
777
0022-1554/91/$3.30The Journal of Histochemistry and Cytochemistry
Copyright t� 1991 by The Histochemical Society. Inc.
Vol. 39, No. 6. pp. 777-785, 1991
Printed in USA.
Original Article
Evaluation of Human Monoclonal Antibody (2-139-1) inCutaneous Melanocytic Neoplasms in Fixed Tissue Sections’
WAIN L. WHITE,2 SAM T TAM, MALCOLM S. MITCHELL, MIMI C. YU,
and JUNE KAN-MITCHELL
Departments of Pathology and Dermatology, Wake Forest University, The Bowman Gray School of Medicine,
Winston-Salem, North Cam/ma (IVLW); Department ofDermatology, Dermatopathology Section, New York University Medical Center�
New York, New York (577); and Departments ofMedicine (MSM), Preventive Medicine (MCY), and Pathology (JK-M),
University ofSouthern California, School ofMedicine, Los Angeles, California.
Received for publication June 18, 1990 and in revised form February 1, 1991; accepted February 6, 1991 (0A2021).
Despite the growing list ofxenogeneic monodonal antibodies
(MAb) that recognize malignant melanoma-associated an-tigens (MAA) in formalin-fixed, paraffin-embedded tissue,none has been able to detect epitopes found in malignantmelanomas and not in melanocytic nevi. A human MAb,2-139-1, that showed promise in this regard was evaluatedagainst 85 melanocytic neoplasms, induding malignantmelanoma and histological simulators, particularly Spitz’snexus. MAb 2-139-1 stained 18 (53%) of34 melanomas, eight(57%) of 14 dysplastic nevi, six (38%) of 16 Spitz’s nevi,and three (14%) of2l banal nevi, which induded three small
congenital nevi. We observed a significant increasing trend
Introduction
The histological distinction between cutaneous malignant mela-
noma and some melanocytic nevi can be extremely difficult and
remains a recurrent fundamental problem in surgical pathology.
The diagnosis relies on imperfect morphological criteria which can-
not predict with certainty the biological behavior of melanocytic
neoplasms. For better understanding, the antigens expressed by
melanocytic neoplasms, benign (melanocytic nevi) and malignant
(melanoma), have been studied with mouse and human monoclo-
nal antibodies (MAb). A rapidly expanding list of melanoma-
associated antigens (MAA) and differentiation antigens has been
described and extensively reviewed (1-13). Although MAb, with
their intrinsic specificity, offer a more precise method of establish-
ing a diagnosis, most cannot distinguish malignant from benign
neoplasms. All of the MAb that are reported to show this capabil-
ity require the use of frozen-section material or tissue culture
in reactivity (% positive cells x intensity) associated with
the potential for malignancy (p for linear trend = 0.002)
We condude that human MAb 2-139-1 is applicable to thestudy of melanocytic neoplasms in routinely processed tis-sue. Although the ability of this MAb to separate benignfrom malignant cells is not absolute, our results suggest thatthe expression ofthe 2-139-1 epitope may be an early eventin melanocytic twnor progression. (JHistochem Cytochem
39:777-785, 1991)
KEY WORDS: Monoclonal antibodies; Malignant melanoma; Melano-
cytic nevus; Melanoma-associated antigens; Tumor progression;
Paraffin-embedded tissue.
(6,7,14-23). None of these MAb is readily applicable to routinely
processed specimens, specifically to formalin-fixed, paraffin-embedded tissues. Mouse MAb that are reactive with paraffin 5cc-
tions, detecting cytoplasmic antigens, cannot discriminate malig-
nant melanoma from melanocytic nevi (8,13,24-36).
A human MAb (2-139-1), produced by a human-mouse hetero-
hybridoma, was reported by some members ofour group to distin-
guish cutaneous malignant melanoma from benign melanocytic
nevi in formalin-fixed, paraffin-embedded sections (37,38). Be-
cause of the promise of these initial reports, we undertook an cx-
panded evaluation ofthis MAb with particular attention to Spitz’s
nevus, which is diagnostically the most difficult lesion to distin-
guish from malignant melanoma (39). Our results, their compari-
son to mouse MAb, and their implication in the issue of tumor
progression will be presented.
Materials and Methods
1 Supported by USPHS grants CA43220 and CA36233 from the NCI
(NIH), a grant from the Concern Foundation, and a gift from Mr. AlanGleitsman.
2 Correspondence to: Wan L. White, MD, Dept. of Pathology, Bow-man Gray School of Medicine, 300 5. Hawthorne Road, Winston-Salem,NC 27103.
Production of Human Monoclonal Antibody. Details of the produc-tion and characterization of the human MAb to MAAs have been previ-
ously reported (37). Six hybnidomas were obtained by the fusion of lym-
phocytes from the regional lymph nodes of patients with metastaticmalignant melanoma with a non-secretory variant of mouse myeloma cellline (M5), a horse serum-adapted subline ofSP2/OAG14. Six human anti-
by guest on February 4, 2015jhc.sagepub.comDownloaded from

778 WHITE, TAM, MITCHELL, YU, KAN-MITCHELL
bodies with reactivities against melanoma have been generated from threefusions and have been partially characterized (37,38). The production of
antibody in nude mouse ascites, purification, and conjugation with biotin
were performed as previously described (40).
Tissue Sections. Formalin-fixed, paraffin-embedded tissue blocks from
85 cases of melanocytic neoplasms were selected from the archives of theDermatopathology Laboratory at New York University Medical Center. These
included 34 malignant melanomas, 14 dysplastic nevi, 16 Spitz’s nevi, 18
banal compound intradermal nevi and blue nevi, and three congenital nevi.The histological criteria used for the diagnosis of each type of melanocyticneoplasm are those elaborated by Maise and Ackerman (39) and Barnhill
et al. (41). The tissues had been fixed in 10% buffered neutral formalinand embedded in paraffin. Tissue blocks were sectioned at a thickness of
5 lim and were picked up on acid-deaned, 0.1% poly-L-lysine(Mw 300,000)-coated slides. These slides were allowed to dry completely on a 37-40’Cslide warmer before placing into a 60C oven for 1 hr. One section was
stained with hematoxylin and cosin to confirm the diagnosis and the other
blanks were used for immunoperoxidase staining.
Avidin-Biotin-Immunoperoxidase Stain. A two-step direct immuno-
peroxidase procedure was performed on all sections (42). In brief,deparaffinized and hydrated slides were washed in PBS (0.01 M NaPO4,
0.15 M NaCI, pH 7.4) containing 0.3% Triton X-100 for 15 mm before the
endogenous peroxidase activity was quenched with 0.3% H202 in metha-
nol for 30 mm. All steps were performed at room temperature unless mdi-cared. Protease digestion occurred in 0.1% trypsin (Sigma, St. Louis, MO;
T8253) with 0.1% CaCl2 in 0.05 M Tnis buffer, pH 7.6, at 37’C for 20 mm.To reduce nonspecific binding of the primary antibodies, sections were in-cubated with 10% normal human serum diluted in PBS, pH 7.4, before
primary antibodies were applied and incubated at 4’C overnight (16-18
hn). Primary antibodies consisted of human biotinylated monoclonal (2-139-1) and a biotinylated normal human polyclonal IgG, which served as
negative control. Both primary antibodies were used at concentrations of25 and 50 �tg/ml in 1% normal human serum. The antibody preparations
were more than 85 % homogeneous, as determined by denaturing poly-
acrylamide gel electrophoresis. Their concentrations were estimated by theBio-Rad protein assay (Bio-Rad; Richmond, CA) before biotinylation aspreviously described (37). ABC reagents (Vector, Bunlingame, CA; Kit PK-
4000) were made up in 1% bovine serum albumin according to kit dilu-
tions and were incubated for 1 hr. Extensive washes were done between
each step with PBS or PBS containing 0.3% Triton X-100, both at pH 7.4.Colordevelopment took place in 0.04% 3-amino-9-ethylcarbazole(AEC)
(Sigma; A5754), with 0.015% H202 in sodium acetate buffer at pH 5.2
for approximately 7 mm. Slides were counterstained in Mayer’s hematoxy-lin (Sigma; MHS-32), covenslipped in Kaiser’s glyceninjelly, and sealed with
nail polish before viewing. A positive reaction consisted of a red granularprecipitate which was chosen to contrast with the brown melanin pigment
found in many melanocytic neoplasms.Although initial attempts to duplicate the earlier experiments with MAb
2-139-1 (38) resulted in a weak signal, staining was restored in many of our
cases with protease digestion, overnight incubation of primary antibody,
and an increase in the concentration ofMAb 2-139-1. These steps, however,
resulted in higher background staining. Blocking sera from rabbit, mouse,horse, human, swine, fetal calf, goat, sheep, and chicken were tested in
an attempt to reduce nonspecific binding of the antibody at these concen-trations. Human serum produced the best signal-to-noise ratio and was
used in this study.
Scoring OfReactivity. The specimens were scored on a scale ofo-4, where
0 denotes no staining; 1, weak staining; 2, definite but moderate staining;
3, strong staining; and 4, maximal staining. In each section, the numberof positive cells per total number of tumor cells was counted at x 400 and
a percentage of positivity was derived. This percentage was multiplied by
the staining intensity of the cells to calculate an immunohistochemical re-activity score for each neoplasm (43).
Statistical Analysis. Since the reactivity scores were cleanly not normally
distributed, we used a non-parametric method to test for possible differ-
ences in reactivity scores between the four groups of melanocytic lesions.
We first ranked all specimens in increasing order of their reactivity scores,
assigning the value 1 to the lowest rank, 2 to the second lowest rank, and
so on. Tied values were assigned the mean of the corresponding ranks. Theone-way analysis ofvaniance test was performed on the ranks; a test for Iin-ear trend in group means was also carried out (44,45). All values quoted
were two-sided.The linear trend analysis was intended to test the hypothesis that neac-
tivity with MAb 2-139-1 in these melanocytic neoplasms could be correlated
with malignancy or their different potential for malignant transformation,
from the low of the banal melanocytic nevus to the high of melanoma,
which can metastasize and kill. Dysplastic melanocytic nevus was rankedimmediately below melanoma because of its established role as an inter-mediate precursor in the evolution ofmalignant melanoma (46-49). Spitz’snevus was ranked below dysplastic nevus but above banal nevus because,despite its atypical histological features, it is a benign lesion, yet prelimi-
nary data suggest that it has some capacity for limited tumor progressionbut not for widespread metastasis and death (50). To avoid the conflicting
and confounding data on the potential for malignant transformation incongenital nevus (51,52), only very small (<1.0 cm in diameter) congenital
nevi were studied. These very small lesions have no well-established increasedrisk ofmelanoma as compared with other types ofmelanocytic nevi (51,52),
and were therefore included with the banal nevi.
Results
Immunostaining ofMal:gnant Melanoma
The cases of malignant melanoma consisted of 28 primary lesions
and six metastatic tumors (Table 1). Of the primary melanomas,
18 were common lesions involving the epidermis and dermis, three
were melanoma in situ only, three were with a pre-existing nevus,
two were spindle-cell melanomas, and two were undifferentiated
neoplasms (5-100 positive). One of the primary lesions showed fo-
cal regression.
MAb 2-139-1 reacted with 18 (54%)ofthe 34 melanomas. Twelve
of the melanomas stained intensely (Y/4�) over most of the neo-
plasm (50% or greater), with reactivity score indices of 1.5 to 4.0
(mean 2.3) (Figures la and ib). In eleven of these cases, 80% or
more of the lesion was reactive with the MAb. Three other cases
stained intensely positive but only focally (15% or less). The re-
maiming positive cases stained weakly (1�/2�) and varied from focal
staining (one case) to staining most of the neoplasm (two cases).
In one case, malignant melanoma in situ was present in associ-
ation with an intradermal melanocytic nevus. The antibody reacted
strongly against the malignant melanoma in situ but did not stain
the intradermal melanocytic nevus cells (Figures 2a, 2b, 3a, and
3b). In no case did MAb 2-139-1 stain any normal melanocytes or
Langerhans’ cells. The biotin-conjugated human IgG did not stain
any of the lesions; therefore, nonspecific binding of 2-139-1 and
of the ABC complex and nonspecific interactions caused by en-
dogenous biotin were excluded. Of particular interest was the as-
sociation of the epitope identified by the MAb with melanin in
many ofthe more heavily pigmented lesions. The antigen was pres-
ent not only in tumor cells in these lesions but also appeared within
by guest on February 4, 2015jhc.sagepub.comDownloaded from
HUMAN MONOCLONAL ANTIBODY IN MELANOCYTIC TUMORS 779
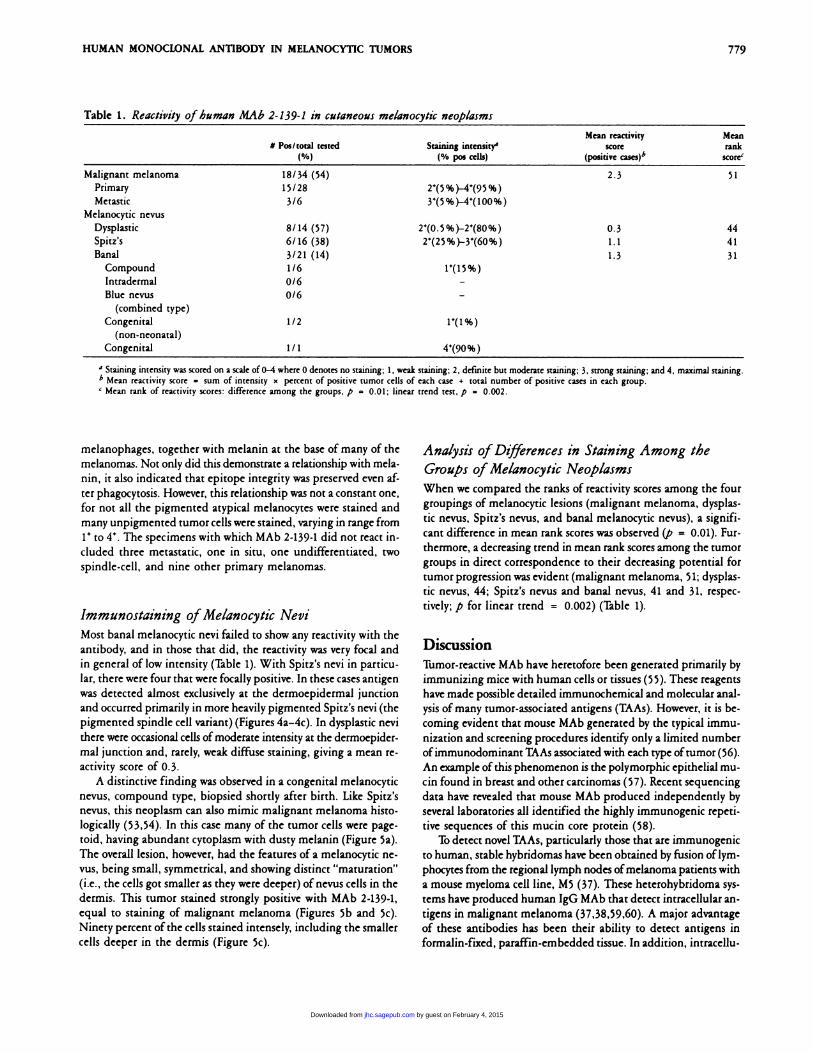
Table 1 . Reactivity of human MAb 2- 139- 1 in cutaneous melanocytic neoplasms
C Pos/total tested(%)
Staining intensity’
(% pos cells)
Mean reactivityscore
(positive cases)�’
Meanrank
score’
Malignant melanoma 18/34 (54) 2.3 51
Primary 15/28 2�(5%)-4(95%)
Metastic 3/6 Y(5%)-4(100%)Melanocytic nevus
Dysplastic 8/14 (57) 2’(0.5%)-2�(80%) 0.3 44Spitz’s 6/16 (38) 2’(25%)-Y(60%) 1.1 41
Banal 3/21 (14) 1.3 31Compound 1/6 P(15%)
Intradermal 0/6 -Blue nevus 0/6 -
(combined type)
Congenital 1/2 1’(1%)(non-neonatal)
Congenital 1 / 1 4(90%)
a Staining intensity was scored on a scale ofo-4 where 0 denotes no staining; 1, weak staining; 2, definite but moderate staining; 3. strong staining; and 4, maximal staining.
b Mean reactivity score = sum of intensity x percent of positive tumor cells of each case + total number of positive cases in each group.
C Mean rank of reactivity scores: difference among the groups, p = 0.01; linear trend test. p = 0.002.
melanophages, together with melanin at the base of many of the
melanomas. Not only did this demonstrate a relationship with mela-
fin, �t also indicated that epitope integrity was preserved even af-
ter phagocytosis. However, this relationship was not a constant one,
for not all the pigmented atypical melanocytes were stained and
many unpigmented tumor cells were stained, varying in range from
1� to 4� The specimens with which MAb 2-139-1 did not react in-
cluded three metastatic, one in situ, one undifferentiated, two
spindle-cell, and nine other primary melanomas.
Immunostaining ofMelanocytic Nevi
Most banal melanocytic nevi failed to show any reactivity with the
antibody, and in those that did, the reactivity was very focal and
in general of low intensity (Table 1). With Spitz’s nevi in particu-
lan, there were four that were focally positive. In these cases antigen
was detected almost exclusively at the dermoepidermal junction
and occurred primarily in more heavily pigmented Spitz’s nevi (the
pigmented spindle cell variant) (Figures 4a-4c). In dysplastic nevi
there were occasional cells of moderate intensity at the dermoepider-
mal junction and, rarely, weak diffuse staining, giving a mean re-
activity score of 0.3.
A distinctive finding was observed in a congenital melanocytic
nevus, compound type, biopsied shortly after birth. Like Spitz’s
nevus, this neoplasm can also mimic malignant melanoma histo-
logically (53,54). In this case many of the tumor cells were page-
toid, having abundant cytoplasm with dusty melanin (Figure 5a).
The overall lesion, however, had the features of a melanocytic ne-
vus, being small, symmetrical, and showing distinct “maturation”
(i.e. , the cells got smaller as they were deeper) of nevus cells in the
dermis. This tumor stained strongly positive with MAb 2-139-1,
equal to staining of malignant melanoma (Figures 5b and Sc).
Ninety percent of the cells stained intensely, including the smaller
cells deeper in the dermis (Figure Sc).
Analysis ofD:fferences in Staining Among the
Groups ofMelanocytic Neoplasms
When we compared the ranks of reactivity scores among the four
groupings of melanocytic lesions (malignant melanoma, dysplas-
tic nevus, Spitz’s nevus, and banal melanocytic nevus), a signifi-
cant difference in mean rank scores was observed (p = 0.01). Fur-
thermore, a decreasing trend in mean rank scores among the tumor
groups in direct correspondence to their decreasing potential for
tumor progression was evident (malignant melanoma, 5 1; dysplas-
tic nevus, 44; Spitz’s nevus and banal nevus, 41 and 3 1, respec-
tively; p for linear trend = 0.002) (Table 1).
DiscussionTumor-reactive MAb have heretofore been generated primarily by
immunizing mice with human cells or tissues (55). These reagents
have made possible detailed immunochemical and molecular anal-
ysis of many tumor-associated antigens (TkAs). However, it is be-
coming evident that mouse MAb generated by the typical immu-
nization and screening procedures identify only a limited number
ofimmunodominant TAAs associated with each type oftumor (56).
An example ofthis phenomenon is the polymorphic epithelial mu-
cm found in breast and other carcinomas (57). Recent sequencing
data have revealed that mouse MAb produced independently by
several laboratories all identified the highly immunogenic repeti-
tive sequences of this mucin core protein (58).
To detect novel TkAs, particularly those that are immunogenic
to human, stable hybnidomas have been obtained by fusion of lym-
phocytes from the regional lymph nodes ofmelanoma patients with
a mouse myeloma cell line, MS (37). These heterohybridoma sys-
tems have produced human IgG MAb that detect intracellular an-
tigens in malignant melanoma (37,38,59,60). A major advantage
of these antibodies has been their ability to detect antigens in
formalin-fixed, paraffin-embedded tissue. In addition, intracellu-
by guest on February 4, 2015jhc.sagepub.comDownloaded from
.,� �
‘1
.. �... � �.;: � �
; � ...‘-. . � . .., I-
� - ;,k.w�. ‘4,j�.. P�.
,..�w,. � .:,-
,. �
‘-: � � � ‘ .#{149}� � �‘� � . . . ., . . ,�1� �
. .. .. . ‘��.;:‘� � � � . � . �
� . ,�,: ��‘: � . #{149}�..
780 WHITE, TAM, MITCHELL, YU, KAN-MITCHELL
by guest on February 4, 2015jhc.sagepub.comDownloaded from

HUMAN MONOCLONAL ANTIBODY IN MELANOCYTIC TUMORS 781
lar antigens are more diffusely distributed and are much more read-
ily visualized in histologic sections than are exterior cell surface an-
tigens.
The present study confirms the earlier observation that, among
melanocytic lesions, reactivity of the human MAb 2-139-1 is most
selective for malignant melanoma (38). When we ranked the
melanocytic lesions according to their malignant potential, we found
a significant increasing trend in reactivity with MAb 2-139-1 (1�ble
1), from the least reactive banal melanocytic nexus to the most reac-
tive malignant melanoma. Although there were individual mela-
noma cells that did not stain and nevus cells that did, the differ-
ence in overall staining (the percent and intensity of cells stained)
among the four groups of lesions was statistically significant.
The benign Spitz’s nevus has more histological features in com-
mon with melanoma than differences, including scattered mela-
nocytes throughout all reaches of the epidermis, striking cytologi-
cal atypia, and mitotic figures (39). Six of 16 (38%) Spitz’s nevi
were reactive with MAb 2-139-1. The reactivity was, however, limited
to the dermoepidermal junction and, in most cases, the intensity
of staining was less than for melanoma (Figures 4b and 4c).
Of particular interest was another notorious simulator of mela-
noma, some congenital nevi biopsied shortly after birth or in child-
hood (53,54). Our lesion from a neonate showed intense staining
not only ofthe pagetoid melanocytes within the epidermis (Figure
Sb) but also of the small nevus cells in the dermis and around cc-
crine ducts and glands (Figure Sc). This finding suggests that the
antigen defined by 2-139-1 is analogous with antigens, such as ma-
jon histocompatibility class II antigens, that are expressed during
early melanocyte differentiation (5). Our previous results with a
variety of fetal tissues and other neoplasms indicate, however, that
the antigen detected by 2-139-1 is internal and is neither a differ-
entiation antigen of neural crest cells nor an oncofetal antigen
(37,59,60).
In dysplastic nevi, 59% stained to some degree with MAb 2-139-1,
but only a small proportion of the cells were reactive, and/or cells
showed weak intensity reflected in the low mean positive index of
0.3. In addition, the preliminary conclusion that MAb 2-139-1 was
not reactive in banal intradermal melanocytic nevi was confirmed
by this study (38).
Several factors can explain the failure of MAb 2-139-1 to stain
some melanomas. Phenotypic heterogeneity has been established
as a fundamental characteristic ofmalignant melanoma (3,61-64).
Yet in earlier series, the MAb 2-139-1 was reactive against almost
all melanomas tested (38), so that heterogeneity would be unlikely
to account for the lack of staining. Alternatively, the variability of
formalin fixation may be significant in explaining the negative reac-
tion with some ofthe melanomas tested. Prolonged exposure could
result in its destruction or masking by more stable cross-linkage
ofantigenic sites by formalin (65). This possibility is currently un-
der investigation.
Like the human IgG, MAb 2-139-1, several mouse MAb have
recently been reported to identify internal antigens in human mela-
noma in routine paraffin sections (8,24-36), and a composite of
the published results of these antibodies is presented in lible 2.
However, all seem to be different from MAb 2-139-1. NK1/C3 is
directed against a plasma membrane glycoprotein and often reacts
with intradermal nevus cells, all dysplastic nevi, and with histo-
cytes (25-29,35,36,66,67). Four of the mouse MAb (ME 491, ME
492, HMSA-1, HMSA-2)were raised against preparations of melano-
somes and the molecular weights of their target antigens have been
determined (l#{224}ble 2). HMSA-1 and -2 differ from MAb 2-139-1
by their uniform positive staining of intradermal nevus cells
(8,30,31). ME 491 and 492 react with many normal adult tissues
(including thyroid and lung), normal melanocytes, and nexus cells
at the dermoepidermal junction in common banal compound nevi
(33,34), which MAb 2-139-1 does not.
Another mouse MAb, HMB-45, reacts very similarly to MAb
2-139-1. Like the MAb 2-139-1 epitope, the HMB-45 antigen re-
mains incompletely characterized biochemically, but it is thought
to be a 7 KD glycopeptide in the cytoplasm and inner cell mem-
brane (24,68-70). Palozzo and Duray have demonstrated HMB-45
reactivity in agminated congenital Spitz nevi within nevus cells deep
within the dermis (71). This pattern parallels our finding with MAb
2-139-1 in the congenital nevus biopsied shortly after birth (Figure
Sc). Furthermore, just as our linear trend analysis indicates that
the 2-139-1 epitope may be related to tumor progression, the pres-
ence of the HMB-45 protein in the dermal component of some
dysplastic nevi likewise supports its association with tumor progres-
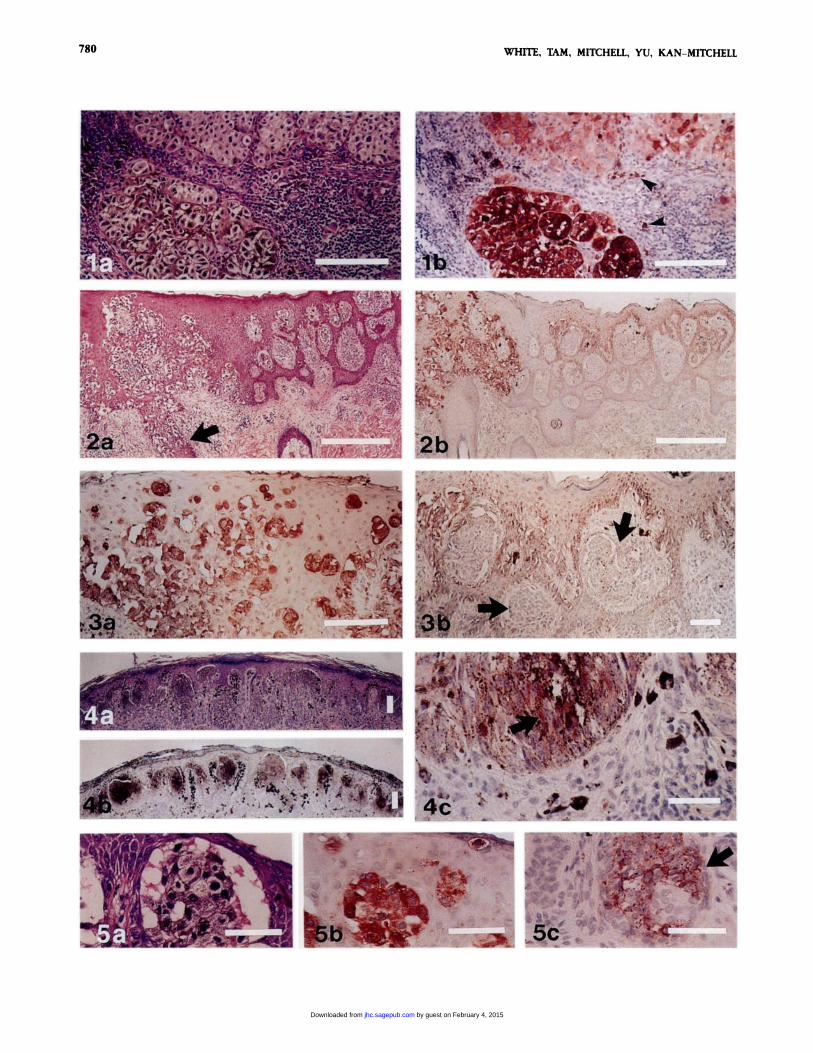
Figure 1 . (a) Large pagetoid melanocytes with dusty melanin are present at the base of a malignant melanoma. (b) Intense cytoplasmic staining of the atypicalmelanocytes is seen with MAb 2-139-1. In this field 71% of tumor cells are positive at 4, 29% at 1-3. If this field were representative of the entire neoplasm,the reactivity score would be approximately 1.7. Reaction product is also present in melanophages (arrowheads). Original magnification x 400. Bars - 100 �tm.
Figure 2. (a) Malignant melanoma in situ arising in association with an intradermal melanocytlc nevus has pagetoid cells in a pagetold pattern throughout alllevels ofthe epidermis and involves follicular epithelium (arrow). H & E. (b) MAb 2-139-1 strongly decorates the melanoma in situ. Original magnification x 100.Bars = 500 �m.
Figure a (a) Detail ofthe malignant melanoma in situ (in Figure 2)showsthe intraepidermal melanocytes positively stained with MAb 2-139-1. (b) Onlythe atypicalmelanocytes at the dermoepidermal junction are stained with MAb 2-139-1. The nests of nevus cells (arrows) are completely negatIve. The pigment In the nevuscells is melanin, not reaction product. Original magnifications: a x 400; b x 200. Bars - 100 �sm.
Figure 4. (a) Spitz’s nevus, pigmented spindle-cell variant shows typical histologicalfeatures of symmetry, mostly nested at thejunction, with epidermal hyperpla-sia, hypergranulosis, and hyperkeratosis. H & E. (b) Despite heavy pigmentation, a blush ofposltive reactivity is seen with MAb 2-139-1. (c)A detail shows posItivestaining junctional nevus cells (arrow) in addition to heavy pigmentation. Original magnifications: a, b x 40; C x 400. Bars - 100 �tm.
Figure 5. (a) A congenital nevus biopsied shortly after birth has large pagetoid cells in the epidermis that stimulate melanoma In situ. H & E. (b) MAb 2-139-1
decorates the large melanocytes in the epidermis with the same intensity as malignant melanoma. (C) The small nevus cells in the dermis and around adnexalstructures (arrow) stain intensely with MAb 2-139-1 . Original magnification x 400. Bars - 100 �sm.
by guest on February 4, 2015jhc.sagepub.comDownloaded from
782 WHITE, TAM, MITCHELL, YU, KAN-MITCHELL
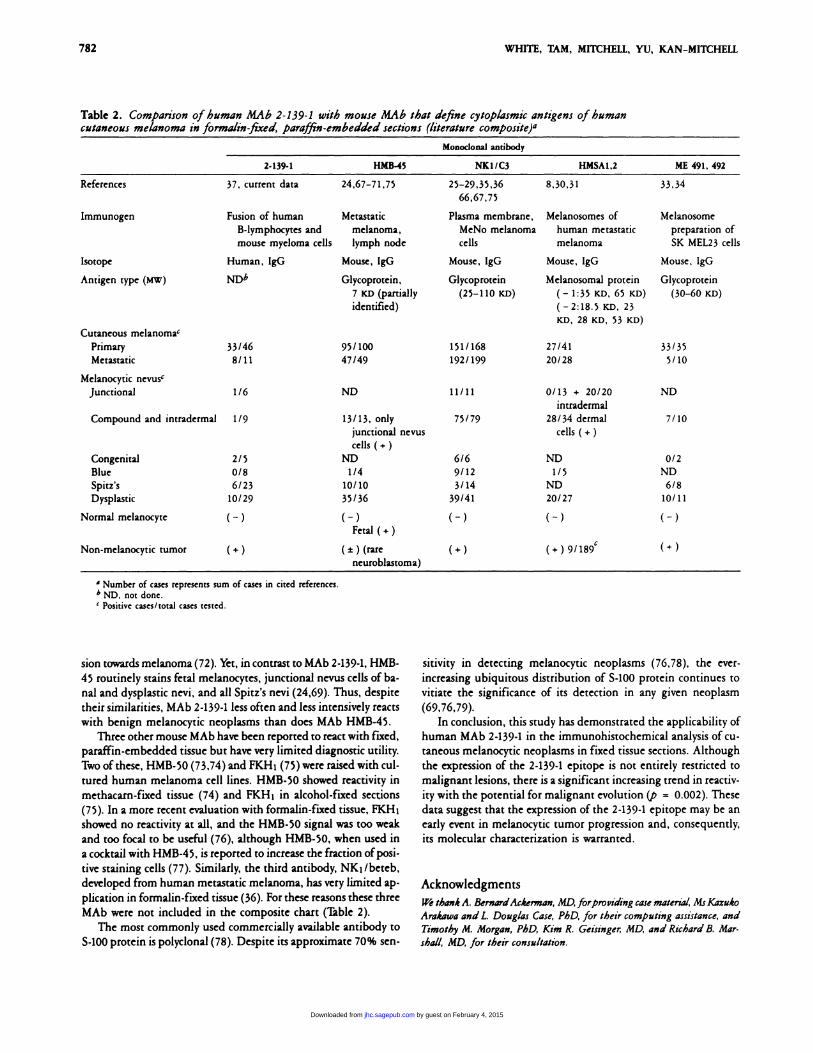
Table 2. Comparison ofhuman MAb 2-139-1 with mouse MAb that define cytoplasmic antigens of humancutaneous melanoma in formalin-fixed, paraffin-embedded sections (literature composite)a
Monoclonal antibody
2-139-1 H.MB-45 NK1/C3 HMSA1,2 ME 491, 492
References 37, current data 24,67-71,75 25-29,35,36
66,67,75
8,30,31 33,34
Immunogen Fusion of humanB-lymphocytes andmouse mycloma cells
Metastaticmelanoma,lymph node
Plasma membrane,MeNo melanomacells
Melanosomes ofhuman metastaticmelanoma
Melanosomepreparation of5K MEL23 cells
Isotope Human, IgG Mouse, IgO Mouse, IgG Mouse, IgG Mouse, lgG
Antigen type (MW) Glycoprotein,
7 KD (partiallyidentified)
Glycoprotein
(25-110 lcD)
Melanosomal protein
( - 1:35 KD, 65 lcD)
(-2:18.5 KD, 23KD, 28 KD, 53 KD)
Glycoprotein
(30-60 KD)
Cutaneous melanoma’Primary 33/46 95/100 151/168 27/41 33/35
Metastatic 8/11 47/49 192/199 20/28 5/10
Melanocytic nevusc
Junctional 1/6 ND 11/11 0/13 + 20/20 ND
intradermal
Compound and intradermal 1/9 13/13, onlyjunctional nevus
cells ( +)
75/79 28/34 dermal
cells ( +)7/10
Congenital 2/5 ND 6/6 ND 0/2
Blue 0/8 1/4 9/12 1/5 ND
Spitz’s 6/23 10/10 3/14 ND 6/8
Dysplastic 10/29 35/36 39/41 20/27 10/11
Normal melanocyte ( - ) ( - )Fetal ( +)
( - ) ( - ) ( -)
Non-melanocytic tumor (+) (±) (rareneuroblastoma)
(+) (+) 9/189C (+)
a Number of cases represents sum of cases in cited references.b ND, not done.C Positivecases/totalcases tested.
sion towards melanoma (72). Yet, in contrast to MAb 2-139-1, HMB-
45 routinely stains fetal melanocytes, junctional nevus cells of ba-
nal and dysplastic nevi, and all Spitz’s nevi (24,69). Thus, despite
their similarities, MAb 2-139-1 less often and less intensively reacts
with benign melanocytic neoplasms than does MAb HMB-45.
Three other mouse MAb have been reported to react with fixed,
paraffin-embedded tissue but have very limited diagnostic utility.
Two ofthese, HMB-50 (73,74) and FKHi (75)were raised with cul-
tured human melanoma cell lines. HMB-50 showed reactivity in
methacarn-fixed tissue (74) and FKH1 in alcohol-fixed sections
(75). In a more recent evaluation with formalin-fixed tissue, FKHi
showed no reactivity at all, and the HMB-S0 signal was too weak
and too focal to be useful (76), although HMB-50, when used in
a cocktail with HMB-45, is reported to increase the fraction of posi-
tive staining cells (77). Similarly, the third antibody, NK�/beteb,
developed from human metastatic melanoma, has very limited ap-
plication in formalin-fixed tissue (36). For these reasons these three
MAb were not included in the composite chart (Thble 2).The most commonly used commercially available antibody to
5-100 protein is polyclonal (78). Despite its approximate 70% sen-
sitivity in detecting melanocytic neoplasms (76,78), the ever-
increasing ubiquitous distribution of 5-100 protein continues to
vitiate the significance of its detection in any given neoplasm
(69,76,79).
In conclusion, this study has demonstrated the applicability of
human MAb 2-139-1 in the immunohistochemical analysis of cu-
taneous melanocytic neoplasms in fixed tissue sections. Although
the expression of the 2-139-1 epitope is not entirely restricted to
malignant lesions, there is a significant increasing trend in rcactiv-
ity with the potential for malignant evolution (p = 0.002). These
data suggest that the expression ofthe 2-139-1 epitope may be an
early event in melanocytic tumor progression and, consequently,
its molecular characterization is warranted.
Acknowledgments
w� thankA. BernardAckerman, MD,forproviding case material, MsKazuko
Arakawa and L Douglas Case, PhD, for their computing assistance, andTimothy M. Morgan, PhD, Kim R. Geisinger, MD, and Richard B. Mar-
shall, MD, for their consultation.
by guest on February 4, 2015jhc.sagepub.comDownloaded from

HUMAN MONOCLONAL ANTIBODY IN MELANOCYTIC TUMORS 783
Literature Cited1. Koprowski H, Steplewski Z, Herlyn D, Herlyn M. Studies of antibod-
ies against human melanoma produced by somatic cell hybrids. Proc
NatI Acad Sci USA 1978;75:3405
2. ThompsonJJ, Henlyn MF, Elder DE, Clark WH, Steplewski Z, Kopnowski
H. Use ofmonoclonal antibodies in detection of melanoma-associatedantigens in cultured rapidly dividing human melanocytes. AmJ Pathol
1982;107:357
3. Houghton AN, Cordon-Cardo C, Eisinger M. Differentiation antigens
of melanomas and melanocytes. Int Rev Exp Pathol 1986;28:317
4. Reisfeld RA, Cheresh DA. Human tumor antigens. Adv Immunol
1987;40:323
5. Herlyn M, Koprowski H. Melanoma antigens: immunological and bio-logical characterization and clinical significance. Annu Rev Immunol1988;6:283
6. Rodeck U, Herlyn M. Characteristics of cultured human melanocytesfrom different stages of tumor progression. In Nathanson L, ed. Malig-
nant melanoma: biology, diagnosis and therapy. Boston: Kiuwer Aca-
demic Publishers, 1988:3
7. Ravindranath MH, Inc BY. Gangliosides as antigens of human mela-
noma. In Nathanson L, ed. Malignant melanoma: biology, diagnosis,
and therapy. Boston: Kluwer Academic Publishers, 1988:17
8. Maeda K, Macda K,Jimbow K. Specification and use ofa mouse mono-clonal antibody raised against melanosomes for the histopathologic di-
agnosis of amelanotic malignant melanoma. Cancer 1988;62:926
9. Thomson DM, Real FX, Munakami 5, Cordon-Cardo C, Old U, Hough-
ton AN. Differentiation antigens ofmelanocytes and melanoma: analysis
ofmelanosome and cell surface markers ofhuman pigmented cells with
monoclonal antibodies. 3 Invest Dermatol 1988;90:459
10. Ganin-Chesa P, Beresford HR, Carrato-Mena A, Oettgen HF, Old U,
Melamed MR, Rettig WJ. Cell surface molecules of human melanoma:immunohistochemical analysis of the gp57, GD3, and meL-CSPG an-
tigenic systems. Am J Pathol 1989;134:295
1 1. Cordon-Cardo C. Biochemical and immunologic diagnosis of cancer.Melanoma Tumor Biol 1987;8:151
12. Nance DV, Siegal CP. The use of monoclonal antibodies in the search
for tumor-specific antigens. In Wick MR, Siegal GR. eds. Monoclonalantibodies in diagnostic immunohistochcmistry. New York: Marcel Dek-ken, 1988:593
13. Natali PG, Fawwaz R, Ruiter DJ, Bigotti A, Kageshita T, Temponi
Fenrone S. Immunodiagnostic and immunotherapeutic applications ofantihuman melanoma associated antigen monoclonal antibodies. In
Conti CJ, Slaga TJ, Klein-Szanto AJP, eds. Skin tumors: experimental
and clinical aspects. Carcinogenesis - a comprehensive survey. Vol 11.New York: Raven Press, 1989:133
14. Ruiter D, Gemma D, Steijlen P. Antibodies selected to discriminate
between malignant melanomas and nevocellular nevi. 3 Invest Denmatol
1985;85:4
15. Holzmann B,JohnsonJ, Kaudewitz P, Riethm#{252}ller G. In situ analysis
of antigens on malignant and benign cells of the melanocyte lineage:
differential expression of two surface molecules, gp. 75 and p. 89. JExp Med 1985;161:366
16. Real FX, Houghton AN, Albino AP, Cordon-Cardo C, Melamed MR.
Oettgen HF, Old U. Surface antigens of melanomas and melanocytesdefined by mouse monoclonal antibodies. Specificity analysis and corn-
panison of antigen expression in cultured cells and tissues. Cancer Res
1985;45:4401
17. LehmannJM, Holzmann B, Eckhard WB, Schmiegelow P, Riethm#{252}llerG,JohnsonJP. Discrimination between benign and malignant cells ofmelanocyte lineage by two novel antigens, a glycoprotein with a mo-
lecular weight of 113,000 and a protein with a molecular weight of
76,000. Cancer Res 1987;47:841
18. Brocken EB, Suter U, Bruggen J, Ruitten D). Phenotypic dynamics oftumor progression in human malignant melanoma. mt J Cancer1985;39:29
19. Holzmann B, Brocker EB, Uehmann)M, Ruiter DJ, Song C, Riethm#{252}llerG, Johnson JP. Tumor progression in human malignant melanoma:five stages defined by their antigenic phenotypes. mt J Cancer1987;39:466
20. Suter U, BrUggenJ, Br#{246}ckenEB. Song C. Tumor-associated antigen cx-
pressed in melanoma cells with lower malignant potential. Eur Int JCancer 1985;35:787
21. JohnsonJP, Strade BG, Holzmann B, &hwable W, RiethmUller G.novo expression of cell adhesion molecule 1 in melanoma correlates
with increased risk ofmetastasis. Proc NatI Acad Sci USA 1989;86:641
22. Henlyn M, Henlyn D, Elder DE, Bondi E, LaRossa D, Hamilton R, SearsHF, Balaban G, Guenry IV D, Clark WH, Koprowski H. Phenotypiccharacteristics of cells derived from precursors of human melanoma.
Cancer Res 1983;43:5502
23. Stade BG, Holzmann B, RiethmOller G, Johnson JP. cDNA cloning
of the progression associated melanoma antigen p3.58: identity toICAM-1, a leukocyte intracellular adhesion molecule. In Metzgar RS,Mitchell MS, eds. Human tumor antigens and specific tumor therapy.
UCLA symposia on molecular and cellular biology, new series. Vol 99.
New York: Alan Liss, 1988:73
24. Gown AM, Vogel AM, Hoak D, Gough F, McNutt MA. Monoclonal
antibodies specific for melanocytic tumors distinguish subpopulationsof melanocytes. Am J Pathol 1986;123:195
25. Vennegoor C, CalafatJ, Hageman PH, Van Buitenen F,Janssen H, Kolk
A, R#{252}mkePh. Biochemical characterization and cellular localizationofa formalin-resistant melanoma-associated antigen reacting with mono-clonal antibody NK1/C-3. Int J Cancer 1985;35:287
26. Mackie RM, Campbell I, Turbitt ML. Use ofNKIC3 monoclonal anti-
body in the assessment of benign and malignant melanocytic lesions.
J Clin Pathol 1984;37:367
27. Palmer AA, Hall BE, Lew A. A comparison ofsome methods for iden-tifying amelanotic and oligomelanotic melanoma metastases in paraf-
fin sections. Pathology 1985;17:335
28. Garter KC, Ralkiaer E, Skinner), Brown D, Heryct A, Pulford KAF,
Hou-)ensen K, Mason DY. An immunocytochemical study of malig-
nant melanoma and its differential diagnosis from other malignanttumors. J Clin Pathol 1985;38:1353
29. Hagen EC, Vennegoor C, Schlingemann RO, Van Den Velde EA, Ruiter
DJ. Correlation of histopathological characteristics with staining pat-terns in human melanoma assessed by monoclonal antibodies reactiveon paraffin sections. Histopathology 1986;1O:689
30. Akutsu Y, Jimbow K. Development and characterization of a mousemonoclonal antibody, MoAb HMSA-1, against melanosomal fractionof human malignant melanoma. Cancer Res 1986;46:2904
31. Macda K,Ji.mbOW K. Development ofMoAb HMSA-2 for melanosomesof human melanoma and its application to immunohistopathologicdiagnosis of neoplastic melanocytes. Cancer 1987;59:415
32. Macda K, Macda K, )imbow K. Positive reactivity of dysplastic mela-nocytes with a monoclonal antibody against malignant melanosomes,MoAb HMSA-2. J Invest Dermatol 1988;91:247
33. Atkinson B, Ernst CS, Ghnist BFD, Herlyn M, Biaszczyk M, Ross AH,
Herlyn D, Steplewski Z, Kopnowski H. Identification of melanoma-
associated antigens using fixed tissue screening of antibodies. CancerRes 1984;44:2577
34. Atkinson B, Ernst CS, Ghnist BFD, Ross AH, Clark WH, Herlyn M,
Herlyn D, Maul G, Steplewski Z, Koprowski H. Monoclonal antibodyto a highly glycosylated protein reacts in fixed tissue with melanomaand other tumors. Hybnidoma 1985;4:243
35. van Duinen SG, Ruiter DJ, Hageman P, Vennegoor C, Dickersin GR,
by guest on February 4, 2015jhc.sagepub.comDownloaded from

784 WHITE, TAM, MITCHELL, YU, KAN-MITCHELL
57. Gendler 5, BurchellJM, Duhig T, Lamport D, White R, Parker M, Tay-
Scheffer E, RUmke P. Immunohistochemical and histochemical toolsin the diagnosis of amelanotic melanoma. Cancer 1984;53:1566
36. Vennegoor C, Hageman P, Van Nouhuijs H, Ruiter DJ, Calafat), Rin-
gens PJ, R#{252}mkeP. A monoclonal antibody specific for cells of themelanocyte lineage. Am ) Pathol 1988;130:179
37. Kan Mitchell), Imam A, KempfR, 1�ylor C, Mitchell M. Human mono-clonal antibodies directed against melanoma tumor-associated anti-gens. Cancer Res 1986;46:2490
38. Imam A, Mitchell MS, Modlin RI, Taylor CR, KempfRA, Kan-Mitch-
ellJ. Human monoclonal antibodies that distinguish cutaneous malig-nant melanomas from benign nevi in fixed tissue sections. ) Invest Den-matol 1986;86:145
39. Maize)C, Ackerman AB. Pigmented lesions ofthe skin. Philadelphia:
Lea & Febiger, 1987
40. Imam A, Drushella M, Taylor C, T#{246}k#{233}sZ. Generation and immuno-
histological characterization ofhuman monoclonal antibodies to mam-
many carcinoma cells. Cancer Res 1985;45:263
41. Bannhill RU, Roush GC, Duray PH. Correlation of histologic architec-
tural and cytoplasmic features with nuclear atypia in atypical (dysplas-
tic) nevomelanocytic nevi. Hum Pathol 1990:21:51
42. Sheibani K, Tubbs RR. Enzyme immunohistochemistry: technical
aspects. Semin Diag Pathol 1984;1:235
43. McCarty KS)r, Miller IS. Cox EB, Konrath), McCarty KS Sr. Estrogen
receptor analysis. Correlation ofbiochemical and immunohistochemi-cal methods using monoclonal antireceptor antibodies. Arch PatholLab Med 1985;109:716
44. Afifi AA, Azen SP. Statistical analysis, a computer oriented approach.2nd Ed. New York: Academic Press, 1979
45. SAS User’s Guide: Statistics, version 5 edition. Cary, NC: SAS Insti-
tute Inc. 1985:651
46. Tong AKF, Murphy GF, Mihm MC. Dysplastic nevus: a formalhistogenetic precursor of malignant melanoma. In Mihm MC, Mur-
phy GF, Kaufman N, eds. Pathobiology and recognition of malignant
melanoma. Baltimore: Williams & Wilkins, 1988:10
47. Elder DE, Clark WH. Malignant melanoma. In Thiens BH, DobsonRL, eds. Pathogenesis ofskin disease. New York: Churchill Uivingstone,1986:445
48. Clank WHJr, Elder DE, Guerry D IV, Epstein MN, Greene MH, Van
Horn M. A study of tumor progression: the precursor lesions of super-ficial spreading and nodular melanoma. Hum Pathol 1984;15:1147
49. Albert US. Rhodes AR, Sober A). Dysplastic melanocytic nevi and cu-taneous melanoma: markers of increased melanoma risk for affectedpersons and blood relatives. ) Am Acad Dermatol 1990;22:69
50. Smith K), Seab)A, Lupton GP, Gnaham)H. Spindle cell and epithelioid
cell nevi with atypia and metastasis. J Cutan Pathol 1988;15:342
51. Elder DE. The blind men and the elephant: different views of small
congenital nevi. Arch Dermatol 1985;121:1263
52. KopfAW. Response [management ofcongenital nevocytic nevi]. ) Am
Acad Dermatol 1984;11:633
53. Cullity G. Intra-epithelial changes in childhood nevi simulating malig-
nant melanoma. Pathology 1984;16:307
54. Silvers D, Helwig E. Melanocytic nevi in neonates. ) Am Acad Den-
matol 1981;4:166
55. Resifeld BA. Immunotherapy ofcancer: a perspective view. ) Clin Lab
Anal 1988;2:124
56. Kan-Mitchell). Human monoclonal antibodies. In Metzgar RS, Mitchell
MS, eds. Human tumor antigens and specific tumor therapy. UCLAsymposia on molecular and cellular biology. New Series. Vol 99. NewYork: Alan Liss, 1988:105
lor-Papdimitniou J. Cloning of partial cDNA encoding differentia-tion and tumor-associated mucin glycopnoteins expressed by human
mammary epithelium. Proc NatI Acad Sci USA 1988;84:6060
58. Gendler 5, 1�ylor-PapadimitniouJ, Burchell), Duhig T A polymorphicepithelial mucin expressed by breast and other carcinomas: immuno-logical and molecular studies. In Metzgar 83, Mitchell MS, eds. Hu-man tumor antigens and specific tumor therapy. UCLA symposia onmolecular and cellular biology. New Series. Vol 99. New York: AlanLiss, 1988:11
59. Kan-Mitchell), White WL, Mitchell MS. Tumor-reactive human IgGmonoclonal antibody from a melanoma patient. Cancer Res 1989;49:4536
60. Kan-Mitchell), Mitchell MS. Human monoclonal antibodies and theirusefulness in diagnosis. In Kupchick HZ, ed. In vitro diagnosis of hu-
man tumors using monoclonal antibodies. Immunology series. New
York: Marcell Dekker, 1988;39:289
61. Houghton AN, Real FX, Davis U), Cordon-Cardo C, Old L). Pheno-typic heterogeneity of melanoma: relationship to the differentiation
program of melanoma cells. ) Exp Med 1987;165:812
62. Hellstr#{246}mI, Garnigues H), Cabasco U, Mosely GH, Brown JP, Hell-strom KE. Studies of a high molecular weight human melanoma-associated antigen. ) Immunol 1983;130:1467
63. Natali PG, Cavaliere R, Bigotti A, Nicotna MR. Russo C, Ng AK, Gia-comini P, Ferrone S. Antigenic heterogeneity ofsurgically removed pni-
mary and autologous metastatic human melanoma cells. ) Immunol
1983;130:1462
64. Albino AP, Lloyd KO, Houghton AN, Oettgen HF, Old U. Hetenoge-neity in surface antigen expression and glycoprotein expression of celllines derived from different metastases of the same patient: implica-
tions for the study of tumor antigens. ) Exp Med 1981;154:1764
65. Visscher D, Cooper PH, Zarbo R), Cnissman )D. Cutaneous neuroen-docnine (Merkel cell) carcinoma: an immunophenotypic, clinicopatho-
logic, and flow cytometnic study. Mod Pathol 1989;2:331
66. Paul E, Cochran A), Wen D-R. Immunohistochemical demonstrationofS-100 protein and melanoma-associated antigens in melanocytic nevi.
) Cutan Pathol 1988;15:161
67. Hageman PC, Vennegoon C, Van der Valk MA, Van der Esch EP, Vanden Tweel)G, Ruitter D), Van Duinen 5G. R#{252}mkeP. Reactivity of mono-clonal antibody NK1 /C3 directed against a melanoma-associated an-
tigen with formalin-fixed paraffin-embedded normal and tumor tis-
sues. Eur) Cancer Clin Oncol 1988;24(suppl 2):S33
68. Vennegoor C, Hageman PH, Van Nouhuijs H, Ruiter DJ, Calafat J,Ringens P), R#{252}mkePH. A monoclonal antibody specific for cells ofthe melanocyte lineage. Am J Pathol 1988;130:179
69. Corwin D), Gown AM. Review ofselected lineage-directed antibodiesuseful in routinely processed tissues. Arch Pathol Lab Med 1989;113:645
70. Wick MR. Swanson PE, Rocamona A. Recognition of malignant mela-
noma by monoclonal antibody HMB-45: an immunohistochemical studyof200 paraffin-embedded cutaneous tissues.J Cutan Pathol 1988;15:201
71. Palazzo)P, Duray PH. Congenital agminated Spitz nevi: immunoreac-
tivity with a melanoma-associated monoclonal antibody. J Cutan Pathol1988;15:166
72. Smoller BR, McNutt NS, Hsu A. HMB-45 staining of dysplastic nevi:support for a spectrum of progression toward melanoma. Am J Surg
Pathol 1989;13:680
73. Esclamado RM, Gown AM, Vogel AM. Unique proteins defined bymonoclonal antibodies specific for human melanoma. Am ) Surg
1986;152:376
74. Vogel AM, Esclamado RM. Identification ofa secreted Mr 95,000 gly-
coprotein in human melanocytes and melanomas by a melanocytic spe-
cific monoclonal antibody. Cancer Res 1988;48:1286
75. Fukaya T, Hashimoto K, Eto H, Mehregan A. Mouse monoclonal anti-
by guest on February 4, 2015jhc.sagepub.comDownloaded from

HUMAN MONOCLONAL ANTIBODY IN MELANOCYTIC TUMORS 785
body (FKH1) detecting human melanoma-associated antigens. Can-
cer Res 1986;46:5195
76. Thomson W, Mackie RM. Comparison offive anti-melanoma antibodies
for identification ofmelanocytic cells in tissue sections in routine den-
matopathology. ) Am Acad Dermatol 1989;21:1280
77. Zanbo RJ, Visscher DW, Bacchi CE, Gown AM. Combination of mono-
clonal antibodies HMB-45 and HMB-50 in immunocytochemical con-firmation of the diagnosis of malignant melanoma. Lab Invest
1990;62:11A
78. Cochran A), Wen D-R, Henschman HR, Gaynor RB. Detection of S-100
protein as an aid to the identification ofmelanocytic tumors. Int) Cancer
1982;30:295
79. AbdelatifOMA, Khankhanian NK, Crosby)H, Chamberlain R, Seig-
len MM, Tom GD. Malignant fibrous histiocytoma and malignant mela-noma: the role of immunohisto-chemistry and electron microscopy in
the differential diagnosis. Mod Pathol 1989;2:477
by guest on February 4, 2015jhc.sagepub.comDownloaded from
Recommended

![[Incidence of haematological neoplasms in Castilla y Leon.]](https://img.pdfslide.net/doc/110x75/634c4a087e26824f4400ef99/incidence-of-haematological-neoplasms-in-castilla-y-leon.jpg)















