
Ada3 Requirement for HAT Recruitment to Estrogen Receptors and
Estrogen-Dependent Breast Cancer Cell Proliferation
Aleksandra Germaniuk-Kurowska,1,3Alo Nag,
1,3Xiangshan Zhao,
1,3Manjari Dimri,
2,3
Hamid Band,2,3,4
and Vimla Band1,3,4
Divisions of 1Cancer Biology and 2Molecular Oncology, Evanston Northwestern Healthcare Research Institute; 3Department of Medicine,Feinberg School of Medicine; 4Department of Biochemistry, Molecular Biology and Cell Biology, Weinberg College of Arts,Northwestern University, Evanston, Illinois
Abstract
We have previously shown that evolutionarily conservedalteration/deficiency in activation (Ada) protein associateswith and promotes estrogen receptor (ER)–mediated targetgene expression. Here, we examined the role of endogenousAda3 to recruit histone acetyl transferases (HAT) to an ER-responsive promoter and its role in estrogen-dependent cellproliferation and malignant phenotype. Using a combinationof glycerol gradient cosedimentation and immunoprecipita-tion analyses, we show that Ada3, ER, and three distinct HATs[p300, (p300/CBP-associated factor) PCAF, and general con-trol nonrepressed 5 (Gcn5)] are present in a complex. Usingchromatin immunoprecipitation analysis, we show that shorthairpin RNA (shRNA)–mediated knockdown of Ada3 inER-positive breast cancer cells significantly reduced theligand-dependent recruitment of p300, PCAF, and Gcn5 tothe ER-responsive pS2 promoter. Finally, we use shRNAknockdown to show that Ada3 is critical for estrogen-dependent proliferation of ER-positive breast cancer cell linesin two-dimensional, as well as three-dimensional, culture.Knockdown of Ada3 in ER-positive MCF-7 cells inducedreversion of the transformed phenotype in three-dimensionalculture. Thus, our results show an important role of Ada3in HAT recruitment to estrogen-responsive target genepromoters and for estrogen-dependent proliferation of breastcancer cells. [Cancer Res 2007;67(24):11789–97]
Introduction
Estrogens play pivotal functional roles in the development,differentiation, and oncogenesis in female reproductive organs,such as the mammary gland and uterus (1, 2). The effects ofestrogens are mediated through estrogen receptor a (ERa) andERh, members of the nuclear hormone receptor (NR) family. Thesereceptors serve as sequence-specific transcriptional regulators bybinding to specific estrogen-responsive elements (ERE) withintarget gene promoters (3, 4). In the absence of bound ligands,NRs repress target gene transcription by associating with histonedeacetylase–containing corepressor complexes. Ligand bindingtriggers the release of corepressors and subsequent association
with coactivator complexes that are needed for crucial steps ingene transcription, including chromatin modification, remodeling,and recruitment of the RNA polymerase II holoenzyme (reviewed inrefs. 4–6).
Two groups of NR coactivators have been widely studied: (a) thep300 and the highly related cAMP-responsive element bindingprotein–binding protein (CBP) and (b) the mammalian Mediatorcomplexes (TRAP, DRIP, ARC, CRSP, SMCC, etc.; refs. 4–6). The p300and CBP proteins are recruited to ligand-activated, DNA-boundNRs by the steroid receptor coactivator (SRC) family (SRC-1, SRC-2,and SRC-3) of bridging factors (4–6). The SRC proteins havereceptor interaction domains containing LXXLL motifs thatcontact the ligand-binding domains (7, 8). The p300 and CBPproteins are histone acetyltransferases (HAT) that mediate theacetylation of nucleosomal histones, a covalent modificationgenerally associated with the enhancement of transcription.Furthermore, the HAT-dependent acetylation of transcriptionalactivators themselves promotes their activity (9).
Recent studies have identified an evolutionarily conservedcoactivator complex as a potential regulator of ER and otherNRs. Initially identified through genetic studies in yeast, the corecomponents of the alteration/deficiency in activation (Ada)complex include the adapter proteins Ada3 and Ada2 and generalcontrol nonrepressed 5 (Gcn5), a HAT (10). Mammalian Ada3exists as a component of a yeast Ada-like complex that includesAda2 and Gcn5, indicating evolutionary conservation of itsfunction (11). Initial studies of mammalian retinoid X receptor(RXR) and GR expressed in the yeast indicated a requirementof yeast Ada complex, including the yAda3 gene product, fortranscriptional activation (12). We and others have shown inmammalian cell systems that Ada3 indeed interacts with NRs,including RXR and ER (13, 14), as well as some non-NRtranscriptional factors, such as p53 (15–17). Notably, overexpres-sion of Ada3 enhanced ER-mediated target gene expression,whereas down-regulation of its expression had an opposite effect(14). These studies have raised the possibility that Ada3, as acomponent of ER coactivator complexes, may control ER-mediatedbiological outputs in mammalian cells. However, whether Ada3 atphysiologically expressed levels indeed functions in this capacity isnot known. This issue is particularly important because othermultiple adaptor proteins, such as SRCs (see above), have beenpreviously shown to control ER function in mammalian cells(1–6, 18). Furthermore, mammalian cells express Gcn5, as well asclosely related HAT, called p300/CBP-associated factor (PCAF;ref. 19); the known interaction of PCAF with p300 might suggestthat Ada3 may not play an important role in ER function.Alternatively, the ability of PCAF and apparently Gcn5 to interactwith Ada3, as well as p300, might suggest that these proteins mayfunction in concert and that Ada3 may play a crucial role in their
Note: Current address for X. Zhao and V. Band: Department of Genetics, CellBiology, and Anatomy, Nebraska Medical Center, Omaha, NE 68198-5805.
Current address for H. Band: Eppley Institute in Cancer & Allied Diseases, EppleyCancer Center/UNMC, Omaha, NE 68198-5805.
Requests for reprints: Vimla Band, Department of Genetics, Cell Biology &Anatomy, 985805 Nebraska Medical Center, Omaha, NE 68198-5805. Phone: 402-559-8565; Fax: 402-559-7328; E-mail: [email protected].
I2007 American Association for Cancer Research.doi:10.1158/0008-5472.CAN-07-2721
www.aacrjournals.org 11789 Cancer Res 2007; 67: (24). December 15, 2007
Research Article
Research. on November 29, 2015. © 2007 American Association for Cancercancerres.aacrjournals.org Downloaded from
Research. on November 29, 2015. © 2007 American Association for Cancercancerres.aacrjournals.org Downloaded from
Research. on November 29, 2015. © 2007 American Association for Cancercancerres.aacrjournals.org Downloaded from

recruitment to ligand-bound ER on target gene promoters andsubsequent functional responses.
In this study, we show that endogenously expressed Ada3 ispresent in a complex that includes ER and multiple HATs (p300,Gcn5, and PCAF) and that Ada3 is required for the recruitmentof these HATs to an ER target gene promoter in the context ofintact chromatin. We also show Ada3 is required for ER-mediatedproliferation of breast cancer cells in two-dimensional, as well asthree-dimensional, cultures, with a reversal of the transformedphenotype in three-dimensional cultures of MCF-7 cells upon Ada3knockdown. These studies underscore a crucial role of Ada3 inorchestrating ER-mediated gene transcription, as well as down-stream biological outputs in the context of breast cancer cells.
Materials and Methods
Cells and media. The ER-positive human breast cancer cell lines MCF-7and ZR-75-1 were grown in a-MEM medium (Life Technologies)
supplemented with 10% fetal bovine serum (FBS). For some experiments,
the ER-positive cell lines were cultured for 72 h in a phenol red–free
a-medium (Hyclone) supplemented with 10% charcoal/dextran-treated FBS(Hyclone) to remove residual 17h-estradiol (E2).
Antibodies. Generation of anti-hAda3 rabbit polyclonal antiserum has
been described previously (15). Antibodies against p53 (DO1), ERa, p300,
PCAF, Gcn5, and h-actin were purchased from Santa Cruz Biotechnology,Inc. Anti–Flag–horseradish peroxidase (HRP) antibodies were from Sigma.
h-Actin antibodies were from Abcam, Inc.
Generation of stable Ada3 short hairpin RNA knockdown cells. The
hAda3-specific RNA interference sequences used in short hairpin RNA(shRNA) constructs are GCAATCAGAACAAGCCCTT (#1) and GGTGACA-
GACGATTCCTGA (#2). The oligonucleotides were cloned in the pSUPER-
Retro vector (OligoEngine). MCF-7 and ZR-75-1 cells were infected withhAda3 RNA interference retroviral supernatants as described previously
(14, 17). Virally transduced cells were selected and maintained in 0.5 Ag/mL
puromycin for 72 h, and expression of endogenous hAda3 was assessed
in the whole-cell lysate using anti-hAda3 polyclonal antibody followed byWestern blotting.
Cell proliferation assay. Cells (2 � 104) from MCF-7 or ZR-75-1, stably
expressing scrambled shRNA or two independent shRNA against hAda3,
were plated in six-well plates for growth analyses. Cells were seeded inregular a-MEM medium supplemented with 0.5 Ag/mL of puromycin. After
24 h, medium was replaced with phenol red–free a-medium (plus 0.5 Ag/mL
puromycin) to remove residual E2 and, after that, treated with either vehicleor 100 pmol/L E2. The cells were trypsinized and counted at the indicated
time points.
Three-dimensional culture on Matrigel. For the three-dimensional
tissue culture system, the procedure was essentially as described previously(20, 21) with some modifications. Briefly, single-cell suspensions were
prepared by trypsinization and 2.5 � 103 cells in 0.4 mL of 2% Matrigel
(phenol red–free Matrigel from BD Biosciences) in phenol red–free a-
medium. For stable cells containing shRNAs, medium was supplementedall the time with 0.5 Ag/mL puromycin. Cells were plated in eight-well
chamber slides (BD Biosciences) on top of a 40-AL polymerized layer
of 100% Matrigel. The cultures were fed every 3 days. For some samplesafter E2 deprivation, 100 pmol/L E2 was included in the feeding medium
through the end of experiment. Phase contrast images were documented
periodically during the entire growth period.
Separation of native hAda3-associated complexes. MCF-7 cells werecultured for 72 h in phenol red–free a-medium (Hyclone) to remove all
residual E2 and, after that, treated for 24 h with either vehicle or 100 pmol/L
E2. Whole-cell extracts (f4 mg of protein) were dialyzed 3 h at 4jC against
buffer containing 50 mmol/L HEPES (pH 7.9), 50 mmol/L NaCl, 0.1 mmol/LEDTA, 1 mmol/L phenylmethylsulfonyl fluoride (PMSF). The dialyzed
material was centrifuged to remove all insoluble material and then layered
onto 9 mL of a 10% to 30% linear glycerol gradient and centrifuged for
18 h at 41,000 rpm at 4jC using a TH641 rotor (Sorvall). Gradients were
fractionated by collecting 360 AL aliquots. Fractions were trichloroaceticacid precipitated and then analyzed by Western blotting using specific
antibodies, as indicated in the figure. Additionally, high molecular weight
markers (HMW; Sigma) were centrifuged on the separate gradients, and one
of five from every fraction was separated by 10% SDS-PAGE and stainedwith Coomassie Brilliant Blue. This allowed the calculation of the apparent
molecular weights corresponding to particular fractions of the gradient.
Chromatin immunoprecipitation analysis. The chromatin immuno-
precipitation (ChIP) assay was performed as described previously (22). Inbrief, MCF-7 cells were cultured in phenol red–free medium (Hyclone) for
3 days followed by either mock treatment or 10 nmol/L E2 for 30 min. After
that, cells were treated with 1% formaldehyde to cross-link the DNA-protein
complexes. The cross-linking was quenched by adding glycine solution.Cells were washed with cold PBS. A small portion of the cross-linked,
sheared chromatin solution was saved as input DNA (5%), and the
remainder was used for immunoprecipitation with anti-ERa antibody(Ab-10, Neo Marker), anti-p300 (RW128, Upstate), anti-RNA polymerase II
(8WG16, Abcam), anti-PCAF (E-8, Santa Cruz), anti-Gcn5 (H-75, Santa
Cruz), and hAda3 (15). Immunoprecipitated DNA was deproteinized by
phenol-chloroform extraction, precipitated by ethanol, and resuspended inTE buffer. PCR amplifications were performed with primers covering pS2
promoter (region from �353 to �30): forward primer 5¶-GGCCATCTCT-
CACTATGAATCACTTCTGC-3¶ and reverse primer 5¶-GGCAGGCTCTGTT-
TGCTTAAAGAGCG-3¶. Control PCR was performed using primers coveringthe region upstream to pS2 promoter (region from �679 to �501): for-
ward primer 5¶-GTGATTCTCCTGACTTAACC-3¶ and reverse primer 5¶-TG-
GCGCAGTGGCTCACGCTG-3¶. Glyceraldehyde-3-phosphate dehydrogenase(GAPDH) primers were used as control (14).
Immunoblots and immunoprecipitation. For immunoblots, 80 Ag of
cell lysate was fractionated by SDS-PAGE, transferred to a polyvinylidene
difluoride membrane, and probed with antibodies. Enhanced chemilumi-nescence reagents were used for signal detection. For immunoprecipita-
tion experiments, MCF-7 cells were maintained in a phenol red–free
a-medium supplemented with 10% charcoal/dextran-treated FBS (Hyclone)
for 72 h and then treated with either 100 pmol/L E2 or vehicle for 24 h.
Results
Ada3 cosediments with multiple HATs upon E2 treatment ofbreast cancer cell lines. Whereas yeast Ada3 functions as a com-ponent of the ADA complex with Gcn5 as the HAT (23), mammalianAda3 was found to associate with p300 (16, 17) and was present inthe same complex as PCAF in a cervical cancer cell line (24). Thus,Ada3 may form multiple independent coactivator complexes withvarious HATs or a larger complex containing multiple HATs. We usedcosedimentation analysis on glycerol gradients to isolate intactAda3-containing complexes from untreated and E2-treated breastcancer cell line MCF-7 to address this issue. Cells were lysed undermild conditions (50 mmol/L NaCl) to ensure that even potentiallyweak protein-protein interactions remained intact. Equal amountsof protein extracts were subjected to glycerol gradient centrifugationand Ada3-containing complexes were identified by Westernimmunoblotting of individual fractions. Internal standards wereused to determine the relative molecular sizes of isolated complexes.
In estrogen-deprived MCF-7 cells, the majority of Ada3 wasfound in fractions 11 to 14 (Fig. 1A–D). Upon E2 treatment, theAda3 protein showed a shift toward higher molecular weightfractions ( fractions 13–16), suggestive of association with addi-tional proteins. As expected, a proportion of the ER protein, as wellthe Ada3-binding partner p53, was also present in the samefractions as Ada3 (Fig. 1A–D), confirming our previous studies(15, 17). Whereas p53 remained relatively unchanged in its locationin the gradients, the ER peak shifted into higher molecular weightfractions ( fractions 11–14 versus fractions 13–18).
Cancer Research
Cancer Res 2007; 67: (24). December 15, 2007 11790 www.aacrjournals.org
Research. on November 29, 2015. © 2007 American Association for Cancercancerres.aacrjournals.org Downloaded from

We also immunoblotted the fractionated proteins to determinethe location of potential HAT partners of Ada3: p300, Gcn5, andPCAF. Each of these HATs showed partial cosedimentation withAda3, which, with PCAF and Gcn5, was relatively modest before E2treatment (Fig. 1A); however, the cosedimentation was more clearlyobserved upon E2 treatment (Fig. 1B), consistent with theirincorporation into an Ada3-containing complex. The peak p300
fractions did not show a significant shift upon E2 treatment;however, we have frequently noticed increased intensity of p300bands in E2 versus untreated cell fractions (Fig. 1). Cosedimenta-tion of the three HAT proteins (p300, Gcn5, and PCAF), as well asthe ER, with Ada3 and their shift into overlapping fraction uponE2 treatment supports the idea that Ada3 is present in a complexwith these proteins.
Figure 1. Biochemical fractionation ofnative Ada3-containing complexes inbreast cells. Whole-cell extracts fromMCF-7, either vehicle-treated (A) or treatedwith 100 pmol/L of E2 for 24 h (B ; f4 mgof protein), as well as HMW (Sigma),were dialyzed 3 h at 4jC against buffer X[50 mmol/L HEPES (pH 7.9), 50 mmol/LNaCl, 0.1 mmol/L EDTA, 1 mmol/L PMSF]and then layered on top of 9 mL of a 10%to 30% linear glycerol gradient andcentrifuged O/N at 41,000 rpm at 4jC,using a TH641 rotor (Sorvall). Gradientswere fractionated by collecting 360 ALaliquots from the top of the gradients.Fractions were TCA precipitated and thenanalyzed by Western blotting using specificantibodies, as indicated in the figure. Onefifth from every fraction of the separatedHMW markers was separated by 10%SDS-PAGE and stained with CoomasieBrilliant Blue (data not shown). Thisallowed the calculation of the apparentmolecular weight corresponding toparticular fractions of the gradient.The intensity of the bands in particularfractions of the gradients of cells, eithervehicle-treated (C ) or treated with 100pmol/L of E2 for 24 h (D), was quantifiedby densitometry using Scion ImageSoftware. The graphs show the intensity ofthe bands (Y axis) versus fraction number(X axis). The experiments were repeatedat least thrice.
Ada3 Requirement for ER-Dependent Cell Proliferation
www.aacrjournals.org 11791 Cancer Res 2007; 67: (24). December 15, 2007
Research. on November 29, 2015. © 2007 American Association for Cancercancerres.aacrjournals.org Downloaded from

Ada3 associates with PCAF, Gcn5, and p300 in breast cancercell lines. To further substantiate the association of Ada3 withvarious HATs, we performed coimmunoprecipitation analysesusing lysates of untreated or E2-treated MCF-7 cells expressing aFlag-Ada3. Ectopically expressed Flag-Ada3 was used for theseanalyses to overcome the uncertainty about the efficiency ofimmunoprecipitation because of the comigration of endogenousAda3 with the heavy chain of immunoprecipitation antibody(data not shown). Anti-Flag immunoprecipitation followed by anti–Flag-HRP immunoblotting showed efficient immunoprecipitationof Ada3 protein (Fig. 2). As expected, anti-ER immunoblottingshowed a clear association between Ada3 and ER (Fig. 2). Notably,all three HATs, p300, PCAF, and Gcn5, were also coimmunopre-cipitated with the Ada3 protein. These results complement thecosedimentation analyses, indicating that Ada3 associates with allthree HATs in addition to the ER.
Ligand-dependent recruitment of Ada3 and HATs to an ERtarget gene promoter. Given the critical role of coactivatorrecruitment to transcriptional activators bound to target genepromoters (25) and our previous findings that Ada3 is recruitedto native estrogen-responsive promoter pS2 (14), we performedChIP analysis of untreated and E2-treated MCF-7 cell chromatinextracts to assess the corecruitment of Ada3 and HATs on pS2promoter. As expected, both ER and Ada3 were found to bebound to pS2 promoter, and treatment with E2 led to asignificant increase in the pS2 promoter occupancy by bothERa and Ada3 (Fig. 3). Parallel ChIP analysis of HATs showed thatp300, Gcn5, and PCAF are recruited to the pS2 promoter upon E2treatment (Fig. 3); concurrently, we also observed that RNApolymerase II was present in ChIPs, as anticipated (26). Nosignificant signals were detected from either the preimmuneserum or mouse or rabbit IgG (Fig. 3). Control PCR analysis of aregion upstream of the ERE element in the pS2 promoter andGAPDH promoter also showed no detectable signals (Fig. 3).These results clearly indicated that all three HATs examined are
recruited to the pS2 promoter in addition to Ada3 and ER andraised the possibility that Ada3 may play a role in the recruitmentprocess (addressed below).
Ada3 is required for optimal association of HATs and ERAto the pS2 promoter in a native chromatin context. As Ada3serves as an adapter within the ADA complexes, we surmised thatit might play a role in Gcn5 recruitment to ER target promoters;as the mechanisms of PCAF and p300 recruitment to ER targetpromoter remain to be determined, we also asked if Ada3 playsany role in their recruitment. For this purpose, we generatedMCF-7 cells stably expressing either a scrambled shRNA or twoindependent shRNAs against Ada3. Anti-Ada3 immunoblotting ofcell lysates showed the specific knockdown of Ada3 in Ada3shRNA but not in control shRNA-expressing cells (Fig. 4). Extractsof untreated or E2-treated cells were then subjected to ChIPanalyses to assess recruitment of various proteins to pS2promoter. MCF-7 cells expressing the scrambled shRNA controlshowed the anticipated ligand-promoted recruitment of Ada3,ERa, p300, PCAF, Gcn5, and RNA polymerase II to the pS2promoter with no signals in negative control lanes (preimmuneserum, mouse or rabbit IgG, or PCR of upstream region in pS2promoter or GAPDH promoter; Fig. 5). Notably, ChIPs of cellsexpressing shRNAs against hAda3 showed a substantial decreasein the recruitment of ER and all three HATs to the pS2 promotercompared with cells expressing the control shRNA (Fig. 5). Theseresults suggest that endogenous Ada3 plays an essential role inthe recruitment of all three HATs examined to an ER targetpromoter. Interestingly, Ada3 knockdown also resulted in reducedChIP with anti-ER antibodies, indicating that Ada3 also contrib-utes to the stability of ER association with a target promoter innative chromatin context.
Ada3 is required for ER-dependent cell proliferation. Inview of the critical role of endogenous Ada3 in the recruitment ofHATs (and apparently to the stability of ER occupancy) to an ERtarget promoter, as revealed by ChIP analyses of Ada3knockdown cells, we hypothesized that Ada3 will be requiredfor downstream functional consequences of ER-dependenttranscriptional activation. Therefore, we compared the level ofE2-induced cell proliferation in control versus Ada3 shRNA-expressing MCF-7 cells, as well as a similar pair of another ER-positive breast cancer cell line, ZR-75-1. Western blot analysis ofcell lysates showed the specific knockdown of Ada3 protein inAda3 shRNA-expressing cells compared with scrambled shRNA-expressing cells, with more substantial knockdown with Ada3shRNA #2 versus #1 (Fig. 4). Assessment of cell proliferation inresponse to E2 over time showed the expected E2-inducedproliferation of control shRNA-expressing MCF-7 and ZR-75-1cells scrambled shRNA, whereas no significant proliferation wasobserved in the absence of E2 (Fig. 4). In contrast, cells withAda3 knockdown showed a substantial reduction in E2-inducedcell proliferation at all times; the inhibition was more marked(f50% reduction at day 7) in cells expressing shRNA #2, inwhich a higher degree of knockdown was observed comparedwith those expressing shRNA #1 (f25% reduction at day 7;Fig. 4). These results underscore the important role of Ada3 inestrogen-dependent cell proliferation.
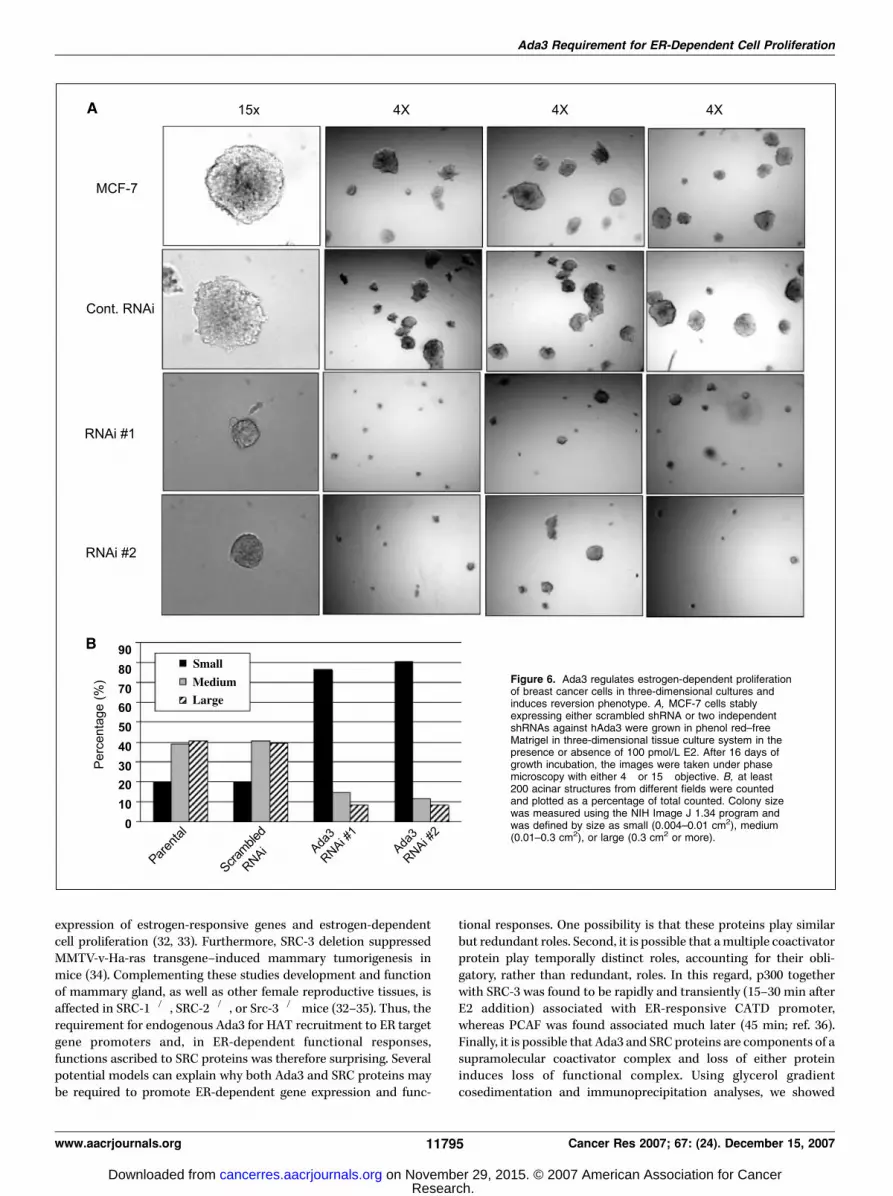
Essential role of Ada3 in estrogen-dependent malignantphenotype of breast cancer cells in three-dimensional Matrigelculture. Compared with the conventional two-dimensional tissueculture system, the Matrigel-supported three-dimensional culturesystem allows analysis of normal and cancerous breast epithelial
Figure 2. Ada3 interacts with PCAF, Gcn5, and p300 in breast cancer cells.MCF-7 cells stably expressing Flag-Ada3 were starved for 72 h, eithervehicle-treated or treated with 100 pmol/L of E2 for 24 h, and then lysed. Lysates(3 mg) were incubated with either anti-flag or nonspecific antibodies for 2 h at4jC. After adding protein G–Sepharose beads at 4jC for 1 h, the bound proteinswere subjected to Western blotting analysis with anti–Flag-HRP to detecthAda3 or other indicated antibodies to detect various proteins.
Cancer Research
Cancer Res 2007; 67: (24). December 15, 2007 11792 www.aacrjournals.org
Research. on November 29, 2015. © 2007 American Association for Cancercancerres.aacrjournals.org Downloaded from

cell proliferation that more closely mimics that observed in vivo(27). We therefore assessed the role of endogenous Ada3 in theE2-dependent proliferation of MCF-7 cells in three-dimensionalculture. For this purpose, MCF-7 cells expressing scrambled orAda3 shRNA were cultured for 4 days in phenol red–free Matrigel,using previously established methods (28), followed by feeding withmedium containing E2 (100 pmol/L) for a period of 2 weeks. Inthe absence of exogenous E2, neither the control nor the Ada3knockdown cells displayed any proliferation (data not shown).However, when E2 was included in the growth medium, the controlshRNA-expressing cells formed large irregular colonies typical ofbreast cancer cell growth on Matrigel (Fig. 6A ; refs. 20, 21, 27).In contrast, MCF-7 cell lines expressing two independent shRNAsdeveloped markedly smaller structures (Fig. 6A). The extent ofreduction in colony size was quantified by enumerating small(0.004–0.01 cm2), medium (0.01–0.3 cm2), or large (0.3 cm2 or more)colonies. Whereas parental and control shRNA-expressing MCF-7cells form predominantly large and medium-sized colonies, thosewith Ada3 shRNA form primarily small colonies (Fig. 6B). Notably,the colonies formed by Ada3 knockdown MCF-7 cells were moreregular and acinus-like, similar to those reported by the Bisselllaboratory for other treatments that induce a reversal of themalignant phenotype of breast cancer cells on Matrigel (Fig. 6A;
ref. 29). These results provide evidence that Ada3 protein isrequired for E2-dependent proliferation under three-dimensionalgrowth conditions that closely mimic in vivo cell proliferation.
Discussion
Estrogens play crucial roles in the development and function ofvarious female reproductive tissues and are critically important inhuman cancer (1, 2). For example, nearly 70% to 80% breast cancersare ER-positive and antiestrogens, and aromatase inhibitors arenow used as a first line of treatment for such patients (1, 2, 30).Similar to NRs in general, the ability of estrogens to serve asactivating switches to turn on ER-responsive genes and the ensuingcontrol of cellular functions requires coregulatory proteins;corepressors keep ER targets repressed, whereas coactivatorspromote target gene expression (5–8). The latter is mediatedthrough the recruitment of HATs, which allow chromatinremodeling required for optimal transcriptional activation (5–8, 25).Thus far, the best characterized ER coactivators are members ofthe SRC family, which have been linked to the recruitment ofHAT proteins p300 and PCAF (5–8, 31–34). We have previouslyidentified Ada3, an evolutionarily conserved component of ADA3coactivator complex, as a coactivator of ER-mediated target gene
Figure 3. Ada3 is recruited to the endogenousestrogen promoter along with HATs in aligand-dependent manner. Soluble chromatinwas prepared from MCF-7 cells, eithervehicle-treated or treated with 10 nmol/L E2 for30 min, and immunoprecipitated with antibodiesagainst ERa, Ada3, p300, Gcn5, PCAF, or RNApolymerase II. The final DNA extractions wereamplified using pairs of primers that cover theregion of pS2 promoter. PCR amplification ofthe GAPDH promoter or region upstream to pS2promoter was used as specificity control.
Ada3 Requirement for ER-Dependent Cell Proliferation
www.aacrjournals.org 11793 Cancer Res 2007; 67: (24). December 15, 2007
Research. on November 29, 2015. © 2007 American Association for Cancercancerres.aacrjournals.org Downloaded from

expression (14). Here, we show that Ada3 associates with multipleHATs (p300, PCAF, and GCN5) and that endogenous Ada3 inhuman ER-positive breast cancer cell lines is essential for therecruitment to ER target gene promoters, suggesting its role as akey organizer of HAT recruitment and ER-dependent transcription.We further show that Ada3 is essential for ER-dependent functionalresponses, as shown by a marked inhibition of estrogen-dependentproliferation of human breast cancer cell lines with Ada3knockdown and reversal of the malignant behavior of such cells
when grown in three-dimensional culture on Matrigel. Thus, ourresults suggest a critical role of Ada3 as a regulator of ER functionin the context of human ER-positive breast cancer cell proliferationand malignant phenotype.
It is noteworthy that estrogen-regulated gene transcription,proliferation, and oncogenesis in mammary cell systems requiremembers of the SRC family (31–34). For example, estrogen-dependent proliferation of MCF-7 cells was reduced upon SRC-3depletion (35). Similarly, depletion of SRC-1 or SRC-2 inhibit the
Figure 5. Ada3 regulates estrogen-dependentproliferation of breast cancer cells. ER-positive breastcancer cells (MCF-7 and ZR-75-1) stably expressingeither scrambled shRNA or two independent shRNAsagainst hAda3 were cultured for 72 h in a phenolred–free medium followed by stimulation with100 pmol/L of E2 for 7 d. At the indicated timepoints, cells were trypsinized and counted usinghemocytometer. Cells from the parallel cultures wereharvested, and total cell lysates were subjected toWestern blot analysis using antibodies againsthAda3 and h-actin (as a loading control). Next toeach graph are the levels of Ada3 protein in cellsused in this experiment. Points , mean of triplicates;bars , SD.
Figure 4. Ada3 regulates recruitment ofHATs to the promoter of estrogen-responsivegene. MCF-7 cells were infected with eitherscrambled shRNA or two independentshRNAs against hAda3. Cells were eithervehicle-treated or treated with 10 nmol/L ofE2 for 30 min. Equal amounts of solublechromatin were subjected forimmunoprecipitation using indicatedantibodies. The final DNA extractions wereamplified using pairs of primers that coverthe region of pS2 promoter. PCR amplificationof the GAPDH promoter or region upstreamto pS2 promoter was used as specificitycontrol.
Cancer Research
Cancer Res 2007; 67: (24). December 15, 2007 11794 www.aacrjournals.org
Research. on November 29, 2015. © 2007 American Association for Cancercancerres.aacrjournals.org Downloaded from
expression of estrogen-responsive genes and estrogen-dependentcell proliferation (32, 33). Furthermore, SRC-3 deletion suppressedMMTV-v-Ha-ras transgene–induced mammary tumorigenesis inmice (34). Complementing these studies development and functionof mammary gland, as well as other female reproductive tissues, isaffected in SRC-1�/�, SRC-2�/�, or Src-3�/� mice (32–35). Thus, therequirement for endogenous Ada3 for HAT recruitment to ER targetgene promoters and, in ER-dependent functional responses,functions ascribed to SRC proteins was therefore surprising. Severalpotential models can explain why both Ada3 and SRC proteins maybe required to promote ER-dependent gene expression and func-
tional responses. One possibility is that these proteins play similarbut redundant roles. Second, it is possible that a multiple coactivatorprotein play temporally distinct roles, accounting for their obli-gatory, rather than redundant, roles. In this regard, p300 togetherwith SRC-3 was found to be rapidly and transiently (15–30 min afterE2 addition) associated with ER-responsive CATD promoter,whereas PCAF was found associated much later (45 min; ref. 36).Finally, it is possible that Ada3 and SRC proteins are components of asupramolecular coactivator complex and loss of either proteininduces loss of functional complex. Using glycerol gradientcosedimentation and immunoprecipitation analyses, we showed
Figure 6. Ada3 regulates estrogen-dependent proliferationof breast cancer cells in three-dimensional cultures andinduces reversion phenotype. A, MCF-7 cells stablyexpressing either scrambled shRNA or two independentshRNAs against hAda3 were grown in phenol red–freeMatrigel in three-dimensional tissue culture system in thepresence or absence of 100 pmol/L E2. After 16 days ofgrowth incubation, the images were taken under phasemicroscopy with either 4� or 15� objective. B, at least200 acinar structures from different fields were countedand plotted as a percentage of total counted. Colony sizewas measured using the NIH Image J 1.34 program andwas defined by size as small (0.004–0.01 cm2), medium(0.01–0.3 cm2), or large (0.3 cm2 or more).
Ada3 Requirement for ER-Dependent Cell Proliferation
www.aacrjournals.org 11795 Cancer Res 2007; 67: (24). December 15, 2007
Research. on November 29, 2015. © 2007 American Association for Cancercancerres.aacrjournals.org Downloaded from

that Ada3 is present in the same complex as ER, as well as three HATs(p300, PCAF, and Gcn5), consistent with the model that Ada3 is acritical component of a large complex (composed of Ada3, Ada2,Gcn5, p300, and PCAF), which plays an essential role in trans-criptional activation by ER and potentially other transcriptionalactivators. Future studies using combinatorial knockdown in cellsand compound mouse knockouts should help test these models.
Ada3 was initially identified as a core component of the yeastADA coactivator complex that includes an adaptor protein Ada2,which in turn associates with a HAT protein Gcn5 (37). Ada3interacts directly with the activation domains of transcriptionalcomplexes, thus serving as a key component in the recruitment ofAda complex and its associated HAT activity to transcriptionalactivators bound to specific promoters. Given our currentobservations that Ada3 facilitates the recruitment of multipleHATs, including Gcn5, to ER target promoters, and recent studiesindicating that Ada3 (and other Ada components) assemble intomultiple complexes that can vary significantly from the trimolec-ular yeast ADA complex, Ada3 may play multiple functional roles inthe context of different transcriptional activators. Adding to thiscomplexity, mammalian cells seem to contain a Gcn5 complex thatdoes not include Ada2 (38). Similarly, two distinct Ada2 proteinsexist in higher eukaryotic cells, and studies in Drosophila show thatboth Ada2 proteins are essential for development, indicative ofdistinct functions (39, 40). In general, however, Ada3 has beenfound as an invariant component of many ADA-related complexesin mammals and other higher eukaryotic cells, suggesting a pivotalrole of Ada3 in coactivator functions of these complexes. Inaddition, Ada3 has emerged as a binding partner of the human
papilloma virus E6 oncoprotein (15), suggesting the possibility thatviral oncoproteins may alter the function of Ada3 duringoncogenesis. Thus, detailed analysis of Ada3 and mechanisms ofits function are likely to elucidate basic regulation of geneexpression, as well as its aberrations in cancer.
In conclusion, we provide evidence that Ada3 associates with andis required for the recruitment of multiple HAT proteins (p300, Gcn5,and PCAF) to ER target gene promoters and that Ada3 is requiredfor ER-mediated cell proliferation. Future studies using total andmammary gland targeted knockout mice should further delineatethe important role of Ada3 versus other known ER coactivators, suchas SRC1 to SRC3 in ER function, either as redundant regulators or aspart of a single complex of temporally coordinated coactivatorcomplexes. Elucidation of the role of coactivators in ER functionshould provide insights relevant to normal reproductive tissuedevelopment and function, as well as oncogenesis.
Acknowledgments
Received 7/18/2007; revised 9/27/2007; accepted 10/12/2007.Grant support: NIH grants CA94143, CA96844, CA81076 (V. Band), CA87986,
CA76118, CA99900, and CA99163 (H. Band), National Cancer Institute (NCI) Center ofCancer Nanotechnology Excellence grant NCI 1U54 CA119341-01 (H. Band and V.Band), NCI Breast Cancer Specialized Programs of Research Excellence and AVONBreast Cancer Fund at Robert H. Lurie Cancer Center, Northwestern University,Duckworth Family Chair for Breast Cancer Research (V. Band), and Jean Ruggles-Romoser Chair for Cancer Research (H. Band).
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.
We thank Michael A. Sassack for help in this project and Drs. Qingshen Gao andGoberdhan Dimri and the members of Band laboratories for helpful suggestions anddiscussion.
Cancer Research
Cancer Res 2007; 67: (24). December 15, 2007 11796 www.aacrjournals.org
References
1. Nilsson S, Makela S, Treuter E, et al. Mechanisms ofestrogen action. Physiol Rev 2001;81:1535–65.
2. Russo J, Russo IH. The role of estrogen in the initiationof breast cancer. J Steroid Biochem Mol Biol 2006;102:89–96.
3. Robinson-Rechavi M, Escriva Garcia H, Laudet V.The nuclear receptor superfamily. J Cell Sci 2003;116:585–6.
4. Singh RR, Kumar R. Steroid hormone receptorsignaling in tumorigenesis. J Cell Biochem 2005;96:490–505.
5. Barnes CJ, Vadlamudi RK, Kumar R. Novel estrogenreceptor coregulators and signaling molecules in humandiseases. Cell Mol Life Sci 2004;61:281–91.
6. Smith CL, O’Malley BW. Coregulator function: a key tounderstanding tissue specificity of selective receptormodulators. Endocr Rev 2004;25:45–71.
7. Heery DM, Kalkhoven E, Hoare S, Parker MG. Asignature motif in transcriptional co-activators medi-ates binding to nuclear receptors. Nature 1997;387:733–6.
8. Needham M, Raines S, McPheat J, et al. Differentialinteraction of steroid hormone receptors with LXXLLmotifs in SRC-1a depends on residues flanking themotif. J Steroid Biochem Mol Biol 2000;72:35–46.
9. Katzenellenbogen BS, Choi I, Delage-Mourroux R, etal. Molecular mechanisms of estrogen action: selectiveligands and receptor pharmacology. J Steroid BiochemMol Biol 2000;74:279–85.
10. Marcus GA, Silverman N, Berger SL, Horiuchi J,Guarente L. Functional similarity and physical associ-ation between GCN5 and ADA2: putative transcriptionaladaptors. EMBO J 1994;13:4807–15.
11. Horiuchi J, Silverman N, Marcus GA, Guarente L.ADA3, a putative transcriptional adaptor, consists oftwo separable domains and interacts with ADA2 and
GCN5 in a trimeric complex. Mol Cell Biol 1995;15:1203–9.
12. vom Baur E, Harbers M, Um SJ, Benecke A,Chambon P, Losson R. The yeast Ada complex mediatesthe ligand-dependent activation function AF-2 ofretinoid X and estrogen receptors. Genes Dev 1998;12:1278–89.
13. Zeng M, Kumar A, Meng G, et al. Human papillomavirus 16 E6 oncoprotein inhibits retinoic X receptor-mediated transactivation by targeting human ADA3coactivator. J Biol Chem 2002;277:45611–8.
14. Meng G, Zhao Y, Nag A, et al. Human ADA3 binds toestrogen receptor (ER) and functions as a coactivatorfor ER-mediated transactivation. J Biol Chem 2004;279:54230–40.
15. Kumar A, Zhao Y, Meng G, et al. Human papil-lomavirus oncoprotein E6 inactivates the transcrip-tional coactivator human ADA3. Mol Cell Biol 2002;22:5801–12.
16. Wang T, Kobayashi T, Takimoto R, et al. hADA3 isrequired for p53 activity. EMBO J 2001;20:6404–13.
17. Nag A, Germaniuk-Kurowska A, Dimri M, et al. Anessential role of human Ada3 in p53 acetylation. J BiolChem 2007;282:8812–20.
18. Shao W, Brown M. Advances in estrogen receptorbiology: prospects for improvements in targeted breastcancer therapy. Breast Cancer Res 2004;6:39–52.
19. Marmorstein R. Structure of histone acetyltrans-ferases. J Mol Biol 2001;311:433–44.
20. Debnath J, Muthuswamy SK, Brugge JS. Morphogen-esis and oncogenesis of MCF-10A mammary epithelialacini grown in three-dimensional basement membranecultures. Methods 2003;30:256–68.
21. Dimri M, Naramura M, Duan L, et al. Modeling breastcancer-associated c-Src and EGFR overexpression inhuman MECs: c-Src and EGFR cooperatively promoteaberrant three-dimensional acinar structure and inva-sive behavior. Cancer Res 2007;67:4164–72.
22. Christova R, Oelgeschlager T. Association of humanTFIID-promoter complexes with silenced mitotic chro-matin in vivo . Nat Cell Biol 2002;4:79–82.
23. Eberharter A, Sterner DE, Schieltz D, et al. TheADA complex is a distinct histone acetyltransferasecomplex in Saccharomyces cerevisiae . Mol Cell Biol1999;19:6621–31.
24. Ogryzko VV, Kotani T, Zhang X, et al. Histone-likeTAFs within the PCAF histone acetylase complex. Cell1998;94:35–44.
25. Dilworth FJ, Chambon P. Nuclear receptors coordi-nate the activities of chromatin remodeling complexesand coactivators to facilitate initiation of transcription.Oncogene 2001;20:3047–54.
26. Eeckhoute J, Keeton EK, Lupien M, Krum SA, CarrollJS, Brown M. Positive cross-regulatory loop ties GATA-3to estrogen receptor a expression in breast cancer.Cancer Res 2007;67:6477–83.
27. Nelson CM, Bissell MJ. Modeling dynamic reciprocity:engineering three-dimensional culture models of breastarchitecture, function, and neoplastic transformation.Semin Cancer Biol 2005;15:342–52.
28. Li T, Sotgia F, Vuolo MA, et al. Caveolin-1 mutationsin human breast cancer: functional association withestrogen receptor a-positive status. Am J Pathol 2006;168:1998–2013.
29. Itoh M, Nelson CM, Myers CA, Bissell MJ. Rap1integrates tissue polarity, lumen formation, and tumor-igenic potential in human breast epithelial cells. CancerRes 2007;67:4759–66.
30. Ellis MJ, Rigden CE. Initial versus sequential adjuvantaromatase inhibitor therapy: a review of the currentdata. Curr Med Res Opin 2006;22:2479–87.
31. Liao L, Kuang SQ, Yuan Y, Gonzalez SM, O’Malley BW,Xu J. Molecular structure and biological function of thecancer-amplified nuclear receptor coactivator SRC-3/AIB1. J Steroid Biochem Mol Biol 2002;83:3–14.
32. Xu J, Qiu Y, DeMayo FJ, Tsai SY, Tsai MJ, O’Malley BW.
Research. on November 29, 2015. © 2007 American Association for Cancercancerres.aacrjournals.org Downloaded from

Ada3 Requirement for ER-Dependent Cell Proliferation
www.aacrjournals.org 11797 Cancer Res 2007; 67: (24). December 15, 2007
Partial hormone resistance in mice with disruption ofthe steroid receptor coactivator-1 (SRC-1) gene. Science1998;279:1922–5.
33. Xu J, Li Q. Review of the in vivo functions of the p160steroid receptor coactivator family. Mol Endocrinol2003;17:1681–92.
34. Xu J, Liao L, Ning G, Yoshida-Komiya H, Deng C,O’Malley BW. The steroid receptor coactivator SRC-3(p/CIP/RAC3/AIB1/ACTR/TRAM-1) is required for nor-mal growth, puberty, female reproductive function, andmammary gland development. Proc Natl Acad SciU S A 2000;97:6379–84.
35. List HJ, Lauritsen KJ, Reiter R, Powers C, Wellstein A,Riegel AT. Ribozyme targeting demonstrates that thenuclear receptor coactivator AIB1 is a rate-limitingfactor for estrogen-dependent growth of human MCF-7breast cancer cells. J Biol Chem 2001;276:23763–8.
36. Shang Y, Hu X, DiRenzo J, Lazar MA, Brown M.Cofactor dynamics and sufficiency in estrogen receptor-regulated transcription. Cell 2000;103:843–52.
37. Saleh A, Lang V, Cook R, Brandl CJ. Identification ofnative complexes containing the yeast coactivator/repressor proteins NGG1/ADA3 and ADA2. J Biol Chem1997;272:5571–8.
38. Forsberg EC, Lam LT, Yang XJ, Nakatani Y, BresnickEH. Human histone acetyltransferase GCN5 exists in astable macromolecular complex lacking the adapterADA2. Biochemistry 1997;36:15918–24.
39. Kusch T, Guelman S, Abmayr SM, Workman JL.Two Drosophila Ada2 homologues function in dif-ferent multiprotein complexes. Mol Cell Biol 2003;23:3305–19.
40. Muratoglu S, Georgieva S, Papai G, et al. Twodifferent Drosophila ADA2 homologues are present indistinct GCN5 histone acetyltransferase-containingcomplexes. Mol Cell Biol 2003;23:306–21.
Research. on November 29, 2015. © 2007 American Association for Cancercancerres.aacrjournals.org Downloaded from

Correction: Ada3 Requirement for ER-Dependent CellProliferation
In the article on Ada3 requirement for ER-dependent cellproliferation in the December 15, 2007 issue of Cancer Research (1),the figure legends on page 11794 are switched; the legend for Fig. 4is next to Fig. 5, and the legend for Fig. 5 is next to Fig. 4.
I2008 American Association for Cancer Research.doi:10.1158/0008-5472.CAN-68-5-COR1
1. Germaniuk-Kurowska A, Nag A, Zhao X, Dimri M, Band H, Band V. Ada3requirement for HAT recruitment to estrogen receptors and estrogen-dependentbreast cancer cell proliferation. Cancer Res 2007;67:11789–97.
www.aacrjournals.org 1609 Cancer Res 2008; 68: (5). March 1, 2008
Correction

2007;67:11789-11797. Cancer Res Aleksandra Germaniuk-Kurowska, Alo Nag, Xiangshan Zhao, et al. ProliferationReceptors and Estrogen-Dependent Breast Cancer Cell Ada3 Requirement for HAT Recruitment to Estrogen
Updated version
http://cancerres.aacrjournals.org/content/67/24/11789
Access the most recent version of this article at:
Cited articles
http://cancerres.aacrjournals.org/content/67/24/11789.full.html#ref-list-1
This article cites 40 articles, 19 of which you can access for free at:
Citing articles
http://cancerres.aacrjournals.org/content/67/24/11789.full.html#related-urls
This article has been cited by 9 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications
Research. on November 29, 2015. © 2007 American Association for Cancercancerres.aacrjournals.org Downloaded from
Recommended

















