
ARTICLE IN PRESS
0044-5231/$ - se
doi:10.1016/j.jc
�Correspond
Likovrissis, Gre
E-mail addr
Zoologischer Anzeiger 243 (2005) 181–209
www.elsevier.de/jcz
Praon Haliday (Hymenoptera: Braconidae: Aphidiinae) of Southeastern
Europe: key, host range and phylogenetic relationships
Nickolas G. Kavallieratosa,�, Zeljko Tomanovicb, Petr Staryc, Christos G. Athanassioud,Constantin Fassease, Olivera Petrovicf, Ljubisa Z. Stanisavljevicb,Maria Anagnou Veronikig
aLaboratory of Agricultural Entomology, Department of Entomology and Agricultural Zoology, Benaki Phytopathological Institute,
8 Stefanou Delta, 14561 Kifissia, Attica, GreecebInstitute of Zoology, Faculty of Biology, University of Belgrade, Studentski trg 16, 11000 Belgrade, Serbia and MontenegrocInstitute of Entomology, Academy of Sciences of the Czech Republic, Branisovska 31, 37005 Ceske Budejovice, Czech RepublicdLaboratory of Agricultural Zoology and Entomology, Agricultural University of Athens, 75 Iera Odos,
11855 Athens, Attica, GreeceeElectron Microscopy Laboratory, Agricultural University of Athens, 75 Iera Odos, 11855 Athens, Attica, GreecefDepartment of Plant Protection, Faculty of Agriculture, University of Belgrade, Nemanjina 6, 11081 Zemun,
Serbia and MontenegrogInsect Pathology and Microbiology Laboratory, Department of Entomology and Agricultural Zoology, Benaki Phytopathological
Institute, 8 Stefanou Delta str., 14561 Kifissia, Attica, Greece
Received 31 October 2003; received in revised form 20 October 2004; accepted 2 November 2004Corresponding editor: M. Schmitt
Abstract
A review of species in the genus Praon Haliday, 1833 is presented. Twenty described species are keyed and illustratedwith scanning electron micrographs and line drawings. The Praon species presented in this work have been identifiedfrom 67 aphid taxa occurring on 120 plant taxa. Furthermore, 87 original parasitoid–aphid–plant associations of thespecies mentioned in the key are presented. Phylogenetic relationships among Praon species are reconstructed usingparsimony and cladistic distance methods. Praon abjectum is the sister taxon to the remaining Praon species. Werecognized three species group: ‘‘Parapraon’’, ‘‘dorsale-yomenae’’ and ‘‘rosaecola’’. Monophyly is suggested for‘‘Parapraon’’ species group and paraphyly for ‘‘dorsale-yomenae’’ group. Finally, by phylogenetic reconstruction, aclose phylogenetic relationship between ‘‘Parapraon’’ and ‘‘dorsale-yomenae’’ species group was found.r 2004 Elsevier GmbH. All rights reserved.
Keywords: Review; Taxonomy; Morphological characters; Phylogeny; Southeastern Europe
e front matter r 2004 Elsevier GmbH. All rights reserved.
z.2004.11.001
ing author. Current address: P.O. Box 50803, 14123,
ece.
ess: [email protected] (N.G. Kavallieratos).
1. Introduction
With over 50 species described worldwide, thegenus Praon Haliday, 1833 is a large one in Aphidiinae.All species are solitary endoparasitoids of aphidsin Aphidinae, Myzocallidinae, Chaitophorinae and

ARTICLE IN PRESSN.G. Kavallieratos et al. / Zoologischer Anzeiger 243 (2005) 181–209182
Lachninae (Mackauer 1959; Stary 1970, 1981; Remau-diere and Remaudiere 1997). Unlike other aphidiinegenera, larval pupation takes place under the parasitizedaphid (Mackauer 1959; Stary 1970).
During the last several years many papers werepublished regarding Aphidiinae or Braconidae phylo-geny based on molecular data or great numbers ofmorphological or life history characters (Belshaw andQuicke 1997; Dowton et al. 1998; Smith et al. 1999;Quicke and Belshaw 1999; Sanchis et al. 2000; Kamb-hampati et al. 2000). Phylogenetic analyses of moleculardata suggest that the Praini clade (Praon+Dyscritulus)is basal among aphidiines (Dowton et al. 1998; Smithet al. 1999; Kambhampati et al. 2000). Praon hasretained many plesiomorphic characters such as: maleand female antennae with different number of segments,triangular lacinia, occipital carina complete, submargi-nal cell of hindwing present, host mummy light (seeQuicke and Belshaw 1999 for review of characters). Wetried to reconstruct the phylogenetic relationship withinthe genus Praon using 11 of the most informativecharacters for the species level. We compiled somecharacters from the phylogenetic literature (Stary 1976a;Gardenfors 1986; Quicke and van Achterberg 1990;Quicke and Belshaw 1999), and we used several originalcharacters with original coding strategies.
Due to great intraspecific variability there are manytaxonomic problems in the genus (Mackauer 1959; Stary1966, 1971); Tremblay and Pennacchio 1985; Mescheloffand Rosen 1988; Tomanovic et al. 2003a, b). Manyspecies are important parasitoids of aphids in variousagroecosystems. Some of them have been introducedworldwide against pest aphids (Bosch et al. 1959; Carver1984; Stary 1995; Dahlsten et al. 1999) and arecommercially reared as biological control agents (Hag-var and Hofsvang 1991; Heimpel and Lundgren 2000).
Mackauer (1959) revised the European species inPraon. Other important papers which studied Praon
systematics in Europe are Stary (1961, 1966, 1971,1976b, 1983), Stary et al. (1971), Tremblay andPennacchio (1985) and Tremblay et al. (1986). TheAsian species of Praon have been revised by Stary andSchlinger (1967), Takada (1968) and Mescheloff andRosen (1988). The most important treatments of NorthAmerican species of Praon are Smith (1944), Mackauer(1963), Johnson (1987) and Pike et al. (2000).
Several species in Praon were described and many newPraon–aphid–plant associations were reported in South-eastern Europe over the last 15 years (Tomanovic et al.1998; Kavallieratos and Lykouressis 1999–2000; Kaval-lieratos et al. 2001, 2003; Tomanovic and Kavallieratos2002; Tomanovic et al. 2003a b). In this paper, wereview 20 Praon species found in Southeastern Europe,with detailed external morphology by scanning electronmicroscope and drawings, and an original key for theidentification of the species is provided. Furthermore,
the Praon species presented in this work have beenidentified from 67 aphid taxa occurring on 120 planttaxa. We report 87 parasitoid–aphid associations, whichmight provide useful information about the host rangepattern of Praon spp., which is important in IPMprograms.
2. Material and methods
2.1. Collection and deposition of specimens
Specimens were collected during 1960–2002 in manylocalities in Serbia and Montenegro, Bosnia andHerzegovina, The Former Yugoslav Republic of Mace-donia, Bulgaria, Greece and Turkey (a detailed list oflocalities, sampling dates and sex ratios are availablefrom the authors). Samples from various host plantsbearing aphid colonies, consisting of both live andmummified aphids, were collected. Live aphids werekilled and preserved in 90% ethanol and 75% lactic acid2:1 (Eastop and van Emden 1972) for identification at alater date. The remaining aphid colonies were main-tained in the laboratory until parasitoid emergence.Mummies, each attached on a small leaf piece, wereplaced separately in small plastic boxes and were putinside a growth cabinet. On the lid of each box there wasa circular opening covered with muslin for ventilation inorder to maintain similar conditions inside the boxes tothose in the growth cabinet (22.5 1C, relative humidity65%, 16L:8D) (Kavallieratos et al. 2001, 2002a, b,2003). The external structure of the emerged parasitoidswas studied using an OLYMPUS SZX 9 stereomicro-scope. Several specimens were gold coated with a sputtercoater and examined using a Cambridge S 150 scanningelectron microscope. Material examined in this study isdeposited in the collection of the Belgrade NaturalHistory Museum (Serbia and Montenegro), the Instituteof Zoology, Faculty of Biology, University of Belgrade(Serbia and Montenegro) and the Laboratory ofAgricultural Zoology and Entomology, AgriculturalUniversity of Athens (Greece). Terminology used inthis paper regarding the diagnostic characters of theaphidiines is based on Huber and Sharkey (1993), Pikeand Stary (1995) and Kavallieratos et al. (2001).
2.2. Diagnostic characters used in the key
Pubescence of the face, number of antennal segments,number of longitudinal placodes on second flagellomere( ¼ F2), length of first flagellomere ( ¼ F1), lengthof F2, width of F1, colour of F1 and F2, length ofstigma, width of stigma, length of distal abscissa of R1
( ¼ metacarpus), degree of development and colour ofm-cu vein, development of radial sector vein ( ¼ Rs),

ARTICLE IN PRESSN.G. Kavallieratos et al. / Zoologischer Anzeiger 243 (2005) 181–209 183
pubescence of lateral lobes of mesonotum, pubescenceof propodeum, pubescence of tergite 1, shape ofovipositor sheath; shape of dorsal margin of ovipositorsheath, prominence of ovipositor sheath, number ofconical apical spines of ovipositor sheath.
2.3. Phylogenetic analyses
An unweighted cladystic analysis of the Praon specieswas undertaken. We used 50% majority-rule parsimonyanalysis and distance methods. Ephedrus persicae wasused as an outgroup. Eleven characters were coded (tenmorphological and one life history character) and arepresented in Table 1. The data matrix was analyzed usingPAUP* version 4(10b) (Swofford 2003). The tree wasvisualized and printed using TREE VIEW (Page 1996).
2.4. Characters description
1.
Tab
Tax
P. g
P. n
P. r
P. m
P. p
P. v
P. b
P. a
P. fl
P. b
P. e
P. y
P. l
P. u
P. r
P. a
P. s
P. a
P. o
P. n
E. p
Ephe
Host range patterns: 0 ¼ Lachninae; 1 ¼ Myzo-callidinae; 2 ¼ Aphidinae.We presumed a plesiomorphous character stateparasitism on more basal aphid lineages as Lachni-nae and Myzocallidinae rather than Aphidinae(Stary, 1981).
2.
Forewing stigma: 0 ¼ less than 3.5 time as long aswide (proportion of long of forewing stigma andwide at Radial cector (Rs)); 1 ¼ elongated, morethan 3.5 time as long as wide.le 1. Character matrix used in cladistic analysis of Praon species
a Characters
1 2 3 4 5
allicum 2 0 2 1 1
ecans 2 0 2 1 1
etusae 2 1 2 1 1
egourae 2 1 1 1 0
ubescens 2 ? 0 1 0
olucre 2 ? 0 0 1
arbatum 2 1 1 1 1
bjectum 2 1 0 1 0
avinode 1 1 0 1 0
icolor 0 1 0 1 0
xsoletum 2 0 2 0 0
omenae 2 ? 1 1 1
ongicorne 2 0 0 1 0
roleucon 2 ? 1 1 0
osaecola 2 0 0 1 0
bsinthii 2 0 1 1 1
taryi 2 ? 0 1 0
thenaeum 2 1 1 0 0
rpheusi 2 0 0 1 0
onveilleri 2 ? 1 1 0
ersicae ? 1 0 0 0
drus persicae was used as outgroup.
Some species have a large range of variation of thischaracter around limit value. In these unresolvedcases we put question mark.
3.
Forewing vein m-cu: 0 ¼ fully sclerotized andcolored; 1 ¼ colorless, partly sclerotized or partlyeffaced; 2 ¼ absent or significant reduced.4.
Forewing vein Rs+M: 0 ¼ partly sclerotized andcolored; 1 ¼ colorless.5.
Ovipositor sheath: 0 ¼ straight dorsaly; 1 ¼ dorsalmargin clearly concave.We presumed that a curved and a longer ovipositorsheath is apomorphous character state because ofthe easy host aphid accessibility during oviposition(Stary, 1976a).6.
Ovipositor sheath: 0 ¼ short; 1 ¼ elongated. Seeexplanation in 5.7.
Conical apical spines: 0 ¼ absent; 1 ¼ one; 2 ¼ two.This character is specific for Praon and represents anadaptive advantage during ovipostion (Mackauer,1959).8.
Tergite I: 0 ¼ subquadrate, 1.1–1.3 time as long aswide at spiracles; 1 ¼ more elongated, about 1.5time as long as wide at spiracles.9.
Number of longitudinal placodes on flagellomere 2(F2): 0 ¼ 4 and more; 1 ¼ 1–3; 2 ¼ 0.10.
Flagellomere 1 (F1): 0 ¼ shorter than 5.0 times aslong as wide (proportion between length of F1 andwidth at the basal part); 1 ¼ longer than 5.0 times aslong as wide.6 7 8 9 10 11
1 2 0 2 0 1
1 2 0 1 0 1
1 2 0 1 1 1
1 1 1 1 ? 1
1 2 0 1 1 0
1 2 0 1 1 0
1 2 0 1 1 0
0 0 0 1 0 1
0 2 1 1 1 1
0 2 1 1 1 0
1 2 0 1 ? 1
1 2 0 1 1 1
1 2 1 2 1 0
1 2 0 0 ? 1
0 2 0 1 1 0
1 2 0 1 1 1
1 2 0 1 1 0
1 1 0 1 1 1
1 2 0 1 0 0
1 2 0 ? 1 1
1 0 0 ? 0 1

ARTICLE IN PRESSN.G. Kavallieratos et al. / Zoologischer Anzeiger 243 (2005) 181–209184
11.
Lateral lobes of mesonotum: 0 ¼ densely pubescent;1 ¼ with large hairless areas.3. Results
3.1. Key to the species of Praon Haliday, 1833 in
Southeastern Europe (based on adult females)
1
Rs vein of forewing point like (Fig. 2) P.orpheusi Kavallieratos, Athanassiou andTomanovic, 2003
1*
Rs vein of forewing normally developed(Figs. 9, 16, 23, 27, 33, 40, 46, 53, 58, 61, 66,72, 79, 85, 91, 98, 105, 112)............................. 22
Forewing vein m-cu not developed (Figs. 16,23, 27) or weakly developed (Fig. 9).............. 32*
Forewing vein m-cu coloured throughout(Figs. 33, 40, 46, 53, 61, 66, 72, 79); colourlessthroughout (Figs. 58, 85, 91, 112); erased atits second part and colourless (Fig. 98);coloured at about first third and colourless atthe rest part (Fig. 105)................................... 63
Propodeum covered with few long hairs (Fig.11); tergite 1 almost hairless (Fig. 12); dorsaloutline of ovipositor sheath almost straight(Fig. 13); antennae 17–18-segmented P.exsoletum (Nees, 1811)
3* Propodeum densely pubescent (Figs. 18, 25,29); tergit 1 covered with long or short hairsalong the sides (Figs. 19, 26, 30); dorsaloutline of ovipositor sheath concave (Figs.20, 31); antennae 15-18-segmented.................
44
F1 and F2 yellow; F2 without longitudinalplacodes; antennae 16-17 (18)-segmentedP.gallicum Stary, 1971
4* F1 yellow or brown; F2 brown; F2 with onelongitudinal placode......................................
5 5 F1 brown; antennae15-16-segmented;proportion between length and width offorewing stigma about 3.2 (Fig. 23);tergite 1 covered with long hairs along thesides (Fig. 26) P. necans Mackauer, 1959
5*
F1 yellow; antennae 18-segmented;proportion between length and width offorewing stigma about 4.0 (Fig. 27); tergite 1covered with short hairs along the sides(Fig. 30) P. retusae Tomanovic andKavallieratos, 20026
Forewing vein m-cu coloured throughout(Figs. 33, 40, 46, 53, 61, 66, 72, 79)................. 76*
Forewing vein m-cu colourless throughout(Figs. 58, 85, 91, 112, 119); erased at itssecond part and colourless (Fig. 98);coloured at about first-third and colourless atthe rest part (Fig. 105)................................... 147
Antennae (20) 21–23-segmented;F2 without longitudinal placodesP. longicorne Marshall, 18917*
Antennae 15–20-segmented; F2 with 1 ormore longitudinal placodes............................ 88
Mesonotum with hairless areas (Figs. 41,47).................................................................. 98*
Mesonotum densely pubescent (Figs. 54, 62,67, 73, 80)...................................................... 109
Antennae 15–16-segmented; F1 brown;proportion between length and width of F1about 4.0–4.5 P. abjectum (Haliday, 1833)
9* Antennae 18–20-segmented; F1 yellow;proportion between length and width of F1
about 5.0–5.5 P. flavinode (Haliday, 1833)
10 Dorsal outline of ovipositor sheath almoststraight (Figs. 57, 64, 70); tergite 1 coveredwith sparse long hairs along sides (Figs. 56,63, 69)...........................................................
1110*
Dorsal outline of ovipositor sheath concave(Figs. 76, 83); tergite 1 covered with longdense hairs along sides (Figs. 75, 82)............ 1311
Antennae 19-segmented; F1 yellow;proportion between length and width offorewing stigma about 4.0 (Fig. 53) P. bicolorMackauer, 1959
11* Antennae (15)16–17-segmented; F1 brown;proportion between length and width offorewing stigma 3.3–3.5 (Figs. 61 and66)..................................................................
1212
Forewing vein Rs subequal to distal abscissaof R1 (Fig. 61); proportion between length offorewing stigma and length of distal abscissaof R1 1.9–2.2 (Fig. 61); ovipositor sheathrounded (Fig. 64); proportion between lengthand width of F1 5.5–6.0; parasitoid ofMacrosiphum rosae P. rosaecola Stary, 196112*
Forewing vein Rs longer than distal abscissa ofR1 (Fig. 66); proportion between length offorewing stigma and length of distal abscissa ofR1 about 1.8 (Fig. 66); ovipositor sheathlanceolate (Fig. 70); proportion between lengthand width of F1 about 5.5; parasitoid of Myzuspersicae P. staryi Kavallieratos and Lykouressis,2000
13
Antennae 16–17-segmented; forewingmedian vein shorter than Rs vein (Fig. 72);propodeum covered with long sparse hairs onupper part (Fig. 74); ovipositor sheathrounded (Figs. 76 and 77) P. pubescens Stary,196113*
Antennae 17–18 (19)-segmented; forewingmedian vein longer than Rs vein (Fig. 79);propodeum covered with short, dense hairson upper part (Fig. 81); ovipositor sheathlanceolate (Fig. 83) P. volucre (Haliday, 1833)
ARTICLE IN PRESSN.G. Kavallieratos et al. / Zoologischer Anzeiger 243 (2005) 181–209 185
14
Face and propodeum densely haired(Figs. 84, 87, 90, 93)..................................... 1514*
Face and propodeum normally haired(Figs. 97, 100, 104, 107, 111, 114)................. 1615
Antennae 18–19-segmented; ovipositorsheath with one conical apical spine(Fig. 89); proportion between length andwidth of F1 about 5.0 P. megourae Stary,197115*
Antennae 20–21-segmented; ovipositor sheathwith two conical apical spines (Figs. 95 and 96);proportion between length and width of F16.0–6.3P. barbatum (Mackauer, 1967)
16 Antennae 20–21-segmented; forewing veinm-cu colourless proximally and effaceddistally (Fig. 98) or coloured at about firstthird and colourless at the rest part(Fig. 105); ovipositor sheath with oneconical apical spine (Figs. 102 and103)................................................................
1716*
Antennae 16–19 (20)-segmented; forewingvein m-cu colourless (Figs. 58, 112);ovipositor sheath with two conical apicalspines (Figs. 116 and 117)............................... 1817
Forewing vein m-cu colourless proximallyand effaced distally (Fig. 98); proportionbetween length and width of forewingstigma about 3.5 (Fig. 98); proportionbetween length of stigma and lengthof distal abscissa of R1 about 1.5(Fig. 98); proportion between lengthand width of F1 about 5.0; F2 with forelongitudinal placode P. uroleuconTomanovic and Kavallieratos, 2003
17* Forewing vein m-cu coloured at about firstthird and colourless at the rest part (Fig.105); proportion between length and width offorewing stigma about 4.0 (Fig. 105);proportion between length of stigma andlength of distal abscissa of R1 about 1.8 (Fig.105); proportion between length and width ofF1 about 6.0; F2 without longitudinalplacodes P. athenaeum Kavallieratos andLykouressis, 2000
18
F1 yellow P. yomenae Takada, 1968 18* F1 brown........................................................ 19 19 Antennae (17)18–19-segmented; median veinshort (Fig. 58); ovipositor sheath concave;parasitoid of Macrosiphoniella tanacetaria
P. absinthii Bignell, 1894
19* Antennae 16–17-segmented; median; longwith colourless the middle third (Fig. 120);ovipositor sheath very slightly concave(Fig. 124); parasitoid of Uroleucon
inulicola P. nonveilleri Tomanovic andKavallieratos, 2003
3.2. Parasitoid–aphid–plant associations
Abbreviations
SER—Serbia, MNG—Montenegro, FYRM—TheFormer Yugoslav Republic of Macedonia, BOS—Bosnia and Herzegovina, BUL—Bulgaria, GRE—Greece, TUR—Turkey.
The references in this section are abbreviated asfollows: Kavallieratos and Lykouressis (1999–2000):[K1999-2000]; Tomanovic and Kavallieratos (2002):[T2002]; Kavallieratos et al. (2003): [K2003]; Tomanovicet al. (2003a): [T2003]; Tomanovic et al. (2003b):[TK2003]; Kavallieratos et al. (2004): [K2004].
An asterisk states new information.
Genus Praon Haliday, 1833
Praon Haliday, 1833Achoristus Ratzeburg, 1852Aphidaria Provancher, 1886Parapraon Stary, 1983Praon abjectum (Haliday, 1833) (Figs. 39–44)Aphidius (Praon) abjectus Haliday, 1833Bracon (Achoristus) aphidiiformis Ratzeburg, 1852Host records: Aphis craccivora on Robinia pseudacacia
(SER, BOS) [K2004]; Aphis elegantula Szelegiewicz onEpilobium sp. (MNG) [K2004]; Aphis fabae Scopoli onPhylladelphus coronarius (SER) [K2004], Pisum sativum
(SER) [K2004], Spiraea sp. (BUL) [K2004]; Aphisgrossulariae Kaltenbach on Epilobium hirsutum (SER)[K2004]; Aphis sambuci L. on Sambucus nigra (SER);Aphis spiraephaga Muller on Spiraea media (SER)[K2004], Spiraea sp. (SER) [K2004]; Aphis sp. onunknown plant (BUL) [K2004]; Brachycaudus helichrysi(Kaltenbach) on Tanacetum macrophyllum (SER)[K2004]; Liosomaphis berberidis Kaltenbach on Berberis
vulgaris (SER) [K2004], Mahonia aquifolium (SER)[K2004]; Longicaudus trirhodus (Walker) on Thalictrum
aquilegifolium (SER, MNG) [K2004].Praon absinthii Bignell, 1894 (Figs. 58–60)Host records: Macrosiphoniella tanacetaria (Kalten-
bach) on Tanacetum vulgare (SER) [K2004].Praon athenaeum Kavallieratos and Lykouressis, 2000
(Figs. 104–110)Host records: Hyperomyzus lactucae (L.) on Sonchus
oleraceus L. (GRE) [K1999-2000].Praon barbatum Mackauer, 1967 (Figs. 90–96)Praon barbatum Mackauer in: Mackauer and Finlay-
son (1967)Host records: Acyrthosiphon pisum (Harris) on Med-
icago sativa (SER, MNG) [K2004], Pisum sativum (SER)[K2004], Trifolium pratense (SER) [K2004].
Praon bicolor Mackauer, 1959 (Figs. 52–57)Host records: Eulachnus rileyi (Williams) on Pinus
nigra (SER) [K2004]; Schizolachnus pineti (Fabricius) onPinus nigra (SER) [K2004], P. sylvestris (SER) [K2004].
Praon exsoletum (Nees, 1811) (Figs. 8–14)

ARTICLE IN PRESSN.G. Kavallieratos et al. / Zoologischer Anzeiger 243 (2005) 181–209186
Praon palitans Muesebeck, 1956Host records: Therioaphis trifolii (Monell) on Medicago
sativa (SER, GRE) [K2004], unknown plant (BUL)*.Praon flavinode (Haliday, 1833) (Figs. 45–51)Aphidius (Praon) flavinodis Haliday, 1833Host records: Eucallipterus tiliae (L.) on Tilia sp.
(SER) [K2004]; Calaphis flava (Mordvilko) on Betula sp.(SER) [K2004] (Figs. 1–51).
Praon gallicum Stary, 1971 (Figs. 15–21)Host records: Metopolophium dirhodum (Walker) on
Avena sativa (SER) [K2004], Triticum aestivum (SER)[K2004], Zea mays (SER) [K2004], Hordeum vulgare
(SER) [K2004], H. murinum (SER) [K2004]; Pleotricho-phorus glandulosus (Kaltenbach) on Artemisia vulgaris
(SER) [K2004]; Rhopalosiphum padi (L.) on Zea mays
(SER) [K2004]; Schizaphis graminum Rondani onTriticum aestivum (SER) [K2004]; Sitobion avenae(Fabricius) on Triticum aestivum (SER) [K2004], Zea
mays (SER) [K2004].Praon longicorne Marshall, 1891 (Figs. 32–38)Praon grossum Stary, 1971Host records: Amphorophora spp. on Rubus hirsutus
(SER, MNG) [K2004]; Macrosiphum cholodkovskyi(Mordvilko) on Filipendula ulmaria (MNG) [K2004];Macrosiphum funestum (Macchiati) on Rubus hirsuta
(SER) [K2004], (MNG*); Macrosiphum oredonensisRemaudiere on Lonicera nigra (MNG) [K2004]; Macro-siphum prenanthidis (Borner) on Cicerbita pancicii (MNG)[K2004].
Praon megourae Stary, 1971 (Figs. 84–89)Host records: Megoura viciae Buckton on Lathyrus
pratensis (MNG) [K2004].Praon necans Mackauer, 1959 (Figs. 22–26)Host records: Rhopalosiphum nymphaeae L. on Ra-
nunculus aquatilis (SER) [K2004].Praon nonveilleri Tomanovic and Kavallieratos, 2003
(Figs. 118–124)Praon nonveilleri Tomanovic and Kavallieratos in:
Tomanovic et al. (2003b)Host records: Uroleucon inulicola (Hille Ris Lambers)
on Inula ensifolia (SER) [TK2004], Inula helenium (SER)[TK2004].
Praon orpheusi Kavallieratos, Athanassiou and Toma-
novic, 2003 (Figs. 1–7)Host records: Macrosiphum rosae (L.) on Rosa
sempervirens (GRE) [K2003], R. caesia (BUL) [K2003].Praon pubescens Stary, 1961 (Figs. 71–77)Host records: Nasonovia ribisnigri (Mosley) on Crepis
viscidula (SER), [K2004] Hieracium sp. (SER, MNG)[K2004]; Nasonovia spp. on Hieracium spp. (SER,MNG) [K2004].
Praon retusae Tomanovic and Kavallieratos, 2002
(Figs. 27–31)Host records: Acyrthosiphon sp. on Salix retusa
(MNG) [T2002].Praon rosaecola Stary, 1961 (Figs. 61–64)
Host records: Macrosiphum rosae L. on Rosa sp.(SER) [K2004].
Praon staryi Kavallieratos and Lykouressis, 2000
(Figs. 65–70)Host records: Myzus persicae (Sulzer) on Nicotiana
tabacum (GRE) [K1999-2000].Praon volucre (Haliday, 1833) (Figs. 78–83)Aphidius (Praon) volucris Haliday, 1833Blacus angulator Nees, 1834Aphidius aphidivorus Ratzeburg, 1844Praon pruni Ivanov, 1925Host records: Acyrthosiphon lactucae (Passerini) on
Lactuca serriola (SER, GRE) [K2004]; Acyrthosiphonmalvae (Mosley) on Geranium sp. (SER, MNG) [K2004];Acyrthosiphon pisum (Harris) on Lathyrus cicera (GRE)[K2004], Medicago arabica (GRE) [K2004], Medicago
sativa (SER, GRE) [K2004], Pisum sativum (SER)[K2004], Vicia narbonensis (GRE) [K2004], V. sativa
ssp amphicarpa (GRE) [K2004]; Amphorophora rubi(Kaltenbach) on Rubus caesius (SER) [K2004], R.
fruticosus (GRE) [K2004]; Aphis craccivora Koch onIpomoea batatas (GRE) [K2004], Ipomoea sp. (GRE)[K2004], Melilotus sulcata (GRE) [K2004]; Aphis crepi-dis (Borner) on Crepis biennis (SER) [K2004]; Aphisfabae Scopoli on Amaranthus retroflexus (SER) [K2004],Beta vulgaris (SER) [K2004], Conium maculatum (TUR)[K2004], Galium aparine (GRE) [K2004], Papaver
somniferum (SER) [K2004], Phaseolus vulgaris (SER)[K2004], Pittosporum tobira (GRE, TUR) [K2004],Rumex crispus (GRE, TUR) [K2004]; R. hydrolapathum
(BUL, TUR) [K2004], Vicia faba (GRE) [K2004]; A.fabae cirsiiacanthoidis Scopoli on Carduus australis
(GRE) [K2004], Carlina corymbosa (BUL, GRE)[K2004], Cirsium creticum (TUR) [K2004], Onopordum
illyricum (TUR) [K2004], Silybum marianum (GRE)[K2004]; Aphis fabae solanella Theobald on Solanum
nigrum (GRE) [K2004]; Aphis gosypii Glover on Citrus
aurantium (GRE), C. deliciosa (GRE) [K2004], Vibur-
num tinus (GRE) [K2004]; Aphis spiraephaga (Muller)on Spiraea sp. (SER) [K2004]; Aphis umbrella (Borner)on Malva neglecta (GRE) [K2004]; Aphis viticis Ferrarion Vitex agnus-castus (GRE) [K2004]; Aulacorthumsolani (Kaltenbach) on Cirsium vulgare (GRE) [K2004],Citrus aurantium (GRE) [K2004], Malva neglecta (GRE)[K2004], Sonchus oleraceus (GRE) [K2004], Urtica
pilulifera (GRE) [K2004], U. urens (GRE) [K2004];Brachycaudus cardui (L.) on Carduus pycnocephalus
(GRE) [K2004]; Brachycaudus helichrysi (Kaltenbach)on Carduus crispus (GRE) [K2004], Chrysanthemum
coronarium (GRE) [K2004], Helianthus annuus (SER*);Onopordum illyricum (GRE) [K2004], Prunus cerasifera
(SER) [K2004], Brachycaudus helichrysi var. warei(Theobald) on Vicia sativa (GRE) [K2004]; Brevicorynebrassicae on Brassica oleracea (SER) [K2004]; Capito-phorus elaeagni (del Guercio) on Carduus crispus (GRE)[K2004]; Corylobium avellanae (Schrank) on Corylus

ARTICLE IN PRESS
Figs. 1–7. Praon orpheusi Kavallieratos, Athanassiou and Tomanovic, 2003, female. 1: Head, anterior aspect, 2: Forewing, 3:
Mesonotum, dorsal aspect, 4: Propodeum (arrow), dorsal aspect 5: Tergite 1, dorsal aspect, 6: Ovipositor sheath, lateral aspect, 7:
Apex of ovipositor sheath, lateral aspect.
N.G. Kavallieratos et al. / Zoologischer Anzeiger 243 (2005) 181–209 187

ARTICLE IN PRESS
Figs. 8–14. Praon exsoletum (Nees, 1811), female. 8: Head, anterior aspect, 9: Forewing, 10: Mesonotum, dorsal aspect, 11:
Propodeum (arrow), dorsal aspect, 12: Tergite 1, dorsal aspect, 13: Ovipositor sheath, lateral aspect, 14: Apex of ovipositor sheath,
lateral aspect.
N.G. Kavallieratos et al. / Zoologischer Anzeiger 243 (2005) 181–209188

ARTICLE IN PRESS
Figs. 15–21. Praon gallicum Stary, 1971, female. 15: Head, anterior aspect, 16: Forewing, 17: Mesonotum, dorsal aspect, 18:
Propodeum (arrow), dorsal aspect, 19: Tergite 1, dorsal aspect, 20: Ovipositor sheath, lateral aspect, 21: Apex of ovipositor sheath,
lateral aspect.
N.G. Kavallieratos et al. / Zoologischer Anzeiger 243 (2005) 181–209 189
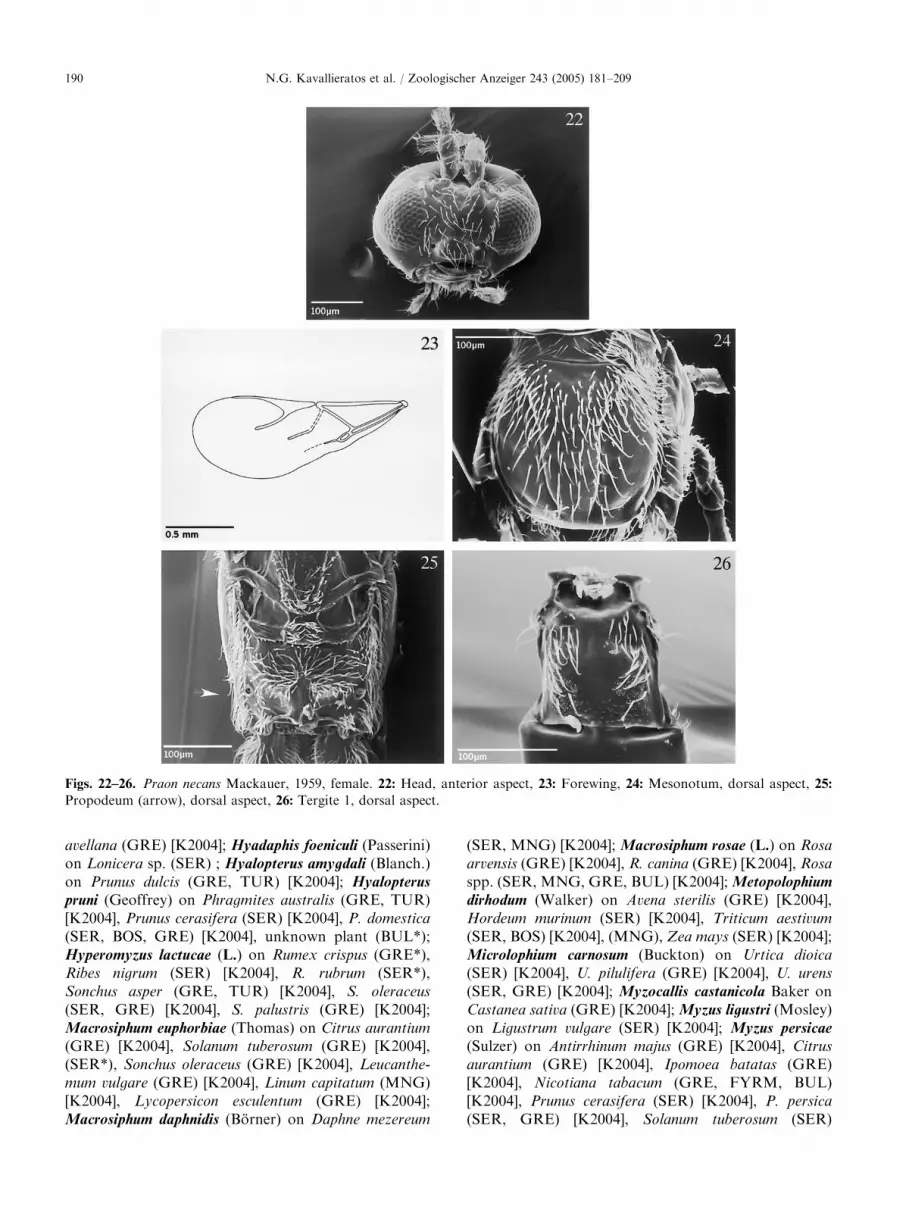
ARTICLE IN PRESS
Figs. 22–26. Praon necans Mackauer, 1959, female. 22: Head, anterior aspect, 23: Forewing, 24: Mesonotum, dorsal aspect, 25:
Propodeum (arrow), dorsal aspect, 26: Tergite 1, dorsal aspect.
N.G. Kavallieratos et al. / Zoologischer Anzeiger 243 (2005) 181–209190
avellana (GRE) [K2004]; Hyadaphis foeniculi (Passerini)on Lonicera sp. (SER) ; Hyalopterus amygdali (Blanch.)on Prunus dulcis (GRE, TUR) [K2004]; Hyalopteruspruni (Geoffrey) on Phragmites australis (GRE, TUR)[K2004], Prunus cerasifera (SER) [K2004], P. domestica
(SER, BOS, GRE) [K2004], unknown plant (BUL*);Hyperomyzus lactucae (L.) on Rumex crispus (GRE*),Ribes nigrum (SER) [K2004], R. rubrum (SER*),Sonchus asper (GRE, TUR) [K2004], S. oleraceus
(SER, GRE) [K2004], S. palustris (GRE) [K2004];Macrosiphum euphorbiae (Thomas) on Citrus aurantium
(GRE) [K2004], Solanum tuberosum (GRE) [K2004],(SER*), Sonchus oleraceus (GRE) [K2004], Leucanthe-
mum vulgare (GRE) [K2004], Linum capitatum (MNG)[K2004], Lycopersicon esculentum (GRE) [K2004];Macrosiphum daphnidis (Borner) on Daphne mezereum
(SER, MNG) [K2004]; Macrosiphum rosae (L.) on Rosa
arvensis (GRE) [K2004], R. canina (GRE) [K2004], Rosa
spp. (SER, MNG, GRE, BUL) [K2004]; Metopolophiumdirhodum (Walker) on Avena sterilis (GRE) [K2004],Hordeum murinum (SER) [K2004], Triticum aestivum
(SER, BOS) [K2004], (MNG), Zea mays (SER) [K2004];Microlophium carnosum (Buckton) on Urtica dioica
(SER) [K2004], U. pilulifera (GRE) [K2004], U. urens
(SER, GRE) [K2004]; Myzocallis castanicola Baker onCastanea sativa (GRE) [K2004]; Myzus ligustri (Mosley)on Ligustrum vulgare (SER) [K2004]; Myzus persicae(Sulzer) on Antirrhinum majus (GRE) [K2004], Citrus
aurantium (GRE) [K2004], Ipomoea batatas (GRE)[K2004], Nicotiana tabacum (GRE, FYRM, BUL)[K2004], Prunus cerasifera (SER) [K2004], P. persica
(SER, GRE) [K2004], Solanum tuberosum (SER)

ARTICLE IN PRESS
Figs. 27–31. Praon retusae Tomanovic and Kavallieratos, 2002, female. 27: Forewing, 28: Mesonotum, dorsal aspect, 29:
Propodeum, dorsal aspect, 30: Tergite 1, dorsal aspect, 31: Ovipositor sheath, lateral aspect.
N.G. Kavallieratos et al. / Zoologischer Anzeiger 243 (2005) 181–209 191
[K2004], Urtica urens (GRE) [K2004]; Myzusvarians Davidson on Clematis vitalba (SER, GRE)[K2004]; Ovatus crataegarius (Walker) on Mentha
spicata (GRE) [K2004]; Phorodon humuli (Schrank)on Prunus cerasifera (SER) [K2004]; Rhopalomyzuslonicerae (Siebold) on Lonicera sp. (SER) [K2004];Rhopalosiphum padi (L.) on Hordeum murinum (GRE)[K2004], Piptatherum miliaceum (GRE) [K2004],Triticum durum (GRE) [K2004], Zea mays (SER)[K2004]; Schizaphis graminum (Rondani) on Avena
sativa (SER) [K2004]; Sitobion avenae (Fabricius) onBromus nemoralis (SER) [K2004], Dactylis glomerarata
(SER*), Triticum aestivum (SER, MNG, BOS,GRE, BUL) [K2004], T. durum (GRE) [K2004];Sitobion fragariae (Walker) on Bromus madritensis
(GRE) [K2004], Dactylis glomerata (SER*), Hordeum
murinum (SER) [K2004]; Sitobion sp. on unknownplant (BUL*); Toxoptera auranti (Boyer de Fonsco-lombe) on Citrus aurantium (GRE) [K2004]; Uroleuconaeneum (Hille Ris Lambers) on Carlina corymbosa
(GRE, TUR) [K2004]; Uroleucon cichorii (Koch) onCichorium intybus (SER) [K2004]; Uroleucon jaceae (L.)on Carthamus dentatus (GRE*), Carthamus lanatus
(GRE*); Uroleucon sonchi (L.) on Sonchus oleraceus
(GRE, TUR) [K2004], S. palustris (GRE, TUR) [K2004](Figs. 52–103).
Praon uroleucon Tomanovic and Kavallieratos, 2003
(Figs. 97–103)Praon uroleucon Tomanovic and Kavallieratos in:
Tomanovic et al. (2003a)

ARTICLE IN PRESS
Figs. 32–38. Praon longicorne Marshall, 1891, female. 32: Head, anterior aspect, 33: Forewing, 34: Mesonotum, dorsal aspect, 35:
Propodeum (arrow), dorsal aspect, 36: Tergite 1, dorsal aspect, 37: Ovipositor sheath, lateral aspect, 38: Apex of ovipositor sheath,
lateral aspect.
N.G. Kavallieratos et al. / Zoologischer Anzeiger 243 (2005) 181–209192
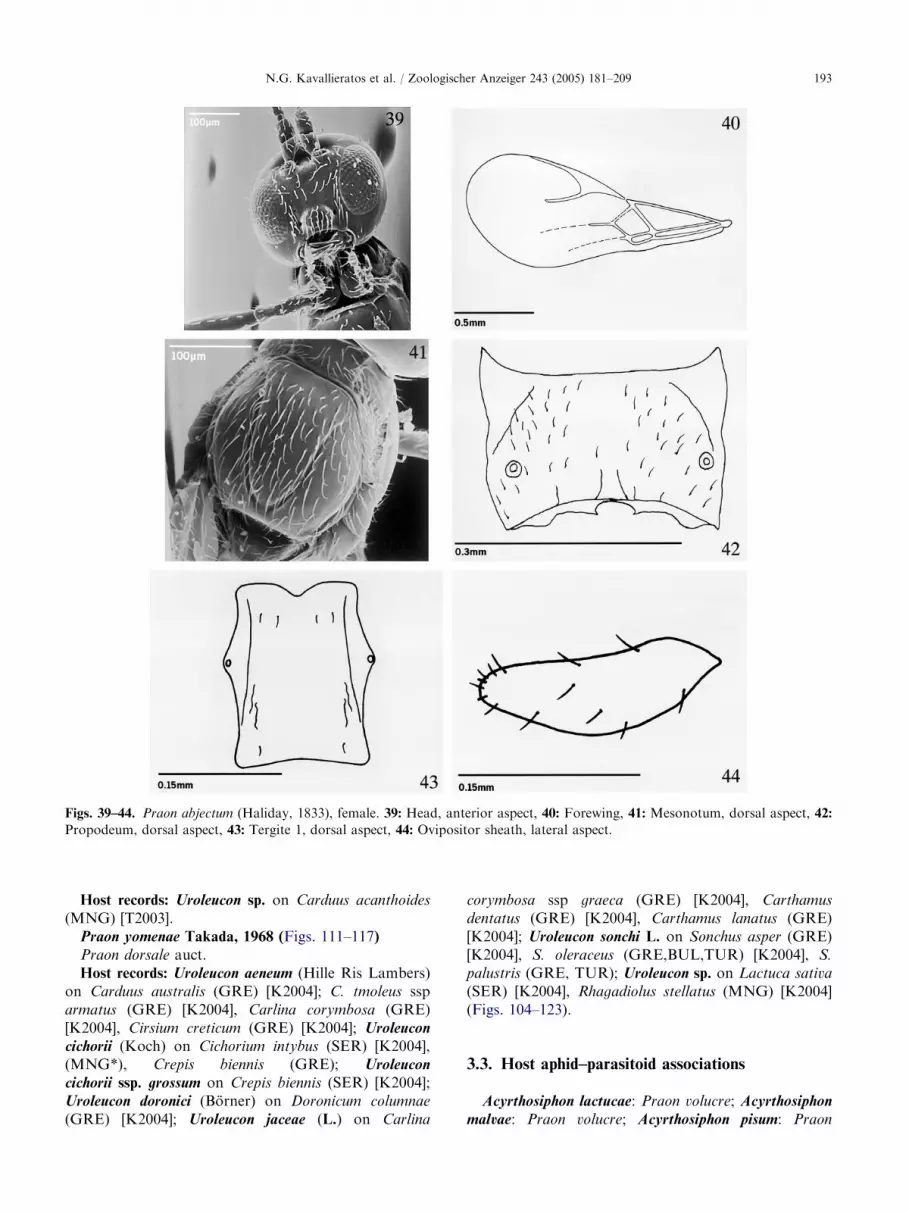
ARTICLE IN PRESS
Figs. 39–44. Praon abjectum (Haliday, 1833), female. 39: Head, anterior aspect, 40: Forewing, 41: Mesonotum, dorsal aspect, 42:
Propodeum, dorsal aspect, 43: Tergite 1, dorsal aspect, 44: Ovipositor sheath, lateral aspect.
N.G. Kavallieratos et al. / Zoologischer Anzeiger 243 (2005) 181–209 193
Host records: Uroleucon sp. on Carduus acanthoides
(MNG) [T2003].Praon yomenae Takada, 1968 (Figs. 111–117)Praon dorsale auct.Host records: Uroleucon aeneum (Hille Ris Lambers)
on Carduus australis (GRE) [K2004]; C. tmoleus ssparmatus (GRE) [K2004], Carlina corymbosa (GRE)[K2004], Cirsium creticum (GRE) [K2004]; Uroleuconcichorii (Koch) on Cichorium intybus (SER) [K2004],(MNG*), Crepis biennis (GRE); Uroleuconcichorii ssp. grossum on Crepis biennis (SER) [K2004];Uroleucon doronici (Borner) on Doronicum columnae
(GRE) [K2004]; Uroleucon jaceae (L.) on Carlina
corymbosa ssp graeca (GRE) [K2004], Carthamus
dentatus (GRE) [K2004], Carthamus lanatus (GRE)[K2004]; Uroleucon sonchi L. on Sonchus asper (GRE)[K2004], S. oleraceus (GRE,BUL,TUR) [K2004], S.
palustris (GRE, TUR); Uroleucon sp. on Lactuca sativa
(SER) [K2004], Rhagadiolus stellatus (MNG) [K2004](Figs. 104–123).
3.3. Host aphid–parasitoid associations
Acyrthosiphon lactucae: Praon volucre; Acyrthosiphonmalvae: Praon volucre; Acyrthosiphon pisum: Praon

ARTICLE IN PRESS
Figs. 45–51. Praon flavinode (Haliday, 1833), female. 45: Head, anterior aspect, 46: Forewing, 47: Mesonotum, dorsal aspect, 48:
Propodeum (arrow), dorsal aspect, 49: Tergite 1, dorsal aspect, 50: Ovipositor sheath, lateral aspect, 51: Apex of ovipositor sheath,
lateral aspect.
N.G. Kavallieratos et al. / Zoologischer Anzeiger 243 (2005) 181–209194

ARTICLE IN PRESSN.G. Kavallieratos et al. / Zoologischer Anzeiger 243 (2005) 181–209 195
volucre; Acyrthosiphon sp.: Praon retusae; Amphorophorarubi: Praon volucre; Amphorophora spp.: Praon long-
icorne; Aphis craccivora: Praon abjectum, Praon volucre;Aphis crepidis: Praon volucre; Aphis epilobii: Praon
abjectum; Aphis fabae: Praon abjectum, Praon volucre;Aphis fabae cirsiiacanthoidis: Praon volucre; Aphis fabaesolanella: Praon volucre; Aphis gossypii: Praon volucre;Aphis grossulariae: Praon abjectum; Aphis sambuci:Praon abjectum; Aphis spiraephaga: Praon abjectum,Praon volucre; Aphis umbrella: Praon volucre; Aphisviticis: Praon volucre; Aulacorthum solani: Praon volucre;Brachycaudus cardui: Praon volucre; Brachycaudus heli-chrysi; Praon abjectum, Praon volucre; Brachycaudushelichrysi var. warei: Praon volucre; Brachycaudus sp.:
Figs. 52–57. Praon bicolor Mackauer, 1959, female. 52: Head, ante
Propodeum (arrow), dorsal aspect, 56: Tergite 1, dorsal aspect, 57:
Praon volucre; Callaphis flava: Praon flavinode; Capito-phorus elaeagni: Praon volucre; Corylobium avellanae:Praon volucre; Eucallipterus tiliae: Praon flavinode;Eulachnus rileyi: Praon bicolor, Hyadaphis foeniculi:Praon volucre; Hyalopterus amygdali: Praon volucre;Hyalopterus pruni: Praon volucre; Hyperomyzus lactucae:Praon athenaeum, Praon volucre; Liosomaphis berberidis:Praon abjectum; Longicaudus trirhodus: Praon abjectum;Macrosiphoniella tanacetaria: Praon absinthii; Macro-siphum cholodkovskyi: Praon longicorne; Macrosiphumeuphorbiae: Praon volucre; Macrosiphum daphnidis:Praon volucre; Macrosiphum funestum: Praon longicorne;Macrosiphum oredonense: Praon longicorne; Macrosi-phum prenanthidis: Praon longicorne; Macrosiphum
rior aspect, 53: Forewing, 54: Mesonotum, dorsal aspect, 55:
Ovipositor sheath, lateral aspect.

ARTICLE IN PRESS
Figs. 58–64. Praon absinthii Bignell, 1894, female. 58: Forewing, 59: Mesonotum, dorsal aspect, 60: Propodeum, dorsal aspect.
Praon rosaecola Stary, 1961, female 61: Forewing, 62: Mesonotum, dorsal aspect, 63: Tergite 1, dorsal aspect, 64: Ovipositor sheath,
lateral aspect.
N.G. Kavallieratos et al. / Zoologischer Anzeiger 243 (2005) 181–209196

ARTICLE IN PRESS
Figs. 65–70. Praon staryi Kavallieratos and Lykouressis, 2000, female. 65: Head, anterior aspect. 66: Forewing, 67: Mesonotum,
dorsal aspect, 68: Propodeum (arrow), dorsal aspect, 69: Tergite 1, dorsal aspect, 70: Ovipositor sheath, lateral aspect.
N.G. Kavallieratos et al. / Zoologischer Anzeiger 243 (2005) 181–209 197
rosae: Praon orpheusi, Praon rosaecola, Praon volucre;Megoura viciae: Praon megourae; Metopolophium dirho-dum: Praon gallicum, Praon volucre; Microlophium
carnosum: Praon volucre; Myzocallis castanicola: Praon
volucre; Myzus ligustri: Praon volucre; Myzus nicotianae:Praon staryi; Myzus persicae: Praon volucre; Myzus

ARTICLE IN PRESS
Figs. 71–77. Praon pubescens Stary, 1961, female. 71: Head, anterior aspect, 72: Forewing, 73: Mesonotum, dorsal aspect, 74:
Propodeum (arrow), dorsal aspect, 75: Tergite 1, dorsal aspect, 76: Ovipositor sheath, lateral aspect, 77: Apex of ovipositor sheath,
lateral aspect.
N.G. Kavallieratos et al. / Zoologischer Anzeiger 243 (2005) 181–209198

ARTICLE IN PRESS
Figs. 78–83. Praon volucre (Haliday, 1833), female. 78: Head, anterior aspect, 79: Forewing, 80: Mesonotum, dorsal aspect, 81:
Propodeum (arrow), dorsal aspect, 82: Tergite 1, dorsal aspect, 83: Ovipositor sheath, lateral aspect.
N.G. Kavallieratos et al. / Zoologischer Anzeiger 243 (2005) 181–209 199
varians: Praon volucre; Nasonovia ribisnigri: Praon
pubescens; Nasonovia spp.: Praon pubescens; Ovatuscrataegarius: Praon volucre; Phorodon humuli: Praon
volucre; Pleotrichophorus glandulosus: Praon gallicum;Rhopalomyzus lonicerae: Praon volucre; Rhopalosiphumnymphaeae: Praon necans; Rhopalosiphum padi: Praon
gallicum, Praon volucre; Schizaphis graminum: Praon
gallicum, Praon volucre; Schizolachnus pineti: Praon
bicolor; Sitobion avenae: Praon gallicum, Praon volucre;Sitobion fragariae: Praon volucre; Therioaphis trifolii:Praon exsoletum; Toxoptera aurantii: Praon volucre;Uroleucon aeneum: Praon volucre, Praon yomenae;
Uroleucon cichorii: Praon yomenae, Praon volucre;Uroleucon cichorii grossum: Praon yomenae; Uroleucondoronici: Praon yomenae; Uroleucon inulicola: Praon
nonveilleri; Uroleucon jaceae: Praon yomenae; Uroleuconsonchi: Praon volucre, Praon yomenae; Uroleucon spp.:Praon uroleucon.
3.4. Phylogenetic inference
The tree in Fig. 124 shows the reconstructedphylogenetic relationships among 20 Praon species from

ARTICLE IN PRESS
Figs. 84–89. Praon megourae Stary, 1971, female. 84: Head, anterior aspect, 85: Forewing, 86: Mesonotum, dorsal aspect, 87:
Propodeum, dorsal aspect, 88: Tergite 1, dorsal aspect, 89: Ovipositor sheath, lateral aspect.
N.G. Kavallieratos et al. / Zoologischer Anzeiger 243 (2005) 181–209200
southeastern Europe. A strict consensus tree by 50%majority rule is based on 31 equally most parsimonioustrees. The tree has a length of 31 steps, Consistencyindex (CI) 0.484, Retention index (RI) 0.680 andRescaled consistency index (RC) 0.329. As shown inFig. 124, Praon abjectum is the sister taxon to theremaining Praon species. This species has a lot ofplesiomorphic character states, especially in wing vena-tion and morphology of ovipositor sheath (Table 1). Wefound that ‘‘Parapraon’’ group (Fig. 124), that originallycomprised P. gallicum, P. necans and two Nearctic
species (Stary 1983) is monophyletic. Stary (1983) placedthese species in a separate genus Parapraon based on thereduced wing venation [the lack of recurrent (m-cu)vein]. Johnson (1987) in his revision of American Praon
pointed out the great variability of character states of m-cu vein and consequently synonymized Parapraon withPraon. Recently described P. retusae (Tomanovic andKavallieratos 2002) and P. exsoletum also belong to‘‘Parapraon’’ group. All species of the European‘‘Parapraon’’ group share the following synapomor-phies: absent or significantly reduced m-cu vein;

ARTICLE IN PRESS
Figs. 90–96. Praon barbatum Mackauer, 1967, female. 90: Head, anterior aspect, 91: Forewing, 92: Mesonotum, dorsal aspect, 93:
Propodeum (arrow), dorsal aspect, 94: Tergite 1, dorsal aspect, 95: Ovipositor sheath, lateral aspect, 96: Apex of ovipositor sheath,
lateral aspect.
N.G. Kavallieratos et al. / Zoologischer Anzeiger 243 (2005) 181–209 201

ARTICLE IN PRESS
Figs. 97–103. Praon uroleucon Tomanovic and Kavallieratos, 2003, female. 97: Head, anterior aspect, 98: Forewing, 99:
Mesonotum, dorsal aspect, 100: Propodeum (arrow), dorsal aspect, 101: Tergite 1, dorsal aspect, 102: Ovipositor sheath, lateral
aspect, 103: Apex of ovipositor sheath, lateral aspect.
N.G. Kavallieratos et al. / Zoologischer Anzeiger 243 (2005) 181–209202

ARTICLE IN PRESS
Figs. 104–110. Praon athenaeum Kavallieratos and Lykouressis, 2000, female. 104: Head, Anterior aspect, 105: Forewing, 106:
Mesonotum, dorsal aspect, 107: Propodeum (arrow), dorsal aspect, 108: Tergite 1, dorsal aspect, 109: Ovipositor sheath, lateral
aspect, 110: Apex of ovipositor sheath, lateral aspect.
N.G. Kavallieratos et al. / Zoologischer Anzeiger 243 (2005) 181–209 203

ARTICLE IN PRESS
Figs. 111–117. Praon yomenae Takada, 1968, female. 111: Head, anterior aspect, 112: Forewing, 113: Mesonotum, dorsal aspect,
114: Propodeum (arrow), dorsal aspect, 115: Tergite 1, dorsal aspect, 116: Ovipositor sheath, lateral aspect, 117: Apex of ovipositor
sheath, lateral aspect.
N.G. Kavallieratos et al. / Zoologischer Anzeiger 243 (2005) 181–209204

ARTICLE IN PRESS
Figs. 118–123. Praon nonveilleri Tomanovic and Kavallieratos, 2003, female. 118: Female head, anterior aspect, 119: Female
forewing, lateral aspect, 120: Female mesonotum, dorsal aspect, 121: Female propodeum (arrow), 122: Female metasomal tergum 1
(arrow), dorsal aspect, 123: Female ovipositor seath, lateral aspect.
N.G. Kavallieratos et al. / Zoologischer Anzeiger 243 (2005) 181–209 205
elongated ovipositor sheath; two conical apical spineson ovipositor sheath and large hairless area on laterallobes of mesonotum.
The species group closely related to ‘‘Parapraon’’ isthe ‘‘dorsale-yomenae’’ group, which originally com-prised Uroleucon aphid parasitoids (P. yomenae) andthe recently re-established species P. dorsale andP. longicorne (Tremblay and Pennacchio 1985; Trem-blay et al. 1986), P. uroleucon, P. nonveilleri andP. unitum (Tomanovic et al. 2003a, b; Mescheloffand Rosen 1988). Close relations, based on morphology,between ‘‘dorsale-yomenae’’ group and P. absinthi,
P. megourae and P. athenaeum were pointed out byMackauer (1959), Mackauer and Finlayson (1967),Stary (1971), Johnson (1987), Kavallieratos andLykouressis (1999–2000), Tomanovic et al. (2003a, b).Therefore, our parsimony analysis supported theirplacement inside the ‘‘dorsale-yomenae’’ group, butwith basal a position of P. megourae and P. athenaeum.Also, we concluded that the ‘‘dorsale-yomenae’’
group is paraphyletic, since P. longicorne belongs tothe P. rosaecola clade. Praon barbatum is sister to theremaining species of the ‘‘rosaecola’’ group. Also,rosaecola clade includes P. flavinode and P. bicolor,

ARTICLE IN PRESS
Figs. 124. Cladogram of 20 Praon species based on 11 characters and unweighted 50% majority-rule consensus tree parsimony
analysis of 31208 trees (31 the most parsimonious trees) [tree length 31; consistency index (CI) 0.484, retention index (RI) 0.680 and
rescaled consistency index (RC) 0.329]. E. persicae was used as outgroup. Frequency of occurence is shown.
N.G. Kavallieratos et al. / Zoologischer Anzeiger 243 (2005) 181–209206
which parasitize more basal host aphid lineages(Myzocallidinae and Lachninae). Both species sharethe following synapomorphies: elongated stigma (morethan 3.5 times as long as wide), forewing veinRs+M colorless, two conical apical spines on thetop of ovipositor sheath, tergite I elongated (morethan 1.5 time as long as wide at spiracles), flagellomere1 elongated (longer than 5.0 times as long aswide) and synplesiomorphies: fully sclerotized andcolored m-cu vein, ovipositor sheath straight dorsallyand short.
The tree in Fig. 125 shows the reconstructedphylogenetic relationships among Praon speciesbased on cladistic analysis by distance methods.
The tree has a length of 32 steps, CI 0.469, RI 0.660and RC 0.309. Also, this tree shows generallysimilar results to those of the strict consensus tree(Fig. 124), but with some diferent positions ofseparate species inside species groups. Main resultswhich we obtained by distance methods: basalposition for P. abjectum, as sister taxon to the remainingPraon species, basal position of P. megourae and P.
athenaeum for ‘‘Parapraon’’, ‘‘dorsale-yomenae’’ and‘‘rosaecola’’ group, close relation between ‘‘Parapraon’’
and ‘‘dorsale-yomenae’’ group with monophyletic posi-tion of ‘‘Parapraon’’ group (P. gallicum, P. necans, P.
retusae and P. exsoletum) and paraphyletic of ‘‘dorsale-
yomenae’’ group.

ARTICLE IN PRESS
Figs. 125. A cladistic analysis by distance methods of the internal phylogeny of 20 Praon species. E. persicae was used as outgroup.
Bootstrap values were calculated from 100 replicates. More than 50% of the bootstrap values are shown.
N.G. Kavallieratos et al. / Zoologischer Anzeiger 243 (2005) 181–209 207
Acknowledgements
The research was supported by the grant 1547 (TheMinistry of Science and Environmental Protection ofthe Republic of Serbia), A6007105 and S5007102 (GrantAgency, Academy of Sciences of the Czech Republic)and finally by the Entomology Institute ProjectZ5007907 (Academy of Sciences of the Czech Republic).
References
Belshaw, R., Quicke, D.L.J., 1997. A molecular phylogeny
of the Aphidiinae (Hymenoptera: Braconidae). Mol.
Phylogenet. Evol. 7, 281–293.
Bosch, R.V.D., Schlinger, E.I., Dietrick, E.J., Hagen, K.S.,
Holloway, J.K., 1959. The colonization and establishment
of imported parasites of the spotted alfalfa aphid in
California. J. Econ. Entomol. 52, 136–141.
Carver, M., 1984. The potential host ranges in Australia of
some imported aphid parasites (Hymenoptera: Ichneumo-
noidea: Aphidiidae). Entomophaga 29, 351–359.
Dahlsten, D.L., Zuparko, R.L., Hajek, A.E., Rowney, D.L.,
Dreistadt, S.H., 1999. Long-term sampling of Eucallipterus
tiliae (Homoptera: Drepanosiphidae) and associated nat-
ural enemies in a northern California site. Environ.
Entomol. 28, 845–850.
Dowton, M., Austin, A.D., Antolin, M.F., 1998. Evolutionary
relationships among the Braconidae (Hymenoptera: Ich-
neumonoidea) inferred from partial 16S rDNA gene
sequences. Insect Mol. Biol. 7, 129–150.

ARTICLE IN PRESSN.G. Kavallieratos et al. / Zoologischer Anzeiger 243 (2005) 181–209208
Eastop, V.F., van Emden, H.F., 1972. The insect material. In:
van Emden, H.F. (Ed.), Aphid Technology. Academic
Press, London, pp. 1–45.
Gardenfors, U., 1986. Taxonomic and biological revision of
Palaearctic Ephedrus (Haliday) (Hymenoptera, Braconidae,
Aphidiinae). Entomol. Scand. Suppl. 27, 1–95.
Hagvar, E.B., Hofsvang, T., 1991. Aphid parasitoids (Hyme-
noptera: Aphidiidae) biology, host selection and use in
biological control. Biocontrol News Inform. 12, 13–41.
Heimpel, G.E., Lundgren, J.G., 2000. Sex ratios of commer-
cially reared biological control agents. Biol. Cont. 19,
77–93.
Huber, J.T., Sharkey, M.J., 1993. Structure. In: Goulet, H.,
Huber, J.T. (Eds.), Hymenoptera of the World: An
Identification Guide to Families. Research Branch, Agri-
culture Canada, Ottawa, Ontario (Publication 1894/E),
pp. 13–59.
Johnson, J.W., 1987. A revision of the species of Praon
Haliday in North America, North of Mexico (Hymenop-
tera: Aphidiidae). Can. Entomol. 119, 999–1025.
Kambhampati, S., Volkl, W., Mackauer, M., 2000. Phyloge-
netic relationships among genera of Aphidiinae (Hyme-
noptera: Braconidae) based on DNA sequence of the
mitochondrial 16S rRNA gene. Syst. Entomol. 25,
437–445.
Kavallieratos, N.G., Lykouressis, D.P., 1999–2000. Two new
species of Praon Haliday (Hymenoptera: Aphidiidae) from
Greece. Entomol. Hell. 13, 5–12.
Kavallieratos, N.G., Lykouressis, D.P., Sarlis, G.P., Stathas,
G.J., Sanchis Segovia, A., Athanassiou, C.G., 2001. The
Aphidiinae (Hymenoptera: Ichneumonoidea: Braconidae)
of Greece. Phytoparasitica 29, 306–340.
Kavallieratos, N.G., Athanassiou, C.G., Stathas, G.J., Toma-
novic, Z., 2002a. Aphid parasitoids (Hymenoptera: Braco-
nidae: Aphidiinae) on citrus: seasonal abundance,
association with the species of host plant and sampling
indices. Phytoparasitica 30, 365–377.
Kavallieratos, N.G., Stathas, G.J., Athanassiou, C.G.,
Papadoulis, G.Th., 2002b. Dittrichia viscosa and Rubus
ulmifolius as reservoirs of aphid parasitoids (Hymenoptera:
Braconidae: Aphidiinae) and the role of certain coccinellid
species. Phytoparasitica 30, 231–242.
Kavallieratos, N.G., Athanassiou, C.G., Tomanovic, Z., 2003.
A new species and a key to Greek Praon Haliday
(Hymenoptera: Braconidae: Aphidiinae). Dtsch. Entomol.
Z. 50, 13–22.
Kavallieratos, N.G., Tomanovic, Z., Stary, P., Athanassiou,
C.G., Sarlis, G.P., Petrovic, O., Niketic, M.,
Anagnou-Veroniki, M., 2004. A survey of aphid parasitoids
(Hymenoptera: Braconidae: Aphidiinae) of Southeastern
Europe and their aphid—plant associations. Appl. Entomol.
Zool. 39, 527–563.
Mackauer, M., 1959. Die europaischen Arten der Gattungen
Praon und Areopraon (Hymenoptera: Braconidae: Aphidii-
nae). Beitr. Entomol. 9, 810–865.
Mackauer, M., 1963. A re-examination of C.F. Baker’s
collection of aphid parasites (Hymenoptera: Aphidiidae).
Can. Entomol. 95, 921–935.
Mackauer, M., Finlayson, T., 1967. The hymenopterous
parasites (Hymenoptera: Aphidiidae et Aphelinidae) of
the pea aphid in Eastern North America. Can. Entomol. 99,
1051–1082.
Mescheloff, E., Rosen, D., 1988. Biosystematic studies on the
Aphidiidae of Israel (Hymenoptera: Ichneumonoidea) 2.
The Genera Ephedrus and Praon. Israel J. Entomol. 22,
75–100.
Page, R.D.M., 1996. TREEVIEW: an application to display
phylogenetic trees on personal computers. Comput. Appl.
Biosci. 12, 357–358.
Pike, K.S., Stary, P., 1995. New species of parasitic wasps
attacking cereal aphids in the Pacific Northwest (Hyme-
noptera: Braconidae: Aphidiinae). J. Kansas Entomol. Soc.
68, 408–414.
Pike, K.S., Stary, P., Miller, T., Graf, G., Allison, D.,
Boydston, L., Miller, R., 2000. Aphid parasitoids (Hyme-
noptera: Braconidae: Aphidiinae) of Northwest USA. Proc.
Entomol. Soc. Wash. 102, 688–740.
Quicke, D.L.J., Belshaw, R., 1999. Incongruence between
morphological data sets: an example from the evolution of
endoparasitism among parasitic wasps (Hymenoptera:
Braconidae). Syst. Biol. 48, 436–454.
Quicke, D.L.J., van Achterberg, C., 1990. Phylogeny of the
subfamilies of the family Braconidae (Hymenoptera:
Ichneumonoidea). Zool. Verh. 258, 1–95.
Remaudiere, G., Remaudier, M., 1997. Catalogue des
Aphididae du Monde. INRA, Paris.
Sanchis, A., Latorre, A., Gonzalez-Candelas, F., Michelena,
J.M., 2000. An 18S rDNA based molecular phylogeny of
Aphidiinae (Hymenoptera: Braconidae). Mol. Phylogenet.
Evol. 14, 180–194.
Smith, C.F., 1944. The Aphidiinae of North America
(Braconidae: Hymenoptera). Ohio State Univ. Contr. Zool.
Entomol. 6, 1–154.
Smith, P.T., Kambhampati, S., Volkl, W., Mackauer, M.,
1999. A phylogeny of aphid parasitoids (Hymenoptera:
Braconidae: Aphidiinae) inferred from mitochondrial
NADH 1 dehydrogenase gene sequence. Mol. Phylogenet.
Evol. 11, 236–245.
Stary, P., 1961. Two new species of Praon Haliday from
Czechoslovakia (Hymenoptera: Aphidiidae). Acta Soc.
Entomol. Cechoslov. 58, 340–343.
Stary, P., 1966. Aphid Parasites of Czechoslovakia. Dr. W.
Junk, The Hague.
Stary, P., 1970. Biology of Aphid Parasites (Hymenoptera:
Aphidiidae) with Respect to Integrated Control. Dr. W.
Junk, The Hague.
Stary, P., 1971. New aphid parasites from Central Europe
(Hymenoptera: Aphidiidae). Acta Entomol. Bohemoslov.
68, 310–318.
Stary, P., 1976a. External female genitalia of the Aphidiidae
(Hymenoptera). Acta Entomol. Bohemoslov. 73,
102–112.
Stary, P., 1976b. Aphid Parasites (Hymenoptera: Aphidiidae)
of the Mediterranean Area. Dr. W. Junk, The Hague.
Stary, P., 1981. On the strategy, tactics and trends
of host specificity evolution in aphid parasitoids (Hyme-
noptera: Aphidiidae). Acta Entomol. Bohemoslov. 78,
65–75.
Stary, P., 1983. Parapraon, a new genus of the Aphidiidae
(Hymenoptera). Acta Entomol. Bohemoslov. 80, 206–209.

ARTICLE IN PRESSN.G. Kavallieratos et al. / Zoologischer Anzeiger 243 (2005) 181–209 209
Stary, P., 1995. The Aphidiidae of Chile (Hymenoptera:
Ichneumonoidea: Aphidiidae). Dtsch. Entomol. Z. 1,
113–138.
Stary, P., Schlinger, E., 1967. A Revision of the Far East Asian
Aphidiidae (Hymenoptera). Dr. W. Junk, The Hague.
Stary, P., Remaudiere, G., Leclant, F., 1971. Les Aphidiidae
(Hymenoptera) de France et leurs hotes (Homoptera:
Aphididae). Entomophaga, Mem. H. S. 5, 1–74.
Swofford, D.L., 2003. PAUP*. Phylogenetic Analysis Using
Parsimony (*and Other Methods). Version 4 (10b). Sinauer
Associates, Sunderland, MA.
Takada, H., 1968. Aphidiidae of Japan (Hymenoptera). Ins.
Mats. 30, 67–124.
Tomanovic, Z., Kavallieratos, N.G., 2002. Two new aphidiine
wasps (Hymenoptera: Braconidae: Aphidiinae) from the
southeastern Europe. Reichenbachia 34, 341–345.
Tomanovic, Z., Brajkovic, M., Krunic, M., 1998. A checklist
of aphid parasitoids (Hymenoptera: Aphidiidae) in Yugo-
slavia. Acta Entomol. Serbica 3, 95–106.
Tomanovic, Z., Kavallieratos, N.G., Athanassiou, C.G.,
Petrovic, O., 2003a. A new Praon species (Hymenoptera:
Braconidae: Aphidiinae) of the Uroleucon parasitoid com-
plex from the Mediterranean area. Phytoparasitica 31,
19–26.
Tomanovic, Z., Kavallieratos, N.G., Athanassiou, A., Stani-
savljevic, L.Z., 2003b. A review of the West Palaearctic
aphidiines (Hymenoptera: Braconidae: Aphidiinae) para-
sitic on Uroleucon spp. with a description of a new species.
Ann. Soc. Entomol. Fr. 39, 343–353.
Tremblay, E., Pennacchio, F., 1985. Taxonomic status of some
species of the genus Praon Haliday (Hymenoptera:
Braconidae: Aphidiinae). Boll. Lab. Entomol. Agr. ‘‘Filip-
po Silvestri’’ 42, 143–147.
Tremblay, E., Pennacchio, F., Matrone, L., Piccolo, D., 1986.
Redescription of Praon dorsale (Haliday, 1833) and
resurrection of Praon longicorne Marshall, 1891: a statis-
tical analysis of morphological data (Hymenoptera: Bra-
conidae). Entomotaxonomia 8, 31–42.
Recommended










![Defensive microbial symbionts in Hymenoptera [2014]](https://img.pdfslide.net/doc/110x75/63323b944e01430403009303/defensive-microbial-symbionts-in-hymenoptera-2014.jpg)






