
R E S EA RCH AR T I C L E
Immune stimulating and therapeutic potential oftuftsin-incorporated nystatin liposomes against Cryptococcus
neoformans in leukopenic BALB/C mice
Masood Alam Khan1,2, Ahmed Aljarbou2, Arif Khan1,2 & Mohammad Owais1
1Interdisciplinary Biotechnology Unit, Aligarh Muslim University, Aligarh, India; and 2Department of Pharmaceutics, College of Pharmacy, Qassim
University, Buraidah, Saudi Arabia
Correspondence: Mohammad Owais,
Interdisciplinary Biotechnology Unit,
Aligarh Muslim University, Aligarh-202002,
India. Tel.: + 91 571 2720388;
fax: + 91 571 2721776;
e-mail: [email protected]
Received 28 November 2011; revised 14
March 2012; accepted 13 May 2012.
Final version published online 11 July 2012.
DOI: 10.1111/j.1574-695X.2012.00992.x
Editor: Richard Marconi
Keywords
cryptococcal meningitis; drug delivery;
immunomodulator; nystatin.
Abstract
Cryptococcus neoformans infection is a common fungal infection in persons
infected with human immune deficiency virus (HIV) or those with defective
cell-mediated immunity. Since treatment of cryptococcal meningitis poses a big
challenge, the present study aimed to develop a novel liposomal therapeutic
formulation against cryptococcosis. Treatment with tuftsin-incorporated lipo-
somes increased the anti-cryptococcal activity of murine peritoneal macro-
phages. Prophylactic treatment of mice with tuftsin-incorporated liposomes
reduced the dissemination of C. neoformans to brain tissues. Moreover, the
co-administration of tuftsin with nystatin liposomes augmented the anti-
cryptococcal activity of nystatin, as mice treated with tuftsin-incorporated
nystatin liposomes showed the highest survival and least fungal burden in
their brain tissues. The results of the present study favour the use of immune-
stimulating molecules along with antifungal agents in the treatment of oppor-
tunistic fungal infections.
Introduction
The opportunistic fungal pathogens such as Candida,
Aspergillus, Cryptococcus and Histoplasma spp. occur most
commonly in patients with impaired body immunity
(Ruhnke, 2004). The emergence of HIV infections, the
use of anti-cancer chemotherapy and intra-vascular
devices, and the persistent use of antibiotics and immu-
nosuppressive drugs for organ transplantation have
increased the frequency of fungal infections (Vilchez
et al., 2002; Kontoyiannis et al., 2003; Olszewski et al.,
2010). Most fungal pathogens cause diseases in immuno-
compromised subjects, but Cryptococcus neoformans can
cause infection in both immunocompetent and immuno-
compromised individuals (Rude et al., 2002). Cryptococcus
neoformans adopts unique virulence factors and strategies
to evade the host immune mechanisms (Olszewski et al.,
2010; Voelz & May, 2010).
Cryptococcus neoformans causes mild to severe pneu-
monia-like disease depending upon the virulence of the
isolate as well as on the status of the host immune sys-
tem (Mitchell & Perfect, 1995). Host resistance to
C. neoformans is largely dependent on the active partici-
pation of the innate and adaptive immune responses
(Shoham & Levitz, 2005; Zhou & Murphy, 2006). Anti-
bodies are less effective in elimination of C. neoformans,
but they function as opsonins in antibody-dependent
cellular cytotoxicity (Casadevall & Pirofski, 2005). The
cell-mediated immune response has been shown to play
a crucial role in controlling Cryptococcus infection (Sho-
ham & Levitz, 2005; Olszewski et al., 2010; Voelz &
May, 2010).
Liposomal formulations of polyene antibiotics such as
amphotericin B and nystatin have been proved to be the
most effective agents in the treatment of various fungal
infections (Nq et al., 2003). However, the toxicities
caused by polyene antibiotics limit their use in a clinical
setting. Liposomal formulations minimize the toxic mani-
festations associated with these drugs and are widely used
in the treatment of deep-seated fungal infections
(Dupont, 2002; Nasti et al., 2006). Moreover, liposomal
nystatin was an effective treatment against aspergillosis in
ª 2012 Federation of European Microbiological Societies FEMS Immunol Med Microbiol 66 (2012) 88–97Published by Blackwell Publishing Ltd. All rights reserved
IMM
UN
OLO
GY
& M
EDIC
AL
MIC
ROBI
OLO
GY

patients not responding to or intolerant of amphotericin
B therapy (Offner et al., 2004).
Tuftsin is a tetrapeptide (Thr-Lys-Pro-Arg) fraction of
the immunoglobulin G (IgG) molecule (Siemion &
Kluczyk, 1999). It binds to neutrophils and macrophages
to stimulate their phagocytic activity (Siemion & Kluc-
zyk, 1999) and also increases tumor necrosis factor
alpha (TNF-a) release from human Kupffer cells (Siem-
ion & Kluczyk, 1999). Tuftsin-macrophage interaction
results in activation of macrophages expressing nitric
oxide (NO) synthase to produce NO. Murine peritoneal
macrophages activated by tuftsin were able to kill the
intracellular protozoan Leishmania major (Siemion &
Kluczyk, 1999).
The use of immunomodulator-based therapy could be
of great significance in the successful treatment of infec-
tious diseases caused by opportunistic fungal pathogens
(Casadevall & Pirofski, 2001; Romani, 2001; Hamad,
2008). The immune-stimulatory agents not only help to
restore the normal immune response in immunocompro-
mised hosts, but also increase the efficacy of anti-
microbial agents (Kullberg et al., 2004; Matthews &
Burnie, 2004). Because of the presence of its receptors on
macrophages, liposomized-tuftsin facilitates the targeting
of drugs to macrophages, which helps eliminate the intra-
cellular infections (Gupta & Haq, 2005). The use of tuft-
sin has been shown to increase the efficacy of antibiotics
against protozoan, bacterial and fungal infections (Agar-
wal et al., 1994; Agrawal et al., 2002; Khan et al., 2003).
Tuftsin-incorporated nystatin liposomes have been shown
to be more effective, safer and more stable than conven-
tional nystatin liposomes without tuftsin (Khan et al.,
2006). In the present study, we showed that liposomal
tuftsin reduces the dissemination of C. neoformans in the
brain tissues of leukopenic mice.
Materials and methods
Mice
BALB/C mice used in this study were purchased from
National JALMA Research Institute for Leprosy, Agra,
India. The techniques used for bleeding, injection and
sacrifice of animals were approved by the Animal Ethics
Committee [Committee for the Purpose of Control and
Supervision of Experiments on Animals (CPCSEA), Gov-
ernment of India].
Materials
Nystatin was purchased from Sigma Chemical Co.
(St. Louis, MO). Cholesterol was bought from Centron
Research Laboratory (Bombay, India) and used after
crystallization with methanol. Egg phosphatidylcholine
(egg PC) was isolated and purified as described earlier
(Singleton et al., 1965). Tuftsin modified at the C-termi-
nus was prepared as described earlier (Gupta & Haq,
2005).
Test strain
The clinical isolate of C. neoformans was obtained from
the Department of Microbiology, J. N. Medical College,
A.M.U. Aligarh, India, as described earlier (Nasti et al.,
2006). The isolate was subcultured on Sabouraud Dex-
trose Agar at 37 °C for 48–72 h.
Antifungal susceptibility testing
The minimum inhibitory concentration (MIC) of nystatin
was determined by the broth macrodilution method as
described by the National Committee for Clinical Labora-
tory Standards recommendations (NCCLS, 1997). Nystatin
was tested over a concentration range of 0.02–5 lg mL�1
as described earlier (Nasti et al., 2006) Testing was
performed in 96-well round-bottom microtitre plates. A
cell suspension of C. neoformans was prepared in RPMI-
1640 medium and adjusted to give a final inoculum of
2 9 103 cells mL�1. The wells containing fungal inoculum
with different concentrations of the nystatin and proper
controls were incubated for 48 h.
Quantitative analysis of leukocytes
The number of leukocytes was determined by total leuko-
cyte count (TLC). The cells were counted on day 3 after
cyclophosphamide treatment in both control and tuftsin-
treated mice. Blood samples (20 lL) were taken from
mice and mixed with Turk’s fluid (380 lL) and the total
number of leukocytes was counted using a Neubaur
chamber (Brown, 1980).
Preparation of tuftsin-incorporated and
tuftsin-free nystatin liposomes
Liposomes were prepared from egg PC (49 lmol) and
cholesterol (21 lmol) with or without modified tuftsin
(7–8% by PC weight) by the sonication method (Owais
& Gupta, 2000). All the ingredients including nystatin
(Drug: Lipid, 1 : 20 molar ratio) were dissolved in a
round-bottomed flask in a mixture of chloroform/metha-
nol (1: 1, v/v). The solvents were evaporated under
reduced pressure to form a thin film of lipids and the
traces of the solvents were removed by subjecting the
flask to vacuum. Subsequently, the lipid film (consisting
of egg PC/cholesterol, nystatin) was hydrated with 2.0 mL
FEMS Immunol Med Microbiol 66 (2012) 88–97 ª 2012 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
Tuftsin liposomes control murine cryptococcosis 89

of sterile normal saline with intermittent stirring followed
by sonication (1 h, 4 °C) in a bath type sonicator under
N2 atmosphere. After sonication, the lipid formulations
of drug were centrifuged at 10 000 g for 1 h at 4 °C to
remove traces of undispersed lipids, and finally dialyzed
against normal saline for 24 h at 4 °C in the dark.
Determination of size, lamellerity and
structure of liposomes
The liposome lamellerity, size and structure were evalu-
ated by transmission electron microscopy as described
earlier (Khan et al., 2006).
Estimation of liposome-incorporated nystatin
and tuftsin
Nystatin incorporation in liposomes was determined
spectrophotometrically and by high performance liquid
chromatography (HPLC) as described earlier (Khan et al.,
2006). A standard curve of nystatin was plotted at
320 nm. Nystatin associated with liposomes was esti-
mated by dissolving the liposomal formulation in metha-
nol and determining the absorbance at 320 nm using an
equal amount of lipid in methanol as a blank. The
amount of nystatin entrapped in liposomes was calculated
from the standard curve of the drug. The intercalation
efficiency of nystatin in liposomes was also estimated by
the HPLC method. Briefly, the sample (20 lL) was
injected in a Hypersil octyldecyl-silane 5 lm particle size
analytical column (150 9 4.6 mm internal diameter).
A standard curve was plotted by calibrating peak area
vs. amount of the drug injected in the column. The
intercalation efficiency of nystatin in both plain egg PC
and tuftsin-bearing liposomes was found out to be of the
same order as estimated by spectrophotometrically
(90 ± 4%).
To facilitate the incorporation of tuftsin into liposome
bilayers, tuftsin was modified at its C-terminus by attach-
ing a long fatty acyl residue through an ethylenediamine
spacer arm [Thr-Lys-Pro-Arg-NH-(CH2)2-NH-COC15H31]
according to the published procedure (Singhal et al.,
1984). The tuftsin incorporated in nystatin liposomes was
estimated by the bicinchoninic acid (BCA) method.
Briefly, the liposomes (given volume) were lysed with 10%
Triton X-100 solution (the final concentration of Triton
X-100 was maintained at 1%). The mixture of solutions A
(containing sodium carbonate, sodium bicarbonate,
BCA and sodium tartrate in 0.1 M sodium hydroxide)
and B (4% cupric sulphate) of BCA reagent was added
to an aliquot of tuftsin-incorporated liposomes digested
with Triton X-100. The reaction mixture was incubated
at 37 °C for 45 min. The absorbance was measured at
600 nm and the content of tuftsin incorporated in lipo-
somes was determined using a standard curve of tuftsin
plotted in the presence of Triton X-100. The percent
intercalation efficiency of the incorporated tuftsin was
found to be about 95%.
Preparation of C. neoformans cells for
infection
Yeast cells of C. neoformans were harvested from agar
plates into YPD (1% yeast extract, 2% peptone, 5% dex-
trose) medium at 37 °C for 48 h as described earlier (Nasti
et al., 2006). The cells were washed with sterile normal
saline at low speed centrifugation (670 g) and diluted to
the appropriate concentrations in saline prior to use. Each
mouse was infected intravenously with 1 9 105 cells of
C. neoformans to determine the kinetics of cryptococcal
meningitis in mice. In survival studies, each mouse was
infected with 7 9 105 cells of C. neoformans.
Assessment of anticryptococcal activity of
liposomal tuftsin-treated macrophages
Macrophages (1 9 105 cells per well) were seeded in trip-
licates in 24-well costar plates containing complete med-
ium supplemented with 10% fetal bovine serum as
described earlier (Khan et al., 2003). The plates were
incubated at 37 °C in 5% CO2 for 24 h. Medium was
taken out and cells were washed. The macrophages were
then treated with sham liposomes or tuftsin-bearing lipo-
somes (the concentration of tuftsin at 1 lg mL�1) for
4 h. Cells were washed and C. neoformans (5 9 104
cells per well) was added to tuftsin-treated or untreated
macrophages and incubated for 2 h. After 2 h of infec-
tion, non-phagocytosed C. neoformans cells were rinsed
with cold phosphate-buffered saline (PBS). The number
of colony forming units (CFUs) recovered from the lysis
of macrophages (no tuftsin treatment) after 2 h of infec-
tion was considered baseline. After 24 h of incubation,
macrophages were lysed using 0.1% Tween-20 to deter-
mine the numbers of phagocytosed C. neoformans cells.
The various dilutions of lysates were spread on SD agar
plates and incubated at 37 °C for 48 h.
Induction of leukopenia in mice
Mice were injected intravenously with a single dose of
cyclophosphamide (250 mg kg�1) to induce leukopenia.
Leukopenia temporarily persisted for a week and mice
were infected on day 3 post-cyclophosphamide injection.
Day 0 was the day of infection of mice with C. neofor-
mans. In prophylactic studies, pretreatment with tuftsin
(50 lg per mouse, intraperitoneal) was started 8 h after
ª 2012 Federation of European Microbiological Societies FEMS Immunol Med Microbiol 66 (2012) 88–97Published by Blackwell Publishing Ltd. All rights reserved
90 M.A. Khan et al.

cyclophosphamide treatment for three consecutive days
(day �3 to day �1).
Kinetics of the dissemination of C. neoformans
in the brain of immunocompetent and
leukopenic mice pretreated with sham
liposomes or tuftsin-incorporated liposomes
The prophylactic role of immunomodulator tuftsin
against C. neoformans was investigated in immunocompe-
tent and leukopenic mice. Mice were treated with lipos-
omized-tuftsin (50 lg per mouse) intraperitoneally. Each
mouse was infected with C. neoformans (1 9 105 cells per
mouse in a volume of 100 lL sterile saline) by the intra-
venous route on day 3 post-cyclophosphamide treatment.
Brain tissues were collected aseptically from infected mice
at 3, 6, 12, 24 and 48 h post-C. neoformans infection. The
organs were washed with sterile PBS and homogenized in
sterile PBS. Appropriate dilutions of homogenates were
plated (100 lL) in triplicate on chloramphenicol contain-
ing Sabouraud-dextrose agar plates and CFU were
counted after 48 h of growth at 37 °C. Chloramphenicol
was used to prevent any bacterial contamination.
Treatment with tuftsin-incorporated or
tuftsin-free nystatin liposomes
Immunocompetent and leukopenic mice were administered
tuftsin-incorporated or tuftsin-free nystatin liposomes
intraperitoneally on days 1, 3 and 5 post-Cryptococcus
infection. Each treatment group contained 10 mice, divided
into the following groups: (1) saline, (2) sham liposomes, (3)
tuftsin liposomes, (4) liposomal nystatin (3 and 5 mg kg�1)
and (5) tuftsin-incorporated liposomal nystatin (3 and
5 mg kg�1).
Prophylactic use of liposomal tuftsin followed
by treatment with tuftsin-free or tuftsin-
incorporated nystatin liposomes against
C. neoformans in leukopenic mice
Since treatment with tuftsin-free or tuftsin-incorporated
nystatin liposomes did not completely cure C. neoformans-
infected leukopenic mice, the prophylactic effect on the
efficacy of liposomal nystatin in leukopenic mice of tuft-
sin was determined. Each mouse was treated with 50 lgof liposomized-tuftsin intraperitoneally for three con-
secutive days as described earlier (Khan et al., 2003).
Pretreatment with tuftsin liposomes (PT-tuftsin-liposomes)
in leukopenic mice was started 8 h after cyclophospha-
mide injection. After 3 days of tuftsin treatment, each
mouse was infected with 5 9 105 cells of C. neoformans.
Mice were divided into the following groups: (1) saline,
(2) sham liposomes, (2) PT-tuftsin-liposomes + sham
liposomes, (4) tuftsin liposomes, (5) PT-tuftsin-liposomes +tuftsin liposomes, (6) liposomal nystatin (5 mg kg�1), (7)
PT-tuftsin-liposomes + liposomal nystatin (5 mg kg�1),
(8) tuftsin-bearing liposomal nystatin (5 mg kg�1), (9) PT-
tuftsin-liposomes + tuftsin-liposomal nystatin (5 mg kg�1).
Quantitative analysis of C. neoformans in the
brain tissues
The prophylactic and therapeutic role of tuftsin was evalu-
ated by determining the fungal load in brain tissues. Three
mice from each group were sacrificed on day 3 post-
C. neoformans infection (8 h after the 2nd dose of treat-
ment) and their brains were taken out aseptically as
described earlier (Nasti et al., 2006). Briefly, weighed
portions of the brain tissues were homogenized in 5 mL of
sterile normal saline and different dilutions of the suspen-
sion were plated on SD agar plates containing chloramphe-
nicol. The plates were incubated at 37 °C for 48–72 h. The
numbers of colonies were counted and the fungal load was
determined by multiplying them by the dilution factor.
Statistical analyses
Analysis of the survival of mice was performed using
Kaplan–Meier curve, and various groups were compared
by log-rank test. Fungal burden in organs was analyzed
by one-way ANOVA followed by the Bonferroni post hoc
test using GRAPHPAD PRISM software version 3.0.
Results
In vitro antifungal susceptibility testing
The minimum inhibitory concentration (MIC) was defined
as the lowest concentration of nystatin at which there was
complete inhibition of the fungal growth. The MIC of
nystatin for C. neoformans was found to be 1.5 lg mL�1.
Tuftsin facilitates the early recovery of the
depleted leukocytes in cyclophosphamide-
treated mice
Immuno-potential effect of tuftsin was analyzed by treating
leukopenic mice with tuftsin-bearing liposomes, and mice
treated with saline or sham liposomes acted as controls.
The number of leukocytes was counted in the blood of
tuftsin-treated or untreated mice. Our results demonstrated
that treatment with tuftsin liposomes induced early recov-
ery of leukocytes in leukopenic mice. We found that leuko-
penic mice treated with tuftsin-liposomes have higher
counts of leukocytes (1220–1956) on day 4 post-cyclophos-
FEMS Immunol Med Microbiol 66 (2012) 88–97 ª 2012 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
Tuftsin liposomes control murine cryptococcosis 91

phamide injection compared with mice treated with saline
(166–272) or sham liposomes (183–344) (P < 0.001).
Moreover, the group of leukopenic mice treated with
tuftsin-incorporated liposomes showed complete recovery
of leukocytes on day 8 post-cyclophosphamide treatment,
whereas saline- or sham liposome-treated mice showed
only 40–60% recovery in leukocyte counts.
Tuftsin-treated macrophages show more
resistance against C. neoformans
Cryptococcus neoformans can multiply inside macrophages
as shown by an increase in the numbers of CFUs after
24 h of infection (Fig. 1). Tuftsin binds to macrophages
and activates them to kill the phagocytosed C. neoformans.
Liposomized-tuftsin showed an increased effect on macro-
phage-mediated killing of intracellular pathogens, as tuft-
sin-incorporated liposomes were taken up rapidly by
macrophages compared with similar liposomes without
tuftsin. Macrophages treated with tuftsin-incorporated
liposomes showed increased anti-cryptococcal activity
compared with tuftsin-free liposomes.
Treatment with tuftsin-liposomes reduces the
dissemination of C. neoformans to brain
tissues of immunocompetent and
leukopenic mice
The kinetics of C. neoformans infection to the brain
tissues was determined in immunocompetent and
leukopenic mice prophylactically treated or untreated
with tuftsin-incorporated liposomes. Mice were eutha-
nized and brain tissues were taken at 3, 6, 12, 24 and
48 h post-C. neoformans infection. There was a reduced
C. neoformans burden in the brain tissues of mice treated
with tuftsin-incorporated liposomes compared with mice
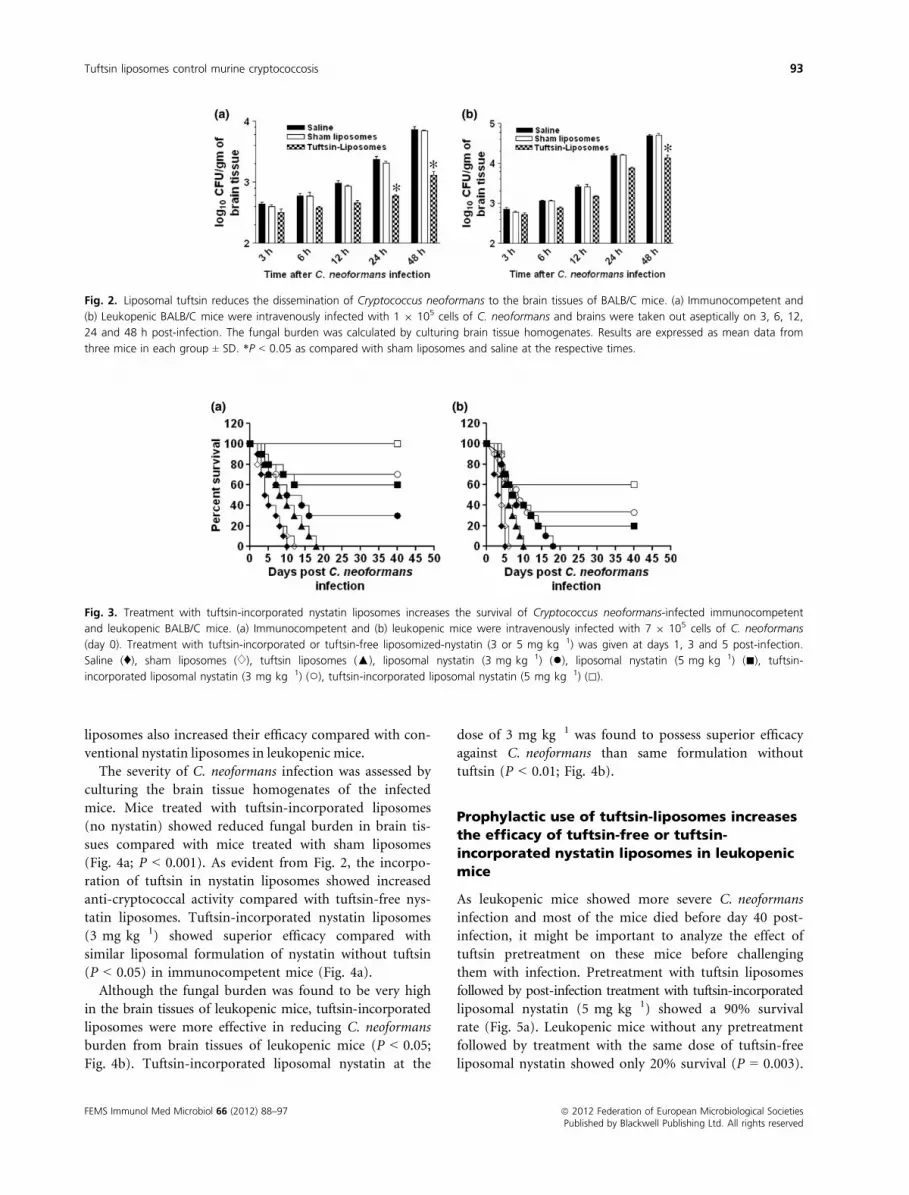
treated with sham liposomes or saline (Fig. 2a).
The C. neoformans infection was found to be more
severe in leukopenic mice compared with immunocom-
petent mice as shown by higher numbers of CFUs in the
brain tissues of leukopenic mice (Fig. 2b). Leukopenic
mice pretreated with tuftsin-incorporated liposomes also
showed reduced burden of C. neoformans in the brain
tissues, particularly at 24 and 48 h post-infection
(Fig. 2b). This suggests that activation of the immune
system by tuftsin liposomes plays an important role in
reducing the dissemination of C. neoformans to the brain
tissues.
Incorporation of tuftsin augments the activity
of liposomal nystatin against C. neoformans in
BALB/C mice
The tuftsin-incorporated liposomal nystatin at the dose
of 3 and 5 mg kg�1 on days 1, 3 and 5 post-infection
showed increased efficacy against systemic C. neoformans
in both immunocompetent and leukopenic mice. Immu-
nocompetent mice treated with tuftsin-incorporated nys-
tatin liposomes (5 mg kg�1) showed 100% survival rate
on day 40 post-infection, whereas the mice treated with
liposomized-nystatin (5 mg kg�1) without tuftsin showed
a 60% survival rate (P = 0.029; Fig. 3a). Mice treated with
tuftsin-incorporated liposomal nystatin at the dose of
3 mg kg�1 showed a 70% survival, whereas liposomized-
nystatin at the same dose showed only 30% survival.
Thus tuftsin-incorporated nystatin liposomes showed
superiority over tuftsin-free nystatin liposomes in treating
C. neoformans-infected mice.
Cryptococcus neoformans-infected leukopenic mice were
also treated with tuftsin-free or tuftsin-incorporated liposo-
mal nystatin on days 1, 3 and 5 post-infection. Like immu-
nocompetent mice, leukopenic mice showed an increased
survival upon treatment with tuftsin-incorporated nystatin
liposomes. The group of leukopenic mice treated with tuft-
sin-incorporated nystatin liposomes (5 mg kg�1) showed a
60% survival rate on day 40 post-C. neoformans challenge,
whereas mice treated with tuftsin-free nystatin liposomes at
the same dose showed only 20% survival (Fig. 3b). Mice
treated with tuftsin-incorporated nystatin liposomes at
3 mg kg�1 showed a 30% survival rate, whereas mice trea-
ted with tuftsin-free nystatin liposomes (3 mg kg�1) died
before day 40 post-infection (Fig. 3b). These results showed
that incorporation of tuftsin on the surface of nystatin
Fig. 1. Treatment of macrophages with tuftsin liposomes increases
the anti-cryptococcal activity of macrophages. Peritoneal macrophages
were untreated or treated with sham liposomes or tuftsin liposomes
for 2 h as described in Materials and methods. Macrophages were
infected with Cryptococcus neoformans for 24 h. Cells were lysed
and intracellular yeasts were plated on Sabouraud dextrose agar for
48 h at 37 °C. Medium vs. tuftsin liposomes (P < 0.05); Sham
liposomes vs. tuftsin liposomes (P < 0.05).
ª 2012 Federation of European Microbiological Societies FEMS Immunol Med Microbiol 66 (2012) 88–97Published by Blackwell Publishing Ltd. All rights reserved
92 M.A. Khan et al.
liposomes also increased their efficacy compared with con-
ventional nystatin liposomes in leukopenic mice.
The severity of C. neoformans infection was assessed by
culturing the brain tissue homogenates of the infected
mice. Mice treated with tuftsin-incorporated liposomes
(no nystatin) showed reduced fungal burden in brain tis-
sues compared with mice treated with sham liposomes
(Fig. 4a; P < 0.001). As evident from Fig. 2, the incorpo-
ration of tuftsin in nystatin liposomes showed increased
anti-cryptococcal activity compared with tuftsin-free nys-
tatin liposomes. Tuftsin-incorporated nystatin liposomes
(3 mg kg�1) showed superior efficacy compared with
similar liposomal formulation of nystatin without tuftsin
(P < 0.05) in immunocompetent mice (Fig. 4a).
Although the fungal burden was found to be very high
in the brain tissues of leukopenic mice, tuftsin-incorporated
liposomes were more effective in reducing C. neoformans
burden from brain tissues of leukopenic mice (P < 0.05;
Fig. 4b). Tuftsin-incorporated liposomal nystatin at the
dose of 3 mg kg�1 was found to possess superior efficacy
against C. neoformans than same formulation without
tuftsin (P < 0.01; Fig. 4b).
Prophylactic use of tuftsin-liposomes increases
the efficacy of tuftsin-free or tuftsin-
incorporated nystatin liposomes in leukopenic
mice
As leukopenic mice showed more severe C. neoformans
infection and most of the mice died before day 40 post-
infection, it might be important to analyze the effect of
tuftsin pretreatment on these mice before challenging
them with infection. Pretreatment with tuftsin liposomes
followed by post-infection treatment with tuftsin-incorporated
liposomal nystatin (5 mg kg�1) showed a 90% survival
rate (Fig. 5a). Leukopenic mice without any pretreatment
followed by treatment with the same dose of tuftsin-free
liposomal nystatin showed only 20% survival (P = 0.003).
Fig. 2. Liposomal tuftsin reduces the dissemination of Cryptococcus neoformans to the brain tissues of BALB/C mice. (a) Immunocompetent and
(b) Leukopenic BALB/C mice were intravenously infected with 1 9 105 cells of C. neoformans and brains were taken out aseptically on 3, 6, 12,
24 and 48 h post-infection. The fungal burden was calculated by culturing brain tissue homogenates. Results are expressed as mean data from
three mice in each group ± SD. *P < 0.05 as compared with sham liposomes and saline at the respective times.
Fig. 3. Treatment with tuftsin-incorporated nystatin liposomes increases the survival of Cryptococcus neoformans-infected immunocompetent
and leukopenic BALB/C mice. (a) Immunocompetent and (b) leukopenic mice were intravenously infected with 7 9 105 cells of C. neoformans
(day 0). Treatment with tuftsin-incorporated or tuftsin-free liposomized-nystatin (3 or 5 mg kg�1) was given at days 1, 3 and 5 post-infection.
Saline (♦), sham liposomes (♢), tuftsin liposomes (▲), liposomal nystatin (3 mg kg�1) (●), liposomal nystatin (5 mg kg�1) (■), tuftsin-
incorporated liposomal nystatin (3 mg kg�1) (○), tuftsin-incorporated liposomal nystatin (5 mg kg�1) (□).
FEMS Immunol Med Microbiol 66 (2012) 88–97 ª 2012 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
Tuftsin liposomes control murine cryptococcosis 93

Pretreatment with liposomal tuftsin also increases the sur-
vival of C. neoformans infected leukopenic mice from
20% to 60% after treatment with liposomal nystatin
(5 mg kg�1) (Fig. 5a).
The effect of tuftsin pretreatment on the therapeutic
efficacy of tuftsin-free or tuftsin-incorporated liposomal
nystatin was also assessed by culturing the brain tissue
homogenates. The fungal burden was found to be the
least in mice pretreated with tuftsin liposomes followed
by treatment with tuftsin-incorporated liposomal nystatin
(Fig. 5b). Tuftsin pretreated leukopenic mice showed
greater resistance to C. neoformans infection compared
with mice not pretreated with tuftsin (P < 0.01). The
fungal burden was much reduced in the brain tissues of
Fig. 4. Treatment with tuftsin-incorporated nystatin liposomes showed higher efficacy in reducing fungal burden from the brain tissues of
Cryptococcus neoformans-infected immunocompetent and leukopenic BALB/C mice. (a) Immunocompetent and (b) leukopenic mice were
infected with 7 9 105 cells of C. neoformans (day 0). Treatment with tuftsin-incorporated or tuftsin-free liposomized-nystatin (3 or 5 mg kg�1)
was given at days 1, 3 and 5 post-infection. On day 3, three mice from each group were sacrificed and their brains were taken out and
homogenized. The brain tissue homogenates were cultured to determine fungal load. For immunocompetent mice: sham liposomes vs. tuftsin
liposomes (P < 0.001); liposomal nystatin (3 mg kg�1) vs. tuftsin-incorporated liposomal nystatin (3 mg kg�1) (P < 0.05). For leukopenic mice:
sham liposomes vs tuftsin liposomes (P < 0.05); liposomal nystatin (3 mg kg�1) vs. tuftsin-incorporated liposomal nystatin (3 mg kg�1) (P < 0.01).
Fig. 5. Prophylactic use of tuftsin-incorporated liposomes increases the efficacy of liposomal nystatin against Cryptococcus neoformans in
leukopenic BALB/C mice. (a) Leukopenic mice were pretreated with liposomized-tuftsin (50 lg per mouse) by the intraperitoneal route for three
consecutive days. Mice were infected with 7 9 105 cells of C. neoformans (day 0). Treatment with tuftsin-incorporated or tuftsin-free
liposomized-nystatin (5 mg kg�1) was given at days 1, 3 and 5 post-infection. Mice were observed for 40 days to check their survival. Saline (9),
sham liposomes (□), PT-tuftsin + sham liposomes (■), tuftsin liposomes (D), PT-tuftsin + tuftsin liposomes (▲), liposomal nystatin (5 mg kg�1)
(∇), PT-tuftsin + liposomal nystatin (5 mg kg�1) (▼), tuftsin-incorporated liposomal nystatin (5 mg kg�1) (♢), PT-tuftsin + tuftsin-incorporated
liposomal nystatin (5 mg kg�1) (♦). (b) On day 3, three mice from each group were sacrificed and their brains were taken out and homogenized.
The brain tissue homogenates were cultured to determine fungal load. Liposomal nystatin vs. PT-tuftsin + liposomal nystatin (P < 0.01),
Liposomal nystatin vs. PT-tuftsin + tuftsin-incorporated liposomal nystatin (P < 0.001).
ª 2012 Federation of European Microbiological Societies FEMS Immunol Med Microbiol 66 (2012) 88–97Published by Blackwell Publishing Ltd. All rights reserved
94 M.A. Khan et al.

leukopenic mice pretreated with tuftsin liposomes fol-
lowed by therapy with tuftsin-incorporated liposomal
nystatin compared with mice treated with liposomal nys-
tatin without tuftsin (P < 0.01).
Residual fungal burden
To determine the clearance of infection, the residual
fungal burden in the brain tissues of the surviving mice
was analyzed on day 40 post-C. neoformans infection. It
fell in the range of 0–75 CFUs per brain, which suggested
that infection in these mice had subsided.
Discussion
Among the fungal pathogens, C. neoformans is one of the
leading causes of morbidity and mortality in immunologi-
cally compromised persons (Zhou & Murphy, 2006). The
severity of the infection and disease depends largely on the
competence of the host immune system, inoculum and the
virulence of the pathogen. Neutrophils and macrophages
play an important role in controlling C. neoformans infec-
tion (Romani, 2001). On the other hand, in order to estab-
lish itself, C. neoformans tries to modulate or subvert the
host immune system to establish the infection (Vecchiarelli,
2007).
Tuftsin is known to stimulate the phagocytic activity of
neutrophils and macrophages, which play a role in the
protection against fungal infections (Siemion & Kluczyk,
1999). Tuftsin was incorporated on the surface of lipo-
somes. The results of the present study show that treat-
ment with tuftsin-incorporated liposomes reduces the
dissemination of C. neoformans to brain tissues. There
was a reduced burden of C. neoformans in the brain
tissues of mice prophylactically treated with tuftsin lipo-
somes compared with mice treated with liposomes with-
out tuftsin (Fig. 1a). Similar results were found in
cyclophosphamide-treated mice, although the severity of
C. neoformans infection was greater in them (Fig. 1b). As
tuftsin activates the phagocytic activities of neutrophils
and macrophages (Siemion & Kluczyk, 1999), it is quite
possible that neutrophils and macrophages control the
dissemination of C. neoformans to brain tissues of tuftsin-
pretreated mice.
The use of immunomodulators in combination with
chemotherapy imparts greater protection against fungal
infections (Dutta, 2002; Khan et al., 2003). Immunomod-
ulator tuftsin stimulates the respiratory burst phenome-
non in macrophages and neutrophils by producing free
nitrogen and oxygen species (Siemion & Kluczyk, 1999).
Tuftsin-incorporated liposomal nystatin at a dose of
5 mg kg�1 was found to produce a 100% cure rate
against systemic lethal infection of C. neoformans in
immunocompetent mice, whereas C. neoformans-infected
mice treated with liposomal nystatin (without tuftsin) at
the same dose showed only 60% survival. The fungal bur-
den also supported the survival data, as C. neoformans-
infected mice treated with tuftsin-incorporated nystatin
liposomes showed the fewer CFUs in brain tissues.
As opportunistic fungal infections are commonly found
in immunocompromised persons, mice were made leuko-
penic by injecting cyclophosphamide. Quantitative or qual-
itative defects in neutrophils are predisposing factors to
disseminated fungal infections. Neutrophils play a very
important role in limiting the early multiplication of the
fungal pathogens, and during the infection macrophages in
the tissues become activated and prevent the dissemination
of the infection. Leukopenic mice treated with tuftsin-
incorporated nystatin liposomes at the doses of 3 and
5 mg kg�1 showed a 30 and 60% survival rate, respectively,
on day 40 post-C. neoformans infection, whereas the group
of mice treated with nystatin liposomes (without tuftsin) at
the dose of 5 mg kg�1 showed only 20% survival. The
increased severity of C. neoformans infection may be due to
reduced numbers of leukocytes in cyclophosphamide-
injected mice. The results of the present study showed that
combination therapy of tuftsin with liposomal nystatin is
more effective in increasing the survival of C. neoformans-
infected leukopenic mice. The increased effect of tuftsin-
incorporated liposomal nystatin against C. neoformans in
leukopenic mice can be ascribed to the tuftsin-induced
activation of tissue macrophages and residual leukocytes.
Moreover, tuftsin-incorporated nystatin liposomes can
specifically target the drug to macrophages, which may act
as drug depots and reach the site of infection. Thus the
status of innate immune cells at the time of infection plays
a role in determining the anti-cryptococcal efficacy of lipo-
somal nystatin in tuftsin-treated or untreated leukopenic
mice. Tuftsin-incorporated nystatin liposomes stay in the
systemic circulation for a prolonged time, in comparison
with the tuftsin-free nystatin liposomes (Khan et al., 2006).
The present study shows the increased efficacy of liposomal
nystatin in leukopenic mice pretreated with tuftsin-incor-
porated liposomes. This may be due to the stimulating
effect of tuftsin in the recovery of leukocytes or activated
state of tissue macrophages or residual neutrophils in
leukopenic mice. Tuftsin-macrophage interaction results in
the activation of macrophages expressing NO synthase to
produce NO, which plays an important role in killing intra-
cellular C. neoformans (Ghosn et al., 2006). Inducible nitric
oxide synthase-deficient mice showed more susceptibility
to C. neoformans infection compared with control mice
(Aguirre & Gibson, 2000).
The results of the present study clearly demonstrate the
important role of tuftsin in augmenting the anti-crypto-
coccal activity of liposomal nystatin in murine model
FEMS Immunol Med Microbiol 66 (2012) 88–97 ª 2012 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
Tuftsin liposomes control murine cryptococcosis 95

because of its ability to specifically target nystatin to intra-
cellular residing sites of the pathogen in macrophages.
Tuftsin also has the ability simultaneously to activate neu-
trophils, monocytes and macrophages for increased phago-
cytosis and killing of pathogens by these cells. Furthermore,
tuftsin can also be used to rejuvenate the immune system
of the patients suffering from immune-deficiency diseases
or undergoing cancer chemotherapy.
Acknowledgements
We are very grateful to Aligarh Muslim University and
Interdisciplinary Biotechnology Unit (IBU) for providing
all the necessary facilities. We are also grateful to Prof.
Abida Malik, Department of Microbiology, JNMC,
Aligarh, for identification and characterization of the
Cryptococcus neoformans isolate. Masood A. Khan thanks
the University Grant Commission (UGC), Govt. of India
for awarding financial assistance in the form of research
fellowship. There is no conflict of interest among authors.
References
Agarwal A, Kandpal H, Gupta HP, Singh NB & Gupta CM
(1994) Tuftsin-bearing liposomes as rifampin vehicles in
treatment of tuberculosis in mice. Antimicrob Agents
Chemother 38: 588–593.Agrawal AK, Agrawal A, Pal A, Guru PY & Gupta CM (2002)
Superior chemotherapeutic efficacy of amphotericin B in
tuftsin-bearing liposomes against Leishmania donovani
infection in hamsters. J Drug Target 10: 41–45.Aguirre KM & Gibson GW (2000) Differing requirement for
inducible nitric oxide synthase activity in clearance of
primary and secondary Cryptococcus neoformans infection.
Med Mycol 38: 343–353.Brown BA (1980) Hematology: Principles and Procedures, 3rd
edn. Lea & Febiger, Philadelphia, PA.
Casadevall A & Pirofski LA (2001) Adjunctive immune therapy
for fungal infections. Clin Infect Dis 33: 1048–1056.Casadevall A & Pirofski L (2005) Insights into mechanisms of
antibody-mediated immunity from studies with Cryptococcus
neoformans. Curr Mol Med 5: 421–433.Dupont B (2002) Overview of lipid formulations of
amphotericin B. J Antimicrob Chemother 49(Suppl 1): 31–36.Dutta RC (2002) Peptide immunomodulators versus infection:
an analysis. Immunol Lett 83: 153–161.Ghosn EE, Russo M & Almeida SR (2006) Nitric oxide-
dependent killing of Cryptococcus neoformans by B-1-derived
mononuclear phagocyte. J Leukoc Biol 80: 36–44.Gupta CM & Haq W (2005) Tuftsin-bearing liposomes as
antibiotic carriers in treatment of macrophage infections.
Methods Enzymol 391: 291–304.Hamad M (2008) Antifungal immunotherapy and
immunomodulation: a double-hitter approach to deal with
invasive fungal infections. Scand J Immunol 67: 533–543.
Khan MA, Syed FM, Nasti HT, Saima K, Haque W, Shehbaz A
& Owais M (2003) Use of tuftsin bearing nystatin
liposomes against an isolate of Candida albicans showing
less in vivo susceptibility to amphotericin B. J Drug Target
11: 93–99.Khan MA, Faisal SM & Mohammed O (2006) Safety, efficacy
and pharmacokinetics of tuftsin-loaded nystatin liposomes
in murine model. J Drug Target 14: 233–241.Kontoyiannis DP, Mantadakis E & Samonis G (2003) Systemic
mycoses in the immunocompromised host: an update in
antifungal therapy. J Hosp Infect 53: 243–258.Kullberg BJ, Oude Lashof AM & Netea MG (2004) Design of
efficacy trials of cytokines in combination with antifungal
drugs. Clin Infect Dis 39: S218–S223.Matthews RC & Burnie JP (2004) Recombinant antibodies: a
natural partner in combinatorial antifungal therapy. Vaccine
22: 865–871.Mitchell TG & Perfect JR (1995) Cryptococcosis in the era of
AIDS – 100 years after the discovery of Cryptococcus
neoformans. Clin Microbiol Rev 8: 515–548.Nasti TH, Khan MA & Owais M (2006) Enhanced efficacy of pH-
sensitive nystatin liposomes against Cryptococcus neoformans
in murine model. J Antimicrob Chemother 57: 349–352.National Committee for Clinical Laboratory Standards (1997).
Reference Method for Broth Dilution Antifungal Susceptibility
Testing of Yeasts: Approved Standard M27-A. NCCLS,
Villanova, PA.
Nq AW, Wasan KM & Lopez-Berestein G (2003) Development
of liposomal polyene antibiotics: a historical perspective.
J Pharm Pharm Sci 6: 67–83.Offner F, Krcmery V, Boogaerts M et al. (2004) Liposomal
nystatin in patients with invasive aspergillosis refractory to
or intolerant of amphotericin B. Antimicrob Agents
Chemother 48: 4808–4812.Olszewski MA, Zhang Y & Huffnagle GB (2010) Mechanisms
of Cryptococcal virulence and persistence. Future Microbiol
5: 1269–1288.Owais M & Gupta CM (2000) Liposome mediated cytosolic
delivery of macromolecules and its possible use in vaccine
development. Eur J Biochem 267: 3946–3956.Romani L (2001) Host immune reactivity and antifungal
chemotherapy: the power of being together. J Chemother 13:
347–353.Rude TH, Toffaletti DL, Cox GM & Perfect JR (2002)
Relationship of the glyoxylate pathway to the
pathogenesis of Cryptococcus neoformans. Infect Immun 70:
5684–5694.Ruhnke M (2004) Mucosal and systemic fungal infections in
patients with AIDS: prophylaxis and treatment. Drugs 64:
1163–1180.Shoham S & Levitz SM (2005) The immune response to fungal
infections. Br J Haematol 129: 569–582.Siemion IZ & Kluczyk A (1999) Tuftsin: on the 30-year
anniversary of Victor Najjar’s discovery. Peptides 20: 645–674.Singhal A, Bali A, Jain RK & Gupta CM (1984) Specific
interactions of liposomes with PMN leukocytes upon
ª 2012 Federation of European Microbiological Societies FEMS Immunol Med Microbiol 66 (2012) 88–97Published by Blackwell Publishing Ltd. All rights reserved
96 M.A. Khan et al.

incorporating tuftsin in their bilayers. FEBS Lett 178:
109–113.Singleton WS, Gray MS & Brown ML (1965) A method for
adsorbent fractionation of cottonseed oil for experimental
intravenous fat emulsions. J Am Oil Chem Soc 42:
53–56.Vecchiarelli A (2007) Fungal capsular polysaccharide and
T-cell suppression: the hidden nature of poor
immunogenicity. Crit Rev Immunol 27: 547–557.
Vilchez RA, Fung J & Kusne S (2002) Cryptococcosis in organ
transplant recipients: an overview. Am J Transplant 2:
575–580.Voelz K & May RC (2010) Cryptococcal interactions with the
host immune system. Eukaryot Cell 9: 835–846.Zhou Q & Murphy WJ (2006) Immune response and immune
therapy to Cryptococcus infections. Immunol Res 35:
191–208.
FEMS Immunol Med Microbiol 66 (2012) 88–97 ª 2012 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
Tuftsin liposomes control murine cryptococcosis 97
Recommended

















