
doi: 10.1152/ajprenal.00255.2012305:F216-F226, 2013. First published 1 May 2013;Am J Physiol Renal Physiol
Mariliza V. Rodrigues, José Eduardo Krieger and Adriana C. C. GirardiBruna H. Inoue, Daniel F. Arruda-Junior, Luciene C. G. Campos, Ana Luiza T. Barreto,endocytic machinery in the renal proximal tubulewith lower expression of critical components of the apical Progression of microalbuminuria in SHR is associated
You might find this additional info useful...
60 articles, 23 of which you can access for free at: This article citeshttp://ajprenal.physiology.org/content/305/2/F216.full#ref-list-1
including high resolution figures, can be found at: Updated information and serviceshttp://ajprenal.physiology.org/content/305/2/F216.full
found at: can beAmerican Journal of Physiology - Renal Physiology about Additional material and information
http://www.the-aps.org/publications/ajprenal
This information is current as of July 16, 2013.
1522-1466. Visit our website at http://www.the-aps.org/. 9650 Rockville Pike, Bethesda MD 20814-3991. Copyright © 2013 the American Physiological Society. ESSN: volume and composition. It is published 24 times a year (twice monthly) by the American Physiological Society,relating to the kidney, urinary tract, and their respective cells and vasculature, as well as to the control of body fluid
publishes original manuscripts on a broad range of subjectsAmerican Journal of Physiology - Renal Physiology
by Adriana G
irardi on July 16, 2013http://ajprenal.physiology.org/
Dow
nloaded from

Progression of microalbuminuria in SHR is associated with lower expressionof critical components of the apical endocytic machinery in the renalproximal tubule
Bruna H. Inoue, Daniel F. Arruda-Junior, Luciene C. G. Campos, Ana Luiza T. Barreto,Mariliza V. Rodrigues, José Eduardo Krieger, and Adriana C. C. GirardiHeart Institute (InCor), University of São Paulo Medical School, São Paulo, Brazil
Submitted 3 May 2012; accepted in final form 26 April 2013
Inoue BH, Arruda-Junior DF, Campos LC, Barreto AL, Ro-drigues MV, Krieger JE, Girardi AC. Progression of microalbu-minuria in SHR is associated with lower expression of critical com-ponents of the apical endocytic machinery in the renal proximaltubule. Am J Physiol Renal Physiol 305: F216–F226, 2013. Firstpublished May 1, 2013; doi:10.1152/ajprenal.00255.2012.—Cumula-tive epidemiological evidence indicates that the presence of mi-croalbuminuria predicts a higher frequency of cardiovascular events,peripheral disease, and mortality in essential hypertension. Microalbu-minuria may arise from increased glomerular permeability and/orreduced proximal tubular reabsorption of albumin by receptor-medi-ated endocytosis. This study aimed to evaluate the temporal pattern ofurinary protein excretion and to test the hypothesis that progressionof microalbuminuria is associated with decreased protein expressionof critical components of the endocytic apparatus in the renal proxi-mal tubule of spontaneously hypertensive rats (SHR). We found thaturinary albumin excretion increased progressively with blood pressurein SHR from 6 to 21 wk of age. In addition, SDS-PAGE analysis ofurinary proteins showed that microalbuminuric SHR virtually ex-creted proteins of the size of albumin or smaller (�70 kDa), typical oftubular proteinuria. Moreover, the protein abundance of the endocyticreceptors megalin and cubilin as well as of the chloride channel ClC-5progressively decreased in the renal cortex of SHR from 6 to 21 wkof age. Expression of the vacuolar H�-ATPase B2 subunit was alsoreduced in the renal cortex of 21-wk-old compared with both 6- and14-wk-old SHR. Collectively, our study suggests that enhanced uri-nary protein excretion, especially of albumin, may be due, at least inpart, to lower expression of key components of the apical endocyticapparatus in the renal proximal tubule. Finally, one may speculate thatdysfunction of the apical endocytic pathway in the renal proximaltubule may contribute to the development of microalbuminuria inessential hypertension.
receptor-mediated endocytosis; megalin; cubilin; ClC-5; vacuolarH�-ATPase; hypertension
THE IDENTIFICATION AND QUANTITATION of proteinuria, particularly ofalbumin, are of great importance in the initial diagnosis and atsubsequent follow-up in kidney disease patients (40, 48, 60).Massive and sustained proteinuria is an unequivocal sign ofestablished nephropathy while detection of small increases in thenormal amount of urinary albumin excretion, known as mi-croalbuminuria (30–300 mg/24 h or 20–200 �g/min) (40, 60), isindicative of renal dysfunction and renal disease progression.
Increased urinary albumin excretion results from enhancedglomerular filtration and/or decreased reabsorption of this mac-
romolecule in the renal proximal tubule (14). Under physio-logical conditions, about 170 mg-9 g of albumin are filtereddaily by human glomeruli, but �30 mg are indeed excreted inthe urine largely due to protein reabsorption in the renalproximal tubule via clathrin-dependent, receptor-mediated en-docytosis, an essential mechanism for the transport of macro-molecules into the cells as well as across the epithelia (8, 42,57). Receptor-mediated endocytosis in the apical membraneof the proximal tubule involves binding of albumin, amonga number of other ligands, to the receptor megalin and to itsintracellular binding partner cubilin (5, 17, 47). Once theligand-receptor complex is internalized, it is directed to theendosomal compartment where the complex is dissociated;the receptors megalin/cubilin recycle back to the apical plasmamembrane whereas albumin is degraded in the lysosomes to itsconstituent amino acids that are released into the bloodstream(13, 17).
In addition to megalin and cubilin, the endocytosis of albuminrequires the coordinated action of a macromolecular complex,which includes the Na�/H� exchanger isoform 3 (NHE3), thevacuolar proton pump (v-H�-ATPase), and the chloride chan-nel ClC-5 (17, 22, 24, 26, 32, 36, 47, 53). These transportproteins are mainly involved in the maintenance of pH homeo-stasis of endosomal compartments, which is essential to theendocytic process, and influences not only the ligand-receptordissociation but also the vesicle trafficking, endosomal fusionevents, and recycling of receptors to the plasma membrane (20,38, 42, 54). The role of megalin, cubilin, NHE3, ClC-5, andv-H�-ATPase as critical components of the proximal tubuleendocytic machinery is supported by several studies conductedin knockout mice (22, 24, 46, 62). These experimental animalmodels (6, 52, 58) as well as a number of diseases character-ized by tubular proteinuria have been shown to be associatedwith decreased renal expression of these receptors and/ortransporters (2, 32, 44, 56).
Microalbuminuria is an independent risk factor for cardio-vascular disease and renal disease progression (4, 11, 37, 40,49). In essential hypertension, glomerular endothelial dys-function, intraglomerular hypertension, and hemodynamicmaladjustment as well as podocyte injury are believed tocontribute to microalbuminuria (11, 16, 41, 49). Alterna-tively, Russo and colleagues (50) have found that the pres-ence of microalbuminuria in spontaneously hypertensiverats (SHR) is independent of changes in glomerular perme-ability, suggesting that defective uptake of filtered albuminby the renal proximal tubule may represent an importantmechanism underlying the development of microalbumin-uria in this experimental model. The present study was
Address for reprint requests and other correspondence: A. C. C. Girardi,Laboratory of Genetics and Molecular Cardiology, Heart Institute, Univ. of SãoPaulo Medical School, Avenida Dr. Enéas de Carvalho Aguiar, 44, 10° andar,Bloco II, 05403-000 São Paulo, Brazil (e-mail: [email protected]).
Am J Physiol Renal Physiol 305: F216–F226, 2013.First published May 1, 2013; doi:10.1152/ajprenal.00255.2012.
1931-857X/13 Copyright © 2013 the American Physiological Society http://www.ajprenal.orgF216
by Adriana G
irardi on July 16, 2013http://ajprenal.physiology.org/
Dow
nloaded from
design to evaluate the temporal pattern of urinary proteinexcretion and to test the hypothesis that progression ofmicroalbuminuria is associated with lower renal expressionof critical components of the endocytic machinery thatmediate the reabsorption of albumin in the renal proximaltubule of spontaneously hypertensive rats.
MATERIALS AND METHODS
Materials. A monoclonal antibody directed to megalin was akind gift from Dr. Daniel Biemesderfer (Yale University School ofMedicine, New Haven, CT). We purchased polyclonal antibodiesto cubilin, transferrin, podocin, and nephrin, and a monoclonalantibody to v-ATPase B2 subunit from Santa Cruz Biotechnology(Santa Cruz, CA), a polyclonal antibody to ClC-5 from AlphaDiagnostic International (San Antonio, TX), and a monoclonalantibody to actin from Merck (Darmstadt, Germany). Horseradishperoxidase-conjugated goat anti-mouse, goat anti-rabbit, and rabbitanti-goat secondary antibodies were purchased from Life Technol-ogies (Carlsbad, CA). All other reagents and chemicals wereacquired from Sigma-Aldrich (St. Louis, MO), unless otherwisespecified.
Experimental animals. All procedures were carried out in accor-dance with the ethical principles of animal research of the BrazilianCollege of Animal Experimentation and were approved by the Insti-tutional Animal Care and Use Committee. Studies were performed in
male SHR and age-matched Wistar rats. Rats were housed at theUniversity of São Paulo Medical School animal facility under stan-dardized conditions (constant temperature of 22°C, 12:12-h light-darkcycle, and relative humidity of 60%). Blood pressure and renalfunction evaluation were performed at 6, 14, and 21 wk of age.Arterial blood was collected from the right ventricle at the time ofdeath for measurements of serum creatinine levels. Kidneys wereremoved, weighted, and used for preparation of renal cortical mem-branes or RNA extraction immediately after the rats had been eutha-nized by decapitation.
Blood pressure measurement. Blood pressure was measured bytail-cuff plethysmography (BP-2000 Blood Pressure Analysis System,Visitech Systems, Apex, NC). Before these measurements, rats weretrained in the blood pressure device to become adapted to the exper-imental procedures.
Renal function evaluation. Rats were individually placed intometabolic cages during 4 consecutive days: the first day to adapt therats to the cages and the following days to assess urine function. Urinesamples collected during each 24-h period were used to determineurine output, glomerular filtration rate (GFR), urinary creatinine, andsodium and protein excretion. Urine output was measured gravimet-rically. Creatinine clearance was used to estimate GFR. Serum andurinary creatinine concentrations were measured by a kinetic method(Labtest, Minas Gerais, Brazil) using a ThermoPlate Analyzer Plus(ThermoPlate, São Paulo, Brazil). Sodium was measured on a Radi-ometer ABL800 Flex (Radiometer Medical, Brønshøj, Denmark).Urinary protein excretion was determined using a Sensiprot kit(Labtest).
Urine albumin excretion. Urinary albumin concentration was de-termined with an ELISA kit specific for rat urine albumin (Nephratkit; Exocell, Philadelphia, PA). Experiments were carried out follow-ing the manufacturer’s instructions. Additionally, albumin was iso-lated from urine by SDS-PAGE and quantified by densitometry usingImageJ Software (Scion, Frederick, MD).
Preparation of renal cortical membranes. Rat renal cortices wereseparated at 4°C, minced with opposing razor blades, and homoge-nized in a Potter-Elvehjem-style tissue grinder (Polymix PX-SR 50;Kinematica, Bohemia, NY) for 25 strokes in ice-cold PBS (10 mMphosphate, 140 mM NaCl, pH 7.4) containing protease (1 mMpepstatin, 1 mM leupeptin, and 230 mM PMSF) and phosphataseinhibitors (15 mM NaF and 50 mM sodium pyrophosphate). Renalcortical membranes were prepared by differential centrifugation ofrenal cortical homogenates from individual animals as describedpreviously (12). Protein concentration was measured by the method ofLowry (33).
SDS-PAGE. Equivalent protein amounts of renal cortical mem-branes or a volume of urine containing 5 or 40 �g of creatininewere solubilized in SDS sample buffer (2% SDS, 10% glycerol,0.1% bromophenol blue, 50 mM Tris, pH 6.8), and proteins wereseparated by SDS-PAGE using 7.5 or 10% polyacrylamide gels
Table 1. Primers used for real-time RT-PCR and predictedamplicon size
GeneNCBI
Reference Sequence Size, bp
Lrp2 NM_030827 S-TCCATCCTTCTTCGTTGGA 131 bpAS-CCGGTTTGCTGTGATAACC
Cubn NM_053332 S-CACCAGGGGCTACAACAGGCT 154 bpAS-GGGTACCCAGGGCTGCTGAA
Clcn5 NM_017106 S-TTACGCCAATGGAGATCGT 183 bpAS-CAAAACAACCCCATGCTTC
Atp6v1b2 NM_057213 S-GAAAGCTGTGGTGGGAGAA 140 bpAS-CCAGCCAATGTCCAAAGTC
Nphs1 NM_022628 S-TGAGACGCTTCACAACACCCGT 92 bpAS-CCAGATGTCAGCCTGAGTCCTCT
Nphs2 NM_130828 S-AAGAGCATTGCCCAAGATG 146 bpAS-TCTTTGTGCCTCAGCTTCC
Ppia NM_017101 S-AATGCTGGACCAACACAAA 101 bpAS-CCTTCTTTCACCTTCCCAAA
Lrp2, megalin receptor; cubn, cubilin receptor; Atp6vl2, Clcn5, chloridechannel 5 (ClC-5); v-H�-ATPase transporter B2 subunit; Nphs1, nephrin;Nphs2, podocin; Ppia, peptidylprolyl isomerase A (cyclophilin A); S, sense;AS, antisense.
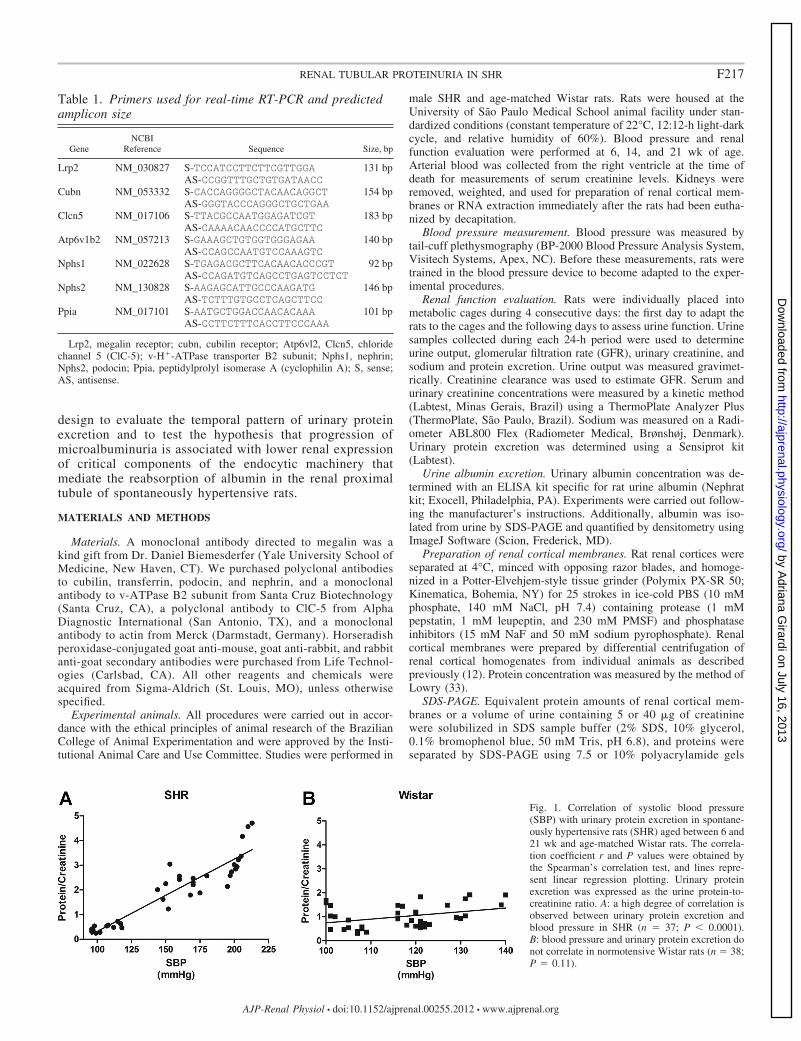
Fig. 1. Correlation of systolic blood pressure(SBP) with urinary protein excretion in spontane-ously hypertensive rats (SHR) aged between 6 and21 wk and age-matched Wistar rats. The correla-tion coefficient r and P values were obtained bythe Spearman’s correlation test, and lines repre-sent linear regression plotting. Urinary proteinexcretion was expressed as the urine protein-to-creatinine ratio. A: a high degree of correlation isobserved between urinary protein excretion andblood pressure in SHR (n � 37; P � 0.0001).B: blood pressure and urinary protein excretion donot correlate in normotensive Wistar rats (n � 38;P � 0.11).
F217RENAL TUBULAR PROTEINURIA IN SHR
AJP-Renal Physiol • doi:10.1152/ajprenal.00255.2012 • www.ajprenal.org
by Adriana G
irardi on July 16, 2013http://ajprenal.physiology.org/
Dow
nloaded from
according to Laemmli (31). Following electrophoresis, some gelswere silver stained using a ProteoSilver Plus kit to detect urinaryproteins.
Immunoblotting. Renal cortical membrane proteins were trans-ferred to polyvinylidene difluoride membranes (PVDF; Immobilon-P,Millipore) from polyacrylamide gels at 500 mA for 5 h at 4°C with aTE 62 Transfer Cooled Unit (GE HealthCare, Piscataway, NJ) andstained with Ponceau S in 0.5% trichloroacetic acid. PVDF mem-branes containing transferred proteins were incubated first in blotto(5% nonfat dry milk and 0.1% Tween 20 in PBS, pH 7.4) for 1 h,followed by overnight incubation in primary antibody. Primary anti-bodies were diluted in blotto in concentrations ranging from 1:500 to1:50,000. The membranes were then washed five times in blotto andincubated for 1 h with horseradish peroxidase-conjugated immuno-globulin secondary antibody (1:2,000). Bound antibody was detected
using an enhanced chemiluminescence system (GE Healthcare) ac-cording to the manufacturer’s protocols. The signals were capturedusing an ImageQuant LAS 4000 mini (GE HealthCare) and quantifiedusing ImageJ Software (Scion).
RNA extraction and real-time RT-PCR. Total RNA was isolatedfrom rat renal cortices by using TRIzol Reagent (Life Technologies)according to the manufacturer’s specifications. First-strand cDNA wassynthesized using a random High Capacity cDNA Archive kit (Ap-plied Biosystems, Foster City, CA) following the manufacturer’sguidelines. Quantitative real-time RT-PCRs were carried out usingSYBR Green PCR Master Mix-PE (Applied Biosystems) on the ABIPrism 7700 Sequence Detection System (Applied Biosystems). Theoligonucleotide primers used to detect megalin, cubilin; ClC-5, v-H�-ATPase B2-subunit, nephrin, podocin, and the internal control cyclo-philin are shown in Table 1. mRNA analyses of target genes were
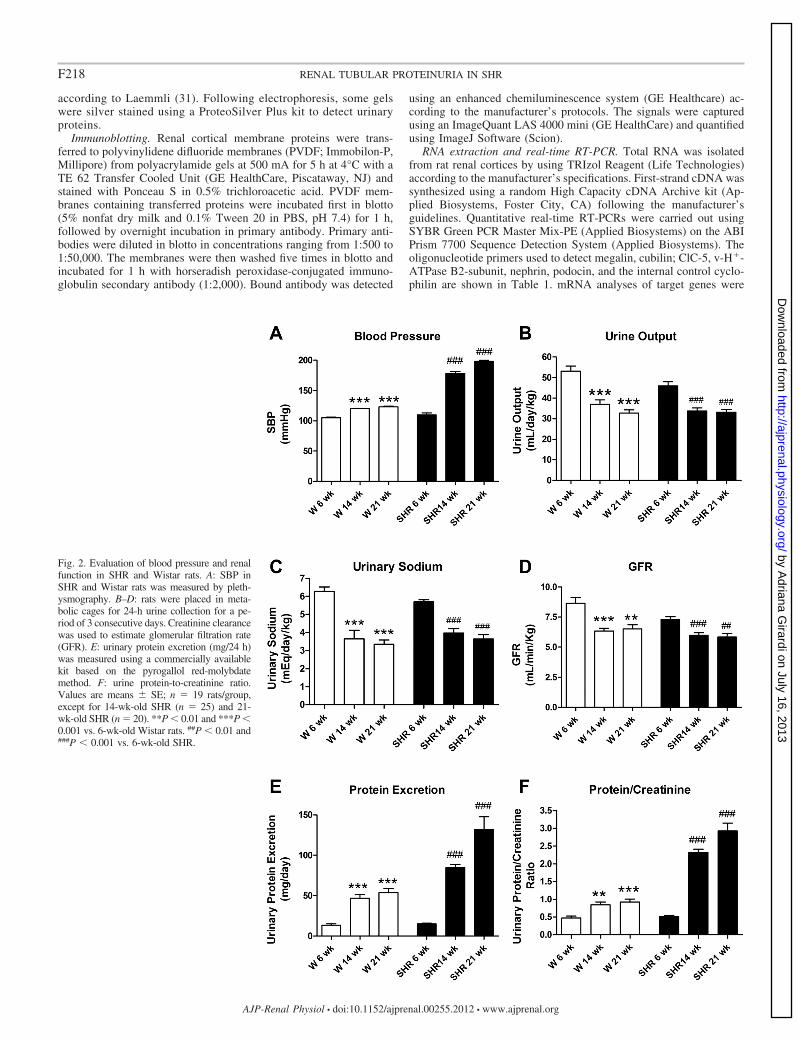
Fig. 2. Evaluation of blood pressure and renalfunction in SHR and Wistar rats. A: SBP inSHR and Wistar rats was measured by pleth-ysmography. B–D: rats were placed in meta-bolic cages for 24-h urine collection for a pe-riod of 3 consecutive days. Creatinine clearancewas used to estimate glomerular filtration rate(GFR). E: urinary protein excretion (mg/24 h)was measured using a commercially availablekit based on the pyrogallol red-molybdatemethod. F: urine protein-to-creatinine ratio.Values are means � SE; n � 19 rats/group,except for 14-wk-old SHR (n � 25) and 21-wk-old SHR (n � 20). **P � 0.01 and ***P �0.001 vs. 6-wk-old Wistar rats. ##P � 0.01 and###P � 0.001 vs. 6-wk-old SHR.
F218 RENAL TUBULAR PROTEINURIA IN SHR
AJP-Renal Physiol • doi:10.1152/ajprenal.00255.2012 • www.ajprenal.org
by Adriana G
irardi on July 16, 2013http://ajprenal.physiology.org/
Dow
nloaded from
assayed in triplicate. The comparative threshold (CT) cycle methodwas used for data analyses.
Histological and morphometric analysis. The kidneys from 6–8rats/group were perfused with PBS and with 4% paraformaldehyde inPBS via the left ventricle. The kidneys were removed, weighed, fixedovernight in 4% paraformaldehyde in PBS, and embedded in paraffin.For morphometric analysis, 3-�m-thick sections were stained withhematoxylin-eosin (HE). Renal sections were subsequently examinedunder light microscopy, and the images and measurements wereacquired using a Leica DM2700 M microscope and Leica Qwinsoftware (version 2.2), respectively. Twenty glomeruli per slide werechosen for analysis. The major (D) and minor (d) diameters and areaof glomeruli and Bowman’s capsule were measured. Determination ofthe mean glomerular diameter (M) and volume (V) of glomeruli andBowman’s capsule was calculated using the following equations M �� D � d and V � �/6 � M3. The formula A � �/4 � M2 was usedto confirm the area measured.
Statistical analysis. Data are expressed as means � SE, with nindicating the number of observations. Comparisons among groupswere made by one-way ANOVA, unless indicated otherwise. Adjust-ment for multiple comparisons was made with the Bonferroni method.Differences were considered significant if P � 0.05.
RESULTS
Correlation between urinary protein excretion and bloodpressure in SHR. Regression analysis was performed to exam-ine whether urinary protein excretion was correlated withsystolic blood pressure (SBP) at early ages in SHR. To thisend, we plotted the values of SBP against the urinary protein/creatinine ratio from 25 SHR ranging from 6 to 21 wk of age.As shown in Fig. 1, urinary protein excretion increases signif-icantly with SBP in SHR during the analyzed period (Spear-man correlation coefficient, r � 0.9244; P � 0.0001) (Fig. 1A).Correlation between these variables was not observed in age-matched normotensive Wistar rats (r � 0.3147, P � 0.11) (Fig.1B). To evaluate the pattern of urinary protein excretion and totest the hypothesis that progression of proteinuria is associatedwith diminished expression of critical components of the apicalendocytic apparatus in the renal proximal tubule of SHR, allthe following experiments were undertaken in 6-, 14-, and21-wk-old SHR. Age-matched Wistar rats served as the refer-ence age control.
Evaluation of blood pressure and renal function in SHR andWistar rats. The temporal evaluation of blood pressure andrenal function in 6-, 14-, and 21-wk-old SHR and Wistar ratsof the corresponding age is illustrated in Fig. 2. Figure 2Ashows that SBP values increased from 6 to 14 wk of age
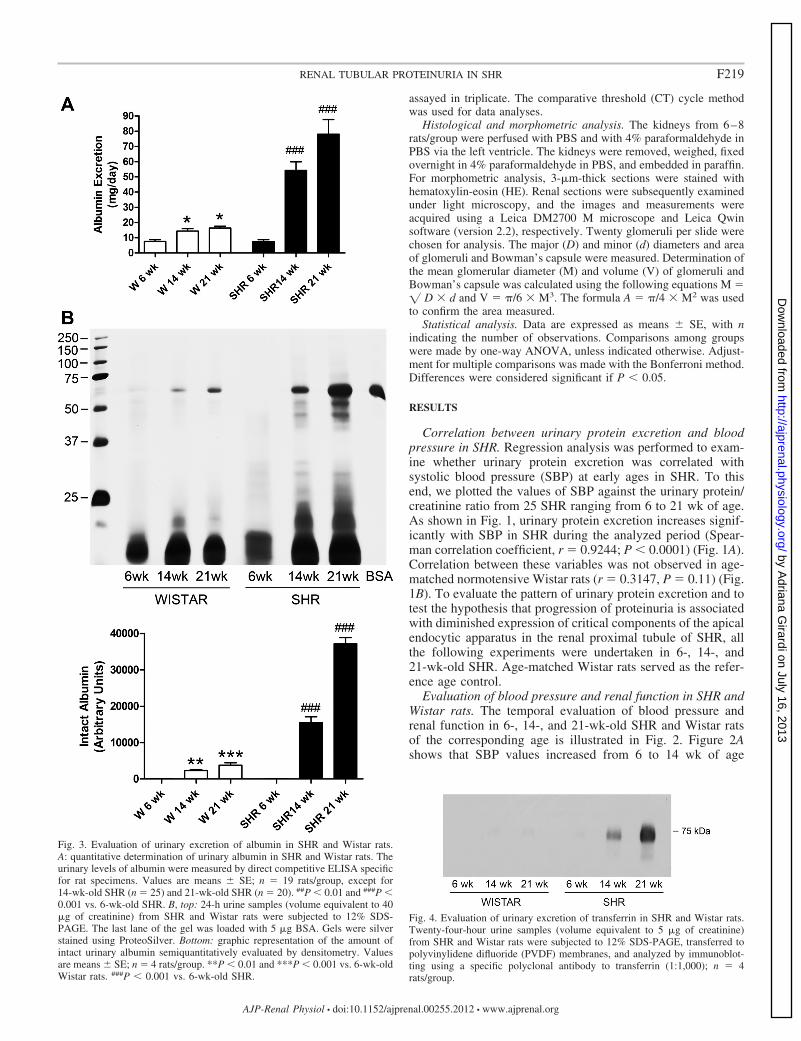
Fig. 4. Evaluation of urinary excretion of transferrin in SHR and Wistar rats.Twenty-four-hour urine samples (volume equivalent to 5 �g of creatinine)from SHR and Wistar rats were subjected to 12% SDS-PAGE, transferred topolyvinylidene difluoride (PVDF) membranes, and analyzed by immunoblot-ting using a specific polyclonal antibody to transferrin (1:1,000); n � 4rats/group.
Fig. 3. Evaluation of urinary excretion of albumin in SHR and Wistar rats.A: quantitative determination of urinary albumin in SHR and Wistar rats. Theurinary levels of albumin were measured by direct competitive ELISA specificfor rat specimens. Values are means � SE; n � 19 rats/group, except for14-wk-old SHR (n � 25) and 21-wk-old SHR (n � 20). ##P � 0.01 and ###P �0.001 vs. 6-wk-old SHR. B, top: 24-h urine samples (volume equivalent to 40�g of creatinine) from SHR and Wistar rats were subjected to 12% SDS-PAGE. The last lane of the gel was loaded with 5 �g BSA. Gels were silverstained using ProteoSilver. Bottom: graphic representation of the amount ofintact urinary albumin semiquantitatively evaluated by densitometry. Valuesare means � SE; n � 4 rats/group. **P � 0.01 and ***P � 0.001 vs. 6-wk-oldWistar rats. ###P � 0.001 vs. 6-wk-old SHR.
F219RENAL TUBULAR PROTEINURIA IN SHR
AJP-Renal Physiol • doi:10.1152/ajprenal.00255.2012 • www.ajprenal.org
by Adriana G
irardi on July 16, 2013http://ajprenal.physiology.org/
Dow
nloaded from
(110 � 3 vs. 180 � 2 mmHg, P � 0.001) and continued to risefrom 14 to 21 wk of age in SHR (178 � 3 vs. 198 � 2 mmHg,P � 0.001). In contrast, a small but significant increase inblood pressure was observed from 6 to 14 wk of age in Wistar
rats (105 � 1 vs. 120 � 1 mmHg, P � 0.001), but noadditional increment was observed from 14 to 21 wk of age inthe lineage of normotensive animals [120 � 1 vs. 123 � 2mmHg, not significant (NS)].
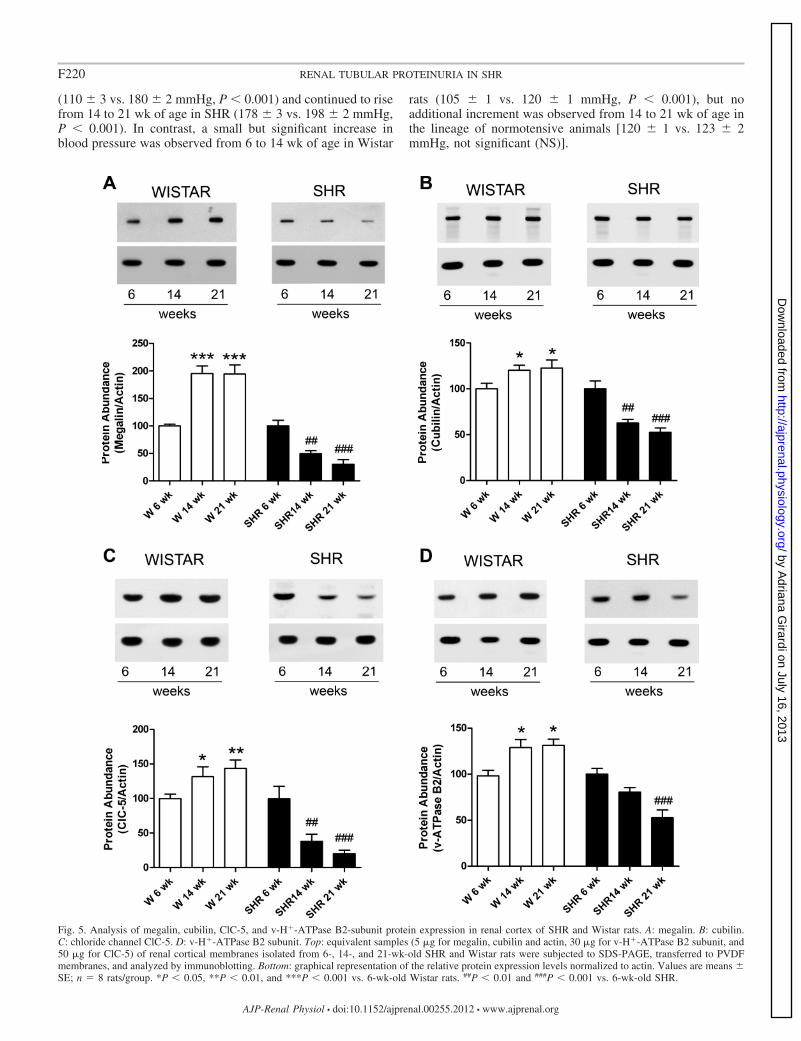
Fig. 5. Analysis of megalin, cubilin, ClC-5, and v-H�-ATPase B2-subunit protein expression in renal cortex of SHR and Wistar rats. A: megalin. B: cubilin.C: chloride channel ClC-5. D: v-H�-ATPase B2 subunit. Top: equivalent samples (5 �g for megalin, cubilin and actin, 30 �g for v-H�-ATPase B2 subunit, and50 �g for ClC-5) of renal cortical membranes isolated from 6-, 14-, and 21-wk-old SHR and Wistar rats were subjected to SDS-PAGE, transferred to PVDFmembranes, and analyzed by immunoblotting. Bottom: graphical representation of the relative protein expression levels normalized to actin. Values are means �SE; n � 8 rats/group. *P � 0.05, **P � 0.01, and ***P � 0.001 vs. 6-wk-old Wistar rats. ##P � 0.01 and ###P � 0.001 vs. 6-wk-old SHR.
F220 RENAL TUBULAR PROTEINURIA IN SHR
AJP-Renal Physiol • doi:10.1152/ajprenal.00255.2012 • www.ajprenal.org
by Adriana G
irardi on July 16, 2013http://ajprenal.physiology.org/
Dow
nloaded from
The patterns of variation of urine output, urinary sodiumexcretion, and GFR were similar between the two rat strains.Urine output (Fig. 2B), urinary sodium excretion (Fig. 2C), andGFR (Fig. 2D) were higher in 6-wk-old animals compared with14- and 21-wk-old rats of the same lineage. These renalfunction parameters remain unchanged from 14 to 21 wk of agein both SHR and Wistar rats.
Consistent with the findings shown in Fig. 1, daily urinaryexcretion of protein progressively increased in SHR throughoutthe experimental period (Fig. 2E). A significant increase in thedaily urinary excretion of protein was noted between 6- and14-wk-old Wistar rats, but no further changes were observedbetween 14- and 21-wk-old normotensive animals (Fig. 2E).To minimize the differences in GFR and urinary losses thatmay result in underestimated values of protein excretion; wenormalized protein excretion by creatinine (Fig. 2F). As ex-pected, the urinary protein-to-creatinine ratio progressivelyincreased in SHR (0.51 � 0.04; 2.32 � 0.09; and 2.93 � 0.22at 6, 14, and 21 wk of age, respectively), whereas the ratioincreased from 6 to 14 wk (0.47 � 0.05 vs. 0.61 � 0.06, P �0.05) and remained unchanged between 14- and 21-wk-oldWistar rats (0.84 � 0.07 vs. 0.92 � 0.08, NS).
Evaluation of albumin excretion in SHR and Wistar rats.Urinary albumin excretion was quantified by ELISA (Fig. 3A)and analyzed by SDS-PAGE (Fig. 3B). SHR exhibited aprogressive increase in urinary albumin excretion from 6 to 21wk of age (Fig. 3A), while albumin excretion remained withinthe normal range (�30 mg/day) in normotensive Wistar ratsand in 6-wk-old SHR.
The profile of urinary proteins excreted by SHR and Wistaranimals was evaluated by SDS-PAGE, and the amount of
intact albumin was semiquantitatively determined by densi-tometry (Fig. 3B). The results from these experiments werevery similar to the ones obtained in Fig. 3A. As seen in thebottom panel of Fig. 3B, there was a continuing increase in thelevels of intact albumin in the urine of Wistar rats throughoutthe experimental period (94 � 6, 2,318 � 250, and 3,778 �665 in 6-, 14-, and 21-wk-old Wistar rats, respectively), but nostatistical difference was observed between 14- and 21-wk-oldnormotensive animals (P 0.05). The increase in the urinarylevels of intact albumin was much more pronounced in SHR(70 � 30, 15,456 � 1,663, and 37,166 � 1,692 arbitrary unitsin 6-, 14-, and 21-wk-old SHR, respectively). Most impor-tantly, the molecular weights of proteins excreted in the urineof the animals displaying microalbuminuria (14- and 21-wk-old SHR) were similar in size to albumin or smaller, typical oftubular proteinuria (Fig. 3B).
Evaluation of transferrin excretion in SHR and Wistar rats.The qualitative change in the levels of urinary transferrin, aprotein that is known to be reabsorbed from the glomerularultrafiltrate by receptor-mediated endocytosis (30), was alsoevaluated in 24-h urine samples from SHR and Wistar rats.Figure 4 shows the results of immunoblot analyses of trans-ferrin in the urine of these experimental animal models. Vir-tually no excretion of transferrin could be observed in 24 urinesamples obtained from normotensive Wistar rats and from6-wk-SHR (n � 4 animals/group) while transferrin contentprogressively increased in the urine of 14- and 21-wk-old SHR.
Evaluation of protein and mRNA expression of megalin,cubilin, ClC-5, and the v-ATPase B2 subunit in the renal cortexof SHR and Wistar rats. The results obtained above suggesttubular proteinuria in SHR, so we next evaluated whether
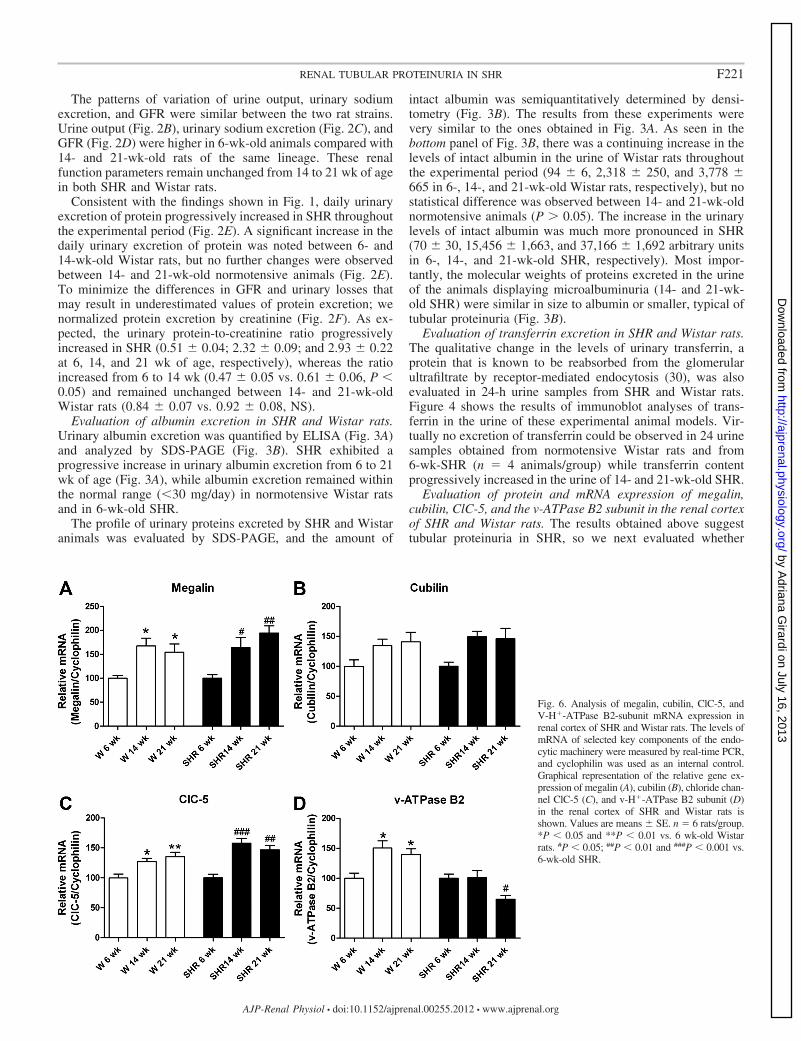
Fig. 6. Analysis of megalin, cubilin, ClC-5, andV-H�-ATPase B2-subunit mRNA expression inrenal cortex of SHR and Wistar rats. The levels ofmRNA of selected key components of the endo-cytic machinery were measured by real-time PCR,and cyclophilin was used as an internal control.Graphical representation of the relative gene ex-pression of megalin (A), cubilin (B), chloride chan-nel ClC-5 (C), and v-H�-ATPase B2 subunit (D)in the renal cortex of SHR and Wistar rats isshown. Values are means � SE. n � 6 rats/group.*P � 0.05 and **P � 0.01 vs. 6 wk-old Wistarrats. #P � 0.05; ##P � 0.01 and ###P � 0.001 vs.6-wk-old SHR.
F221RENAL TUBULAR PROTEINURIA IN SHR
AJP-Renal Physiol • doi:10.1152/ajprenal.00255.2012 • www.ajprenal.org
by Adriana G
irardi on July 16, 2013http://ajprenal.physiology.org/
Dow
nloaded from
progression of microalbuminuria was associated with a lowerexpression of selected proteins involved in the process ofreceptor-mediated endocytosis in renal proximal tubule ofgenetic hypertensive animals. Protein abundance (Fig. 5) andthe mRNA levels (Fig. 6) of megalin, cubilin, ClC-5, and thev-H�-ATPase B2 subunit were examined in the renal cortex ofSHR and Wistar rats. Regulation of the B2 subunit of thev-H�-ATPase was examined, as opposed to other subunits,because it is known that B2 is expressed in the apical mem-brane of the proximal tubule where it plays a role in theacidification of intracellular organelles and possibly in trans-epithelial proton secretion (45), and because we have previ-ously found that the activity of the v-H�-ATPase is regulatedby changes of the B2 subunit in response to angiotensin II (9)and by both ClC-5 and CFTR in the renal proximal tubule (10).
We noticed a similar pattern for renal cortical protein abun-dance of megalin, cubilin, ClC-5, and the v-H�-ATPase B2
subunit in normotensive Wistar rats (Fig. 5, A–D). The relativeexpression of these four proteins normalized to actin increasedin both 14- and 21-wk-old rats compared with 6-wk-old Wistarrats. Conversely, the protein abundance of megalin (Fig. 5A),cubilin (Fig. 5B), and ClC-5 (Fig. 5C) progressively decreasedin the renal cortex of SHR. Reduction of megalin proteinexpression was 50% at 14 wk of age and 70% at 21 wk of agecompared with 6-wk-old SHR. The relative protein expressionlevels of cubilin decayed 40% at 14 wk of age and 50% at 21wk of age compared with 6-wk-old SHR. Similarly, the relativeprotein levels of ClC-5 were 60% lower at 14 wk of age and80% lower at 21 wk of age compared with 6-wk-old SHR.Regarding cortical expression of the v-H�-ATPase B2 subunit,no statistical difference was observed between 6-wk-old and14-wk-old SHR; however, a significant decrease was observedin 21-wk-old SHR (47 � 7% vs. 6 wk-old SHR; P 0.001 and29 � 3% vs. 14 wk-old SHR, P � 0.05) (Fig. 5D).
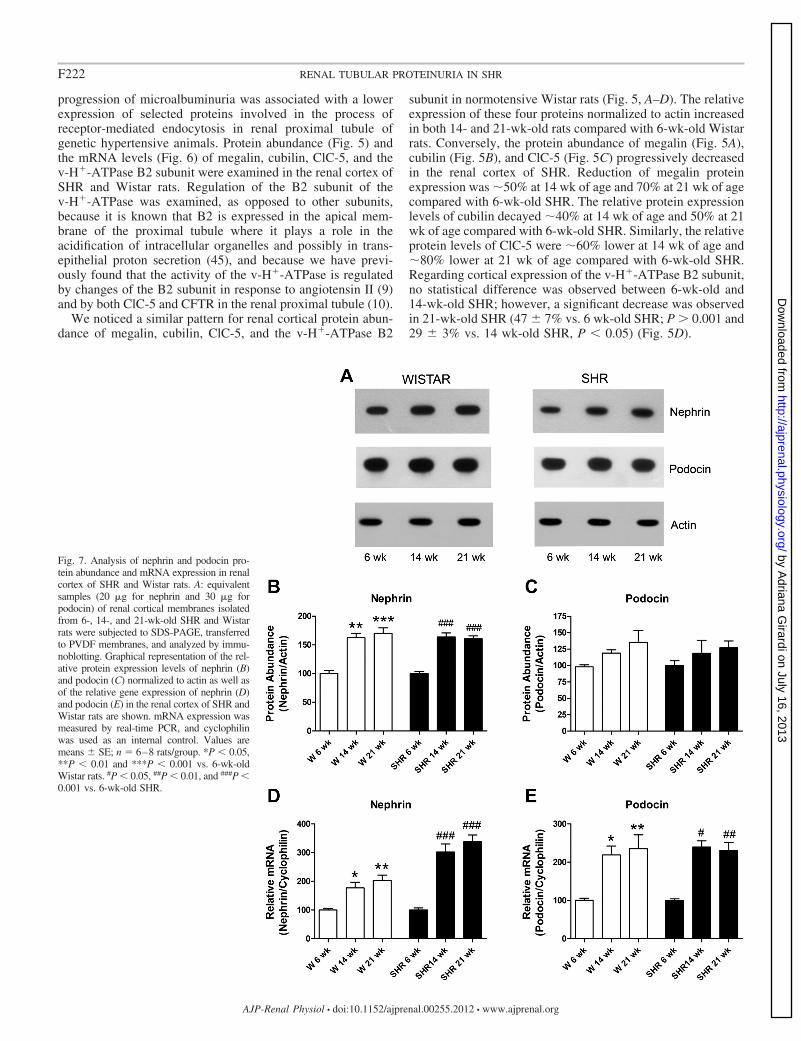
Fig. 7. Analysis of nephrin and podocin pro-tein abundance and mRNA expression in renalcortex of SHR and Wistar rats. A: equivalentsamples (20 �g for nephrin and 30 �g forpodocin) of renal cortical membranes isolatedfrom 6-, 14-, and 21-wk-old SHR and Wistarrats were subjected to SDS-PAGE, transferredto PVDF membranes, and analyzed by immu-noblotting. Graphical representation of the rel-ative protein expression levels of nephrin (B)and podocin (C) normalized to actin as well asof the relative gene expression of nephrin (D)and podocin (E) in the renal cortex of SHR andWistar rats are shown. mRNA expression wasmeasured by real-time PCR, and cyclophilinwas used as an internal control. Values aremeans � SE; n � 6–8 rats/group. *P � 0.05,**P � 0.01 and ***P � 0.001 vs. 6-wk-oldWistar rats. #P � 0.05, ##P � 0.01, and ###P �0.001 vs. 6-wk-old SHR.
F222 RENAL TUBULAR PROTEINURIA IN SHR
AJP-Renal Physiol • doi:10.1152/ajprenal.00255.2012 • www.ajprenal.org
by Adriana G
irardi on July 16, 2013http://ajprenal.physiology.org/
Dow
nloaded from

The relative mRNA expression of megalin, cubilin, ClC-5,and the v-H�-ATPase B2 subunit in renal cortex from SHRand Wistar rats was determined by real-time RT-PCR using thespecific primers depicted in Table 1. The expression of thesetranscripts was normalized by cyclophilin. As illustrated in Fig.6, the pattern of mRNA expression of these four genes wasconsistent with the one observed at the protein level in the renalcortex of Wistar rats. Interestingly, this pattern was also ob-served for megalin, cubilin, and ClC-5 transcripts in the renalcortex of SHR. As opposed to the progressive decrease inprotein levels (Figs. 5, A–C), the mRNA expression of megalin(Fig. 6A), cubilin (Fig. 6B), and ClC-5 (Fig. 6C) was signifi-cantly higher in both 14- and 21-wk-old rats compared with6-wk-old SHR. On the other hand, changes in v-H�-ATPaseB2 subunit mRNA (Fig. 6D) were associated with changes inprotein expression (Fig. 5D) in the renal cortex of SHR.
Evaluation of protein and mRNA expression of nephrin andpodocin in the renal cortex of SHR and Wistar rats. Recentevidence suggests that the most selective barrier for the ultra-filtration of proteins resides in the glomerular slit diaphragm(14, 39, 59). Because a decrease in the expression of the slitdiaphragm molecular components, nephrin and/or podocin (1,15, 25, 29, 34), has been associated with proteinuria, we alsoevaluated whether nephrin and podocin expression diminisheswith a blood pressure increase in SHR.
As illustrated in Fig. 7, a similar temporal expression profilefor renal cortical nephrin and podocin protein and mRNA wasobserved in both strains. Immunoblot analysis showed that therelative protein abundance of nephrin, normalized by actin,was significantly higher in adult Wistar and SHR rats (at 14and 21 wk of age) compared with rats of the same lineage at 6wk of age (Fig. 7, A and B). On the other hand, the proteinabundance of podocin remained unaltered with age in bothSHR and Wistar rats (Fig. 7, A and C). Analysis of the relativeabundance of nephrin (Fig. 7D) and podocin (Fig. 7E) tran-scripts in the renal cortex of SHR and Wistar rats indicates thatthe expression of these slit diaphragm components is signifi-cantly higher in both 14- and 21-wk-old compared with 6-wk-old rats. Interestingly, whereas Wistar rats had approximatelytwofold greater nephrin mRNA levels at 14 and 21 wk com-pared with 6 wk, nearly a fourfold increase was observed inSHR during the same experimental period (Fig. 7D).
Evaluation of renal hypertrophy in SHR. Hypertension maylead to renal hypertrophy and glomerular injury that are notalways associated with altered renal hemodynamics (3, 27).We therefore evaluated whether renal hypertrophy, assessed as
the kidney weight/body weight ratio, as well as glomerularvolume (43), a key determinant of glomerular structuralchanges, were altered in SHR throughout the experimentalperiod compared with normotensive Wistar rats.
As depicted in Table 2, body weight and kidney weightprogressively increased with age in both SHR and Wistar rats.SHR displayed significantly lower body weight and kidneyweight than that of age-matched Wistar rats (P � 0.001). Thekidney weight/body weight ratio was higher in 6-wk-oldWistar rats and in SHR compared with 14- and 21-wk-old ratsof the same lineage. There was no difference between SHR andage-matched Wistar rats in terms of the kidney weight/bodyweight ratio, suggesting that there was no renal hypertrophy inSHR during this experimental period.
The results of glomerular volume estimation are shown inTable 2. In both SHR and Wistar rats, mean glomerular volumesignificantly increased from 6 to 14 wk of age, but remainedunchanged from 14 to 21 wk of age. Similarly, Bowman’scapsule volume was higher in 14- and 21-wk-old than in6-yr-old rats. In concert, these findings suggest that increasedurinary excretion of albumin in SHR, among other proteins,was not associated with glomerular hypertrophy during thisexperimental period.
DISCUSSION
The results from the present study provide evidence support-ing the view that tubular dysfunction represents an importantcontributing mechanism underlying microalbuminuria in hy-pertension. First, despite the progressive increase in microalbu-minuria and urinary transferrin excretion between 14- and21-wk-old SHR, GFR and mean glomerular volume remainedunchanged during this period, thus minimizing the role ofglomerular hyperfiltration of proteins. Second, analyses ofurine samples by SDS-PAGE showed that microalbuminuricSHR only excreted proteins with molecular weight similar orsmaller than albumin (70 kDa), which resembles the profile ofurinary proteins excreted by typical models of tubular protein-uria observed in megalin and ClC-5 knockout mice. Third,progression of microalbuminuria in SHR throughout the ex-perimental period was associated with lower expression of keycomponents of the apical endocytic machinery in the renalproximal tubule. Finally, we found no evidence for changes inthe protein abundance of the glomerular proteins nephrin (28)or podocin (7) over the 21-wk experimental period.
Table 2. Physiological and renal morphometric parameters
Wistar SHR
5 wk 14 wk 21 wk 5 wk 14 wk 21 wk
Body wt, g 181 � 3 (19) 358 � 11*** (19) 417 � 18*** (19) 136 � 3 (19) 292 � 4### (25) 321 � 3### (20)Kidney wt, mg 2089 � 51 (8) 3199 � 121*** (8) 3428 � 106*** (8) 999 � 16 (8) 1815 � 86### (13) 2128 � 87### (12)Kidney wt/body wt, mg/g 9.37 � 0.16 (8) 6.94 � 0.19* (8) 7.26 � 0.19* (8) 8.70 � 0.10 (8) 6.71 � 0.10# (13) 6.78 � 0.11# (12)Glomerular volume, �m3 � 104 12.7 � 1.6 (6) 29.5 � 1.0** (6) 34.6 � 6.0** (6) 14.9 � 1.2 (6) 27.8 � 3.2# (8) 32.0 � 7.1# (7)Glomerular area, �m2 � 103 2.9 � 0.24 (6) 5.2 � 0.11** (6) 5.7 � 0.64** (6) 3.3 � 0.18 (6) 5.0 � 0.39# (8) 5.5 � 0.82# (7)Bowman’s capsule volume,
�m3 � 104 21.2 � 2.5 (6) 50.3 � 1.8* (6) 58.9 � 10.1** (6) 25.5 � 2.2 (6) 45.1 � 3.3## (8) 55.6 � 10.7## (7)Bowman’s capsule area, �m2 � 103 4.2 � 0.30 (6) 7.5 � 0.1* (6) 8.2 � 0.89** (6) 4.7 � 0.26 (6) 7.0 � 0.34## (8) 8.0 � 1.0## (7)
Values are means � SE. Number of animals per group is indicated in parenthesis. SHR, spontaneously hypertenisve rats. *P � 0.05, **P � 0.01 and ***P �0.001 vs. 6-wk-old Wistar rats. #P � 0.5, ##P � 0.01, and ###P � 0.001 vs. 6-wk-old SHR.
F223RENAL TUBULAR PROTEINURIA IN SHR
AJP-Renal Physiol • doi:10.1152/ajprenal.00255.2012 • www.ajprenal.org
by Adriana G
irardi on July 16, 2013http://ajprenal.physiology.org/
Dow
nloaded from

Uptake of albumin by the proximal tubules from glomerularfiltrate occurs via receptor-mediated endocytosis, which re-quires the formation of the megalin/cubilin receptor complexand several accessory plasma membrane transport proteins. Wefound that the protein expression of the endocytic receptorsmegalin and cubilin, as well as of the chloride channel ClC-5and of the v-H�-ATPase B2-subunit progressively decreases inSHR from 14 to 21 wk of age, indicating that receptor-mediated endocytosis is compromised at this stage of hyper-tension. To our knowledge, this is the first study that shows thatmicroalbuminuria in SHR is associated with lower renal cor-tical expression of megalin, cubilin, and the v-ATPase B2subunit. On the other hand, reduced renal expression of ClC-5in microalbuminuric SHR has been previously demonstratedby Tanaka and Nakaki (55). These authors found that therelative expression of ClC-5 in the renal cortex of SHR was60% lower than in Wistar-Kyoto (WKY) rats at 11 wk of ageand 70% lower compared with 14-wk-old animals. Our find-ings are consistent with this previous study, since we have alsofound that the expression of the ClC-5 channel progressivelydecreases in the renal cortex of SHR.
One intriguing finding of our study is the occurrence of thedissociation between protein and mRNA expression of selectedcomponents of the endocytic machinery in the renal proximaltubule of SHR. The progressive decrease in the protein expres-sion of megalin, cubilin, and ClC-5 in the renal cortex of SHRis not dependent on transcription regulation. In fact, proteinexpression decreased despite increased mRNA expression ofthese genes. This dissociation might be due to posttranscrip-tional events such as modulation of translation by RNA bind-ing proteins (61) and/or by increased degradation of thesemembrane proteins. However, we have no direct explanation asto why this dissociation occurs.
Numerous in vitro studies (18, 19, 21) as well as in vivoobservations (22) have demonstrated that NHE3 is one of theplasma membrane transport proteins necessary for albuminendocytosis in the renal proximal tubule. Regulation of NHE3in genetic models of essential hypertension has been exten-sively studied (23); however, whether there are changes onNHE3-mediated acidification at the early endosomal compart-ment and whether it is correlated to tubular dysfunction in SHRremains to be determined. We have previously demonstratedthat NHE3 transport activity is inhibited in the renal proximaltubule of SHR after development of hypertension (12). Inhibi-tion of NHE3-mediated-NaHCO3 reabsorption in 14-wk-oldSHR compared with age-matched WKY rats is mainly due toredistribution of the transporter from the body to the base of themicrovilli with no significant changes in NHE3 total proteinabundance. Furthermore, changes in NHE3 transport activitywere accompanied by changes in the phosphorylation level ofthe transporter at the PKA consensus site serine 552 (12, 23,35). In this regard, receptor-mediated endocytosis has beenshown to be downregulated by cAMP/PKA in proximal tubulecells via inhibition of NHE3 (21). Further work needs to bedone to detect whether increases in the phosphorylation statusof NHE3, especially at the PKA consensus sites, may lead toalkalinization of the endosomal compartment, and conse-quently to decreased uptake of albumin by proximal tubulecells.
There is a growing body of evidence suggesting that prox-imal tubular dysfunction may play an important role in the
development of microalbuminuria in the early stages of dia-betic nephropathy (51, 58). The tubular reabsorption of albu-min is decreased in streptozotocin (STZ)-induced-diabetic-rats(58). This drop in albumin reabsorption occurs independentlyof changes in glomerular permeability (50, 51), and it isassociated with a reduction of megalin expression in the renalproximal tubules (58). In a similar fashion, the results of ourcurrent study suggest that the causal events leading to mi-croalbuminuria in hypertension are mainly of tubular origin. Itis noteworthy to mention that microalbuminuria progressed toovert proteinuria in 48-wk-old SHR, when total urinary proteinexcretion was 500 mg/day. Moreover, analyses of urinaryproteins by SDS-PAGE revealed that these 12-mo-old SHRexcreted proteins with molecular weights both smaller andlarger than albumin (data not shown). These observationssuggest that dysfunction of the glomerular filtration barriermay occur at the macroalbuminuric stage. In this regard,previous studies have demonstrated that the integrity of theglomerular filtration barrier is normal in SHR during the first 9mo of age. However, disturbances in glomerular protein siev-ing of a size-selective nature progressively develops in olderhypertensive rats (3, 27).
In conclusion, these results demonstrate that the urinaryexcretion of albumin and transferrin, among other low-molec-ular-weight proteins, progressively increases in SHR from 6 to21 wk of age and that this enhanced urinary protein excretionis associated with lower expression of key components of theapical endocytic apparatus in the renal proximal tubule. More-over, it is tempting to speculate that dysfunction of the apicalendocytic pathway in the renal proximal tubule may be acontributing mechanism underlying the development of mi-croalbuminuria in hypertension.
GRANTS
This work was supported by Fundação de Amparo a Pesquisa do Estado deSão Paulo (FAPESP) Grants 07/52945-8 and 12/10146-0.
DISCLOSURES
No conflicts of interest, financial or otherwise, are declared by the authors.
AUTHOR CONTRIBUTIONS
Author contributions: B.H.I., J.E.K., and A.C.C.G. contributed conception anddesign of research; B.H.I., D.F.A.-J., L.C.G.C., A.L.T.B., and M.V.R. performedexperiments; B.H.I., D.F.A.-J., L.C.G.C., A.L.T.B., M.V.R., and A.C.C.G. ana-lyzed data; B.H.I., D.F.A.-J., L.C.G.C., A.L.T.B., M.V.R., and A.C.C.G. inter-preted results of experiments; B.H.I., D.F.A.-J., L.C.G.C., and A.C.C.G. preparedfigures; B.H.I. and A.C.C.G. drafted manuscript; B.H.I., J.E.K., and A.C.C.G.edited and revised manuscript; B.H.I., D.F.A.-J., L.C.G.C., A.L.T.B., M.V.R.,J.E.K., and A.C.C.G. approved final version of manuscript.
REFERENCES
1. Agrawal V, Prasad N, Jain M, Pandey R. Reduced podocin expressionin minimal change disease and focal segmental glomerulosclerosis isrelated to the level of proteinuria. Clin Exp Nephrol. [Epub ahead of print].
2. Aminoff M, Carter JE, Chadwick RB, Johnson C, Grasbeck R,Abdelaal MA, Broch H, Jenner LB, Verroust PJ, Moestrup SK, de laChapelle A, Krahe R. Mutations in CUBN, encoding the intrinsicfactor-vitamin B12 receptor, cubilin, cause hereditary megaloblastic anae-mia 1. Nat Genet 21: 309–313, 1999.
3. Bakoush O, Tencer J, Torffvit O, Tenstad O, Skogvall I, Rippe B.Increased glomerular albumin permeability in old spontaneously hyper-tensive rats. Nephrol Dial Transplant 19: 1724–1731, 2004.
4. Bianchi S, Bigazzi R, Campese VM. Microalbuminuria in essentialhypertension: significance, pathophysiology, and therapeutic implications.Am J Kidney Dis 34: 973–995, 1999.
F224 RENAL TUBULAR PROTEINURIA IN SHR
AJP-Renal Physiol • doi:10.1152/ajprenal.00255.2012 • www.ajprenal.org
by Adriana G
irardi on July 16, 2013http://ajprenal.physiology.org/
Dow
nloaded from

5. Birn H, Christensen EI. Renal albumin absorption in physiology andpathology. Kidney Int 69: 440–449, 2006.
6. Birn H, Fyfe JC, Jacobsen C, Mounier F, Verroust PJ, Orskov H,Willnow TE, Moestrup SK, Christensen EI. Cubilin is an albuminbinding protein important for renal tubular albumin reabsorption. J ClinInvest 105: 1353–1361, 2000.
7. Boute N, Gribouval O, Roselli S, Benessy F, Lee H, Fuchshuber A,Dahan K, Gubler MC, Niaudet P, Antignac C. NPHS2, encoding theglomerular protein podocin, is mutated in autosomal recessive steroid-resistant nephrotic syndrome. Nat Genet 24: 349–354, 2000.
8. Brown MS, Goldstein JL. A receptor-mediated pathway for cholesterolhomeostasis. Science 232: 34–47, 1986.
9. Carraro-Lacroix LR, Girardi AC, Malnic G. Long-term regulation ofvacuolar H�-ATPase by angiotensin II in proximal tubule cells. PflügersArch 458: 969–979, 2009.
10. Carraro-Lacroix LR, Lessa LM, Bezerra CN, Pessoa TD, Souza-Menezes J, Morales MM, Girardi AC, Malnic G. Role of CFTR andClC-5 in modulating vacuolar H�-ATPase activity in kidney proximaltubule. Cell Physiol Biochem 26: 563–576, 2010.
11. Cerasola G, Cottone S, Mule G. The progressive pathway of microalbu-minuria: from early marker of renal damage to strong cardiovascular riskpredictor. J Hypertens 28: 2357–2369, 2010.
12. Crajoinas RO, Lessa LM, Carraro-Lacroix LR, Davel AP, PachecoBP, Rossoni LV, Malnic G, Girardi AC. Posttranslational mechanismsassociated with reduced NHE3 activity in adult vs. young prehypertensiveSHR. Am J Physiol Renal Physiol 299: F872–F881, 2010.
13. Czekay RP, Orlando RA, Woodward L, Lundstrom M, FarquharMG. Endocytic trafficking of megalin/RAP complexes: dissociation of thecomplexes in late endosomes. Mol Biol Cell 8: 517–532, 1997.
14. D’Amico G, Bazzi C. Pathophysiology of proteinuria. Kidney Int 63:809–825, 2003.
15. Furness PN, Hall LL, Shaw JA, Pringle JH. Glomerular expression ofnephrin is decreased in acquired human nephrotic syndrome. Nephrol DialTransplant 14: 1234–1237, 1999.
16. Futrakul N, Sridama V, Futrakul P. Microalbuminuria—a biomarker ofrenal microvascular disease. Ren Fail 31: 140–143, 2009.
17. Gekle M. Renal tubule albumin transport. Annu Rev Physiol 67: 573–594,2005.
18. Gekle M, Drumm K, Mildenberger S, Freudinger R, Gassner B,Silbernagl S. Inhibition of Na�-H� exchange impairs receptor-mediatedalbumin endocytosis in renal proximal tubule-derived epithelial cells fromopossum. J Physiol 520: 709–721, 1999.
19. Gekle M, Freudinger R, Mildenberger S. Inhibition of Na�-H� ex-changer-3 interferes with apical receptor-mediated endocytosis via vesiclefusion. J Physiol 531: 619–629, 2001.
20. Gekle M, Mildenberger S, Freudinger R, Silbernagl S. Endosomalalkalinization reduces Jmax and Km of albumin receptor-mediated endocy-tosis in OK cells. Am J Physiol Renal Fluid Electrolyte Physiol 268:F899–F906, 1995.
21. Gekle M, Serrano OK, Drumm K, Mildenberger S, Freudinger R,Gassner B, Jansen HW, Christensen EI. NHE3 serves as a moleculartool for cAMP-mediated regulation of receptor-mediated endocytosis. AmJ Physiol Renal Physiol 283: F549–F558, 2002.
22. Gekle M, Volker K, Mildenberger S, Freudinger R, Shull GE, Wi-emann M. NHE3 Na�/H� exchanger supports proximal tubular proteinreabsorption in vivo. Am J Physiol Renal Physiol 287: F469–F473, 2004.
23. Girardi AC, Di Sole F. Deciphering the mechanisms of the Na�/H�
exchanger-3 regulation in organ dysfunction. Am J Physiol Cell Physiol302: C1569–C1587, 2012.
24. Gunther W, Piwon N, Jentsch TJ. The ClC-5 chloride channel knock-out mouse - an animal model for Dent’s disease. Pflügers Arch 445:456–462, 2003.
25. Hara A, Wada T, Furuichi K, Sakai N, Kawachi H, Shimizu F,Shibuya M, Matsushima K, Yokoyama H, Egashira K, Kaneko S.Blockade of VEGF accelerates proteinuria, via decrease in nephrin ex-pression in rat crescentic glomerulonephritis. Kidney Int 69: 1986–1995,2006.
26. Hryciw DH, Ekberg J, Pollock CA, Poronnik P. ClC-5: a chloridechannel with multiple roles in renal tubular albumin uptake. Int J BiochemCell Biol 38: 1036–1042, 2006.
27. Hultstrom M. Development of structural kidney damage in spontaneouslyhypertensive rats. J Hypertens 30: 1087–1091, 2012.
28. Kestila M, Lenkkeri U, Mannikko M, Lamerdin J, McCready P,Putaala H, Ruotsalainen V, Morita T, Nissinen M, Herva R, Kashtan
CE, Peltonen L, Holmberg C, Olsen A, Tryggvason K. Positionallycloned gene for a novel glomerular protein—nephrin—is mutated incongenital nephrotic syndrome. Mol Cell 1: 575–582, 1998.
29. Koop K, Eikmans M, Baelde HJ, Kawachi H, De Heer E, Paul LC,Bruijn JA. Expression of podocyte-associated molecules in acquiredhuman kidney diseases. J Am Soc Nephrol 14: 2063–2071, 2003.
30. Kozyraki R, Fyfe J, Verroust PJ, Jacobsen C, Dautry-Varsat A,Gburek J, Willnow TE, Christensen EI, Moestrup SK. Megalin-dependent cubilin-mediated endocytosis is a major pathway for the apicaluptake of transferrin in polarized epithelia. Proc Natl Acad Sci USA 98:12491–12496, 2001.
31. Laemmli UK. Cleavage of structural proteins during the assembly of thehead of bacteriophage T4. Nature 227: 680–685, 1970.
32. Lloyd SE, Pearce SH, Fisher SE, Steinmeyer K, Schwappach B,Scheinman SJ, Harding B, Bolino A, Devoto M, Goodyer P, RigdenSP, Wrong O, Jentsch TJ, Craig IW, Thakker RV. A commonmolecular basis for three inherited kidney stone diseases. Nature 379:445–449, 1996.
33. Lowry OH, Rosebrough NJ, Farr AL, Randall RJ. Protein measure-ment with the Folin phenol reagent. J Biol Chem 193: 265–275, 1951.
34. Luimula P, Ahola H, Wang SX, Solin ML, Aaltonen P, Tikkanen I,Kerjaschki D, Holthofer H. Nephrin in experimental glomerular disease.Kidney Int 58: 1461–1468, 2000.
35. Magyar CE, Zhang Y, Holstein-Rathlou NH, McDonough AA. Prox-imal tubule Na transporter responses are the same during acute and chronichypertension. Am J Physiol Renal Physiol 279: F358–F369, 2000.
36. Marshansky V, Vinay P. Proton gradient formation in early endosomesfrom proximal tubules. Biochim Biophys Acta 1284: 171–180, 1996.
37. Martinez F, Mansego ML, Chaves FJ, Redon J. Genetic bases ofurinary albumin excretion and related traits in hypertension. J Hypertens28: 213–225, 2010.
38. Mellman I, Fuchs R, Helenius A. Acidification of the endocytic andexocytic pathways. Annu Rev Biochem 55: 663–700, 1986.
39. Miner JH. Renal basement membrane components. Kidney Int 56: 2016–2024, 1999.
40. Mogensen CE. Microalbuminuria as a predictor of clinical diabeticnephropathy. Kidney Int 31: 673–689, 1987.
41. Mountokalakis TD. The renal consequences of arterial hypertension.Kidney Int 51: 1639–1653, 1997.
42. Mukherjee S, Ghosh RN, Maxfield FR. Endocytosis. Physiol Rev 77:759–803, 1997.
43. Najafian B, Basgen JM, Mauer M. Estimating mean glomerular volumeusing two arbitrary parallel sections. J Am Soc Nephrol 13: 2697–2705,2002.
44. Obermuller N, Kranzlin B, Blum WF, Gretz N, Witzgall R. Anendocytosis defect as a possible cause of proteinuria in polycystic kidneydisease. Am J Physiol Renal Physiol 280: F244–F253, 2001.
45. Paunescu TG, Da Silva N, Marshansky V, McKee M, Breton S, BrownD. Expression of the 56-kDa B2 subunit isoform of the vacuolar H�-ATPase in proton-secreting cells of the kidney and epididymis. Am JPhysiol Cell Physiol 287: C149–C162, 2004.
46. Piwon N, Gunther W, Schwake M, Bosl MR, Jentsch TJ. ClC-5 Cl�
channel disruption impairs endocytosis in a mouse model for Dent’sdisease. Nature 408: 369–373, 2000.
47. Pollock CA, Poronnik P. Albumin transport and processing by theproximal tubule: physiology and pathophysiology. Curr Opin NephrolHypertens 16: 359–364, 2007.
48. Remuzzi G, Ruggenenti P, Benigni A. Understanding the nature of renaldisease progression. Kidney Int 51: 2–15, 1997.
49. Rodicio JL, Campo C, Ruilope LM. Microalbuminuria in essentialhypertension. Kidney Int Suppl 68: S51–S54, 1998.
50. Russo LM, Osicka TM, Brammar GC, Candido R, Jerums G, ComperWD. Renal processing of albumin in diabetes and hypertension in rats:possible role of TGF-beta1. Am J Nephrol 23: 61–70, 2003.
51. Russo LM, Sandoval RM, Campos SB, Molitoris BA, Comper WD,Brown D. Impaired tubular uptake explains albuminuria in early diabeticnephropathy. J Am Soc Nephrol 20: 489–494, 2009.
52. Russo LM, Sandoval RM, McKee M, Osicka TM, Collins AB, BrownD, Molitoris BA, Comper WD. The normal kidney filters nephrotic levelsof albumin retrieved by proximal tubule cells: retrieval is disrupted innephrotic states. Kidney Int 71: 504–513, 2007.
53. Rybak SL, Lanni F, Murphy RF. Theoretical considerations on the roleof membrane potential in the regulation of endosomal pH. Biophys J 73:674–687, 1997.
F225RENAL TUBULAR PROTEINURIA IN SHR
AJP-Renal Physiol • doi:10.1152/ajprenal.00255.2012 • www.ajprenal.org
by Adriana G
irardi on July 16, 2013http://ajprenal.physiology.org/
Dow
nloaded from

54. Storrie B, Desjardins M. The biogenesis of lysosomes: is it a kiss andrun, continuous fusion and fission process? Bioessays 18: 895–903, 1996.
55. Tanaka K, Nakaki T. Reduced renal ClC-5 Cl� channel expression inspontaneously hypertensive rats with microalbuminuria. Eur J Pharmacol501: 185–189, 2004.
56. Thakker RV. The role of renal chloride channel mutations in kidney stonedisease and nephrocalcinosis. Curr Opin Nephrol Hypertens 7: 385–388, 1998.
57. Tojo A, Endou H. Intrarenal handling of proteins in rats using fractionalmicropuncture technique. Am J Physiol Renal Fluid Electrolyte Physiol263: F601–F606, 1992.
58. Tojo A, Onozato ML, Ha H, Kurihara H, Sakai T, Goto A, Fujita T,Endou H. Reduced albumin reabsorption in the proximal tubule ofearly-stage diabetic rats. Histochem Cell Biol 116: 269–276, 2001.
59. Tryggvason K. Unraveling the mechanisms of glomerular ultrafiltration:nephrin, a key component of the slit diaphragm. J Am Soc Nephrol 10:2440–2445, 1999.
60. Viberti GC, Hill RD, Jarrett RJ, Argyropoulos A, Mahmud U, KeenH. Microalbuminuria as a predictor of clinical nephropathy in insulin-dependent diabetes mellitus. Lancet 1: 1430–1432, 1982.
61. Vogel C, Abreu Rde S, Ko D, Le SY, Shapiro BA, Burns SC, SandhuD, Boutz DR, Marcotte EM, Penalva LO. Sequence signatures andmRNA concentration can explain two-thirds of protein abundance varia-tion in a human cell line. Mol Syst Biol 6: 400, 2010.
62. Willnow TE, Hilpert J, Armstrong SA, Rohlmann A, Hammer RE,Burns DK, Herz J. Defective forebrain development in mice lackinggp330/megalin. Proc Natl Acad Sci USA 93: 8460–8464, 1996.
F226 RENAL TUBULAR PROTEINURIA IN SHR
AJP-Renal Physiol • doi:10.1152/ajprenal.00255.2012 • www.ajprenal.org
by Adriana G
irardi on July 16, 2013http://ajprenal.physiology.org/
Dow
nloaded from
Recommended
















