
Vol. 4, No. 5MOLECULAR AND CELLULAR BIOLOGY, May 1984, P. 915-9220270-7306/84/050915-08$02.00/0Copyright C 1984, American Society for Microbiology
Polyamine Starvation Prolongs the S and G2 Phases of Polyamine-Dependent (Arginase-Deficient) CHO Cells
SIW ANEHUS,' PIRKKO POHJANPELTO,2 BO BALDETORP,3 EVA LANGSTROM,3 AND OLLE HEBY1*Department of Zoophysiology, University of Lund, Helgonavdgen 3, S-223 62 Lund, Sweden'; Department of Virology,University of Helsinki, Haartmaninkatu 3, SF-00290 Helsinki 29, Finland2; and Department of Oncology, University
Hospital, S-221 85 Lund, Sweden3
Received 14 November 1983/Accepted 1 February 1984
This study analyzes the effects of polyamine starvation on cell cycle traverse of an arginase-deficientCHO cell variant (CHO-A7). These cells grow well in serum-free medium, provided that it containsornithine or polyamines or both. In the absence of ornithine or polyamines or both, the CHO-A7 cellsdevelop severe polyamine deficiency and, as a consequence, grow more slowly. When grown to a stationaryphase in the presence of ornithine or putrescine or both, the CHO-A7 cells became arrested in GO/early G1.However, when starved for ornithine and polyamines, they accumulated in the S and G2 phases. Ornithineand polyamine starvation of CHO-A7 cells causes an increase in ornithine decarboxylase activity. Whenthis increase was prevented by treatment with DL-a-difluoromethylornithine, an enzyme-activated irrevers-ible inhibitor of ornithine decarboxylase, growth was further suppressed, and a greater fraction of cells werefound in the S and G2 phases of the cell cycle.
A high rate of synthesis of the polyamines putrescine,spermidine, and spermine is associated with cell growth anddivision (6, 18, 27). In eucaryotes, putrescine is formedsolely by decarboxylation of ornithine in a reaction cata-lyzed by ornithine decarboxylase (ODC) (18). Ornithine iseither taken up from the plasma or synthesized intracellular-ly by the action of arginase (18). In fact, arginase may beconsidered the initial enzyme ih the polyamine biosyntheticpathway, at least in cells that do not have a complete set ofurea cycle enzymes (11, 18).An obvious means of determining the physiological func-
tion of the polyamines is to use mutants with specific defectsin the various steps of polyamine biosynthesis. The firstsuccessful isolation of polyamine-dependent mammaliancells was recently reported. Thus, arginase-deficient (8, 21)and ODC-deficient (25) Chinese hamster ovary (CHO) celllines are presently being characterized. In the absence ofpolyamines in the medium, these cells develop severe poly-amine deficiency, and their proliferation gradually decreasesand eventually ceases (21, 25). These findings clearly dem-onstrate the requirement of polyamines for growth.Our present conception of cell cycle kinetics of poly-
amine-deficient cells is entirely based on experiments withinhibitors of polyamine biosynthetic enzymes (6). It is irre-futably important to compare these results with results ofexperiments in which polyamine deficiency has beenachieved by other means. Therefore, we have studied theeffects of polyamine starvation on the cell cycle kinetics ofthe arginase-deficient CHO cell variant CHO-A7. Becausethese cells cannot make their own ornithine and are main-tained in the absence of serum (a normal source of arginase)(8, 24), they are dependent on the addition of ornithine forpolyamine synthesis.Flow cytometric analysis revealed a progressive lengthen-
ing of the S and G2 phases, relative to G1, during the courseof starvation for ornithine and polyamines. Since the poly-amine-starved cells exhibited a compensatory increase in
* Corresponding author.
ODC activity, further polyamine depletion was attempted bysimultaneous treatment with tlL-a-difluoromethylornithine(DFMO), an enzyme-activated irreversible inhibitor of ODC(15). This treatment potentiated the inhibition of growth byfurther interference with S and G2 phase progression.
MATERIALS AND METHODSCell line. CHO-A7, an arginase-deficient cell line (8, 21),
was used in these experiments. These cells maintain anessentially normal rate of proliferation in serum-free medi-um, provided that it is supplemented with albumin andpolyamines (or ornithine).
Polyamine-starved CHO-A7 cells were checked for myco-plasma contamination by autoradiography after [3H]thymi-dine exposure (16), by fluorescence staining with Hoechst33258 dye (22), and by culture in mycoplasma medium. Alltests were negative.
Routine cultivation. The cells were grown as monolayercultures in gelatinized 90-mm plastic petri dishes (Nunc,Roskilde, Denmark) at 37°C in a high humidity 5% C02-95%air incubator. They were grown in Ham nutrient mixture F12(containing 1 ,uM putrescine) and Eagle minimum essentialmedium (1:1) supplemented with 0.2% bovine serum albu-min, penicillin (50 IU/ml)-streptomycin (50 p,gIml) mixture,0.1 mM L-ornithine, and 20 mM NaHCO3 (pH 7.2 to 7.3).The final concentration of putrescine in the growth mediumwas 0.5 p.M. The cells were subcultivated (1:20 to 1:40 split)every 3 to 4 days.Polyamine starvation. To deplete cells of their polyamines,
they were cultured in ornithine- and polyamine-free growthmedium. Further polyamine depletion was attempted byincluding 5 mM DFMO in the growth medium. In controlexperiments, the ornithine- and putrescine-free growth me-dium was supplemented with 0.1 mM L-ornithine, which issufficient to maintain a normal polyamine content. For theexperiments, 2.5 x 105 cells were seeded per dish in 15 ml ofgrowth medium. All growth media were changed every 2days.During the experimental period (8 days), cells were re-
moved every 24 h by trypsinization in the cold. Single-cell
915
on August 29, 2015 by guest
http://mcb.asm
.org/D
ownloaded from

916 ANEHUS ET AL.
suspensions in 5 to 10 ml of growth medium were obtainedby pipetting. Cells were pooled from two to six dishes, andthe cell number was counted in a hemacytometer. Sampleswere removed for flow cytometric analysis, enzyme assays,
and polyamine analysis.When polyamine starvation was extended for a longer
time period, cells were seeded at a lower density. In thisparticular experiment, plaque-purified CHO-A7 cells were
used. They were plated in 90-mm plastic petri dishes at a
density of 6 x 103 to 8 x 103 cells per cm2. Polyaminedepletion was accomplished essentially as described above,by growing the cells in ornithine- and polyamine-free medi-um in the absence or presence of 1 mM DFMO. Control cellswere grown in medium suppletnented with 10 ,uM putres-
cine. All growth media were changed every 2 days. Growthcurves were established for each experimental condition bycounting the cells every 2 to 3 days during a 14-day period.Samples were removed during stationary-phase growth (day10) for flow cytometric and polyamine analyses.
Analysis of enzyme activities and polyamine content. Theactivities of ODC and S-adenosylmethionine decarboxylase(SAMDC) and the contents of the polyamines were deter-mined as described previously (13).
Cell preparative procedure for flow cytometric analysis.Single-cell suspensions (1 x 106 to 2 x 106 cells) were fixedin 1 ml of absolute ethanol (-20°C). After fixation, the cellswere treated with RNase and pepsin (2). The resulting cellnuclei were passed through a nylon mesh with a pore size of50 ,um and finally stained with propidium iodide (2).
Flow cytometric analysis. Propidium iodide-strained nuclei ata density of about 1 x 106 nuclei per ml were analyzed on a
Cytofluorograph System 50-H (Ortho Instruments,Westwood, Mass.) equipped with a 4-W argon-ion laser(model 95; Lexel Corp., Palo Alto, Calif.). The laser line at
488 nm (450-mW power output) was used for excitation.Fluorescence emission was detected between 625 nm andinfrared. About 2 x 104 nuclei from each preparation were
analyzed. The impulses were sorted by a 512 multichannelanalyzer and processed with a PDP 11/34 computer (DigitalEquipment Corp., Maynard, Mass.). Nuclear doublets were
excluded by electronic threshold settings.The peak at approximately twice the modal channel num-
ber of Gl cell nuclei represents G2 cell nuclei. Obviously,mitotic cell nuclei should appear in the second peak. Howev-er, since most mitotic cells lack a nuclear membrane, theyare likely to be destroyed by pepsin (1). The nuclei of late-telophase cells may separate and appear in the first peak ofthe histogram (1). Pepsin treatment is also likely to dispersethe nuclei of multinucleate cells, which become frequentduring the course of polyamine starvation. The percentagesof cells in each phase of the cell cycle were derived from theDNA distributions essentially as described by Dean (3).
[3Hlthymidine incorporation. CHO-A7 cells were grown inmonolayer cultures in the presence of 0.1 mM ornithine and0.5 ,uM putrescine. On day 6, the cells were trypsinized andthen suspended and diluted in fresh growth medium. Aportion (2 x 106 cells in 1 ml) was added to each of 40 petridishes (90 mm) containing 14 ml of growth medium.
During the subsequent 20 h of growth, cells were pulse-labeled at 1-h intervals with 2.79 MBq of [methyl-3H]thymi-dine (specific activity, 1.48 TBq/mmol) for 30 min. Corre-sponding flow cytometric data were obtained in a parallelexperiment in which no [3H]thymidine was added. Theamount of label incorporated into acid-insoluble materialwas determined by liquid scintillation counting.
Chemicals. All growth media and antibiotics were pur-
w= [
0 1 2 3 4 5 6 7
DAYS AFTER PLATING
FIG. 1. Proliferation of CHIO-A7 (arginase-deficient) cells sub-jected to ornithine and polyamine starvation. Cells were grown inputrescine-free growth medium containing 0.1 mM L-ornithine (O),in ornithine- and putrescine-free growth mediunm (A), or in orni-thine- and putrescine-free growth medium supplemented with S mMDFMO (O). All growth media were changed every 2 days (dashedlines).
chased from GIBCO Europe, Paisley, Scotland. Bovineserum albumin (fraction V, 96 to 99% pure) atid RNase A(type III-A, bovine pancreas) were purchased from SigmaChemical Co., St. Louis, Mo_. Trypsin (crystallized twice,bovine pancreas) was purchased from Worthington Diagnos-tics, Freehold, N.J., pepsin was purchased from E. Merck
TABLE 1. Effect of orhithine and polyamine starvation on theprogression of CHO-A7 cells through the cell cycle
Supplement"~ Popula- Duration (h) of celltion cycle phase'
Ornithine Putres- DFMO doublingOrnithinecine
DFMO time (h) 61 S 62
+ - - 12.5 3.9 5.9 1.7- - - 16.5 5.3 8.0 2.2- - + 20.0 6.6 9.8 2.6
a Cells were plated in putrescine-free growth medium in theabsence or presence of ornithine (0.1 mM) or DFMO (5 mM).
b The G1, S, and G2 phase durations were calculated from cellcycle phase fractions (Fig. 5) derived from DNA histograms ofexponentially growing cells (day 2) (Fig. 1), assuming a growthfraction of 1.0 and a mitotic phase duration of 1.0 h during all threeculture conditions.
MOL. CELL. BIOL.
on August 29, 2015 by guest
http://mcb.asm
.org/D
ownloaded from

S AND G2 PROLONGATION DUE TO POLYAMINE DEFICIENCY 917
AG, Darmstadt, Federal Republic of Germany, and propi-dium iodide was purchased from Calbiochem-Behring, LaJolla, Calif.
RESULTSCell proliferation. To determine the effect of ornithine and
polyamine starvation on cell proliferation, CHO-A7 cellswere cultured in putrescine-free medium containing 0.1 mMornithine (control cells) and in ornithine- and putrescine-freemedium with or without 5 mM DFMO (Fig. 1). When thecells were starved for ornithine and polyamines (withoutaddition of DFMO), their growth rate decreased as com-pared to that of the control. The population doubling timesduring exponential growth (day 2) were 12.5 h for the controland 16.5 h for the starved cells (Table 1). The growth ratedecreased even further when the ornithine- and putrescine-free medium was supplemented with 5 mM DFMO. Thepopulation doubling time during exponential growth (day 2)was 20 h in these cells (Table 1). The cell density reached bycontrol cells in stationary phase (day 7) was 6 x 105 cells percm2. In polyamine-starved cultures, the corresponding cell
ODC
2 1 2
and -SAMDCaciiy(iooeof4 prhux 0cls)n
CHO-A7 cells subjected to ornit p i
symbols~~~~~~~ ~~~~~~ar epandithleedtFg.1$mila atrso
changesr o
milligram-of-protein bs.
4-~~~~~~~~~~
2-~~~~~~~~~~
OL~~~~~~~~~I12 4 5
DASAFE PAINGFIG.2.ODC activity (nanomole Of IC2prhu x1cls
andSAMDC activity (p~~~~icooesO IO pe orxI el)iCHO-A7cells subjected to om~~ithn an poymn traionh1yblr xlie ntelgn to Fi.1 iirptesochangeswere ohserved when the dat weeepese napr
milligram-of-protein bas~~~is.
2-~~ ~ ~ ~ ~ \
2 i c
0
E
0- 1 2 3I
z A N. P (
-J
0-
contents in CHO-A7 cells subjected to ornithine and polyaminestarvation. The symbols are explained in the legend to Fig. 1.Similar patterns of changes were observed when the data wereexpressed on a per-milligram-of-protein basis.
densities were only 33 and 26% of the control for untreatedand DFMO-treated cells, respectively.Polyamine synthesis. The effects of ornithine and poly-
amine depletion on the activities of the two rate-controllingenzymes in the polyamine biosynthetic pathway are shownin Fig. 2. During the first 2 days of growth, in the absence ofornithine and polyamines in the medium, the cellular ODCactivity increased 38-fold (as compared to the day 0 value).The ODC activity in the control cells increased only slightly(about twofold) during the same time period. It reached amaximum on day 3 (ninefold higher than the day 0 value).When the cells were grown in the absence of both ornithine
VOL. 4, 1984
on August 29, 2015 by guest
http://mcb.asm
.org/D
ownloaded from

918 ANEHUS ET AL.
ALLI
B1LL
i liI
ULU0 2 4 6 8
DAYS AFTER PLATINGFIG. 4. DNA histograms of CHO-A7 cells subjected to ornithine
and polyamine starvation. Cells were grown in (A) putrescine-freegrowth medium containing 0.1 mM L-ornithine, (B) ornithine- andputrescine-free growth medium, or (C) ornithine- and putrescine-free growth medium supplemented with 5 mM DFMO. The DNAdistributions were determined by flow cytometric analysis. Thevertical and horizontal dimensions represent cell number and cellu-lar DNA content, respectively, scaled arbitrarily. The estimated cellcycle phase fractions of these cell populations are shown in Fig. 5.
and polyamines and in the presence of 5 mM DFMO, theODC activity remained essentially at the day-0 levelthroughout the experiment.
In the two ornithine- and polyamine-starved cultures (withand without DFMO), there were eightfold increases in theSAMDC activities during the first 2 days of growth (ascompared to the day 0 value) (Fig. 2). The SAMDC activitieswere elevated in both cultures during the remainder of theexperimental period. In the control culture, the SAMDCactivity increased slightly (about threefold) during the firstday of growth, but then decreased continuously.Polyamine content. The changes in intracellular polyamine
content were followed during the course of ornithine andpolyamine starvation (Fig. 3). The putrescine content ofcontrol cells remained at about the same level for the first 5days of growth, but decreased toward the end of the experi-ment. In the two ornithine- and polyamine-starved cultures(with and without DFMO), however, the cellular putrescinecontent decreased below detectable levels during day 1.The spermidine content of control cells increased by 97%
during the first 2 days (Fig. 3). It remained elevated foranother 3 days before decreasing toward the day-0 level. Inthe two ornithine- and polyamine-starved cultures (with andwithout DFMO), the cellular spermidine content remained atthe initial level during the first day of growth. During day 2,the spermidine content decreased precipitously. From days2 to 7, the spermidipe content was 4 to 9% of the control inthe two ornithine- and polyamine-starved cultures (with andwithout DFMO).The spermine content of control cells showed a pattern of
changes similar to that of spermidine, but the increase wasless pronounced (38%) (Fig. 3). During day 1 of ornithine and
polyamine starvation, there was a 99 to 106% increase inspermine content in the two starved cultures (in the absenceand presence of DFMO). Thereafter, their spermine contentdecreased continuously toward the basal level.
Cell cycle kinetics. To reveal whether the deceleration ofgrowth in polyamine-starved cultures is due to interferencewith the progression through a particular phase of the cellcycle, the cellular DNA content was determined by flowcytometric analysis. Figure 4 shows the DNA distributionsof cell populations withdrawn at various stages of ornithineand polyamine starvation. The fractions of cells in the G1, S,and G2 phases of the cell cycle were estimated essentially asdescribed by Dean (3). The phase fractions of the variouscell populations were then plotted in time sequence (Fig. 5).From the time of plating until the cells exhibited an
exponential rate of growth (day 2) there was a decrease inthe G1 fraction and a corresponding increase in the S and G2fractions (Fig. 5). This was true for the control cultures aswell as for the two ornithine- and polyamine-starved cultures
80
60
40
2C
z0I-
LA.w
0.
SCR
20[
-0 1 2 3 4 5 6 7
DAYS AFTER PLATING
FIG. 5. G1, S, and G2 phase fractions in populations of CHO-A7cells subjected to omithine and polyamine starvation. The cell cyclephase fractions were derived from the DNA distributions shown inFig. 4, essentially as described by Dean (3). The symbols 0, A, andO are explained in the legend to Fig. 1 and correspond to A, B, andC, respectively, in Fig. 4.
Gl
G2
NOMM6--dML - -Alb.bM&AMM
II-s50-
to
?0-
0.1 - it
0 I
MOL. CELL. BIOL.
6
4
2
6=,.
on August 29, 2015 by guest
http://mcb.asm
.org/D
ownloaded from

S AND G2 PROLONGATION DUE TO POLYAMINE DEFICIENCY 919
A B
I I
r
FIG. 6. DNA histograms of plaque-purified CHO-A7 cells sub-jected to extended ornithine and polyamine starvation. Cells weregrown for 10 days in (A) ornithine- and putrescine-free growthmedium or (B) ornithine- and putrescine-free growth medium sup-plemented with 1 mM DFMO. The estimated cell cycle phasefractions of these cell populations are shown in Table 2. All growthmedia were changed twice a week.
(with and without DFMO). Thus, during the exponentialphase of growth (day 2) 35% of the cells were in G1, 52%were in S, and 13% were in G2. Assuming a growth fractionof 1.0 and a mitotic time of 1.0 h for cells in exponentialgrowth, one can estimate the durations of the G1, S, and G2phases by using the cell cycle phase fractions obtained onday 2 (Table 1).
In the control cultures, the G1 phase fraction increasedand the S and G2 phase fractions decreased during subse-quent growth (days 2 to 6) (Fig. 5). When the cells reachedtheir stationary phase (by day 6), about 95% of the cells werein G1. Of the remaining 5%, S-phase cells constituted 2%and G2-phase cells 3%.
In the ornithine- and polyamine-starved cultures (grown inthe absence of DFMO), the G1 phase fraction continued todecrease (days 2 to 4) (Fig. 5). From its minimum of 28%(day 4), the G1 phase fraction increased progressively to avalue of 64% during late-stationary phase (day 8). The S-phase fraction continued to increase and reached a maxi-
mum on day 4. From its maximum of 64%, the S-phasefraction decreased continuously and constituted only 31% inlate-stationary phase (day 8). The G2 phase fraction de-creased from day 2 (13%) and constituted only 5% of thecells in late-stationary phase (day 8).
In those ornithine- and polyamine-starved cultures thatwere treated with DFMO, the Gl-phase cells constituted a
somewhat larger fraction on days 4 and 6 (43 and 40%,respectively) than on day 2 (35%) (Fig. 5). The G1-phasecells constituted only 18% of the population, however, bylate-stationary phase (day 8). The S-phase fraction remainedessentially constant (50 to 52%) from days 2 through 6. Inlate-stationary phase, however, the S-phase cells constitutedmore than 67% of the entire population. The G2-phasefraction decreased from day 2 (13%) and constituted 7 to 8%of the cells on days 4 to 6. In late-stationary phase, however,it almost doubled (15%).
It is thus apparent that polyamine depletion slows growthby interfering with S-phase (and possibly G2-phase) progres-sion rather than by stopping the cells at their normal point ofarrest in GO/early G1.
Figure 6 shows the DNA distributions of plaque-purifiedCHO-A7 cells subjected to extended ornithine and poly-amine starvation (10 days). The corresponding cell cyclephase fractions are shown in Table 2 as are the polyaminecontents of these polyamine-starved cells. Cells were platedat a low cell density (6 x 103 to 8 x 103 cells per cm2) topermit growth for a 14-day period. The stationary phase wasreached on day 10, and the density of cells grown in theabsence of putrescine was 2.6 x 105 cells per cm2. Treatmentwith 1 mM DFMO in the absence of putrescine in themedium resulted in an even lower cell density in stationaryphase, namely, 1.5 x 105 cells per cm2. The growth curvesfor these experiments are not shown since they conveybasically the same information as those in Fig. 1.
It is apparent from these data that the cell cycle distribu-tion changes toward a higher average cellular DNA contentupon extended polyamine starvation (Fig. 6A, Table 2).Simultaneous inhibition of polyamine synthesis by DFMOtreatment potentiated this effect even further, and only 10%of the cells remained in Gl (Fig. 6B, Table 2).
After 10 days of polyamine starvation (in the presence orabsence of DFMO), the cellular polyamine content (Table 2)was suppressed to approximately the same extent as in theexperiment shown in Fig. 3.To determine whether control cells (growing in ornithine-
and putrescine-supplemented medium) become arrested inGO/early Gl or at a restriciton point in mid- or late G1, astationary-phase population (day 6) containing 85% of thecells in G1 was diluted, and the cells were plated in freshgrowth medium and then pulse-labeled with [3H]thymidine
TABLE 2. Effect of extended ornithine and polyamine starvation on polyamine content and cell cycle phase progression of plaque-purified CHO-A7 cells
Supplementa Polyamine content (nmol/106 cells)' Phase fraction (%)Ornithine Putrescine DFMO Putrescine Spermidine Spermine Gl S G2
- - - ND 0.561 1.465 28.1 41.4 30.5- - + ND 0.342 1.295 10.2 42.4 47.4
a Cells were grown for 10 days in the absence of ornithine and putrescine. To further decrease the cellular polyamine content, thecompensatory increase in ODC activity was inhibited by exposing cells to DFMO (1 mM final concentration) during the entire course ofornithine and polyamine starvation. All growth media were changed every 2 days.
b Putrescine, spermidine, and spermine content in cells subjected to 10 days of ornithine and polyamine starvation. ND, Not detectable.c Cell cycle phase fractions in cell populations subjected to 10 days of ornithine and polyamine starvation. The cell cycle phase fractions
were derived from the DNA distributions shown in Fig. 6.
VOL. 4, 1984
on August 29, 2015 by guest
http://mcb.asm
.org/D
ownloaded from
920 ANEHUS ET AL.
10
4
I0.i-u
2I-
O
cL
8z
cj
I
6
4
2
0
0
000iLL,iJ_I a,0J L.. L Lb L10 15
C)
100 -
z
a80 C)
60CD,mm
40 >0-I
z20_0e
0
20
HOURS AFTER PLATING
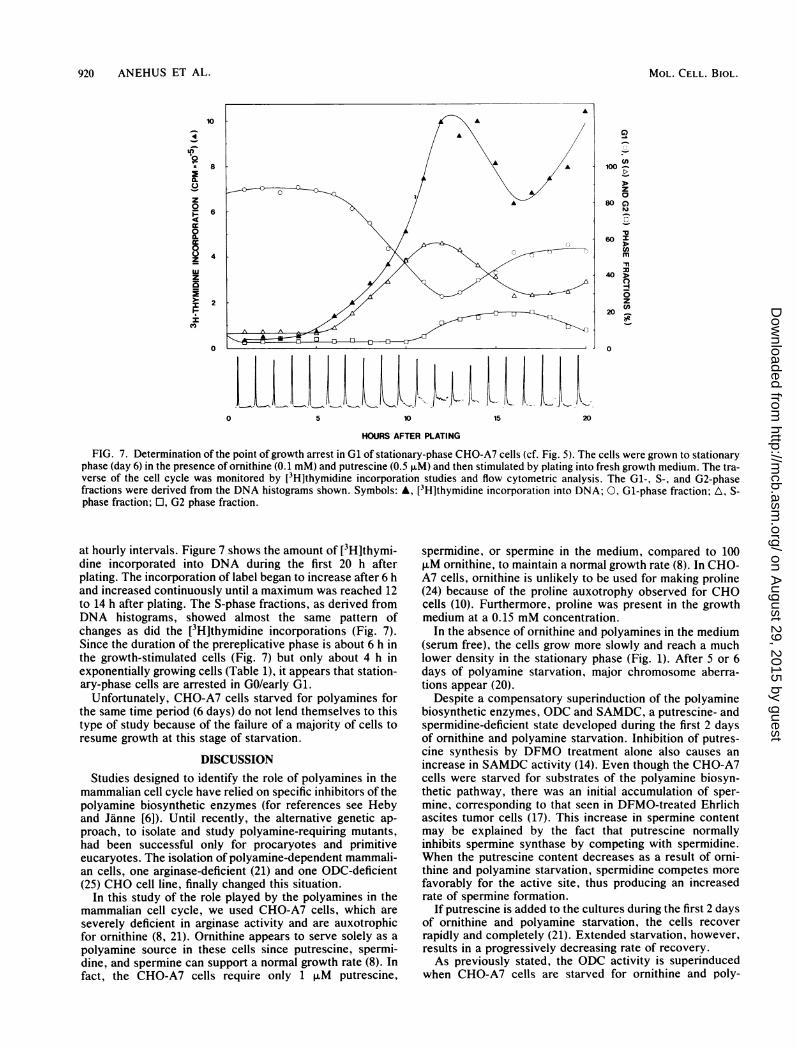
FIG. 7. Determination of the point of growth arrest in Gl of stationary-phase CHO-A7 cells (cf. Fig. 5). The cells were grown to stationaryphase (day 6) in the presence of ornithine (0.1 mM) and putrescine (0.5 ,uM) and then stimulated by plating into fresh growth medium. The tra-verse of the cell cycle was monitored by [3H]thymidine incorporation studies and flow cytometric analysis. The Gl-, S-, and G2-phasefractions were derived from the DNA histograms shown. Symbols: A, [3H]thymidine incorporation into DNA; 0, Gl-phase fraction; A, S-phase fraction; O, G2 phase fraction.
at hourly intervals. Figure 7 shows the amount of [3H]thymi-dine incorporated into DNA during the first 20 h afterplating. The incorporation of label began to increase after 6 hand increased continuously until a maximum was reached 12to 14 h after plating. The S-phase fractions, as derived fromDNA histograms, showed almost the same pattern ofchanges as did the [3H]thymidine incorporations (Fig. 7).Since the duration of the prereplicative phase is about 6 h inthe growth-stimulated cells (Fig. 7) but only about 4 h inexponentially growing cells (Table 1), it appears that station-ary-phase cells are arrested in GO/early G1.
Unfortunately, CHO-A7 cells starved for polyamines forthe same time period (6 days) do not lend themselves to thistype of study because of the failure of a majority of cells toresume growth at this stage of starvation.
DISCUSSIONStudies designed to identify the role of polyamines in the
mammalian cell cycle have relied on specific inhibitors of thepolyamine biosynthetic enzymes (for references see Hebyand Janne [6]). Until recently, the alternative genetic ap-proach, to isolate and study polyamine-requiring mutants,had been successful only for procaryotes and primitiveeucaryotes. The isolation of polyamine-dependent mammali-an cells, one arginase-deficient (21) and one ODC-deficient(25) CHO cell line, finally changed this situation.
In this study of the role played by the polyamines in themammalian cell cycle, we used CHO-A7 cells, which areseverely deficient in arginase activity and are auxotrophicfor ornithine (8, 21). Ornithine appears to serve solely as apolyamine source in these cells since putrescine, spermi-dine, and spermine can support a normal growth rate (8). Infact, the CHO-A7 cells require only 1 ,uM putrescine,
spermidine, or spermine in the medium, compared to 100,uM ornithine, to maintain a normal growth rate (8). In CHO-A7 cells, ornithine is unlikely to be used for making proline(24) because of the proline auxotrophy observed for CHOcells (10). Furthermore, proline was present in the growthmedium at a 0.15 mM concentration.
In the absence of ornithine and polyamines in the medium(serum free), the cells grow more slowly and reach a muchlower density in the stationary phase (Fig. 1). After 5 or 6days of polyamine starvation, major chromosome aberra-tions appear (20).
Despite a compensatory superinduction of the polyaminebiosynthetic enzymes, ODC and SAMDC, a putrescine- andspermidine-deficient state developed during the first 2 daysof ornithine and polyamine starvation. Inhibition of putres-cine synthesis by DFMO treatment alone also causes anincrease in SAMDC activity (14). Even though the CHO-A7cells were starved for substrates of the polyamine biosyn-thetic pathway, there was an initial accumulation of sper-mine, corresponding to that seen in DFMO-treated Ehrlichascites tumor cells (17). This increase in spermine contentmay be explained by the fact that putrescine normallyinhibits spermine synthase by competing with spermidine.When the putrescine content decreases as a result of orni-thine and polyamine starvation, spermidine competes morefavorably for the active site, thus producing an increasedrate of spermine formation.
If putrescine is added to the cultures during the first 2 daysof ornithine and polyamine starvation, the cells recoverrapidly and completely (21). Extended starvation, however,results in a progressively decreasing rate of recovery.As previously stated, the ODC activity is superinduced
when CHO-A7 cells are starved for ornithine and poly-
MOL. CELL. BIOL.
on August 29, 2015 by guest
http://mcb.asm
.org/D
ownloaded from

S AND G2 PROLONGATION DUE TO POLYAMINE DEFICIENCY 921
amines. At high ODC activities, not only ornithine but alsolysine may be used as a substrate (18). Thus, when CHO-A7cells were grown in the absence of ornithine, cadaverine wasgenerated at an increasing rate during the course of starva-tion (9). Since cadaverine acts as an aminopropyl groupacceptor in the spermidine synthase-catalyzed reaction,aminopropylcadaverine is formed. The production of theseunusual polyamines is likely, at least partly, to circumventgrowth arrest. Thus, cadaverine exerts a certain growth-stimulatory effect at concentrations (1 to 10 ,uM) close tothose observed in CHO-A7 cells during ornithine and poly-amine starvation (about 20 to 40 ,uM) (9). It should bestressed, however, that cadaverine is much less effective instimulating growth of CHO-A7 cells than are putrescine,spermidine, and spermine (8). Most probably, DFMO poten-tiates the suppression of growth of the ornithine- and poly-amine-starved CHO-A7 cells by irreversibly inhibiting thecatalytic activity of ODC (15), i.e., by preventing thesynthesis of cadaverine and its aminopropyl derivatives (9).When CHO-A7 cells reach a high density, they cease to
proliferate, and most of them are arrested in GO/early G1.Growth retardation caused by ornithine and polyamine star-vation, however, was not the result of GO/early Gl arrest.Instead, a large fraction of the CHO-A7 cells were found inthe S phase. If the duration of polyamine starvation wasextended and polyamine synthesis was simultaneously sup-pressed by DFMO, a majority of the cells exhibited a G2-phase (4C) DNA content. It has to be taken into account,however, that some of the cells with a 4C DNA content maybe tetraploid cells in the Gl phase of the tetraploid cell cycle.Large cell nuclei have been found in polyamine-starvedcultures (12).The effects of polyamine depletion on cell cycle progres-
sion, as studied by inhibitors of the polyamine biosyntheticenzymes, do not show a consistent pattern (4-7, 23, 26). Ithas been suggested that normal cells become arrested in G1,whereas transformed cells become arrested in S or G2 orboth upon polyamine limitation (26). This generalization didnot hold true, however. Notably, 9L rat brain tumor cellswere found to accumulate in G1 and not in S (7, 23). CHOcells were found mainly in the G1 and S phases of the cellcycle when blocked in their synthesis of polyamines (4, 26).Whether extended polyamine synthesis inhibition wouldcause a shift toward the G2 phase similar to that observed inthe present study remains to be determined for other celllines.Many experiments show that polyamine deficiency results
in a cell cycle distribution that is similar to that whichcharacterizes the particular cell type when it is in a state ofgeneral nutritional depletion (6). The CHO-A7 cells clearlyconstitute an exception, however, because when they aregrown to a high density in the presence of ornithine orputrescine in the medium, they become arrested in GO/earlyG1, whereas in the absence of polyamines they accumulatein the S and G2 phases. These data are taken to denote thatthe polyamines are not merely nutrients but exert a specificfunction in DNA replication.
That DNA replication is generally more sensitive to poly-amine starvation than are other events in the cell cycle isalso consistent with the fact that putrescine treatment short-ens the S phase in human fibroblasts (19). Data obtained inother laboratories indicate that polyamine depletion causes areduction in the number of active replicons as well as areduced rate of replication fork movement (reviewed byHeby and Janne [6]). The fact that cells accumulate in G2upon prolonged polyamine starvation suggests that cells may
accumulate errors in their DNA during replication, whichmake them incapable of going through mitosis. This explana-tion is also consistent with the continuous accumulation ofcells exhibiting chromosome aberrations (20). The fact thatpolyamine starvation results in disappearance of actin fila-ments and microtubules (21) suggests that polyamines mayalso have a role in cytokinesis. This possible function hasbeen proposed also on the basis of experiments with poly-amine synthesis inhibitors (26) and may explain the accumu-lation of multinucleate cells in polyamine-deficient cultures(21, 26).
ACKNOWLEDGMENTS
This investigation was supported by grants from the SwedishNatural Science and Medical Research Councils, the John andAugusta Persson and the Sigrid Juselius Foundations, and the RoyalPhysiographical Society (Lund).
DL-a-Difluoromethylornithine monohydrochloride monohydrate(RMI 71,782-A25) was a generous gift from Centre de Recherche,Merrell International. Strasbourg, France.
LITERATURE CITED
1. Beck, H.-P. 1977. Effect of pepsin pretreatment on pulse-cytophotometric DNA histograms. Cell Tissue Kinet. 10:265-268.
2. Crissman, H. A., A. P. Stevenson, R. J. Kissane, and R. A.Tobey. 1979. Techniques for quantitative staining of cellularDNA for flow cytometric analysis, p. 243-261. In M. R.Melamed, P. F. Mullaney, and M. L. Mendelsohn (ed.), Flowcytometry and sorting. John Wiley & Sons, Inc., New York.
3. Dean, P. N. 1980. A simplified method of DNA distributionanalysis. Cell Tissue Kinet. 13:299-308.
4. Harada, J. J., and D. R. Morris. 1981. Cell cycle parameters ofChinese hamster ovary cells during exponential, polyamine-limited growth. Mol. Cell. Biol. 1:594-599.
5. Heby, O., G. Andersson, and J. W. Gray. 1978. Interferencewith S and G2 phase progression by polyamine synthesisinhibitors. Exp. Cell Res. 111:461-464.
6. Heby, O., and J. Janne. 1981. Polyamine antimetabolites: bio-chemistry, specificity. and biological effects of inhibitors ofpolyamine synthesis. p. 243-310. In D. R. Morris and L. J.Marton (ed.). Polyamines in biology and medicine. MarcelDekker, New York.
7. Heby, O., L. J. Marton, C. B. Wilson, and J. W. Gray. 1977.Effect of methylglyoxal-bis(guanylhydrazone), an inhibitor ofspermidine and spermine synthesis. on cell cycle traverse. Eur.J. Cancer 13:1009-1017.
8. Holtta, E., and P. Pohjanpelto. 1982. Polyamine dependence ofChinese hamster ovary cells in serum-free culture is due todeficient arginase activity. Biochim. Biophys. Acta 721:321-327.
9. Holttia, E., and P. Pohjanpelto. 1983. Polyamine starvationcauses accumulation of cadaverine and its derivatives in apolyamine-dependent strain of Chinese hamster ovary cells.Biochem. J. 210:945-948.
10. Kao, F., and T. T. Puck. 1967. Genetics of somatic mammaliancells. IV. Properties of Chinese hamster cell mutants withrespect to the requirements for proline. Genetics 55:513-524.
11. Klein, D., and D. R. Morris. 1978. Increased arginase activityduring lymphocyte mitogenesis. Biochem. Biophys. Res. Com-mun. 81:199-204.
12. Knuutila, S., and P. Pohjanpelto. 1983. Polyamine starvationcauses parallel increase in nuclear and chromosomal aberrationsin a polyamine-dependent strain of CHO. Exp. Cell Res.145:222-225.
13. Lundquist, A., B. Lowkvist, M. Linden, and 0. Heby. 1983.Polyamines in early embryonic development: their relationshipto nuclear multiplication rate, cell cycle traverse, and nucleolarformation in a dipteran egg. Dev. Biol. 95:253-259.
14. Mamont, P. S., C. Danzin, J. Wagner, M. Siat, A.-M. Joder-Ohlenbusch, and N. Claverie. 1982. Accumulation of decarbox-ylated S-adenosyl-L-methionine in mammalian cells as a conse-
VOL. 4, 1984
on August 29, 2015 by guest
http://mcb.asm
.org/D
ownloaded from

922 ANEHUS ET AL.
quence of the inhibition of putrescine biosynthesis. Eur. J.Biochem. 123:499-504.
15. Metcalf, B. W., P. Bey, C. Danzin, M. J. Jung, P. Casara, andJ. P. Vevert. 1978. Catalytic irreversible inhibition of mammali-an ornithine decarboxylase (EC 4.1.1.17) by substrate andproduct analogues. J. Am. Chem. Soc. 100:2551-2553.
16. Nardone, R. M., J. Todd, P. Gonzalez, and E. V. Gaffney. 1965.Nucleoside incorporation into strain L cells: inhibition of pleu-ropneumonia-like organisms. Science 149:1100-1101.
17. Oredsson, S., S. Anehus, and 0. Heby. 1980. Irreversibleinhibition of the early increase in ornithine decarboxylaseactivity following growth stimulation is required to block Ehr-lich ascites tumor cell proliferation in culture. Biochem.Biophys. Res. Commun. 94:151-158.
18. Pegg, A. E., and H. G. Williams-Ashman. 1981. Biosynthesis ofputrescine, p. 3-42. In D. R. Morris and L. J. Marton (ed.),Polyamines in biology and medicine. Marcel Dekker, NewYork.
19. Pohjanpelto, P. 1975. Putrescine shortens the S-period in humanfibroblasts. Biomed. Express 23:350-352.
20. Pohjanpelto, P., and S. Knuutila. 1982. Polyamine deprivationcauses major chromosome aberrations in a polyamine-depen-dent Chinese hamster ovary cell line. Exp. Cell Res. 141:333-339.
21. Pohjanpelto, P., I. Virtanen, and E. Holtti. 1981. Polyamine
starvation causes disappearance of actin filaments and microtu-bules in polyamine-auxotrophic CHO cells. Nature (London)293:475-477.
22. Russell, W. C., C. Newman, and D. H. Williamson. 1975. Asimple cytochemical technique for demonstration of DNA incells infected with mycoplasmas and viruses. Nature (London)253:461-462.
23. Seidenfeld, J., J. W. Gray, and L. J. Marton. 1981. Depletion of9L rat brain tumor cell polyamine content by treatment with DL-a-difluoromethylornithine inhibits proliferation and the G1 to Stransition. Exp. Cell Res. 131:209-216.
24. Smith, R. J., and J. M. Phang. 1979. The importance ofornithine as a precursor for proline in mammalian cells. J. Cell.Physiol. 98:475-482.
25. Steglich, C., and I. E. Scheffler. 1982. An ornithine decarboxyl-ase-deficient mutant of Chinese hamster ovary cells. J. Biol.Chem. 257:4603-4609.
26. Sunkara, P. S., and P. N. Rao. 1981. Role of the polyamines inthe regulation of the cell cycle in normal and transformedmammalian cells, p. 267-291. In I. L. Cameron and T. B. Pool(ed.), The transformed cell. Academic Press, Inc., New York.
27. Williams-Ashman, H. G., and A. E. Pegg. 1981. Aminopropylgroup transfers in polyamine biosynthesis, p. 43-73. In D. R.Morris and L. J. Marton (ed.), Polyamines in biology andmedicine. Marcel Dekker, New York.
MOL. CELL. BIOL.
on August 29, 2015 by guest
http://mcb.asm
.org/D
ownloaded from
Recommended

















