, M., 2008. Biochronological, taphonomical and paleoenvironmental background of the fossil great ape Pierolapithecus catalaunicus (Primate hominidae). Journal of Human Evolution 55, 589–603.
Cuenca-Bescos, G., 1988. Revisión de los Sciuridae del Aragoniense y del Rambliense en la fosa de Calatayud – Montalban. Scripta Geologica 87, 2–115.
14

Hír, Prieto, tiuca – Geobios Miocene rodent faunas from Com ne ti 1 and Tau
Daams, R., 1999. Family Gliridae. In: Rössner, G., Heissig, K. (Eds), The Miocene Land Mammals of Europe. Verlag Dr. Friedrich Pfeil, München, pp. 301–318.
Daams, R., Bruijn, de H., 1995. A classification of the Gliridae (Rodentia) on the basis of dental morphology. Hystrix 6, 3–50.
Dahlmann, T., 2001. Die Kleinsäuger der unter-pliozänen Fundstelle Wölfersheim in der Wetterau (Mammalia: Lipotyphla, Chiroptera, Rodentia). Courier Forschungsinstitut Senckenberg 227, 1–129.
Daxner-Höck, G., 1967. Ein neuer Cricetodontide (Rodentia, Mammalia) aus dem Pannon des Wiener Beckens. Annalen des Naturhistorischen Museums in Wien 77, 27–36.
Daxner-Höck, G., 1972. Cricetinae aus dem alt-Pilozän vom Eichkogel bei Mödling (Niederösterreich) und von Vösendorf bei Wien. Paläontologische Zeitschrift 46, 133–150.
Daxner-Höck, G., 1996. Faunenwandel im Obermiozän und Korrelation der MN-„Zonen” mit den Biozonen des Pannons der Zentralen Paratethys. Beiträge zur Paläontologie 21, 1–9.
Daxner-Höck, G., 1998. Säugetiere (mammalia) aus dem Karpat des Korneuburger Beckens 3. Rodentia und Carnivora. Beiträge zur Paläontologie 23, 367–407.
Daxner-Höck, G., 2004a. Flying Squirrels (Pteromyinae, Mammalia) from the Upper Miocene of Austria. Annalen Naturhistorisches Museum Wien 106A, 387–423.
Daxner-Höck, G., 2004b. Biber und ein Zwerghamster aus Mataschen (Unter-Pannonium, Steirisches Becken). Jonnea-Geologie und Paläontologie 5, 19–33.
Daxner-Höck, G., 2005. Eomyidae and Gliridae from Rudabánya. Palaeontographia Italica 90, 149–161. Daxner-Höck, G., 2010. Sciuridae, Gliridae and Eomyidae (Rodentia, Mammalia) from the Middle Miocene of St.
Stefan in the Gratkorn Basin (Styria, Austria). Annalen des naturhistorischen Museums in Wien Serie A 112, 507–536.
Daxner-Höck, G., Bruijn, H. de, 1981. Gliridae (Rodentia, Mammalia) des Eichkogels bei Mödling (Niederösterreich). Paläontologische Zeitschrift 55, 157–172.
Daxner-Höck, G., Höck, E., 2009. New data on Eomyidae and Gliridae (Rodentia, Mammalia) from the Late Miocene of Austria. Annalen des Naturhistorisches Museums Wien 111A, 375–444.
Daxner-Höck, G., Mein, P., 1975. Taxonomische Probleme um das Genus Miopetaurista Kretzoi, 1962 (Fam. Sciuridae). Paläontologische Zeitschrift 49, 75–77.
de Bruijn, H., 1966. Some new Gliridae from the Calatayud area (Prov. Zaragoza, Spain). I, II. Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen, Ser. B 69 (2), 58–78.
de Bruijn, H., 1995. The Vertebrate locality Maramena (Macedonia, Greece) at the Turolian-Ruscinian Boundary (Neogene). 8. Sciuridae, Petauristidae and Eomyidae (Rodentia, Mammalia). Münchner Geowissenschaftliche Abhandlungen 28, 87–102.
de Bruijn, H., 1998. Vertebrates from the Early Miocene lignite deposits of the opencast mine Oberdorf (wester Styrian Basin, Austria). 6. Rodentia 1 (mammalia). Annalen Naturhistorisches Museum Wien 99A, 99–137.
de Bruijn, H., 1999. Superfamiliy Sciuroidea. In: Rössner, G., Heissig, K. (Eds.), The miocene land mammals of Europe. Pfeil, München, pp. 271–280.
de Bruijn, H., 2009. The Eumyarion (Mammalia, Rodentia, Muridae) assemblage from Sandelzhausen (Miocene, Southern Germany): a test on homogeneity. Paläontologische Zeitschrift 83, 77–83.
de Bruijn, H., Hoek Ostende, L. van den., Kristkoiz-Boon, E., Rummel, M., Theocharopoulos, C., Ünay, E., 2003. Rodent, lagomorphs and insectivores, from the middle Miocene hominoid locality of Çandir (Turkey). Courier Forschungsinstitut Senckenberg 240, 51–87.
de Bruijn, H., Mayda, S., Hoek Ostende, L.W. van den, Kaya, T., Saraç, G., 2006. Small mammals from the Early Miocene of Sabuncubeli (Manisa, S.W. Anatolia, Turkey). Beiträge zur Paläontologie 30, 57–87.
de Bruijn, H., Mein, P., 1968. On the mammalian fauna of the Hipparion-Beds in the Calatayud-Teruel Basin (Prov. Zaragoza, Spain). Part V. The Sciurinae. Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen B 71, 73–90.
de Bruijn, H., Meulekamp, J.E., 1972. Late Miocene rodents from the Pandanassa formation (Prov. Rethymnon), Crete, Greece. Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen B 75, 54–60.
de Bruijn, H., Meulen, A.J. van der, Katsikatsos, G., 1980. The Mammals from the Lower Miocene of Aliveri (Island of Evia, Greece). Part 1. The Sciuridae. Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen B 83, 241–261.
de Bruijn, H., Saraç, G., 1991. Early Miocene Rodent Faunas from the Eastern Mediterranean Area. Part 1. the genus Eumyarion. Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen B 94, 1–36.
15

Hír, Prieto, tiuca – Geobios Miocene rodent faunas from Com ne ti 1 and Tau
de Bruijn, H., Ünay, E., 1996. On the Evolutionary History of the Cricetodontini from Europe and Asia Minor and its Bearing on the reconstruction of Migrations and the Continental Biotope During the Neogene. In: Bernor, R., Fahlbusch, V., Mittmannn, H. (Eds.), The Evolution of Eastern Eurasian Neogene Mammal Faunas. Columbia University, pp. 227–234.
Ellermann, J.R., 1941. The families and genera of living rodents. British Museum 1, London. Engesser, B., 1972. Die obermiozäne Säugertierefauna von Anwil (Baselland). Tätigkeitsberichte der
Naturforschenden Gesellschaft Baselland 28, 37–363. Engesser, B., 1979. Relationships of some insectivores and rodents from the Miocene of North America and Europe.
Bulletin Carnegie Museum Natural History 14, 1–68. Engesser, B., 1990. Die Eomyidae (Rodentia, Mammalia) der Molasse der Schweiz und Savoyens. Shweizerische
Paläontologische Abhandlungen 112, 1–144. Fahlbusch, V., 1964. Die Cricetiden der Oberen Süßwassermolasse Bayerns. Bayerische Akademie der
Wissenschaften; mathematisch-naturwissenschaftliche Klasse, Abhandlungen, neue Folge 118, 1–136. Fahlbusch, V., 1979. Flughörnchen-Schädel. Jahresbericht 1978 und Mitteilungen der Freunde der Bayerischen
Staatssammlung für Paläontologie und Geologie München 7, 18–19. Fejfar, O., 1974. Die Eomyiden und Cricetiden (Rodentia, Mammalia) des Miozäns der Tschechoslowakei.
Paleontographica (A) 146, 99–180. Fejfar, O., 1990. The Neogene VP Sites of Czechoslovakia: A Contribution to the Neogene Terrestric Biostratigraphy
of Europe Based on Rodents. In: Lindsay, E.H., Falhbusch, V., Mein, P. (Eds.), European Neogene Mammal Chronology. NATO ASI-Series (A) 180, pp. 211–236.
Fejfar, O., Storch, G., 1990. Eine pliozäne (ober-ruscinishe) Kleinsäugerfauna aus Gundersheim, Rheinhesse. 1. Nagetiere: Mammalia, Rodentia. Senckenbergiana lethaea 71, 139–184.
Feru, M., R dulescu, C., Samson, P., 1979. La faune de Micromammifères du Miocène de Tau (département d’Arad). Travaux de l’Institut de Spéologie « Emil Racovitz » 18, 185–190.
Feru, M., R dulescu, C., Samson, P., 1980. La faune de Micromammiféres du Miocéne de Com nesti (département d’Arad). Travaux de l’Institut de Spéologie « Emil Racovitz » 19, 171–190.
Freudenthal, M., 1966. On the Mammalian Fauna of the Hipparion-Beds in the Calatayud-Teruel Basin. Part 1. The genera Cricetodon and Rotundomys (Rodentia). Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen B 69, 269–317.
Freudenthal, M., 1967. On the mammalian fauna of the Hipparion-beds in the Calatayud-Teruel Basin. Part II3. Democricetodon and Rotundomys (Rodentia). Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen B 70, 298–315.
Freudenthal, M., 2006. The status of Democricetodon, Fahlbuschia, Pseudofahlbuschia and Renzimys (Cricetidae, Mammalia). A reply to Van der Meulen et al. (2004). Geobios 39, 43–55.
Freudenthal, M., Martín-Suárez, E., 2006. Gliridae (Rodentia, Mammalia) from the Late Miocene fissure filling Biancone 1 (Gargano, Province of Foggia, Italy). Palaeontologia electronica 9, 6A, 1–23.
Gál, E., Hír, J., Kessler, E., Kókay, J., Venczel, M., 2000. Middle Miocene fossils from the road cut, Rákóczi-kápolna. II. The Mátrasz l s 2nd locality. Folia Historico-naturalia Musei Matraensis 24, 39–75 (in Hungarian with English abstract).
Garapich, A., 2002. An overview of Miocene rodents from Be chatów (Poland). Folia Zoologica 51 (supp. 1), 59–66. García-Alix, A., Minwer-Barakat, R., Martín-Suárez, E., Freudenthal, M., 2008. Cricetidae and Gliridae (Rodentia,
Mammalia) from the Miocene and Pliocene of southern Spain. Scripta Geologica 136, 1–37. Gross, M., Böhme, M., Prieto, J., in press. Gratkorn – a benchmark locality for the continental Sarmatian s.str. of the
Central Paratethys. International Journal of Earth Sciences (doi: 10.1007/s00531-010-0615-1). Gross, M., Fritz, I., Piller, W.E., Soliman, A., Harzhauser, M., Hubmann, B., Moser, B., Schloger, R., Suttner, T.J.,
2007a. The Neogene of the Styrian Basin. Guide to Excursion. Jonnea-Geologie und Paläontologie 9, 117–193. Gross, M., Harzhauser, M., Mandic, O., Piller, W.E., Rögl, F., 2007b. A Stratigraphic Enigma: the age of the Neogene
Deposits of Graz (Styrian Basin; Austria). Guide to Excursion. Jonnea-Geologie und Paläontologie 9, 195–120. Harzhauser, M., Gross, M., Binder, H., 2008. Biostratigraphy of Middle Miocene (Samartian) wetland systems in an
Eastern Alpine intramontane basin (Gratkorn Basin, Austria): the terrestrial gatropod approach. Geologica Carpathica 59, 45–58.
Heissig, K., 1995. Die Entwicklung der großen Democricetodon-Arten und die Gattung Collimys (Cricetidae, Mamm.) im späten Mittelmiozän. Mitteilungen der Bayerischen Staatssammlung für Paläontologie und historische Geologie 35, 87–108.
16

Hír, Prieto, tiuca – Geobios Miocene rodent faunas from Com ne ti 1 and Tau
Heissig, K., 2006. Die Gattung Miodyromys (Gliridae, Mammalia) im tiefen Mittelmiozän der Oberen Süßwassermolasse Bayerns. Beiträge zur Paläontologie 30, 143–153.
Hír, J., 2003. The Middle Miocene (Late Astaracian, MN7-8) rodent fauna of Fels tárkány 3/2 (Hungary). Acta Palaeontologica Romaniae 5, 125–136.
Hír, J., 2006. Late Astaracian (Late Sarmatian) Lagomorphs and Rodents from Fels tárkány – Felnémet (Northern Hungary). Beiträge zur Paläontologie 30, 155–173.
Hír, J., 2007. Cricetodon klariankae n. sp. (Cricetodontini, Rodentia) from Fels tárkány-Felnémet (Northern Hungary). Fragmenta Palaeontologica Hungarica 24/25, 15–24.
Hír, J., Kókay, J., 2004. Middle Miocene molluscs and rodents from Mátrasz l s (Mátra Mountains, Hungary). Fragmenta Palaeontologica Hungarica 22, 83–97.
Hír, J., Kókay, J., 2009. Middle Miocene Molluscs, Lagomorphs and Rodents of Fels tárkány 1 and Fels tárkány 2. Fragmenta Palaeontologica Hungarica 27, 81–89.
Hír, J., Kókay, J., 2010. A systematic study of the middle-late Miocene rodents and lagomorphs (Mammalia) of Fels tárkány 3/8 and 3/10 (Northern Hungary). Geodiversitas 32, 307–329.
Hír, J., Kókay, J., Venczel, M., 2001. Middle Miocene molluscs and microvertebrata from T ad (Bihor County, Romania). Acta Paleontologica Romaniae 3, 161–172.
Hír, J., Mészáros, L., 2002. Middle Miocene insectivores and rodents (Mammalia) from Sámsonháza (Northern Hungary). Fragmenta Palaeontologica Hungarica 20, 9–23.
Hír, J., Venczel, M., 2005. New Middle Miocene vertebrate localities from Subpiatra (Bihor County, Romania). Acta Palaeontologica Romaniae 5, 211–221.
Hugueney, M., Mein, P., 1965. Lagomorphes et Rongeurs du Néogène de Lissieu (Rhòne). Travaux du Laboratoire Géolologique de la Faculté des Sciences de Lyon 12, 109–123.
Joniak, P., 2005. New Rodent assemblages from the Upper Miocene deposits of the Vienna Basin and Danube Basin. Ph.D. Thesis, Comenius University, Faculty of Natural Sciences, Department of Geology and Paleontology (unpublished).
Jung, W., Mayr, H., 1980. Neuere Befunde zur Biostratigraphie der Oberen Süßwassermolasse Süddeutschlands und ihre palökologische Deutung. Mitteilungen der Bayerischen Staatssammlung für Paläontologie und historische Geologie 20, 159–173.
Kälin, D., Engesser, B., 2001. Die jungmiozäne Säugetierfauna vom Nebelbergweg bei Nunningen (Kanton Solothurn, Schweiz). Schweizerische Paläontologische Abhandlungen 121, 1–61.
Kälin, D., Kempf, O., 2009. High-resolution stratigraphy from the continental record oft he Middle Miocene Northern Alpine Foreland Basin of Switzerland. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 254, 177–235.
Kälin, D., Weidmann, M., Engesser, B., Berger, J.-P., 2001. Paléontologie et âge de la Molasse d’eau douce supérieure (OSM) du Jura Neuchâtelois. Schweizerische Paläontologische Abhandlungen 121, 63–101.
Kordos, L., 1986. Upper Miocene Hamsters (Cricetidae, Mammalia) of Hasznos and Szentendre: a taxonomic and stratigraphic study. Annual Report of the Hungarian Geological Institute on the year 1984, 523–553.
Kowalski, K., 1967. Rodents from the Miocene of Opole. Acta zoological Cracoviensia 12, 1–18. Kowalski, K., 1997. Gliridae (Mammalia: Rodentia) from the Miocene of Be chatów in Poland. Acta zoologica
Cracoviensia 40, 173–198. Kretzoi, M., 1959. Insectivoren, Nagetiere und Lagomorphen der jüngpliozänen Fauna von Csarnóta im Villányer
Gebirge (Südungarn). Vertebrata Hungarica 1, 237–246. Kretzoi, M., 1962. Fauna und Faunenhorizont von Csarnóta. Annual Report of the Hungarian Geological Institute on
the year 1959, 297–395. Kretzoi, M., Fejfar, O., 2005. Sciurids and Cricetids (Mammalia, Rodentia) from Rudabánya. Palaeontographia Italica
90, 113–148. Lopez-Antoñanzas, R., Mein, P., 2009. First detailed description of Hispanomys bijugatus Mein and Freudenthal,
1971 (Rodentia, Cricetodontinae) from the Upper Aragonian of La Grive-Saint Alban (France): Biostratigraphical implications. Geobios 42, 783–796.
López-Guerrero, P., Álvarez-Sierra, M.A., López-Antoñanzas, R., Oliver-Pérez, A., Peláez-Campomanes, P., 2008. Cricetodontini (Cricetidae, Rodentia) del Aragoniense superior (Mioceno medio) de Nombrevilla 2 (Zaragoza, España). Paleontologica Nova 8, 259–271.
Lungu, A., 1981. [The Hipparion fauna of the Middle Sarmatian from Moldavia (Insectivora, Lagomorpha, Rodentia)]. Izdatelstvo Štiinca, 1–118.
17

Hír, Prieto, tiuca – Geobios Miocene rodent faunas from Com ne ti 1 and Tau
Maridet, O., 2003. Révision du genre Democricetodon (Mammalia, Rodentia, Cricetinae) et dynamique des faunes de rongeurs du Néogène d’Europe occidentale: évolution, paléodiversité et paléogéographie. Ph.D. thesis, Université Claude-Bernard Lyon 1 (unpublished).
Maridet, O, Berthet, D., Mein, P., 2000. Un nouveau gisement karstique polyphasé miocène moyen de Four (Isère) : étude des Cricetidae (Mammalia, Rodentia) et description de Democricetodon fourensis nov. sp.. Géologie de la France 2, 71–79.
Markovi , Z., 2003. The Miocene small mammals of Serbia, a review. Deinsea 10, 393–398. Mayr, H., 1979. Gebissmorphologische Untersuchungen an miozänen Gliriden (Mammalia, Rodentia)
Süddeutschlands. Ph.D. thesis, Universität München (unpublished). Mayr, H., Fahlbusch, V., 1975. Eine unterpliozäne Kleinsäugerfauna aus der Oberen Süßwasser-Molasse Bayerns.
Mitteilungen der Bayerischen Staatssammlung für Paläontologie und historische Geologie 15, 91–111. McGuire, J.L., 2010. Geometric morphometrics of vole (Microtus californicus) dentition as a new paleoclimate proxy:
Shape changes along geographic and climatic clines. Quaternary International 212, 198–205. Mein, P., 1970. Les Sciuroptères (Mammalia, Rodentia) néogènes d’Europe Occidentale. Geobios 3, 1–56. Mein, P., Freudenthal, M., 1971. Une nouvelle classification des Cricetidae (Mammalia, Rodentia) du Tertiaire de
l’Europe. Scripta Geologica 2, 1–37. Mein, P., Ginsburg, L., 2002. Sur l’âge relatif des différents dépôts karstiques miocènes de la Grive-Saint-Alban
(Isère). Cahiers scientifiques du Muséum d’histoire naturelle de Lyon 2, 7–47. Meulen, A.J. van der, Bruijn, H. de, 1982. The mammals from the Lower Miocene of Aliveri (Island of Evia, Greece).
Part 2. The Gliridae. Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen B 85, 485–524. Nesin, V., Kowalski, K., 1997. Miocene Gliridae (Mammalia:Rodentia) from Grytsiv (Ukraine). Acta Zoologica
Cracoviensia 40, 209–222. Peláez-Campomanes, P., Daams, R., 2002. Middle Miocene rodents from Pa alar, Anatolia, Turquey. Acta
Palaeontologica Polonica 47, 125–132. Prieto, J., 2007. Kleinsäuger-Biostratigraphie und Paläoökologie des höheren Mittelmiozäns (MN8) Bayerns;
Spaltenfüllungen der Fränkischen Alb und Lokalitäten der Oberen Sü wassermolasse im Vergleich. Ph.D. thesis, Ludwig-Maximilians-Universität München (unpublished).
Prieto, J., Böhme, M., Gross, M., 2010a. The cricetid rodent fauna from Gratkorn (Austria, Styria): A benchmark locality for the continental Sarmatian s. str. (late Middle Miocene) in the central Paratehys. Geologica Carpathica 61, 419–436.
Prieto, J., Gross, M., Böhmer, C, Böhme, M., 2010b. Insectivores and bat (Mammalia) from the late Middle Miocene of Gratkorn (Austria): biostratigraphic and ecologic implications. Neues Jahrbuch für Geologie und Paläontologie 258, 107–119.
Prieto, J., Rummel, M., 2009a. The genus Collimys Daxner-Höck, 1972 (Rodentia, Cricetidae) in the Middle Miocene fissure fillings of the Frankian Alb (Germany). Zitteliana 48/49, 75–88.
Prieto, J., Rummel, M., 2009b. Small and medium sized Cricetidae (Mammalia, Rodentia) from the Middle Miocene fissure filling Petersbuch 68 (South Germany). Zitteliana 48/49, 89–102.
Qiu, Z.D., 1996. Middle Miocene micromammalian fauna from Tunggur, Nei Mongol. Beijing Science Press, Beijing. Qiu, Z.D., 2002. Sciurids from the Late Miocene Lufeng hominioid locality, Yunnan. Vertebrata PalAsiatica 40, 177–
193. R dulescu, C., Samson, P., 1988. Les Cricétides (Rodentia, Mammalia) du Miocène (Astarcien Supérieur) du
Roumanie. Travaux de l’Institut de Spéologie « Emil R covi z » 37, 67–78. Reumer, J., van den Hoek Ostende, L.W., 2003. Petauristidae and Sciuridae (Mammalia, Rodentia) from tegelen,
Zuurland, and the Maasvlakte (the Netherlands). In: Reumer, J., Wessels, W. (Eds.), Distribution and Migration of Tertiary Mammals in Europe. Deinsea 10, pp. 455–467.
Rummel, M., 1998. Die Cricetiden aus dem Mittel- und Obermiozän der Türkei unter besonderer Berückschtigung der Sickenberg/Tobien’schen Aufsammlung (1968-1970). Documenta naturae 123, 1–300.
Sabol, M., Joniak, P., Holec, P., 2004. Succession(-s) of mammalian assemblages during the Neogene, a case study from the Slovak part of the Western Carpathians. Scripta Facultatis Scientarium Naturalis Universitatis Masaryk Brunensis 31/32, 65–84.
Sarica-Filoreau, N., 2002. Faunes de Rongeurs Néogènes et Quaternaires des Grabens d’Anatolie occidentale. Systématique, biochronologie et implications tectoniques. Ph.D. thesis, Muséum national d’histoire naturelle de Paris (unpublished).
18

Hír, Prieto, tiuca – Geobios Miocene rodent faunas from Com ne ti 1 and Tau
Seehuber, U., 2009. Litho- und biostratigraphische Untersuchungen in der Oberen Süßwassermolasse in der Umgebung von Kirschheim in Schwaben. Documenta naturae 175, 1–355.
Sen, S., Ünay, E., 1979. Sur quelques Cricetodontini (Rodentia) du Miocène moyen d’Anatolie. Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen B 82, 293–301.
Tobien, H., 1978. New Species of Cricetodontini (Rodentia, Mammalia) from the Miocene of Turkey (Känozoikum un Braunkohlen der Turkei. 21). Mainzer geowissenschaftliche Mitteilungen 6, 209–219.
Tong, H., Jaeger, J.-J., 1993. Muroid rodents from the Middle Miocene Fort Tenan locality (Kenya) and their contribution to the phylogeny of Muroids. Palaeontographica (A) 229, 51–73.
Ünay, E., 1980. The Cricetodontini (Rodentia) from the Bayraktepe section (Çanakkale, Turkey). Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen B 83, 399–418.
Ünay, E., 1989. Rodents from the Middle Oligocene of Turkish Thrace. Utrecht Micropaleontological Bulletins, Special Publication 5, 1–119.
Ünay, E., 1990. Turkomys pasalarensis Tobien, its range of variation in the type locality at Pa alar, Turkey. Journal of Human Evolution 19, 437–443.
Ünay, E., Bruijn, H. de, Saraç, G., 2003. A preliminary zonation of the continental Neogene of Anatolia based on rodents. In: Reumer, J.W.F., Wessels, W. (Eds.), Distribution and migration of Tertiary mammals in Eurasia. A volume in honour of Hans de Bruijn, DEINSEA 10, pp. 539–547.
Venczel, M., Hír, J., Huza, R., Popa, E., Golban, D., 2005. A new Middle Miocene vertebrate fauna from Subpiatr (Bihor County, Romania). Nymphaea, Folia Naturae Bihariae 32, 23–38.
Vianey-Liaud, M., 2003. Gliridae (Mammalia, Rodentia) de l’Oligocène européen: origine des trois genres miocènes. Coloquios de Paleontologia, Volumen Extraordinario 1, 669–698.
Wessels, W., 1996. Myocricetodontinae from the miocene of Pakistan. Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen B 99, 253–312.
Weerd van de, A., 1979. Early Ruscinian rodents and lagomorphs (Mammalia) from the lignites near Prolemais (Macedonia, Greece). I. proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen B 82, 127–170.
Wu, W., 1990. Die Gliriden (Mammalia, Rodentia) aus der Oberen Süßwasser-Molasse von Puttenhausen (Niederbayern). Mitteilungen der Bayerischen Staatssammlung für Paläontologie und historische Geologie 30, 65–105.
Wu, W., 1993. Neue Gliridae (Rodentia, Mammalia) aus untermozänen (orleanischen) Spaltenfüllungen Süddeutschlands. Documenta Naturae 81, 1–149.
19
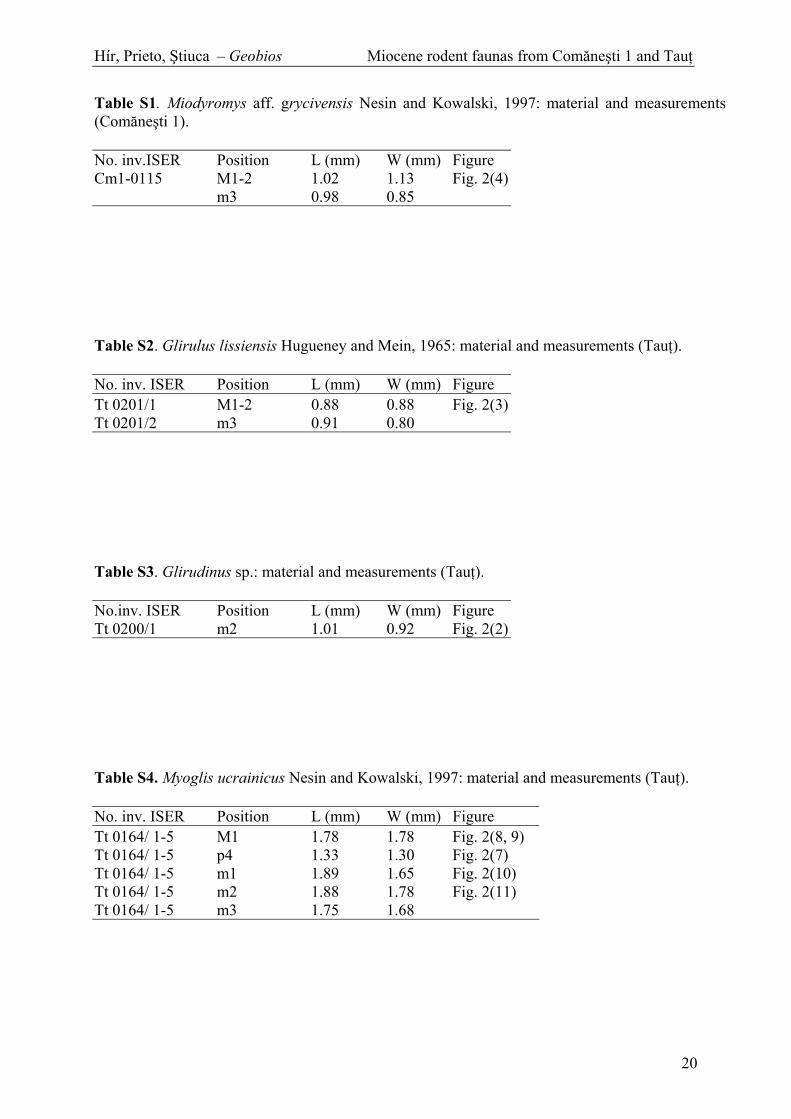
Hír, Prieto, tiuca – Geobios Miocene rodent faunas from Com ne ti 1 and Tau
Table S1. Miodyromys aff. grycivensis Nesin and Kowalski, 1997: material and measurements (Com ne ti 1). No. inv.ISER Position L (mm) W (mm) Figure Cm1-0115 M1-2 1.02 1.13 Fig. 2(4) m3 0.98 0.85 Table S2. Glirulus lissiensis Hugueney and Mein, 1965: material and measurements (Tau ). No. inv. ISER Position L (mm) W (mm) Figure Tt 0201/1 M1-2 0.88 0.88 Fig. 2(3) Tt 0201/2 m3 0.91 0.80 Table S3. Glirudinus sp.: material and measurements (Tau ). No.inv. ISER Position L (mm) W (mm) Figure Tt 0200/1 m2 1.01 0.92 Fig. 2(2) Table S4. Myoglis ucrainicus Nesin and Kowalski, 1997: material and measurements (Tau ). No. inv. ISER Position L (mm) W (mm) Figure Tt 0164/ 1-5 M1 1.78 1.78 Fig. 2(8, 9) Tt 0164/ 1-5 p4 1.33 1.30 Fig. 2(7) Tt 0164/ 1-5 m1 1.89 1.65 Fig. 2(10) Tt 0164/ 1-5 m2 1.88 1.78 Fig. 2(11) Tt 0164/ 1-5 m3 1.75 1.68
20
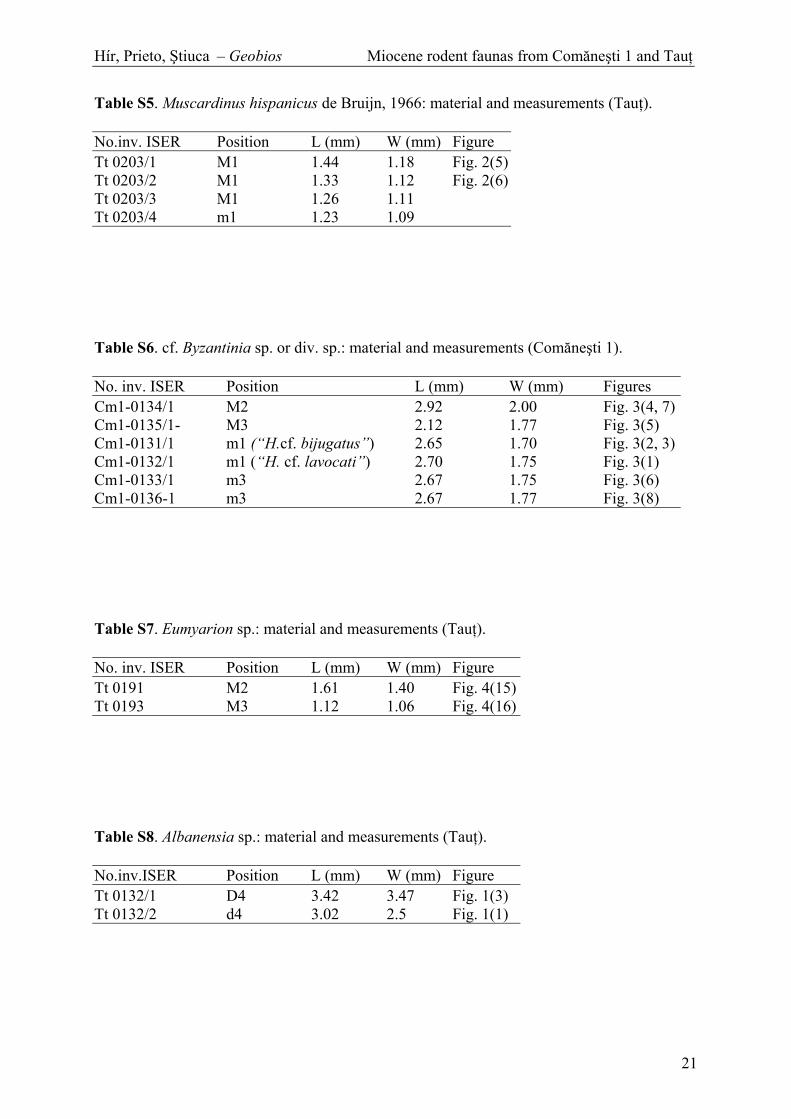
Hír, Prieto, tiuca – Geobios Miocene rodent faunas from Com ne ti 1 and Tau
Table S5. Muscardinus hispanicus de Bruijn, 1966: material and measurements (Tau ). No.inv. ISER Position L (mm) W (mm) Figure Tt 0203/1 M1 1.44 1.18 Fig. 2(5) Tt 0203/2 M1 1.33 1.12 Fig. 2(6) Tt 0203/3 M1 1.26 1.11 Tt 0203/4 m1 1.23 1.09 Table S6. cf. Byzantinia sp. or div. sp.: material and measurements (Com ne ti 1). No. inv. ISER Position L (mm) W (mm) Figures Cm1-0134/1 M2 2.92 2.00 Fig. 3(4, 7) Cm1-0135/1- M3 2.12 1.77 Fig. 3(5) Cm1-0131/1 m1 (“H.cf. bijugatus”) 2.65 1.70 Fig. 3(2, 3) Cm1-0132/1 m1 (“H. cf. lavocati”) 2.70 1.75 Fig. 3(1) Cm1-0133/1 m3 2.67 1.75 Fig. 3(6) Cm1-0136-1 m3 2.67 1.77 Fig. 3(8) Table S7. Eumyarion sp.: material and measurements (Tau ). No. inv. ISER Position L (mm) W (mm) Figure Tt 0191 M2 1.61 1.40 Fig. 4(15) Tt 0193 M3 1.12 1.06 Fig. 4(16) Table S8. Albanensia sp.: material and measurements (Tau ). No.inv.ISER Position L (mm) W (mm) Figure Tt 0132/1 D4 3.42 3.47 Fig. 1(3) Tt 0132/2 d4 3.02 2.5 Fig. 1(1)
21
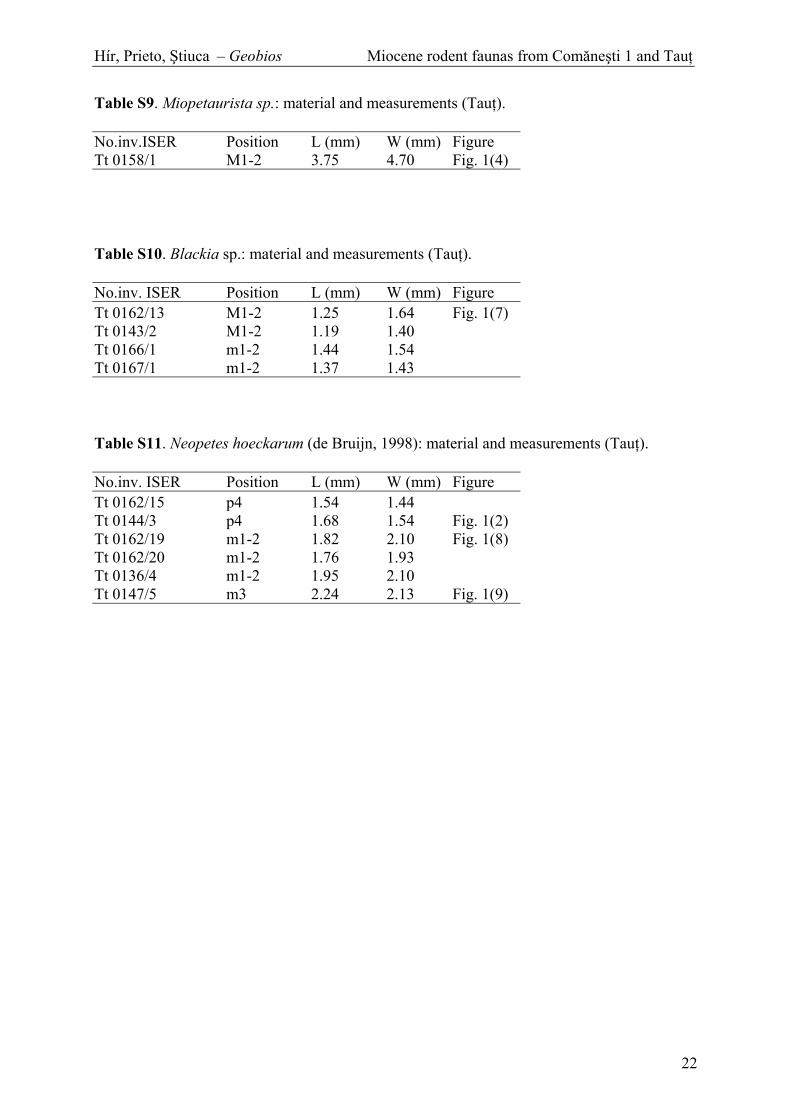
Hír, Prieto, tiuca – Geobios Miocene rodent faunas from Com ne ti 1 and Tau
Table S9. Miopetaurista sp.: material and measurements (Tau ). No.inv.ISER Position L (mm) W (mm) Figure Tt 0158/1 M1-2 3.75 4.70 Fig. 1(4) Table S10. Blackia sp.: material and measurements (Tau ). No.inv. ISER Position L (mm) W (mm) Figure Tt 0162/13 M1-2 1.25 1.64 Fig. 1(7) Tt 0143/2 M1-2 1.19 1.40 Tt 0166/1 m1-2 1.44 1.54 Tt 0167/1 m1-2 1.37 1.43 Table S11. Neopetes hoeckarum (de Bruijn, 1998): material and measurements (Tau ). No.inv. ISER Position L (mm) W (mm) Figure Tt 0162/15 p4 1.54 1.44 Tt 0144/3 p4 1.68 1.54 Fig. 1(2) Tt 0162/19 m1-2 1.82 2.10 Fig. 1(8) Tt 0162/20 m1-2 1.76 1.93 Tt 0136/4 m1-2 1.95 2.10 Tt 0147/5 m3 2.24 2.13 Fig. 1(9)
22
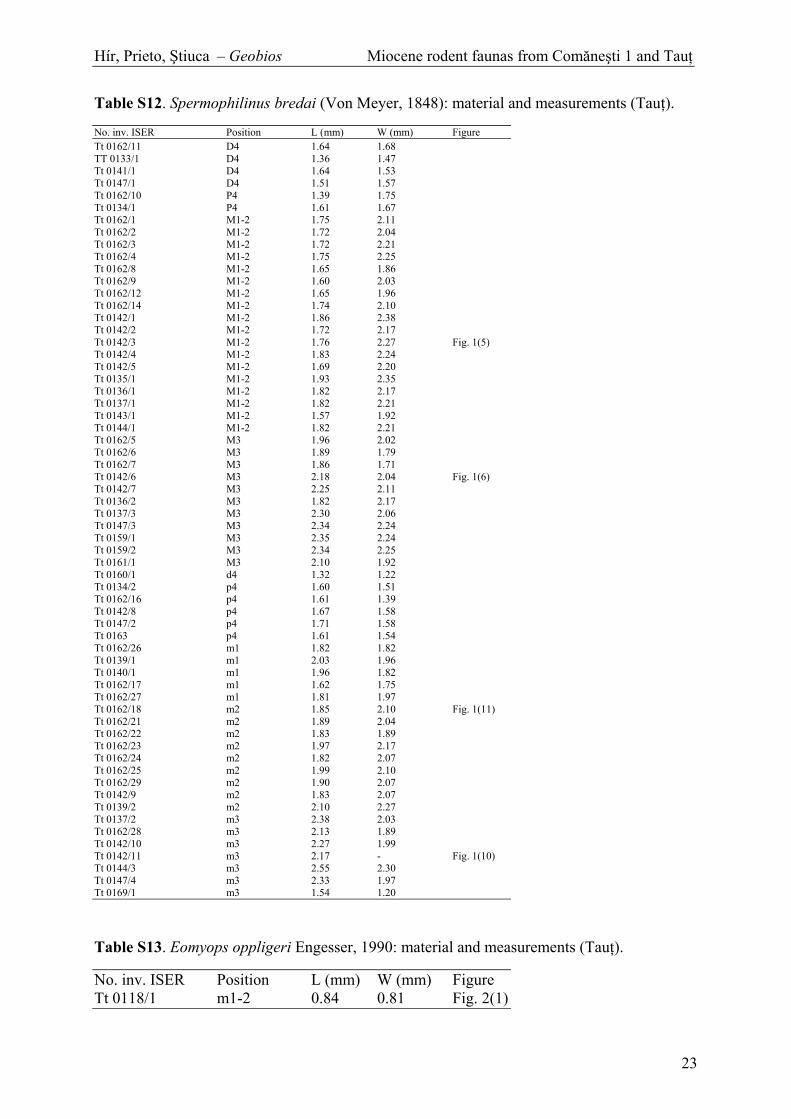
Hír, Prieto, tiuca – Geobios Miocene rodent faunas from Com ne ti 1 and Tau
Table S12. Spermophilinus bredai (Von Meyer, 1848): material and measurements (Tau ). No. inv. ISER Position L (mm) W (mm) Figure Tt 0162/11 D4 1.64 1.68 TT 0133/1 D4 1.36 1.47 Tt 0141/1 D4 1.64 1.53 Tt 0147/1 D4 1.51 1.57 Tt 0162/10 P4 1.39 1.75 Tt 0134/1 P4 1.61 1.67 Tt 0162/1 M1-2 1.75 2.11 Tt 0162/2 M1-2 1.72 2.04 Tt 0162/3 M1-2 1.72 2.21 Tt 0162/4 M1-2 1.75 2.25 Tt 0162/8 M1-2 1.65 1.86 Tt 0162/9 M1-2 1.60 2.03 Tt 0162/12 M1-2 1.65 1.96 Tt 0162/14 M1-2 1.74 2.10 Tt 0142/1 M1-2 1.86 2.38 Tt 0142/2 M1-2 1.72 2.17 Tt 0142/3 M1-2 1.76 2.27 Fig. 1(5) Tt 0142/4 M1-2 1.83 2.24 Tt 0142/5 M1-2 1.69 2.20 Tt 0135/1 M1-2 1.93 2.35 Tt 0136/1 M1-2 1.82 2.17 Tt 0137/1 M1-2 1.82 2.21 Tt 0143/1 M1-2 1.57 1.92 Tt 0144/1 M1-2 1.82 2.21 Tt 0162/5 M3 1.96 2.02 Tt 0162/6 M3 1.89 1.79 Tt 0162/7 M3 1.86 1.71 Tt 0142/6 M3 2.18 2.04 Fig. 1(6) Tt 0142/7 M3 2.25 2.11 Tt 0136/2 M3 1.82 2.17 Tt 0137/3 M3 2.30 2.06 Tt 0147/3 M3 2.34 2.24 Tt 0159/1 M3 2.35 2.24 Tt 0159/2 M3 2.34 2.25 Tt 0161/1 M3 2.10 1.92 Tt 0160/1 d4 1.32 1.22 Tt 0134/2 p4 1.60 1.51 Tt 0162/16 p4 1.61 1.39 Tt 0142/8 p4 1.67 1.58 Tt 0147/2 p4 1.71 1.58 Tt 0163 p4 1.61 1.54 Tt 0162/26 m1 1.82 1.82 Tt 0139/1 m1 2.03 1.96 Tt 0140/1 m1 1.96 1.82 Tt 0162/17 m1 1.62 1.75 Tt 0162/27 m1 1.81 1.97 Tt 0162/18 m2 1.85 2.10 Fig. 1(11) Tt 0162/21 m2 1.89 2.04 Tt 0162/22 m2 1.83 1.89 Tt 0162/23 m2 1.97 2.17 Tt 0162/24 m2 1.82 2.07 Tt 0162/25 m2 1.99 2.10 Tt 0162/29 m2 1.90 2.07 Tt 0142/9 m2 1.83 2.07 Tt 0139/2 m2 2.10 2.27 Tt 0137/2 m3 2.38 2.03 Tt 0162/28 m3 2.13 1.89 Tt 0142/10 m3 2.27 1.99 Tt 0142/11 m3 2.17 - Fig. 1(10) Tt 0144/3 m3 2.55 2.30 Tt 0147/4 m3 2.33 1.97 Tt 0169/1 m3 1.54 1.20
Table S13. Eomyops oppligeri Engesser, 1990: material and measurements (Tau ).
No. inv. ISER Position L (mm) W (mm) Figure Tt 0118/1 m1-2 0.84 0.81 Fig. 2(1)
23
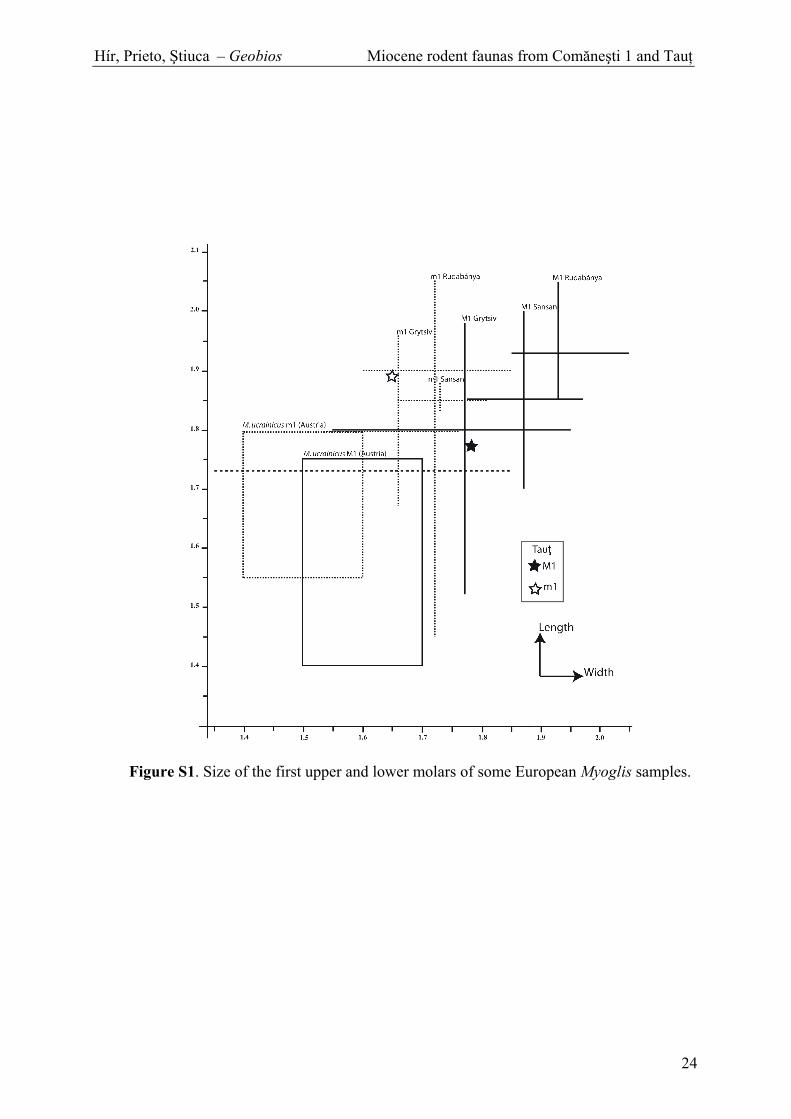
Hír, Prieto, tiuca – Geobios Miocene rodent faunas from Com ne ti 1 and Tau
Figure S1. Size of the first upper and lower molars of some European Myoglis samples.
24
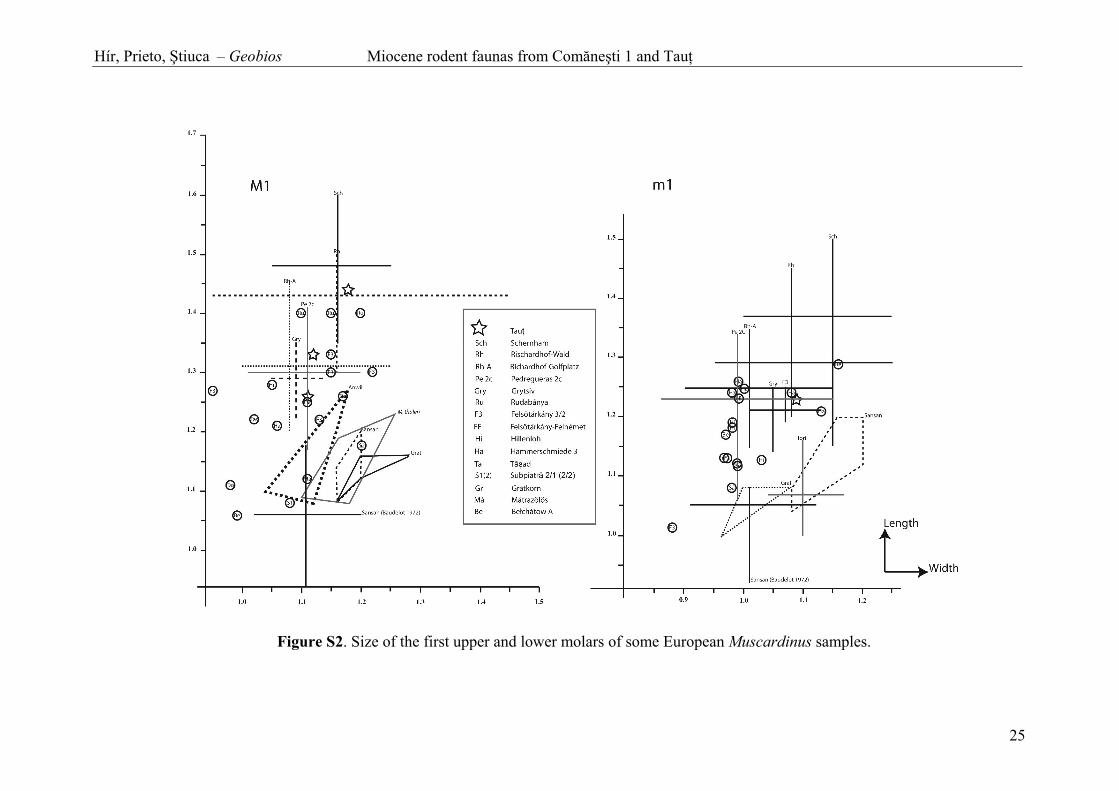
Hír, Prieto, tiuca – Geobios Miocene rodent faunas from Com ne ti 1 and Tau
Figure S2. Size of the first upper and lower molars of some European Muscardinus samples.
25
Recommended

















