
583
Critical Reviews in Plant Sciences, 21(6):583–606 (2002)
0735-2689/02/$.50© 2002 by CRC Press LLC
Endophytic Bacteria and Their Potential Applications
Cindy Lodewyckx,1,2 Jaco Vangronsveld,2 Fiona Porteous,3 Edward R.B.Moore,3 Safieh Taghavi,1,4 Max Mezgeay,1,5 and Daniel van der Lelie1,2,4*
1Environmental Technology, Flemish Institute for Technological Research (Vito), Boeretang 200, 2400 Mol,Belgium; 2 Environmental Biology, Limburgs Universitair Centrum, Universitaire Campus, 3590 Diepenbeek,Belgium;3Soil Quality and Protection Programme, Macaulay Land use Research Institute, Craigiebuckler,Aberdeen, AB15 8QH, UK; 4Present address: Biology Department, Bldg. 463, Brookhaven National Laboratory,Upton, NY11973-5000, USA; 5Lab. for Microbiology, Center of Studies for Nuclear Energy, SCK/CEN,Boeretang 200, B 2400 Mol, Belgium
* Corresponding author: Daniel van der Lelie ([email protected])
I. INTRODUCTION
Although the term “endophyte” is most com-monly associated with fungi, there is a signifi-cant amount of literature pertaining to bacteriaas endophytes as well, some of which are be-lieved to impart a beneficial effect, whereas oth-ers are regarded to have a neutral or detrimentaleffect on plants. For more than 50 years, bacteriahave been observed to exist inside plants with-out causing apparent disease symptoms (Tervetand Hollis, 1948; Hollis, 1951). Various reportsindicate that such bacteria exist in a variety oftissue types within numerous plant species, sug-gesting a ubiquitous existence in most if not allhigher plants.
A variety of definitions have been applied tothe term endophyte and consideration of eachleads to different interpretations. Kloepper et al.(1992) called bacteria found within tissues inter-nal to the epidermis endophytes. Because quies-cent endophytic bacteria can become pathogenicunder certain conditions and within different hostgenotypes (Misaghi and Donndelinger, 1990),James and Olivares (1997) adjusted the definitionand stated that all bacteria that colonize the inte-rior of plants, including active and latent patho-gens, can be considered to be endophytes. Con-sidering all bacteria that colonize the interior ofplants, one should also take into account thosebacteria that reside within living plant tissues
without doing substantive harm or gaining benefitother than securing residency (Kado, 1992), aswell as those bacteria that establish endosymbio-sis with the plant, whereby the plant receives anecological benefit from the presence of the sym-biont (Quispel, 1992). The definition of endo-phytic bacteria should, in accordance with thedefinition of endomycorrhizal fungi, also includebacteria that reside in the cortex of the root. Thesesubdefinitions may provide an operational over-view of what is considered to be an “endophyte”,and consequently this might be regarded as themost general definition of for which the term“endophyte” stands. In practice, the differencebetween endophytic bacteria residing in the rootand bacteria colonizing the rhizoplane is oftendetermined by the surface sterilization techniqueapplied.
II. ECOLOGY OF ENDOPHYTICBACTERIA
Endophytic bacteria have been isolated fromboth monocotyledonous and dicotyledonous plants,ranging from woody tree species, such as oak(Brooks et al., 1994) and pear (Whitesides andSpotts, 1991), to herbaceous crop plants, such assugar beets (Jacobs et al., 1985) and maize (Fischeret al., 1992; Gutierrez-Zamora and Martinez-Romero, 2001; Lalande et al., 1989; McInroy and

584
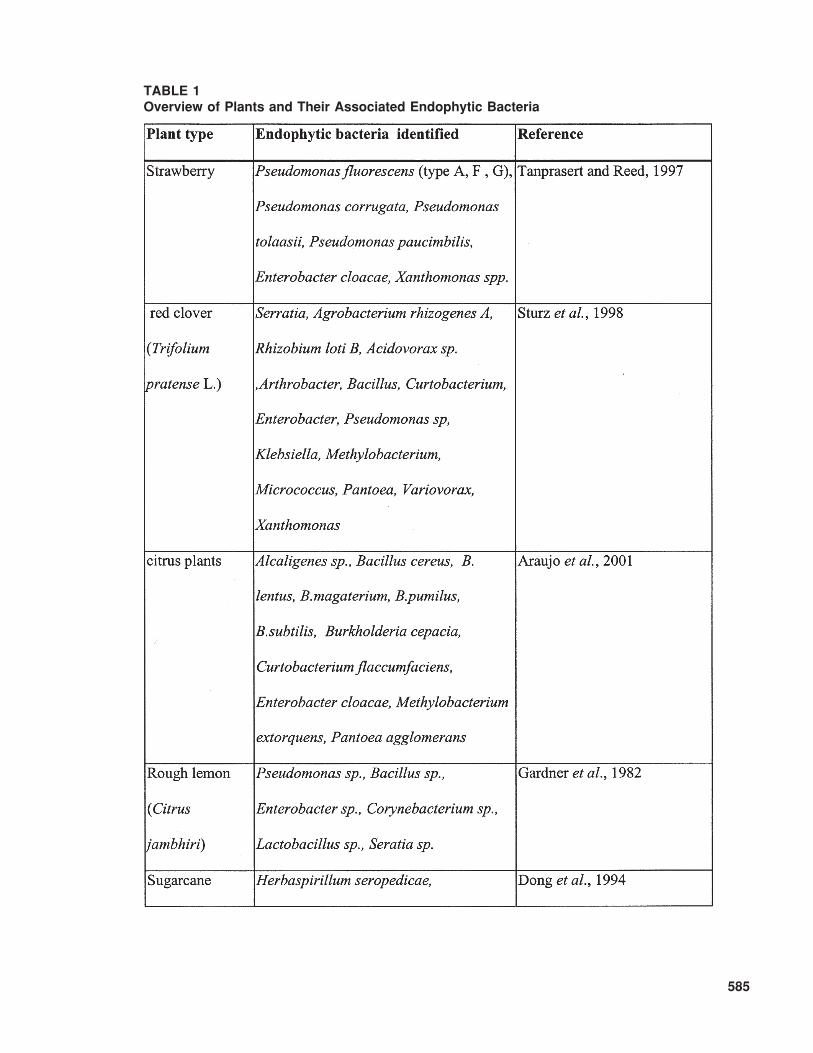
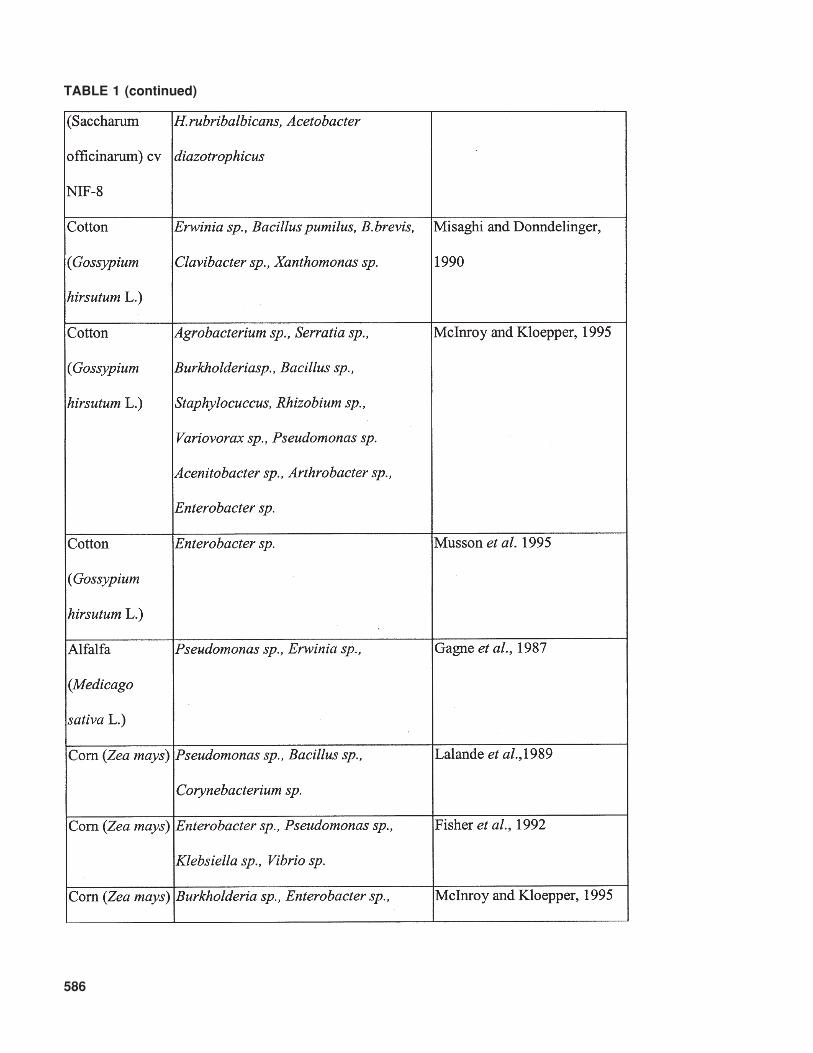
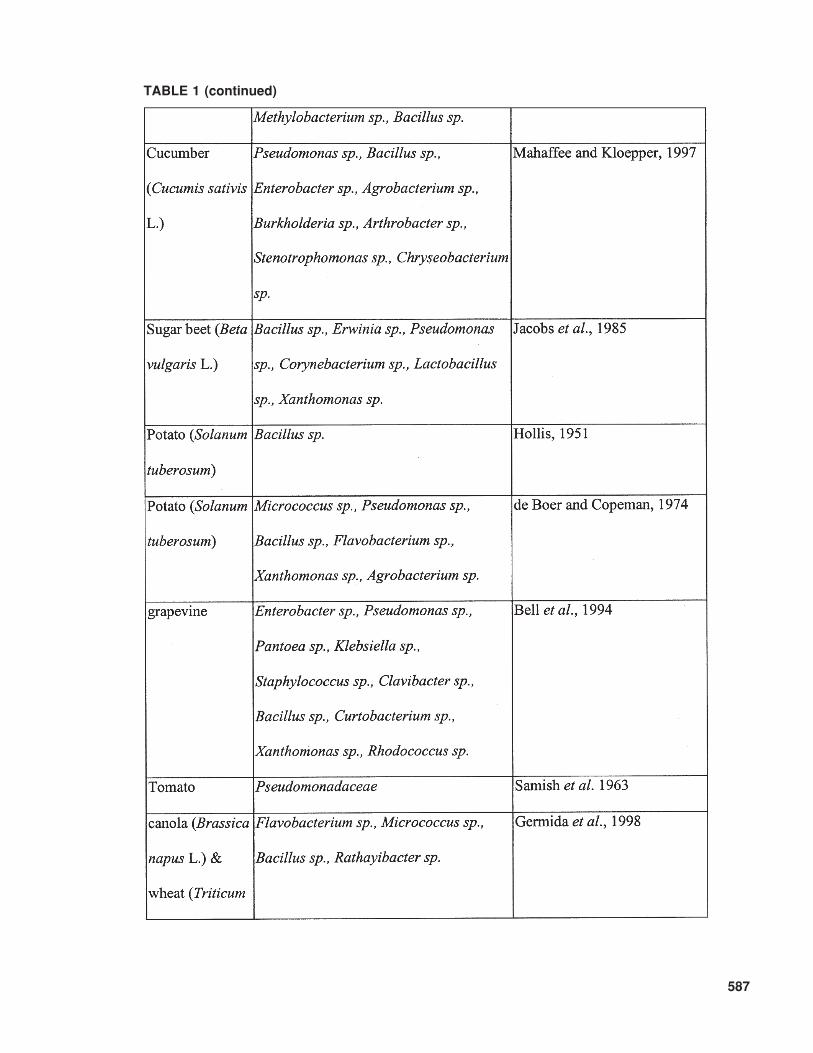
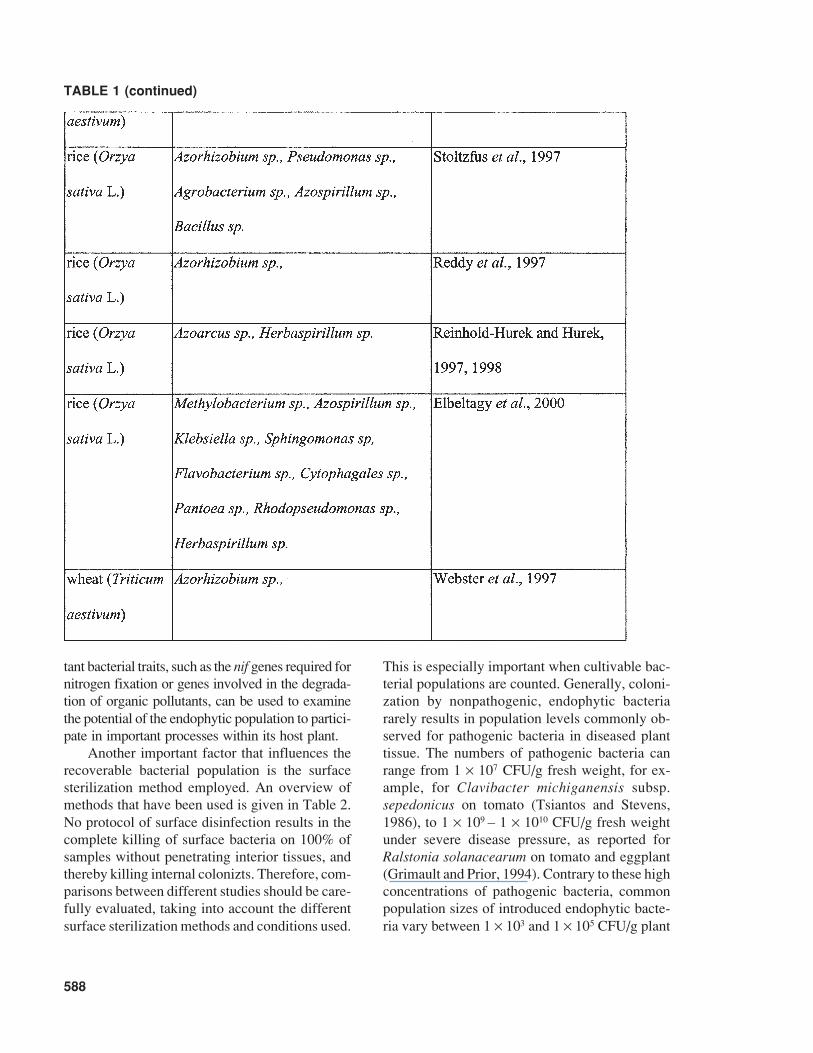
Kloepper, 1995). Diversity associated with bacte-rial endophytes exists, not only in the plant speciescolonized but also in the colonizing bacterial taxa.As shown in Table 1, plants can be colonizedsimultaneously by a large variety of endophyticbacteria. The variation in bacteria that has beenreported as endophytes spans a significant range ofGram-positive and Gram-negative bacteria andinclude genera such as Acidovorax, AcinetobacterActinomyces, Aeromonas, Afipia, Agrobacterium,Agromonas, Alcaligenes, Alcanivorax, Allorhizobium,Alteromonas, Aminobacter, Aquaspirillum,Arthrobacter, Aureobacterium, Azoarcus, Azo-monas, Azorhizobium, Azotobacter, Azospirillum,Bacillus, Beijerinckia, Blastobacter, Blastomonas,Brachymonas, Bradyrhizobium, Brenneria,Brevundimonas, Burkholderia, Chelatobacter,Chromobacterium, Chryseomonas, Comamonas,Corynebacterium, Delftia, Derxia, Devosia,Enterobacter, Flavimonas, Flavobacterium,Flexibacter, Frankia, Halomonas, Herbaspirillum,Matsuebacter, Mesorhizobium, Moraxella, Nevskia,Nocardia, Ochrobactrum, Pantoea, Pectobacte-rium, Phenylbacterium, Phyllobacterium, Photo-bacterium, Porphyrobacter, Pseudoalteromonas,Pseudomonas, Psychrobacter, Ralstonia, Renibac-terium Rhizobacter, Rhizobium, Rhizomonas,Rhodanobacter, Rhodococcus, Shewanella,Sinorhizobium, Sphingobacterium, Sphingomonas,Spirillum, Stenotrophomonas, Streptomyces,Thauera, Variovorax, Vibrio, Xanthomonas,Xylella, Zoogloea, Zymobacter, Zymomonas, ormembers of the group of the pink-pigmented fac-ultatively methylotrophic bacteria, such asMethylobacterium. Even though it remains diffi-cult to compare earlier and more recent studies thatidentify bacteria, certain trends are apparent withpredominant bacterial types isolated as endophytes(Kobayashi and Palumbo, 2000).
Plants can be considered to be complexmicroecosystems where different habitats are ex-ploited by a wide variety of bacteria (McInroyand Kloepper, 1994). These habitats are not onlyrepresented by plant external surfaces, where epi-phytic bacteria predominate, but also by internaltissues, where many microorganisms penetrateand survive. Inside the plant microecosystem,different microbial species, both bacterial andfungal (Fisher et al., 1992), are able to interact
and to establish an equilibrium. Some of thesemicroorganisms can be considered to be domi-nant species (Van Peer et al., 1990) and may berepresented by those that are most frequently, andin large numbers, isolated from the host plant. Aswell as the dominants, there is a large variety ofspecies that cannot be isolated easily because oftheir low numerical consistencey. These are con-sidered to be rare species. This distribution amongendophytic bacteria was first observed by Gardneret al. (1982), who identified bacteria present inthe xylem fluid from the roots of the rough lemonrootstock of the Florida citrus tree. Among the 13genera found, the most frequently occurring gen-era, Pseudomonas (40%) and Enterobacter (18%),could be regarded as the dominant species, whilethe others might be classified as rare species. Thisstable biodiversity is considered to be the mostimportant condition in the establishment of anyecosystem.
It should be noted that attempts to evaluatetotal populations of bacteria in plants may pro-duce varied results, depending on the growth mediaused for isolation, variations in the growth condi-tions of the host plant, and the manner in whichthe plant tissue was used. This is also the case forplant-associated rhizosphere bacterial populations(Kloepper and Beauchamp, 1992). Molecularapproaches, based on amplification of the 16SrDNA may be employed to overcome the limita-tions of classic isolation procedures that are de-pendent on the isolation of bacteria. After theisolation of total DNA from the plant or specificplant parts, such as roots, vascular fluid, or leaves,oligonucleotide primers are employed to hybrid-ize at conserved sites on the DNA and specifi-cally amplify the bacterial 16S rRNA genes(Weisburg, 1991). The PCR-amplifed 16S rDNAmay be cloned and further analyzed by sequencingor T-RFIP in order to identify the bacteria. Sucha strategy was used by Sessitsch et al. (2001) toanalyze the populations of endophytic bacteria inthree potato cultivars. Additionally, molecular eco-logical techniques that are routinely applied tostudy population composition and activity, in-cluding DGGE, TGGE, and T-RFLP, can be ap-plied to study the population dynamics of endo-phytic bacteria. Furthermore, primers that allowthe specific amplification of genes encoding impor-
585
TABLE 1Overview of Plants and Their Associated Endophytic Bacteria
586
TABLE 1 (continued)
587
TABLE 1 (continued)
588
tant bacterial traits, such as the nif genes required fornitrogen fixation or genes involved in the degrada-tion of organic pollutants, can be used to examinethe potential of the endophytic population to partici-pate in important processes within its host plant.
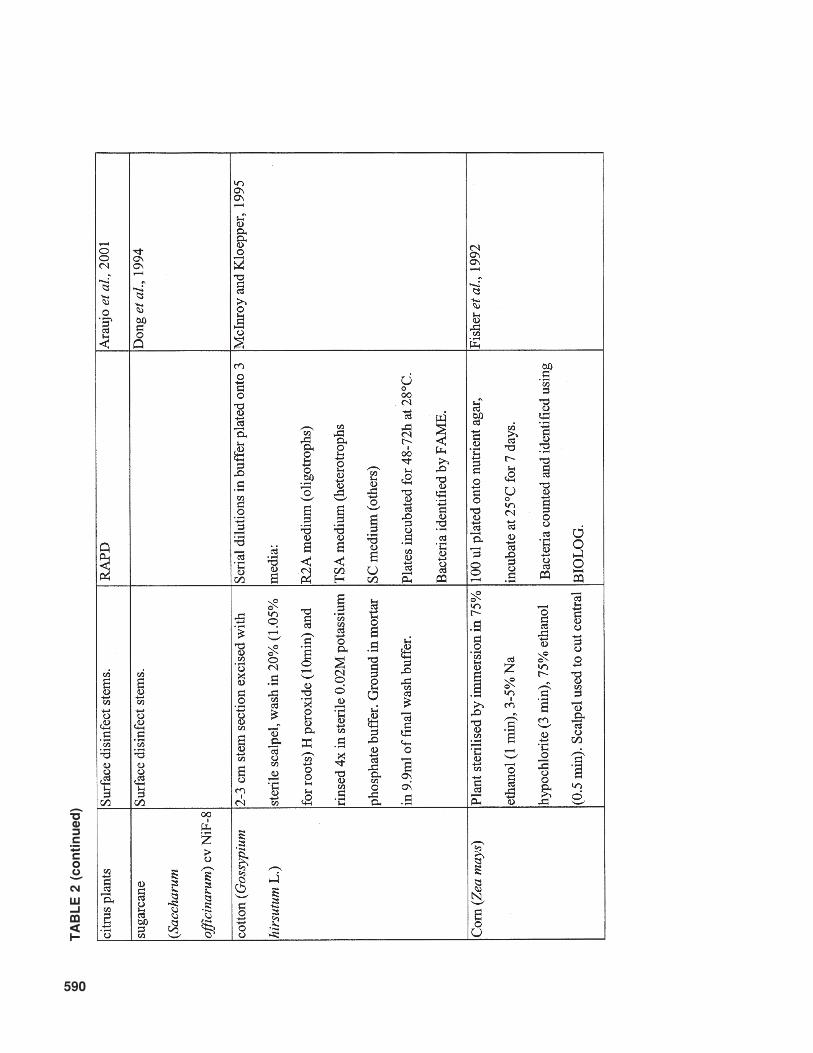
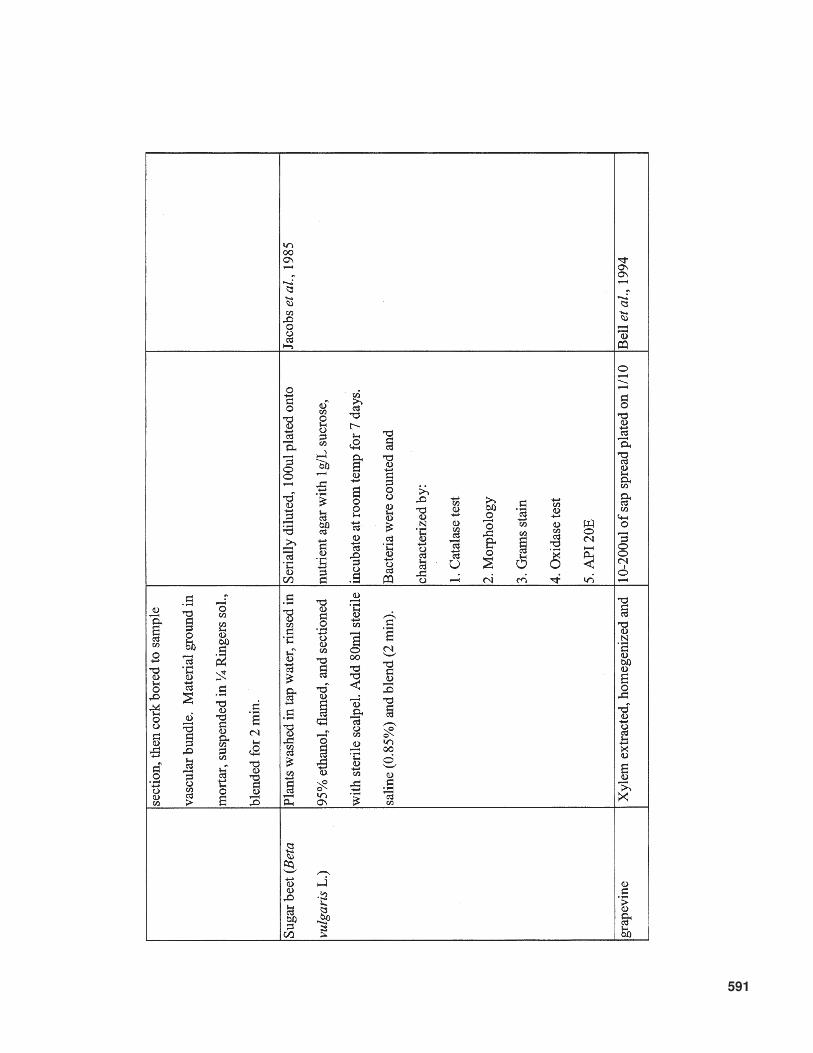
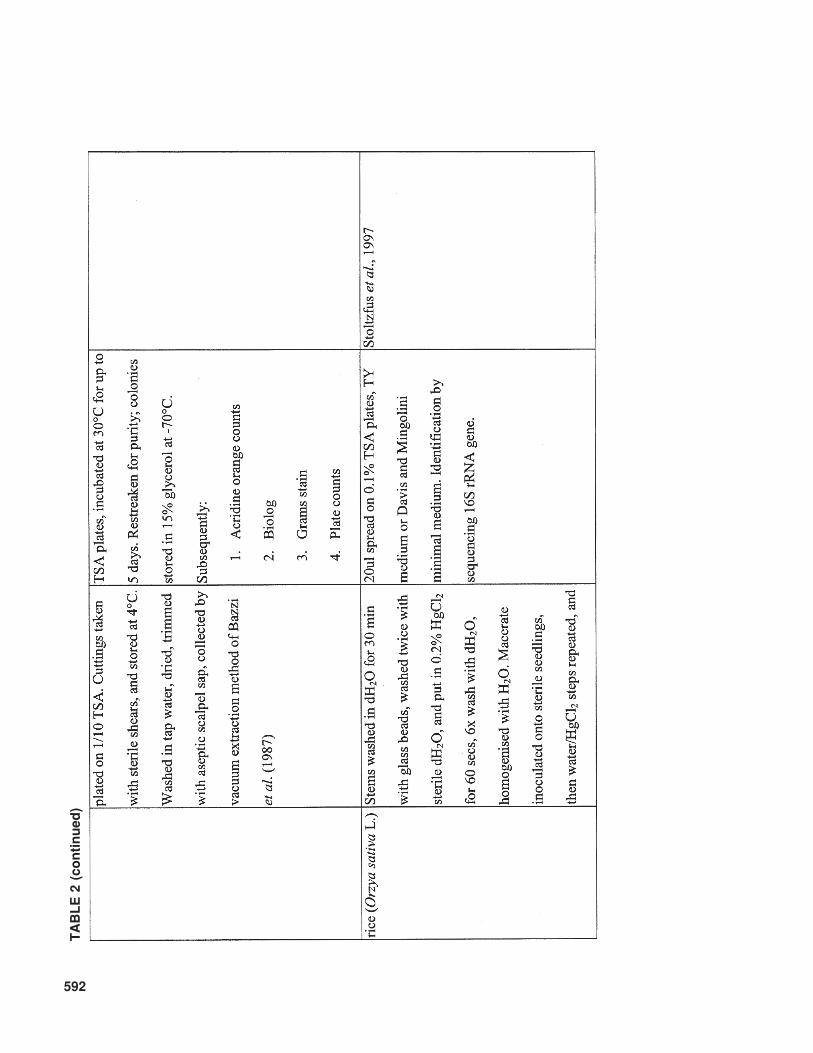
Another important factor that influences therecoverable bacterial population is the surfacesterilization method employed. An overview ofmethods that have been used is given in Table 2.No protocol of surface disinfection results in thecomplete killing of surface bacteria on 100% ofsamples without penetrating interior tissues, andthereby killing internal colonizts. Therefore, com-parisons between different studies should be care-fully evaluated, taking into account the differentsurface sterilization methods and conditions used.
TABLE 1 (continued)
This is especially important when cultivable bac-terial populations are counted. Generally, coloni-zation by nonpathogenic, endophytic bacteriararely results in population levels commonly ob-served for pathogenic bacteria in diseased planttissue. The numbers of pathogenic bacteria canrange from 1 × 107 CFU/g fresh weight, for ex-ample, for Clavibacter michiganensis subsp.sepedonicus on tomato (Tsiantos and Stevens,1986), to 1 × 109 – 1 × 1010 CFU/g fresh weightunder severe disease pressure, as reported forRalstonia solanacearum on tomato and eggplant(Grimault and Prior, 1994). Contrary to these highconcentrations of pathogenic bacteria, commonpopulation sizes of introduced endophytic bacte-ria vary between 1 × 103 and 1 × 105 CFU/g plant

589
TA
BL
E 2
Ove
rvie
w o
f M
eth
od
s U
sed
to
Iso
late
an
d C
har
acte
rize
En
do
ph
ytic
Bac
teri
a fr
om
Dif
fere
nt
Pla
nt
Sp
ecie
s
590
TA
BL
E 2
(co
nti
nu
ed)
591
592
TA
BL
E 2
(co
nti
nu
ed)

593

594
tissue for most investigated plant species (Donget al., 1994; Frommel et al., 1991; Quadt-Hallmannand Kloepper, 1996; Lamb et al., 1996). Similarto indigenous endophytes, which are usually abun-dant in the same concentration range as the intro-duced endophytes, the highest bacterial densitiesare usually observed in the roots and decreaseprogressively from the stem to the leaves (Quadt-Hallman and Kloepper, 1996; Lamb et al., 1996).Independent of the initial inoculum size, endo-phytic populations tend to approach optimal den-sities depending on the plant tissue. For potatoesinoculated with an endophytic Pseudomonas sp.,the root population increased to a final populationdensity of 1 × 106 CFU/cm, whereas the stempopulation decreased from an initial concentra-tion of 3.3 × 105 to 1.9 × 103 CFU/cm (Frommelet al., 1991). The fact that colonization is espe-cially abundant in root tissue may reflect the factthat the root is the primary site where endophytesgain entry into plants. With the exception of seed-transmitted bacteria, which are present in the plant,potential endophytes must first colonize the rootsurface prior to entering the plant. This mightexplain the close relationship between endophyticand rhizosphere colonizing bacteria, that is, manyfacultative endophytic bacteria can also surviveas rhizosphere bacteria. Potential internal colo-nists find their host by chemotaxis, electrotaxis,or accidental encounter. The main entry for endo-phytic bacteria appears to be through wounds thatoccur naturally as a result of plant growth orthrough root hairs and at epidermal junctions(Sprent and de Faria, 1988). This was confirmedby several microscopic studies (Wiehe et al., 1994;Benhamou et al., 1996a, 1996b; Pan et al., 1997).Artificial wounding was shown to contribute toincreased endophytic colonization in roots of plantscompared with intact roots (Gagné et al., 1987).Besides providing entry avenues, wounds alsocreate favorable conditions for the approachingbacteria by allowing leakage of plant exudates,which serve as a nutrient source for the bacteria.
Wounds and lateral roots are not, however,absolutely required for entrance of endophyticbacteria. Seedlings grown with minimal distur-bance in liquid media or on water agar were pen-etrated by endophytic bacteria long before lateralroot emergence (Levanoy et al., 1989; Quadt-
Hallmann et al., 1997). Because untreated controlplants were endophyte free, the observed bacte-rial behavior might indicate active penetration.This hypothesis is supported by the presence ofcellulytic and pectinolytic enzymes produced bynumerous endophytic bacteria, such as Azoarcussp. (Hurek et al., 1994), Azospirillum irakense(Khammas and Kaiser, 1991), and Pseudomonasfluorescens (Duijff et al., 1997; Benhamou et al.,1996a; Quadt-Hallmann et al., 1997). Enzymaticdegradation of plant cell walls by these bacteriawas only observed when they colonized the rootepidermis but never after colonizing intercellularspaces of the root cortex. These results suggestthat the endophyte induced production of cellu-lase and pectinase specifically for penetration intothe host plant. Although these observations dem-onstrate the possibility of active penetrationmechanisms for some endophytic bacteria, verylittle is known about the origin and regulation ofthese enzymes. It is assumed that these bacteriamust possess some regulatory mechanism to spe-cifically regulate their enzyme production in termsof quantity and time of expression.
Although the root zone offers the most obvi-ous site of entry for many endophytes, entry mayalso occur at sites on aerial portions of plants.Sharrock et al. (1991) suggested that, in somecases, endophytic populations within fruit mayarise by entry through flowers. Endophyte pen-etration is also believed to occur through naturalopenings on the leaves (e.g., stomata) or throughstem lenticels (Kluepfel, 1993). A completelydifferent way of penetration is described byAshbolt and Inkerman (1990) for sugar cane, viathe mealybug, and by Kluepfel (1993), via a rangeof different insects.
Once inside the plant tissue, endophytic bac-teria remain localized in a specific plant tissue,such as the root cortex, or colonize the plantsystematically by transport or active migrationthrough the conducting elements or the apoplast(Hurek et al., 1994; James et al., 1994; Mahaffeeet al., 1997; Quadt-Hallmann et al., 1997, Patriquinand Döbereinner, 1978). The different mecha-nisms of distribution might be due to interactionswith other bacteria or to the different require-ments of each microorganism that allows them toinhabit different niches, represented by tissue and,

595
more specifically, by the intercellular spaces in-side each tissue (Di Fiori and Del Gallo, 1995).There are a few studies that have reported on theintracellular presence of endophytes. This seemsto be an occasional finding, except for particularcases, such as those involving Rhizobium or Al-caligenes faecalis (You et al., 1983 and 1991),where both bacteria are enveloped by the plant inspecialized structures.
The fact that bacteria seem to be capable ofcolonizing the internal tissues of plants couldconfer an ecological advantage over bacteria thatcan only colonize plants epiphytically. The inter-nal tissues of plants are thought to provide a moreuniform and protective environment for microor-ganisms than plant surfaces, where exposure toextreme environmental conditions, such as tem-perature, osmotic potentials, and ultraviolet ra-diation, are major factors limiting long-term bac-terial survival. The plant interior could also be ahideout from grazing by soil protozoa. However,there are probably other limiting factors that mustbe overcome when establishing populations in theinternal tissues of plants. Thus, establishing andmaintaining an introduced bacterial populationwould still be limited and influenced by the samefactors that limit plant disease.
III. BENEFICIAL EFFECTS
The beneficial effects of bacterial endophyteson their host plant appear to occur through similarmechanisms described for plant growth-promot-ing rhizobacteria (PGPR) (Kloepper et al., 1991a;Höflich et al., 1994). This makes sense becausemost of the bacterial endophytes isolated fromtrees, healthy crops, and weeds can be consideredto be facultatively endophytic and are capable ofliving outside plant tissues as rhizospheric bacte-ria (Di Fiori and Del Gallo, 1995). Additionally,many endophytic bacterial taxa from sweet cornor cotton were reported to be common soil bacte-ria (McInroy and Kloepper, 1994).
Plant growth-promoting bacteria can affectplant growth directly or indirectly. The directpromotion of plant growth by PGPR, for the mostpart, entails providing the plant with a compoundthat is synthesized by the bacterium or facilitating
the uptake of certain nutrients from the environ-ment. The indirect promotion of plant growthoccurs when PGPR decrease or prevent the del-eterious effects of one or more phytopathogenicorganisms.
A. Direct Plant Growth-PromotingActivity
There are several ways in which differentPGPR have been reported to directly facilitate theproliferation of their plant hosts: PGPR can fixatmospheric nitrogen and supply it to plants; theysynthesize siderophores that can solubilize andsequester iron from the soil and provide it to theplant; they synthesize several different phytohor-mones that can act to enhance various stages ofplant growth; they may have mechanisms for thesolubilization of minerals such as phosphorus thatwill become more available for plant growth; andthey may synthesize some less well-character-ized, low-molecular-mass compounds or enzymesthat can modulate plant growth and development(Brown, 1974; Davison, 1988; Kloepper et al.,1986, 1989; Lambert and Joos, 1989; Glick et al.,1994; Patten and Glick, 1996). A particular PGPRmay affect plant growth and development by us-ing any one or more of these mechanisms. It isprobable that the same is true for endophytic bac-teria.
Direct evidence for plant growth-promotingactivity by endophytic bacteria came from Sturz(1995). According to his study, approximately10% of bacterial isolates recovered from withinpotato tubers were shown to promote plant growth.Other experiments with clover and potatoes in acrop rotation revealed that 21% of the isolatedendophytic bacteria promoted plant growth, re-flected in increased shoot height (63%), shoot wetweight (66%), and increased root wet weight(55%) (Sturz et al., 1998).
Probably, the most studied aspect of plantgrowth promotion in endophytic bacteria, as wellas in PGPR, is nitrogen fixation. Diazotrophy iscommon in prokaryotes and the extensive bio-chemical and molecular biological studies of sym-biotic diazotrophs, such as rhizobia, have servedas a starting point for understanding the mecha-

596
nisms of growth promotion involved. Becauseone of the major benefits that rhizobia provide toplants is fixed nitrogen in exchange for fixedcarbon, it was thought initially that diazotrophicPGPR might also function in this way. However,not all PGPR are diazotrophic, and many of thosethat are diazotrophic fix only limited amounts ofnitrogen and not nearly enough to provide fortheir own as well as the host plant’s nitrogenrequirements (Hong et al., 1991). The generallack of bacterial nitrogen release is thought to bethe main reason why nitrogen fixation in the rhizo-sphere only poorly contributes to the nitrogensupply for the whole plant.
Endophytic diazotrophs are presumed to havean advantage over root-associated diazotrophs, suchas Azospirillum and Azotobacter, in that they colo-nize the interior rather than the surface of the plantsand, hence, are better placed to exploit carbonsubstrates supplied by the plant (Döbereinner etal., 1995a, 1995b; McInroy and Kloepper, 1995;Boddey et al., 1995a, 1995b; Sprent and James,1995; Triplett, 1996). Moreover, as they are oftenlocated within underground roots and/or dense planttissue (stem nodes and xylem vessels), the bacteriaare likely to be growing within a low pO2 environ-ment, which is necessary for the expression andoperation of nitrogenase (Patriquin et al., 1983;Gallon, 1992; Baldani et al., 1997).
As all of these assumptions seem to supportthe conditions for nitrogen fixation by endophyticbacteria, there is still no direct evidence that en-dophytic diazotrophs actually are the responsibleagents of biological nitrogen fixation. Althoughsome agriculturally important grasses, such assugar cane (Saccharum sp.), rize (Oryza sativa),wheat (Triticum aestivum), and maize (Zea mays),contain numerous diazotrophic bacteria such asAcetobacter diazotrophicus, Herbaspirillum sp.,and Azospirillum sp., there is little evidence thatthese bacteria actually fix N2 in their host plants(James and Olivares, 1997; Olivares et al., 1996).In the case of sugarcane, no correlation betweenbacterial numbers and biological nitrogen fixa-tion has been demonstrated, and the expression ofnitrogenase by any bacterium has yet to be shownunambiguously (James and Olivares, 1997). Thesituation may be complicated by the possibilitythat endophytic diazotrophs may affect plants in
a manner other than via N2 fixation. Inoculationof rice with the diazotrophic endophytic bacte-rium Azoarcus BH72 significantly promoted plantgrowth (Hurek et al., 1994). In this particularcase, growth promotion also occurred with Nif–
mutants, indicating that N2 fixation by Azoarcussp. � was apparently not involved in plant growthpromotion. Therefore, the authors speculated thatthe observed plant growth promotion might havebeen caused by enhanced plant mineral uptakeand improved plant water relationships associ-ated with the colonization by Azoarcus.
Plant growth regulators, such as ethylene,auxins, or cytokinins, produced by some strainsof Pseudomonas, Enterobacter, Staphylococcus,Azotobacter, and Azospirillum, may also be con-sidered to be causal agents for altering plant growthand development (Arshad and Frankenberger,1991; Bashan and Holguin, 1997; Leifert et al.,1994). Azospirillum, for instance, is generallyregarded as being a rhizosphere bacterium thatcolonizes mainly the elongation and root hair zonesof roots (Okon and Kapulnik, 1986; Döbereinneret al., 1995b; Vande Broek et al., 1998; Bashanand Holguin, 1995). Some Azospirillum strainscan also be endophytic, being found within theroots of some Gramineae. The observed plantgrowth promotion after inoculation of plant rootswith Azospirillum is thought to be the conse-quence of bacterial auxin production and excre-tion (Barbieri and Galli, 1993).
Another way in which plant-associated bacte-ria might influence the host plant has been dis-cussed by Glick et al. (1995, 1998). They demon-strated that many plant growth- promoting bacteriacontain the enzyme 1-aminocyclopropane-1-car-boxylic acid (ACC) deaminase. It was hypoth-esized that this enzyme, which has no knownfunction in bacteria, might be part of a mecha-nism used by certain bacteria to stimulate plantgrowth (Glick et al., 1998). This could occur byACC deaminase modulating the level of ethylenein developing plants (Glick et al., 1994, Glick etal., 1997; Hall et al., 1996). It is also well docu-mented that plants respond to a variety of differ-ent environmental stresses by synthesizing “stress”ethylene (Abeles et al., 1992; Hyodo 1991). Infact, a significant portion of the damage to plantsfrom environmental stress, such as infection with

597
fungal pathogens, may occur as a direct result ofthe response of the plant to the increased level ofstress ethylene (Van Loon, 1984).
Because ACC deaminase may act to ensurethat increased ethylene levels are lowered in adeveloping or stressed plant, it may enhance theplant’s fitness, and thus can be considered to actas a plant growth-promoting characteristic.
B. Indirect Plant Growth-PromotingActivity
Endophytic bacteria, as well as PGPR, seemto be able to lessen or prevent the deleteriouseffects of phytopathogenic organisms, and thisability can be considered to be an indirect promo-tion of plant growth. For instance, seed treatmentof maize with a culture of Burkholderia cepacia,isolated from the rhizosphere of maize, resultedin disease suppression and growth promotion(Bevivino et al., 1998). Similar observations weremade by Hebbar et al. (1992 a and b), who de-scribed an antagonistic effect of B. cepacia againstFusarium moniliforme, a soil-borne fungal patho-gen of maize. A general way in which rhizosphereinhabiting PGPR can prevent the proliferation ofphytopathogens and thereby facilitate plant growthis through the production of siderophores withvery high affinities for iron (Castignetti andSmarrelli, 1986). The secreted siderophore mol-ecules bind most of the Fe3+ that is available in therhizosphere and as a result effectively prevent anypathogens in the immediate vicinity from prolif-erating because of a lack of iron (O’Sullivan andO’Gara, 1992; Buysens et al., 1994). Unlike mi-crobial or fungal phytopathogens, plants aregenerally not harmed by the localized depletionof iron caused by PGPR and a number of plantshave mechanisms for binding the bacterial iron-siderophore complex, transporting it through theplant, and, subsequently, reductively releasing theiron from the siderophore to be used by the plant(Wang et al., 1993).
Another way that a PGPR can be employed toprevent proliferation of phytopathogens is throughthe synthesis of antibiotics. Genetically engineeredstrains of Pseudomonas spp. that produced en-hanced amounts of additional antibiotic metabo-
lites showed improved disease control (Fenton etal., 1992; Bangera and Thomashow, 1996). Incontrast, nonantibiotic-producing mutants of sev-eral different disease-suppressive bacterial strainswere no longer able to prevent phytopathogen-caused damage to plants (Keel et al., 1992). Theseresults supported the hypothesis that antibioticscontribute to the defense against phytopathogens.
Other mechanisms of disease suppression havebeen reviewed by Glick (1995) and involve theproduction of several enzymes, competition fornutrients, and suitable niches on the root surfaceand the induction of systemic resistance by PGPR.
In the case of endophytic bacteria, some studiesreported the ability of fungal suppression. Severalendophytic bacteria isolated from rice seeds exhibitedstrong antifungal activity against Rhizoctonia solani,Pythium myriotylum, Gaeumannomyces graminis, andHeterobasidium annosum (Mukhopadhyay et al.,1996). Enterobacter cloaca, an endophyte isolatedfrom corn, seemed to be involved in antibiosis againstFusarium moniliforme (Hinton and Bacon, 1995),and this result could be added to the experimentsperformed by Chen et al. (1994), who showed thatwilt disease symptoms (caused by a Fusarium sp.)could be reduced successfully when cotton plantswere artificially inoculated with naturally occurringendophytes isolated from the same host plant.
Antagonistic activity of endophytic bacteriaagainst bacterial pathogens has been reported forClavibacter michiganensis subsp. sepedonicum,the causal agent for bacterial rot on tomato (VanBuren et al., 1993). Additionally, Pseudomonasfluorescens 89B-27 and Serratia marcescens 90-166 were observed to induce resistance in cucum-ber to Pseudomonas syringae pv. lachrymans (Liuet al., 1995).
There is also some evidence that endophytesmay contribute to the control of plant-parasiticnematodes (Hallmann et al., 1995) and insects(Dimock et al., 1988). Rhizospheric and endo-phytic bacteria, associated with some crops withantagonistic properties to phytopathogenic nema-todes, have been demonstrated to be involved incontrolling these pests (Kloepper et al., 1991b). Itshould be mentioned that control of these para-sites seems to be more complex and difficult thanfor fungal and bacterial pathogens, because dam-age from nematodes and insects occurs as a result

598
of their feeding habits and migration, thus limit-ing the efficacy of bacterial antagonism.
C. Endophytic Bacteria from Thlaspicaerulescens
A special niche for plant-associated bacteriais constituted by heavy metal-accumulating plants.The presence of plant-associated bacteria, eitherin the rhizosphere or as endophytes, will requirespecific adaptation of these bacteria, such as ad-aptation to high levels of heavy metals. Addition-ally, characterization of these bacteria and theirheavy metal resistance phenotypes can also pro-vide valuable information on the bioavailabilityof heavy metals in the rhizosphere and within thehyperaccumulator plant. Delorme et al. (2001)studied the effects of Thlaspi caerulescens ontotal (culturable) bacteria, total fungi, as well ascadmium- and zinc-resistant populations innonrhizosphere and rhizosphere soil. Microbialnumbers in rhizosphere soil were increased, com-pared with those in nonrhizosphere soil. Also, theratios of metal-resistant bacteria found in the pres-ence of T. caerulescens were significantly higherthan those observed in the rhizosphere of Trifo-lium pratense L. It was hypothesized thatT. caerutescens acidifies its rhizosphere, whichwould affect metal uptake by increasing availablemetals around the roots and, consequently, in-crease the selection for metal-resistant bacteria.Whiting et al. (2001) demonstrated that the rhizo-sphere bacteria associated with T. caerulescensincreased the amount of water-soluble zinc in thesoil and, as such, helped to enhance the zincaccumulation in T. caerulescens shoots.
Lodewyckx et al. (2002) studied the bacterialpopulations associated with Thlaspi caerulescenssubsp. calaminaria grown in soil collected fromthe abandoned Zn and Pb mining and smelter siteof Plombières (Belgium). The nonrhizosphericsoil representing population consisted of bacteriaequipped with multiple heavy metal resistancecharacteristics: 7.8% and 4% of the cultivablebacteria were able to survive elevated levels ofthe major pollutants Zn (1 mM) and Cd (0.8 mM).As observed by Delorme et al. (2001), the rhizo-sphere population was well adapted to heavy
metals and showed a survival rate of 88% and78% on the respective selective heavy metals,which indicates a difference in metal availabilityin the vicinity of the root compared to thenonrhizospheric soil. Characterization and identi-fication of the inhabitants of the roots and shootsdemonstrated that although similar species wereisolated in both tissues, some differences could benoted. In the presence of Zn and Cd, rhizoplaneand root endophytic isolates were observed tohave a much lower survival rate under the sameconditions and root endophytic bacteria evenseemed to have different growth requirements.Additionally, only some bacteria residing in theseplant root compartments were able to producesiderophores under iron limiting conditions. Con-trary to the root-residing inhabitants, the shootrepresented a niche rich in metal-resistant bacte-ria and even seemed to contain species that wereexclusively abundant in this environment. Thesedifferences in the characteristics of the bacterialmicroflora associated with T. caerulescens mightreflect altered metal speciation in the differentcompartments studied (Lodewyckx et al., 2002).
IV. PRACTICAL APPLICATIONS
The potential that endophytes have to offerfor agricultural gains has been realized beyondtheir use as natural biocontrol agents. The inher-ent nature of certain endophytes to potentiallycolonize plants in a systemic manner provides anovel approach as a delivery system to plants forvarious beneficial traits.
A. Enhanced Pest Control
The use of genetically engineered strains ofendophytic bacteria for enhanced pest control hasbeen envisioned for some time (Dimock et al.,1988; Misaghi and Donndelinger, 1990). How-ever, several limitations hinder their rapid devel-opment. The selection of endophytes to constructgenetically engineered organisms requires a vari-ety of traits necessary for the construction of ef-fective strains. First, the endophyte must resideat, or deliver, the pest-controlling factor to a site

599
within the plant that is accessible to the pest.Second, the endophyte must not have significantcrop-threatening disease capabilities. Third, theendophyte must be readily amenable to geneticmanipulation. Several additional factors must beconsidered in the construction of recombinantmicrobes, including the ability to readily expressforeign genes and the long-term maintenance ofthe gene(s) within the endophyte. This is mosteasily performed by stable integration into thechromosome of the endophyte, but it requiresintegration of a compatible form of the gene at asite that does not affect the fitness of the bacte-rium. Once incorporated, the gene product mustbe expressed in a form that is high enough inconcentration and functionally accessible to thepest.
The most intensive study of a recombinantbacterial endophyte constructed for pest controlpurposes was performed with Clavibacter xylisubsp. cynodontis, expressing the cryIA gene fromBacillus thuringiensis (Lampel et al., 1994;Tomasino et al., 1995). In this example, the re-sultant protein was an endotoxin that is activeagainst the European corn borer. The recombi-nant bacterium was effective in reducing insectboring in corn in laboratory, greenhouse, and fieldstudies. However, significant increases in overallyield were not detected compared with plants thatwere not treated with the endophyte.
B. Improved Phytoremediation
The use of biological techniques can stronglyreduce the costs of remediating sites contami-nated with organic xenobiotics. For large con-taminated sites, bioremediation is the only alter-native economically and socially acceptable. Inparticular, phytoremediation, one of the “soft”bioremediation techniques, is becoming an ac-ceptable alternative for the treatment of contami-nated sites and wastewater. Phytoremediation oforganic contaminants is based on the combinedaction between plants and their associated micro-organisms, such as mycorrhizal fungi and bacte-ria. Soil contaminants, especially organicxenobiotics with a log Kow between 0.5 and 3.5and weak electrolytes (weak acids and bases or
amphoteres such as herbicides) are readily takenup by plants (Trapp et al., 1994; Trapp, 2000).Recent unpublished evidence suggests that nu-merous compounds enter the xylem faster thanthe soil microflora can degrade them, even if therhizosphere is enriched with degrader bacteria.Plant-bacterial combinations can increase con-taminant degradation in the rhizosphere, althoughthe role played by indigenous root-associatedbacteria during plant growth in contaminated soilsis unclear. Siciliano et al. (2001) examined whetherplants had the ability to selectively enhance theprevalence of endophytes containing pollutantcatabolic genes in unrelated environments con-taminated with different pollutants. Their resultsindicated that the enrichment of catabolic geno-types in the root interior is both plant and con-taminant dependent (Siciliano et al., 2001): selec-tive enrichment for endophytic bacteria containingthe appropriate catabolic genes was observed inplants grown on petroleum hydrocarbon-contami-nated sites, on field sites contaminated withnitroaromatics, and after the addition of petroleumto sediment. Similarly, the numbers of alkB- orndoB-positive endophytes in Festuca arundinaceawere positively correlated with the concentrationof creosote in the soil but not with the numbers ofalkB- or ndoB-positive bacteria in the bulk soil.
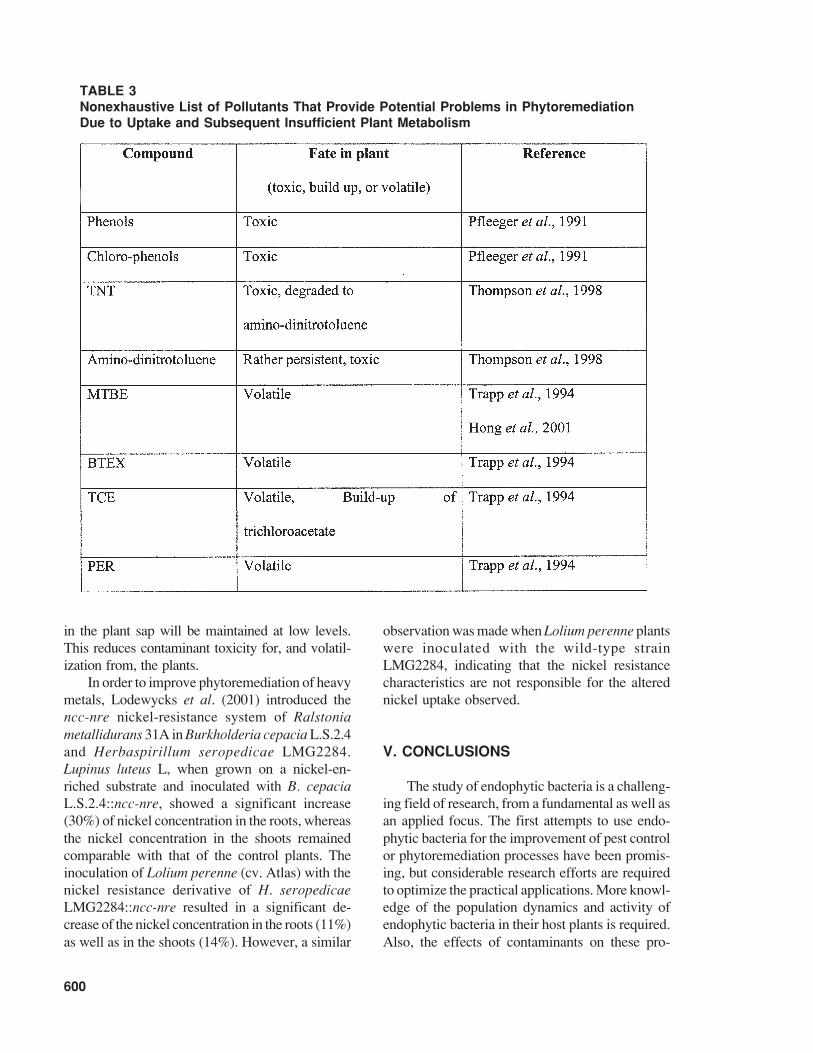
Certain organic pollutants may not be degraded,but may be accumulated in the plant (see also Table 3).Alternatively, in the case of volatile pollutants, plantsrelease these compounds, or their metabolites,through the stomata, which questions the merits ofphytoremediation in such cases. Additionally, wa-ter-soluble and volatile organic pollutants might bepartially degraded by plants and, subsequently, ac-cumulation of toxic metabolites can occur. Recently,the European Commision funded a project(ENDEGRADE, QLK3-CT2000-00164 [http://endegrade.dmu.dk]) that has as its objective to as-certain whether genetically modified endophyticbacteria equipped with catabolic pathways can beutilized to improve phytoremediation of organicpollutants. It is hypothesized that endophytic bacte-ria can be made to degrade toxic organic compoundswithin the plant vascular system, thereby assistingthe plants in metabolizing soil contaminants. As aresult, even when the plants are taking up pollutantsat a high rate, the concentration of toxic compounds
600
in the plant sap will be maintained at low levels.This reduces contaminant toxicity for, and volatil-ization from, the plants.
In order to improve phytoremediation of heavymetals, Lodewycks et al. (2001) introduced thencc-nre nickel-resistance system of Ralstoniametallidurans 31A in Burkholderia cepacia L.S.2.4and Herbaspirillum seropedicae LMG2284.Lupinus luteus L, when grown on a nickel-en-riched substrate and inoculated with B. cepaciaL.S.2.4::ncc-nre, showed a significant increase(30%) of nickel concentration in the roots, whereasthe nickel concentration in the shoots remainedcomparable with that of the control plants. Theinoculation of Lolium perenne (cv. Atlas) with thenickel resistance derivative of H. seropedicaeLMG2284::ncc-nre resulted in a significant de-crease of the nickel concentration in the roots (11%)as well as in the shoots (14%). However, a similar
observation was made when Lolium perenne plantswere inoculated with the wild-type strainLMG2284, indicating that the nickel resistancecharacteristics are not responsible for the alterednickel uptake observed.
V. CONCLUSIONS
The study of endophytic bacteria is a challeng-ing field of research, from a fundamental as well asan applied focus. The first attempts to use endo-phytic bacteria for the improvement of pest controlor phytoremediation processes have been promis-ing, but considerable research efforts are requiredto optimize the practical applications. More knowl-edge of the population dynamics and activity ofendophytic bacteria in their host plants is required.Also, the effects of contaminants on these pro-
TABLE 3Nonexhaustive List of Pollutants That Provide Potential Problems in PhytoremediationDue to Uptake and Subsequent Insufficient Plant Metabolism

601
cesses should be evaluated further. For the con-struction of endophytic strains with new catabolicfunctions, natural gene transfer offers great poten-tial. Many catabolic functions are found in soilbacteria, where they are encoded on self-transfer-able plasmids or transposons. These can be easilyexchanged through natural gene transfer. Heterolo-gous expression of the catabolic functions mightnot be a major problem, especially when the donorstrain and the recipient endophytic strain are closelyrelated, as is frequently the case. However, for thesuccessful treatment of organic xenobiotics byendophytic bacteria, it might be necessary to usemutant strains that constitutively express meta-bolic pathways. This to avoid problems of induc-tion due to low concentrations of the contaminant.
A considerable research effort is also required todesign strategies for the reinoculation of endophyticbacteria. In order to guarantee reproducibility, reliablemethods of inoculum delivery should be developed.This is especially the case for the inoculation of trees,such as poplar, with endophytic bacteria. Intense test-ing of different delivery systems has indicated that theefficacy of the application method for introducing en-dophytic bacteria into plant tissue is strain specific(Musson et al., 1995). Some methods that have provento be successful include the infusion of a bacterialsuspension into imbibed seeds (Turner et al., 1993) orthe application of bacteria via alginate beads (Bashan1986), which have the advantage of adding bacteria-specific nutrients to the alginate to improve bacterialsurvival rates. It should be noted that the developmentof successful application technologies would fully de-pend on improving our understanding of how bacterialendophytes enter and colonize plants. This remarkcould be applied to all aspects of the ecology of bacte-rial endophytes and only under those circumstancescan the potential use of bacterial endophytes for plantbeneficial purposes be fully evaluated.
ACKNOWLEDGMENT
The authors wish to acknowledge the Euro-pean Commission for their support to theENDEGRADE project (EU PROJECT: QLK3-CT2000-00164). DvdL and ST are presently be-ing supported by Laboratory Directed Researchand Development funds at the Brookhaven
National Laboratory under contract with the U.S.Department of Energy.
REFERENCES
Abeles, F.B., Morgan, P.W., and Saltveit, M.E. 1992. Regu-lation of ethylene production by internal, environ-mental and stress factors. In: Ethylene in Plant Biol-ogy, 2nd ed. pp. 56–119. Academic Press, San Diego,CA.
Araujo, W.L., Maccheroni, W., Aguilar-Vildosa, C.I.,Barrosa, P.A.V., Saridakis, H.O., and Azevedo, J.L.2001. Variability and interactions between endophyticbacteria and fungi isolated from leaf tissues of citrusrootstocks. Can. J. Microbiol. 47:229–236.
Arshad, M. and Frankenberger, W.T. 1991. Microbial pro-duction of plant hormones. In: The Rhizosphere andPlant Growth. pp. 327–334. Keister, P.B. and Cregan,P.B., Eds., Kluwer Academic Publishers, Dordrecht,the Netherlands.
Ashbolt, N.J. and Inkerman, P.A. 1990. Acetic acid bacterialbiota of the pink sugar cane mealybug Saccharococcussacchari, and its environs. Appl. Environ. Microbiol.56:707–712.
Baldani, J.I., Caruso, L., Baldani, V.L.D., Goi, S.R., andDöbereinner, J. 1997. Recent advances in BNF withnon-legume plants. Soil Biol. Biochem. 29:911–922.
Bangera, M.G. and Thomashow, L.S. 1996. Characterizationof a genomic locus required for synthesis of the anti-biotic 2,4–diacetylphloroglucinol by the biologicalcontrol agent Pseudomonas fluorescens Q2–87. Mol.Plant-Microbe Interact. 9:83–90.
Barbieri, P. and Galli, E. 1993. Effect on wheat developmentof inoculation with an Azospirillum brasilense mutantwith altered indole-3–acetic acid production. Res.Microbiol. 144:69–75.
Bashan, Y. 1986. Alginate beads as synthetic inoculant car-riers for slow release of bacteria that affect plantgrowth. Appl. Environ. Microbiol. 51:1089–1098.
Bashan, Y. and Holguin, G. 1995. Inter-root movement ofAzospirillum brasilense and subsequent root coloni-zation of crop and weed seedlings growing in soil.Microbiol. Ecol. 29:269–281.
Bashan, Y. and Holguin, G. 1997. Azosprillum-plant rela-tionships:environmental and physiological advances(1990–1996). Can. J. Microbiol. 43:103–121.
Bell, C.R., Dickie, G.A., Harvey, W.L.G., and Chan, J.W.Y.F.1994. Endophytic bacteria in grapevine. Can. J.Microbiol. 41:46–53.
Benhamou, N., Bélanger, R.R., and Paulitz, T.C. 1996a. Pre-inoculation of Ri T-DNA-transformed pea roots withPseudomonas fluorescens inhibits colonization byPhyhium ultimum Trow:an ultrastructural and cy-tochemical study. Planta 199:105–117.
Benhamou, N., Kloepper, J.W., Quadt-Hallmann, A., andTuzun, S. 1996b. Induction of defense related ultra-

602
structural modifications in pea root tissues inoculatedwith endophytic bacteria. Plant Physiol. 112:919–929.
Bevivino, A., Sarrocco, S., Dalmastri, C., Tabacchioni, C.,Cantale, C., and Chiarini, L. 1998. Characterizationoff a free-living maize-rhizosphere population ofBurkholderia cepacia:effect of seed treatment on dis-ease suppression and growth promotion of maize.FEMS Microbiol. Ecol. 27:225–237.
Boddey, R.M., Urquiaga, S., de Oliveira, O.C., Baldani,V.L.D., and Döbereinner, J. 1995a. Biological nitro-gen fixation associated with sugar cane and rice:con-tributions and prospects for improvement. Plant Soil174:195–209.
Boddey, R.M., Reis, V.M., Urquiaga, S., da Silva, L.G., dosReis, F.B., Baldani, J.I., and Döbereinner, J. 1995b.N2 fixation in sugar cane:the role of Acetobacterdiazotrophicus. In: Nitrogen Fixation: Fundamentalsand Applications. pp. 641–646. Tikhonovich, I.A.,Rovorov, N.A., and Newton, W.E., Eds., Kluwer Aca-demic, Dordrecht.
Brooks, D.S., Gonzalez, C.F., Appel, D.N., and Filer, T.H.1994. Evaluation of endophytic bacteria as potentialbiological control agents for oak wilt. Biol. Control.4:373–381.
Brown, M.E. 1974. Seed and root bacterization. Annu. Rev.Phytopathol. 12:181–197.
Buysens, S., Poppe, J., and Höfte, M. 1994. Role ofsiderophores in plant growth stimulation and antago-nism by Pseudomonas aeruginosa 7NSK2. In: Im-proving Plant Productivity with Rhizosphere Bacte-ria. pp. 139–141. Ryder, M.H., Stephens, P.M., andBowen, G.D., Eds., CSIRO, Adelaide, Australia.
Castignetti, D. and Smarelli, J., Jr. 1986. Siderophores, theiron nutrition of plants, and nitrate reductase. FEBSLett. 209:147–151.
Chen, C., Banske, E.M., Musson, and G., Kloepper, J.W.1994. Biological conrol potential and population dy-namics of endophytic bacteria in a cotton/ Fusariumwilt system. In: Improving Plant Productivity withRhizosphere Bacteria. pp. 191–193. Ryder, M.H.,Stephens, P.M., and Bowen, G.D., Eds., CSIRO,Adelaide, Australia.
Davison, J. 1988. Plant beneficial bacteria. Biotechnology6:282–286.
De Boer, S.H. and Copeman, R.J. 1974. Endophytic bacte-rial flora in Solanum tuberosum and its significance inbacterial ring rot diagnosis. Can. J. Plant Sci. 54:115–122.
Delorme, T. A., Gagliardi, J. V., Angle, J. S., and Chaney, R.L. 2001. Influence of the zinc hyperaccumulatorThlaspi caerulescens J. & C. Presl. and the nonmetalaccumulator Trifolium pratense L. on soil microbialpopulations. Can. J. Microbiol. 47:773–776.
Di Fiori, S. and Del Gallo, M. 1995. Endophytic bacteria:theirpossible role in the host plant. In: Azospirillum VI andRelated Microorganisms. pp. . Fendrik, I., Del Gallo,M., Vanderleyden, J., and de Zamaroczy, M., Eds.,Springer-Verlag, Berlin.
Dimock, M.B., Beach, R.M., and Carlson, P.S. 1988. Endo-phytic bacteria for the delivery of crop protectionagents. In: Biotechnology, Biological Pesticides andNovel Plant-Pest Resistance for Insect Pest Manage-ment. pp. 88–92. Roberts, D.W. and Granados, R.R.,Eds., Boyce Thompson Institute for Plant Research,Ithaca, NY.
Döbereinner, J., Urquiaga, S., and Boddey, R.M. 1995a.Alternatives for nitrogen nutrition of crops in tropicalagriculture. Fert. Res. 42:339–346.
Döbereinner, J., Baldani, V.L.D., and Reis, V.M. 1995b.Endophytic occurrence of diazotrophic bacteria in non-leguminous crops. In: Azospirillum VI and RelatedMicroorganisms. pp. 3–14. Fendrik, I., Del Gallo, M.,Vanderleyden, J., and de Zamaroczy, M., Eds.,Springer-Verlag, Berlin.
Dong, Z., Canny, M.J., Mc Cully, M.E., Roboredo, M.R.,Cabadilla, C.F., Ortega, E., and Rodés, R. 1994. Anitrogen fixing endophyte of sugarcane stems. PlantPhysiol. 105:1139–1147.
Duijff, B.J., Gianinazzi-Pearson, V., and Lemanceau, P. 1997.Involvement of the outer membrane lipopolysaccha-rides in the endophytic colonization of tomato rootsby biocontrol Pseudomonas fluorescens strainWCS417r. New Phytologist 135:325–334.
Elbeltagy, A., Nishioka, K, Suzuki, H., Sato, T., Sato, Y-I.,Morisaki, H., Mitsui, H., and Minamisawa, K. 2000.Isolation and characterisation of endophytic bacteriafrom wild and traditionally cultivated rice varieties.Soil Sci. Plant Nutrition 46:617–629.
Fenton, A.M., Stephens, P.M., Crowley, J., O’Callaghan,M., and O’Gara, F. 1992. Exploitation of gene(s)involved in 2,4–diacetylphloroglucinol biosynthesisto confer a new biocontrol capability to a Pseudomo-nas strain. Appl. Environ. Microbiol. 58:3873–3878.
Fisher, P.J., Petrini, O., and Lappin, S.H.M., 1992. Thedistribution of some fungal and bacterial endophytesin maize (Zea mays L.). New Phytologist 122:299–305.
Frommel, M.I., Nowak, J., and Lazarovits, G. 1991. Growthenhancement and developmental modifications of invitro grown potato (Solanum tuberosum ssp.Tuberosum) as affected by a nonfluorescent Pseudomo-nas sp. Plant Physiol. 96:928–936.
Gagné, S., Richard, C., Rousseau, H., and Antoun, H. 1987.Xylem residing bacteria in alfalfa roots. Can. J.Microbiol. 33:996–1000.
Gallon, J.R. 1992. Reconciling the incompatible:N2 fixationand O2. New Phytol. 122:571–609.
Gardner, J.M., Feldman, A.W., and Zablotowicz, M. 1982.Identity and behavior of xylem-residing bacteria inrough lemon roots of Florida citrus trees. Appl. Environ.Microbiol. 43:1335–1342.
Germida, J.J., Siciliano, S.D., and Seib, A.M. 1998. Pheno-typic plasticity of Pseudomonas aureofasciens (lacZY) introduced into and recovered from field andlaboratory microcosm soils. FEMS Microbiol. Ecol.27:133–139.

603
Glick, B.R, Jacobson, C.B., Schwarze, M.K., and Pasternak,J.J. 1994. 1–Aminocyclopropane-1–carboxylic aciddeaminase mutants of the plant growth promotingrhizobacterium Pseudomonas putida GR 12–2 do notstimulate canola root elongation. Can. J. Microbiol.40:911–915.
Glick, B.R. 1995. The enhancement of plant growth by free-living bacteria. Can. J. Microbiol. 41:109–117.
Glick, B.R, Liu, C., Ghosh, S., and Dumbroff, E.B. 1997.Early development of canola seedlings in the presenceof the plant growth-promoting rhizobacteriumPseudomonas putida GR 12–2. Soil Biol. Biochem.29:1233–1239
Glick, B.R., Penrose, D.M., and Li, J., 1998. A model for thelowering of plant ethylene concentrations by plantgrowth promoting bacteria. J. Theor. Biol. 190:63–68.
Grimault, V. and Prior, P. 1994. Invasiveness of Pseudomo-nas solanacearum in tomato, eggplant, and pepper:acomparative study. Eur. J. Plant Pathol. 100:259–267.
Gutierrez-Zamora, M. L. and Martinez-Romero, E. 2001.Natural endophytic association between Rhizobiumetli and maize (Zea mays L.). J. Biotechnol. 91:117–126.
Hall, J.A., Peirson, D., Ghosh, S., Glick, B.R. 1996. Rootelongation in various agronomic crops by the plantgrowth promoting rhizobacterium Pseudomonas putidaGR12–2. Isr. J. Plant Sci. 44:37–42.
Hallmann, J., Kloepper, J.W., Rodriguez-Kabana, R., andSikora, R.A. 1995. Endophytic rhizobacteria as an-tagonists of Meloidogyne incognita on cucumber.Phytopathology 85:1136.
Hebbar, K.P., Davey, A.G., Merrin, J., and Dart, P.J. 1992a.Rhizobacteria of maize antagonistic to Fusariummoniliforme, a soil-borne fungal pathogen:coloniza-tion of rhizosphere and roots. Soil Biol. Biochem.24:989–997.
Hebbar, K.P., Davey, A.G., Merrin, J., Mc Loughlin, T.J.,and Dart, P.J. 1992b. Pseudomonas cepacia, a poten-tial suppressor of maize soil-borne disease-seed in-oculation and maize root colonization. Soil Biol.Biochem. 24:999–1007.
Hinton, D.M. and Bacon, C.W. 1995. Enterobacter cloacaeis an endophytic symbiont of corn. Mycopathology129:117–125.
Höflich, G., Wiehe, W., and Kühn, G. 1994. Plant growthstimulation by inoculation with symbiotic and associa-tive rhizosphere mechanisms. Experientia 50:897–905.
Hollis, J.P 1951. Bacteria in healthy potato tissue. Phytopa-thology 41:350–366.
Hong M.S., Farmayan, W.F., Dortch, I. J., Chiang, C. Y.,McMillan, S. K., and Schnoor, J. L. 2001.Phytoremediation of MTBE from a groundwaterplume. Environ. Sci. Technol. 35:1231–1239.
Hong, Y., Pasternak, J.J., and Glick, B.R. 1991. Biologicalconsequences of plasmid transformation of the plantgrowth promoting rhizobacterium Pseudomonas putidaGR12–2. Can. J. Microbiol. 37:796–799.
Hurek, T., Reinhold-Hurek, B., Van Montagu, M., andKellenberger, E. 1994. Root colonization and sys-temic spreading of Azoarcus sp. strain BH72 in grasses.J. Bacteriol. 176:1913–1923.
Hyodo, H. 1991. Stress/wound ethylene. In: The Plant Hor-mone Ethylene. pp. 151–189. Mattoo, A.K. and Suttle,J.C., Eds., CRC Press, Inc., Boca Raton, Fla., USA.
Jacobs, M.J., Bugbee, W.M., and Gabrielson, D.A. 1985.Enumeration, location, and characterization of endo-phytic bacteria within sugar beet roots. Can. J. Botany63:1262–1265.
James, E.K., Reis, V.M., Olivares, F.L., Baldani, J.I., andDöbereinner, J. 1994. Infection of sugar cane by thenitrogen fixing bacterium Acetobacter diazotrophicus.J. Exp. Bot. 45:757–766.
James, K. and Olivares, F.L. 1997. Infection and colonizationof sugar cane and other Graminaceous plants by endo-phytic diazotrophs. Crit. Rev. Plant Sci. 17:77–119.
Kado, C.I. 1992. Plant pathogenic bacteria. In: The Prokary-otes. pp. 660–662. Ballows, A., Trüper, G.G., Dworkin,M., Harder, W., and Schleifer, K.-H., Eds., Springer-Verlag, New York.
Keel, C., Schnider, U., Maurhofer, M., Voisard, C., Laville,J., Burger, U., Wirthner, P., Haas, D., and Défago, G.1992. Suppression of root diseases by Pseudomonasfluorescens CHAO:importance of the bacterial sec-ondary metabolite 2,4–diacetylphloroglucinol. Mol.Plant-Microbe Interact. 5:4–13.
Khammas, K.M. and Kaiser, P. 1991. Characterization of apectinolytic activity in Azospirillum irakense. PlantSoil 137:75–79.
Kloepper, J.W., Scher, F.M., Laliberte, M., and Tipping, B.1986. Emergence-promoting rhizobacteria: descriptionand implications for agriculture. In: Iron, Siderophores,and Plant Disease. pp. 155–164. Swinburne, Eds., Ple-num Press, New York.
Kloepper, J.W., Lifshitz, R., and Zablotowicz, R.M. 1989.Free-living bacterial inocula for enhancing crop pro-ductivity. Trends Biotechnol. 7:39–43.
Kloepper, J.W., Tipping, E.M., and Lifshitz, R. 1991a. Plantgrowth promotion mediated by bacterial rhizospherecolonizers. In: The Rhizosphere and Plant Growth. pp.315–326. Keister, D.L. and Cregan, P.B., Eds., KluwerAcademic Publishers, Dordrecht, The Netherlands.
Kloepper, J.W., Rodriguez-Kabana, R., McInroy, J.A., andCollins, D.J. 1991b. Analysis of populations and physi-ological characterization of microorganisms in rhizo-spheres of plants with antagonistic properties tophytopathogneic nematodes. Plant Soil 136:95–102.
Kloepper, J.W. and Beauchamp, C.J. 1992. A review ofissues related to measuring colonization of plant rootsby bacteria. Can. J. Microbiol. 38:1219–1232.
Kloepper, J.W., Schippers, B., and Bakker, P.A.H.M. 1992.Proposed elimination of the term endorhizosphere.Phytopathology 82:726–727.
Kluepfel, D.A. 1993. The behavior and tracking of bacteriain the rhizosphere. Ann. Rev. Phytopathol. 31:441–472.

604
Kobayashi, D.Y. and Palumbo, J.D. 2000. Bacterial Endo-phytes and Their Effects on Plants and Uses in Agri-culture. Bacon, C.W. and White, J.F. Jr., Eds., MarcelDekker, New York.
Lalande, R., Bissonnette, N., Coutlée, D., and Antoun, H.1989. Identification of rhizobacteria from maize anddetermination of their plant-growth promoting poten-tial. Plant Soil 115:7–11.
Lamb, T.G., Tonkyn, D.W., and Kluepfel, D.A. 1996. Move-ment of Pseudomonas aureofaciens from the rhizo-sphere to aerial plant tissue. Can. J. Microbiol.42:1112–1120.
Lambert, B. and Joos, H. 1989. Fundamental aspects ofrhizobacterial plant growth promotion research. TrendsBiotechnol. 7:215–219.
Lampel, J.S., Canter, G.L., Dimock, M.B., Kelly, J.L., Ander-son, J.J., Uratani, B.B., Foulke, J.S., Jr., and Turner,J.T. 1994. Integrative cloning, expression, and stabil-ity of the cryIA. gene from Bacillus thuringiensissubsp. kurstaki in a recombinant strain of Clavibacterxyli subsp. cynodontis. Appl. Environ. Microbiol.60:501–508.
Leifert, C., Morris, C.E., and Waites, W.M. 1994. Ecologyof microbial saprophytes and pathogens in tissue cul-ture and field grown plants:reasons for contaminationproblems in vitro. Crit. Rev. Plant Sci. 13:139–183.
Levanoy, H., Bashan, Y., Romano, B., and Klein, E. 1989.Ultrastructural localization and identification ofAzospirillum brasilense on and within wheat roots byimmunogold labeling. Plant Soil 117:207–218.
Liu, L., Kloepper, J.W., and Tuzun, S. 1995. Induction ofsystemic resistance in cucumber against bacterial angu-lar leaf spot by plant growth-promoting rhizobacteria.Phytopathology 85:843–847.
Lodewyckx, C., Taghavi, S., Mergeay, M., Vangronsveld, J.,Clijsters, H., and van der Lelie, D. 2001. The effect ofrecombinant heavy metal resistant endophytic bacte-ria on heavy metal uptake by their host plant. Int. J.Phytoremed. 3:173–187.
Lodewyckx, C., Mergeay, M., Vangronsveld, J., Clijsters,H., and van der Lelie, D. 2002. Isolation, characteriza-tion and identification of bacteria associated to thezinc hyperaccumulator Thlaspi caerulescens subsp.calaminaria. Int. J. Phytorem. 4:101–115.
Mahaffee, W.F., Kloepper, J.W., Van Vuurde, J.W.L., Vander Wolf, J.M., and Van den Brink, M. 1997. Endo-phytic colonization of Phaseolus vulgaris byPseudomonas fluorescens strain 89B-27 andEnterobacter asburiae strain JM22. In: ImprovingPlant Productivity in Rhizosphere Bacteria. RyderM.H., Stephens P.M., and Bowen G.D., Eds., CSIRO,Melbourne, Australia.
Mahaffee, W.F. and Kloepper, J.W. 1997. Temporal changesin the bacterial communities of soil, rhizosphere, andendorhiza associated with field-grown cucumber(Cucumis sativus L.). Microb. Ecol. 34:210–223.
McInroy, J.A. and Kloepper, J.W. 1994. Novel bacterial taxainhabiting internal tissue of sweet corn and cotton. In:
Improving Plant Productivity with Rhizosphere Bac-teria. pp. 190. Ryder, M.H., Stephens, P.M., andBowen, G.D., Eds., CSIRO, Melbourne, Australia.
McInroy, J.A. and Kloepper, J.W. 1995. Survey of indig-enous bacterial endophytes from cotton and sweetcorn. Plant Soil 173:337–342.
Misaghi, I.J. and Donndelinger, C.R. 1990. Endophytic bac-teria in sympton free cotton plants. Phytopathology80:808–811.
Mukhopadhyay, N.K., Garrison, N.K., Hinton, D.M., Bacon,C.W., Khush, G.S., Peck, H.D., and Datta, N. 1996.Identification and characterization of bacterial endo-phytes of rice. Mycopathology 134:151–159.
Musson, G., McInroy, J.A., and Kloepper, J.W. 1995. Devel-opment of delivery systems for introducing endophyticbacteria into cotton. Biocontrol Sci. Technol. 5:407–416.
Olivares, F.L., Baldani, V.L.D., Reis, V.M., Baldani, J.I.,and Döbereiner J. 1996. Occurrence of the endophyticdiazotrophs Herbaspirillum spp. in roots, stems, andleaves predominantly of Gramineae. Biol. Fertil. Soils21:197–200.
Okon, Y. and Kapulnik, Y. 1986. development and functionof Azospirillum-inoculated roots. Plant Soil 90:3–16.
O’Sullivan, D.J. and O’Gara, F. 1992. Traits of fluorescentPseudomonas spp. involved in suppression of plantroot pathogens. Microbiol. Rev. 56:662–676.
Pan, M.J., Rademan, S., Kuner, K., and Hastings, J.W. 1997.Ultrastructural studies on the colonization of bananatissue and Fusarium oxysporum f. sp. cubense race 4by the endophytic bacterium Burkholderia cepacia. J.Phytopathol. 145:479–486.
Patten, C.L. and Glick, B.R. 1996. Bacterial biosynthesis ofindole-3–acetic acid. Can. J. Microbiol. 42:207–220
Patriquin, D.G. and Döbereinner, J. 1978. Light microscopyobservations of tetrazolium-reducing bacteria in theendorhizosphere of maize and other grasses in Brazil.Can. J. Microbiol. 24:734–742.
Patriquin, D.G. Döbereinner, J., and Jain, D.K. 1983. Sitesand processes of association between diazotrophs andgrasses. Can. J. Microbiol. 29:900–915.
Pfleeger, T. 1991. A short-term bioassay for whole planttoxicity. In: Plants for Toxicity Assessment: SecondVolume, ASTM STP 1115. pp. 355–364. Gorsuch,J.W., Lower, R.W., Wang, W., and Lewis, M.A.,Eds., American Society for Testing and Materials,Philadelphia, USA.
Quadt-Hallmann, A. and Kloepper, J.W. 1996. Immunologi-cal detection and localization of the cotton endophyteEnterobacter asburiae JM22 in different plant spe-cies. Can. J. Microbiol. 42:1144–1154.
Quadt-Hallmann, A., Benhamou, N., and Kloepper, J.W.,1997. Bacterial endophytes in cotton:mechanismsentering the plant. Can. J. Microbiol. 43:577–582.
Quispel, A. 1992. A search for signals in endophytic micro-organisms. In: Molecular Signals in Plant-MicrobeCommunications. pp. 471–490. D.P.S. Verma, D.P.S.,Eds., CRC Press, Boca Raton, FL.

605
Reddy, P.M., Ladha, J.K., SO, R.B., Hernandez, R.J., Ramos,M.C., Angeles, O.R., Dazzo, F.B., and De Bruijn, F.J.1997. Rhizobial communications with rice roots:in-ductions of phenotypic changes, mode of invasion andextent of colonization. Plant Soil 194:81–98.
Reinhold-Hurek, B. and Hurek, T. 1997. Azoarcus spp. andtheir interactions with grass roots. Plant Soil 194:57–64.
Reinhold-Hurek, B. and Hurek, T. 1998. Interactions ofGramineous plants with Azoarcus spp. and otherdiazotrophs:identification, localization, and perspec-tives to study their function. Crit. Rev. Plant Sci.17:29–54.
Samish, Z., Etinger-Tulczynska, R., and Bick, M., 1963. Themicroflora within the tissue soft fruit and vegetables.J. Food Sci. 28:259–266.
Sessitsch, A., Reiter, B., Pfeifer, U., and Wilhelm, E. 2001.Analysis of endophytic bacteria in three potato culti-vars. In: Proceedings 9th meeting of the InternationalSociety of Molecular Ecology, Amsterdam, August2001:abstract nr.:Tu.052.
Sharrock, K.R., Parkes, S.L., Jack, H.K., Rees-George, J.,and Hawthorne, B.T. 1991. Involvement of bacterialendophytes in storage rots of buttercup squash(Cucurbita maxima D. hybrid ‘Delicia’). New ZealandJ. Crop Horticultural Sci. 19:157–165.
Siciliano, S. D., Fortin, N, Mihoc, A., Wisse, G., Labelle, S.,Beaumier, D., Ouelette, D., Roy, R., Whyte, L. G.,Banks, M. K., Schwab, P., Lee, K., and Greer, C. W.2001. Selection of specific endophytic bacterial geno-types by plants in response to soil contamination.Appl. Environ. Microbiol. 67:2469–2475.
Sprent, J.I. and de Faria, S.M. 1988. Mechanisms of infec-tion of plants by nitrogen fixing organisms. Plant Soil110:157–165.
Sprent, J.I. and James, E.K. 1995. N2 fixation by endophyticbacteria:questions of entry and operation. In:Azospirillum VI and Related Microorganisms. pp. 15–30. Fendrik, I., Del Gallo, M., Vanderleyden, J., andde Zamaroczy, M., Eds., Springer-Verlag, Berlin.
Stoltzfus, J.R., SO, R., Malarvithi, P.P., Ladha, J.K., and deBruijn, F.J. 1997. Isolation of endophytic bacteriafrom rice and assessment of their potential for supply-ing rice with biologically fixed nitrogen. Plant Soil194:25–36.
Sturdy, M.L. and Cole, L.J. 1974. Studies on endogenousbacteria in potato tubers infected by Phytophorainfestans Pethybr. Ann. Bot. 8:121–127.
Sturz, A.V. 1995. The role of endophytic bacteria duringseed piece decay and tomato tuberization. Plant Soil175:257–263.
Sturz, A.V., Christie, B.R., and Matheson, B.G. 1998. Asso-ciations of bacterial endophyte populations from redclover and potato crops with potential for beneficialallelopathy. Can. J. Microbiol. 44:162–167.
Tanprasert, P. and Reed, B.M. 1997. Detection and identifi-cation of bacterial contaminants from strawberry run-
ner explants. In Vitro Cellular Developmental Biol.33:221–226.
Tervet, I.W. and Hollis, J.P. 1948. Bacteria in the storageorgans of healthy plants. Phytopathology 38:960–967.
Thompson, P. L., Ramer, L. A., Guffey, A. P., and Schnoor,J. L. 1998. Decreased transpiration of poplar treesexposed to 2,4,6–trinitrotoluene. Environ. Toxicol.Chem. 17:902–906.
Tomasino, S.F., Leister, R.T., Dimock, M.B., Beach, R.M.,and Kelly, J.L. 1995. Field performance of Clavibacterxyli subsp. cynodontis expressing the insecticidal pro-tein gene cryIA. of Bacillus thuringiensis against Eu-ropean corn borer in field corn. Biol. Con. 5:442–448.
Trapp, S. 2000. Modeling uptake into roots and subsequenttranslocation of neutral and ionisable organic com-pounds. Pest Manage. Sci. 56:767–778.
Trapp, S., Mc Farlane, J. C., and Matthies, M. 1994. Modelfor uptake of xenobiotics into plants — validationwith bromacil experiments. Environ. Toxicol. Chem.13:413–422.
Triplett, E.W. 1996. Diazotrophic endophytes:progress andprospects for nitrogen fixation in monocots. Plant Soil186:29–38.
Tsiantos, J.T. and Stevens, W.A. 1986. The population dy-namics of Corynebacterium michiganense pv.Michiganensis and other selected bacteria in tomatoleaves. Phytopathol. Mediterr. 25:160–162.
Turner, J.T., Jeffrey, L.K., and Carlson, P.S. 1993. Endo-phytes:an alternative genome for crop improvement.In: International Crop Science. pp. 555–560. Buxton,D.R., Shibles, R., Forsberg, R.A., Blad, B.L., Asay,K.H., Paulsen, G.M., and Wilson, R.F., Eds., CropScience Society of America, Madison, WI.
Van Buren, A.M., Andre, C., and Ishimaru, C.A. 1993.Biological control of the bacterial ring rot pathogenby endophytic bacteria isolated from potato. Phytopa-thology 83:1406.
Vande Broek, A., Lambrecht, M., and Vanderleyden, J. 1998.Bacterial chemotactic mobility is important for theinitiation of wheat root colonization by Azospirillumbrasilense. Microbiology 144:2599–2606.
Van Loon, L.C. 1984. Regulation of pathogenesis and symp-tom expression in diseased plants by ethylene. In:Ethylene: Biochemical, Physiological and Applied As-pects. Fuchs, Y. and Chalutz, E., Eds., MartinusNijhoff/ W., Junk, the Hague, The Netherlands.
Van Peer, R., Punte, H.L.M., De Wega, L.A., and Schippers,B. 1990. Characterization of root surface andendorhizosphere Pseudomonads in relation to theircolonization of roots. Appl. Environ. Microbiol.56:2462–2470.
Wang, Y., Brown, H.N., Crowley, D.E., and Szaniszlo, P.J.1993. Evidence for direct utilization of a siderophore,ferrioxamine B. in axenically grown cucumber. PlantCell Environ. 16:579–585.
Webster, G., Gough, C., Vasse, J., Batchelor, C.A.,O’Callaghan, K.J., Kothar, S.L., Davey, M.R., De–

606
narie, J. and Cocking, E.E. 1997. Interactions of rhizo-bia with rice and wheat. Plant Soil 194:115–122.
Weisburg, W.G. 1991. 16S ribosomal DNA amplifica-tion for phylogenetic study. J. Bacteriol. 173:697–703
Whitesides, S.K. and Spotts, R.A. 1991. Frequency, distri-bution, and characteristics of endophytic Pseudomo-nas syringe in pear trees. Phytopathology 81:453–457.
Whiting, S. N., de Souza, M. P., and Terry, N. 2001. Rhizo-sphere bacteria mobilize Zn for hyperaccumulationby Thlaspi caerulescens. Environ. Sci. Technol.35:3144–3150.
Wiehe, W., Hecht-Buchholz, C., and Höflich, G. 1994. Elec-tron microscopic investigations on root colonizationof Lupinus albus and Pisum sativum with two associa-tive plant growth promoting rhizobacteria, Pseudomo-nas fluorescens and Rhizobium leguminosarum bv.trifolii. Symbiosis 17:15–31.
You, C.B., Song, W., Wang, H.X., Li, J.P., Lin, M., and Hai,W.L. 1991. Association of Alcaligenes faecalis withwetland rice. Plant Soil 137:81–85.
You, C.B., Li, X., Wang, H.X., Qiu, Y.S., Mo, X.Z., andZhang, Y.L. 1983. Associative nitrogen fixation ofAlcaligenes faecalis with rice plant. Biol N2 fixationNewsl. Sydney 11:92–103.

Recommended

















