
Hum Genet (1985) 71 : 62-74
© Springer-Verlag 1985
Genetic linkage relationships of seven DNA probes with Duchenne and Becker muscular dystrophy
C, S. B r o w n I , N . S. T . T h o m a s 1 , M . Sarfarazi 1 , K. E . D a v i e s 2 , L . K u n k e l 3 , P . L . P e a r s o n 4, H . M . K i n g s t o n 1 , D . J . S h a w 1 , a n d P . S. H a r p e r I
1Section of Medical Genetics, University of Wales College of Medicine, Heath Park, Cardiff, UK 2Nuffield Department of Clinical Medicine, John Radcliffe Hospital, Headington, Oxford, England 3Genetics Division, Children's Hospital Medical Centre, Boston, USA 4Department of Human Genetics, Sylvius Laboratoria des Rijksuniversiteit Leiden, The Netherlands
S u m m a r y . The inheritance of seven restriction fragment length polymorphisms detected by DNA probes has been studied in families with Duchenne and Becker muscular dys- trophies (DMD and BMD). The probes used have all been mapped to the short arm of the X-chromosome, four being distal and three proximal to the disease loci located within the Xp21 region. Linkage analysis of the DNA polymorphisms in relation to the two disorders showed similar genetic distances. Data obtained from DMD and BMD families have been com- bined to give more precise values for the different recombina- tion fractions. Combined use of these polymorphic DNA markers will be of practical value in the genetic counselling of women at risk for Duchenne and Becker muscular dystrophy.
I n t r o d u c t i o n
Duchenne muscular dystrophy (DMD) and Becker muscular dystrophy (BMD) are X-linked recessive disorders with DMD being the most common and the most severe, with a frequency of about 1 in 5000 newborn males. BMD, first described by Becker and Kiener in 1955, is clinically similar in the pattern of muscle involvement, but follows a more benign course and has a lower frequency of around 1 in 30,000 liveborn males.
Since the biochemical defects that cause these X-linked muscular dystrophies are unknown, there has been no direct means of identifying the responsible gene or genes. It has, however, become possible to approach this goal through gene mapping. Localisation of DMD to the p21 region of the X chromosome has been achieved by observations of girls with the disease who have de novo balanced X;autosome transloca- tions in which the p21 region of the X chromosome is consist- ently involved, although the autosomal region is variable (Zatz et al. 1981). Independent evidence has come from the use of restriction fragment length polymorphisms (RFLPs) as genetic markers which can be typed in DMD and BMD kin- dreds, to give genetic linkage data between the marker and disease loci.
The first polymorphic markers shown to be linked to DMD were the DNA probes RC8 (Murray et al. 1982) and L1.28
Offprint requests to: N.S.T. Thomas, Section of Medical Genetics, University of Wales College of Medicine, Heath Park, Cardiff, UK
(Davies et al. 1983), these being situated on either side of the disease locus. A study of these markers in BMD families (Kingston et al. 1983, 1984) also showed genetic linkage to these probes, suggesting that BMD and DMD may be closely linked or allelic. In this study we have investigated genetic linkage between DMD, BMD, and a further five DNA polymorphisms, two proximal to the disease loci and three dis- tal; the earlier probe data have been updated and included. A total of 59 kindreds have been analysed, 33 with DMD and 26 with BMD, and the total data using seven DNA probes located on the short arm of the X chromosome are presented.
Mater ia l s a n d m e t h o d s
Origin of probes
RC8 (DXS9) was selected as a lambda clone containing a single copy DNA sequence from a flow-sorted X chromosome specific library (Davies et al. 1981). It detects a polymorphism when genomic DNA is cut with the restriction enzyme Taql and its physical location is Xp21-Xp223. The rare allele fre- quency in the normal British population is 0.11.
L1.28 (DXS7) was isolated as a single copy sequence from a library of random human genomic DNA cloned into plasmid pBR322 (Pearson et al. unpublished work). It also detects a polymorphism with Taql and its physical location is Xp21- Xp110. It has a rare allele frequency of 0.33 in the British population.
782 and 754 were selected from a flow-sorted X-library (Kunkel et al. 1982; HoNer et al. 1985) as single-copy sequences and cloned into plasmid pAT153. 782 detects a polymorphism with EcoR1 and maps to the same region as RC8. Its rare allele frequency is 0.50 in our sample. 754 detects a polymorphism with Pstl and has been localised to Xp21-Xp113. It has a rare allele frequency of 0.39.
D2 and 99.6 were also isolated from the above X-library and these unique sequences were cloned into plasmid pBR322. D2 detects a polymorphism with PvuII and has been localised to Xp21-Xp223, as has 99.6. The rare allele fre- quency for D2 is 0.32. 99.6 shows a polymorphism with Pstl and its rare allele has a frequency of 0.37.
OTC is a single-copy cDNA sequence obtained by screen- ing a human cDNA library with a synthetic oligonucleotide

63
Fig.la-L Alleles detected by the seven polymorphic probes when DNA is cut with the appropriate restriction enzyme. DNA transfer onto Zetaprobe (Biorad) was carried out overnight in 10 x SSC. Prehybridisation was at 65°C for 4 -5h in 10ml of 5 x SSC; 10 x Dcnhardts; 0.5mg/ml denatured herring sperm DNA; 5% dextran sulphate and 50mM potassium phosphate (pH 6.5). Hybridisation was overnight at 65°C in 10ml of 5xSSC; 2xDenhard ts ; 100gg/ml denatured herring sperm DNA; 10% dextran sulphate; 20mM potassium phosphate (pH 6.5) and the denatured radiolabelled probe. Posthybridisation washes were at 65°C in 3 x SSC plus 0.1% sodium dodecyl sulphate (SDS) for 10 to 15 min. The stringency of washing was increased by incubation in decreasing SSC concentrations (1 x SSC-0.1 x SSC) plus 0.1% SDS at 65°C for 15 to 30 min, dependent on which probe was being used. Autoradiography onto Fuji X-ray film was at - 70°C for 1 to 5 days with intensifying screens

64
corresponding to 27 bases of the rat ornithine transcarbamyl- ase (OTC) sequence. OTC was cloned into plasmid pBR322 and localised to the region Xp21-Xpl14 (Davies et al. 1985). It detects a polymorphism with Mspl and for this purpose the OTC insert was cut out of plasmid pBR322 and labelled inde- pendently. OTC has a rare allele frequency of 0.36.
All seven probes have been mapped as either proximal or distal to the DMD locus using rodent-human cell hybrids made from a female with muscular dystrophy due to an X;21 translocation, considered to represent a breakpoint on the X chromosome at or near the Duchenne gene (Worton et al. 1984).
Preparation of human DNA
Total human DNA was extracted from whole blood in EDTA by the method of Kunkel et al. (1977). DNA samples of 3gg were digested to completion using the appropriate restriction enzyme and the fragments were separated according to molecular weight by agarose gel electrophoresis (0.8% gels). Blotting onto Zeta-probe membrane (Biorad) was carried out by the method of Southern (1975) with some modifications (see legend to Fig. 1). Figure l a - f shows the alleles detected by each probe which are defined by the presence or absence of restriction enzyme recognition sites.
DNA probe labelling
All the DNA probes, except OTC, were radioactively labelled by nick translation, to a specific activity of 0.5-2.0 x 108 dpm/ gg, using a commercial kit (Amersham). The OTC insert was excised from its vector and radio-labelled using an oligo- nucleotide-primed reaction (Feinberg and Vogelstein 1984) to high specific activity. Probe prepared by either radiolabelling method was separated from unincorporated nucleotide by G-50 chromatography, denatured by boiling for 10rain, chil- led on ice, and mixed with the hybridisation solution.
Families
Pedigrees in which DMD or BMD had occurred in at least two males were studied, thus excluding isolated cases possibly aris- ing by new mutation. In the case of BMD, only families with two or more generations affected were studied to avoid possi- ble confusion with autosomal recessive dystrophies. Only those females who were obligatory carriers, together with affected and unaffected males were used for linkage analysis.
Linkage analysis
For all these linkage studies two point analysis was carried out using LIPED 3 (Ott 1974), and Lod scores calculated at recombination fraction (0) intervals of 0.01. All the pedigrees have also been examined manually to record the number of recombinant events occurring between the two-paired loci under study.
Results
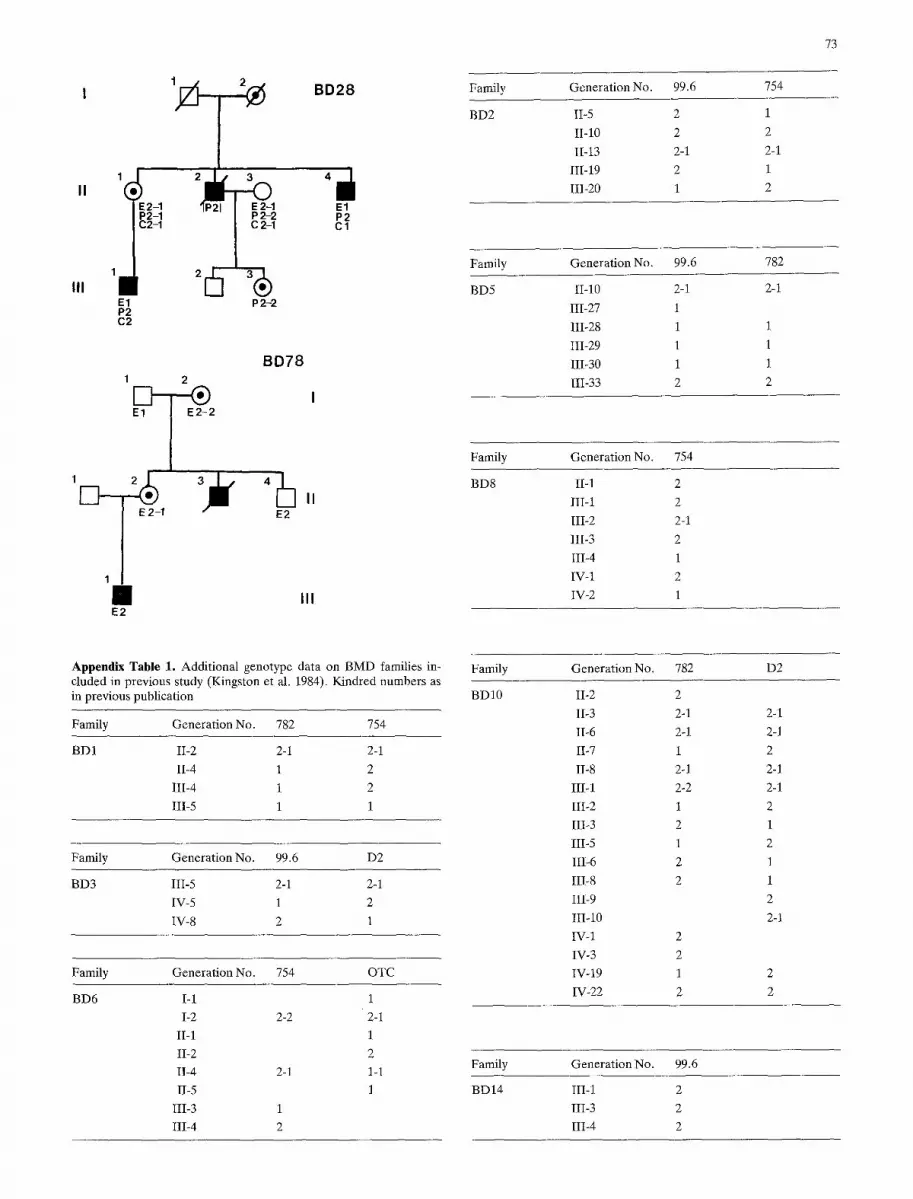
Thirty three informative DMD families and 26 informative BMD families were analysed for genetic linkage to each of the probe loci. The genotypes of informative individuals used in the linkage analysis are shown on the pedigrees of the 33 Duchenne families (Appendix Fig. 1) and five Becker families (Appendix Fig.2), and in the Appendix Table i for the remaining Becker families previously documented by King- ston et al. (1984).
The Lod scores obtained at 0.05 intervals of the recombi- nation fraction 0, and the 95% confidence limits of 6 for link- age between the seven polymorphic loci and the disease loci are shown in Table 1 for BMD, Table 2 for DMD, and Table 3 for the combined BMD and DMD families. In each case the maximum Lod score obtained from analysing 0 intervals of 0.01 is also presented.
Table 1. Lod scores for Becker muscular dystrophy and marker loci
0 782 RC8 ~ 99.6 D 2 754 b OTC L 1.28 a n = 8 n = 6 n = 14 n = 10 n = 14 n = 7 n = 18
0.01 -3.285 -2.769 -1.394 -7.719 -5.740 -1.215 -5.508 0.05 -0.167 -0.290 2.198 -2.018 -0.236 1.740 1.266 0.10 0.851 0.503 3.194 -0.070 1.459 2.546 3.382
0.15 1.222 0.781 3.392 0.729 2.005 2.700 4.071 0.20 1.316 0.841 3.234 1.044 2.063 2.573 4.147
0.25 1.248 0.781 2.871 1.091 1.861 2.285 3.867
0.30 1.074 0.644 2.369 0.966 1.505 1.894 3.349 0.35 0.827 0.465 1.781 0.738 1.072 1.443 2.663 0.40 0.541 0.277 1.154 0.459 0.630 0.960 1.855 0.45 0.255 0.111 0.546 0.190 0.255 0.473 0.958
Maximum 0 0.20 0.19 0.15 0.24 0.18 0.15 0.18
Maximum scores 1.316 0.842 3.392 1.098 2.079 2.700 4.170
95% confidence limits of 0 0.07-0.41 0.06-0.50 0.06-0.28 0.11-0.44 0.09-0.33 0.05-0.30 0.10-0.30
0 Recombination fraction n Number of informative families a Includes some data published by Kingston et al. (1983, 1984) b Includes data given by Brown et al. (1985)

Table 2. Lod scores for Duchenne muscular dystrophy and marker loci
0 782 RC8a 99.6 D2 754b OTC L1.28a n - - 10 n = 10 n = 14 n = 13 n = 17 n = 9 n = 20
65
0.01 -5.975 -1.290 -2.945 -3.319 -5.810 0.720 -6.475
0.05 - 1.511 0.768 0.679 -0.338 -0.822 2.982 -0.845
0.10 0.041 1.433 1.752 0.504 0.804 3.494 1.007
0.15 0.701 1.615 2.057 0.717 1.411 3.479 1.716
0.20 0.980 1.578 2.049 0.677 1.587 3.233 1.953
0.25 1.049 1.414 1.860 0.523 1.528 2.843 1.932
0.30 0.983 1.170 1.568 0.337 1.318 2.362 1.744
0.35 0.824 0.886 1.211 0.181 1.020 1.812 1.447
0.40 0.599 0.584 0.823 0.070 0.677 1.220 1.052
0.45 0.323 0.290 0.414 0.012 0.326 0.609 0.573
Maximum 0 0.25 0.17 0.17 0.16 0.21 0.12 0.22
Maximum scores 1.049 1.623 2.082 0.722 1.594 3.529 1.968
95% confidence limits of 0 0.11-0.47 0.05-0.36 0.07-0.35 0.06-0.50 0.10-0.39 0.04-0.26 0.10-0.39
0 Recombination fraction n Number of informative families a Includes some data published by Davies et al. (1983) b Includes data given by Brown et al. (1985)
Table 3. Lod scores for Duchenne and Becket muscular dystrophy and marker loci (combined)
0 782 RC8 99.6 D2 754 OTC L1.28 n = 18 n = 16 n = 28 n = 23 n = 31 n = 16 n = 38
0.01 - 9.260 - 4.059 - 4.339 - 11.038 - 11.550 - 0.495 - 11.983
0.05 -1.678 0.478 2.877 - 2.356 - 1.058 4.722 0.421
0.10 0.892 1.936 4.946 0.434 2.263 6.040 4.389
0.15 1.923 2.396 5.449 1.446 3.416 6.179 5.787
0.20 2.296 2.420 5.283 1.721 3.650 5.806 6.100
0.25 2.297 2.195 4.731 1.614 3.389 5.128 5.799
0.30 2.057 1.814 3.937 1.303 2.823 4.256 5.093
0.35 1.651 1.351 2.992 0.919 2.092 3.255 4.110
0.40 1.140 0.861 1.977 0.529 1.307 2.180 2.907
0.45 0.578 0.401 0.960 0.202 0.581 1.082 1.531
Maximum 0 0.22 0.17 0.16 0.21 0.19 0.13 0.20
Maximum scores 2.333 2.449 5.457 1.726 3.653 6.208 6.100
95% confidence limits of 0 0.12-0.36 0.08-0.32 0.08-0.26 0.12-0.35 0.11-0.30 0.07-0.23 0.12-0.29
0 Recombination fraction n Number of informative Duchenne and Becker muscular dystrophy families
Discussion
Our da ta show genet ic l inkage b e t w e e n D M D , B M D , and five new po lymorph ic D N A probes (782, 99.6, D2, 754, and OTC) , in addi t ion to the two originally s tudied (RC8 and L1.28). The mos t l ikely genet ic dis tances range b e t w e e n 12 cM ( D M D - O T C in Tab le 2) and 2 5 c M (DMD-782 in Tab le 2); the l inkage da ta for L1.28 and D M D have b e e n upda ted , slightly increas ing the mos t likely overal l l inkage dis tance to 20cM.
Mapp ing of the seven p robes using a hybr id cell l ine wi th a b r e a k p o i n t at p21 divides t h e m into two groups , one distal to D M D and B M D (782, 99.6, D2, and RC8) , the o the r proxi- mal (754, OTC, L1.28). A l t h o u g h the value of 0 wi th respect
to D M D and B M D gives some indica t ion of the i r relat ive o rder ing wi th in these groups, the conf idence limits are wide, and a more precise o rder ing will need to be based on mul- t ipoin t analysis of the p r o b e - p r o b e data , on obse rva t ion of individual crossovers in families in format ive for m o r e t han one po lymorph i sm and on analysis of cell l ines showing dele- t ions in the p21 region.
Our cur ren t results are of par t icular in te res t in conf i rming tha t the gene for B M D is loca ted in the same region of the shor t a rm as tha t for D M D , as suggested by the prev ious da ta of Kings ton et al. (1983, 1984). C o m p a r i s o n of the distances in Tab les 1 and 2 shows essential ly s imilar values of 0 for all seven p robes and makes it l ikely tha t the two disorders may be

66
allelic. Combining the D M D and B M D data (Table 3) does not significantly alter the most likely genetic distances, but does allow a more precise localisation with narrower confi- dence limits; the data also are strongly against any significant multi-locus heterogenei ty within ei ther B M D and D M D .
Apar t f rom being tools for mapping the disease loci, these l inked probes can be used in the calculation of carrier risks of females f rom affected families, even though none of them are very closely l inked to D M D and B M D . Previously, L1.28 and RC8 have been used for this purpose, in conjunction with creatine kinase data (Harper et al. 1983; Pembrey et al. 1984; Kingston et al. (1985). The detailed application of the present probes in carrier predict ion will be discussed elsewhere, but if the heterozygote frequencies of all seven probes are taken into account a total of 98% of women will be heterozygous for at least one locus and could potential ly benefit f rom improved estimates of carrier risk.
Acknowledgements. We would like to thank Dr. Tom O'Brien, Dr. Hywel Williams, and our colleagues in other centres for obtaining blood samples as well as the many families who provided them. Dr. Victor Ionasescu of Iowa City kindly provided samples from family D173, the only kindred from outside the UK. We also thank Dr. Linda Meredith for practical assistance, and Mr. D. M. Katibeh for his help in computer data preparation. Dr. Ron Worton kindly supplied DNA from his cell hybrids containing the translocated X:21 chromo- some. This work was supported by grants from the Muscular Dys- trophy Group of Great Britain and the Muscular Dystrophy Associa- tion of America.
References
Becker PE, Kiener F (1955) Eine neue X-chromosomale Muskeldys- trophie. Arch Psychiatr Nervenkr 193 : 427-448
Brown CS, Pearson PL, Thomas NST, Sarfarazi M, Harper PS, Shaw DJ (1985) Linkage analysis of a DNA polymorphism proximal to the Duchenne and Becker muscular dystrophy loci on the short arm of the X-chromosome. J Med Genet 22:179-181
Davies KE, Young BD, Elles RG, Hill ME, Williamson R (1981) Cloning of a representative genomic library of the human X chromosome after sorting by flow cytometry. Nature 293 : 374-376
Davies KE, Pearson PL, Harper PS, Murray JM, O'Brien T, Sar- farazi M, Williamson R (1983) Linkage analysis of two cloned DNA sequences flanking the Duchenne muscular dystrophy locus on the short arm of the human X-chromosome. Nucleic Acids Res 11:2303-2312
Davies KE, Briand P, Ionasescu V, Ionasescu G, Williamson R, Brown CS, Cavard C, Cathelinean L (1985) Gene for OTC: Characterisafion and linkage to Duchenne muscular dystrophy. Nucleic Acids Res 13 : 155-184
Feinberg AP, Vogelstein B (1984) A technique for radiolabelling DNA restriction fragments to high specific activity. Anal Biochem 137 : 266-267
Harper PS, O'Brien T, Murray JM, Davies KE, Pearson P, William- son R (1983) The use of linked DNA polymorphism for genotype prediction in families with Duchenne muscular dystrophy. J Med Genet 20 : 252-254
HoNer MH, Wapenaar MC, Goor N, Bakker B, Van Ommen GJB, Pearson PL (1985) Isolation of probes detecting restriction frag- ment length polymorphisms from X chromosome specific librar- ies: Potential use for diagnosis of Duchenne muscular dystrophy. Hum Genet 70:148-156
Kingston HM, Thomas NST, Pearson PL, Sarfarazi M, Harper PS (1983) Genetic linkage between Becker muscular dystrophy and a polymorphic DNA sequence on the short arm of the X chromo- some. J Med Genet 20: 255-258
Kingston HM, Sarfarazi M, Thomas NST, Harper PS (1984) Localisa- tion of the Becker muscular dystrophy gene on the short arm of the X-chromosome by linkage to cloned DNA sequences. Hum Genet 67: 6-17
Kingston HM, Sarfarazi M, Newcombe RG, Willis N, Harper PS (1985) Carrier detection in Becker muscular dystrophy using creatine kinase estimation and DNA analysis. Clin Genet 27 : 383- 391
Kunkel LM, Smith KD, Boyer SH, Borgaonkar DS, Watchell SS, Miller OJ, Breg WR, Jones HW Jr, Rary JM (1977) Analysis of human Y-chromosome-specific reiterated DNA in chromosome variants. Proc Natl Acad Sci USA 74:1245-1249
Kunkel L, Tantrahavi U, Eisenhard M, Latt S (1982) Regional locali- sation on the human X of DNA sequences cloned from flow sorted chromosomes. Nucleic Acids Res 10 : 1557-1561
Murray JM, Davies KE, Harper PS, Meredith L, Mueller CR, Wil- liamson R (1982) Linkage relationship of a cloned DNA sequence of the short arm of the X-chromosome to Duchenne muscular dys- trophy. Nature 300 : 69-71
Ott J (1974) Estimation of the recombination fraction in human link- age studies. Am J Hum Genet 26 : 588-597
Pembrey ME, Davies KE, Winters RM, Elles R, Williamson R, Faz- zone TA, Walker C (1984) Clinical use of DNA markers linked to the gene for Duchenne muscular dystrophy. Arch Dis Child 59 : 208-216
Southern EM (1975) Detection of specific sequences among DNA fragments separated by gel electrophoresis. J Mol Biol 98:503- 517
Worton RG, Duff C, Sylvester JE, Schmickel RD, Huntington FW (1984) Duchenne muscular dystrophy involving translocation of the dmd gene next to ribosomal RNA genes. Science 224 : 1447- 1449
Zatz M, Viannia-Morgante AM, Campos P, Diament AJ (1981) Translocation (X;6) in a female with Duchenne muscular dys- trophy: Implications for the localisation of the DMD locus. J Med Genet 18: 442-447
Received May 8, 1985

D3
F2-1 I IF21 C2-1 P2-1
"
III F2 C1 P2
Appendix Fig. 1. Pedigrees with genotypes of DMD families
Key to pedigrees (Appendix)
B RC8 (DXS9) alleles C L1.28 (DXS7) alleles E 99.6 alleles F D2 alleles ( - ) inferred genotype ] 782 alleles L OTC alleles P 754 alleles Probe alleles in heterozygous females are written with the rarer allele first
1 2
1 1 2
] P2-2 I L I - I
II 1j 2 ~ j 2_ 1 B1 I B2-1 P1 I E 2 - 1 L1 I F2-1
I P2 -1 ~-' III 11~
J1 B1 E2 F2 P2 L1
E 1 - 1 P2-1 L2-1 C2-2
II
III
IV
E 2 P2 L1 C1
LC2J I P2-1 IEli I L2-1 [P21 I C2-1
E1-1 P2-2 L1-1
1
E1 P2 L1 C2
C2-2
2 q
E1 P2 L1 C1
D4
D13
61 .,o~
67
J 2 J2 -1 B1 B2-1 C2 E2-1
F 2 - 1 :1
,,, '~ ~ ~i '~ J 2 J2 B2 B2 E2 E1 F1 F1 C2 C1
1 ~ . ~ D28
~-~6 'm ~i '-"r~
III L1 C1
C1 I L2_1 I C2-1
J 2 L1 C1
1 2
1
3r~
J1 L2 C2
D 3 4
J2 L2 C2

68
,, '~] B2
III
. ~ D48 D83 D93
1 2 1 2
/ i F2-1 P2 i P2-1
B2-1 I [ ~ E ~ ' ' 1 ~ 2 ~ , t 3 (~ " 1 2 3
1 P2-1/ F2 F1 P2 P2 J1 J2 C1 C1 C1 B1 B2 E2 E1
1 I I I
B2 ~'B2 P2 C1 C2
D103
,-2 b 3D_
1 / ~ ~ 3 4 ( ~ ~ 6 6 7L-L, I I I [ ] I R2-~ , - - B2
,, " ' 1-3(~ 4~._] b (~ 6 6 '~(~ 8 ~,] 9 ~10-11(~ 12-'13~
B1 B2 C1 C1
14
[ ] 'l 9
'I 10
I I
I I I
IE l l E 2 - 1 C1 E 2 - 1 [P2} F2-1 F2-1 P2-1 p1-1
C1-1 C2-1
1 6 2 / 3 i 4 6 E1-1 E2 E2 P2-2 F 1 F2
P1 P1 C1 C2
-1
5 6 -
D106
II 1 ( ~ 2 3
1 2 ~ 4 III
IV
1 2 J2-1 p2-1
'I % J1 P2 c2
lO 11(~
D129
8 9 10
J1 P1 C2
12 14(~ 156 16 / J2 C1

69
I
1
III 1 ~- J2 E2
E2-1
J2 E2
1 2
D 1 3 5
1 2 3 4 5 II
B2~ 1 B1
1 5
III
IV B2 B2
E2-1 F2-1 D 1 3 8 D 1 3 9 p1-1 C2"1
1 2
B1 B2-1 II C2 C2-1
I E2-1 I F2 -1 I F2-.1 I P2-1
J C2-1 I C2-1
I m 2 1 B 2 ~ ' ~ I" [_L] 2~_] 3 i ,, / ~J, " ~, ' J 2 J1 C2 C2 E2 E2 E2 E 1 E2 IF2I IF2] IF2] F1 F2 P2 P2 P1 C2 C2 C1 C2 C2
1 2
I ~ [L21 I I-1-1 iC2] ] C1-1
1 ~ ,, ' ~ ~il I ~ D144 j ~ 1 B1 I B 1-1 " L2 I !_2-1
IC
' ~ ~ ~ 4~, I I B2 I J2-1 J21 I J1
J2 J1 J1 J2 B1 B1 B1 B1 L1 L1 L2 L1 C1 C1
1 2
, E] 0 D147
D137
B1 B1
D 1 4 0
3 [ ]
IF21 C ~,
E1 F2 Pl C1
D 1 4 6
C1 C2-2 C2
p '@ c c~ ' D - ~ ' 6 ' 6 9 6 o / 1@
I I | C1

"70
D150
C2
,, ~ b 'b C2
DI51
1 2 3
"6 % 6,,I, C1-1 C1
I I
I I I J1 E1 L1
1
F1 J1 IEll P2 L1
1
E] [Fl l L2
J2 -1 E2 -1 F2-2 P2-2 L2-1
E1 L2
J2 E2 F1 P1 L1
J1 E2 L2
J2 E2 L2
® J2-1 E2"2 F2"1 P2"1 L2-1
D152
,Q J1 E2 F1 P1 L1
s-8~
5 6
D155
1 2
E2 E2-1 IF2t F 1-1 L2 L1-1
II r__]l | l E2-1 E1 F2-1 L1 L2"1
III E1 F1 L1
II
III ! 6
I P2-2
F1 F2 P2 P2
3 6 '6
F2 F2 P1 P1
D156
[ ~ ~ F 2 2
Z P2
'6

71 I 2
D157
I F 2":1 I P 2-1
,,, '.1, ~6 ~6 ",i ' ~6 °rt 'rh E1 E2-2 E2 F2 P1 P2
8 6
1 2 3 [ ] ® [ ] D16o E2-1 E1 P2-1 P1
E1 E1 E1 E2 P2 P1 P1 P2
II
1 III (
1
iv [
26
1 D ICll I
36 'F~
.I,, 6 6 P2 P 2 - 2 C2
2 ® lC2-11
D162
I~: I 7 E
1 2
I E 2 P L1
2-1 J 2 2-1 E2 I E 2-1
I P2-2 P1 IP2-1 ,L,-1 L~ ,L~4 L ~ C~ IC~-1 ,,, : E ~ ' i
J2 J2 J1 E2 E2 E1 P2 P2 P1 L1 L1 L2 C1 C1 C2
D 1 6 3
J2 E2 P1 L2 C2

72
D164 D168 D169 1 2 1 2 1 2 1 2
I / / _ I ~ . I ~ . i i i~ ~ ~ ~ F 2 - 1 C2 -1
II II II II P1 P1 E1 E1 P1 P1 F2 F1
C2 C2
D170
1 2 I ~ D173
-~ ~ ' r ~ 5 °m ~_9~ IF1] F2-1 . F2 L1 F2 2-2 L1 L2-2 L1 L1
III 1 s 617 L 2 F 1-1 F2 2-1
L2-1 L2
2 i 3 5
F1 F1 L2 L2
7 t
iI l
III
BD7
2 1 P1 E1 E, 2 - , 1-1
'D ~;, 'm ' i E2 E1 III F2 F1 E2 E1 P1 P2 P1
III
IV
1 1 BD19
I E2-1 i 2 -~
I J2-1 E2
J2 E1 P2
Appendix Fig.2. Pedigrees with genotypes of BMD families not included in previous study (Kingston et al. 1984)
I I
I I I II E1 P2 C2
E2-1 P2-1 C2-1
P2-2
BD28
' i E1 P2 C1
-1
2 E3-- E1 E2-2
BD78
E2
E2
Appendix Table 1. Additional genotype data on BMD families in- cluded in previous study (Kingston et al. 1984). Kindred numbers as in previous publication
Family Generation No. 782 754
BD1 II-2 2-1 2-1
II-4 1 2
III-4 1 2
III-5 1 1
Family Generation No. 99.6 D2
BD3 III-5 2-1 2-1
IV-5 1 2
IV-8 2 1
Family Generation No. 754 OTC
BD6 I-1 1
I-2 2-2 2-1
II-1 1
II-2 2
II-4 2-1 1-1
II-5 1
III-3 1
III-4 2
Family Generation No. 99.6 754
BD2 II-5 2 1
II-10 2 2
II-13 2-1 2-1
III-19 2 1
111-20 1 2
Family Generation No. 99.6 782
BD5 II-10 2-1 2-1
III-27 1
III-28 1 1
III-29 1 1
III-30 1 1
1II-33 2 2
Family Generation No. 754
BD8 I1-1 2
III-1 2
III-2 2-1
III-3 2
[II-4 1
IV-1 2
IV-2 1
Family Generation No. 782 D2
BD10 II-2 2
II-3 2-1 2-I
II-6 2-1 2-I
II-7 1 2
II-8 2-1 2-1
III-1 2-2 2-I
III-2 1 2
III-3 2 1
III-5 1 2
III-6 2 1
III-8 2 1
III-9 2
III-10 2-1
IV-1 2
IV-3 2
IV-19 1 2
IV-22 2 2
Family Generation No. 99.6
BD14 III-1 2
III-3 2
III-4 2
73

74
Appendix. Table 1 (continued)
Family Generation No. 782 754
BD17 I-1 1 1
II-1 1-1 2-1
II-3 2 1
III-2 1
Family Generation 782 99.6 OTC No.
B D l l II-2 2-1
II-5 2-1
II-6 1
II-7
II-8 1-1 1-1 2-1
II-9 2
III-1 1
III-13 1
III-16 2-1 2-1 1-1
III-19 1
III-20 2
IV-15 1 1 1
Farriily Generation 782 D2 OTC No.
BD16 II-7 1-1 2-2 2-2
III-12 2 1 1
III-13 1-1 2-2 2-1
IV-11 2-1 2-1 2-1
Family Generation 782 754 OTC No.
BD20 I-2 2-1 2-1
II-1 2 I
II-2 2-2 2-1 2-1
III-1 2 2
III-2 2-2 2-2 2-1
1V-1 2 2 2
IV-2 2 2 1
Family Generation No. D2
BD22 II-1 2-1
II-5 1-1
III-1 2
III-2 2
III-3 2
III-4 2
Family Generation 782 D2 OTC No.
BD24 I-1 2
I-2 1-1 2-1 1-1
II-1 2-1 2-1 2-1
II-4 2-1 2-1 2-1
III-1 1 1 1
III-5 1 1
III-7 1
Family Generation No. 99.6 D2
V-8 1 2 2 BD30 II-3 1
lII-2 2-2
III-3 2-1
IV-6 2 Family Genera- 782 99.6 D2 754 OTC
tion No. 1V-7 2 IV-13 2
BD18 II-7 2-1 2-1 2-1 2-1 2-1
III-1 1-1 2-2 2-1 III-4 2 2 2 1 1 Family Generation No. 754
III-5 1-1 1-1 2-1 2-2 2-1 BD23 I-2 2-2
III-10 1 2 1 II-3 2-1
III-11 2-1 2-2 2-2 2-I 2-1 II-7 2-1
III-14 2 2 1 1 III-26 2
III-15 1-1 2-1 2-1 2-I 2-1
III-16 2 2 2 1 1 IV-2 1-1 1-1 2-1 2-1 1-1 Family Generation No. 754
IV-4 1 1 2 2 BD25 III-1 2
IV-12 1 2 2 III-2 2
IV-13 2 2 III-4 2
IV-14 1 2 1 1 III-6 2
IV-15 2 1 2 IIL7 2
IV-16 2 2 1 l IV-4 2-2
IV-25 2 2 2 2 IV-5 2
IV-26 1 2 2 IV-6 2-1
V-1 1 1 V-1 2
V-3 2 V-3 2
2
2-1
2-2
2
1 2
Recommended

















