
RED CELLS, IRON, AND ERYTHROPOIESIS
Ligation of complement receptor 1 increases erythrocyte membrane deformability*Aleksandra M. Glodek,1 *Rossen Mirchev,2 David E. Golan,2,3 Joseph A. Khoory,4 Jennie M. Burns,5
Sergey S. Shevkoplyas,5 Anne Nicholson-Weller,1,6 and Ionita C. Ghiran1
1Division of Allergy and Inflammation and Division of Infectious Diseases, Beth Israel Deaconess Medical Center, Harvard Medical School, Boston, MA;2Department of Biologic Chemistry and Molecular Pharmacology, Harvard Medical School, Boston MA; 3Division of Hematology, Brigham and Women’sHospital, Boston, MA; 4Department of Biology and Biotechnology, Worcester Polytechnic Institute, Worcester, MA; 5Department of Biomedical Engineering,Tulane University, New Orleans, LA; and 6Divsion of Infectious Diseases, Beth Israel Deaconess Medical Center, Harvard Medical School, Boston, MA
Microbes as well as immune complexesand other continuously generated inflam-matory particles are efficiently removedfrom the human circulation by red bloodcells (RBCs) through a process calledimmune-adherence clearance. During thisprocess, RBCs use complement receptor1 (CR1, CD35) to bind circulatingcomplement-opsonized particles andtransfer them to resident macrophages inthe liver and spleen for removal. We here
show that ligation of RBC CR1 by anti-body and complement-opsonized par-ticles induces a transient Ca�� influx thatis proportional to the RBC CR1 levels andis inhibited by T1E3 pAb, a specific inhib-itor of TRPC1 channels. The CR1-elicitedRBC Ca�� influx is accompanied by anincrease in RBC membrane deformabilitythat positively correlates with the numberof preexisting CR1 molecules on RBCmembranes. Biochemically, ligation of
RBC CR1 causes a significant increase inphosphorylation levels of �-spectrin thatis inhibited by preincubation of RBCswith DMAT, a specific casein kinase IIinhibitor. We hypothesize that the CR1-dependent increase in membrane deform-ability could be relevant for facilitatingthe transfer of CR1-bound particles fromthe RBCs to the hepatic and splenicphagocytes. (Blood. 2010;116(26):6063-6071)
Introduction
In primates, in contrast to other vertebrates, clearing the intravascu-lar space of complement-opsonized inflammatory particles (eg,microbes and immune complexes) is mediated by circulating redblood cells (RBCs) using complement receptor 1 (CR1, CD35).1,2
During this process, known as immune-adherence clearance, RBCsimmobilize complement-tagged particles and transport them to theliver and spleen where resident macrophages remove the comple-ment-tagged particles and leave the RBCs intact. Immune-adherence clearance acts as a “buffer system,” preventing deposi-tion of circulating immune complexes in susceptible organs, suchas the kidney, and preventing activation of circulating leukocytesby inflammatory particles.3,4 We and others have also shown thatCR1-mediated immune-adherence promotes more efficient phago-cytosis and intracellular killing of complement-opsonized patho-gens compared with opsonized pathogens that are free-floating inplasma and not RBC-bound.5,6
We have previously found that, in circulating human RBCs,CR1 is disperse in RBC plasma membranes, and, after ligation byimmune particles, interacts with Fas-associated phosphatase-1 andrearranges into large clusters.7 Beneath the plasma membrane ofRBCs, the spectrin cytoskeleton defines a series of “corrals” thatare critical for maintaining RBC shape and deformability and forregulating the range and magnitude of lateral diffusion of mosttransmembrane proteins.8 The mechanical attributes of the spectrinmeshwork depend critically on the transient phosphorylation of�-spectrin, adducin, and protein 4.1R.9-11 Therefore, we hypoth-esized that ligation-mediated CR1 clustering is an active processwith CR1 directly affecting the phosphorylation status of cytoskel-
etal proteins and thus the mechanical properties of RBCs. We herereport that, in human RBCs, CR1 ligation induces a transient Ca��
influx that depends on stretch-activated transient receptor potentialchannel-1 (TRPC-1). In addition, CR1 ligation and Ca�� influxpromote phosphorylation of the cytoskeletal proteins, �-adducinand �-spectrin, which correlates with increased membrane deform-ability. Our study identifies CR1 ligation as an important eventaffecting RBC membrane deformability, which in itself could havean important role during the immune-adherence clearance process.
Methods
Antibodies and reagents
Antibodies (Abs) were obtained as follows: anti-CR1 monoclonal Abs(mAb): 1F11 (gift of Henry Marsh, Celldex Therapeutics, Needham, MA),YZ-1,12 and 2B11,13 rabbit polyclonal anti-CR1,2 nonimmune immunoglobu-lin G1 (IgG1; BD Biosciences); anti-TRPC1 rabbit polyclonal (Santa CruzBiotechnology); anti-TRPC1, T1E3 (gift of Yao Xiaoqiang, University ofHong Kong), anti-TRPC1 rabbit monoclonal anti-actin, anti-CD47, anti-adducin, anti-phospho-adducin (serine 726), anti-phospho serine/threoninemAbs, and anti–human glycophorin C (GPC) mAb (BRIC10; InternationalBlood Group Reference Laboratory; Abcam). Secondary Abs included:AlexaFluor488 goat anti–mouse IgG, AlexaFluor488 goat anti–rabbit IgG,AlexaFluor594 goat anti–rabbit IgG “highly cross absorbed,” and Alex-aFluor594 goat anti–mouse IgG “highly cross absorbed” (Invitrogen);horseradish peroxidase (HRP)-goat anti–mouse IgG, HRP-donkey anti–goat IgG, and HRP-donkey anti–rabbit IgG (Jackson ImmunoResearchLaboratories), GsMTx-4 (Peptide Institute). Reagents were obtained as
Submitted April 26, 2010; accepted September 8, 2010. Prepublished online asBlood First Edition paper, September 22, 2010; DOI 10.1182/blood-2010-04-273904.
*A.M.G. and R.M. contributed equally to this work.
The online version of this article contains a data supplement.
The publication costs of this article were defrayed in part by page chargepayment. Therefore, and solely to indicate this fact, this article is herebymarked ‘‘advertisement’’ in accordance with 18 USC section 1734.
© 2010 by The American Society of Hematology
6063BLOOD, 23 DECEMBER 2010 � VOLUME 116, NUMBER 26
For personal use only.on June 13, 2016. by guest www.bloodjournal.orgFrom

follows: Fluo-4-AM, eosin 5 maleimide (Invitrogen); IgG-free bovineserum albumin (BSA; Jackson ImmunoResearch Laboratories); inhibitorsfor casein kinase I, D4476, and casein kinase II, 2-dimethylamino-4,5,6,7-tetrabromo-1H-benzimidazole (DMAT; EMD Chemicals); phorbol 12-myristate 13-acetate (PMA), 2-(N-morpholino) ethanesulfonic acid (MES),and 2-aminoethoxydiphenyl borate (2-APB; Sigma-Aldrich).
Analysis of RBC calcium influx
RBCs (108) were preloaded with Fluo-4 AM for 15 minutes at roomtemperature (RT), washed, and resuspended in Hank balanced salt solution(HBSS) with Ca�� and Mg��. RBCs were incubated at RT for anadditional 10 minutes and washed once to remove any uncleaved Fluo-4AM. Due to the ATP-depleting effect of the acetoxymethyl group, allexperiments were performed within 1 hour from Fluo-4 AM loading.Fluorescence levels of RBCs were acquired for 15 seconds using a LSRIIflow cytometer (Becton Dickinson) to establish a baseline for intracellularRBC Ca�� concentration. Control or anti-CR1 Ab were then added to theRBCs, mixed briefly, and RBC fluorescence intensity was recorded for2 minutes. Data were exported as FSC 3.0 files without the time dimensionparameter and analyzed using the kinetic module of FlowJo 9.0.1 (Tree-Star). To measure saturated concentration of intracellular Ca��, Fluo-4-loaded RBCs were incubated with 5�M ionomycin. Intracellular Ca��
concentration after CR1 ligation was calculated using the followingformula:
nanomolar[Ca��] � Kd
(F � Fmin)
(Fmax � F)
where Kd is the dissociation constant for Ca��-bound Fluo-4 (around345nM)14; Fmax, the maximum intensity after ionomycin treatment; Fmin, theintensity of Fluo-4 loaded RBCs in the absence of Ca��; and F, theintensity of Fluo-4 after CR1 ligation.
Western blotting analysis
RBCs (2 �L packed cells) incubated with isotype control or anti-CR1 YZ-1mAb (10 �g/mL) were lysed in 100 �L 1 � reducing-loading buffer andboiled for 4 minutes. Samples were run on 10% Tris-HCl gels (Invitrogen)and transferred to nitrocellulose paper (Pierce). Membranes were thenincubated with Ab as noted in Figure legends, followed by appropriateHRP-conjugated secondary Ab, and developed using LAS 4000 imagingsystem (FujiFilm).
Assay of RBC membrane deformability using laser opticaltweezers (LOT)
Anti-GPC mAb was covalently attached to 1-�m polystyrene beads withcarboxyl surfaces (Polysciences). Beads (20 �L) were washed twice in1 mL 20mM MES buffer, pH 6.1. The carboxyl groups on the beads weresensitized using 100mM N-hydroxysuccinimide and 0.4% 1-ethyl-3-(3-dimethylaminopropyl) carbodiimide hydrochloride for 20 minutes at RT.Mouse anti-GPC antibody (10 �L of a 5 �g/mL solution) was added to thebead suspension and the mixture was incubated with constant shaking for2 hours at RT. BSA was then added to 1% (final concentration), and themixture was incubated for an additional 15 minutes. The conjugation reactionwas terminated by adding ethanolamine (100mM final concentration) for another15 minutes. The mixture was then washed twice in HBSS with 0.1% BSA.The conjugated beads were stored at 4°C and used within 1 week.
Anti-GPC-conjugated polystyrene beads were mixed with RBCs inHBSS with 0.5% BSA and placed between 2 glass coverslips to make a30-�m–thick sample. Most of the RBCs attached to the glass by a smallarea on their rim. LOT (stiffness 7.7 10�5N/m, calibrated using harmonicpotential approximation applied to the motion of an unattached trappedbead in solution) was used to trap a bead from the solution and bring thetrapped bead into contact with a RBC for 5 seconds, which usually resultedin strong attachment. A computer-controlled procedure translated thetrapped bead 2 �m at a rate of 20 �m/min extending the RBC into anelliptical shape, and then switched off the LOT, which released the bead andthe pulled RBC recovered its circular (discocytic) shape. The time required
for the shape recovery was calculated based on time-lapse images recordedat 30 frames/s. The first 0.5 seconds of each trajectory was fitted to anexponential with recovery time constant tc. This characteristic time forrelaxation is given by tc � �/�, where � is the viscosity of the RBCmembrane, and � is the shear elastic modulus of the membrane.15
Differences between the control and various experimental conditionswere compared using the nonparametric one-way analysis of variance(ANOVA) followed by Tukey-Kramer multiple comparison procedureperformed using MATLAB (MathWorks) and P values .05 wereconsidered statistically significant.
Assay of RBC deformability using a filtration device
To test the ability of RBCs to undergo capillary-like deformations, wefabricated a filter comprising an array of posts with 5 � 5-�m openings(channels) between them using polydimethylsiloxane. The design andfabrication of this microchannel 2-dimensional (2D) filter device andconfiguration of the experimental setup have been described previouslyin detail.16
To establish the flow of sample through the 2D filter device, the outlet ofthe device was connected to a waste reservoir (60-mL syringe barrel;Becton Dickinson) with a 60-cm long piece of PE-60 tubing (InstechLaboratories) filled with HBSS with Ca�� and Mg��. The differencebetween the level of liquid in the inlet reservoir of the 2D filter device andthe level of liquid in the waste reservoir provided the driving pressure. Thezero pressure difference corresponded to the absence of movement of RBCswithin the device. RBCs (8 �L, 20% hematocrit) were loaded into the inletreservoir and allowed to enter the network (approximately 1 minute) bylowering the waste reservoir tubing to a position located approximately15 cm below the zero pressure point. Once RBCs entered the capillary area,the waste reservoir tubing was raised to approximately 38 mm below thezero pressure point, which allowed RBCs to pass through the 25 �m lengthof the capillary in approximately 2-3 seconds. The height of the reservoirwas then left unchanged throughout the experiment. The passage of RBCsthrough the 2D filter was recorded using a 40�/0.75 (NA) Ph2 Plan Fluoriteobjective on a TE300 Nikon inverted microscope, using a Retiga Exi(QImaging) camera controlled by iVision 4.01 (BioVision) at a rate of10 frames/s. The images were analyzed frame by frame to measure the timefrom RBC channel contact to egress, and the results expressed as secondsper passage. Differences in passage time between the control and CR1-ligated RBCs with and without inhibitors were compared using theMann-Whitney test (Prism 4.0). A P value � .05 was considered statisti-cally significant.
Flow cytometry
RBCs were fixed and permeabilized as described in,7 and incubated for15 minutes with Ab (as noted in the figure legends) in 0.5% BSA in HBSSbuffer at 4°C, followed by 2 washes and incubation for 15 minutes withAlexaFluor488–labeled secondary Ab specific for each primary Ab at adilution recommended by the manufacturer. In each experiment, at least10 000 events were recorded using FACScan or LRSII (Becton Dickinson)and analyzed using CellQuest Pro Version 4.0.1 software (BD Biosciences).
Statistical analysis
The statistical analysis tests were performed using Prism Version 4.0(GraphPad Software) and are detailed for each experiment.
Results
Ligation of RBC CR1 induces Ca�� influx
RBCs are classically described as passive carriers of immunecomplexes from blood to resident macrophages in the liver andspleen. We have previously reported that the ligation of RBC CR1by complement-opsonized particles reorganizes CR1 into largeclusters.7 Here, we asked whether ligation of CR1 would trigger a
6064 GLODEK et al BLOOD, 23 DECEMBER 2010 � VOLUME 116, NUMBER 26
For personal use only.on June 13, 2016. by guest www.bloodjournal.orgFrom

RBC Ca�� influx. Unlike any other CR1-bearing cell type, theexpression patterns of CR1 on RBCs are genetically determinedwith RBCs expressing approximately 100 CR1/cell for low CR1expressors, 500/cell on intermediate CR1 expressors, and 1000/cellfor high CR1 expressors.17 To determine whether the levels of CR1expression on RBCs correlate with the magnitude of CR1-mediatedCa�� influx, we used RBCs from donors with known low and highRBC CR1 expression.7 RBCs were loaded with Fluo-4 andincubated with either irrelevant match control Ab, anti-CD47, oranti-CR1 mAb, and the results analyzed by flow cytometry. Ourresults show that ligation of RBC CR1 with mAb (Figure 1A)promoted a sustained Ca�� influx, with RBCs from high CR1donors displaying a more pronounced Ca�� influx (300nM)compared with RBCs from low CR1 expressors (75nM). Ligationof RBCs from either donor with anti-CD47 mAb did not trigger anymeasurable Ca�� influx (data not shown). When experiments wererepeated using complement-opsonized beads as physiologic CR1ligands (Figure 1B), a similar RBC Ca�� influx pattern wasobserved, although the amplitude and the differences between RBCCa�� influx of high (100nM) and low CR1 expressors (60nM)were less pronounced. Similarly, ligation of RBC CR1 with Absrendered a higher ratio between CR1 levels of high CR1 donors(mean fluorescence intensity [MFI] � 108) and low CR1 donors(MFI � 7.23, control Ab MFI � 2.23) (Figure 1C) compared withbinding of the fluorescently labeled complement opsonized beadsto high CR1 RBCs (MFI � 440; Figure 1D) and low CR1 RBCs(MFI � 110, control beads MFI � 3.55; Figure 1D). Our resultssuggest that, although the RBC CR1 levels vary by approximately10-fold between high and low CR1 expressors, from a functionalperspective the gap is less pronounced (Figures 1E-F). Becausecarrying immune-complexes and complement-opsonized particlessuch as viruses and bacteria could augment the Ca�� influxpromoted by shear stress experienced by circulating RBCs,18,19 we
next searched for an RBC Ca�� channel that is also amechanoreceptor.
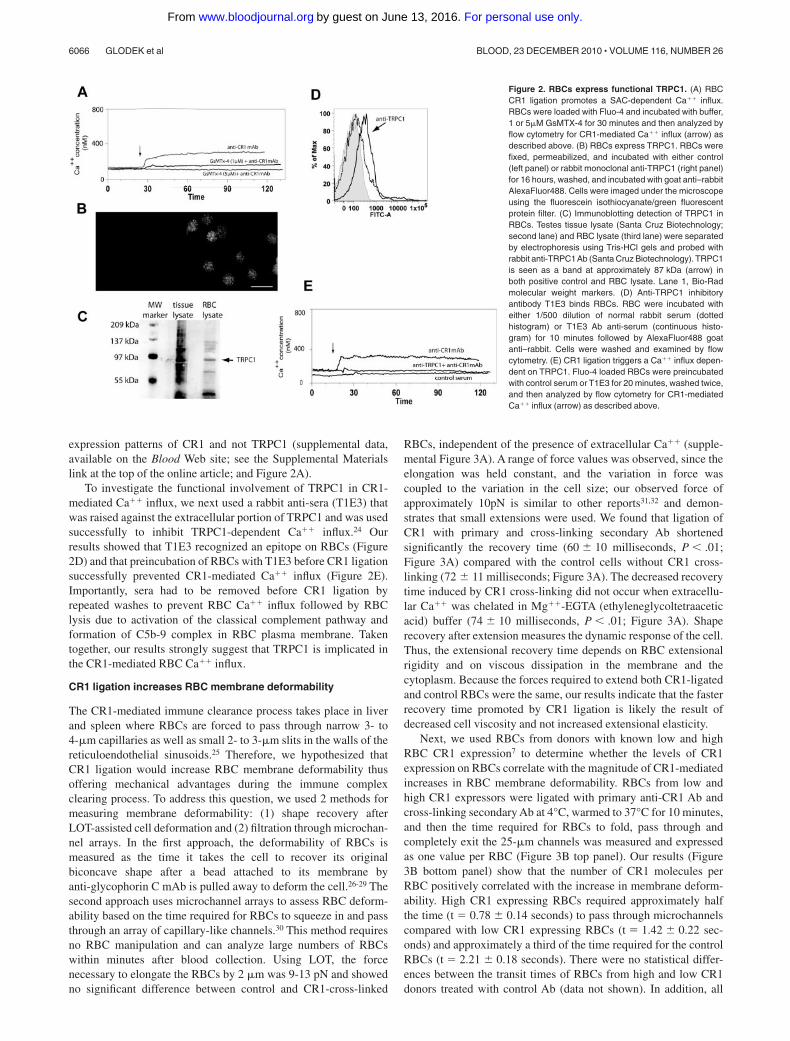
TRPC1 is involved in the Ca�� influx induced by CR1 ligationand shear stress
To verify the putative involvement of a stretch-activated cationchannel (SAC) in CR1-mediated Ca�� influx, we first usedGsMTx-4, a nonspecific peptide blocker of cationic SAC.20 Ourdata showed that preincubation of RBCs with increasing concentra-tions of GsMTx-4 (1�M and 5�M) progressively inhibited CR1mediated Ca�� influx as measured by flow cytometry (Figure 2A).We next focused on the widely expressed stretch-activated cationchannel, TRPC1, which was shown to promote Ca�� influx aftermechanical stimulation of cell membranes.21,22 We started byinvestigating the presence of TRPC1 in RBCs by using immunoflu-orescence microscopy, flow cytometry, and immunoblotting meth-ods. Fixed RBCs were permeabilized and reacted with either rabbitcontrol or rabbit anti-TRPC1 mAb, followed by goat anti-rabbitAlexaFluor488 conjugated IgG. Immunofluorescence microscopydemonstrated a distinct punctate pattern at the plasma membrane,uniformly distributed throughout the RBC population (Figure 2B).To further confirm the presence of TRPC1 in RBCs, we usedimmunoblot analysis of RBC lysates probed with rabbit anti-TRPC1 mAb (Santa Cruz Biotechnology). Our results show thatanti-TRPC1 pAb detected a protein running at a MW around85 kDa (Figure 2C right lane), which corresponded to the MW ofTRPC1 identified in testes tissue lysate (Santa Cruz Biotechnol-ogy) used as a positive control (Figure 2C left lane).23 In addition,analysis of RBCs from 2 low and 3 high CR1 donors by flowcytometry showed no consistent differences in RBC TRPC1expression levels, further suggesting that the differences measuredin Ca�� influx between various RBC donors are due to different
Figure 1. Ligation of RBC CR1 promotes Ca�� influx that depends on the genetically determined CR1 levels. (A) Antibody ligation of RBC CR1 triggers a Ca�� influx.Intra-RBC Ca�� concentration of Fluo-4–loaded RBCs from known high and low CR1 expressors was measured by flow cytometric analysis for 20 seconds before RBC CR1was ligated (arrow) by anti-CR1 mAb. Changes in intra-RBC Ca�� concentration were measured for a total of 2 minutes. (B) Ligation of CR1 by complementopsonized-particles promotes RBC Ca influx. Intra-RBC Ca�� concentration of Fluo-4–loaded RBCs from known high and low CR1 expressors was measured by flowcytometry for 20 seconds before RBC CR1 was ligated (arrow) by control or complement opsonized beads. Each line represents the average RBC fluorescence analyzed at arate of approximately 1000 RBC/second. (C) Characterization of RBC CR1 levels by antibody shows large gap between low and high CR1 expressors. RBCs from known lowand high CR1 expressors were incubated with anti-CR1 mAb followed by AlexaFluor488 secondary Ab and analyzed by flow cytometry. (D) Functional characterization of RBCCR1 using complement-opsonized beads identifies a narrower gap between RBCs from high and low expressors. RBCs from known low and high CR1 expressors wereincubated with fluorescently labeled complement opsonized beads for 30 minutes, washed, and analyzed by flow cytometry. (E) Fluorescence microscopy overlapped withphase contrast microscopy of RBCs from a high CR1 expressor after incubation with fluorescently labeled complement opsonized beads. (F) Fluorescence microscopyoverlapped with phase contrast microscopy of RBCs from a low CR1 expressor after incubation with fluorescently labeled complement opsonized beads. Bar represents10 �m. The results shown here are from the same individuals and are representative of 3 independent experiments using 3 different high CR1 and 2 different low CR1expressors.
CR1 INCREASES RBC MEMBRANE DEFORMABILITY 6065BLOOD, 23 DECEMBER 2010 � VOLUME 116, NUMBER 26
For personal use only.on June 13, 2016. by guest www.bloodjournal.orgFrom
expression patterns of CR1 and not TRPC1 (supplemental data,available on the Blood Web site; see the Supplemental Materialslink at the top of the online article; and Figure 2A).
To investigate the functional involvement of TRPC1 in CR1-mediated Ca�� influx, we next used a rabbit anti-sera (T1E3) thatwas raised against the extracellular portion of TRPC1 and was usedsuccessfully to inhibit TRPC1-dependent Ca�� influx.24 Ourresults showed that T1E3 recognized an epitope on RBCs (Figure2D) and that preincubation of RBCs with T1E3 before CR1 ligationsuccessfully prevented CR1-mediated Ca�� influx (Figure 2E).Importantly, sera had to be removed before CR1 ligation byrepeated washes to prevent RBC Ca�� influx followed by RBClysis due to activation of the classical complement pathway andformation of C5b-9 complex in RBC plasma membrane. Takentogether, our results strongly suggest that TRPC1 is implicated inthe CR1-mediated RBC Ca�� influx.
CR1 ligation increases RBC membrane deformability
The CR1-mediated immune clearance process takes place in liverand spleen where RBCs are forced to pass through narrow 3- to4-�m capillaries as well as small 2- to 3-�m slits in the walls of thereticuloendothelial sinusoids.25 Therefore, we hypothesized thatCR1 ligation would increase RBC membrane deformability thusoffering mechanical advantages during the immune complexclearing process. To address this question, we used 2 methods formeasuring membrane deformability: (1) shape recovery afterLOT-assisted cell deformation and (2) filtration through microchan-nel arrays. In the first approach, the deformability of RBCs ismeasured as the time it takes the cell to recover its originalbiconcave shape after a bead attached to its membrane byanti-glycophorin C mAb is pulled away to deform the cell.26-29 Thesecond approach uses microchannel arrays to assess RBC deform-ability based on the time required for RBCs to squeeze in and passthrough an array of capillary-like channels.30 This method requiresno RBC manipulation and can analyze large numbers of RBCswithin minutes after blood collection. Using LOT, the forcenecessary to elongate the RBCs by 2 �m was 9-13 pN and showedno significant difference between control and CR1-cross-linked
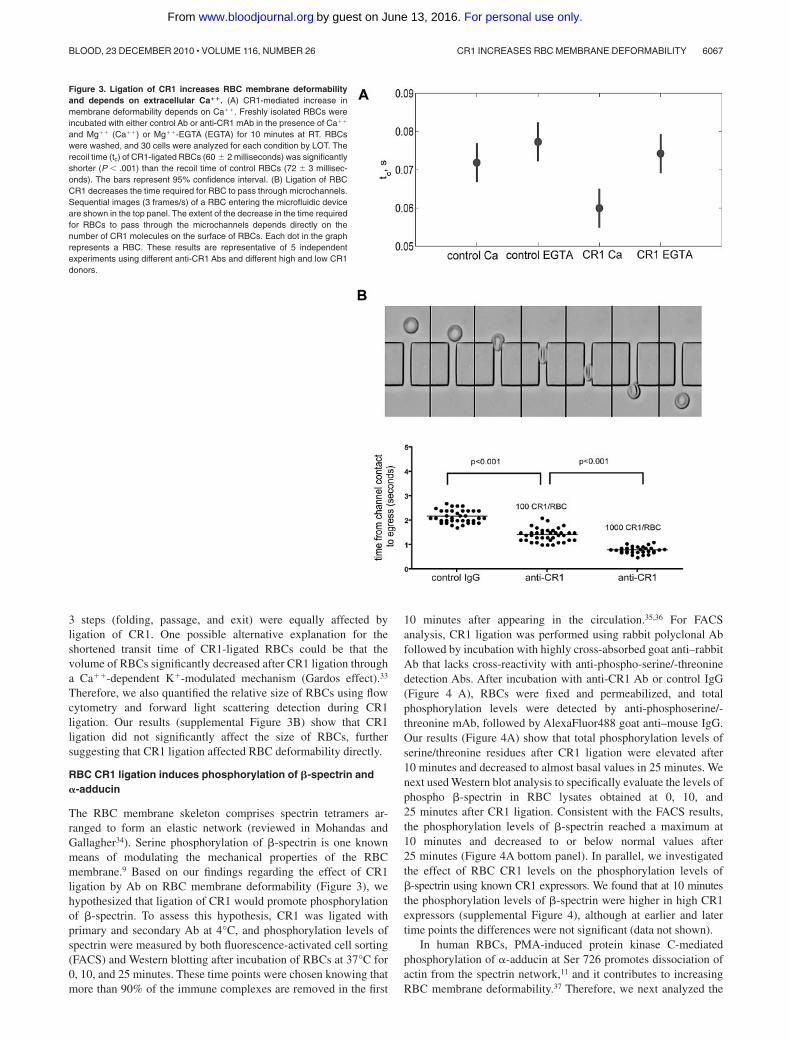
RBCs, independent of the presence of extracellular Ca�� (supple-mental Figure 3A). A range of force values was observed, since theelongation was held constant, and the variation in force wascoupled to the variation in the cell size; our observed force ofapproximately 10pN is similar to other reports31,32 and demon-strates that small extensions were used. We found that ligation ofCR1 with primary and cross-linking secondary Ab shortenedsignificantly the recovery time (60 10 milliseconds, P .01;Figure 3A) compared with the control cells without CR1 cross-linking (72 11 milliseconds; Figure 3A). The decreased recoverytime induced by CR1 cross-linking did not occur when extracellu-lar Ca�� was chelated in Mg��-EGTA (ethyleneglycoltetraaceticacid) buffer (74 10 milliseconds, P .01; Figure 3A). Shaperecovery after extension measures the dynamic response of the cell.Thus, the extensional recovery time depends on RBC extensionalrigidity and on viscous dissipation in the membrane and thecytoplasm. Because the forces required to extend both CR1-ligatedand control RBCs were the same, our results indicate that the fasterrecovery time promoted by CR1 ligation is likely the result ofdecreased cell viscosity and not increased extensional elasticity.
Next, we used RBCs from donors with known low and highRBC CR1 expression7 to determine whether the levels of CR1expression on RBCs correlate with the magnitude of CR1-mediatedincreases in RBC membrane deformability. RBCs from low andhigh CR1 expressors were ligated with primary anti-CR1 Ab andcross-linking secondary Ab at 4°C, warmed to 37°C for 10 minutes,and then the time required for RBCs to fold, pass through andcompletely exit the 25-�m channels was measured and expressedas one value per RBC (Figure 3B top panel). Our results (Figure3B bottom panel) show that the number of CR1 molecules perRBC positively correlated with the increase in membrane deform-ability. High CR1 expressing RBCs required approximately halfthe time (t � 0.78 0.14 seconds) to pass through microchannelscompared with low CR1 expressing RBCs (t � 1.42 0.22 sec-onds) and approximately a third of the time required for the controlRBCs (t � 2.21 0.18 seconds). There were no statistical differ-ences between the transit times of RBCs from high and low CR1donors treated with control Ab (data not shown). In addition, all
Figure 2. RBCs express functional TRPC1. (A) RBCCR1 ligation promotes a SAC-dependent Ca�� influx.RBCs were loaded with Fluo-4 and incubated with buffer,1 or 5�M GsMTX-4 for 30 minutes and then analyzed byflow cytometry for CR1-mediated Ca�� influx (arrow) asdescribed above. (B) RBCs express TRPC1. RBCs werefixed, permeabilized, and incubated with either control(left panel) or rabbit monoclonal anti-TRPC1 (right panel)for 16 hours, washed, and incubated with goat anti–rabbitAlexaFluor488. Cells were imaged under the microscopeusing the fluorescein isothiocyanate/green fluorescentprotein filter. (C) Immunoblotting detection of TRPC1 inRBCs. Testes tissue lysate (Santa Cruz Biotechnology;second lane) and RBC lysate (third lane) were separatedby electrophoresis using Tris-HCl gels and probed withrabbit anti-TRPC1Ab (Santa Cruz Biotechnology). TRPC1is seen as a band at approximately 87 kDa (arrow) inboth positive control and RBC lysate. Lane 1, Bio-Radmolecular weight markers. (D) Anti-TRPC1 inhibitoryantibody T1E3 binds RBCs. RBC were incubated witheither 1/500 dilution of normal rabbit serum (dottedhistogram) or T1E3 Ab anti-serum (continuous histo-gram) for 10 minutes followed by AlexaFluor488 goatanti–rabbit. Cells were washed and examined by flowcytometry. (E) CR1 ligation triggers a Ca�� influx depen-dent on TRPC1. Fluo-4 loaded RBCs were preincubatedwith control serum or T1E3 for 20 minutes, washed twice,and then analyzed by flow cytometry for CR1-mediatedCa�� influx (arrow) as described above.
6066 GLODEK et al BLOOD, 23 DECEMBER 2010 � VOLUME 116, NUMBER 26
For personal use only.on June 13, 2016. by guest www.bloodjournal.orgFrom
3 steps (folding, passage, and exit) were equally affected byligation of CR1. One possible alternative explanation for theshortened transit time of CR1-ligated RBCs could be that thevolume of RBCs significantly decreased after CR1 ligation througha Ca��-dependent K�-modulated mechanism (Gardos effect).33
Therefore, we also quantified the relative size of RBCs using flowcytometry and forward light scattering detection during CR1ligation. Our results (supplemental Figure 3B) show that CR1ligation did not significantly affect the size of RBCs, furthersuggesting that CR1 ligation affected RBC deformability directly.
RBC CR1 ligation induces phosphorylation of �-spectrin and�-adducin
The RBC membrane skeleton comprises spectrin tetramers ar-ranged to form an elastic network (reviewed in Mohandas andGallagher34). Serine phosphorylation of �-spectrin is one knownmeans of modulating the mechanical properties of the RBCmembrane.9 Based on our findings regarding the effect of CR1ligation by Ab on RBC membrane deformability (Figure 3), wehypothesized that ligation of CR1 would promote phosphorylationof �-spectrin. To assess this hypothesis, CR1 was ligated withprimary and secondary Ab at 4°C, and phosphorylation levels ofspectrin were measured by both fluorescence-activated cell sorting(FACS) and Western blotting after incubation of RBCs at 37°C for0, 10, and 25 minutes. These time points were chosen knowing thatmore than 90% of the immune complexes are removed in the first
10 minutes after appearing in the circulation.35,36 For FACSanalysis, CR1 ligation was performed using rabbit polyclonal Abfollowed by incubation with highly cross-absorbed goat anti–rabbitAb that lacks cross-reactivity with anti-phospho-serine/-threoninedetection Abs. After incubation with anti-CR1 Ab or control IgG(Figure 4 A), RBCs were fixed and permeabilized, and totalphosphorylation levels were detected by anti-phosphoserine/-threonine mAb, followed by AlexaFluor488 goat anti–mouse IgG.Our results (Figure 4A) show that total phosphorylation levels ofserine/threonine residues after CR1 ligation were elevated after10 minutes and decreased to almost basal values in 25 minutes. Wenext used Western blot analysis to specifically evaluate the levels ofphospho �-spectrin in RBC lysates obtained at 0, 10, and25 minutes after CR1 ligation. Consistent with the FACS results,the phosphorylation levels of �-spectrin reached a maximum at10 minutes and decreased to or below normal values after25 minutes (Figure 4A bottom panel). In parallel, we investigatedthe effect of RBC CR1 levels on the phosphorylation levels of�-spectrin using known CR1 expressors. We found that at 10 minutesthe phosphorylation levels of �-spectrin were higher in high CR1expressors (supplemental Figure 4), although at earlier and latertime points the differences were not significant (data not shown).
In human RBCs, PMA-induced protein kinase C-mediatedphosphorylation of �-adducin at Ser 726 promotes dissociation ofactin from the spectrin network,11 and it contributes to increasingRBC membrane deformability.37 Therefore, we next analyzed the
Figure 3. Ligation of CR1 increases RBC membrane deformabilityand depends on extracellular Ca��. (A) CR1-mediated increase inmembrane deformability depends on Ca��. Freshly isolated RBCs wereincubated with either control Ab or anti-CR1 mAb in the presence of Ca��
and Mg�� (Ca��) or Mg��-EGTA (EGTA) for 10 minutes at RT. RBCswere washed, and 30 cells were analyzed for each condition by LOT. Therecoil time (tc) of CR1-ligated RBCs (60 2 milliseconds) was significantlyshorter (P .001) than the recoil time of control RBCs (72 3 millisec-onds). The bars represent 95% confidence interval. (B) Ligation of RBCCR1 decreases the time required for RBC to pass through microchannels.Sequential images (3 frames/s) of a RBC entering the microfluidic deviceare shown in the top panel. The extent of the decrease in the time requiredfor RBCs to pass through the microchannels depends directly on thenumber of CR1 molecules on the surface of RBCs. Each dot in the graphrepresents a RBC. These results are representative of 5 independentexperiments using different anti-CR1 Abs and different high and low CR1donors.
CR1 INCREASES RBC MEMBRANE DEFORMABILITY 6067BLOOD, 23 DECEMBER 2010 � VOLUME 116, NUMBER 26
For personal use only.on June 13, 2016. by guest www.bloodjournal.orgFrom
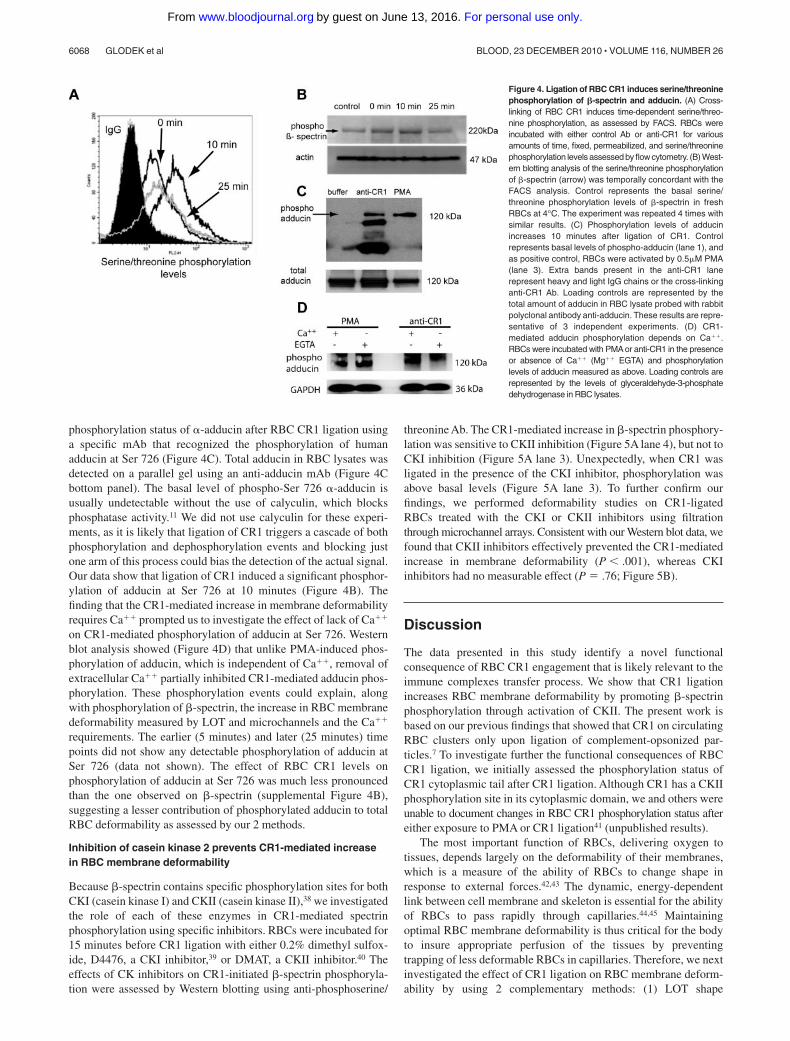
phosphorylation status of �-adducin after RBC CR1 ligation usinga specific mAb that recognized the phosphorylation of humanadducin at Ser 726 (Figure 4C). Total adducin in RBC lysates wasdetected on a parallel gel using an anti-adducin mAb (Figure 4Cbottom panel). The basal level of phospho-Ser 726 �-adducin isusually undetectable without the use of calyculin, which blocksphosphatase activity.11 We did not use calyculin for these experi-ments, as it is likely that ligation of CR1 triggers a cascade of bothphosphorylation and dephosphorylation events and blocking justone arm of this process could bias the detection of the actual signal.Our data show that ligation of CR1 induced a significant phosphor-ylation of adducin at Ser 726 at 10 minutes (Figure 4B). Thefinding that the CR1-mediated increase in membrane deformabilityrequires Ca�� prompted us to investigate the effect of lack of Ca��
on CR1-mediated phosphorylation of adducin at Ser 726. Westernblot analysis showed (Figure 4D) that unlike PMA-induced phos-phorylation of adducin, which is independent of Ca��, removal ofextracellular Ca�� partially inhibited CR1-mediated adducin phos-phorylation. These phosphorylation events could explain, alongwith phosphorylation of �-spectrin, the increase in RBC membranedeformability measured by LOT and microchannels and the Ca��
requirements. The earlier (5 minutes) and later (25 minutes) timepoints did not show any detectable phosphorylation of adducin atSer 726 (data not shown). The effect of RBC CR1 levels onphosphorylation of adducin at Ser 726 was much less pronouncedthan the one observed on �-spectrin (supplemental Figure 4B),suggesting a lesser contribution of phosphorylated adducin to totalRBC deformability as assessed by our 2 methods.
Inhibition of casein kinase 2 prevents CR1-mediated increasein RBC membrane deformability
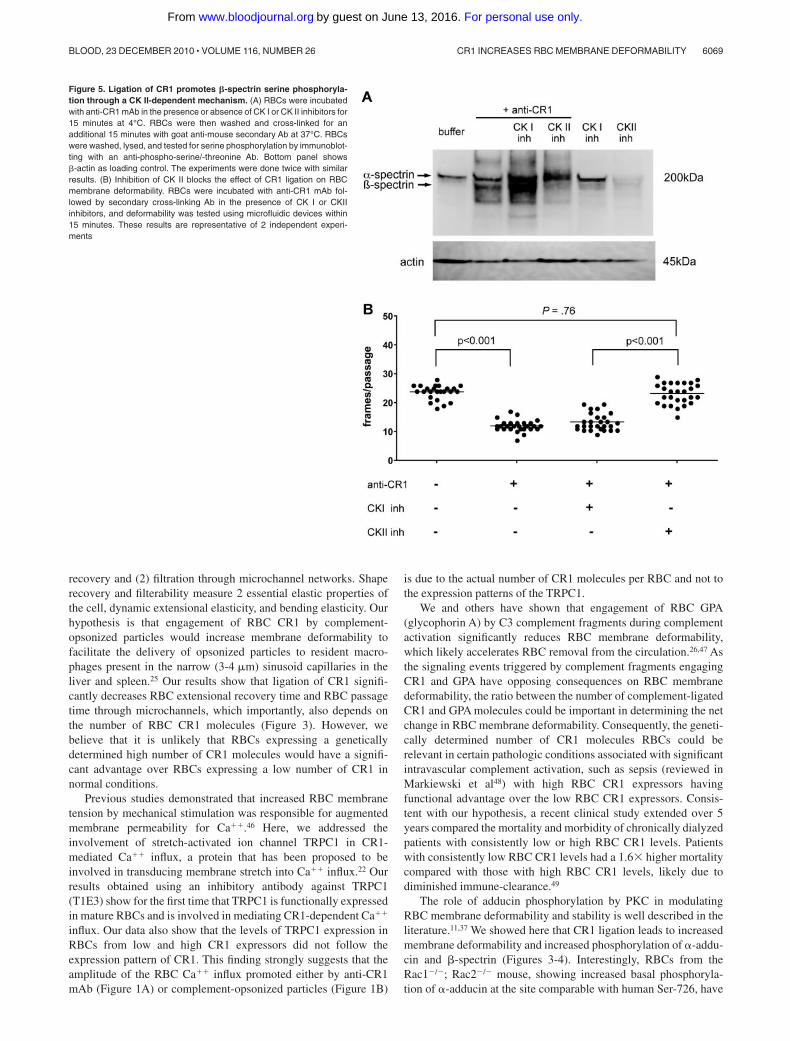
Because �-spectrin contains specific phosphorylation sites for bothCKI (casein kinase I) and CKII (casein kinase II),38 we investigatedthe role of each of these enzymes in CR1-mediated spectrinphosphorylation using specific inhibitors. RBCs were incubated for15 minutes before CR1 ligation with either 0.2% dimethyl sulfox-ide, D4476, a CKI inhibitor,39 or DMAT, a CKII inhibitor.40 Theeffects of CK inhibitors on CR1-initiated �-spectrin phosphoryla-tion were assessed by Western blotting using anti-phosphoserine/
threonine Ab. The CR1-mediated increase in �-spectrin phosphory-lation was sensitive to CKII inhibition (Figure 5A lane 4), but not toCKI inhibition (Figure 5A lane 3). Unexpectedly, when CR1 wasligated in the presence of the CKI inhibitor, phosphorylation wasabove basal levels (Figure 5A lane 3). To further confirm ourfindings, we performed deformability studies on CR1-ligatedRBCs treated with the CKI or CKII inhibitors using filtrationthrough microchannel arrays. Consistent with our Western blot data, wefound that CKII inhibitors effectively prevented the CR1-mediatedincrease in membrane deformability (P .001), whereas CKIinhibitors had no measurable effect (P � .76; Figure 5B).
Discussion
The data presented in this study identify a novel functionalconsequence of RBC CR1 engagement that is likely relevant to theimmune complexes transfer process. We show that CR1 ligationincreases RBC membrane deformability by promoting �-spectrinphosphorylation through activation of CKII. The present work isbased on our previous findings that showed that CR1 on circulatingRBC clusters only upon ligation of complement-opsonized par-ticles.7 To investigate further the functional consequences of RBCCR1 ligation, we initially assessed the phosphorylation status ofCR1 cytoplasmic tail after CR1 ligation. Although CR1 has a CKIIphosphorylation site in its cytoplasmic domain, we and others wereunable to document changes in RBC CR1 phosphorylation status aftereither exposure to PMA or CR1 ligation41 (unpublished results).
The most important function of RBCs, delivering oxygen totissues, depends largely on the deformability of their membranes,which is a measure of the ability of RBCs to change shape inresponse to external forces.42,43 The dynamic, energy-dependentlink between cell membrane and skeleton is essential for the abilityof RBCs to pass rapidly through capillaries.44,45 Maintainingoptimal RBC membrane deformability is thus critical for the bodyto insure appropriate perfusion of the tissues by preventingtrapping of less deformable RBCs in capillaries. Therefore, we nextinvestigated the effect of CR1 ligation on RBC membrane deform-ability by using 2 complementary methods: (1) LOT shape
Figure 4. Ligation of RBC CR1 induces serine/threoninephosphorylation of �-spectrin and adducin. (A) Cross-linking of RBC CR1 induces time-dependent serine/threo-nine phosphorylation, as assessed by FACS. RBCs wereincubated with either control Ab or anti-CR1 for variousamounts of time, fixed, permeabilized, and serine/threoninephosphorylation levels assessed by flow cytometry. (B) West-ern blotting analysis of the serine/threonine phosphorylationof �-spectrin (arrow) was temporally concordant with theFACS analysis. Control represents the basal serine/threonine phosphorylation levels of �-spectrin in freshRBCs at 4°C. The experiment was repeated 4 times withsimilar results. (C) Phosphorylation levels of adducinincreases 10 minutes after ligation of CR1. Controlrepresents basal levels of phospho-adducin (lane 1), andas positive control, RBCs were activated by 0.5�M PMA(lane 3). Extra bands present in the anti-CR1 lanerepresent heavy and light IgG chains or the cross-linkinganti-CR1 Ab. Loading controls are represented by thetotal amount of adducin in RBC lysate probed with rabbitpolyclonal antibody anti-adducin. These results are repre-sentative of 3 independent experiments. (D) CR1-mediated adducin phosphorylation depends on Ca��.RBCs were incubated with PMA or anti-CR1 in the presenceor absence of Ca�� (Mg�� EGTA) and phosphorylationlevels of adducin measured as above. Loading controls arerepresented by the levels of glyceraldehyde-3-phosphatedehydrogenase in RBC lysates.
6068 GLODEK et al BLOOD, 23 DECEMBER 2010 � VOLUME 116, NUMBER 26
For personal use only.on June 13, 2016. by guest www.bloodjournal.orgFrom
recovery and (2) filtration through microchannel networks. Shaperecovery and filterability measure 2 essential elastic properties ofthe cell, dynamic extensional elasticity, and bending elasticity. Ourhypothesis is that engagement of RBC CR1 by complement-opsonized particles would increase membrane deformability tofacilitate the delivery of opsonized particles to resident macro-phages present in the narrow (3-4 �m) sinusoid capillaries in theliver and spleen.25 Our results show that ligation of CR1 signifi-cantly decreases RBC extensional recovery time and RBC passagetime through microchannels, which importantly, also depends onthe number of RBC CR1 molecules (Figure 3). However, webelieve that it is unlikely that RBCs expressing a geneticallydetermined high number of CR1 molecules would have a signifi-cant advantage over RBCs expressing a low number of CR1 innormal conditions.
Previous studies demonstrated that increased RBC membranetension by mechanical stimulation was responsible for augmentedmembrane permeability for Ca��.46 Here, we addressed theinvolvement of stretch-activated ion channel TRPC1 in CR1-mediated Ca�� influx, a protein that has been proposed to beinvolved in transducing membrane stretch into Ca�� influx.22 Ourresults obtained using an inhibitory antibody against TRPC1(T1E3) show for the first time that TRPC1 is functionally expressedin mature RBCs and is involved in mediating CR1-dependent Ca��
influx. Our data also show that the levels of TRPC1 expression inRBCs from low and high CR1 expressors did not follow theexpression pattern of CR1. This finding strongly suggests that theamplitude of the RBC Ca�� influx promoted either by anti-CR1mAb (Figure 1A) or complement-opsonized particles (Figure 1B)
is due to the actual number of CR1 molecules per RBC and not tothe expression patterns of the TRPC1.
We and others have shown that engagement of RBC GPA(glycophorin A) by C3 complement fragments during complementactivation significantly reduces RBC membrane deformability,which likely accelerates RBC removal from the circulation.26,47 Asthe signaling events triggered by complement fragments engagingCR1 and GPA have opposing consequences on RBC membranedeformability, the ratio between the number of complement-ligatedCR1 and GPA molecules could be important in determining the netchange in RBC membrane deformability. Consequently, the geneti-cally determined number of CR1 molecules RBCs could berelevant in certain pathologic conditions associated with significantintravascular complement activation, such as sepsis (reviewed inMarkiewski et al48) with high RBC CR1 expressors havingfunctional advantage over the low RBC CR1 expressors. Consis-tent with our hypothesis, a recent clinical study extended over 5years compared the mortality and morbidity of chronically dialyzedpatients with consistently low or high RBC CR1 levels. Patientswith consistently low RBC CR1 levels had a 1.6� higher mortalitycompared with those with high RBC CR1 levels, likely due todiminished immune-clearance.49
The role of adducin phosphorylation by PKC in modulatingRBC membrane deformability and stability is well described in theliterature.11,37 We showed here that CR1 ligation leads to increasedmembrane deformability and increased phosphorylation of �-addu-cin and �-spectrin (Figures 3-4). Interestingly, RBCs from theRac1�/�; Rac2�/� mouse, showing increased basal phosphoryla-tion of �-adducin at the site comparable with human Ser-726, have
Figure 5. Ligation of CR1 promotes �-spectrin serine phosphoryla-tion through a CK II-dependent mechanism. (A) RBCs were incubatedwith anti-CR1 mAb in the presence or absence of CK I or CK II inhibitors for15 minutes at 4°C. RBCs were then washed and cross-linked for anadditional 15 minutes with goat anti-mouse secondary Ab at 37°C. RBCswere washed, lysed, and tested for serine phosphorylation by immunoblot-ting with an anti-phospho-serine/-threonine Ab. Bottom panel shows�-actin as loading control. The experiments were done twice with similarresults. (B) Inhibition of CK II blocks the effect of CR1 ligation on RBCmembrane deformability. RBCs were incubated with anti-CR1 mAb fol-lowed by secondary cross-linking Ab in the presence of CK I or CKIIinhibitors, and deformability was tested using microfluidic devices within15 minutes. These results are representative of 2 independent experi-ments
CR1 INCREASES RBC MEMBRANE DEFORMABILITY 6069BLOOD, 23 DECEMBER 2010 � VOLUME 116, NUMBER 26
For personal use only.on June 13, 2016. by guest www.bloodjournal.orgFrom

significantly decreased membrane deformability.44 There could be2 reasons for this apparent discrepancy between normal human andmouse Rac1�/�; Rac2�/� RBCs. First, the Rac1�/�; Rac2�/�
mouse may have an abnormally high actin-to-spectrin ratio thatcould directly cause cytoskeletal rearrangement; and second, themicrocytic anemia observed in the Rac1�/�; Rac2�/� mouse maycause an apparent decrease in membrane deformability. Impor-tantly, the effect of CR1 ligation on membrane deformability lastedsignificantly longer than its effect on spectrin phosphorylation(Figure 4), which would suggest that additional regulatory mecha-nisms such as interaction of spectrin with certain lipids fromplasma membrane are also important in modulating RBC mem-brane deformability and stability.50,51
Our studies using cell-permeant specific inhibitors for caseinkinases identified CKII as the main kinase responsible for phosphor-ylation of �-spectrin promoted by CR1 ligation and detected noeffect due to CKI inactivation (Figure 5). However, there areconflicting reports in the literature regarding the identity of theRBC CK responsible for phosphorylation of �-spectrin, with onereport identifying CK II38 and another one CKI.9 This seemingcontradiction may be explained by the fact in the paper identifyingCKI as the main �-spectrin kinase, the authors used: (1) [�32P] ATPas a phosphoryl donor, which can be used by both CKI and CKIIand (2) N-(2-aminoethyl)-5-chloroisoquinoline-sulfonamide, aninhibitor now known to inhibit ribosomal S6 kinase, serum/glucocorticoid regulated kinase, and stress/mitogen-activated ki-nase, in addition to CKI.39,52 Intriguingly, our data showed thatincubation of RBCs with CKI inhibitor increased the levels of�-spectrin phosphorylation, both in control and CR1-ligated RBCs(Figure 5). This unexpected result could be explained by theregulatory activity of CKI on serine/threonine protein phosphatasespresent in RBCs and responsible for dephosphorylation ofspectrin.53,54
In conclusion, we have demonstrated that ligation of RBC CR1triggers a complex Ca��-dependent signaling cascade leading to asignificant increase in plasma membrane deformability. In addition,
recognition of CR1 ligation, as well as its expression levels asimportant factors involved in regulating RBC membrane deform-ability will allow us a better understanding of immune adhesion aswell as the immune transfer process. Based on our work, one canalso speculate that in critically ill patients transfusion with RBCsfrom high CR1 expressors could be potentially advantageousespecially in pathologies associated with significant complementactivation.
Acknowledgments
We thank Henry Marsh for providing the anti-CR1 mAb, Xiao-qiang Yao for providing the T1E3 anti-TRPC1 pAb, and PeterWeller for critically reviewing the manuscript.
This work was supported by National Institutes of Health grantsAI42987 (A.N.W) and R01HL32854 (D.E.G). Part of the work wassupported by the W81XWH 07-1-0286 grant from the MedicalResearch and Material Command (MRMC) and FA9550 08-1-0364grant from the Defense Advanced Research Projects Agency.
Authorship
Contribution: I.C.G. designed the research, performed the majorityof the experiments, analyzed the data, and wrote the manuscript;A.M.G., R.M., S.S.S., and J.A.K. designed the research andperformed experiments; A.N.-W. and D.E.G. analyzed data andcritically reviewed the manuscript; and J.B. provided technicalassistance for red cell deformability experiments.
Conflict-of-interest disclosure: The authors declare no compet-ing financial interests.
Correspondence: Ionita C. Ghiran, Division of Allergy &Inflammation, Beth Israel Deaconess Medical Center, HarvardMedical School, Center for Life Science, CLS 930, 3 Blackfan Cir,Boston, MA 02115; e-mail: [email protected].
References
1. Nelson RAJ. The immune adherence phenom-enon: an immunologically specfic reaction be-tween microorganisms and erythrocytes leadingto enhanced phagocytosis. Science. 1953;118(3077):733-737.
2. Fearon DT. Identification of the membrane glyco-protein that is the C3b receptor of the humanerythrocyte, polymorphonuclear leukocyte, B lym-phocyte and monocyte. J Exp Med. 1980;152(1):20-30.
3. Ng YC, Schifferli JA, Walport MJ. Immune com-plexes and erythrocyte CR1 (complement recep-tor type 1): effect of CR1 numbers on binding andrelease reactions. Clin Exp Immunol. 1988;71(3):481-485.
4. Hess C, Schifferli JA. Immune adherence revis-ited: novel players in an old game. News PhysiolSci. 2003;18:104-108.
5. Nelson RAJ. The immune-adherence phenom-enon. A hypothetical role of erythrocytes in de-fense against bacteria and viruses. Proc RoyalSoc Med. 1956;49:55-58.
6. Pilsczek FH, Nicholson-Weller A, Ghiran I.Phagocytosis of Salmonella montevideo by hu-man neutrophils: immune adherence increasesphagocytosis, whereas the bacterial surface de-termines the route of intracellular processing.J Infect Dis. 2005;192(2):200-209.
7. Ghiran I, Glodek AM, Weaver G, Klickstein LB,Nicholson-Weller A. Ligation of erythrocyte CR1
induces its clustering in complex with scaffoldingprotein FAP-1. Blood. 2008;112(8):3465-3473.
8. Brown FL. Regulation of protein mobility via ther-mal membrane undulations. Biophys J. 2003;84(2 Pt 1):842-853.
9. Manno S, Takakuwa Y, Nagao K, Mohandas N.Modulation of erythrocyte membrane mechanicalfunction by beta-spectrin phosphorylation anddephosphorylation. J Biol Chem. 1995;270(10):5659-5665.
10. Matsuoka Y, Hughes CA, Bennett V. Adducinregulation. Definition of the calmodulin-bindingdomain and sites of phosphorylation by proteinkinases A and C. J Biol Chem. 1996;271(41):25157-25166.
11. Manno S, Takakuwa Y, Mohandas N. Modulationof erythrocyte membrane mechanical function byprotein 4.1 phosphorylation. J Biol Chem. 2005;280(9):7581-7587.
12. Changelian PS, Jack RM, Collins LA, Fearon DT.PMA induces the ligand-independent internaliza-tion of CR1 on human neutrophils. J Immunol.1985;134(3):1851-1858.
13. Chen CH, Ghiran I, Beurskens FJ, et al. AntibodyCR1-2B11 recognizes a non-polymorphic epitopeof human CR1 (CD35). Clin Exp Immunol. 2007;148(3):546-554.
14. Gee KR, Brown KA, Chen WN, Bishop-Stewart J,Gray D, Johnson I. Chemical and physiologicalcharacterization of fluo-4 Ca(2�)-indicator dyes.Cell Calcium. 2000;27(2):97-106.
15. Hochmuth RM, Worthy PR, Evans EA. Red cell ex-tensional recovery and the determination of mem-brane viscosity. Biophys J. 1979;26(1):101-114.
16. Shevkoplyas SS, Yoshida T, Gifford SC, BitenskyMW. Direct measurement of the impact of im-paired erythrocyte deformability on microvascularnetwork perfusion in a microfluidic device. LabChip. 2006;6(7):914-920.
17. Wilson JG, Murphy EE, Wong WW, Klickstein LB,Weis JH, Fearon DT. Identification of a restrictionfragment length polymorphism by a CR1 cDNAthat correlates with the number of CR1 on eryth-rocytes. J Exp Med. 1986;164(1):50-59.
18. Larsen FL, Katz S, Roufogalis BD, Brooks DE.Physiological shear stresses enhance the Ca2�permeability of human erythrocytes. Nature.1981;294(5842):667-668.
19. Brain MC, Pihl C, Robertson L, Brown CB. Evi-dence for a mechanosensitive calcium influx intored cells. Blood Cells Mol Dis. 2004;32(3):349-352.
20. Suchyna TM, Johnson JH, Hamer K, et al. Identi-fication of a peptide toxin from Grammostolaspatulata spider venom that blocks cation-selec-tive stretch-activated channels. J Gen Physiol.2000;115(5):583-598.
21. Riccio A, Medhurst AD, Mattei C, et al. mRNAdistribution analysis of human TRPC family inCNS and peripheral tissues. Brain Res Mol BrainRes. 2002;109(1-2):95-104.
22. Maroto R, Raso A, Wood TG, Kurosky A,
6070 GLODEK et al BLOOD, 23 DECEMBER 2010 � VOLUME 116, NUMBER 26
For personal use only.on June 13, 2016. by guest www.bloodjournal.orgFrom

Martinac B, Hamill OP. TRPC1 forms the stretch-activated cation channel in vertebrate cells. NatCell Biol. 2005;7(2):179-185.
23. Clapham DE, Julius D, Montell C, Schultz G. In-ternational Union of Pharmacology. XLIX. No-menclature and structure-function relationships oftransient receptor potential channels. PharmacolRev. 2005;57(4):427-450.
24. Kwan HY, Shen B, Ma X, et al. TRPC1 associateswith BK(Ca) channel to form a signal complex invascular smooth muscle cells. Circ Res. 2009;104(5):670-678.
25. Young B. Wheater’s Functional Histology: A Textand Colour Atlas (5th ed.) Edinburgh: ChurchillLivingstone/Elsevier; 2006.
26. Karnchanaphanurach P, Mirchev R, Ghiran I, etal. C3b deposition on human erythrocytes in-duces the formation of a membrane skeleton-linked protein complex. J Clin Invest. 2009;119(4):788-801.
27. Mirchev R, Golan DE. Single-particle tracking andlaser optical tweezers studies of the dynamics ofindividual protein molecules in membranes of in-tact human and mouse red cells. Blood Cells MolDis. 2001;27(1):143-147.
28. Bambardekar K, Dharmadhikari AK, DharmadhikariJA, Mathur D, Sharma S. Measuring erythrocytedeformability with fluorescence, fluid forces, andoptical trapping. J Biomed Opt. 2008;13(6):064021.
29. Brandao MM, Fontes A, Barjas-Castro ML, et al.Optical tweezers for measuring red blood cellelasticity: application to the study of drug re-sponse in sickle cell disease. Eur J Haematol.2003;70(4):207-211.
30. Shevkoplyas SS, Gifford SC, Yoshida T, BitenskyMW. Prototype of an in vitro model of the micro-circulation. Microvasc Res. 2003;65(2):132-136.
31. Henon S, Lenormand G, Richert A, Gallet F. Anew determination of the shear modulus of thehuman erythrocyte membrane using optical twee-zers. Biophys J. 1999;76(2):1145-1151.
32. Li J, Dao M, Lim CT, Suresh S. Spectrin-levelmodeling of the cytoskeleton and optical twee-zers stretching of the erythrocyte. Biophys J.2005;88(5):3707-3719.
33. Gardos G. The function of calcium in the potas-sium permeability of human erythrocytes. Bio-chim Biophys Acta. 1958;30(3):653-654.
34. Mohandas N, Gallagher PG. Red cell membrane:past, present, and future. Blood. 2008;112(10):3939-3948.
35. Schifferli JA, Ng YC, Estreicher J, Walport MJ.The clearance of tetanus toxoid-anti-tetanus tox-oid immune complexes from the circulation of hu-mans. Complement- and erythrocyte complementreceptor 1-dependent mechanisms. J Immunol.1988;140(3):899-904.
36. Halma C, Daha MR, Camps JA, Evers-SchoutenJH, Pauwels EK, Van E. Deficiency of comple-ment component C3 is associated with acceler-ated removal of soluble 123I-labelled aggregatesof IgG from the circulation. Clin Exp Immunol.1992;90(3):394-400.
37. de Oliveira S, Silva-Herdade AS, Saldanha C.Modulation of erythrocyte deformability by PKCactivity. Clin Hemorheol Microcirc. 2008;39(1-4):363-373.
38. Wei T, Tao M. Human erythrocyte casein kinaseII: characterization and phosphorylation of mem-brane cytoskeletal proteins. Arch Biochem Bio-phys. 1993;307(1):206-216.
39. Rena G, Bain J, Elliott M, Cohen P. D4476, a cell-permeant inhibitor of CK1, suppresses the site-specific phosphorylation and nuclear exclusion ofFOXO1a. EMBO Rep. 2004;5(1):60-65.
40. Hora R, Bridges DJ, Craig A, Sharma A. Erythro-cytic casein kinase II regulates cytoadherence ofPlasmodium falciparum-infected red blood cells.J Biol Chem. 2009;284(10):6260-6269.
41. Changelian PS, Fearon DT. Tissue-specific phos-phorylation of complement receptors CR1 andCR2. J Exp Med. 1986;163(1):101-115.
42. Vaya A, Lopez JM, Contreras MT, et al. Erythro-cyte deformability in survivors of acute myocar-dial infarction measured by two different method-ologies. Clin Hemorheol Microcirc. 2002;27(1):17-25.
43. Koksal C, Ercan M, Bozkurt AK. Hemorrheologi-cal variables in critical limb ischemia. Int Angiol.2002;21(4):355-359.
44. Kalfa TA, Pushkaran S, Mohandas N, et al. RacGTPases regulate the morphology and deform-ability of the erythrocyte cytoskeleton. Blood.2006;108(12):3637-3645.
45. Chasis JA, Mohandas N, Shohet SB. Erythrocytemembrane rigidity induced by glycophorin A-li-gand interaction. Evidence for a ligand-inducedassociation between glycophorin A and skeletalproteins. J Clin Invest. 1985;75(6):1919-1926.
46. Johnson RM. Membrane stress increases cationpermeability in red cells. Biophys J. 1994;67(5):1876-1881.
47. Parker CJ, Soldato CM, Rosse WF. Abnormalityof glycophorin-� on paroxysmal nocturnal hemo-globinuria erythrocytes. J Clin Invest. 1984;73(4):1130-1143.
48. Markiewski MM, DeAngelis RA, Lambris JD.Complexity of complement activation in sepsis.J Cell Mol Med. 2008;12(6A):2245-2254.
49. Ohi H, Tamano M, Okada N. Low CR1 (C3b re-ceptor) level on erythrocytes is associated withpoor prognosis in hemodialysis patients. NephronClin Pract. 2008;108(1):c23-c27.
50. An X, Debnath G, Guo X, et al. Identification andfunctional characterization of protein 4.1R andactin-binding sites in erythrocyte beta spectrin:regulation of the interactions by phosphatidylino-sitol-4,5-bisphosphate. Biochemistry. 2005;44(31):10681-10688.
51. An X, Guo X, Sum H, Morrow J, Gratzer W,Mohandas N. Phosphatidylserine binding sites inerythroid spectrin: location and implications formembrane stability. Biochemistry. 2004;43(2):310-315.
52. Bain J, Plater L, Elliott M, et al. The selectivity ofprotein kinase inhibitors: a further update. Bio-chem J. 2007;408(3):297-315.
53. Gross SD, Anderson RA. Casein kinase I: spatialorganization and positioning of a multifunctionalprotein kinase family. Cell Signal. 1998;10(10):699-711.
54. Usui H, Imazu M, Maeta K, Tsukamoto H, AzumaK, Takeda M. Three distinct forms of type 2A pro-tein phosphatase in human erythrocyte cytosol.J Biol Chem. 1988;263(8):3752-3761.
CR1 INCREASES RBC MEMBRANE DEFORMABILITY 6071BLOOD, 23 DECEMBER 2010 � VOLUME 116, NUMBER 26
For personal use only.on June 13, 2016. by guest www.bloodjournal.orgFrom
online September 22, 2010 originally publisheddoi:10.1182/blood-2010-04-273904
2010 116: 6063-6071
S. Shevkoplyas, Anne Nicholson-Weller and Ionita C. GhiranAleksandra M. Glodek, Rossen Mirchev, David E. Golan, Joseph A. Khoory, Jennie M. Burns, Sergey deformabilityLigation of complement receptor 1 increases erythrocyte membrane
http://www.bloodjournal.org/content/116/26/6063.full.htmlUpdated information and services can be found at:
(717 articles)Red Cells, Iron, and Erythropoiesis Articles on similar topics can be found in the following Blood collections
http://www.bloodjournal.org/site/misc/rights.xhtml#repub_requestsInformation about reproducing this article in parts or in its entirety may be found online at:
http://www.bloodjournal.org/site/misc/rights.xhtml#reprintsInformation about ordering reprints may be found online at:
http://www.bloodjournal.org/site/subscriptions/index.xhtmlInformation about subscriptions and ASH membership may be found online at:
Copyright 2011 by The American Society of Hematology; all rights reserved.of Hematology, 2021 L St, NW, Suite 900, Washington DC 20036.Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly by the American Society
For personal use only.on June 13, 2016. by guest www.bloodjournal.orgFrom
Recommended

















