
Behavioral Neuroscience 1987, Vol. 101, No.4, 576-586
Copyright 1987 by the American Psychological Association, Inc. 0735-7044/87/$00.75
Effect of Naloxone on Conditioned Suppression in Rats
Michael Vigorito Brooklyn College of CUNY
John J. B. Ayres University of Massachusetts, Amherst
Painful stimuli are known to engage an endorphin analgesic system that can be reversed by the opiate antagonist, naloxone. Naloxone, then, should increase the effectiveness of aversive unconditioned stimuli (USs) in Pavlovian fear conditioning. Consistent with this hypothesis, naloxone administered during the acquisition of conditioned suppression in rats enhanced posttrial suppression and preconditioned stimulus (pre-CS; context-controlled) suppression. Furthermore, it enhanced CS-elicited suppression during extinction when administered during acquisition but not when administered only during extinction. Thus naloxone does not enhance an already existing fear nor enhance the memory of previous conditioning; instead, it enhances the conditioning of fear presumably by making the aversive US more painful.
Over the past several years, numerous studies have demonstrated that the endogenous opiates play an important role in the modulation of pain (see Basbaum & Fields, 1978; Sherman & Liebeskind, 1980, for reviews). This finding has generated an interest in research seeking to determine the role that these substances play in aversive learning (e.g., Bolles & Fanselow, 1982; Riley, Zellner, & Duncan, 1980). Recent theoretical models of fear and fear conditioning have incorporated an endogenous analgesic process as an integral part (Bolles & Fanselow, 1980; Fanselow, 1981; Schull, 1979). In the perceptual-defensive-recuperative (PDR) model of fear and pain (Bolles & Fanselow, 1980), for example, the anticipation of an aversive event is assumed to produce fear. Fear is described as a central motive state that organizes an animal's species-specific-defense reactions (Bolles, 1970) and inhibits other motivational systems. Pain, also a motivational system, has tissue insult as its antecedent condition and leads to recuperative behavior. Because recuperative behavior competes with defensive behavior, fear prevents the occurrence of recuperative behavior by inhibiting pain. Fanselow and Bolles ( 1979a) proposed that fear inhibits pain by triggering an endorphin analgesic process. Consistent with this hypothesis are several studies that suggest that fear causes analgesia (e.g., Chance, 1980; Chance, Krynock, & Rosecrans, 1978; Fanselow & Baackes, 1982; Fanselow & Bolles, 1979a; MacLennan, Jackson, & Maier, 1980) and that conditioned fear increases brain endorphin activity (Chance, White, Krynock, & Rosecrans, 1978; Scallet, 1982).
The conditioned opponent-process theory (Schull, 1979) also predicts that fear produces analgesia. This model combines certain features of Solomon and Corbit's (1974) oppo-
This article is based on a thesis submitted by Michael Vigorito in partial fulfillment of the requirements for the Master of Science degree at the University of Massachusetts.
We would like to thank Jerrold S. Meyer, John W. Moore, and Michael S. Fanselow for their valuable comments.
Correspondence concerning this article should be addressed to Michael Vigorito, Department of Psychology, Brooklyn College of CUNY, Brooklyn, New York 11210 or to John J. B. Ayres, Department of Psychology, University of Massachusetts, Amherst, Massachusetts 01003.
576
nent-process theory with the theory of conditioning formulated by Rescorla and Wagner (1972). The Rescorla-Wagner model is formally stated by the equation
~V= K(A- V).
Here there are two parameters that govern the change in associative strength (V) that accrues to a conditioned stimulus (CS). The constant, K, is a learning rate parameter associated with the CS and the unconditioned stimulus (US). The asymptotic level of associative strength that the US can support is represented by A, which increases with US intensity. The equation indicates that the change in conditioned strength during a trial increases as a function of the discrepancy between A and V.
Schull equated the primary a process, which is elicited by the US, with A. He further suggested that the b process is a compensatory conditioned response (CR) that follows the same Pavlovian laws as other CRs. The conditioned opponent b process he equated with the present conditioned strength of the CS paired with the US (i.e., V). Schull suggested that in aversive conditioning situations, the conditioned b process may be the conditioned release of endorphins. Thus, with repeated trials, increments in the strength of anticipatory CRs grow smaller because of the increased release of endorphins (increasing V) which reduces the discrepancy, A- V.
More recently, Fanselow ( 1981) suggested an alternative mechanism that does not require postulating opponent processes. This mechanism is also based on Bolles and Fanselow's ( 1980) assumption that fear triggers an endorphin analgesic system. Thus, during repeated fear conditioning trials using electric shock USs, the A - V discrepancy decreases as a result of increasing V and decreasing A. Increasing V reflects conditioned release of endorphin. Because the endorphin activity reduces the perceived intensity of the shock, A decreases. Since endorphin analgesia is assumed to be a function of fear, the reduction ofA should also be a function ofV.
These theoretical models predict that fear and analgesia are closely linked; any procedure that produces fear should also produce concurrent analgesia. Support for this idea was provided by Fanselow and Baackes (1982; but see Davis & Hendersen, 1985). Fanselow and Baackes reported that fear

NALOXONE AND CONDITIONED SUPPRESSION 577
conditioned to apparatus cues (as measured by freezing) inhibited the rats' paw-licking and paw-lifting responses to formalin injected into the paw. Moreover, because this fearinduced analgesia was reversed by the opiate antagonist naltrexone, an opiate analgesia system was suggested.
These theoretical models suggest that an endogenous opiate analgesic system is activated during Pavlovian fear conditioning and that this system influences the amount oflearning on a given trial.
Because a tightly coupled fear-pain system is a critical part of such models, it seemed feasible to examine the effect of naloxone, a pure opiate antagonist, on conditioned suppression. The conditioned suppression procedure is a popular pamdigm for studying fear conditioning. Administration of naloxone during the acquisition of conditioned suppression should in general enhance the aversiveness of the US. This in tum would enhance fear conditioning to the CS, as well as to background cues, and increase fear of the CS during the extinction of that conditioned fear.
Many investigators have observed nonmonotonicities in the acquisition of conditioned suppression. Within-sessions decrements in suppression (WSDs), for example, are observed under maintained acquisition conditions and are defined by a greater CR on the first reinforced trial of a session than on other reinforced trials of the same session (Ayres, BergerGross, Kohler, Mahoney, & Stone, 1979). An interesting aspect of WSDs is the recovery of the CR on the first trial of a session following a session with decrements. This betweensessions recovery resembles spontaneous recovery that is seen in extinction procedures. A diminution in the CR when CSUS pairings continue to be presented after asymptote has been reached has also been reported by several authors (Goldstein, 1960; Kimmel & Bums, 1975; Libby, 1951; Millenson & Dent, 1971; Overmier, Payne, Brackbill, Linder, & Lawry, 1979). This postasymptomatic performance decrement (PAD) has been attributed to adaptation-like effects-the organism adapts to the US, which therefore loses its ability to maintain a CR. It is possible that an endorphin analgesic system creates WSDs and PADs by reducing the perceived intensity of the US. Thus WSDs may reflect a within-sessions decrease in shock sensitivity that recovers overnight; and PADs may reflect a longer lasting decrease in nociception that does not recover. In Experiment 1, we looked for both WSDs and PADs, seeking to determine whether naloxone had any effect on such nonmonotonicities.
Experiment 1
In Experiment 1, naloxone was administered to mts during the acquisition of conditioned suppression. This drug increases the reaction to electric shock (e.g., Fanselow, 1984a, 1984b; Grau, 1984; Kaplan & Glick, 1979) and has also been observed to reverse fear-induced analgesia (Fanselow & Bolles, 1979a). Presumably, naloxone produces these effects by blocking the endorphin analgesic system. If naloxone increases the aversiveness of shock USs, then rats given the drug during acquisition should show increased conditioned
suppression to the CS and to background cues and should show more suppression during extinction of that suppression than should rats treated with saline (cf. Annau & Kamin, 1961).
It is common in studies of conditioned suppression for ceiling effects to obscure group differences in CS-elicited suppression during acquisition. Annau and Kamin ( 1961 ), for example, could not detect differences during acquisition between groups that received shock intensities ranging from 0.85 rnA to 2.91 rnA. Group differences, however, were readily apparent in a subsequent extinction test. By weakening suppression, extinction moves suppression ratios away from the arbitrary ceiling of 0 (complete suppression) and allows differences among groups to be detected. Because naloxone is postulated here to make the shock US more aversive, it would not be particularly surprising if group differences in CSelicited suppression during acquisition could not be detected, but we would certainly expect such differences to appear in extinction.
There is, however, another measure that might be sensitive to group differences during acquisition, and that is a measure of posttrial suppression. On the basis of recent evidence (Ayres, Axelrod, Mercker, Muchnik, & Vigorito, 1985; Ayres & Vigorito, 1984), we believe that posttrial suppression can reflect CS-elicited effects and that it is less constrained by ceiling effects than is the usual measure of barpress suppression during the CS. The evidence that posttrial suppression is at least in part a measure of CS-elicited effects is as follows. First, when an expected shock US is omitted, specific freezing postures initiated during the CS are often maintained into a 1-min posttrial interval. Second, on nonreinforced trials, there is a significant correlation between measures of CS and posttrial freezing and between CS freezing and posttrial barpress suppression. Third, CSs in different modalities may elicit different degrees of freezing even though they have received identical conditioning treatments. In such a case, the CS that elicits more freezing also produces more posttrial freezing and barpress suppression, at least on nonreinforced trials. The role of the shock US on reinforced trials appears to be to disrupt freezing initiated during the CS (cf. Bolles & Riley, 1973; Fanselow, 1982), thereby permitting barpressing to resume posttrial. We can take advantage of this possibility. Two groups might freeze enough during the CS to produce complete barpress suppression; yet Group 1 may be more completely frozen than Group 2. If so, then Group 1 's freezing would presumably be harder for the US to disrupt, and this would result in more posttrial barpress suppression in Group l.
Pain reactivity was also measured during conditioning. On odd-numbered Pavlovian conditioning days, each group was given tail-flick tests just before and just after their session. The tail-flick test (Dewey & Harris, 1975) is widely used for assessing nociceptive changes in animals. If the endorphins are activated within a session (possibly producing WSDs), then tail-flick latencies should be longer after a conditioning session than before it. Similarly, if the tonic level of endorphins increases with each session (possibly producing PADs), then tail-flick latencies should be longer after many days of conditioning than prior to those days.

578 MICHAEL VIGORITO AND JOHN J. B. AYRES
Method
Subjects
The subjects were 24 experimentally naive male albino Holtzman rats that were 90 days of age at the start of the experiment. All were kept at 80% of their free-feeding weights and were housed individually in a continuously lighted room. Tap water was available in the home cage at all times.
Apparatus
Eight identical Gerbrands operant conditioning chambers, housed in ventilated plywood cubes lined with acoustical tile, served as the conditioning apparatus. The inside dimensions of each chamber were 23.2 x 20.3 x 19.5 em; the side walls and top were transparent Plexiglas, and the end walls were aluminum. Each chamber had a standard Gerbrands lever, 1.5 x 5.0 em, centrally mounted on one end wall about 8 em above the grid floor. A dipper receptacle, located in the lower left corner of the same wall, measured 5.5 x 5.0 X 5.0 em. The dipper was used to present reinforcement for barpressinga 4-s presentation ofO.l-ml dipper cup containing 32% sucrose (by weight). The floor was composed of 18 stainless steel rods, 2 mm in diameter, mounted 1.3 em apart center to center. An 85-dB (SPL) masking white noise was transmitted through a 10-cm-diameter speaker mounted on the lid of each chamber.
A CS was provided by illuminating (against a normally dark background) a 7.5-W, 110-V ac frosted bulb mounted on the rear wall of the cubicle that housed the conditioning chambers. A large sheet of aluminum foil behind this bulb reflected its light through the Plexiglas walls. The US was a 1-s, !-rnA scrambled shock delivered through the grid floors and was provided by eight Grason-Stadler shock sources (Models E1064GS and 700).
The tail-flick apparatus consisted of a wooden stage with a shallow groove cut along its center. A 300-W General Electric projector lamp was mounted above the stage. A condenser lens about 8 em above the stage and about 20 em below the light focused the light on the tail. The rat was placed in a plastic restrainer and positioned so that the tail rested on the groove. A switch turned on the light source and timer. When the heat became too intense, the rat deflected its tail and activated a photocell located in the groove. This action terminated the trial. The distance between the light source and stage was adjusted to produce control latencies of 6-7 s.
Procedure
A conditioned suppression procedure was used; suppression of barpressing for sucrose reward served to index the strength of conditioned fear. All rats were shaped to barpress for sucrose on Days I through 4. On Days 5 through 10, responding was reinforced on a variable interval (VI) 2-min schedule, and each session was 30 min in duration.
Acquisition. Rats were assigned to three groups matched for weight and baseline response rates. For Groups PC-N (n = 8) and PC-S (n = 7), the next 16 days were Pavlovian conditioning days; the rats received four CS-US pairings each day while barpressing on a VI 2-min schedule. For both groups, a 105-s light CS terminated with the onset of a 1-s, 1-mA shock US. The session length was 30 min with CSs occurring 4, 10, 18, and 25 min into the session. A third group, C-S (n = 8), received exactly the same procedure except that all USs were omitted. This CS-alone group was designed to serve as a tailflick control during Pavlovian conditioning.
The strategy for drug administration was to give Group PC-N an 8 mg/kg ip injection of naloxone hydrochloride immediately before the first session that followed a session with a WSD. Groups PC-S and C-S were injected ip with an equal volume of saline immediately before their session on the same day. The groups continued to receive their respective drug treatments immediately before each session for the rest of the Pavlovian conditioning phase. Thus the first WSD in Group PC-N marked the start of drug administration for all groups.
The tail-flick tests were conducted for all groups on odd-numbered days throughout all phases. All tests occurred (with the rats restrained) just before drug injection and immediately after a barpress session. In each such test, each rat received three tail-flick trials with an intertrial interval (ITI) of about 1 min. During the ITI, the rat remained in the plastic restrainer.
Baseline recovery. After the last day of Pavlovian conditioning, Groups PC-N and PC-S received eight daily 30-min sessions of barpressing on the VI 2-min schedule in the absence of CSs or USs. The aim was to allow Group PC-N to recover its baseline rate, which was depressed during acquisition. Both groups continued receiving drug injections during this training.
Extinction. After 8 days of recovery for Groups PC-N and PC-S, both groups underwent 5 days of extinction to the CS. Extinction was identical to acquisition except that shock USs were omitted. During extinction, Groups PC-N and PC-S continued to receive injections of naloxone and saline, respectively. Drug treatment was continued to prevent possible generalization decrements due to drug-dependent learning (Overton, 1968).
Measures. Suppression to the CS was indexed using the AnnauKamin (1961) suppression ratio, D/(B + D). Here D denotes the response rate during the CS, and B denotes the rate during a 2-min period immediately before the CS (i.e., pre-CS rate). With this ratio, a score of 0 reflects complete suppression to the CS; a score of I to 0.5 reflects no suppression. Note, however, that observed WSDs and PADs can be interpreted as computational artifacts if the pre-CS rates fall across trials or sessions. If the CS rates were to remain constant as the pre-CS rates dropped, the suppression ratio would increase and be misinterpreted as reflecting decreasing suppression to the CS. To avoid this computational artifact when looking for WSDs and PADs, we computed "corrected" suppression ratios by using each rat's preCS score on Trial 1 of the first conditioning day as the baseline rate for all trials. Because the pre-CS rate would then be the same for a given rat on every trial, WSDs and PADs could not be computational artifacts of falling pre-CS rates. Suppression during a 1-min period following the US was also measured, using the ratio 2A/(2A + B); here A denotes the number of responses in the !-min posttrial period, and B denotes the number in the 2-min interval before the CS.
Results
On the last day of VI training, Groups PC-N, PC-S, and CS barpressed at mean rates of 14, 14, and 13 responses per min, respectively. These rates did not differ significantly. (In all subsequent analyses, 1 rat from Group PC-N was dropped because it failed to barpress throughout acquisition and extinction.)
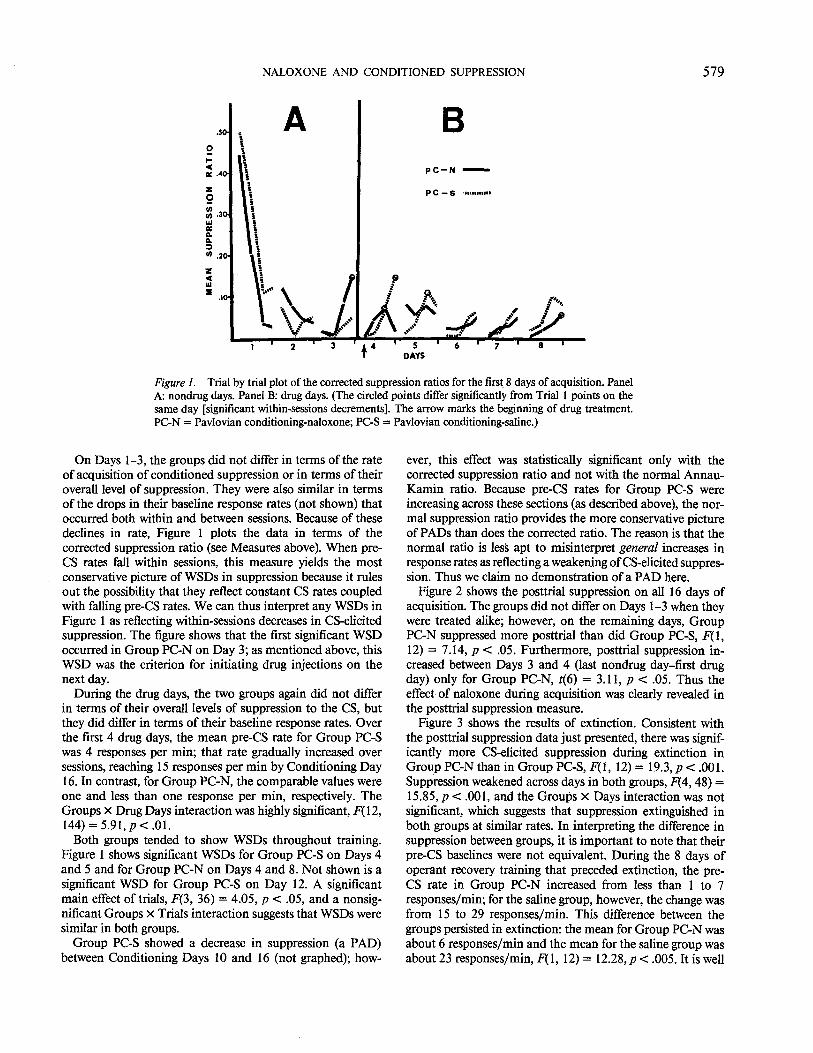
Figure 1 plots the Trial x Trial acquisition of conditioned suppression in Groups PC-N and PC-S over the first 8 of the 16 days of acquisition. Days 1-3 were nondrug days, on which the groups were treated alike. Days 4-16 were drug days, on which Group PC-N received injections of naloxone before each session, whereas Group PC-S received injections of saline.
NALOXONE AND CONDITIONED SUPPRESSION 579
z c w li .1
A
2 3 5 DAYS
B PC-N
PC-S
6
Figure 1. Trial by trial plot of the corrected suppression ratios for the first 8 days of acquisition. Panel A: nondrug days. Panel B: drug days. (The circled points differ significantly from Trial I points on the same day [significant within-sessions decrements]. The arrow marks the beginning of drug treatment. PC-N =Pavlovian conditioning-naloxone; PC-S =Pavlovian conditioning-saline.)
On Days 1-3, the groups did not differ in terms of the rate of acquisition of conditioned suppression or in terms of their overall level of suppression. They were also similar in terms of the drops in their baseline response rates (not shown) that occurred both within and between sessions. Because of these declines in rate, Figure 1 plots the data in terms of the corrected suppression ratio (see Measures above). When preCS rates fall within sessions, this measure yields the most conservative picture of WSDs in suppression because it rules out the possibility that they reflect constant CS rates coupled with falling pre-CS rates. We can thus interpret any WSDs in Figure 1 as reflecting within-sessions decreases in CS-elicited suppression. The figure shows that the first significant WSD occurred in Group PC-N on Day 3; as mentioned above, this WSD was the criterion for initiating drug injections on the next day.
During the drug days, the two groups again did not differ in terms of their overall levels of suppression to the CS, but they did differ in terms of their baseline response rates. Over the first 4 drug days, the mean pre-CS rate for Group PC-S was 4 responses per min; that rate gradually increased over sessions, reaching 15 responses per min by Conditioning Day 16. In contrast, for Group PC-N, the comparable values were one and less than one response per min, respectively. The Groups x Drug Days interaction was highly significant, F(12, 144) = 5.91, p < .01.
Both groups tended to show WSDs throughout training. Figure 1 shows significant WSDs for Group PC-S on Days 4 and 5 and for Group PC-N on Days 4 and 8. Not shown is a significant WSD for Group PC-S on Day 12. A significant main effect of trials, F(3, 36) = 4.05, p < .05, and a nonsignificant Groups X Trials interaction suggests that WSDs were similar in both groups.
Group PC-S showed a decrease in suppression (a PAD) between Conditioning Days 10 and 16 (not graphed); how-
ever, this effect was statistically significant only with the corrected suppression ratio and not with the normal AnnauKamin ratio. Because pre-CS rates for Group PC-S were increasing across these sections (as described above), the normal suppression ratio provides the more conservative picture of PADs than does the corrected ratio. The reason is that the normal ratio is less apt to misinterpret general increases in response rates as reflecting a weakening of CS-elicited suppression. Thus we claim no demonstration of a PAD here.
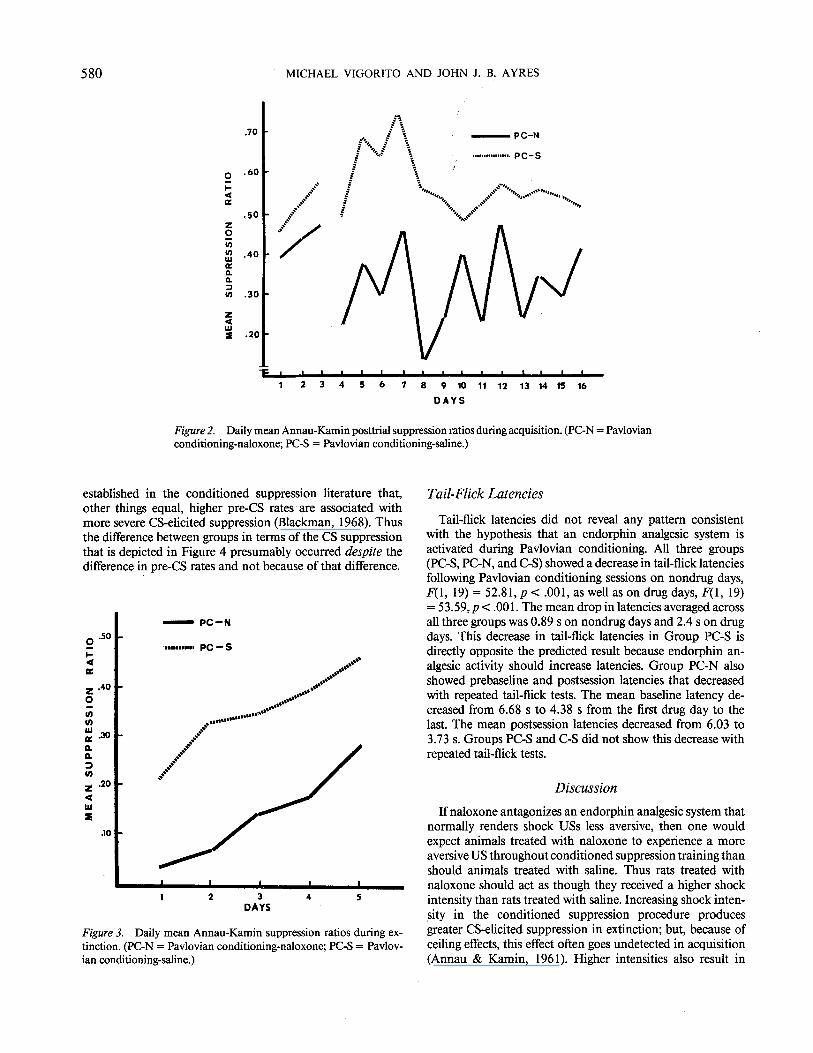
Figure 2 shows the posttrial suppression on all 16 days of acquisition. The groups did not differ on Days 1-3 when they were treated alike; however, on the remaining days, Group PC-N suppressed more posttrial than did Group PC-S, F(1, 12) = 7.14, p < .05. Furthermore, posttrial suppression increased between Days 3 and 4 (last nondrug day-first drug day) only for Group PC-N, t(6) = 3.11, p < .05. Thus the effect of naloxone during acquisition was clearly revealed in the posttrial suppression measure.
Figure 3 shows the results of extinction. Consistent with the posttrial suppression data just presented, there was significantly more CS-elicited suppression during extinction in Group PC-N than in Group PC-S, F(1, 12) = 19.3, p < .001. Suppression weakened across days in both groups, F(4, 48) = 15.85, p < .001, and the Groups X Days interaction was not significant, which suggests that suppression extinguished in both groups at similar rates. In interpreting the difference in suppression between groups, it is important to note that their pre-CS baselines were not equivalent. During the 8 days of operant recovery training that preceded extinction, the preCS rate in Group PC-N increased from less than 1 to 7 responses/min; for the saline group, however, the change was from 15 to 29 responses/min. This difference between the groups persisted in extinction: the mean for Group PC-N was about 6 responses/min and the mean for the saline group was about 23 responses/min, F(l, 12) = 12.28, p < .005. It is well
580 MICHAEL VIGORITO AND JOHN J. B. AYRES
.70
0 .60
.... ........
< .. .. a: .. ..
.50 •..••. ~····/ z 0 ~·
VI / VI .40 w a: Q. Q. :::J VI .30
z < w ::E .20
2 3 4 5 6 7 8 9 10 11 12 13 14 t5 16
DAYS
Figure 2. Daily mean Annau-Kamin posttrial suppression ratios during acquisition. (PC-N =Pavlovian conditioning-naloxone; PC-S =Pavlovian conditioning-saline.)
established in the conditioned suppression literature that, other things equal, higher pre-CS rates ·are associated with more severe CS-elicited suppression (Blackman, 1968). Thus the difference between groups in terms of the CS suppression that is depicted in Figure 4 presumably occurred despite the difference in pre-CS rates and not because of that difference.
0.50
.... < a:: z .40 0 Cl) Cl)
: .30 a. a. ::1 Cl)
z .20
< w :I
.10
-PC-N
..... , .. ,,,. PC-S . .. ~····" •.. .......
·····•·••· ········· ~·············"''"
...
.. .. .. .. •. .. .. .. .. .. .• .. ..
2 3 DAYS
4 5
Figure 3. Daily mean Annau-Kamin suppression ratios during extinction. (PC-N = Pavlovian conditioning-naloxone; PC-S = Pavlovian conditioning-saline.)
Tail-Flick Latencies
Tail-flick latencies did not reveal any pattern consistent with the hypothesis that an endorphin analgesic system is activated during Pavlovian conditioning. All three groups (PC-S, PC-N, and C-S) showed a decrease in tail-flick latencies following Pavlovian conditioning sessions on nondrug days, F(1, 19) = 52.81, p < .001, as well as on drug days, F(l, 19) = 53.59,p < .001. The mean drop in latencies averaged across all three groups was 0.89 son nondrug days and 2.4 son drug days. This decrease in tail-flick latencies in Group PC-S is directly opposite the predicted result because endorphin analgesic activity should increase latencies. Group PC-N also showed prebaseline and postsession latencies that decreased with repeated tail-flick tests. The mean baseline latency decreased from 6.68 s to 4.38 s from the first drug day to the last. The mean postsession latencies decreased from 6.03 to 3.73 s. Groups PC-S and C-S did not show this decrease with repeated tail-flick tests .
Discussion
If naloxone antagonizes an endorphin analgesic system that normally renders shock USs less aversive, then one would expect animals treated with naloxone to experience a more aversive US throughout conditioned suppression training than should animals treated with saline. Thus rats treated with naloxone should act as though they received a higher shock intensity than rats treated with saline. Increasing shock intensity in the conditioned suppression procedure produces greater CS-elicited suppression in extinction; but, because of ceiling effects, this effect often goes undetected in acquisition (Annau & Kamin, 1961 ). Higher intensities also result in

NALOXONE AND CONDITIONED SUPPRESSION 581
more depressed pre-CS baseline rates (Annau & Kamin, 1961 ), presumably reflecting stronger conditioning to the ever-present context or apparatus cues. If WSDs in suppression and PADs are due to within- and between-sessions increases in endogenous opiates, respectively, then naloxone should reduce these WSDs and PADs.
The present results are consistent with many of these expectations. Naloxone did enhance CS-elicited suppression in extinction, and it did result in lower pre-CS rates, suggesting enhanced contextual conditioning. Although naloxone did not enhance suppression during the CS in acquisition, it did enhance posttrial suppression. We have suggested that posttrial suppression is at least in part a measure of CS-elicited effects and that because it is relatively weak, it may be less constrained by ceiling effects than is the more conventional measure of CS-elicited suppression. Also, our finding that naloxone enhances posttrial suppression is consonant with the observation that naloxone increases the active and inactive component of the postshock reaction, that is, the postshock burst followed by postshock freezing (Fanselow, 1984a). Finally, with regard to WSDs and PADs, naloxone did not seem to reduce WSDs, and we were unable to convincingly demonstrate a PAD for it to affect.
If shock engages an endorphin analgesic system, then nondrugged rats might be expected to show reduced pain sensitivity following conditioned suppression sessions. Naloxone should mitigate that effect. The tail-flick tests, however, did not find evidence consistent with these predictions. One reason may be that the tail-flick measure may be insensitive to the small changes that would arise in a mildly stressful situation. Another possibility is that the tail-flick test was conducted outside the training context. Recent evidence suggests that without the presence of some mediating associative cues, analgesia can be lost (Fanselow, 1984a). The failure to detect hypoalgesia may also be due to "nociceptive learning" (Ramabadran, Guillan, & Jacob, 1979). Nociceptive learning refers to a phenomenon in which, upon reexposure to a painful stimulus such as heat to the paws from a hot plate, the animal shows an accelerated response to the source; that is, the animal learns to respond to the source of the pain by decreasing its latency to respond. Because in the present experiment rats were repeatedly exposed to the tail-flick test, nociceptive learning may have masked any increase in hypoalgesia. Moreover, it has been shown that naloxone treatment can facilitate nociceptive learning (Ramabadran et al., 1979). The fact that Group PC-N, but not PC-S and C-S, showed decreased baseline and postsession tail-flick latencies with repeated testing is consistent with the nociceptive learning hypothesis.
Experiment 2
Experiment 1 found that naloxone, when administered during the acquisition of conditioned suppression, acts like an increase in shock US intensity. That is, the drug depresses pre-CS barpress rates, enhances post-US suppression, and enhances CS-elicited suppression during extinction. Because naloxone, as predicted, significantly reduces pre-CS barpress rates, it is difficult, however, to interpret that effect without
knowing the effects of naloxone in situations unlikely to condition fear to contextual cues. Perhaps naloxone simply produces a motor impairment or reduces motivation for food rewards. Current research suggests that endogenous opioid peptides play an important role in the regulation of ingestive behavior (e.g., Morley, Levine, Yim, & Lowy, 1983; Reid, 1985). An important component of this research is the suppression of feeding and drinking by naloxone under a variety of conditions including food and water deprivation (e.g., Brown & Holtzman, 1979; Rodgers, Frank, Taylor, & Liebeskind, 1978) and consumption of highly palatable substances under undeprived and deprived conditions (e.g., Sclafani, Aravich, & Xenakis, 1982).
Experiment 2 was designed, therefore, to assess the effects of naloxone on barpress responding in the absence of shock USs. Barpressing was reinforced with sucrose presented on a VIJ.:.min schedule-the same schedule and the same reinforcer used in Experiment 1.
Method
Subjects and Apparatus
The subjects were 32 male albino Holtzman rats, 120 days old at the start of the experiment, and maintained as before. These rats had been used in a prior experiment in which they received 17 unsignaled 1.0-mA shocks in a shuttlebox. They had also been trained to lick sucrose from a spout in the shuttlebox. The apparatus was unchanged except for the elimination of the light CS and shock US.
Procedure
After an initial session of magazine training, all rats were shaped to barpress for sucrose over the next 3 days. Each response produced a 4-s presentation of sucrose, and each daily session ended after the rat had received 90 presentations. On the next day, responding was reinforced on a VI !-min schedule. The schedule was then increased to VI 2 and so remained throughout the study. Daily sessions were 30 min. On Day 7 of VI training, the rats were divided into two groups (n = 16) matched for response rate and body weight. The naloxone group received 8 mgfkg injections of naloxone just before training on Days 7, 8, and 9. The saline group received an equal volume of saline on the same days. No drugs were given on the next 2 days. On these days the rats received the usual 30-min session of barpressing. However, immediately before each session on the following 2 days, Days 12 and 13, the naloxone group received 4 mg/kg of naloxone, and the saline group received an equal volume of saline.
Results
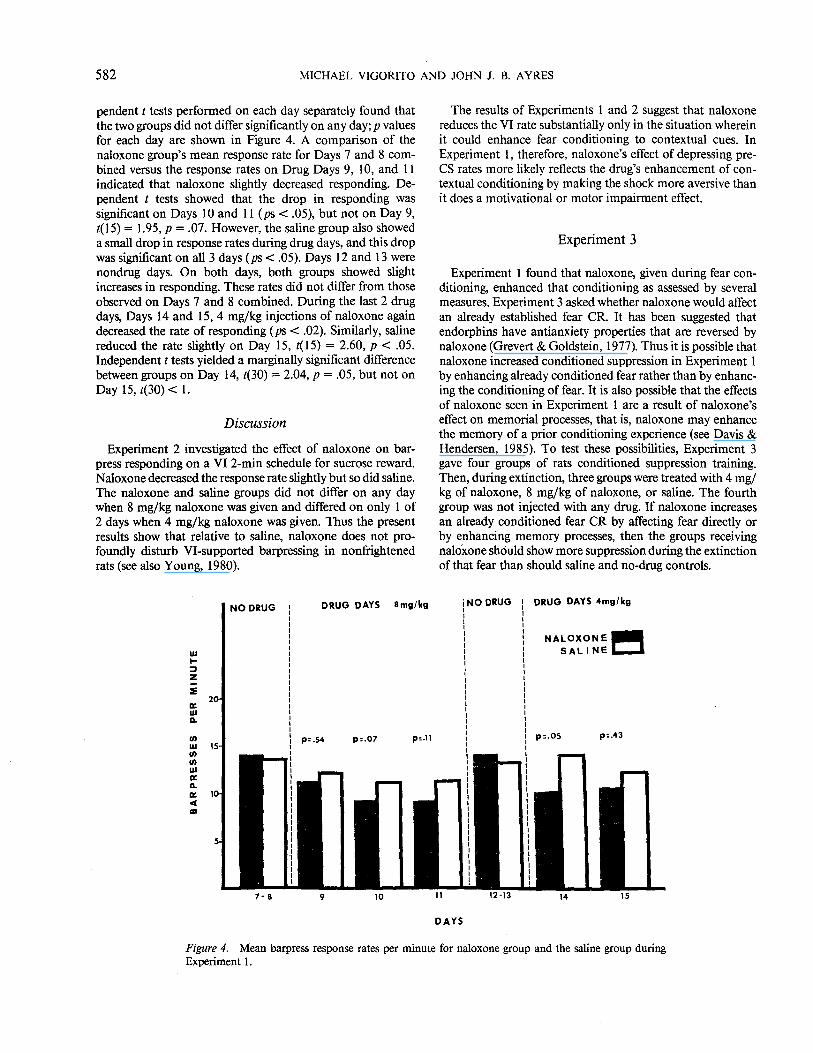
Figure 4 shows the rate of barpressing during the 7th through 15th day of the VI 2-min schedule. The figure is divided into four parts by dashed lines. The first part shows the mean responses per min for Days 7 and 8 combined for the naloxone and saline groups. No drugs were given on these days. As can be seen in the figure, both groups responded at about the same rate, t(30) < 1. The next part of the figure shows the response rate for both groups on Days 9, 10, and 11. Group naloxone received 8 mg/kg of naloxone on these days, and the saline group received saline injections. Inde-
582 MICHAEL VIGORITO AND JOHN J. B. AYRES
pendent t tests performed on each day separately found that the two groups did not differ significantly on any day; p values for each day are shown in Figure 4. A comparison of the naloxone group's mean response rate for Days 7 and 8 combined versus the response rates on Drug Days 9, 10, and 11 indicated that naloxone slightly decreased responding. Dependent t tests showed that the drop in responding was significant on Days 10 and 11 (ps < .05), but not on Day 9, t(l5) = 1.95, p = .07. However, the saline group also showed a small drop in response rates during drug days, and this drop was significant on all3 days (ps < .05). Days 12 and 13 were nondrug days. On both days, both groups showed slight increases in responding. These rates did not differ from those observed on Days 7 and 8 combined. During the last 2 drug days, Days 14 and 15, 4 mg/kg injections of naloxone again decreased the rate of responding (ps < .02). Similarly, saline reduced the rate slightly on Day 15, t(15) = 2.60, p < .05. Independent t tests yielded a marginally significant difference between groups on Day 14, t(30) = 2.04, p = .05, but not on Day 15, t(30) < 1.
Discussion
Experiment 2 investigated the effect of naloxone on barpress responding on a VI 2-min schedule for sucrose reward. Naloxone decreased the response rate slightly but so did saline. The naloxone and saline groups did not differ on any day when 8 mg/kg naloxone was given and differed on only 1 of 2 days when 4 mg/kg naloxone was given. Thus the present results show that relative to saline, naloxone does not profoundly disturb VI-supported barpressing in nonfrightened rats (see also Young, 1980).
Ul 1-::J z :::E
a: Ul D.
Ul Ul I Ul Ul Ul a: D. a:
"" ID
NO DRUG DRUG DAYS Bmg/kg
p:.S4 p:.07 p:.ll
The results of Experiments 1 and 2 suggest that naloxone reduces the VI rate substantially only in the situation wherein it could enhance fear conditioning to contextual cues. In Experiment 1, therefore, naloxone's effect of depressing preCS rates more likely reflects the drug's enhancement of contextual conditioning by making the shock more aversive than it does a motivational or motor impairment effect.
Experiment 3
Experiment 1 found that naloxone, given during fear conditioning, enhanced that conditioning as assessed by several measures. Experiment 3 asked whether naloxone would affect an already established fear CR. It has been suggested that endorphins have antianxiety properties that are reversed by naloxone (Grevert & Goldstein, 1977). Thus it is possible that naloxone increased conditioned suppression in Experiment 1 by enhancing already conditioned fear rather than by enhancing the conditioning of fear. It is also possible that the effects of naloxone seen in Experiment 1 are a result of naloxone's effect on memorial processes, that is, naloxone may enhance the memory of a prior conditioning experience (see Davis & Hendersen, 1985). To test these possibilities, Experiment 3 gave four groups of rats conditioned suppression training. Then, during extinction, three groups were treated with 4 mg/ kg of naloxone, 8 mg/kg of naloxone, or saline. The fourth group was not injected with any drug. If naloxone increases an already conditioned fear CR by affecting fear directly or by enhancing memory processes, then the groups receiving naloxone should show more suppression during the extinction of that fear than should saline and no-drug controls.
iNO DRUG I I I I I I I I I I I
DAYS
DRUG DAYS 4mg/kg
NALOXONESALINE [.._I
p:.OS p:.43
Figure 4. Mean barpress response rates per minute for naloxone group and the saline group during Experiment I.
NALOXONE AND CONDITIONED SUPPRESSION 583.
Method
Subjects and Apparatus
Thirty-nine male albino Holtzman rats, about 120-days-old at the start of the study, were housed and maintained as before. These rats had been used in a prior experiment in which they were shaped to barpress for sucrose reward and given 5 days of barpress training on a VI !-min schedule. After VI training, all the rats received, while blocked from the bar by a false wall, 10-US-alone (shock-alone) presentations and 7-tone-shock pairings. All received extinction to the tone CS before the start of the present experiment. The apparatus in this study was that of Experiments I and 2.
Procedure
Immediately after the tone extinction described above was completed, Experiment 3 began. The rats were assigned to four groups, 4-Nal (n = 10), 8-Nal (n = 10), no-drug (n = 9), and saline (n = 10). All groups received a 2-min light CS that coterminated with a 1-s, 1-mA scrambled footshock US. The light CS was the illumination of the 7.5-W, 110-V ac frosted bulb mounted on the rear wall of the cubicle. This was the same CS used in Experiment 1. The session length was 1,000 s. The light trial could begin in Min 7, 9, or 12 of each session while the rats barpressed on the VI !-min schedule for sucrose. There were seven such sessions given, one per day.
Next, all rats underwent extinction of suppression to the light CS. During each of seven daily sessions, the 2-min light occurred as before but without shock. Drug treatment did not begin until Day 4 of extinction. In earlier work we found that the one-trial-a-day procedure produces a slow rate of extinction; therefore, drug administration began after several days of extinction when conditioned suppression had become weak enough to yield a group mean suppression ratio of approximately 0.20. This procedure should reduce the odds that any drug effect would be obscured by a ceiling effect during early trials. Beginning on Day 4 and continuing throughout extinction, Groups 4-Nal and 8-Nal received 4 mg/kg and 8 mg/kg of naloxone, respectively. The drug was injected ip immediately before each session. The saline group received an equal volume of saline, and the no-drug group received no injections. Thus the no-drug group was treated exactly as on the first 3 days of extinction.
Results
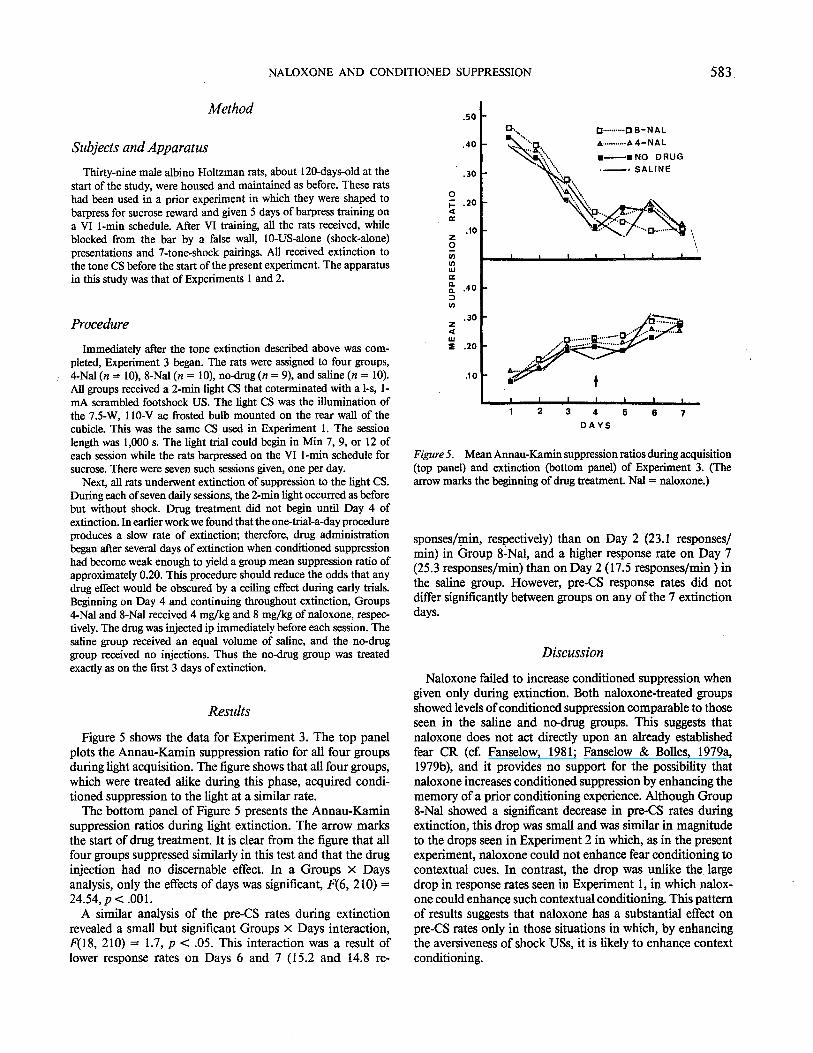
Figure 5 shows the data for Experiment 3. The top panel plots the Annau-Kamin suppression ratio for all four groups during light acquisition. The figure shows that all four groups, which were treated alike during this phase, acquired conditioned suppression to the light at a similar rate.
The bottom panel of Figure 5 presents the Annau-Kamin suppression ratios during light extinction. The arrow marks the start of drug treatment. It is clear from the figure that all four groups suppressed similarly in this test and that the drug injection had no discemable effect. In a Groups x Days analysis, only the effects of days was significant, F(6, 210) = 24.54, p < .00 1.
A similar analysis of the pre-CS rates during extinction revealed a small but significant Groups x Days interaction, F(18, 210) = 1.7, p < .05. This interaction was a result of lower response rates on Days 6 and 7 (15.2 and 14.8 re-
.50
.40
.30
0 1- .20 < a:
.10 z ~ IJ) IJ) w a: ll. .40 ll. ;:)
IJ)
.30 z < w :e .20
.10
2 3 4 DAYS
5 6 7
Figure 5. Mean Annau-Kamin suppression ratios during acquisition (top panel) and extinction (bottom panel) of Experiment 3. (The arrow marks the beginning of drug treatment. Nal = naloxone.)
sponses/min, respectively) than on Day 2 (23.1 responses/ min) in Group 8~Nal, and a higher response rate on Day 7 (25.3 responses/min) than on Day 2 (17 .5 responses/min) in the saline group. However, pre-CS response rates did not differ significantly between groups on any of the 7 extinction days.
Discussion
Naloxone failed to increase conditioned suppression when given only during extinction. Both naloxone-treated groups showed levels of conditioned suppression comparable to those seen in the saline and no-drug groups. This suggests that naloxone does not act directly upon an already established fear CR (cf. Fanselow, 1981; Fanselow & Bolles, 1979a, 1979b ), and it provides no support for the possibility that naloxone increases conditioned suppression by enhancing the memory of a prior conditioning experience. Although Group 8-Nal showed a significant decrease in pre-CS rates during extinction, this drop was small and was similar in magnitude to the drops seen in Experiment 2 in which, as in the present experiment, naloxone could not enhance fear conditioning to contextual cues. In contrast, the drop was unlike the large drop in response rates seen in Experiment 1, in which naloxone could enhance such contextual conditioning. This pattern of results suggests that naloxone has a substantial effect on pre-CS rates only in those situations in which, by enhancing the aversiveness of shock USs, it is likely to enhance context conditioning.

584 MICHAEL VIGORITO AND JOHN J. B. AYRES
General Discussion
The purpose of this research was to assess the effects of naloxone on conditioned suppression. Naloxone given during acquisition increased posttrial suppression to a light CS, increased pre-CS suppression, and enhanced CS-elicited suppression during extinction. Naloxone administered only during the extinction ofCS-elicited suppression, however, did not affect the magnitude of that suppression. These effects are consistent with the hypothesis that naloxone increases the aversiveness of shock USs by blocking an endorphin analgesic system. Under that hypothesis, naloxone should be effective only if administered during the acquisition of conditioned suppression and should have no effect if administered only during the extinction of suppression.
Even though naloxone increased conditioned suppression, it did not eliminate WSDs in that suppression. Why WSDs were not eliminated by naloxone is not clear. Perhaps they reflect short-term CS habituation-like effects. However, the possibility of a nonopioid analgesic mechanism cannot be ruled out. (See Maier, Sherman, Lewis, Terman, & Liebeskind, 1983, and Grau, 1984, for evidence that shock can activate two functionally distinct analgesic mechanisms.)
Some alternative explanations as to why naloxone increased conditioned suppression need to be considered. One might argue that naloxone did not increase the aversiveness of the US but caused a general suppression of motor activity. This suppression of motor activity resulted in less barpressing behavior, which was erroneously taken as increased conditioned fear. This is an unlikely explanation, however, because naloxone increased conditioned suppression only when the drug was present during pairings of CSs and shock USs; naloxone had no effect on conditioned suppression when given only during the extinction of suppression (Experiment 3). Moreover, it is unclear how a simple motor interpretation would explain why greater suppression is seen during the CS and posttrial periods than during the pre-CS period. Also, naloxone had little effect on baseline responding in Experiment 2 in nonfrightened animals.
Another alternative explanation of the effect of naloxone on conditioned suppression is that the drug reduces motivation for sucrose. Experiment 2 systematically examined the effect of two doses of naloxone on barpress responding for sucrose and found that naloxone caused only a small drop in VI response rates. (Similar results were obtained by Vigorito and Sclafani, 1986. They reported that naloxone fails to reduce rats' barpress responding for sucrose reinforcement during 30-min sessions but does reduce intake of the sucrose solution when given 30 min/day access at their home cages.) It is unlikely that naloxone's relatively small effect on baseline responding for sucrose caused the group differences in pre-CS and posttrial suppression found in Experiment 1. Moreover, weaker CS suppression in extinction would be expected if naloxone depressed only baseline responding without enhancing conditioning (Blackman, 1968); the opposite result was in fact observed.
A third alternative explanation is that naloxone affected +J."! acquisition of conditioned suppression by increasing an already established fear CR instead of increasing pain. Such
an increase in fear could be a direct effect of naloxone on the fear CR itself or could be an indirect effect brought about by naloxone's ability to enhance the memory of prior conditioning experiences. The results from Experiment 3 argue against both possibilities. If naloxone increased an already established fear CR either directly or indirectly, then one would expect increased CS-elicited suppression during extinction in rats treated with the drug only during extinction. This did not occur in Experiment 3, nor has it been observed with freezing produced by fear-eliciting context cues (Fanselow, 1982) or with freezing produced by a fear-eliciting discrete CS (Fanselow & Bolles, 1979a).
In Schull's ( 1979) conditioned opponent process theory and Fanselow's ( 1982) model of aversive Pavlovian conditioning, an endorphin analgesic system can be activated by an associative process. Thus a CS that has previously preceded shock can activate endorphins, which in tum would reduce the aversiveness of the shock on subsequent trials. Such a process would limit the amount of fear conditioning that could take place. Naloxone, by antagonizing the endorphin system, would prevent the endorphins from minimizing the aversiveness of the shock and would therefore remove the limits that the endorphin system places on conditioning. Our results are consistent with this view.
Our results are also consistent with a view that emphasizes the role of the US as an activator of an endorphin system. The endorphins, by reducing the aversiveness of subsequent USs, would then limit fear conditioning. By blocking the endorphin system, naloxone would maintain the aversiveness of the US and again remove the limits otherwise placed on conditioning. ·
Our data do not permit us to distinguish between a conditioned or unconditioned elicitation of analgesia. Nevertheless, our results indicate that conditioned suppression increases with both shock intensity and naloxone treatment. This finding can be added to the growing list of aversively motivated behaviors that increase with shock intensity and naloxone treatment, such as freezing (Fanselow & Bolles, 1979b), defensive burying (Whiteside & Devenport, 1985), shock-elicited aggression (Fanselow, Sigmundi, & Bolles, 1980), and suppressive effects of shock punishment (Young, .1980). Collectively, these experiments provide growing support for the hypothesis that aversive stimuli can activate an endogenous analgesic system along with species-specific-defensive behaviors.
References
Annau, Z., & Kamin, L. J. (1961). The conditioned emotional response as a function of intensity of the US. Journal of Comparative and Physiological Psychology, 54, 428-432.
Ayres, J. J. B., Axelrod, H., Mercker, E., Muchnick, F., & Vigorito, M. (1985). Concurrent observations of barpress suppression and freezing: Effects of CS modality and on-line vs. off-line training upon ·posttrial behavior. Animal Learning and Behavior, 13, 44-50.
Ayres, J. J. B., Berger-Gross, P., Kohler, E. A., Mahoney, W. J., & Stone, S. (1979). Some orderly nonmonotonicities in the trial-bytrial acquisition of conditioned suppression: Inhibition with reinforcement? Animal Learning and Behavior, 7, 174-180.

NALOXONE AND CONDITIONED SUPPRESSION 585
Ayres, J. J. B., & Vigorito, M. (1984). Posttrial effects of presenting vs. omitting expected shock USs in the conditioned suppression procedure: Concurrent measurement of barpress suppression and freezing. Animal Learning and Behavior,.I2, 73-78.
Basbaum, A. 1., & Fields, H. L. (1978). Endogenous pain control mechanisms: Review and hypothesis. Annals of Neurology, 4, 451-461.
Blackman, D. E. (1968). Response rate, reinforcement frequency, and conditioned suppression. Journal of the Experimental Analysis of Behavior, 11, 503-516.
Bolles, R. C. ( 1970). Species-specific defense reactions and avoidance learning. Psychological Review, 77, 32-48.
Bolles, R. C., & Fanselow, M. S. (1980). A perceptual-defensiverecuperative model of fear and pain. The Behavioral and Brain Sciences, 3, 291-323.
Bolles, R. C., & Fanselow, M. S. (1982). Endorphins and behavior. Annual Review of Psychology, 33, 87-101.
Bolles, R. C., & Riley, A. L. (1973). Freezing as an avoidance response: Another look at the operant-respondent distinction. Learning and Motivation, 4, 268-275.
Brown, D. R., & Holtzman, S. G. ( 1979). Suppression of deprivationinduced food and water intake in rats and mice by naloxone. Pharmacology, Biochemistry, and Behavior, II, 567-573.
Chance, W. T. (1980). Autoanalgesia: Opiate and non-opiate mechanisms. Neuroscience and Biobehavioral Reviews, 4, 55-67.
Chance, W. T., Krynock, G. M., & Rosecrans, J. A. (1978). Antinociception following lesion-induced hyperemotionality and conditioned fear. Pain, 4, 243-252.
Chance, W. T., White, A. C., Krynock, G. M., & Rosecrans, J. A. ( 1978). Conditioned fear-induced antinociception and decreased binding of (3H) N-Leu-enkephalin to rat brain. Brain Research, 14I, 371-374.
Davis, H. D., & Hendersen, R. W. ( 1985). Effects of conditioned fear on responsiveness to pain: Long-term retention and reversibility by naloxone. Behavioral Neuroscience, 99, 277-289.
Dewey, W. L., & Harris, L. S. (1975). The tail flick test. In S. Ehrenpreis & A. Niedle (Eds.), Methods in narcotic research. New York: Dekker.
Fanselow, M.S. (1981). Naloxone and Pavlovian fear conditioning. Learning and Motivation, 12, 398-419.
Fanselow, M. S. ( 1982). The postshock activity burst. Animal Learning & Behavior, 10, 448-454.
Fanselow, M. S. ( 1984a). Opiate modulation of the active and inactive components of the postshock reaction: Parallels between naloxone pretreatment and shock intensity. Behavioral Neuroscience, 98, 269-277.
Fanselow, M.S. (1984b). Shock-induced analgesia and the formalin test: Effects of shock severity, naloxone, hypophysectomy, and associative variables. Behavioral Neuroscience, 98, 79-95.
Fanselow, M.S., & Baackes, M.P. (1982). Conditioned fear-induced opiate analgesia on the formalin test: Evidence for two aversive motivational systems. Learning and Motivation, 13, 200-221.
Fanselow, M.S., & Bolles, R. C. (1979a). Triggering of the endorphin analgesic reaction by a cue previously associated with shock: Reversal by naloxone. Bulletin of the Psychonomic Society, 14, 88-90.
Fanselow, M. S., & Bolles, R. C. (1979b). Naloxone and shockelicited freezing in the rat. Journal of Comparative and Physiological Psychology, 93, 736-744.
Fanselow, M.S., Sigmundi, R. A., & Bolles, R. C. (1980). Naloxone pretreatment enhances shock-elicited aggression. Physiological Psychology, 8, 369-371.
Goldstein, M. L. ( 1960). Acquired drive strength as a joint function of shock intensity and number of acquisition trials. Journal of Experimental Psychology, 60, 349-358.
Grau, J. W. (1984). Influence of naloxone on shock-induced freezing and analgesia. Behavioral Neuroscience, 98, 278-292.
Grevert, P., & Goldstein, A. (1977). Effects of naloxone on experimentally induced ischemic pain and on mood in human subjects. Proceedings of the National Academy of Sciences of the United States of America, 74, 1291-1294.
Kaplan, R., & Glick, S. D. (1979). Prior exposure to footshock induced naloxone hyperalgesia. Life Sciences, 24, 2309-2312.
Kimmel, H. D., & Bums, R. A. (1975). Adaptational aspects of conditioning. In W. K. Estes (Ed.), Handbook of learning and cognitive processes: Conditioning and behavior theory (vol. 2, pp. 99-142). Hillsdale, NJ: Erlbaum.
Libby, A. (1951 ). Two variables in the acquisition of depressant properties by a stimulus. Journal of Experimental Psychology, 42, 100-107.
MacLennan, A. J., Jackson, R. L., & Maier, S. F. (1980). Conditioned analgesia in the rat. Bulletin of the Psychonomic Society, I5, 387-390.
Maier, S. F., Sherman, J. E., Lewis, J. W., Terman, G. W., & Liebeskind, J. C. (1983). The opioid/nonopioid nature of stressinduced analgesia and learned helplessness. Journal of Experimental Psychology: Animal Behavior Processes, 9, 80-90.
Millenson, J. R., & Dent, J. G. (1971). Habituation of conditioned suppression. Quarterly Journal of Experimental Psychology, 23, 126-134.
Morley, J. E., Levine, A. S., Yim, G. K., & Lowy, M. T. (1983). Opioid modulation of appetite. Neuroscience and Biobehavioral Reviews, 7, 281-305.
Overmier, J. B., Payne, R. J., Brackbill, R. M., Linder, B., & Lawry, J. A. (1979). On the mechanism of the post-asymptotic decrement phenomenon. Acta Neurobiologiae Experimentalis, 39, 603-620.
Overton, D. A. (1968). Dissociated learning in drug states (state dependent learning). In D. H. Efron, J.D. Cole, J. Levine, & J. R. Wittenborn (Eds:), Psychopharmacology: A review of progress I957-I967. Washington, DC: Public Health Service Publication No. 1836.
Ramabadran, K., Guillan, J. C., & Jacob, J. (1979). Actions of hyperalgesic substances(-) naloxone, theophylline, and 5-metoxyN, N-dimethyltryptamine on nociceptive learning. In Leong Way (Ed.), Endogenous and exogenous opiate agonists and antagonists. New York: Permagon Press.
Reid, L. D. (1985). Endogenous opioid peptides and regulation of drinking and feeding. The American Journal of Clinical Nutrition, 42, 1099-1132.
Rescorla, R. A., & Wagner, A. R. (1972). A theory of Pavlovian conditioning: Variations in the effectiveness of reinforcement and nonreinforcement. In A. H. Black & W. F. Prokasy (Eds.), Classical conditioning II: Current research and theory (pp. 64-99). New York: Appleton-Century-Crofts.
Riley, A. L., Zellner, D. A., & Duncan, H. J. (1980). The role of endorphins in animal learning and behavior. Neuroscience and Biobehavioral Reviews, 4, 69-76.
Rodgers, G. H., Frank, H., Taylor, A. N., & Liebeskind, J. C. (1978). Naloxone suppression of food and water intake in deprived rats. Proceedings of the Western Pharmacological Society, 21, 457-460.
Scallet, A. C. (1982). Effects of conditioned fear and environmental novelty on plasma 13-endorphin in the rat. Peptides, 1, 203-206.
Schull, J. (1979). A conditioned opponent theory of Pavlovian conditioning and habituation. In G. Bower (Ed.), The psychology of learning and motivation (Vol. 13, pp. 57-90). New York: Academic Press.
Sclafani, A., Aravich, P. F., & Xenakis, S. (1982). Dopaminergic and endorphinergic mediation of a sweet reward. In B. G. Hoebel & D. Novin (Eds.), The neuratbasis of feeding and reward. Brunswick, ME: Haer Company.

586 MICHAEL VIGORITO AND JOHN J. B. AYRES
Sherman, J. E., & Liebeskind, J. C. (1980). An endorphinergic, centrifugal substrate of pain modulation: Recent findings, current concepts, and complexities. In J. J. Bonica (Ed.), Pain (pp. 191-204). New York: Raven Press.
Solomon, R. L., & Corbit, J.D. (1974). An opponent-process theory of motivation: I. Temporal dynamics of affect. Psychological Review, 81, 119-145.
Vigorito, M., & Sclafani, A. (1986). Naloxone suppression of sucrose intake: Reduced hedonic response or enhanced satiety? Paper pre-
sented at the annual meeting of the Eastern Psychological Association, New York City.
Whiteside, D. A., & Devenport, L. D. ( 1985). Naloxone, preshock, and defensive burying. Behavioral Neuroscience, 99, 436-440.
Young, G. A. (1980). Naloxone enhancement of punishment in the rat. Life Sciences, 26, 1787-1792.
Received August 19, 1985 Revision received April25, 1986 •
Recommended

















