
DOI 10.1007/s00702-003-0070-2
J Neural Transm (2004) 111: 91–100
Hippocampal volume and cell proliferation after acute
and chronic clozapine or haloperidol treatment
A. Schmitt1, S. Weber1, A. Jatzko2, D. F. Braus2, and F. A. Henn1
1 Central Institute of Mental Health, Mannheim, and2 Clinic for Psychiatry and Psychotherapy, University of Hamburg, Hamburg,
Germany
Accepted August 25, 2003Published online December 3, 2003; # Springer-Verlag 2003
Summary. The dentate gyrus contains progenitor cells possessing the capac-ity to proliferate until and throughout adulthood. There is little informationabout the influence of antipsychotics on cell proliferation. To address this, weinvestigated the influence of acute and chronic haloperidol and clozapinetreatment on the total number of newly dividing cells and hippocampalvolume using an animal model with doses equivalent to the therapeutic rangein humans.
Rats were treated with either acute or 28 days haloperidol (1 mg=kg i.p. or1,5 mg=kg=day oral) or clozapine (30 mg=kg i.p. or 45 mg=kg=day oral).After BrdU injection, immunohistochemistry was performed in serial hippo-campal brain sections. Total BrdU-labeled cell number and hippocampusvolume were estimated using stereological methods. Neither neuroleptic alteredtotal number of newly dividing cells in the dentate gyrus. In contrast, chronichaloperidol treatment did increase total hippocampal volume suggesting thathaloperidol alters neuroplastic processes or glial morphology rather than cellproliferation.
Keywords: Haloperidol, clozapine, cell proliferation, hippocampus, stereology.
Introduction
During development, the brain is composed of progenitor cells, which prolif-erate and differentiate into neurons and glial cells. In recent years it has becomeevident, that in addition to the subventricular zone (Conover and Allen, 2002)the subgranular layer of the hippocampal dentate gyrus also contains progenitorcells possessing the capacity to proliferate until and throughout adulthood. Thishas been proven in different mammalian species, such as rats (Cameron et al.,1993), mice (Kempermann et al., 1997), tree shrews (Gould et al., 1997),makakes (Gould and Tanapat, 1999) and humans (Eriksson et al., 1998; Nunes

et al., 2003). In the gentate gyrus of adult rats, division and differentiation of themajority of those progenitor cells to neurons and their migration to the granulecell layer has been demonstrated (Cameron et al., 1993; Kempermann et al.,2003).
There is only little information about the factors influencing the neurogen-esis. While blockade of the glutamatergic NMDA (N-methyl-D-aspartate)receptor and adrenalectomy resulted in increases in the production of granuleneurons, adrenal steroids and NMDA receptor activation diminished neurogen-esis (Cameron et al., 1998). Psychosocial stress has been shown to inhibitproliferation of subgranular neurons in rat, monkey and tree shrew brain (Gouldet al., 1997; Gould and Tanapat, 1999; Tanapat et al., 1998). Because bothcirculating adrenal steroids and glutamate-induced excitatory input to the hip-pocampus are enhanced by stress (Krugers et al., 1993; Moghaddam et al.,1994), the influence of stressful events on cell proliferation and survival ofnewly generated neurons may be mediated by these mechanisms (Czeh et al.,2002; Gould and Tanapat, 1999).
Stress is a common, unspecific factor in psychiatric diseases such asaffective disorder and schizophrenia. In addition, magnetic resonance imagingstudies demonstrate that hippocampal volume is decreased in patients withrecurrent depression and chronic schizophrenia (Bremner et al., 2000;McCarley et al., 1999). This volume loss may be mediated by stress-inducedglucocorticoid neurotoxicity (Arango et al., 2001). Psychopharmacologicaltreatment reduces disease-related stress and chronic antidepressant treatmentas well as ECT is known to increase cell proliferation and neurogenesis inadult hippocampus (Coyle and Duman, 2003; Malberg et al., 2000). A highdose of the traditional neuroleptic haloperidol has been shown to stimulategranule cell proliferation after acute treatment (Dawirs et al., 1998), whilesubchronic haloperidol treatment at lower doses showed no difference in thedentate gyrus (Malberg et al., 2000; Wakade et al., 2002). In the subventric-ular zone, 21 days of treatment with the atypical antipsychotics olanzapineand risperidone increased cell proliferation. However, no differences havebeen found within the hippocampus using two-dimensional cell countingmethods (Wakade et al., 2002). To date, there are no studies investigatingacute and chronic treatment with the atypical antipsychotic clozapine. Clo-zapine is known to support plasticity in animal models (Konradi and Heckers,2001), as well as to enhance learning and cognition in schizophrenic patients(Potkin et al., 2001) and hippocampus-mediated learning has been shown toincrease neurogenesis (Gould, 1999; Shors et al., 2002). Thus, increasedproliferation of neuronal progenitor cells in this area may be one mechanismof antipsychotic drug action and be advantageous for the amelioration ofcognitive symptoms and learning abilities. Two-dimensional cell countingin only a few sections without considering the total hippocampus volumemay have some methological limitations through the influence of medicationon hippocampus volume and different shrinking of tissue after fixation lead-ing to large variations in section thickness. Additionally, problems may becaused by irregular cell shape and size, nonrandom orientation and cutting ofcells during sectioning (Williams and Rakic, 1988). To address this, we
92 A. Schmitt et al.

investigated the influence of acute and chronic haloperidol and clozapinetreatment on hippocampal volume and total number of newly dividing cellsin an animal model with doses equivalent to the therapeutic range in humansusing stereological, three-dimensional cell counting methods.
Material and methods
Animal drug treatment
Animal use procedures were in strict accordance with the NIH guidelines for the care and use oflaboratory animals and had been approved by the local ethics committee.
Male Sprague-Dawley rats (initial weight 200–210 g, Janvier, France) were group-housed 3per cage under a 12 hour light-dark cycle and with food and water ad libitum. In the acutetreatment condition, 9 rats per group received a single dose of either haloperidol (Janssen,Beerse, Belgium) 1 mg=kg i.p., clozapine 30 mg=kg i.p. (Novartis, Basel, Switzerland), dis-solved in 0.9% NaCl, or saline i.p. at 10 a.m. In the chronic treatment condition lasting 28days, 9 rats received haloperidol 1,5 mg=kg=day diluted in minimal acetic acid, while 9 ratsreceived clozapine 45 mg=kg=day diluted in minimal HCl in the drinking water adjusted to pH6,5. 9 age-matched control rats received drinking water adjusted to pH 6.5 by HCl. The drugdosage was calculated twice per week by the amount of ingested water and the animal’s bodyweight. Oral treatment regimes were selected in order to provide continuous levels of theantipsychotics to mimic the clinical situation. The oral doses were set higher than parenteraldosages reported in the literature to account for first-pass metabolic effects on bioavailability(Fitzgerald et al., 1995; See and Ellison, 1990). Haloperidol was given at a dose, whichproduces plasma concentrations in animals equivalent to clinical relevance in human condi-tions. Accordingly, the target dose in rats (1,5 mg=kg=day) has to be set higher than underhuman conditions because rats have faster metabolic rates (Fitzgerald et al., 1995; See andEllison, 1990). Using this dosage as a reference point, we selected the respective clozapinedose on the basis of clinical equivalency (30-fold of haloperidol) (Fitzgerald et al., 1995).After chronic drug administration in the drinking water at 10 a.m. on day 28, rats of thechronic treatment groups received a single dose of saline i.p. to account for the possibleeffects of stress associated with the i.p. injection in the acute and chronic treatment groups.In the acute treatment condition, body and brain weight did not differ between the single i.p.doses of haloperidol, clozapine and controls. Additionally, after four weeks of oral treatment,body weight of rats (405� 32 g) and total brain weight (2.07� 0.24 g) did not differ betweentreatment and control groups.
BrdU injection
One hour after the last i.p. drug or saline administration, bromodeoxyuridine (BrdU, Sigma,Taufkirchen, Germany) dissolved in distilled water, was injected intraperitonally in a volumeof 0.5 ml or less per rat to deliver a dosage of 200 mg=kg, for labeling the newly dividingcells. BrdU, a thymidine analogue, is incorporated in to the DNA of dividing cells during theS-Phase and is detected using a specific, monoclonal antibody (Gratzner, 1982). A two hoursurvival time was selected because it is sufficient for the uptake of BrdU but not for comple-tion of mitosis or migration (Cameron et al., 1993). Two hours after BrdU injection, rats wereanaesthesized under ketamine (100 mg=kg bw)=xylazine (5 mg=kg bw) anaesthesia and sacri-ficed by cardiac perfusion with sodium phosphate buffer (pH 7.4) followed by formaldehyde4% in S€oorensen buffer 0.2 M (NaH2PO4 and Na2HPO4) for rapid brain fixation. Brains wereremoved and the complete brain at the level of the hippocampus was immediately cut byvibratome in the coronal plane into 40mm brain sections for immunostaining using the brainatlas of Paxinos and Watson (Paxinos and Watson, 1986). On an average, we analyzed 27sections of each hippocampus. Generally, in the chronic haloperidol group more sections wererequired than in controls (p<0.1).
Antipsychotics and cell proliferation 93

BrdU-Immunohistochemistry
Every 5th section of the hippocampus was mounted on slides for immunohistochemical stain-ing of incorporated BrdU (Fig. 1). After drying in air at 37�C, brain sections were washed atroom temperature in 0.01 M sodium phosphate buffer (PBS, pH 7.4, 5 ten-minute washes)followed by 15 minutes trypsin denaturation at 37�C. After three 10-minute washes with0.01 M PBS at room temperature, DNA was denaturated with 1,2 N HCl for 60 minutes at37�C. Sections were then washed in 0.01 M PBS (3 ten-minute washes) and air dried. Next,the sections were blocked for 60 minutes in a PBS solution (0.01 M) containing 10% bovineserum albumine (BSA) and 0.03% Triton X-100. They were incubated overnight at 4�C in0.01 M PBS containing 1% BSA, 0.03% Triton X-100 and anti-BrdU first antibody (mouseanti-BrdU 1:700, Vectastain ABC Kit, Linaris, Wertheim, Germany). After washing with0.01% PBS (3 ten-minute washes), endogenous peroxidase acitvity was eliminated with1.4% H2O2 for 15 minutes at room temperature. After washing with 0.01 M PBS (three timesfor 10 minutes), the slices were incubated with the biotinylated second antibody (anti-mouse-IgG 1:200, Vectastain ABC Kit, Linaris, Wertheim, Germany) diluted in 1% normal horseserum and 0.03% Triton X-100 for 60 minutes at room temperature. Sections were washedfive times in 0.01% PBS and subsequently incubated with avidin-biotinylated horseradishperoxidase complex (Vectastain ABC-Kit) for 45 minutes at room temperature (Hsu et al.,1981). After three 10 minute washes with PBS, sections were treated with diaminobenzidine(DAB) 0.025% for 5 minutes at room temperature and subsequently washed twice with 0.01%PBS for 10 minutes. After washing in aqua dest. and hematoxylin staining, sections weredehydrated in ethanol, isopentane and xylol, mounted in Eukitt (Riedel-de Haen, Seelze,Germany) and coverslipped.
Quantification and data analysis
In each animal, we sliced the entire hippocampal region into sections with identical thickness(40mm). Every 5th consecutive section was stained and evaluated. BrdU-labeled nuclei wereidentified in the subgranular and granular layer of the left and right dentate gyrus and countedusing a microscope (Olympus BX-51) connected to a color-video camera and a computer system(Microbrightfield, Germany). Stereological analysis was performed using the optical dissector,which defines the counting volume of a section (a thin optical section inside the thick) and acounting frame in a defined depth according to the rules of stereology. All BrdU-labeled nuclei inthe subgranular and granular layer of the dentate gyrus area were counted. Total estimated cellnumber in the dentate gyrus and the volume of the hippocampus was calculated by the area that
Fig. 1. Photomicrographs of dentate gyrus and BrdU-labeled cells (dark) with brown immu-noprecipitates in the cell nuclei
94 A. Schmitt et al.

was sampled on each section of known thickness and the distance between the correspondingsurfaces of the adjacent systematically sampled sections according to the Cavalieri method(Gundersen et al., 1988; West, 1999).
Statistical analysis
Data were analyzed with bivariate analysis of variance (ANOVA). Means were determined foreach animal and the data were subjected to two-tailed Student’s t-test using SPSS 10 statisticalsoftware. Results are given as mean number and standard deviation of total BrdU-labeled cellnuclei of each treatment group.
Results
In the acute treatment group, haloperidol- and clozapine-treated animalsshowed no difference when compared with controls in the total number ofBrdU-labeled cells. Even after the four week treatment period, the totalBrdU-labeled cell number did not differ between the haloperidol, clozapineand control groups. Furthermore, there was no difference between the acuteand chronic treatment condition (Fig. 2).
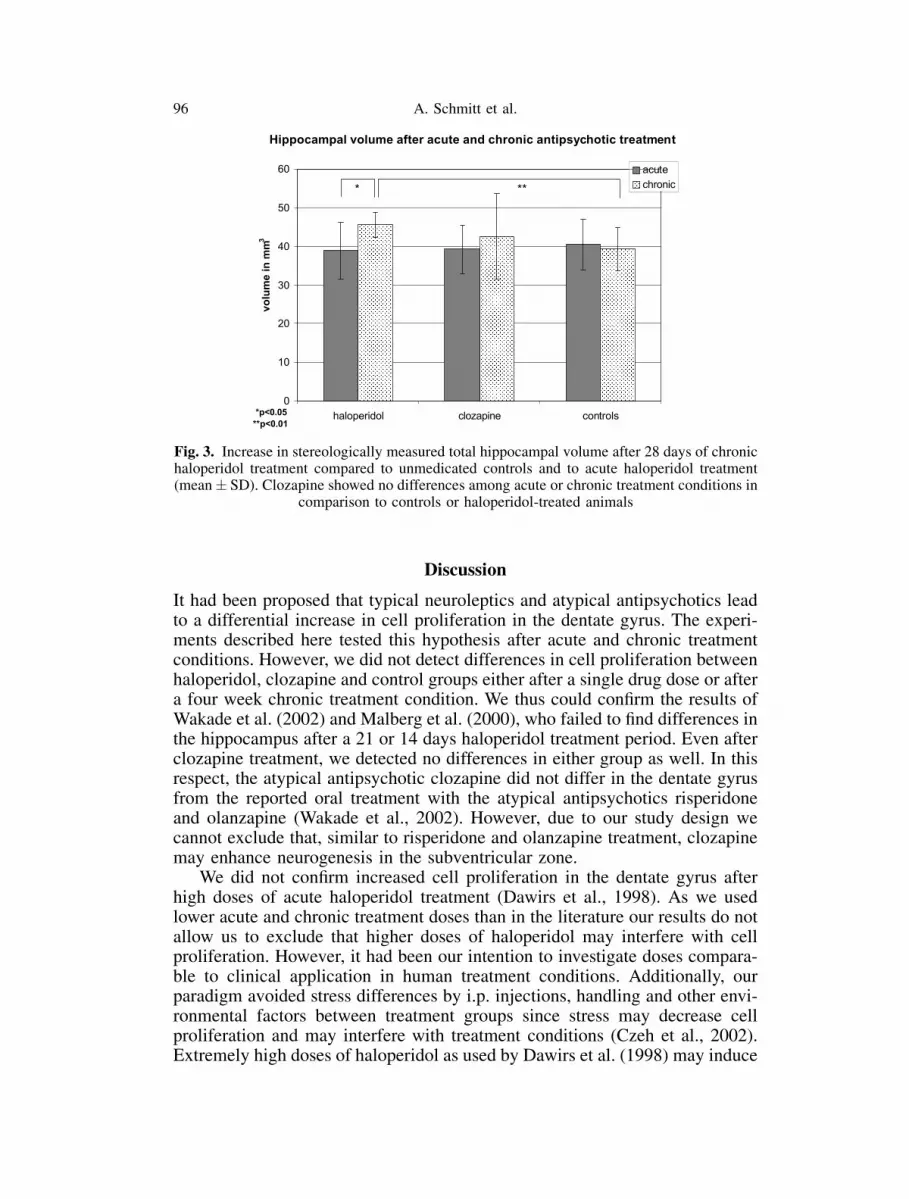
Regarding hippocampal volume, we did not detect any differences afteradministration of a single dose of haloperidol or clozapine. However, after fourweeks of treatment, haloperidol significantly increased the total hippocampalvolume compared to control conditions (by 13.9%, p<0.01), whereas clozapineshowed no difference. Comparing the acute and the chronic haloperidol treat-ment group, the rats in the chronic treatment condition showed a significantlylarger hippocampal volume than those in the acute treatment group (by 14.7%,p<0.05) (Fig. 3).
Fig. 2. There are no differences in estimates of total BrdU-labeled cell nuclei number of thedentate gyrus after single (acute) haloperidol or clozapine injections or 28 days oral haloperidol
or clozapine treatment (chronic) compared to unmedicated controls (mean� SD)
Antipsychotics and cell proliferation 95
Discussion
It had been proposed that typical neuroleptics and atypical antipsychotics leadto a differential increase in cell proliferation in the dentate gyrus. The experi-ments described here tested this hypothesis after acute and chronic treatmentconditions. However, we did not detect differences in cell proliferation betweenhaloperidol, clozapine and control groups either after a single drug dose or aftera four week chronic treatment condition. We thus could confirm the results ofWakade et al. (2002) and Malberg et al. (2000), who failed to find differences inthe hippocampus after a 21 or 14 days haloperidol treatment period. Even afterclozapine treatment, we detected no differences in either group as well. In thisrespect, the atypical antipsychotic clozapine did not differ in the dentate gyrusfrom the reported oral treatment with the atypical antipsychotics risperidoneand olanzapine (Wakade et al., 2002). However, due to our study design wecannot exclude that, similar to risperidone and olanzapine treatment, clozapinemay enhance neurogenesis in the subventricular zone.
We did not confirm increased cell proliferation in the dentate gyrus afterhigh doses of acute haloperidol treatment (Dawirs et al., 1998). As we usedlower acute and chronic treatment doses than in the literature our results do notallow us to exclude that higher doses of haloperidol may interfere with cellproliferation. However, it had been our intention to investigate doses compara-ble to clinical application in human treatment conditions. Additionally, ourparadigm avoided stress differences by i.p. injections, handling and other envi-ronmental factors between treatment groups since stress may decrease cellproliferation and may interfere with treatment conditions (Czeh et al., 2002).Extremely high doses of haloperidol as used by Dawirs et al. (1998) may induce
Fig. 3. Increase in stereologically measured total hippocampal volume after 28 days of chronichaloperidol treatment compared to unmedicated controls and to acute haloperidol treatment(mean� SD). Clozapine showed no differences among acute or chronic treatment conditions in
comparison to controls or haloperidol-treated animals
96 A. Schmitt et al.

neuroleptic analgesia and provide stress differences between haloperidol treatedanimals and controls, accounting for differences in cell proliferation.
Haloperidol is known to be cytotoxic as shown for different neuronal andnon-neuronal cell lines in vitro and in vivo (Behl et al., 1996). In patients takinghaloperidol, pyridinium metabolites are present in low amounts in differentbrain areas including the hippocampus (Eyles et al., 1997). At high acute dosesor chronic exposure with low doses, these metabolites have been shown to beneurotoxic (Bloomquist et al., 1994; Varastet et al., 1994). Based on our datahaloperidol seems not to be toxic on newly dividing cells, since we did not findreduced cell proliferation after haloperidol treatment. However, low haloperidoldoses in the brain could be toxic in vivo after exposure for months or even yearsand our 28-day treatment paradigm may be too short to prove the influence ofneurotoxic metabolites on cell proliferation.
By using the methods described no effects on hippocampal cell proliferationhave been shown, neither by clozapine, nor by haloperidol. However, it cannotbe excluded, that influences on cell proliferation could be detected using longerBrdU-labeling periods (Hildebrandt et al., 1999). Using stereological volumeestimates, we are the first to report hippocampal volume differences in haloper-idol treated animals. According to our results, other mechanisms which do notinfluence cell proliferation may be responsible for the volume difference.
In the striatum, the same dose of haloperidol as used in our study inducedlarger volumes after one month of treatment. This enlargement was associatedwith vacuous chewing movements, an animal paradigm for tardive dyskinesia(Chakos et al., 1996). Additionally, volumetric MRI studies in schizophrenicpatients showed an enlargement of the nucleus caudatus after chronic haloper-idol treatment and a subsequent volume reduction after treatment change toclozapine (Chakos et al., 1995). Anatomical and biochemical effects in thestriatum have been reported after chronic haloperidol treatment. These changesinclude an increase of neuronal size and size of axon terminals (Benes et al.,1985), an increase in asymmetric, glutamatergic synapses and the density ofglutamate immunoreactivity within presynaptic terminals (Meshul et al., 1996).Additionally, dopamine D2 receptors are increased in the striatum after halo-peridol treatment probably related to D2 receptor blockade (Meshul et al.,1994). Dopamine is known to regulate the release of striatal glutamate(Yamamoto and Davy, 1992). This mechanism may not only be restricted tostriatal neurons, but may also occur in the hippocampus. In this region, halo-peridol may act on glutamate synapses by D2 receptor blockade, increasing thevolume of neurons or the neuropil and inducing altered neuroplasticity(Konradi and Heckers, 2001). However, to date, there are no data of neuronalsize or glutamate synapses after haloperidol treatment in the hippocampus.
It has been hypothesized that other biochemical mechanisms may also beinvolved in the volume increase of the hippocampus after haloperidol treatment.Haloperidol has been shown to exert adverse cognitive symptoms and animalstudies report an impaired learning performance after haloperidol treatment,probably related to cholinergic deficits in the hippocampus (Terry et al.,2002). In rabbits, haloperidol treatment induced a blockade of induction oflong-term potentiation in the dentate gyrus (Jibiki et al., 1993), which may
Antipsychotics and cell proliferation 97

be related to altered glutamate function. Additionally, general glial morphologymay be altered after haloperidol treatment, since in the prefrontal cortex, anincrease in glial volume and density has been observed after neuroleptic treat-ment (Selemon et al., 1999).
A reduction of hippocampal volume has been shown in MRI and post-mortem studies of schizophrenic patients (Bogerts, 1999; Pegues et al., 2003;Schulze et al., 2003). Haloperidol may counteract the disease mechanism by abiochemical process increasing the volume of this structure. However, MRIstudies do not report an increased hippocampal volume after haloperidol treat-ment, suggesting that the volume difference in patients may be too small to bedetected. In summary, even comparisons between animal studies and humanconditions have to be considered with caution, since neuroleptics may exertdifferent effects in rat and human brain regions.
Our results suggest that a treatment period of 28 days by either haloperidolor clozapine in doses comparable to clinical application does not influence cellproliferation in the dentate gyrus. In contrast, hippocampal volume wasincreased by chronic haloperidol treatment suggesting that haloperidol influ-ences neuroplastic processes or glial morphology in normal brain tissue. Ourresults encourage future studies investigating these processes to extend ourknowledge about the mechanisms of action of antipsychotic drugs.
Acknowledgement
The work was supported by the Forschungsfonds of the Central Institute of Mental Health.
References
Arango C, Kirkpatrick B, Koenig J (2001) At issue: stress, hippocampal neuronal turnover, andneuropsychiatric disorders. Schizophr Bull 27(3): 477–480
Behl C, Lezoulac’h F, Widmann M, Rupprecht R, Holsboer F (1996) Oxidative stress-resistantcells are protected against haloperidol toxicity. Brain Res 717(1–2): 193–195
Benes FM, Paskevich PA, Davidson J, Domesick VB (1985) The effects of haloperidol onsynaptic patterns in the rat striatum. Brain Res 329(1–2): 265–273
Bloomquist J, King E, Wright A, Mytilineou C, Kimura K, Castagnoli K, Castagnoli N Jr (1994)1-Methyl-4-phenylpyridinium-like neurotoxicity of a pyridinium metabolite derived fromhaloperidol: cell culture and neurotransmitter uptake studies. J Pharmacol Exp Ther 270(2):822–830
Bogerts B (1999) The neuropathology of schizophrenic diseases: historical aspects and presentknowledge. Eur Arch Psychiatry Clin Neurosci 249[Suppl 4]: 2–13
Bremner JD, Narayan M, Anderson ER, Staib LH, Miller HL, Charney DS (2000) Hippocampalvolume reduction in major depression. Am J Psychiatry 157(1): 115–118
Cameron HA, Woolley CS, McEwen BS, Gould E (1993) Differentiation of newly born neuronsand glia in the dentate gyrus of the adult rat. Neuroscience 56(2): 337–344
Cameron HA, Tanapat P, Gould E (1998) Adrenal steroids and N-methyl-D-aspartate receptoractivation regulate neurogenesis in the dentate gyrus of adult rats through a commonpathway. Neuroscience 82(2): 349–354
Chakos MH, Lieberman JA, Alvir J, Bilder R, Ashtari M (1995) Caudate nuclei volumes inschizophrenic patients treated with typical antipsychotics or clozapine. Lancet 345(8947):456–457
Chakos MH, Alvir JM, Woerner MG, Koreen A, Geisler S, Mayerhoff D, Sobel S, Kane JM,Borenstein M, Lieberman JA (1996) Incidence and correlates of tardive dyskinesia in firstepisode of schizophrenia. Arch Gen Psychiatry 53(4): 313–319
98 A. Schmitt et al.

Conover JC, Allen RL (2002) The subventricular zone: new molecular and cellular develop-ments. Cell Mol Life Sci 59(12): 2128–2135
Coyle JT, Duman RS (2003) Finding the intracellular signaling pathways affected by mooddisorder treatments. Neuron 38(2): 157–160
Czeh B, Welt T, Fischer AK, Erhardt A, Schmitt W, Muller MB, Toschi N, Fuchs E, Keck ME(2002) Chronic psychosocial stress and concomitant repetitive transcranial magnetic stimu-lation: effects on stress hormone levels and adult hippocampal neurogenesis. Biol Psychiatry52(11): 1057–1065
Dawirs RR, Hildebrandt K, Teuchert-Noodt G (1998) Adult treatment with haloperidol increasesdentate granule cell proliferation in the gerbil hippocampus. J Neural Transm 105(2–3): 317–327
Eriksson PS, Perfilieva E, Bjork-Eriksson T, Alborn AM, Nordborg C, Peterson DA, Gage FH(1998) Neurogenesis in the adult human hippocampus. Nat Med 4(11): 1313–1317
Eyles DW, Avent KM, Stedman TJ, Pond SM (1997) Two pyridinium metabolites of haloperidolare present in the brain of patients at post-mortem. Life Sci 60(8): 529–534
Fitzgerald LW, Deutch AY, Gasic G, Heinemann SF, Nestler EJ (1995) Regulation of cortical andsubcortical glutamate receptor subunit expression by antipsychotic drugs. J Neurosci 15(3 Pt2): 2453–2461
Gould E (1999) Serotonin and hippocampal neurogenesis. Neuropsychopharmacology 21[2Suppl]: 46S–51S
Gould E, Tanapat P (1999) Stress and hippocampal neurogenesis. Biol Psychiatry 46(11):1472–1479
Gould E, McEwen BS, Tanapat P, Galea LA, Fuchs E (1997) Neurogenesis in the dentate gyrusof the adult tree shrew is regulated by psychosocial stress and NMDA receptor activation.J Neurosci 17(7): 2492–2498
Gratzner HG (1982) Monoclonal antibody to 5-bromo- and 5-iododeoxyuridine: a new reagentfor detection of DNA replication. Science 218(4571): 474–475
Gundersen HJ, Bendtsen TF, Korbo L, Marcussen N, Moller A, Nielsen K, Nyengaard JR,Pakkenberg B, Sorensen FB, Vesterby A et al. (1988) Some new, simple and efficientstereological methods and their use in pathological research and diagnosis. Apmis 96(5):379–394
Hildebrandt K, Teuchert-Noodt G, Dawirs RR (1999) A single neonatal dose of methamphet-amine suppresses dentate granule cell proliferation in adult gerbils which is restored tocontrol values by acute doses of haloperidol. J Neural Transm 106(5–6): 549–558
Hsu SM, Raine L, Fanger H (1981) Use of avidin-biotin-peroxidase complex (ABC) inimmunoperoxidase techniques: a comparison between ABC and unlabeled antibody(PAP) procedures. J Histochem Cytochem 29(4): 577–580
Jibiki I, Wakita S, Kubota T, Kurokawa K, Fukushima T, Yamaguchi N (1993) Haloperidol-induced blockade of induction of long-term potentiation in perforant path-dentate gyruspathway in chronically prepared rabbits. Pharmacol Biochem Behav 46(4): 847–852
Kempermann G, Kuhn HG, Gage FH (1997) More hippocampal neurons in adult mice living inan enriched environment. Nature 386(6624): 493–495
Kempermann G, Gast D, Kronenberg G, Yamaguchi M, Gage FH (2003) Early determination andlong-term persistence of adult-generated new neurons in the hippocampus of mice. Devel-opment 130(2): 391–399
Konradi C, Heckers S (2001) Antipsychotic drugs and neuroplasticity: insights into the treatmentand neurobiology of schizophrenia. Biol Psychiatry 50(10): 729–742
Krugers HJ, Koolhaas JM, Bohus B, Korf J (1993) A single social stress-experience altersglutamate receptor-binding in rat hippocampal CA3 area. Neurosci Lett 154(1–2): 73–77
Malberg JE, Eisch AJ, Nestler EJ, Duman RS (2000) Chronic antidepressant treatment increasesneurogenesis in adult rat hippocampus. J Neurosci 20(24): 9104–9110
McCarley RW, Wible CG, Frumin M, Hirayasu Y, Levitt JJ, Fischer MA, Shenton ME (1999)MRI anatomy of schizophrenia. Biol Psychiatry 45(9): 1099–1119
Meshul CK, Stallbaumer RK, Taylor B, Janowsky A (1994) Haloperidol-induced morphologicalchanges in striatum are associated with glutamate synapses. Brain Res 648(2): 181–195
Antipsychotics and cell proliferation 99

Meshul CK, Bunker GL, Mason JN, Allen C, Janowsky A (1996) Effects of subchronic clozapineand haloperidol on striatal glutamatergic synapses. J Neurochem 67(5): 1965–1973
Moghaddam B, Bolinao ML, Stein-Behrens B, Sapolsky R (1994) Glucocorticoids mediate thestress-induced extracellular accumulation of glutamate. Brain Res 655(1–2): 251–254
Nunes MC, Roy NS, Keyoung HM, Goodman RR, McKhann G 2nd, Jiang L, Kang J, NedergaardM, Goldman SA (2003) Identification and isolation of multipotential neural progenitor cellsfrom the subcortical white matter of the adult human brain. Nat Med 9(4): 439–447
Paxinos G, Watson C (1986) The rat brain in stereotaxic coordinates, 2nd edn. Academic Press,San Diego
Pegues MP, Rogers LJ, Amend D, Vinogradov S, Deicken RF (2003) Anterior hippocampalvolume reduction in male patients with schizophrenia. Schizophr Res 60(2–3): 105–115
Potkin SG, Fleming K, Jin Y, Gulasekaram B (2001) Clozapine enhances neurocognition andclinical symptomatology more than standard neuroleptics. J Clin Psychopharmacol 21(5):479–483
Schulze K, McDonald C, Frangou S, Sham P, Grech A, Toulopoulou T, Walshe M, Sharma T,Sigmundsson T, Taylor M, Murray RM (2003) Hippocampal volume in familial andnonfamilial schizophrenic probands and their unaffected relatives. Biol Psychiatry 53(7):562–570
See RE, Ellison G (1990) Comparison of chronic administration of haloperidol and the atypicalneuroleptics, clozapine and raclopride, in an animal model of tardive dyskinesia. Eur JPharmacol 181(3): 175–186
Selemon LD, Lidow MS, Goldman-Rakic PS (1999) Increased volume and glial density inprimate prefrontal cortex associated with chronic antipsychotic drug exposure. Biol Psy-chiatry 46(2): 161–172
Shors TJ, Townsend DA, Zhao M, Kozorovitskiy Y, Gould E (2002) Neurogenesis may relate tosome but not all types of hippocampal-dependent learning. Hippocampus 12(5): 578–584
Tanapat P, Galea LA, Gould E (1998) Stress inhibits the proliferation of granule cell precursors inthe developing dentate gyrus. Int J Dev Neurosci 16(3–4): 235–239
Terry AV Jr, Hill WD, Parikh V, Evans DR, Waller JL, Mahadik SP (2002) Differential effects ofchronic haloperidol and olanzapine exposure on brain cholinergic markers and spatiallearning in rats. Psychopharmacology (Berl) 164(4): 360–368
Varastet M, Riche D, Maziere M, Hantraye P (1994) Chronic MPTP treatment reproduces inbaboons the differential vulnerability of mesencephalic dopaminergic neurons observed inParkinson’s disease. Neuroscience 63(1): 47–56
Wakade CG, Mahadik SP, Waller JL, Chiu FC (2002) Atypical neuroleptics stimulate neurogen-esis in adult rat brain. J Neurosci Res 69(1): 72–79
West MJ (1999) Stereological methods for estimating the total number of neurons and synapses:issues of precision and bias. Trends Neurosci 22(2): 51–61
Williams RW, Rakic P (1988) Three-dimensional counting: an accurate and direct method toestimate numbers of cells in sectioned material. J Comp Neurol 278(3): 344–352
Yamamoto BK, Davy S (1992) Dopaminergic modulation of glutamate release in striatum asmeasured by microdialysis. J Neurochem 58(5): 1736–1742
Authors’ address: Dr. med. A. Schmitt, Central Institute of Mental Health, P.O. Box 12 21 20,D-68072 Mannheim, Germany, e-mail: [email protected]
100 A. Schmitt et al.: Antipsychotics and cell proliferation
Recommended

















