
ORIGINAL PAPER
Biodegradation of feather waste by extracellular keratinasesand gelatinases from Bacillus spp.
Ana Maria Mazotto • Ana Cristina N. de Melo • Andrew Macrae •
Alexandre Soares Rosado • Raquel Peixoto • Sabrina M. L. Cedrola •
Sonia Couri • Russolina B. Zingali • Ana Lucia V. Villa • Leon Rabinovitch •
Jeane Q. Chaves • Alane B. Vermelho
Received: 30 April 2010 / Accepted: 20 September 2010 / Published online: 13 October 2010
� Springer Science+Business Media B.V. 2010
Abstract In this study, three feather degrading bacterial
strains were isolated from agroindustrial residues from a
Brazilian poultry farm. Three Gram-positive, spore-form-
ing, rod-shaped bacteria and were identified as B. subtilis
1271, B. licheniformis 1269 and B. cereus 1268 using
biochemical, physiologic and molecular methods. These
Bacillus spp. strains grew and produced keratinases and
peptidases using chicken feather as the sole source of
nitrogen and carbon. B. subtilis 1271 degraded feathers
completely after 7 days at room temperature and produced
the highest levels of keratinase (446 U ml-1). Feather
hydrolysis resulted in the production of serine, glycine,
glutamic acid, valine and leucine as the major amino acids.
Enzymography and zymography analyses demonstrated
that enzymatic extracts from the Bacillus spp. effectively
degraded keratin and gelatin substrates as well as, casein,
hemoglobin and bovine serum albumin. Zymography
showed that B. subtilis 1271 and B. licheniformis 1269
produced peptidases and keratinases in the 15–140 kDa
range, and B. cereus produced a keratinase of *200 kDa
using feathers as the carbon and nitrogen source in culture
medium. All peptidases and keratinases observed were
inhibited by the serine specific peptidase inhibitor phen-
ylmethylsulfonyl fluoride (PMSF). The optimum assay
conditions of temperature and pH for keratinase activity
were 40–50�C and pH 10.0 for all strains. For gelatinases
the best temperature and pH ranges were 50–70�C and pH
7.0–11. These isolates have potential for the biodegrada-
tion of feather wastes and production of proteolytic
enzymes using feather as a cheap and eco-friendly
substrate.
Keywords Bacillus spp. � Feather degradation � Feather
keratin � Keratinase � Peptidase
Introduction
In poultry processing industries all over the world, chicken
feathers are generally an unwanted waste byproduct (Su-
zuki et al. 2006). In Brazil 800,000 tons/year of feathers
are discarded by this sector. The accumulation of feathers
can eventually lead to environmental pollution and can also
be considered as a waste of feather protein (Onifade et al.
A. M. Mazotto � A. C. N. de Melo � A. Macrae �A. S. Rosado � R. Peixoto � S. M. L. Cedrola �A. B. Vermelho (&)
Departamento de Microbiologia Geral, Instituto de
Microbiologia Prof. Paulo de Goes (IMPPG), Bloco I, Centro de
Ciencias da Saude (CCS), Universidade Federal do Rio de
Janeiro (UFRJ), Cidade Universitaria, Ilha do Fundao, Rio de
Janeiro, RJ 21941-590, Brazil
e-mail: [email protected]
S. Couri
Instituto Federal de Educacao Ciencia e Tecnologia do Rio de
janeiro, Campus Rio de Janeiro, Rua Senador Furtado no 121,
Maracana, Rio de Janeiro, Brazil
R. B. Zingali
Departamento de Bioquımica, Instituto de Ciencias Biomedicas,
Bloco H, Centro de Ciencias da Saude (CCS), Universidade
Federal do Rio de Janeiro (UFRJ), Cidade Universitaria, Ilha do
Fundao, Rio de Janeiro, RJ 21941-590, Brazil
A. L. V. Villa
Universidade federal do Rio de Janeiro, Campus Macae,
R. Aloısio Gomes da Silva no 50, Granja do Cavaleiros, Macae,
RJ 27930, Brazil
L. Rabinovitch � J. Q. Chaves
Departamento de Bacteriologia, Instituto Oswaldo Cruz,
Fundacao Oswaldo Cruz, Av. Brasil 4365, Rio de Janeiro,
RJ 21045-900, Brazil
123
World J Microbiol Biotechnol (2011) 27:1355–1365
DOI 10.1007/s11274-010-0586-1

1998). Traditional ways to degrade feathers such as alkali
hydrolysis and steam pressure cooking to produce feather
meal may destroy amino acids and they also consume large
amounts of energy (Dalev et al. 1997; Cai et al. 2008). In
addition to the conventional methods, the incineration of
feathers has ecological disadvantages including the release
of large amounts of carbon dioxide and there is an apparent
protein wastage and not to mention energy losses (Matsui
et al. 2009). Feathers are comprised essentially of keratin,
an insoluble structural protein, tightly packed in a b-sheet
polypeptide chain, extensively cross-linked with disulfide,
hydrogen and hydrophobic bonds (Riffel et al. 2003; Fraser
and Parry 2008). Keratinous wastes are not degraded by
commonly known proteases like trypsin, pepsin and papain
due to presence of the disulfide bonds, but are easily
degraded by keratinases (Papadopoulos 1986, Gupta and
Ramnani 2006). These peptidases are largely serine or
metallopeptidases (EC 3.4.21/24) found in several micro-
organisms and have attracted a great deal of attention due
to their multiple applications in industry for the develop-
ment of nonpolluting processes (Onifade et al. 1998; Gupta
and Ramnani 2006).
Keratinolytic microorganisms and their enzymes may
have important applications in biotechnological and
industrial processes involving keratin-containing wastes
from poultry and leather industries. In the pharmaceutical
industry, they have a role in personal care products
involving hair removal and as peeling agents (Grazziotin
et al. 2006; Brandelli 2008; Macedo et al. 2008; Pillai and
Archana 2008). Biotechnological applications involving
keratin and keratinases include bio-hydrogen production,
biodegradable films and keratin composites (Balint et al.
2005; Barone et al. 2005). In medicine, recent studies
indicate that keratinases may have an important role in
deactivating prions and could increase ungual drug deliv-
ery (Mohorcic et al. 2007; Yoshioka et al. 2007). Biodeg-
radation of feathers by keratinase from microorganisms
may provide a viable alternative to produce a digestible
keratin through peptide production. These enzymes are
produced by some species of Bacillus genus, actinomycetes
and fungi (Gupta and Ramnani 2006). Keratinases, from B.
licheniformis and B. subtilis have been studied and shown
to be effective at feather degradation (Rozs et al. 2001;
Thys and Brandelli 2006). These microorganisms are a
source of Versazyme and Valkerase, commercial keratin-
ases which have been used for feather meal improvement
(Odetallah et al. 2005).
Here we describe the isolation and identification of three
keratin degrading Bacillus strains isolated from a poultry
industry. Keratinases and peptidases produced by Bacillus
spp. during submerse cultivation on feather medium were
investigated by qualitative and quantitative analyses using
zymography, spectrophotometry and enzymography. We
also report the partial characterization of these enzymes
produced by three novel Bacillus spp.
Materials and methods
Chemicals
Media constituents were obtained from Oxoid Ltd. (Cam-
bridge, England). Reagents used in electrophoresis and
molecular mass standards were acquired from Amersham
Life Science (Little Chalfont, England). Polyethylenegly-
col 4000 (PEG 4000) was purchased from Vetec (Rio de
Janeiro, Brazil). The peptidase inhibitors (trans-epox-
ysuccinyl l-leucylamido- (4-guanidino) butane [E-64],
phenylmethylsulphonyl fluoride [PMSF], 1,10-phenan-
throline, pepstatin A and EDTA) were obtained from
Sigma Chemical Co. (St. Louis, MO, USA). The protein-
aceous substrates gelatin and casein were purchased from
Merck (Darmstadt, Germany), bovine serum albumin and
hemoglobin from Sigma Chemical Co. (St. Louis, MO,
USA). All other reagents were analytical grade.
Isolation and selection of keratinolytic microorganism
Feather-degrading Bacillus spp. strains were isolated from
industrial residues of a local poultry industry (Rio de
Janeiro, Brazil). One gram of feather wastes was added to a
100 ml aqueous solution comprising 0.5% yeast extract,
0.5% peptone, 2% sucrose and 2% KCl and stored at 28�C
for 48 h. The suspension was streaked on agar plates
containing the same medium composition and incubated at
28�C for 72 h. The single colonies obtained were cultivated
in tubes containing saline solution (0.85% NaCl) supple-
mented with a whole feather for 28 days at 28�C in order to
select keratinolytic microorganisms based on their ability
to hydrolyze feather.
Phenotypic and molecular identification
of the keratinolytic microorganisms
The feather degrading strains were evaluated by specific
biochemical, physiologic and cytomorphologic tests for to
identify bacteria belonging to the Genus Bacillus (Claus
and Berkeley 1986; Vasconcelos and Rabinovitch 1994).
Strains were preserved under refrigeration as spores in
solid Nutrient Agar medium. The strains were lyophilized
and deposited in the Colecao de Culturas do Genero
Bacillus e Generos Correlatos-CCGB (which is affiliated
to the World Federation of Culture Collections) located in
the Laboratorio de Fisiologia Bacteriana, Instituto
Oswaldo Cruz/FIOCRUZ.
1356 World J Microbiol Biotechnol (2011) 27:1355–1365
123

Genomic DNA of the bacterial isolates was extracted
using the Wizard Genomic DNA Purification Kit (Pro-
mega, Madison, USA). Concentrations were determined by
using of a Qubit fluorometer (Invitrogen/Molecular Probes
California, USA). Amplification of the 16S rDNA region
was performed by PCR using universal primers pA and pH
as previously described (Massol-Deya et al. 1995). The
amplified products were purified with Ilustra GFX PCR
DNA and Gel band Purification Kit (Ge Healthcare,
Buckinghamshire, UK) and then directly sequenced in both
directions by using pA and pH primers with a MEGAB-
ACE system (Ge Healthcare, Buckinghamshire, UK).
Preliminary identification of sequences was performed by
blastn against the Genbank database 14/08/2009 (www.
ncbi.nlm.nih.gov/blast). The sequence was submitted to the
GenBank using Sequin Application Version 9.50 (ftp://
ftp.ncbi.nih.gov/sequin/sequin.win.exe).
Growth medium and enzyme extract preparation
The Bacillus sp. strains were cultivated on phosphate-
buffered medium (0.06 M Na2HPO4�7H2O and 0.04 M
KH2PO4, pH 7.2) supplemented with 1% native feather as
the only nitrogen and carbon source for 7 days at 28�C on a
rotary shaker (300 rpm). After incubation, the media were
centrifuged at 2,000g for 20 min at 4�C and the supernatant
solutions were used as enzyme extracts to analyze kera-
tinase and gelatinase activities.
Feather keratin substrate
Chicken feathers obtained from poultry waste were washed
extensively with water and detergent, dried at 60�C over-
night, delipidated with chloroform:methanol (1:1, v/v) and
dried again at 60�C. The Wawrzkiewicz et al. (1991)
method was modified to produce keratin powder from the
lipid free dried feathers. Briefly, 10 g of feathers were
heated with a reflux condenser at 100�C for 80–120 min
with 500 ml of DMSO. Keratin was then precipitated by
the addition of two volumes of acetone and maintained at
4�C for 24–48 h. The keratin precipitates were collected by
centrifugation (2,000g/15 min), washed twice with distilled
water and dried at 4�C. A white powder was obtained for
qualitative and quantitative biochemical analyses related to
keratinases activity and as a keratin standard in feather
degradation studies (Vermelho et al. 2009).
SDS–PAGE
Feather keratin powder was analyzed in 15% SDS–PAGE
using the method of Laemmli (1970). Electrophoresis was
carried out at 170 V. A keratin solution (1 mg/ml) was
added to the SDS–PAGE sample buffer in the proportion of
6:4 (v/v). The gels were silver and coomassie blue stained.
Phosphorylase b (94 kDa), bovine serum albumin (67 kDa),
ovalbumin (43 kDa), carbonic anhydrase (30 kDa), soybean
trypsin inhibitor (20.1 kDa) and a-lactalbumin (14.4 kDa)
were used as molecular mass standards (Pharmacia Biotech).
Matrix-assisted laser desorption/ionization-time
of flight (MALDI-TOF) mass spectrometry (MS)
In order to verify the homogeneity of the keratin substrate
obtained from the feathers, the keratin powder was ana-
lyzed using matrix assisted laser desorption/ionization time
of flight mass spectrometry (MALDI-TOF MS). Immedi-
ately prior to mass spectrometry, acetonitrile/water (5:95,
v/v) and trifluoroacetic acid were added to the samples.
A C18 zip tip was hydrated, the sample loaded, and water
used to wash the sample. An elution solution of acetoni-
trile/water (60:40, v/v), and 0.1% (w/v) trifluoroacetic acid
was then loaded three times, a-cyano-4- Hydroxycinnamic
acid (CCA) matrix was added, and 1 ll of the sample
mixture was spotted directly on a MALDI target for anal-
ysis. Peptide mass spectrometry was carried out with a
Bruker Biflex III MALDI-TOF mass spectrometer in the
reflectron mode. The experiments were performed in trip-
licate using three independent experimental sets.
Feather keratin enzymography
Twenty microliter samples of concentrated culture super-
natant were mixed with an equal volume of feather keratin
powder diluted in phosphate buffer (0.06 M Na2H-
PO4�7H2O and 0.04 M KH2PO4, pH 7.2) to obtain a final
concentration of 0.15 mg/ml. The reaction mixtures were
incubated for 1 h at 37�C. Reactions were terminated by
freezing and the mixtures stored at -20�C until analyzed.
Reaction mixtures (20 ll) were added to 20 ll SDS–PAGE
sample buffer [125 mM Tris, pH 6.8, 4% (w/v) SDS, 20%
(v/v) glycerol, 0.002% (w/v) bromophenol blue] supple-
mented with 5% (v/v) b-mercaptoethanol, and boiled at
100�C for 5 min. Keratin hydrolysis was then analyzed on
15% SDS–PAGE. Electrophoresis was carried out at 170 V
for 2 h at 28�C, and protein bands were silver stained. As a
control of enzymatic activity, aliquots of the concentrated
supernatant were heat-inactivated with the substrates
before incubation. In addition, a second control for keratin
substrate was made by replacing concentrated supernatants
with the same volume of buffer (De Melo et al. 2007).
Quantifying keratinase and gelatinase activities
Enzymatic activity was evaluated at different pH values
(3.0–11.0) and temperatures (10–80�C). Enzyme extract
World J Microbiol Biotechnol (2011) 27:1355–1365 1357
123

(250 ll) was added to 375 ll of feather keratin powder
(6.67%) in a citric acid buffer (0.05 M citric acid pH 3.0,
4.0 and 5.0), phosphate buffer (0.06 M Na2HPO4�7H2O
and 0.04 M KH2PO4 pH 6.0, 7.0 and 8.0) or aminoacetic
acid buffer (0.1 M aminoacetic acid pH 9.0, 10.0 and 11.0).
The reaction mixture was incubated for 1 h at 37�C and
then stopped by the addition of 250 ll of 10% trichloro-
acetic acid. Samples were then put into a refrigerator at 4�C
for 30 min. The supernatant was collected after centrifu-
gation (15 min at 2,500g) and activity measured at 280 nm.
One unit of keratinolytic activity was defined as the amount
of enzyme required to produce an increase of 0.01 absor-
bance unit at 280 nm, under standard assay conditions (1 h
at 37�C).
Gelatinase activity was measured according to the
method of Jones et al. (1998). Briefly, 100 ll of the
enzyme extract and 900 ll of the same buffer solutions
above were added to 1.5 ml of the substrate solution
(gelatin in distilled water, 1% w/v) and the mixture was
incubated at 37�C for 30 min. A 375 ll sample was then
removed from the reaction mixture and added to 500 ll of
isopropanol. This was centrifuged at 2,500g for 15 min and
the supernatant was collected and the absorbance was
measured as described by Lowry et al. (1951). One unit of
gelatinolytic activity was defined as the amount of enzyme
required to produce an increase of 0.01 absorbance unit at
660 nm, under standard assay conditions (30 min at 37�C).
High-performance thin-layer chromatography (HPTLC)
An aliquot of culture supernatant (5 ll) was analyzed by
silica gel 60 HPTLC plates for amino acid detection. The
HPLC plates were run for approximately 1 h at room
temperature in a TLC tank using butanol/acetic acid/dis-
tilled water (4:1:1 v/v/v) as solvent. The amino acid spe-
cific ninhydrin reagent (7.5% in butanol/acetone 1:1 v/v)
was used for development. Commercial amino acids were
used as standard.
Zymography
Culture supernatants were concentrated 20-fold by dialysis
(cut off 9 kDa) against PEG 4000 overnight at 4�C. The
concentrated culture supernatants were mixed with sample
buffer for zymography (125 mM Tris, pH 6.8, 4% SDS,
20% glycerol and 0.002% bromophenol blue) in a sam-
ple:buffer ratio 6:4 (De Melo et al. 2007; Vermelho et al.
2009). Keratinases and gelatinases were assayed and
characterized by 12.5% SDS–PAGE with co-polymerized
keratin feather powder and gelatin (0.1%). Additionally
other substrates such as casein, bovine serum albumin
(BSA) and hemoglobin (0.1%) were incorporated in gel for
substrate specificity studies. Gels were loaded with 30 ll of
concentrated culture supernatant per slot for keratin-SDS–
PAGE, and 20 ll of concentrated culture supernatant per
slot for other substrates co-polymerized in SDS–PAGE.
After electrophoresis at 170 V for 2 h at 4�C the gels were
soaked for 1 h at 28�C in 2.5% Triton X-100. Afterwards,
the gels were then incubated for 48 h at 37�C in proteolysis
buffer Tris–HCl buffer, pH 7.4 (0.5 M Tris). Then, the gels
were stained for 1 h with 0.2% Coomassie brilliant blue R-
250 in methanol-acetic acid–water (50:10:40) and de-
stained in the same solvent (Lopes et al. 2008). For enzy-
matic classification 3 mM phenylmethylsulfonyl fluoride
(PMSF), 0.26 mM ethylenediaminetetraacetic acid
(EDTA), 10 mM 1,10 phenanthroline (Phenan), 10 lM
pepstatin A (Peps) and 5 lM trans-epoxysuccinyl L-leu-
cylamido-(4-guanidino) butane (E-64) were used in prote-
olysis buffer. To quantify the inhibition the bands were
analyzed by ImageJ software.
Results
Selection and identification of keratinolytic Bacillus
spp.
Three aerobic, mesophilic, Gram-positive, and spore-
forming bacilli were selected as keratinolytic after dem-
onstrating complete feather degradation in whole feather
broth. They were identified by biochemical, physiological
and cytomorphological characterization as Bacillus li-
cheniformis (LFB-FIOCRUZ 1269), Bacillus subtilis
(LFB-FIOCRUZ 1271) and Bacillus cereus (LFB-FIO-
CRUZ 1268) at the Laboratorio de Fisiologia Bacteriana,
Instituto Oswaldo Cruz/FIOCRUZ, Brazil. The analyses of
their 16S rDNA sequences confirmed the biochemical
results indicating the identification of Bacillus subtilis,
Bacillus licheniformis and Bacillus cereus. The 16S rDNA
sequences of isolates 1268, 1269 and 1271 showed 99%
sequence similarity with Bacillus cereus, Bacillus licheni-
formis and Bacillus subtilis, respectively. The nucleotide
sequences were deposited at GenBank and their accession
numbers are: GQ482980 (B. subtilis 1271), GQ482981
(B. licheniformis 1269) and GQ482979 (B. cereus 1268).
Production of keratinases and peptidases by Bacillus
species in feather medium
The three Bacillus species were further tested for keratin-
olytic and proteolytic activity. All the strains grew and
produced keratinase and peptidase using chicken feather as
the sole source of nitrogen and carbon. B. subtilis 1271
degraded the feathers completely after 7 days at room
temperature and produced the highest level of keratinase
(446 U/ml), Fig. 1. B. licheniformis 1269 produced lower
1358 World J Microbiol Biotechnol (2011) 27:1355–1365
123

keratinolytic activity and a higher level of proteolytic
activity (394.1 U/ml), as shown in Fig. 1. B. licheniformis
1269 presented only 29.5 U/ml of keratinolytic activity but
was able to degrade 44.2% of the feathers in the medium
(Figs. 1 and 2).
The soluble proteins and free amino acids produced
during feather hydrolysis by Bacillus strains were ana-
lyzed. HTLC analyses of the culture supernatants of the
three strains showed amino acids bands migrating at the
serine, glycine, glutamic acid, valine and leucine positions
(Fig. 2a), indicating that these amino acids are present in
high concentrations. The highest concentration of soluble
protein (4.0 mg/ml) was observed in the B. subtilis 1271
supernatant (Fig. 2b), while B. licheniformis 1269 had the
lowest concentration of soluble protein (1.42 mg/ml).
Feather keratin degradation by keratinases of Bacillus
species
Keratin was successfully extracted from feathers using the
method described and resulted in a white, homogenous
keratin powder. MALDI-TOF and SDS–PAGE analyses
(Fig. 3a, b, respectively) confirmed the presence of pure
keratin. The major fragments were in the m/z 9,000–10,000
range, confirming the presence of a b-keratin (Fig. 3a).
Fragments in m/z 2,000–8,000 range corresponded to the
a-cyno-4-hydroxycinnamic acid matrix. SDS–PAGE
analyses resulted in a single band migrating at *10 kDa
(Fig. 3b). Cell-free supernatants from the three Bacillus
species were then incubated with feather keratin for 1 h at
37�C. After incubation, the single *10 kDa band, char-
acteristic of feather keratin, was no longer observed and
0
100
200
300
400
500
B. cereus 1268
B. lichenifo
rmis 1269
B. subtilis 1271
Act
ivit
y (U
/ml)
Fig. 1 Keratinolytic and proteolytic activity of Bacillus strains
grown in feather medium. The black bar represents the keratinolytic
activity and the gray bars the proteolytic activity
Fig. 2 Degradation of feather in culture medium, demonstrating
protein concentration and TLC of amino acids from culture super-
natant of Bacillus strains. a TLC of amino acid produced during
degradation of feather by B. cereus 1268, B. licheniformis 1269 and
B. subtilis 1271. b Concentration of soluble proteins in culture
medium (bars) and feather degradation (filled diamond) after
incubation with B. cereus 1268, B. licheniformis 1269 and B. subtilis1271 for 7 days at 28�C
Fig. 3 Feather keratin degradation by Bacillus strains. a MALDI
TOF spectrum: major fragments of 9,819.85 and 10,301.27 Da (m/z)
were detected indicating the presence of keratin. b SDS–PAGE of
keratin feather powder: first column from top to bottom Phosphor-
ylase b (97 kDa), bovine serum albumin (66 kDa), ovalbumin
(45 kDa), carbonic anhydrase (30 kDa), soybean trypsin inhibitor
(20.1 kDa) and a-lactalbumin (14.4 kDa) were used as molecular
mass standards (Pharmacia Biotech), second column a single
*10 kDa band. c Enzymography of keratin degradation by extracel-
lular keratinases of B. cereus 1268, B. licheniformis 1269 and
B. subtilis 1271. Feather keratin degradation profiles were analyzed
by 15% SDS–PAGE, and gels were silver stained. In the gel strips on
the left, the enzyme extract solution immediately after the addition of
feather keratin (note 10 kDa band); in strips on the right, the same
solution incubated for 1 h at 37�C (note lack of 10 kDa band)
World J Microbiol Biotechnol (2011) 27:1355–1365 1359
123

considered degraded (Fig. 3c), confirming the presence of
keratinases in the supernatants.
Effect of different substrates on the extracellular
peptidases of Bacillus spp.
The ability of the extracellular peptidases to degrade dif-
ferent proteinaceous substrates was evaluated using kera-
tin, gelatin, casein, BSA and hemoglobin co-polymerized
with 12.5% sodium dodecyl sulfate–polyacrylamide gels
(SDS–PAGE). The gels showed that B. cereus 1268 pro-
duced a single peptidase of *200 kDa able to degrade all
substrates tested (Fig. 4). B. subtilis 1271 and B. licheni-
formis 1269 produced multiple peptidases with the ability
to degrade gelatin (Fig. 4). In B. licheniformis, bands with
an apparent molecular mass of 60 and 100 kDa presented
keratinase activity but these enzymes were more prominent
in B. subtilis 1271 which presented bands migrating in a
range of 15–140 kDa. A broad range of other protein
substrates like casein, BSA and hemoglobin were hydro-
lyzed by B. subtilis and B licheniformis. A peptidase
migrating at 30 kDa in B. licheniformis showed strong
activity with casein, BSA and hemoglobin (Fig. 4).
Effect of temperature and pH on enzyme activity
The effect of temperature and pH on extracellular kera-
tinase and gelatinase activities of the three Bacillus spp.
was determined using gelatin and feather keratin as sub-
strates, respectively. High gelatinase activity was observed
at 60�C for B. subtilis 1271 and 70�C for B. licheniformis
1269 (Fig. 5a). Meanwhile, the gelatinase of B. cereus
1268 was more active at a broader temperature range
(50–70�C). Both keratinases of B. cereus 1268 and B. li-
cheniformis 1269 were optimally active at 40�C. The ker-
atinase of B. subtilis exhibited maximal activity at 50�C
(Fig. 5a). B. subtilis strain 1271 presented maximum gel-
atinase and keratinase activity at pH 9.0 and 10, respec-
tively (Fig. 5b) and in B. cereus the optimum pH was 10
for both substrates (Fig. 5a). In B. licheniformis, the gela-
tinases were active in the range of 7.0–11 and keratinolytic
activity was highest at pH 10 (Fig. 5b), approximately 75%
of the activity was lost at pH 9.0 and 11.0.
Effect of inhibitors on enzyme activity
To characterize the extracellular proteolytic activities of
Bacillus spp. isolates, zymogram gels containing keratin or
gelatin as substrates were incubated in the absence and in
the presence of proteolytic inhibitors of the four major
peptidase classes (aspartic, cysteine, serine and metallo-
peptidase). The proteolytic inhibition results showed that
the profiles of the extracellular peptidases expressed by
these Bacillus spp. are composed of serine peptidases
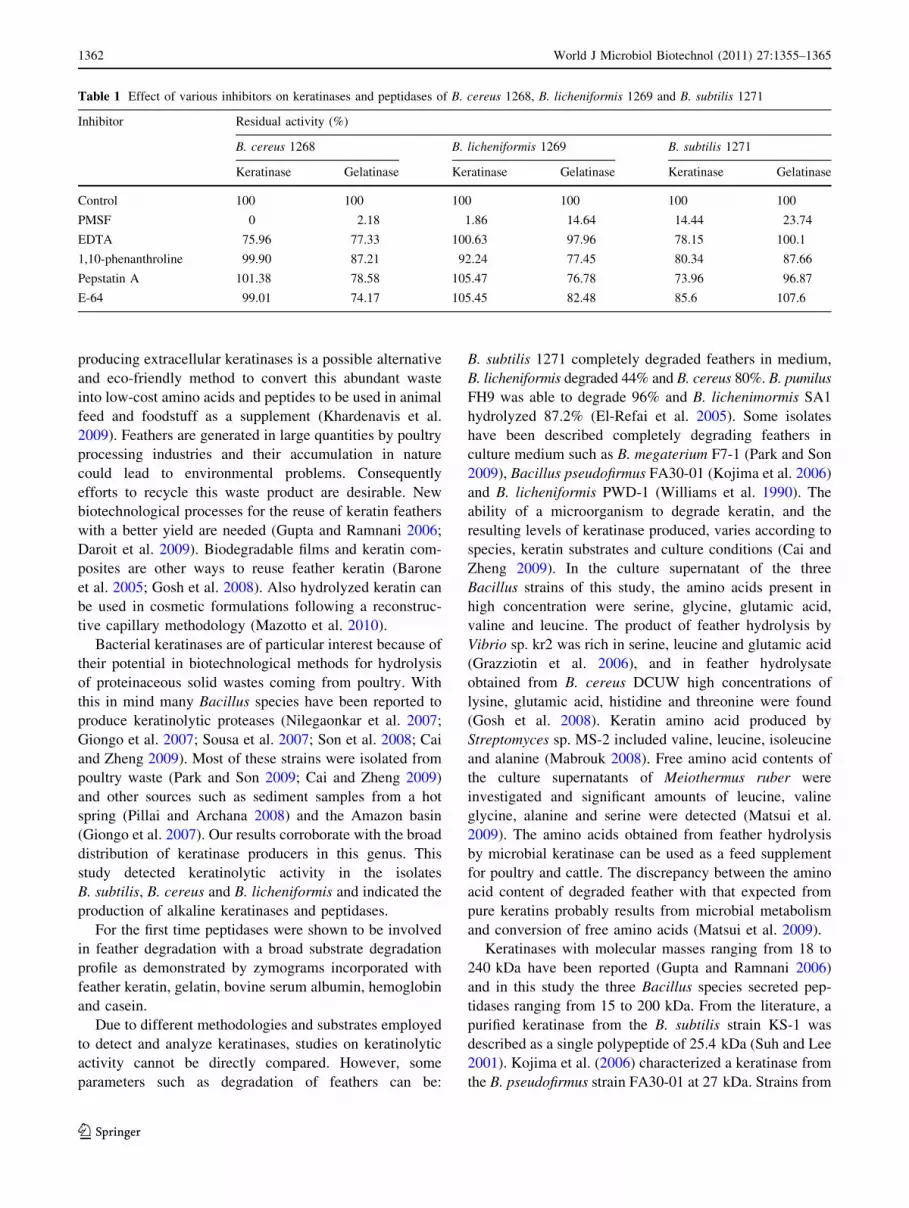
(Fig. 6). As shown in Table 1, keratinases and peptidases
were strongly inhibited by PMFS. E-64 (a cysteine pepti-
dase inhibitor), pepstatin A (an aspartic peptidase inhibi-
tor), EDTA and 1,10-phenanthroline (metallopeptidase
inhibitor or metal-dependent enzyme) did not alter signif-
icantly the behavior of the enzymes.
Fig. 4 Zymograms with co-polymerized gelatin, feather keratin,
casein, BSA or hemoglobin. Enzyme extracts of B. subtilis 1271,B. cereus 1268 and B. licheniformis 1269 were prepared as described
in material and methods. Gel strips containing 30 ll of concentrated
culture supernatant were incubated for 48 h at 37�C in 0.5 M Tris–
HCl, pH 7.4. The molecular masses of the peptidases, expressed in
kDa, are shown on the left
1360 World J Microbiol Biotechnol (2011) 27:1355–1365
123

Discussion
Here we have demonstrated the degradation of intact
feathers and amino acid production by submerse fermen-
tation, using three Bacillus spp. isolated from poultry
waste. Currently some of the feather waste produced by the
poultry industry is transformed into feather meal, however
the final product is not very digestible and some essential
amino acids such as methionine, lysine, and tryptophan are
lost and other non-nutritive amino acids, such as lysino-
alanine and lanthionine are formed (Dalev et al. 1997;
Matsui et al. 2009). In spite of its limitations, this meal is
already incorporated into the diet, as feed, for chicken
poultry, rainbow trout, shrimp and salmon. However this
type of feed needs an amino acid supplement (Bertsch and
Coello 2005). The use of microorganisms capable of
Fig. 5 Effect of temperature (a) and pH (b) on keratinolytic (filled square) and gelatinolytic (filled diamond) activity of extract enzymatic from
B. cereus 1268, B. licheniformis 1269 and B. subtilis 1271
Fig. 6 Effects of proteolytic inhibitors on the extracellular peptidases
and keratinases of B. cereus 1268, B. licheniformis 1269 and
B. subtilis 1271, in gelatin zymogram (a) and keratin zymogram
(b). Gel strips were incubated separately in the absence (control) or in
the presence of different proteolytic inhibitors: 3 mmol l-1
phenylmethylsulfonyl fluoride (PMSF), 0.26 mmol l-1 ethylenedia-
minetetraacetic acid (EDTA), 10 mmol l-1 1,10 phenanthroline
(Phenan), 10 lmol l-1 pepstatin A (Peps) and 5 lmol l-1 M trans-
epoxysuccinyl L-leucylamido-(4-guanidino) butane (E-64)
World J Microbiol Biotechnol (2011) 27:1355–1365 1361
123
producing extracellular keratinases is a possible alternative
and eco-friendly method to convert this abundant waste
into low-cost amino acids and peptides to be used in animal
feed and foodstuff as a supplement (Khardenavis et al.
2009). Feathers are generated in large quantities by poultry
processing industries and their accumulation in nature
could lead to environmental problems. Consequently
efforts to recycle this waste product are desirable. New
biotechnological processes for the reuse of keratin feathers
with a better yield are needed (Gupta and Ramnani 2006;
Daroit et al. 2009). Biodegradable films and keratin com-
posites are other ways to reuse feather keratin (Barone
et al. 2005; Gosh et al. 2008). Also hydrolyzed keratin can
be used in cosmetic formulations following a reconstruc-
tive capillary methodology (Mazotto et al. 2010).
Bacterial keratinases are of particular interest because of
their potential in biotechnological methods for hydrolysis
of proteinaceous solid wastes coming from poultry. With
this in mind many Bacillus species have been reported to
produce keratinolytic proteases (Nilegaonkar et al. 2007;
Giongo et al. 2007; Sousa et al. 2007; Son et al. 2008; Cai
and Zheng 2009). Most of these strains were isolated from
poultry waste (Park and Son 2009; Cai and Zheng 2009)
and other sources such as sediment samples from a hot
spring (Pillai and Archana 2008) and the Amazon basin
(Giongo et al. 2007). Our results corroborate with the broad
distribution of keratinase producers in this genus. This
study detected keratinolytic activity in the isolates
B. subtilis, B. cereus and B. licheniformis and indicated the
production of alkaline keratinases and peptidases.
For the first time peptidases were shown to be involved
in feather degradation with a broad substrate degradation
profile as demonstrated by zymograms incorporated with
feather keratin, gelatin, bovine serum albumin, hemoglobin
and casein.
Due to different methodologies and substrates employed
to detect and analyze keratinases, studies on keratinolytic
activity cannot be directly compared. However, some
parameters such as degradation of feathers can be:
B. subtilis 1271 completely degraded feathers in medium,
B. licheniformis degraded 44% and B. cereus 80%. B. pumilus
FH9 was able to degrade 96% and B. lichenimormis SA1
hydrolyzed 87.2% (El-Refai et al. 2005). Some isolates
have been described completely degrading feathers in
culture medium such as B. megaterium F7-1 (Park and Son
2009), Bacillus pseudofirmus FA30-01 (Kojima et al. 2006)
and B. licheniformis PWD-1 (Williams et al. 1990). The
ability of a microorganism to degrade keratin, and the
resulting levels of keratinase produced, varies according to
species, keratin substrates and culture conditions (Cai and
Zheng 2009). In the culture supernatant of the three
Bacillus strains of this study, the amino acids present in
high concentration were serine, glycine, glutamic acid,
valine and leucine. The product of feather hydrolysis by
Vibrio sp. kr2 was rich in serine, leucine and glutamic acid
(Grazziotin et al. 2006), and in feather hydrolysate
obtained from B. cereus DCUW high concentrations of
lysine, glutamic acid, histidine and threonine were found
(Gosh et al. 2008). Keratin amino acid produced by
Streptomyces sp. MS-2 included valine, leucine, isoleucine
and alanine (Mabrouk 2008). Free amino acid contents of
the culture supernatants of Meiothermus ruber were
investigated and significant amounts of leucine, valine
glycine, alanine and serine were detected (Matsui et al.
2009). The amino acids obtained from feather hydrolysis
by microbial keratinase can be used as a feed supplement
for poultry and cattle. The discrepancy between the amino
acid content of degraded feather with that expected from
pure keratins probably results from microbial metabolism
and conversion of free amino acids (Matsui et al. 2009).
Keratinases with molecular masses ranging from 18 to
240 kDa have been reported (Gupta and Ramnani 2006)
and in this study the three Bacillus species secreted pep-
tidases ranging from 15 to 200 kDa. From the literature, a
purified keratinase from the B. subtilis strain KS-1 was
described as a single polypeptide of 25.4 kDa (Suh and Lee
2001). Kojima et al. (2006) characterized a keratinase from
the B. pseudofirmus strain FA30-01 at 27 kDa. Strains from
Table 1 Effect of various inhibitors on keratinases and peptidases of B. cereus 1268, B. licheniformis 1269 and B. subtilis 1271
Inhibitor Residual activity (%)
B. cereus 1268 B. licheniformis 1269 B. subtilis 1271
Keratinase Gelatinase Keratinase Gelatinase Keratinase Gelatinase
Control 100 100 100 100 100 100
PMSF 0 2.18 1.86 14.64 14.44 23.74
EDTA 75.96 77.33 100.63 97.96 78.15 100.1
1,10-phenanthroline 99.90 87.21 92.24 77.45 80.34 87.66
Pepstatin A 101.38 78.58 105.47 76.78 73.96 96.87
E-64 99.01 74.17 105.45 82.48 85.6 107.6
1362 World J Microbiol Biotechnol (2011) 27:1355–1365
123

the genus Bacillus, in particular the B. licheniformis strains,
have been shown to secrete extracellular keratinases in the
33 to 42 kDa range (Lin et al. 1992; Rozs et al. 2001). B.
lichenifornis 1269 produced a 60 kDa keratinase that
degraded keratin and gelatin, but not the other substrates.
This keratinase has the potential to be used in the dehairing
process in the leather industry. This specific keratinolytic
activity is advantageous in the leather industry, because
collagen, the major leather-forming protein, is not signifi-
cantly degraded by the keratinase (Pillai and Archana
2008). Interestingly, the B. cereus strain used in this
investigation produced an extracellular keratinase with an
apparent molecular mass of 200 kDa. This result is similar
to the results found for Kocuria rosea and Fervidobacte-
rium islandicum keratinases with 240 and 200 kDa,
respectively (Bernal et al. 2006; Nam et al. 2002), but
unknown until now within the genus Bacillus. Other
studies have analyzed extracellular keratinase production
using zymography with non specific substrates such as
gelatin (Nam et al. 2002; Giongo et al. 2007; Gosh et al.
2008), casein (Nilegaonkar et al. 2007; Sousa et al. 2007)
with feather meal (Kojima et al. 2006) incorporated in the
gel. These studies detected only one or two peptidases in
the culture supernatant of keratinolytic microorganisms. In
contrast, in this study a large number of peptidases were
detected in the culture supernatant of B. licheniformis 1269
and B. subtilis 1271 by gelatin zymography. Additionally
five keratinases were detected by keratin zymography in
the B. subtilis 1271 culture supernatant. This is the first
study reporting multiple keratinase production. The results
suggest that feather degradation is not due to a single
keratinase.
Most keratinases from Bacillus spp. described belong to
the serine peptidase class. Therefore, phenylmethane-
sulfonyl fluoride (PMSF) is the potential inhibitor of these
enzymes. However, some Bacillus keratinases can be par-
tially inhibited by EDTA due the importance of cations as
stabilizing agents in these keratinases (Ramnani and Gupta
2006). Keratinases of B. cereus DCUW, Bacillus sp. P13
and B. pseudofirmus FA30-01 were completely inhibited
by PMSF (Kojima et al. 2006; Gosh et al. 2008; Pillai and
Archana 2008), as well as the keratinases produced by
B. subtilis 1271, B. cereus 1268 and B. licheniformis 1269.
In this work, we have shown that feathers are a cheap
and environmentally friendly substrate for the production
of multiple alkaline peptidases by Bacillus strains. Pepti-
dases have been routinely used in industry for various
purposes including bioremediation processes, in the phar-
maceutical industry, cheese making, baking, preparation of
soy hydrolysates, debittering of protein hydrolysates, lea-
ther treatment and mainly in laundry (Rao et al. 1998).
Among these peptidases, alkaline peptidases are the most
appropriate as detergent additives because they digest
various proteinaceous stains (Saeki et al. 2007). The pep-
tidases produced by Bacillus spp. isolated in this study
were able to degrade keratin and other proteins, such as
casein, BSA, hemoglobin and gelatin. Currently, a large
proportion of the commercially available alkaline pepti-
dases are produced by Bacillus species because of their
high pH and temperature stability (Tari et al. 2006). Bac-
teria of the genus Bacillus, usually secrete two types of
extracellular peptidases, a neutral and an alkaline peptidase
(Park et al. 2004; Tang et al. 2004). Significant keratino-
lytic and proteolytic activity was observed for all three
species studied in this work and this activity responded to
changes in pH and temperature. The best pH was in the
7.0–11.0 range and the best temperature in the 40–70�C
range. The keratinase from B. cereus was had optimum
activity at pH 7.0 and 45�C (Sousa et al. 2007), whereas
keratinases from B. pseudofirmus FA30-01, B. lichenifor-
mis AP-1 and B. cereus MCM B-326 had optimum values
at pH between 9.0 and 11.0 and temperature from 50 to
70�C (Nilegaonkar et al. 2007; Tang et al. 2004; Kojima
et al. 2006).
In conclusion we have isolated and described three
keratinolytic Bacillus species that produced several pepti-
dases and keratinases with molecular masses between 15
and 200 kDa. These enzymes were active over a wide pH
and temperature range making them of interest in industrial
processes. High levels of simultaneous proteolytic and
keratinolytic activity from Bacilllus strains are new. The
fact they were isolated from feather waste, although not
necessarily surprising, reinforces their potential application
in processes using feather derivatives and all biotechno-
logical processes involving keratin hydrolysis.
Acknowledgments We would like to thank the technical assistance
of Denise da Rocha de Souza supported by fellowships grants from
MCT/CNPq. This study was supported by grants from Coordenacao
de Aperfeicoamento Pessoal de Nıvel Superior (CAPES), Conselho
Nacional de Desenvolvimento Cientıfico e Tecnologico (MCT/
CNPq), Conselho de Ensino para Graduados e Pesquisas (CEPG/
UFRJ), Fundacao Oswaldo Cruz (FIOCRUZ), Fundacao Carlos
Chagas Filho de Amparo a Pesquisa do Estado do Rio de Janeiro
(FAPERJ) and Fundacao Universitaria Jose Bonifacio (FUJB).
References
Balint B, Bagi Z, Toth A, Rakhely G, Perei K, Kovacs KL (2005)
Utilization of keratin-containing biowaste to produce biohydro-
gen. Appl Microbiol Biotechnol 69:404–410. doi:10.1007/
s00253-005-1993-3
Barone JR, Schmidt WF, Liebner CFE (2005) Compounding and
molding of polyethylene composites reinforced with keratin
feather fiber I. Compos Sci Technol 65:683–692. doi:10.1016/
j.compscitech.2004.09.030
Bernal C, Cairo J, Coello N (2006) Purification and characterization of
a novel exocellular keratinase form Kocuria rosea. Enzyme
Microb Technol 38:49–546. doi:10.1016/j.enzmictec.2005.02.021
World J Microbiol Biotechnol (2011) 27:1355–1365 1363
123

Bertsch A, Coello N (2005) A biotechnological process for treatment
and recycling poultry feathers as a feed ingredient. Bioresour
Technol 96:1703–1708. doi:10.1016/j.biortech.2004.12.026
Brandelli A (2008) Bacterial Keratinases: useful enzymes for
bioprocessing agroindustrial wastes and beyond. Food Bioproc-
ess Technol 1:105–116. doi:10.1007/s11947-007-0025-y
Cai C, Zheng X (2009) Medium optimization for keratinase
production in hair substrate by a new Bacillus subtilis KD-N2
using response surface methodology. J Ind Microbiol Biotechnol
36:875–883. doi:10.1007/s10295-009-0565-4
Cai CG, Chen JS, Qi JJ, Yin Y, Zheng XD (2008) Purification and
characterization of keratinase from a new Bacillus subtilis strain.
J Zhejiang Univ Sci B 9(9):713–720. doi:10.1631/jzus.B08
20128
Claus D, Berkeley RCW (1986) Genus Bacillus Cohn 1872. In:
Sneath PHA, Mair NS, Sharpe ME, Holt JG, Williams ST,
Sharpe ME, Holt JG (eds) Bergey’s manual of systematic
bacteriology, 2nd edn. Williams & Wilkins, Baltimore,
pp 1105–1141
Dalev P, Ivanov I, Liubomirova A (1997) Enzymic modification of
feather keratin hydrolysates with lysine aimed at increasing the
biological value. J Sci Food Agric 73:242–244. doi:10.1002/
(SICI)1097-0010(199702)73:2\242:AID-JSFA712[3.0.CO;2-3
Daroit DJ, Correa APF, Brandelli A (2009) Keratinolytic potential of
a novel Bacillus sp. P45 isolated from the Amazon basin fish
Piaractus mesopotamicus. Int Biodeterior Biodegradation
63:358–363. doi:10.1016/j.ibiod.2008.11.008
De Melo AC, Dornelas-Ribeiro M, Souza EP, Macrae A, Fracalanzza
SEL, Vermelho AB (2007) Peptidase profiles from non-albicansCandida spp. isolated from the blood of a patient with chronic
myeloid leukemia and another with sickle cell disease. FEMS
Yeast Res 7:1004–1012. doi:10.1111/j.1567-1364.2007.00269.x
El-Refai HA, AbdelNaby MA, Gaballa A, El-Araby MH, Abdel
Fattah AF (2005) Improvement of the newly isolated Bacilluspumilus FH9 keratinolytic activity. Process Biochem
40:2325–2332. doi:10.1016/j.procbio.2004.09.006
Fraser RDB, Parry DAD (2008) Molecular packing in the feather
keratin filament. J Struct Biol 162:1–13. doi:10.1016/j.jsb.2008.
01.011
Giongo JL, Lucas FS, Casarin F, Heeb P, Brandelli A (2007)
Keratinolytic proteases of Bacillus species isolated from the
Amazon basin showing remarkable de-hairing activity. World J
Microbiol Biotechnol 23:375–382. doi:10.1007/s11274-006-
9234-1
Gosh A, Chakrabarti K, Chattopadhyay D (2008) Degradation of raw
feather by a novel high molecular weight extracellular protease
from newly isolated Bacillus cereus DCUW. J Ind Microbiol
Biotechnol 35:825–834. doi:10.1007/s10295-008-0354-5
Grazziotin A, Pimentel FA, de Jong EV, Brandelli A (2006)
Nutritional improvement of feather protein by treatment with
microbial keratinase. Anim Feed Sci Technol 126:135–144.
doi:10.1016/j.anifeedsci.2005.06.002
Gupta R, Ramnani P (2006) Microbial keratinases and their prospec-
tive applications: an overview. Appl Microbiol Biotechnol
70:21–33. doi:10.1007/s00253-005-0239-8
Jones LB, Fontamini D, Jarvinen M, Pekkarinen A (1998) Simplified
endoproteinase assays using gelatin or azogelatin. Anal Biochem
263:214–220. doi:10.1006/abio.1998.2819
Khardenavis AA, Kapley A, Purohit HJ (2009) Processing of poultry
feathers by alkaline keratin hydrolyzing enzyme from Serratiasp. HPC 1383. Waste Manage 29:1409–1415. doi:10.1016/
j.wasman.2008.10.009
Kojima M, Kanai M, Tominaga M, Kitazume S, Inoue A, Horikoshi
K (2006) Isolation and characterization of a feather-degrading
enzyme from Bacillus pseudofirmus FA30–01. Extremophiles
10:229–235. doi:10.1007/s00792-005-0491-y
Laemmli VK (1970) Cleavage of structural proteins during the
assembly of the head of bacteriophage T4. Nature 227:680–685
Lin X, Lee CG, Casale ES, Shin JCH (1992) Purification and
characterization of a keratinase from a feather-degrading Bacilluslicheniformis PWD-1. Appl Environ Microbiol 58:3271–3275
Lopes BG, Santos AL, Bezerra CD, Wanke B, Dos Santos LM,
Nishikawa MM, Mazotto AM, Kussumi VM, Haido RM,
Vermelho AB (2008) A 25-kDa Serine Peptidase with Keratin-
olytic activity secreted by Coccidioides immitis. Mycopathol
166:35–40. doi:10.1007/s11046-008-9116-1
Lowry OH, Rosebrough NJ, Farr AL, Randall RJ (1951) Protein
measurement with the Folin phenol reagent. J Biol Chem
193:267–275
Mabrouk MEM (2008) Feather degradation by a new keratinolytic
Streptomyces sp. MS-2. World J Microbiol Biotechnol
24:2331–2338. doi:10.1007/s11274-008-9748-9
Macedo AJ, Beys da Silva WO, Termignoni C (2008) Properties of a
non collagen-degrading Bacillus subtilis keratinase. Can J
Microbiol 54:80–188
Massol-Deya AA, Odelson DA, Hickey RF, Tiedje JM (1995)
Bacterial community fingerprinting of amplified 16S and 16–23S
ribosomal DNA gene sequences and restriction endonuclease
analysis (ARDRA). In: Akkermans ADL (ed) Molecular micro-
bial ecology manual. Kluwer Academic Publishers, Dordrecht,
pp 3.3.2.1.1–3.3.2.1.8
Matsui T, Yamada Y, Mitsuya H, Shigeri Y, Yoshida Y, Saito Y,
Matsui H, Watanabe K (2009) Sustainable and practical
degradation of intact chicken feathers by cultivating a newly
isolated thermophilic Meiothermus ruber H328. Appl Microbiol
Biotechnol 82:941–950. doi:10.1007/s00253-009-1880-4
Mazotto AM, Cedrola SML, Lins U, Rosado AS, Silva KT, Chaves
JQ, Rabinovitch L, Zingali RB, Vermelho AB (2010) Keratin-
olytic activity of Bacillus subtilis AMR using human hair. Lett
Appl Microbiol. doi:10.1111/j.1472-765X.2009.02760.x
Mohorcic M, Torkar A, Friedrich J, Kristl J, Murdan S (2007) An
investigation into keratinolytic enzymes to enhance ungual drug
delivery. Int J Pharm 332:196–201. doi:10.1016/j.ijpharm.2006.
09.042
Nam GW, Lee D, Lee HS, Lee NJ, Kim BC, Choe EA, Hwang JK,
Suhartono MT, Pyun YR (2002) Native-feather degradation by
Fervidobacterium islandicum AW-1, a newly isolated keratin-
ase-producing thermophilic anaerobe. Arch Microbiol 178:
538–547. doi:10.1007/s00203-002-0489-0
Nilegaonkar SS, Zambare VP, Kanekar PP, Dhakephalkar PK,
Sarnaik SS (2007) Production and partial characterization of
dehairing protease from Bacillus cereus MCM B-326. Bioresour
Technol 98:1238–1245. doi:10.1016/j.biortech.2006.05.003
Odetallah NH, Wang JJ, Garlich JD, Shih JC (2005) Versazyme
supplementation of broiler diets improves market growth
performance. Poult Sci 84:858–864. doi:10.1002/9780470385
869.ch12
Onifade AA, Al-Sane NA, Al-Musallan AA, Al-Zarban S (1998) A
review: potential for biotechnological applications of keratin-
degrading microorganisms and their enzymes for nutritional
improvement of feathers and other keratins as livestock feed
resource. Bioresour Technol 66:1–11. doi:10.1016/S0960-
8524(98)00033-9
Papadopoulos MC (1986) The effect of enzymatic treatment on amino
acid content and nitrogen characteristics of feather meals. Anim
Feed Sci Technol 16:151–156. doi:10.1016/0377-8401(86)
90058-1
Park GT, Son HJ (2009) Keratinolytic activity of Bacillus megateriumF7–1, a feather-degrading mesophilic bacterium. Microbiol Res
164:478–485. doi:10.1016/j.micres.2007.02.004
Park CH, Sang JL, Lee SJ, Lee SG, Lee WS, Byun SM (2004) Hetero-
and autoprocessing of the extracellular metalloprotease (Mpr) in
1364 World J Microbiol Biotechnol (2011) 27:1355–1365
123

Bacillus subtilis. J Bacteriol 186:6457–6464. doi:10.1128/JB.
186.19.6457-6464.2004
Pillai P, Archana G (2008) Hide depilation and feather disintegration
studies with keratinolytic serine protease from a novel Bacillussubtilis isolate. Appl Microbiol Biotechnol 78:643–650.
doi:10.1007/s00253-008-1355-z
Rao MB, Tanksale AM, Ghatge MS, Deshpande VV (1998)
Molecular and biotechnological aspects of microbial proteases.
Microbiol Mol Biol Rev 62:597–635
Riffel A, Lucas F, Heeb P, Brandelli A (2003) Characterization of a
new keratinolytic bacterium that completely degrades native
feather keratin. Arch Microbiol 179:258–265. doi:10.1007/
s00203-003-0525-8
Rozs M, Manczinger L, Vagvolgyi C, Kevei F (2001) Secretion of a
trypsin-like thiol protease by a new keratinolytic strain of
Bacillus licheniformis. FEMS Microbiol Lett 205:221–224.
doi:10.1111/j.1574-6968.2001.tb10951.x
Saeki K, Ozaki K, Kobayashi T, Ito S (2007) Detergent Alkaline
Proteases: enzymatic properties, genes, and crystal structures.
J Biosci Bioeng 103:501–508. doi:10.1263/jbb.103.501
Son H, Park H, Kim H, Lee C (2008) Nutritional regulation of
keratinolytic activity in Bacillus pumilis. Biotechnol Lett
30:461–465. doi:10.1007/s10529-007-9567-3
Sousa F, Jus S, Erbel A, Kokol V, Cavaco-Paulo A, Gubitz GM
(2007) A novel metalloprotease from Bacillus cereus for protein
fiber processing. Enzyme Microb Technol 40:1772–1781.
doi:10.1016/j.enzmictec.2006.12.017
Suh HJ, Lee HK (2001) Characterization of a keratinolytic serine
protease from Bacillus subtilis KS-1. Protein Chem 20:165–169.
doi:10.1023/A:1011075707553
Suzuki Y, Tsujimoto Y, Matsui H, Watanabe K (2006) Decomposition
of extremely hard-to-degrade animal proteins by thermophilic
bacteria. J Biosci Bioeng 102:73–81. doi:10.1263/jbb.102.73
Tang X, Lakay FM, ShenW, Shao W, Fang H, Prior BA, WangZ, Zhuge J
(2004) Purification and characterization of an alkaline protease used
in tannery industry from Bacillus licheniformis. Biotechol Lett
26:1421–1424. doi:10.1023/B:BILE.0000045642.19299.3f
Tari C, Genckal H, Tokatli F (2006) Optimization of a growth
medium using a statistical approach for the production of an
alkaline protease from newly isolated Bacillus sp. L21. Proc
Biochem 41:659–665. doi:10.1016/j.procbio.2005.08.012
Thys RCS, Brandelli A (2006) Purification and properties of a
keratinolytic metalloprotease from Microbacterium sp. J Appl
Microbiol 101:1259–1268. doi:10.1111/j.1365-2672.2006.03050.x
Vasconcelos FJM, Rabinovitch L (1994) A new formula for an
alternative culture medium, without antibiotics, for isolation and
presumptive quantification of Bacillus cereus food. J Food
Microbiol 58:235–238
Vermelho AB, Mazotto AM, Nogueira de Melo AC, Vieira FHC,
Duarte TR, Macrae A, Nishikawa MM, Bon EPS (2009)
Identification of a Candida parapsilosis strain producing extra-
cellular serine peptidase with keratinolytic activity. Mycopathol.
doi:10.1007/s11046-009-9231-7
Wawrzkiewicz K, Wolski T, Lobarewski J (1991) Screening the
keratinolytic activity of dermatophytes in vitro. Mycopathologia
114:1–8. doi:10.1007/BF00436684
Williams CM, Richter CS, Mackeinze JM Jr, Shih JCH (1990)
Isolation, identification, and characterization of a feather-
degrading bacterium. Appl Environ Microbiol 56:1509–1515
Yoshioka M, Miwa T, Horii H, Takata M, Nishizawa K, Watanabe M,
Shinagawa M, Murayama Y (2007) Characterization of a
proteolytic enzyme derived from Bacillus strain that effectively
degrades prion protein. J Appl Microbiol 102:509–515. doi:
10.1111/j.1365-2672.2006.03080.x
World J Microbiol Biotechnol (2011) 27:1355–1365 1365
123
Recommended

















