
ORIGINAL ARTICLE
Additions to the cytologically investigated species of Potentilla L.(Rosaceae) from India
Savita Rani • Sanjeev Kumar • Syed Mudassir Jeelani •
Raghbir Chand Gupta • Santosh Kumari
Received: 26 May 2011 / Accepted: 24 October 2011 / Published online: 15 December 2011
� Springer-Verlag 2011
Abstract At present 14 species of Potentilla L. have been
cytologically worked out from different geographical areas
of Kashmir and Himachal Pradesh in the Western Hima-
layas. New chromosome numbers in nine species—
Potentilla argyrophylla (n = 14), P. atrosanguinea
(n = 7, 14), P. desertorum (n = 7), P. gerardiana
(n = 14), P. indica (n = 14), P. micropetala (n = 14),
P. nepalensis (n = 14), P. sibbaldia (n = 14) and
P. thomsonii (n = 7)—have been reported on a worldwide
basis for the first time. Additional chromosomal races of
polyploid cytotypes for P. argyrophylla (n = 28) and
P. desertorum (n = 14) along with a diploid cytotype
for P. micropetala (n = 7) plus diploid cytotypes for the
five species as P. fulgens (n = 7), P. gelida (n = 7),
P. kleiniana (n = 7), P. sibbaldia (n = 7) and P. sundaica
(n = 7) as well as a tetraploid cytotype for P. fruticosa
(n = 14) all have been cytologically worked out from India
for the first time. The course of meiosis varies from normal
to abnormal in different populations of the majority of
the species, such as P. argyrophylla, P. atrosanguinea,
P. desertorum, P. fruticosa, P. fulgens, P. gelida, P. indica,
P. nepalensis, P. sibbaldia and P. sundaica, except for
normal meiosis observed in P. gerardiana, P. kleiniana,
P. micropetala and P. thomsonii. The anomalous taxa are
marked with meiotic abnormalities in the form of cyto-
mixis, chromosomal stickiness, unoriented bivalents, for-
mation of laggards and bridges resulting in abnormal
microsporogenesis, and production of heterogenous-sized
fertile pollen grains along with reduced pollen fertility. All
the taxa with normal meiotic courses show nearly one
hundred percent pollen fertility.
Keywords Cytotypes � Meiotic abnormalities �Pollen fertility � Polyploidy � Potentilla �Himachal Pradesh � Kashmir
Introduction
The genus Potentilla L. includes over 500 species, mainly
distributed in the temperate regions of the Northern
Hemisphere, including 40 species from India (Santapau
and Henry 1973). The species of the genus are character-
ized as perennial or annual herbs or small shrubs usually
with creeping stems rooted at nodes; leaves are pinnate or
palmately compound; the leaflets are usually lobed with
stipules adnate to the petiole. These species prefer open
habitats, especially alpine or sub-alpine, even in xeric
conditions in the mountains. On the basis of leaf mor-
phology and stamen characters, the genus Potentilla is
charecterized in the systematically difficult genera of the
family Rosaceae (Szafer and Pawlowski 1955). Hooker
(1879) classified the genus Potentilla into three sections
while describing the Flora of British India. Because of
ambiguity at the species level, it has been divided into
many subgenera, sections, several subsections, series and
groups of species (Wolf 1908; B all et al. 1968). However,
in the regional Floras of India pertaining to the Western
Himalayas, no such taxonomic segregation is available
(Chaudhary and Wadhwa 1984; Dhaliwal and Sharma
1999; Kaur and Sharma 2004). Recently, molecular studies
have been carried out for analysis of the phylogeny, phy-
logeography, generic circumscription, cytoptype and
genetic diversification in different species of Potentilla
S. Rani � S. Kumar � S. M. Jeelani (&) �R. C. Gupta � S. Kumari
Department of Botany, Punjabi University,
Patiala 147 002, Punjab, India
e-mail: [email protected]
123
Plant Syst Evol (2012) 298:485–497
DOI 10.1007/s00606-011-0560-x

from different regions of the world (Eriksson et al. 1998;
Dobes and Paule 2010; Paule et al. 2011). These studies have
provided a phylogenetic framework for the classification of
different tribes of the subfamily Rosoideae, especially for the
tribe Potentilleae, and proposed placement of the Potentilla
species in different sequenes. Although sexuality is the means
of reproduction in the majority of species of the genus, some of
the species are also well known to reproduce apomictically
(Muntzing 1958; Asker and Frost 1970; Holm and Ghatnekar
1996a, b; Gregor et al. 2002). The documented proof of aga-
mospermy and hybridization accompanying the polyploidi-
zation of genomes has also been considered to have played a
significant role in the evolution of the genus (Gentscheff 1938;
Muntzing and Muntzing 1941; Gustafsson 1947a; Rutishauser
1948; Kashin 1999; Nylehn et al. 2003). Regarding cytolog-
ical studies on the genus from India, until now a total of
eleven species—P. atrosanguinea, P. fragarioides, P. fulgens,
P. indica, P. micropetala, P. nepalensis, P. sundaica
and P. supina from the Western Himalayas; P. fulgens and
P. mooniana from the Eastern Himalayas; and P. indica,
P. kleiniana and P. leschenaultiana from South India—
present only scanty cytological information for Indian
species. To fill the lacuna and to make an addition to
the chromosomal database of lesser known angiosper-
mic genera, the present meiotic studies on 14 species
of Potentilla L. were carried out on a population
basis from different geographical areas of the Western
Himalayas.
Materials and methods
For meiotic studies, flower buds were collected from selected
localities of the far distant areas of the Western Himalayas
(Table 1). Smears of appropriately sized flower buds were
made after fixing them in Carnoy’s fixative, using the standard
acetocarmine technique. Pollen fertility was estimated by
mounting mature pollen grains in a glycero-acetocarmine (1:1)
mixture. Well-filled pollen grains with stained nuclei were
taken as apparently fertile, while shrivelled and unstained
pollen grains were counted as sterile. Photomicrographs of
pollen mother cells and pollen grains were made from freshly
prepared slides using the Nikon 80i Eclipse Digital Imaging
System. Voucher specimens were deposited in the Herbarium,
Department of Botany, Punjabi University, Patiala (PUN).
Results and discussion
Cytological study
Detailed cytological studies were carried out on 45 popu-
lations belonging to 14 species of the genus Potentilla. The
data regarding locality with altitude, accessions and mei-
otic chromosome numbers along with figure numbers of the
presently worked out species are presented in Table 1. The
results for each species with new/varied chromosome
counts and abnormal meiotic courses are discussed below.
Potentilla argyrophylla Wall. ex Lehm.
Of the six populations of the species, two each from Kashmir
and Himachal Pradesh all depict n = 14, and the other two
populations again from Himachal Pradesh show n = 28. The
present counts make new records of the tetraploid cytotype
(n = 14) on a worldwide basis and octaploid (n = 28) cyto-
types on an Indian basis. The species was earlier known to have
cytotypes with higher ploidy levels such as 6x = 2n = 42
(Zhukova 1967), 8x = 2n = 56 (Shimotomai 1930; Popoff
1935; Gustafsson 1947b; Zhukova 1967), 9x = 2n = 63
(Gentscheff 1938), and 10x = 2n = 70, 11x = 2n = 77,
12x = 2n = 84, 13x = 2n = 91 and 14x = 2n = 98 (Pop-
off 1935) from different parts of the USSR.
P. atrosanguinea Raf.
Of the six populations of the species, one from Kashmir
and two from Himachal Pradesh show n = 7 and exist as
diploid cytotypes. Of the remaining three populations from
Himachal Pradesh, two depict n = 14, and only one pop-
ulation reveals n = 28. The present chromosome counts of
n = 7 and n = 14 add new diploid and tetraploid cyto-
types, respectively, for the species on a worldwide basis,
while the octaploid cytotype (n = 28) is in conformity with
the earlier reports from India and outside India. The species
was previously known to have 8x to 12x levels as 2n = 56
(Shimotomai 1930; Christoff and Papasova 1943; Gosw-
ami and Metfield 1978), 2n = 63 (Zhukova 1967) and
2n = 74, 84 (Goswami and Metfield 1978) from different
parts of the world.
P. desertorum Bunge
Of the two populations of species collected from Himachal
Pradesh, one shows n = 7, and the other reveals n = 14. The
diploid cytotype (n = 7) has been cytologically worked out
for the first time on a worldwide basis, while the tetraploid
cytotype (n = 14) is chromosomally known for the first time
from India and confirms the previous reports from Mongolia
and Tadzhikistan (Mesicek and Sojak 1969, 1993) and the
USSR (Guinochet and Lefranc 1981).
P. fruticosa Gray
The single accession of the species collected from Hima-
chal Pradesh depicts n = 14 (a tetraploid cytotype) and is
486 S. Rani et al.
123

Table 1 Data showing the location, altitude, accession number, present chromosome number, figure number, nature of the meiotic course,
ploidy level and pollen size in the presently worked out species of Potentilla L. from the Western Himalayas
Name of taxa Locality Altitude
(m)
Accession
no. (PUN)
Meiotic
chromosome
number (n) with
respective figure
Nature of
meiotic
course/ploidy
level (x)
Pollen size (lm)
P. argyrophylla Wall.
ex Lehm
Thajwas, Distt.
Ganderbal,
Kashmir
3,000 54268 14 (Fig. 1a) Normal/4x 23.95 9 20.89
Gulmarg, Distt.
Baramullah,
Kashmir
2,600 54910 14 Normal/4x 23.89 9 20.91
Chota-Bhangal,
Distt. Kangra,
H.P.
2,300 54860 14 Abnormal/4x 22.61 9 18.37–24.94 9 24.01a
Baragran, Distt.
Kangra, H.P.
3,000 54896 14 Abnormal/4x 22.45 9 18.26–25.05 9 23.92
Chur peaks, Distt.
Sirmaur, H.P.
3,200 54876 28 (Fig. 1b) Normal/8x 26.95 9 24.37
Bhangyanimata,
Distt. Sirmaur,
H.P.
2,800 54878 28 Abnormal/8x 24.35 9 19.76–27.15 9 24.79
P. atrosanguinea Raf. Aharbal, Distt.
Kulgam, Kashmir
2,400 54905 7 Normal/2x 22.71 9 20.21
Biching, Distt.
Kangra, H.P.
3,000 54889 7 Normal/2x 22.63 9 19.97
Bara-Bhangal,
Distt. Kangra,
H.P.
4,000 54880 7 (Fig. 1c) Abnormal/2x 21.14 9 18.58–23.16 9 20.69
Haripurdhar, Distt.
Sirmaur, H.P.
2,600 54907 14 (Fig. 1d) Abnormal/4x 22.23 9 19.65–25.29 9 23.78
Jamnala, Distt.
Sirmaur, H.P.
2,600 54867 14 Normal/4x 24.28 9 22.07
Tisri, Distt.
Sirmaur, H.P.
3,000 54868 28 (Fig. 1e) Abnormal/8x 24.63 9 21.75–26.98 9 24.07
P. desertorum Bunge Triund,
Distt. Kangra,H.P.
3,000 54895 7 (Fig. 1f) Abnormal/2x 20.30 9 18.01–23.93 9 21.14
Biching, Distt.
Kangra, H.P.
2,200 54301 14 (Fig. 1g) Normal/4x 24.21 9 19.60
P. fruticosa Gray Tisri, Distt.
Sirmaur, H.P.
3,000 52668 14 (Fig. 1h) Abnormal/4x 23.14 9 20.13–25.11 9 22.29
P. fulgens Wall. ex D.
Don (=P. lineataTrevir.)
Multan, Distt.
Kangra, H.P.
2,300 54884 7 (Fig. 1i) Abnormal/2x 21.70 9 18.05–23.93 9 19.53
Triund, Distt.
Kangra, H.P.
3,000 54885 14 (Fig. 1j) Normal/4x 26.77 9 21.89
Dal lake, Distt.
Kangra, H.P.
3,800 54893 7 Normal/2x 22.21 9 18.25
Nauradhar, Distt.
Sirmaur, H.P.
1,800 54873 7 Abnormal/2x 21.12 9 18.16–23.88 9 19.49
Shillai, Distt.
Sirmaur, H.P.
2,000 54874 7 Normal/2x 22.32 9 18.17
P. gelida C. A. Mey. Thajwas, Distt.
Ganderbal,
Kashmir
3,100 54227 7 (Fig. 1k) Normal/2x 22.77 9 19.36
Chumnai, Distt.
Anantnag,
Kashmir
3,500 54228 7 Abnormal/2x 21.05 9 18.32–23.67 9 19.36
Additions to the cytologically investigated species of Potentilla L. 487
123

Table 1 continued
Name of taxa Locality Altitude
(m)
Accession
no. (PUN)
Meiotic
chromosome
number (n) with
respective figure
Nature of
meiotic
course/ploidy
level (x)
Pollen size (lm)
P. gerardiana Lindl.
(=P. fragarioides L.)
Haripurdhar, Distt.
Sirmaur,H.P.
2,800 54907 14 (Fig. 1l) Normal/4x 24.59 9 22.38
P. indica (Andrews)
T. Wolf (=Duchesneaindica (Andrews)
Focke
Dara, Distt.
Srinagar, Kashmir
1,900 54200 7 Normal/2x 22.92 9 21.79
Dharmshala, Distt.
Kangra, H.P.
1,800 54199 7 (Fig. 1m) Abnormal/2x 23.10 9 19.76–25.46 9 22.02
Timbi, Distt.
Sirmaur, H.P.
1,800 54199 7 Normal/2x 22.79 9 21.68
Shillai, Distt.
Sirmaur, H.P.
2,000 54862 14 (Fig. 1n) Abnormal/4x 22.90 9 19.56–25.38 9 22.93
Nauradhar, Distt.
Sirmaur, H.P.
1,800 54861 14 Normal/2x 24.11 9 22.28
P. kleiniana Wight &
Arn.
Telangna, Distt.
Sirmaur,H.P.
2,800 55698 7 (Fig. 1o) Normal/2x 23.01 9 21.92
P. micropetala D. Don
(=Sibbaldiamicropetala (D. Don)
Hand.-Mazz.
Churdhar, Sirmaur
H.P.
3,600 54911 7 (Fig. 1p) Normal/2x 22.76 9 20.12
Churdhar, Distt.
Sirmaur, H.P.
3,600 54912 14 (Fig. 1q) Normal/4x 25.13 9 23.74
P. nepalensis Hook. Pehalgam, Distt.
Anantnag,
Kashmir
2,300 54901 21 (Fig. 1r) Abnormal/6x 22.96 9 19.45–24.88 9 21.93
Yusmarg, Distt.
Budgam, Kashmir
2,400 54902 14 Normal/2x 23.96 9 22.33
Triund, Distt.
Kangra, H.P.
2,300 54858 7 (Fig. 1s) Normal/2x 21.88 9 19.02
Lohardarhi, Distt.
Kangra, H.P.
2,500 54894 14 (Fig. 1t) Abnormal/4x 22.36 9 19.22–24.49 9 21.68
Haripurdhar, Distt.
Sirmaur, H.P
2,600 54864 7 Abnormal/2x 20.18 9 18.62–22.73 9 20.67
Shillai, Distt.
Sirmaur, H.P.
2,000 54865 14 Normal/4x 24.18 9 22.51
P. sibbaldia Kurtz.
(=Sibbaldia Hornem.)
Tisri, Distt.
Sirmaur, H.P.
3,000 54425 7 (Fig. 1u) Abnormal/2x 20.35 9 18.63–24.13 9 21.74
Churdhar, Distt.
Sirmaur, H.P.
3,600 55065 14 (Fig. 1v) Normal/4x 25.13 9 23.74
P. sundaica (Miq.)
W. Theob. (=Fragariasundaica Blume)
Karmul -Tral, Distt.
Pulwama,
Kashmir
2,300 54263 7 (Fig. 1w) Normal/2x 21.88 9 19.31
Triund, Distt.
Kangra, H.P.
3,000 52496 7 Normal/2x 22.08 9 19.41
Triund, Distt.
Kangra, H.P.
3,000 54883 7 Normal/2x 22.23 9 19.52
Dal lake, Distt.
Kangra, H.P.
2,800 52497 14 (Fig. 1x) Abnormal/4x 23.24 9 20.98–25.27 9 22.17
P. thomsonii Hand.-
Mazz.
Triund, Distt.
Kangra, H.P.
2,800 54886 7 (Fig. 1y) Normal/2x 22.18 9 19.10
Biching, Distt.
Kangra, H.P.
3,000 54887 7 Normal/2x 21.96 9 19.03
H.P. Himachal Pradesha In case of abnormal taxa the pollen size was calculated for both smaller and larger fertile pollen grains
488 S. Rani et al.
123

reported for the first time for India; it is in conformity with
the earlier report of 2n = 28 (Leht 1999) from Russia. The
diploid cytotypes (n = 7) are also known for the species
from Russia (Guinochet and Lefranc 1981; Leht 1991).
P. fulgens Wall. ex D. Don
Of the five populations of the species collected from
Himachal Pradesh, four exhibit diploid cytotypes (n = 7),
and only one population shows a tetraploid cytotype
(n = 14). The cytotype with n = 7 from India has been
cytologically worked out for the first time and is in con-
formity with the earlier report of 2n = 14 (Ikeda 1989)
from Japan. The tetraploid cytotype, n = 14, conforms to a
previous report from the Eastern Himalayas (Sharma and
Sarkar 1967–1968). The species is also known to have
another aneuploid cytotype with 2n = 34 from the Western
Himalayas (Mehra and Dhawan 1966) and octaploids
(2n = 56) from Russia (Popoff 1939).
P. gelida C. A. Mey.
Both populations of the species collected from Himachal
Pradesh depict n = 7 and exist as diploid cytotypes. The
Indian species has been chromosomally identified for the
first time, and n = 7 is in line with a previous report of
2n = 14 (Guinochet 1968) from France.
P. gerardiana Lindl.
The single accession of the species collected from Hima-
chal Pradesh exhibits n = 14. The tetraploid cytotype is
reported for the first time on a worldwide basis. Earlier the
species was reported to have diploids (2n = 14) from
different parts of the USSR and Japan (Krogulevich 1978;
Probatova N and Sokolovskaya 1981; Iwatsubo and Nar-
uhashi 1991) and octaploids (2n = 56) from the Western
Himalayas in India (Mehra and Dhawan 1966).
P. indica (Andrews) T. Wolf
Of the five populations of the species, one from Kashmir
and two from Himachal Pradesh exhibit diploid cytotypes
(n = 7). The other two populations from Himachal Pradesh
depict tetraploid cytotypes (n = 14). The present chro-
mosome count of n = 14 adds a new tetraploid cytotype
for the species on a worldwide basis, whereas n = 7 is in
conformity with the earlier reports from all over the world.
The species is already known to exhibit 2n = 14 (Subra-
manian 1979), 2n = 42 (Delay 1947), 2n = 70 (Kapil and
Modgil 1990) and 2n = 84 (Mehra and Dhawan 1966;
Naruhashi and Takano 1987; Hill 1989; Zhao et al. 1990;
Naruhashi and Iwatsuboo 1991; Naruhashi et al. 2005).
P. kleiniana Wight & Arn.
The single accession of the species collected from Hima-
chal Pradesh exhibits n = 7. The diploid cytotype is
reported for the first time from India and is in confirmity
with an earlier report of 2n = 14 (Shimotomai 1929) from
the USSR.
P. micropetala D. Don
Two populations of the species collected from Himachal
Pradesh show n = 7 and n = 14 cytotypes, respectively.
The present chromosome count of n = 14 adds a new
tetraploid cytotype for the species on a worldwide basis.
The diploid cytotype is reported for the first time from
India and is in conformity with the earlier single report of
2n = 14 (Mesicek and Sojak 1993) from China and Yun-
nan. The species is also known to have octaploids with
2n = 56 (Mehra and Dhawan 1966) from the northern
Western Himalayas in India.
P. nepalensis Hook.
Of the total six populations of the species, only one from
Kashmir shows n = 21 and exists as a hexaploid cytoytpe.
Among others, one population each from Kashmir and
Himachal Pradesh exhibit diploid cytotypes with n = 7.
The remaining two populations from Himachal Pradesh
reveal tetraploid cytotypes (n = 14). The present chro-
mosome count of n = 14 adds a new tetraploid cytotype to
the species on a worldwide basis. The diploid cytotypes
conform to a single previous chromosomal report of
2n = 14 (Popoff 1939) from Russia. The hexaploid cyto-
type is in conformity with the earlier reports of 2n = 42
(Mehra and Dhawan 1966) from the Western Himalayas in
India and outside India from Russia and England (Christoff
and Papasova 1943; Zhukova 1967; Goswami and Metfield
1975).
P. sibbaldia Kurtz.
The two populations of the species collected from Hima-
chal Pradesh show n = 7 and n = 14, respectively. The
present diploid cytotype is chromosomally reported for the
first time from India, whereas the tetraploid cytotype is
new on a worldwide basis. The species was previously
known to have a single diploid report of 2n = 14 (Guinochet
and Lefranc 1981) from Russia.
P. sundaica (Miq.) W. Theob.
Of the four populations of the species, one from Kashmir
and two from Himachal Pradesh show diploid cytotypes
Additions to the cytologically investigated species of Potentilla L. 489
123

(n = 7). Only one population from Himachal Pradesh
reveals a tetraploid cytotype (n = 14). The present chro-
mosome count of n = 7 is reported for the first time from
India and is in line with the previous reports of 2n = 14
(Shimotomai 1929) from different parts of Europe, whereas
2n = 28 is in conformity with the earlier reports from the
Western Himalayas in India (Mehra and Dhawan 1966) and
outside India (Ikeda 1989).
P. thomsonii Hand.-Mazz.
Both populations of the species collected from Himachal
Pradesh show n = 7 and exist as diploids based on x = 7.
The species has been cytologically worked out for the first
time on a worldwide basis.
Chromosome numbers and polyploidy
A perusal of the cumulative cytological literature brings to
light that 168 species/292 cytotypes of the genus Potentilla,
including 17 species (after the present study) from India,
have been worked out. The chromosome numbers in the
genus vary considerably from 2n = 14 to 112, with
2n = 28 (22.26%) being the most common followed by
2n = 14 (20.20%), 2n = 42 (15.06%), 2n = 56 (9.93%),
2n = 35 (5.47%), 2n = 49 (2.73%), 2n = 70 (2.73%) and
2n = 21 (1.71%), while 2n = 20, 34, 38, 39, 48, 62, 74,
77, 84, 91, 98 and 112 are rarely encountered in certain
species on a worldwide basis (c.f., Fedorov 1974; Kumar
and Subramaniam 1986; Index to Plant Chromosome
Numbers, Web, etc.). However, from India among the
different chromosome counts of 2n = 28, 34, 42 and 56,
the cytotype with 2n = 28 remains the most common and
is in line with the world data. As seen at present and as
previously reported by Delgado et al. (2000), Tomasz and
Kolodziejek (2008) and Lundberg (2011), irrespective of a
wide range of ploidy levels, x = 7 remains the single base
number for the genus both for India and worldwide.
Otherwise also the ploidy based on x = 7 in the different
species of subfamily Rosoideae is relatively high, and the
same is exhibited in the genus Potentilla (Johri et al. 1992),
where it is associated with gametophytic apomixis (Dick-
inson et al. 2007). According to Asker and Jerling (1992),
and Nylehn et al. (2003), Potentilla is one such genus in
which apomixis, sexuality, polyploidy and taxonomic
complexity are all combined. In the presently investigated
taxa also the same polyploidy is seen in nearly half of the
taxa, with the occurrence of tetraploids being the highest
(Table 2). Earlier, Stebbins (1950), and Mesicek and Sojak
(1993) categorized two groups of species within the genus,
with one group having species with stable chromosome
numbers, i.e., a single chromosomal race all over their
distribution and the other group of species with complex
ploidy whose chromosome numbers vary even within the
same species. The same holds true for the Western
Himalayas and India as well (Table 2). Analysis of the
worldwide data reveals that 123 species form the pre-
dominant group belonging to the stable chromosome
number category, and only 45 species are in the complex
ploidy level category, this too with some aneuploid cyto-
types as well. However, for Indian species, on the contrary,
a single chromosomal race either at 2x, 4x or 8x occurs less
frequently than those showing complex polyploidy levels
(Table 2). Comparing the present data with that worldwide,
it can be concluded that higher ploidy levels occur rela-
tively less often in Indian species.
In the presently studied species of Potentilla, the role of
hybridization followed by doubling of the chromosome
numbers resulting in allopolyploidy seems to be a major
mode of speciation in the genus Potentilla, as has been
described previously in many angiosperms (Stebbins 1950;
Grant 1981; Masterson 1994; Solits and Solits 2009).
Table 2 Ploidy levels in presently and previously cytologically worked out species of Potentilla L. from India
Species with single ploidy level Species with complex ploidy levels
Ploidy 2x 4x 8x 2x, 4x 2x, 4x, 5x 2x, 4x, 6x 4x, 8x 2x, 4x, 8x
Ch. no. (n) 2n = 14 2n = 28 2n = 56 2n = 14, 2n = 14, 2n = 28, 2n = 14, 2n = 28, 2n = 14
2n = 18 2n = 34 2n = 28, 2n = 56 2n = 28,
2n = 42 2n = 56
Species P. gelida P. fruticosa P. supina P. desertorum P. fulgens P. nepalensis P. argyrophylla P. atrosanguinea
P. kleiniana P. leschenaultiana P. indica P. gerardiana
P. thomsonii P. mooniana P. micropetala
P. sibbaldia
P. sundaica
Frequency 17.64 17.64 5.88 29.41 5.88 5.88 11.76 5.88
(%) 41.16 58.81
490 S. Rani et al.
123

According to recent molecular studies, the role of apomixis
has been considered to be an important phenomenon for the
evolutionary diversification of apomictic plants (Paule
et al. 2011). Based on the reproductive aspect of P. ar-
gentea, with different cytotypes of the species distributed
in Europe, it has been clearly indicated that a correlation
exists in the form in which sexuality and autogamy are
means of multiplication for diploids, whereas apomixis is
generally associated with polyploids (Holm 1995; Holm and
Ghatnekar 1996a, b; Holm et al. 1997; Paule et al. 2011).
The Indian materials have polyploids at various levels
of ploidy in the form of both inter- and intra-specific
Fig. 1 a Potentillaargyrophylla (n = 14), PMC at
Diakinesis. b P. argyrophylla(n = 28), PMC at Diakinesis.
c P. atrosanguinea (n = 7),
PMC at M-I. d P. atrosanguinea(n = 14), PMC at Diakinesis.
e P. atrosanguinea (n = 28),
PMC at Diakinesis. f P.desertorum (n = 7), PMC at
M-I. g P. desertorum (n = 14),
PMC at A-I. h P. fruticosa(n = 14), PMC at M-I. i P.fulgens (n = 7), PMC at M-I.
j P. fulgens (n = 14), PMC at
Diakinesis. k P. gelida (n = 7),
PMC at M-I. l P. gerardiana(n = 14), PMC at Diakinesis.
m P.indica (n = 7), PMC at
M-I. n P.indica (n = 14), PMC
at M-I. o P. kleiniana (n = 7),
PMC at M-I. p P. micropetala(n = 7) PMC at A-I.
q P.micropetala (n = 14) PMC
at A-I. r P. nepalensis (n = 21),
PMC at Diakinesis.
s P.nepalensis (n = 7) PMC at
A-I. t P.nepalensis (n = 14)
PMC at Diakinesis. u P.sibbaldia (n = 7) PMC at M-I.
v P. sibbaldia (n = 14) PMC at
A-I. w P.sundaica (n = 7) PMC
at M-I. x P. sundaica (n = 14)
PMC at Diakinesis. y P.thomsonii (n = 7) PMC at A-I.
Scale 10 lm
Additions to the cytologically investigated species of Potentilla L. 491
123
variability. During the present study, intercytotypes for
different species were found to be sympatric in distribution
and seem to be the result of both systems, i.e., sexuality and
apomictic mode operative selectively in different cyto-
types. This is in line with previous postulations that such
mixing up of the phenomena may be due to the repro-
ductive mode on the chosen donor pollen (Muntzing and
Muntzing 1945), methodological pitfalls of the applied
crossing methods including unintended self-pollination
(Schwendener 1969), mentor effects (Horandl and Temsch
2009) or dominance of maternal markers (Nosrati et al.
2010). From the present study, it is observed that although
the number of species worked out for the chromosome
numbers is very low from the Western Himalayas, the
amount of inter- and intraspecific chromosomal number
diversity is quite high, with euploid series marked in some
of the species from the region, and a few species studied
from other parts of India support the earlier hypothesis of
Panigrahi and Dikshit (1987) of India as the principal
center of origin of Potentilla.
Further, there appears to be a significant size difference
in meiotic chromosomes in different cytotypes in some
species such as P. argyrophylla (n = 28), P. atrosanguinea
(n = 28), P. desertorum (n = 7), P. fulgens (n = 7) and
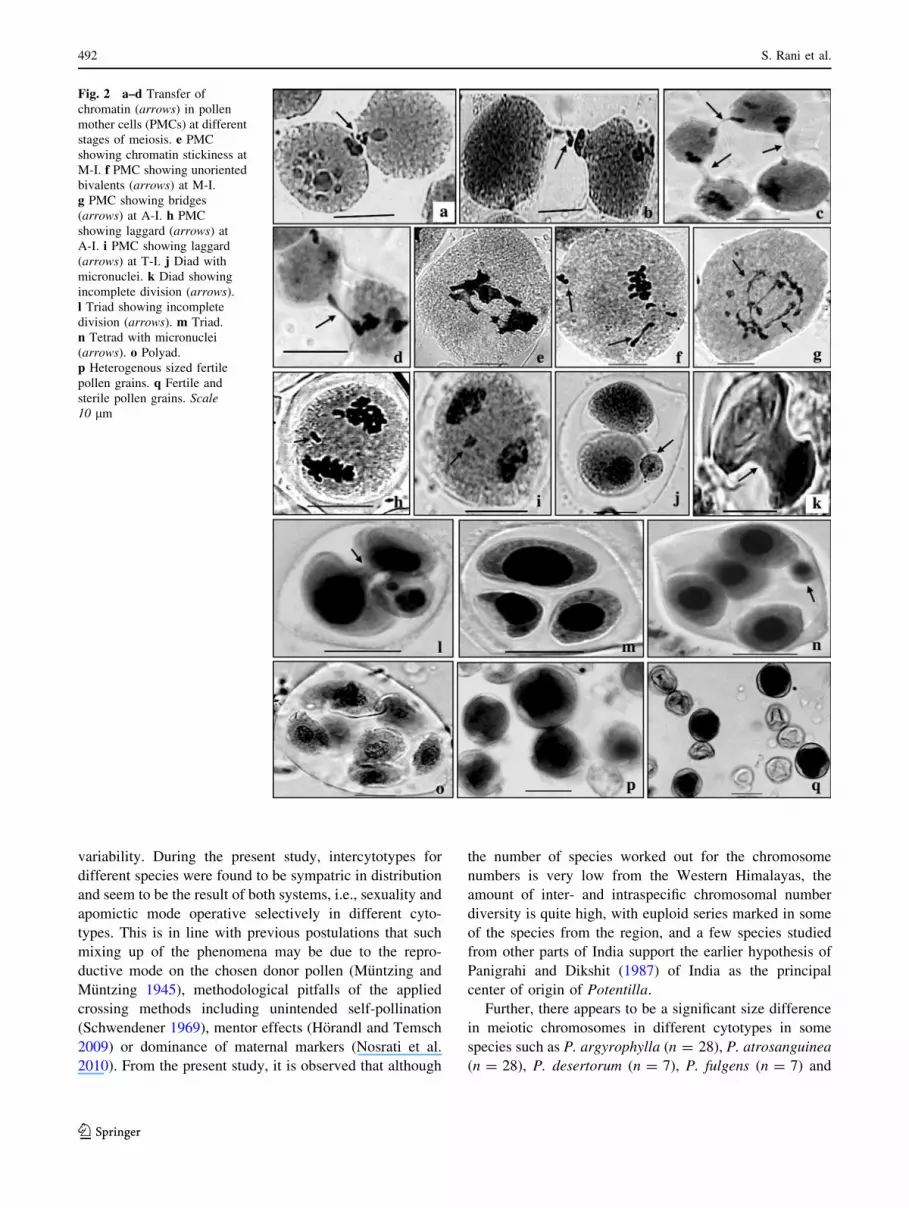
Fig. 2 a–d Transfer of
chromatin (arrows) in pollen
mother cells (PMCs) at different
stages of meiosis. e PMC
showing chromatin stickiness at
M-I. f PMC showing unoriented
bivalents (arrows) at M-I.
g PMC showing bridges
(arrows) at A-I. h PMC
showing laggard (arrows) at
A-I. i PMC showing laggard
(arrows) at T-I. j Diad with
micronuclei. k Diad showing
incomplete division (arrows).
l Triad showing incomplete
division (arrows). m Triad.
n Tetrad with micronuclei
(arrows). o Polyad.
p Heterogenous sized fertile
pollen grains. q Fertile and
sterile pollen grains. Scale10 lm
492 S. Rani et al.
123

P.indica (n = 7), and also at an interspecific level in P.
gelida, whereas for the rest of the intraspecific cytotypes
and at an interspecific level, the chromosomes appear to
be of similar size. Such observations have been seen
earlier in certain species of Potentilla (Delgado et al.
2000).
Table 3 Data on abnormal meiotic course and pollen fertility in different species of Potentilla L. from the Western Himalayas
Taxa/
accessions
Cytomixis Meiotic course showing PMCs with Pollen
fertility
(%)Percentage of PMCs
involved meiosis-I/
meiosis-II
Number of
PMCs
involved
Chromosomal
stickness at M-I
(%)
Unoriented
bivalents at M-I
(%)
Bridges at
meiosis-I/
meiosis-II (%)
Laggards at
meiosis-I/
meiosis-II (%)
P. argyrophylla
54860 23.33 (28/120)/15.38
(20/130)
2–5 8.33 (10/120) 7.69 (10/130) 13.33 (18/135)/
7.69 (10/130)
7.27 (8/110)/5.21
(6/115)
59.79
54896 19.25 (26/135)/15.38
(20/130)
2–3 5.21 (6/115) 5.51 (7/127) 10.71 (15/140)/
6.15 (8/130)
4.44 (6/135)/5.08
(6/118)
60.62
54878 4.95 (6/121)/– 2–3 3.63 (4/110) 4.23 (5/118) 5.69 (7/123)/6.34
(8/126)
7.25 (9/124)/7.87
(10/127)
60.74
P. atrosanguinea
54880 3.63 (4/110)/– 2–3 – – 2.00 (2/100)/2.72
(3/110)
5.71 (6/105)/3.88
(4/103)
55.45
548907 11.66 (14/120)/5.21 (6/
115)
2–4 2.06 (2/97) – 6.95 (8/115)/– 6.03 (7/116)/– 52.47
54868 9.23 (12/130)/5.18 (7/
135)
2–3 3.47 (4/115) 3.44 (4/116) 7.50 (9/120)/6.40
(8/125)
4.23 (5/118)/– 63.90
P. desertorum
54895 11.66 (14/120)/5.21 (6/
115)
2–4 2.06 (2/97) – 1.81 (2/110)/
2.91(3/103)
5.71 (6/105)/3.88
(4/103)
59.98
P. fruticosa
52668 6.49 (10/154)/6.40 (8/
125)
2–3 – 3.34 (4/120) 3.34 (4/120)/– –/5.60 (7/125) 64.87
P. fulgens
54884 –/– – – –- –/– 3.47 (4/115)/
1.81(2/110)
65.00
54873 –/– – – 1.98 (2/101) 2.85 (3/105)/5.83
(7/120)
3.20 (4/125)/2.30
(3/130)
63.48
P. gelida
54228 11.53 (15/130)/– 2–4 – – –/3.70 (5/135) 2.30 (3/130)/4.13
(6/145)
60.70
P. indica
54199 3.20 (4/125)/– 2–3 1.53 (2/130) 0.90 (1/110) –/– –/– 61.62
54862 4.61 (6/130)/3.57 (4/
112)
2–3 – 2.50 (3/120) 3.17 (4/126)/– 3.47 (4/115)/1.66
(2/120)
62.84
P. nepalensis
54901 –/4.80 (6/125) 2–3 4.28 (6/140) – 8.69 (10/115)/
2.40 (3/125)
2.54 (3/118)/– 63.05
54894 –/– 2–3 3.70 (5/135) 4.34 (6/138) 6.15 (8/130)/3.07
(4/130)
–/– 62.45
54864 5.92 (8/135)/4.28 (6/
140)
2–3 – – 5.18 (7/135)/3.67
(5/136)
5.00 (6/120)/3.20
(4/125)
64.07
P. sibbaldia
54425 –/– – – – 5.45(6/110)/
3.57(4/112)
5.08 (6/118)/3.44
(4/116)
61.60
P. sundaica
52497 16.00 (20/125)/8.69 (10/
115)
2–4 3.63 (4/110) 4.95 (5/101) 6.25 (7/112)/3.63
(4/110)
3.36 (4/119)/– 61.58
Figures in parentheses denote the observed number of abnormal PMCs in the numerator and total number of PMCs observed in the denominator
Additions to the cytologically investigated species of Potentilla L. 493
123

Pollen size
The pollen grain size of the presently investigated taxa as given
in Table 1 shows that the polyploids have in general larger
pollen grains than diploids. Furthermore, among polyploids
there is relatively less difference between tetraploid and hexa-
ploid cytotypes, whereas there is relatively more size difference
when tetraploids are compared to octaploids. Such comparable
results have been previously reported for many angiosperms
(Stebbins 1956; Dijkstra and Speckmann 1980; Strivastava and
Strivastava 2002; Zlesak 2009; Omidbaigi et al. 2010).
Meiotic abnormalities
Meiotic abnormalities have been recorded in some popu-
lations of Potentilla argyrophylla, P. atrosanguinea,
P. desertorum, P. fruticosa, P. fulgens, P. gelida, P. indica,
P. nepalensis, P. sibbaldia and P. sundaica. In such pop-
ulations, abnormalities in the form of cytomixis, chromatin
stickiness, unoriented bivalents, bridges and laggards have
been observed at different stages of meiosis (Fig. 2a–i;
Table 3). These meiotic anomalies indicate the existence of
intraspecific genetic diversities. Such genetic differences
have been seen previously in different plant species
(Baptista-Giacomelli et al. 2000; Sheidai et al. 2003). The
phenomenon of chromatin transfer from early prophase to
the pollen formation stage (Fig. 2a–d) has been observed in
most of these populations with the highest percentage
recorded in populations of P. argyrophylla (Table 3).
Cytomixis and chromatin stickiness are considered to be
the result of genetic factors (Bellucci et al. 2003; Ghaffari
2006; Fadaei et al. 2010) and environmental factors
Table 4 Data on abnormal microsporogenesis in different species of Potentilla L. from the Western Himalayas
Taxa/accessions Microsporogenesis
Monads Diads Triads Tetrads
WMN (%) WM (%) WMN (%) WM (%) WMN (%) WM (%) WMN (%) WM (%)
P. argyrophylla
54860 2.19 (2/91) 1.09 (1/91) 3.29 (3/91) – 6.59 (6/91) 4.39 (4/91) 60.43 (55/91) 21.97 (20/91)
54896 2.91 (3/103) 1.94 (2/103) 1.94 (2/103) 0.97 (1/103) 4.85 (5/103) 3.88 (4/103) 63.10 (65/103) 20.38 (21/103)
54878 1.90 (2/105) 2.85 (3/105) 1.90 (2/105) 1.90 (2/105) 2.85 (3/105) 0.95 (1/105) 68.57 (72/105) 19.04 (20/105)
P. atrosanguinea
54880 1.92 (2/104) – 1.92 (2/104) – 3.84 (4/104) 1.92 (2/104) 65.38 (68/104) 25.00 (26/104)
54907 0.90 (1/110) 0.90 (1/110) 0.90 (1/110) 0.90 (1/110) 2.72 (3/110) 3.63 (4/110) 65.45 (72/110) 24.54 (27/110)
54868 – – 0.91 (1/109) – 1.83 (2/109) 0.91 (1/109) 71.55 (78/109) 24.77 (27/109)
P. desertorum
54895 – – – – 0.98 (1/102) 0.98 (1/102) 78.43 (80/102) 19.60 (20/102)
P. fruticosa
52668 1.83 (2/110) 0.91 (1/109) 1.83 (2/109) – 5.50 (6/109) 6.42 (7/109) 64.22 (70/109) 19.26 (21/109)
P. fulgens
54884 1.94 (2/103) 0.97 (1/103) 1.94 (2/103) – 1.94 (2/103 – 73.78 (76/103) 19.41 (20/103)
54873 0.97 (1/103) – 0.97 (1/103) – 1.94 (2/103) – 73.78 (76/103) 19.41 (20/103)
P. gelida
54228 – 1.00 (1/100) – 1.00 (1/100) 1.00 (1/100) 1.00 (1/100) 80.00 (80/100) 16.00 (16/100)
P. indica
54199 0.98 (1/102) 0.98 (1/102) – 0.98 (1/102) 1.96 (2/102) 1.96 (1/102) 76.47 (78/102) 17.64 (18/102)
54862 0.90 (1/111) 0.90 (1/111) 0.90 (1/111) 0.90 (1/111) 3.60 (4/111) 1.80 (2/111) 72.97 (81/111) 18.91 (21/111)
P. nepalensis
54901 – 0.93 (1/107) 0.93 (1/107) – 2.80 (3/107) 0.98 (1/107) 73.83 (79/107) 20.56 (22/107)
54894 0.88 (1/113) – 0.88 (1/113) 0.88 (1/113) – 3.53 (4/113) 73.45 (83/113) 21.23 (24/113)
54864 – – 0.86 (1/115) – 5/115 (4.34) 1.73 (2/115) 73.04 (84/115) 20.00 (23/115)
P. sibbaldia
54425 0.88 (1/113) – 1.76 (2/113) – 3.53 (4/113) 1.76 (2/113) 74.33 (84/113) 17.69 (20/113)
P. sundaica
52497 0.97 (1/103) – 1.76 (2/103) – 1.94 (1/103) 0.97 (1/103) 75.72 (78/103) 20.38 (21/103)
Figures in parentheses denote the observed number of abnormal PMCs in the numerator and total number of PMCs observed in the denominator
WMN without micronuclei; WM with micronuclei
494 S. Rani et al.
123

(Nirmala and Rao 1996) as well as genomic-environmental
interaction (Baptista-Giacomelli et al. 2000), and seems to
be equally applicable to the presently investigated popu-
lations. Cytomixis in these populations leads to the for-
mation of hypo- and hyperploids PMCs (Fig. 2a) and even
empty PMCs in some cases (Fig. 2b). These meiotic
abnormalities lead to the induction of abnormal polarity of
spindles and formation of nuclei of variable numbers and
sizes during microsporogenesis in the form of monads,
diads, triads and polyads along with micronuclei (Fig. 2j–
o; Table 4). In some of the PMCs, diads/triads show
incomplete division (Fig. 2k, l). As a result of these meiotic
abnormalities, large and small-sized pollen grains are
formed along with reduced pollen fertility (52–65%) in
these meiotically abnormal taxa (Fig. 2p–q; Table 3). The
large-sized pollen grains were seen in the range of 5–7% in
each such population. The occurrence of giant pollen
grains, possibly the unreduced 2n pollen grains, has pre-
viously been reported in several species (Vorsa and Bing-
ham1979; Bertagnolle and Thomson 1995; Sheidai et al.
2008; Fadaei et al. 2010). The unreduced gametes are
known to produce higher polyploidy levels through poly-
ploidization (Villeux 1985). According to Bertagnolle and
Thomson (1995) different cytological mechanisms are
responsible for production of 2n gametes. However, the
pollen fertility in the normal populations has been seen to
be quite high (95–99%).
Acknowledgments The authors are grateful to the University
Grants Commission, New Delhi, under the DRS SAP III of UGC,
FIST of the DST programs and the Rajiv Gandhi National Fellowship
Scheme for providing financial assistance. We are very thankful to the
Director of Botanical Surveys of India, DehraDun, for his help in
identification of plant species.
References
Asker S, Frost S (1970) The ‘‘Potentilla collina problem’’ a
chemotaxonomic approach. Hereditas 66:49–70
Asker SE, Jerling L (1992) Apomixis in plants. CRC Press, Boca
Raton
Ball PW, Pawlowski B, Walters SM (1968) Potentilla L. In: Tutin TG
et al (eds) Flora Europea, 2nd edn. Cambridge University Press,
Cambridge, pp 36–47
Baptista-Giacomelli FR, Pagliarini MS, Almeida JL (2000) Meiotic
behavior in several Brazilian oat cultivars (Avena sativa L.).
Cytol 65:371–378
Bellucci M, Roscini C, Mariani A (2003) Cytomixis in pollen mother
cells of Medicago sativa L. J Hered 94:512–516
Bertagnolle F, Thomson JD (1995) Gametes with the somatic
chromosome number: mechanisms of their formation and role
in the evolution of autopolyploid plants. New Phytol 129:1–22
Chaudhary HJ, Wadhwa BM (1984) Flora of Himachal Pradesh
analysis, 1st edn. Botanical Survey of India, Howrah
Christoff M, Papasova G (1943) Die genetischen Grundlagen der
apomicktischen Fortpflanzung in der Gattung Potentilla. Z. Induct.
Abstamm. Vererb-lehre., 8I, I
Delay C (1947) Recherches Sur la structure des noyaux quiescent
chez les Phanerogames. Rev Cytol Cytophysiol Veg 9:169–222;
10:103–229
Delgado L, Gallego F, Rico E (2000) Karyosystematic study of
Potentilla L. subgen. Potentilla (Rosaceae) in the Iberian
Peninsula. Bot J Linn Soc 132:263–280
Dhaliwal DS, Sharma M (1999) Flora of Kullu District (Himachal
Pradesh). Bishen Singh Mahendra Pal Singh, DehraDun
Dickinson TA, Lo E, Talent N (2007) Ploidy, reproductive biology,
and Rosaceae: understanding evolution and making classifica-
tions. Plant Syst Evol 266:59–78
Dijkstra H, Speckmann GI (1980) Autotetrapliody in Caraway
(Carum carvi L.) for the increase of the aetheric oil content of
the seed. Euphytica 29:89–96
Dobes C, Paule J (2010) A comprehensive chloroplast DNA-based
phylogeny of the genus Potentilla (Rosaceae): implications for
its geographic origin, phylogeography and generic circumscrip-
tion. Mol Phylogenet Evol 56:156–175
Eriksson T, Donoghue MJ, Hibbs MS (1998) Phylogenetic analysis of
Potentilla using DNA sequences of nuclear ribosomal internal
transcribed spacers (ITS), and implications for the classification
of Rosoideae (Rosaceae). Plant Syst Evol 211:155–179
Fadaei F, Sheidai M, Asadi M (2010) Cytological study of the genus
Arenaria L. (Caryophyllaceae). Caryol 63(2):149–156
Fedorov ANA (1974) Chromosome number of flowering plants.
Academy of Science of the USSR Komarov Botancial Institute,
Leningard
Gentscheff G (1938) Uber die pseudogame fortpflanzug bei Poten-tilla. Genetica 20:398–408
Ghaffari SM (2006) Occurrence of diploid and polyploid microspores
in Sorghum bicolor (Poaceae) is the result of cytomixis. Afr J
Biotech 5:1450–1453
Goswami DA, Metfield B (1975) Cytogenetic studies in genus
Potentilla L. New Phytol 75:135–146
Goswami DA, Metfield B (1978) Meiotic studies on two Polyploid
Potentillas, P. ambigens Greene and P. atrosanguinea Lodd.
Caryol 31:109–115
Grant V (1981) Plant Speciation 2nd ed. Columbia University Press,
New York, pp 293–297, 324–328
Gregor T, Rolik J, Weising K (2002) RAPD-Untersuchungen und
Chromosomenzahlungen in der Potentilla-collina-Gruppe (Ros-
aceae). Berichte der Bayerischen Gesellschaft zur Er-forschung
der Heimischen Flora 72:159–167
Guinochet M (1968) Nouvelle contribution al’etude cytotaxinomique
des Potentilla. De la section Aureae. Th Wolf CR. Acad Sci
(Paris) 239:1416–1417
Guinochet M, Lefranc M (1981) In Chromosome number reports
LXXIII. Taxon 30:853–854
Gustafsson A (1947a) Apomixis in higher plants. Part III. Biotype andspecies formation. Lunds Univ Arsskr 243:181–320
Gustafsson A (1947) Apomixis in higher plants. Part-II the causal
aspect of apomixis. Acta Univ Lund (Lunds Univ Arsskr) Avd 2,
43(2):71–178
Hill LM (1989) IOPB chromosome data 1. International organization
of plant biosystematists. Newsletter 13:17–19
Holm S (1995) Unexpectedly high levels of genetic variation in
Potentilla argentea (s.1.). in southern Sweden. Hereditas
123:127–139
Holm S, Ghatnekar L (1996a) Apomixis and sexuality in hexaploid
Potentilla argentea. Hereditas 125:53–60
Holm S, Ghatnekar L (1996b) Sexuality and no apomixis found in
crossing experiments with diploid Potentilla argentea. Hereditas
125:77–82
Holm S, Ghatnekar L, Bengtsson BO (1997) Selfing and outcrossing
but no apomixis in two natural populations of diploid Potentillaargentea. J Evol Biol 10:343–352
Additions to the cytologically investigated species of Potentilla L. 495
123

Hooker JD (1879) Flora of British India, vol II. L. Reeve and Co Ltd.,
Ashford
Horandl E, Temsch EM (2009) Introgression of apomixis into sexual
species is inhibited by mentor effects and ploidy barriers in the
Ranunculus auricomus complex. Ann Bot 104:81–89
Ikeda H (1989) Chromosome numbers of Himalayan Potentilla(Rosaceae). J Jap Bot 64:361–367
Iwatsubo Y, Naruhashi N (1991) Karyomorphological and cytoge-
netical studies in Potentilla (Rosaceae) I. Karyotypes of nine
Japanese species. Cytologia 56:1–10
Johri BM, Ambegaokar KB, Srivastava PS (1992) Comparative
embryology of Angiosperms. Springer-Verlag, Berlin
Kapil M, Modgil D (1990) A new report of pentaploid Fragariaindica Andr. from western Himalaya. Cell Chromosom Res
13:24–26
Kashin AS (1999) The structure of agamocomplexes and the problem
of saltatory speciation in angiosperms. Bot Zurn 84:15–29
Kaur H, Sharma M (2004) Flora of Sirmaur. Bishen Singh Mahendra
Pal Singh, Dehra Dun
Krogulevich RE (1978) Kariologicheskij analiz vidov flory Vostoch-
nogo Sajana. V Flora Pribajkal’ja. Nauka, Novosibirsk, pp 19–48
Kumar V, Subramaniam B (1986) Chromosome atlas of flowering
plants of the Indian subcontinent, vol 1 edn. Dicotyledones BSI,
Calcutta
Leht M (1991) Micromorphological characters in the systematic of
Baltic cinquefoils (Potentilla). Botaanilised uurinused. Scripta
Bot Belg 6:150–164
Leht M (1999) Origin, chromosome number and reproduction biology
of Potentilla fruticosa (Rosaceae) in Estonia and Latvia. Ann
Bot Fenn 162:191–196
Lundberg M (2011). Systematics and polyploid evolution in Poten-tilleae (Rosaceae). University dissertation from Stockholm:
Department of Botany, Stockholm University, Sweden
Masterson J (1994) Stomatal size in fossil plants: evidence for
polyploidy in majority of angiosperms. Science 264:421–423
Mehra PN, Dhawan H (1966) Cytological investigation in the N.
W. Himalayan Rosaceae. Proc 53rd Indian Sci Congr 3:276–277
Mesicek J, Sojak J (1969) Chromosome counts of some Mongolian
plants. Folia Gebot Phytotaxon 4:55–86
Mesicek J, Sojak J (1993) Annotated chromosome numbers of
selected Asiatic Potentilla species. Folia Geobot Phytotaxon
28(4):437–446
Muntzing A (1958) Heteroploidy and polymor-phism in some
apomictic species of Potentilla. Hereditas 44:280–329
Muntzing A, Muntzing G (1941) Some new results concerning
apomixis, sexuality and polymorphism in Potentilla. Bot Notiser
94:237–278
Muntzing A, Muntzing G (1945) The mode of reproduction of hybrids
between sexual and apomictic Potentilla argentea. Bot Notiser
98:49–71
Naruhashi N, Iwatsuboo Y (1991) Karyological studies of Duchesnea
(Rosaceae). Cytol 56(3):143–149
Naruhashi N, Takano H (1987) Chromosome numbers and distribu-
tion of Duchesnea (Rosaceae) in Gifu prefecture. Acta Phyto
Geobot 38(8):155–160
Naruhashi N, Nishikawa T, Iwatsubo Y (2005) Taxonomic relation-
ship between Japanese Potentilla anemonefolia and Himalayan
P. sundaica (Rosaceae). J Phytogeogr Taxon 53:1–11
Nirmala A, Rao PN (1996) Genetics of chromosome numerical
mosaism in higher plants. Nucleus 39:151–175
Nosrati H, Price A, Wilcock CC (2010) No evidence of apomixis in
matroclinal progeny from experimental crosses in the genus
Fragaria (strawberry) based on RAPDs. Euphytica 171:193–202
Nylehn J, Hamre E, Nordal I (2003) Facultative apomixis and
hybridization in arctic Potentilla section Niveae (Rosaceae) from
Svalbard. Bot J Linn Soc 142:373–381
Omidbaigi R, Mirzaee M, Hassani ME, Sedghi Moghadam M (2010)
Induction and identification of polyploidy in basil (Ocimumbasilicum L.) medicinal plant by colchicine treatment. Int J Plant
Prod 4(2):87–98
Panigrahi G, Dikshit BK (1987) Systematics of the genus Potentilla L.
(Rosaceae Juss.)—its infrageneric classification and evolutionary
trends. Bull Bot Surv India 27:177–196
Paule J, Sharbal TF, Dobes C (2011) Apomoctic and sexual lineages
of the Potentilla argentea L. group (Rosaceae): cytotype and
molecular genetic differentiation. Taxon 60(3):721–732
Popoff A (1935) Uber die Fortpflanzungsverhaltnisse der Gattung
Potentilla. Planta Berl 24:510
Popoff A (1939) Karyologische Studien iiber die Entwicklungsvorg-
ange in der Gattung Potentilla 9(3):3–58
Probatova NS, Sokolovskaya AP (1981) Kariologicheskoe
issledovanie sosudistykh rastenij ostrovov Dal’nevostocnogo
gosudarstvennogo morskogo sapovednika. Sb. Cvetkaye Ras-
tenija Ostrovov Dalnevostochnogo Morskogo Sapovednika
pp 92–114
Rutishauser A (1948) Pseudogamie und Polymorphie in der Gattung
Potentilla. Arch Julius Klaus-Stiftung Vererbungsf 23(3/4):
267–424
Santapau H, Henry AN (1973) A dictionary of the flowering plants in
India. C.S.I.R, New Delhi
Schwendener J (1969) Experimente zur Evolution von Potentillaprocumbens Sibth. Ber Schweiz Bot Ges 79:49–92
Sharma A, Sarkar AK (eds) (1967–1968) Chromosome number
reports of plants. In: Annual Report, Cytogenet Lab Dept Bot
Univ Calcutta Res Bull 2:38–48
Sheidai M, Koobaz P, Zehzad B (2003) Meiotic studies of some
Avena L. species and populations. Iran J Sci 14:121–131
Sheidai M, Nikoo M, Gholipour A (2008) Cytogenetic variability and
new chromosome number reports in Silene L. species (Sect.
Lasiostemones, Caryophyllaceae). Acta Biol Szedg 52:313–319
Shimotomai N (1929) Uber die Chromosomenzhalen bei einigen
Potentillen. Sci Rep Tohokn Univ Ser 4(4):369–372
Shimotomai N (1930) Chromosomenzahlen u Phylogene bei der
Gattung Potentilla. Y Sci Hirosh Univ, Botany, I, 1
Solits PS, Solits DE (2009) The role of hybridization in plant
speciation. Annu Rev Plant Biol 60:561–588
Stebbins GL (1950) Variation and evolution in plants. New York
Columbia University Press, New York
Stebbins GL (1956) Cytogenetics and evolution of the grass family.
Am J Bot 43:890–905
Strivastava R, Strivastava GK (2002) Autopolyploids of Helianthusannuus L. Var. morden. Cytol 67:213–220
Subramanian D (1979) Cytological studies in South Indian species of
Rosaceae. Soc Adv Bot Annu Conf, Dep Bot Punjab Agric,
Univ, Ludhiana. Abstr 123–124
Szafer W, Pawlowski B (1955) Potentilla L. In: Szafer W, Pawlowski
B (eds) Flora of Poland vol 7. Panstwowe Wydawnictwo
Naukowe, Warszawa, pp 96–43
Tomasz I, Kolodziejek J (2008) Chromosome numbers of Potentillasubsect. Collinae (Rosaceae) from Poland. Caryologia 61(2):
170–175
Villeux R (1985) Diploid and polyploidy gametes in Crop Plants:
mechanisms of formation and utilization in plant breeding. In:
Janick J (ed) Plant Breed Rev 3. AVI, New York, p 442
Vorsa N, Bingham E (1979) Cytology of 2n pollen formation
in diploid alfalfa, Medicago sativa. Can J Genet Cytol 21:525–
530
Web: Missouri Botanical Garden Tropics, Nomenclatural database
http://mobot.mobot.org/cgi-bin/searchvast, accessed on 28
August 2011
Wolf T (1908) Monographie der Gattung Potentilla. Biblliotbeca Bot
71:1–715
496 S. Rani et al.
123

Zhao ZF, Wang YQ, Huang S (1990) Plant chromosome counts (V).
For Res 3:503–508
Zhukova PG (1967) Karyology of some plants cultivated in the
Arctic-Alpine Botanical Garden. In: Avrorin NA (ed) Plantarum
in Zonam Polarem Transportatio II. Leningrad pp 139–149
Zlesak DC (2009) Pollen diameter and guard cell length as predictor
of ploidy in diverse rose cultivars, species, and breeding lines.
In: Zlesak DC (ed) Roses. Floricult ornament biotechnol, vol 3
(special issue 1). Global science books Ltd., UK, pp 53–70
Additions to the cytologically investigated species of Potentilla L. 497
123
Recommended

















