
International Dairy Journal 14 (2004) 889–898
ARTICLE IN PRESS
*Correspondin
E-mail addres
0958-6946/$ - see
doi:10.1016/j.ida
Release of angiotensin converting enzyme-inhibitory peptides bysimulated gastrointestinal digestion of infant formulas
Blanca Hern!andez-Ledesma, Lourdes Amigo, Mercedes Ramos, Isidra Recio*
Instituto de Fermentaciones Industriales (CSIC), Juan de la Cierva 3, Madrid 28006, Spain
Received 14 November 2003; accepted 25 February 2004
Abstract
The angiotensin converting enzyme (ACE)-inhibitory activity of several infant formulas was evaluated. Most of these products
showed moderate inhibitory activity, but two exceptions that corresponded to an extensively hydrolysed whey formula and an
extensively hydrolysed casein formula were detected. Two products (a non-hydrolysed milk protein-based formula and an
extensively hydrolysed whey formula) were subjected to a two-stage in vitro enzymatic procedure, which simulates physiological
digestion, in order to study the impact of digestion on ACE-inhibitory activity. The ACE-inhibitory activity of the non-hydrolysed
formula increased during simulated gastrointestinal digestion, while no significant change was observed in the activity of the
hydrolysed whey formula prior to and after, digestion. The peptides generated from these two products during simulated
physiological digestion were sequenced by tandem spectrometry. At the end of the digestion, most peptides found in the non-
hydrolysed milk protein-based formula were formed during incubation with the pancreatic extract, but, in the hydrolysed whey
formula, many peptides present in the undigested product survived simulated digestion. The potential ACE-inhibitory activity of
these peptides is discussed with regard to their amino-acid sequences.
r 2004 Elsevier Ltd. All rights reserved.
Keywords: ACE-inhibitory peptides; Infant formula; Simulated gastrointestinal digestion; Mass spectrometry; Peptide sequencing
1. Introduction
Infant formulas are designed to simulate not only thecontent of, but also the performance of, human milk asmuch as possible, in order to be an adequate replace-ment of human milk. The most common sources ofprotein in infant formulas are either cows’ milk or soyprotein. In some cases, in order to diminish the risk ofprotein allergy, these products are formulated fromdifferent types of milk protein hydrolysates (Caffarelliet al., 2002).It is now accepted that enzymatic hydrolysis of food
protein releases peptides that may exhibit differentbiological activities. These protein fragments, knownas bioactive peptides, can be formed from the precursorinactive protein during gastrointestinal digestion and/orduring food processing (Meisel, 1997). Among thedifferent groups of bioactive peptides, angiotensinconverting enzyme (ACE)-inhibitory peptides are re-
g author. Tel.: +34-91-5622900; fax: +34-91-5644853.
s: [email protected] (I. Recio).
front matter r 2004 Elsevier Ltd. All rights reserved.
iryj.2004.02.011
ceiving special attention due to their potential beneficialeffects related to hypertension. The formation of ACE-inhibitory peptides by enzymatic hydrolysis (Pihlanto-Lepp.al.a, Koskinen, Piilola, Tupasela, & Korhonen,2000; Hern!andez-Ledesma, Recio, Ramos, & Amigo,2002), by milk fermentation (Nakamura, Yamamoto,Sakai, & Takano, 1995; Gobbetti, Ferranti, Smacchi,Goffredi, & Addeo, 2000; Leclerc, Gauthier, Bachelard,Santure, & Roy, 2002) and during cheese ripening(Saito, Nakamura, Kitazawa, Kawai, & Itoh, 2000;G !omez-Ruiz, Ramos, & Recio, 2002) has extensivelybeen reported.However, once the ACE-inhibitory peptides are
released by food processing they have to be able tosurvive the gastrointestinal system, be absorbed andreach the cardiovascular system in an active form.Several studies have revealed the importance of gastro-intestinal digestion on ACE-inhibitory peptides forma-tion. For instance Maeno, Yamamoto, and Takano(1996) identified a potent in vivo antihypertensivepeptide that exhibits low in vitro ACE-inhibitoryactivity. These authors found that a shorter peptide

ARTICLE IN PRESSB. Hern !andez-Ledesma et al. / International Dairy Journal 14 (2004) 889–898890
with potent ACE-inhibitory activity was liberated bypacreatic digestion from the longer form. Similarly,other studies have showed an increase in ACE-inhibitory activity by the action of digestive enzymeson fermented casein solutions (Pihlanto-Lepp.al.a,Rokka, & Korhonen, 1998; Vermeirssen, Van Camp,Decroos, Van Wijmelbeke, & Verstraete, 2003).However, difficulties in peptide identification limit the
knowledge available on the formation of bioactivepeptides and their release from the precursor proteins.Several chromatographic steps are often necessary topurify the peptides of interest and finally they areidentified by mass spectrometry (MS) alone or MScombined with N-terminal sequence analysis. Thedevelopment of routine and reliable liquid chromato-graphy-MS instruments made it possible to analysepeptides included in complex mixtures without the needto separate the individual components (Papayannopou-los, 1995). If the precursor proteins are known,the fragmentation spectra can be matched to thesequences of the selected peptides with a given mass,thereby allowing the unambiguous identification ofpeptides from unfractionated enzyme digests of proteins(Biemann & Scoble, 1987; McLuckey, Van Berkel,Glish, Huang, & Henion, 1991).The aim of our study was first to investigate the
presence of ACE-inhibitory substances, as naturallyoccurring components in commercial infant formulas.Two of these products were subjected to an enzymatichydrolysis process, which simulates physiological diges-tion, in order to study the formation of ACE-inhibitorypeptides. The peptides generated from both productsduring simulated physiological digestion were sequencedby tandem MS. The potential ACE-inhibitory activity ofthese peptides is discussed in relation to their structure.
2. Materials and methods
2.1. Samples and simulation of gastrointestinal digestion
Milk protein and soy-based infant formulas (IF-1—IF-11) were purchased on the Spanish market. Infantformulas IF-2 and IF-7 corresponded to enzymaticallyhydrolysed casein formula and enzymatically hydro-lysed whey proteins, respectively. Infant formula IF-5was a soy proteins-based formula and the other infantformulas analysed in the study contained mainly non-hydrolysed milk proteins.Two infant formulas (IF-1 and IF-7) were selected to
simulate gastrointestinal digestion. Hydrolysates wereprepared from an aqueous solution of the formulas IF-1and IF-7 (0.7%, w/v, protein). The hydrolysis wascarried out according to Alting, Meijer, and vanBeresteijn (1997). The samples were first hydrolysedwith pepsin (EC 3.4.4.1; 1:60,000, 3400Umg�1) (Sigma
Chemical, St. Louis, MO, USA), which was added at alevel of 20mg g�1 protein, for 90min at 37�C at pH 3.5followed by hydrolysis with Corolase PPs (R .ohm,Darmstadt, Germany), which was added at a level of40mg g�1 protein, at pH approx. 7.5 and 37�C for240min. Corolase PPs is a proteolytic enzyme prepara-tion from pig pancreas glands that contains, in additionto trypsin and chymotrypsin, numerous amino- andcarboxipeptidase activities. Hydrolysis was carried outin a thermally controlled water bath under constantstirring. Aliquots were withdrawn after hydrolysis withpepsin, the pH was raised to 7.5 with 1m NaOH andthey were heated at 95�C for 10min in a water bath.During hydrolysis with Corolase PPs, aliquots werealso taken at 30, 120 and 240min.The enzyme wasinactivated by heating at 95�C for 10min, followed bycooling to room temperature. Each sample was stored at�20�C until further analysis.After the last samples had been taken, aliquots and
the remaining reaction mixtures were centrifuged at10,000� g for 30min and the supernatants weresubjected to ultrafiltration through an hydrophilic3000 Da cut-off membrane (Centripep, Amicon, Inc.,Beverly, MA, USA). The permeates were freeze-driedand kept at �20�C until required.Water-soluble extracts (WSE) of the infant formulas
were obtained by dissolving 4.5 g of the product in30mL of distilled water. The pH was adjusted to 4.6with 1m HCl and the reconstituted formula was thencentrifuged at 12,000� g for 20min at 5�C and filteredthrough a Whatman no. 40 filter.
2.2. Measurement of ACE-inhibitory activity
ACE-inhibitory activity was measured by the spectro-photometric assay of Cushman and Cheung (1971), asmodified by Kim, Yoon, Yu, L .onnerdal, and Chung(1999). Briefly, 20 mL of each sample was added to0.1mL of 0.1m potasium phosphate buffer (pH 8.3)containing 0.3m NaCl, and 5mm hippuryl–histydil–leucine (Sigma Chemical, St. Louis, MO, USA). ACE (5mU) (EC 3.4.15.1, 5.1 Umg�1, Sigma) was added andthe reaction mixture was incubated at 37�C for30min.The reaction was terminated by the addition of0.1mL 1m HCl. The hippuric acid formed was extractedwith ethyl acetate, heat-evaporated at 95�C for 10min,redissolved in distilled water and measured spectro-photometrically at 228 nm.The activity of each samplewas tested in triplicate.The ACE-inhibitory activity of those products with
ACE-inhibitory indexes higher than 50% was alsocalculated as the protein concentration needed to inhibit50% the original ACE activity (IC50), and one unit ofACE-inhibitory activity was expressed as the potencyshowing 50% ACE inhibition under these conditions.
ARTICLE IN PRESS
20
40
60
80
100
IAC
E (
%)
0
50
100
150
200
250
Pep
tide
nitr
ogen
(m
g 10
0g-1
)
IF-1
IF-3
IF-5
IF-7
IF-9
IF-11
IF-2
IF-4
IF-6
IF-8
IF-10
B. Hern !andez-Ledesma et al. / International Dairy Journal 14 (2004) 889–898 891
The total nitrogen content of the WSE was deter-mined by the Kjeldahl method. Amino acid nitrogenwas measured using the Cd-nynhidrin method accordingto Doi, Shibata, and Matoba (1981). The content ofpeptide nitrogen was calculated as the difference of totalnitrogen minus amino-acid nitrogen.
2.3. Analysis by on-line reversed-phase high performance
liquid chromatography coupled on line to tandem mass
spectrometry (RP-HPLC-MS/MS)
RP-HPLC separations of the WSE were performed ona Agilent HPLC system connected on line to an Esquire-LC quadrupole ion trap instrument (Bruker DaltonikGmbH, Bremen, Germany). The HPLC system wasequipped with a quaternary gradient pumping system, anin-line degasser, a variable wavelength absorbancedetector set at 220nm, and an automatic injector (all1100 Series, Agilent Technologies, Waldbronn, Ger-many). The column used in these experiments was a250mm� 4.6mm Widepore C18 column (Bio-Rad, Rich-mond, CA, USA). The injection volume was 50mL.Solvent A was a mixture of water and trifluoroacetic acidat a v/v ratio of 1000–0.37, and solvent B containedacetonitrile and trifluoroacetic acid at a v/v ratio of 1000–0.27. Peptides were eluted with a linear gradient of solventB in A going from 0% to 45% in 60min at a flow rate of0.8mLmin�1. The flow was split post detector byconnecting a T-piece (Valco, Houston, TX, USA) witha 75mm ID peek outlet tube, of an adjusted length, to givea flow of approx. 20mL min�1 which was directed into themass spectrometer via the electrospray interface. Nitrogenwas used as nebulizing agent and drying gas and operatedwith an estimated helium pressure of 5� 10�3 bar. Thecapillary was held at 4 kV. Spectra were recorded overthe mass/charge (m/z) range 100–2500. About 25 spectrawere averaged in the MS analyses and about 5 spectra inthe MS(n) analyses. The signal threshold to perform autoMS(n) analyses was 10000 (i.e., 5% of the total signal)and the precursor ions were isolated within a range of 4.0m/z and fragmented with a voltage ramp going from 0.35to 1.4V. Using Data AnalysisTM (version 3.0; BrukerDaltoniks, GmbH, Bremen, Germany), the m/z spectraldata were processed and transformed to spectra repre-senting mass values. BioTools (version 2.1; BrukerDaltoniks) was used to process the MS(n) spectra andperform peptide sequencing.
0 IF-1
IF-3
IF-5
IF-7
IF-9
IF-11
IF-2
IF-4
IF-6
IF-8
IF-10
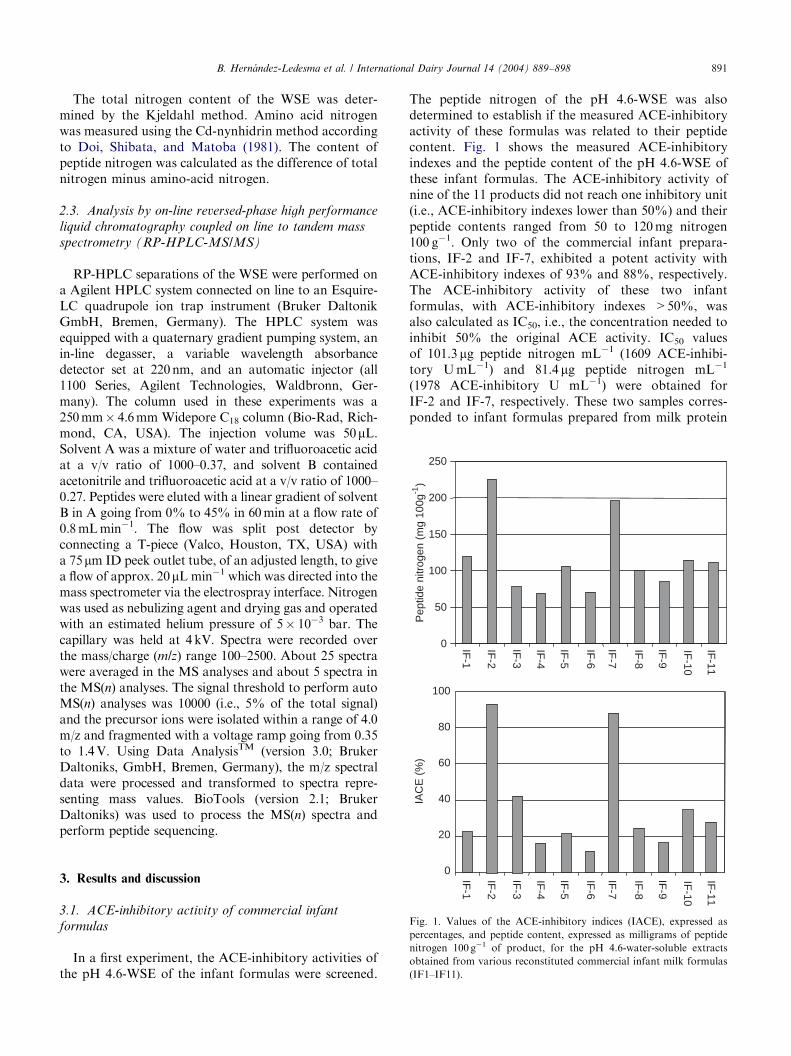
Fig. 1. Values of the ACE-inhibitory indices (IACE), expressed as
percentages, and peptide content, expressed as milligrams of peptide
nitrogen 100 g�1 of product, for the pH 4.6-water-soluble extracts
obtained from various reconstituted commercial infant milk formulas
(IF1–IF11).
3. Results and discussion
3.1. ACE-inhibitory activity of commercial infant
formulas
In a first experiment, the ACE-inhibitory activities ofthe pH 4.6-WSE of the infant formulas were screened.
The peptide nitrogen of the pH 4.6-WSE was alsodetermined to establish if the measured ACE-inhibitoryactivity of these formulas was related to their peptidecontent. Fig. 1 shows the measured ACE-inhibitoryindexes and the peptide content of the pH 4.6-WSE ofthese infant formulas. The ACE-inhibitory activity ofnine of the 11 products did not reach one inhibitory unit(i.e., ACE-inhibitory indexes lower than 50%) and theirpeptide contents ranged from 50 to 120mg nitrogen100 g�1. Only two of the commercial infant prepara-tions, IF-2 and IF-7, exhibited a potent activity withACE-inhibitory indexes of 93% and 88%, respectively.The ACE-inhibitory activity of these two infantformulas, with ACE-inhibitory indexes >50%, wasalso calculated as IC50, i.e., the concentration needed toinhibit 50% the original ACE activity. IC50 valuesof 101.3 mg peptide nitrogen mL�1 (1609 ACE-inhibi-tory UmL�1) and 81.4 mg peptide nitrogen mL�1
(1978 ACE-inhibitory U mL�1) were obtained forIF-2 and IF-7, respectively. These two samples corres-ponded to infant formulas prepared from milk protein
ARTICLE IN PRESSB. Hern !andez-Ledesma et al. / International Dairy Journal 14 (2004) 889–898892
hydrolysates, and consequently, the peptide content ofthese samples (196.3 and 226mg nitrogen 100 g�1) washigher than that observed in the pH 4.6-WSE of theother infant formulas. Results of the RP-HPLC analysesof these pH 4.6-WSE (data not shown) stronglysupported the results of the peptide nitrogen analysis.To date, there is little information available on theACE-inhibitory activity in infant milk formulas, butMeisel, Goepfert, and G .unther (1997) found an ACE-inhibitory index of 31.4% in an hypoallergenic infantfood, a value which concurs with the activity found inmost samples considered in this study.
3.2. Hydrolysis under simulated gastrointestinal
conditions
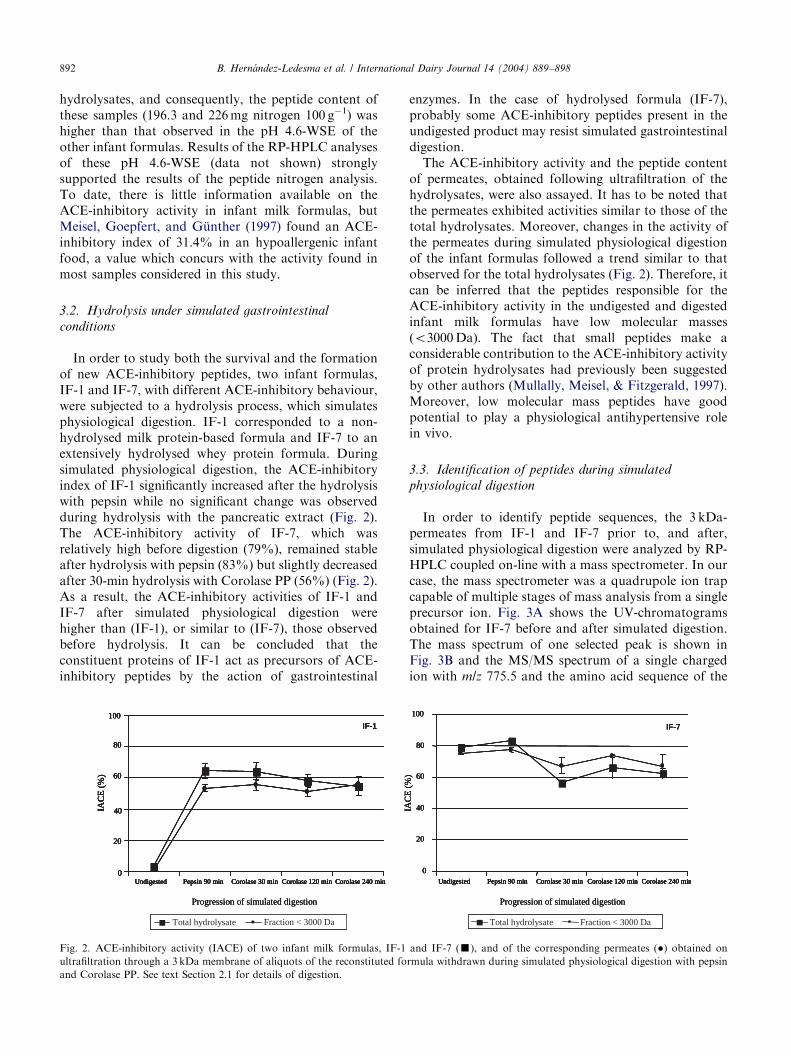
In order to study both the survival and the formationof new ACE-inhibitory peptides, two infant formulas,IF-1 and IF-7, with different ACE-inhibitory behaviour,were subjected to a hydrolysis process, which simulatesphysiological digestion. IF-1 corresponded to a non-hydrolysed milk protein-based formula and IF-7 to anextensively hydrolysed whey protein formula. Duringsimulated physiological digestion, the ACE-inhibitoryindex of IF-1 significantly increased after the hydrolysiswith pepsin while no significant change was observedduring hydrolysis with the pancreatic extract (Fig. 2).The ACE-inhibitory activity of IF-7, which wasrelatively high before digestion (79%), remained stableafter hydrolysis with pepsin (83%) but slightly decreasedafter 30-min hydrolysis with Corolase PP (56%) (Fig. 2).As a result, the ACE-inhibitory activities of IF-1 andIF-7 after simulated physiological digestion werehigher than (IF-1), or similar to (IF-7), those observedbefore hydrolysis. It can be concluded that theconstituent proteins of IF-1 act as precursors of ACE-inhibitory peptides by the action of gastrointestinal
Progression of simulated digestion
Undigested Pepsin 90 min Corolase 30 min Corolase 120 min Corolase 240 min0
IF-1
20
40
60
80
100
IAC
E (
%)
Total hydrolysate Fraction < 3000 Da
Progression of simulated digestion
Undigested Pepsin 90 min Corolase 30 min Corolase 120 min Corolase 240 min0
IF-1
20
40
60
80
100
IAC
E (
%)
Total hydrolysate Fraction < 3000 Da
Progression of simulated digestion
Undigested Pepsin 90 min Corolase 30 min Corolase 120 min Corolase 240 min0
IF-1
20
40
60
80
100
IAC
E (
%)
Undigested Pepsin 90 min Corolase 30 min Corolase 120 min Corolase 240 minUndigested Pepsin 90 min Corolase 30 min Corolase 120 min Corolase 240 min0
IF-1
20
40
60
80
100
IAC
E (
%)
0
IF-1
20
40
60
80
100
IAC
E (
%)
Total hydrolysate Fraction < 3000 DaTotal hydrolysate Fraction < 3000 Da
Fig. 2. ACE-inhibitory activity (IACE) of two infant milk formulas, IF-1
ultrafiltration through a 3 kDa membrane of aliquots of the reconstituted fo
and Corolase PP. See text Section 2.1 for details of digestion.
enzymes. In the case of hydrolysed formula (IF-7),probably some ACE-inhibitory peptides present in theundigested product may resist simulated gastrointestinaldigestion.The ACE-inhibitory activity and the peptide content
of permeates, obtained following ultrafiltration of thehydrolysates, were also assayed. It has to be noted thatthe permeates exhibited activities similar to those of thetotal hydrolysates. Moreover, changes in the activity ofthe permeates during simulated physiological digestionof the infant formulas followed a trend similar to thatobserved for the total hydrolysates (Fig. 2). Therefore, itcan be inferred that the peptides responsible for theACE-inhibitory activity in the undigested and digestedinfant milk formulas have low molecular masses(o3000Da). The fact that small peptides make aconsiderable contribution to the ACE-inhibitory activityof protein hydrolysates had previously been suggestedby other authors (Mullally, Meisel, & Fitzgerald, 1997).Moreover, low molecular mass peptides have goodpotential to play a physiological antihypertensive rolein vivo.
3.3. Identification of peptides during simulated
physiological digestion
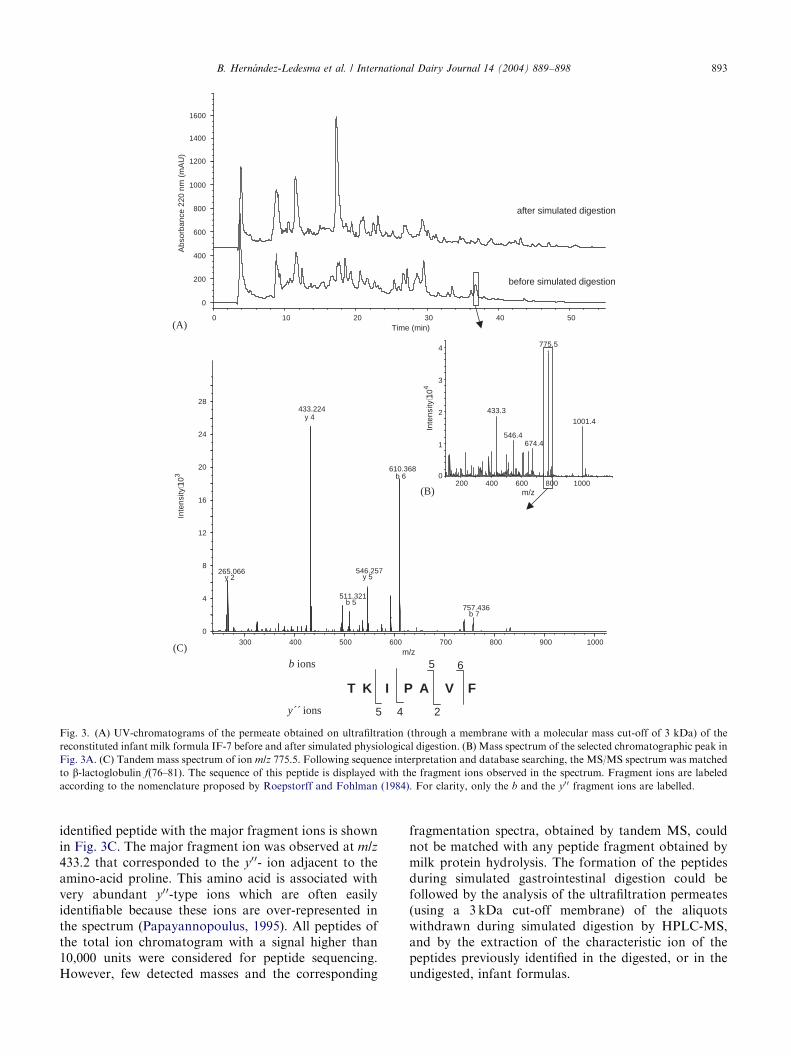
In order to identify peptide sequences, the 3 kDa-permeates from IF-1 and IF-7 prior to, and after,simulated physiological digestion were analyzed by RP-HPLC coupled on-line with a mass spectrometer. In ourcase, the mass spectrometer was a quadrupole ion trapcapable of multiple stages of mass analysis from a singleprecursor ion. Fig. 3A shows the UV-chromatogramsobtained for IF-7 before and after simulated digestion.The mass spectrum of one selected peak is shown inFig. 3B and the MS/MS spectrum of a single chargedion with m/z 775.5 and the amino acid sequence of the
Progression of simulated digestion
Total hydrolysate Fraction < 3000 Da
IF-7
0
20
40
60
80
100
IAC
E (
%)
Undigested Pepsin 90 min Corolase 30 min Corolase 120 min Corolase 240 min
Progression of simulated digestion
Total hydrolysate Fraction < 3000 Da
IF-7
0
20
40
60
80
100
IAC
E (
%)
Undigested Pepsin 90 min Corolase 30 min Corolase 120 min Corolase 240 min
Progression of simulated digestion
Total hydrolysate Fraction < 3000 DaTotal hydrolysate Fraction < 3000 Da
IF-7
0
20
40
60
80
100
IAC
E (
%)
Undigested Pepsin 90 min Corolase 30 min Corolase 120 min Corolase 240 min
IF-7
0
20
40
60
80
100
IAC
E (
%)
Undigested Pepsin 90 min Corolase 30 min Corolase 120 min Corolase 240 minUndigested Pepsin 90 min Corolase 30 min Corolase 120 min Corolase 240 min
and IF-7 (’), and of the corresponding permeates (�) obtained on
rmula withdrawn during simulated physiological digestion with pepsin
ARTICLE IN PRESS
1600
Time (min)0 10 20 30 40 50
0
200
400
600
800
1000
1200
1400
m/z300 400 500 600 700 800 900 1000
0
4
8
12
16
20
24
28
265.066y 2
433.224y 4
511.321b 5
546.257y 5
610.368b 6
757.436b 7
T
b ions
y´´ ions 5
6
2
5
433.3
546.4674.4
775.5
1001.4
0
1
2
3
4
Inte
nsity
⋅104
200 400 600 800 1000m/z
4
Inte
nsity
⋅103
Abs
orba
nce
220
nm (
mA
U)
(A)
(C)
(B)
before simulated digestion
after simulated digestion
FVAPIK
Fig. 3. (A) UV-chromatograms of the permeate obtained on ultrafiltration (through a membrane with a molecular mass cut-off of 3 kDa) of the
reconstituted infant milk formula IF-7 before and after simulated physiological digestion. (B) Mass spectrum of the selected chromatographic peak in
Fig. 3A. (C) Tandem mass spectrum of ion m/z 775.5. Following sequence interpretation and database searching, the MS/MS spectrum was matched
to b-lactoglobulin f(76–81). The sequence of this peptide is displayed with the fragment ions observed in the spectrum. Fragment ions are labeled
according to the nomenclature proposed by Roepstorff and Fohlman (1984). For clarity, only the b and the y0 0 fragment ions are labelled.
B. Hern !andez-Ledesma et al. / International Dairy Journal 14 (2004) 889–898 893
identified peptide with the major fragment ions is shownin Fig. 3C. The major fragment ion was observed at m/z
433.2 that corresponded to the y00- ion adjacent to theamino-acid proline. This amino acid is associated withvery abundant y00-type ions which are often easilyidentifiable because these ions are over-represented inthe spectrum (Papayannopoulus, 1995). All peptides ofthe total ion chromatogram with a signal higher than10,000 units were considered for peptide sequencing.However, few detected masses and the corresponding
fragmentation spectra, obtained by tandem MS, couldnot be matched with any peptide fragment obtained bymilk protein hydrolysis. The formation of the peptidesduring simulated gastrointestinal digestion could befollowed by the analysis of the ultrafiltration permeates(using a 3 kDa cut-off membrane) of the aliquotswithdrawn during simulated digestion by HPLC-MS,and by the extraction of the characteristic ion of thepeptides previously identified in the digested, or in theundigested, infant formulas.
ARTICLE IN PRESS
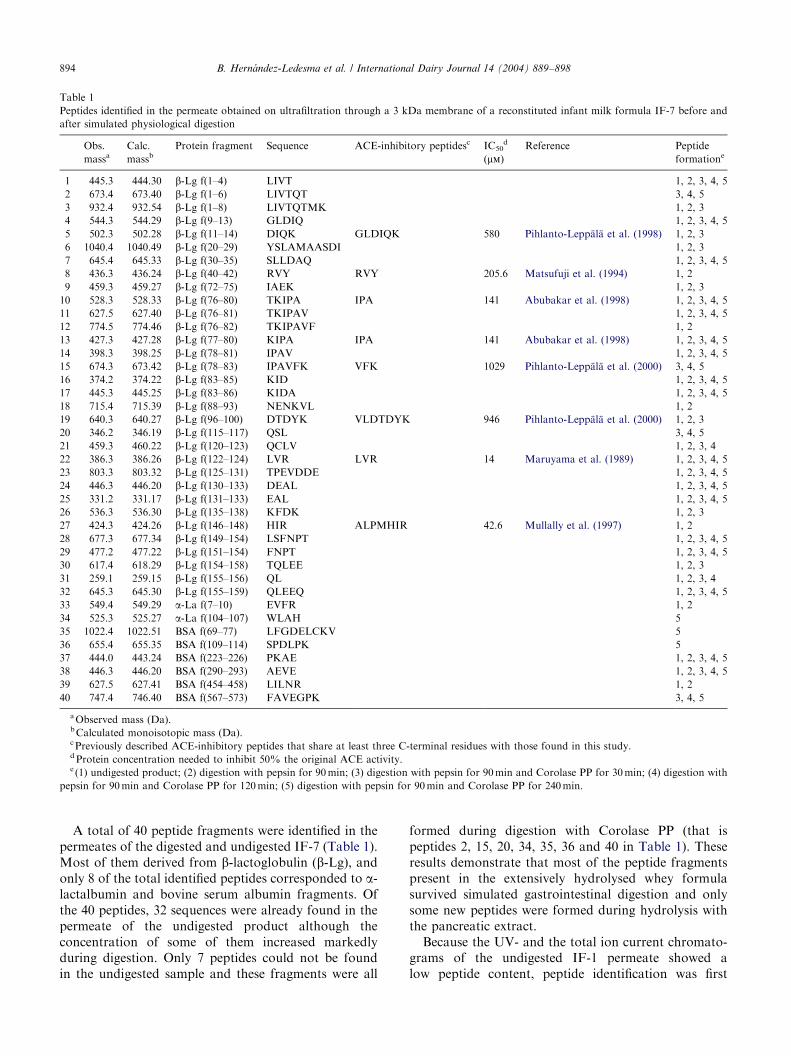
Table 1
Peptides identified in the permeate obtained on ultrafiltration through a 3 kDa membrane of a reconstituted infant milk formula IF-7 before and
after simulated physiological digestion
Obs.
massaCalc.
massbProtein fragment Sequence ACE-inhibitory peptidesc IC50
d
(mm)Reference Peptide
formatione
1 445.3 444.30 b-Lg f(1–4) LIVT 1, 2, 3, 4, 5
2 673.4 673.40 b-Lg f(1–6) LIVTQT 3, 4, 5
3 932.4 932.54 b-Lg f(1–8) LIVTQTMK 1, 2, 3
4 544.3 544.29 b-Lg f(9–13) GLDIQ 1, 2, 3, 4, 5
5 502.3 502.28 b-Lg f(11–14) DIQK GLDIQK 580 Pihlanto-Lepp.al.a et al. (1998) 1, 2, 3
6 1040.4 1040.49 b-Lg f(20–29) YSLAMAASDI 1, 2, 3
7 645.4 645.33 b-Lg f(30–35) SLLDAQ 1, 2, 3, 4, 5
8 436.3 436.24 b-Lg f(40–42) RVY RVY 205.6 Matsufuji et al. (1994) 1, 2
9 459.3 459.27 b-Lg f(72–75) IAEK 1, 2, 3
10 528.3 528.33 b-Lg f(76–80) TKIPA IPA 141 Abubakar et al. (1998) 1, 2, 3, 4, 5
11 627.5 627.40 b-Lg f(76–81) TKIPAV 1, 2, 3, 4, 5
12 774.5 774.46 b-Lg f(76–82) TKIPAVF 1, 2
13 427.3 427.28 b-Lg f(77–80) KIPA IPA 141 Abubakar et al. (1998) 1, 2, 3, 4, 5
14 398.3 398.25 b-Lg f(78–81) IPAV 1, 2, 3, 4, 5
15 674.3 673.42 b-Lg f(78–83) IPAVFK VFK 1029 Pihlanto-Lepp.al.a et al. (2000) 3, 4, 5
16 374.2 374.22 b-Lg f(83–85) KID 1, 2, 3, 4, 5
17 445.3 445.25 b-Lg f(83–86) KIDA 1, 2, 3, 4, 5
18 715.4 715.39 b-Lg f(88–93) NENKVL 1, 2
19 640.3 640.27 b-Lg f(96–100) DTDYK VLDTDYK 946 Pihlanto-Lepp.al.a et al. (2000) 1, 2, 3
20 346.2 346.19 b-Lg f(115–117) QSL 3, 4, 5
21 459.3 460.22 b-Lg f(120–123) QCLV 1, 2, 3, 4
22 386.3 386.26 b-Lg f(122–124) LVR LVR 14 Maruyama et al. (1989) 1, 2, 3, 4, 5
23 803.3 803.32 b-Lg f(125–131) TPEVDDE 1, 2, 3, 4, 5
24 446.3 446.20 b-Lg f(130–133) DEAL 1, 2, 3, 4, 5
25 331.2 331.17 b-Lg f(131–133) EAL 1, 2, 3, 4, 5
26 536.3 536.30 b-Lg f(135–138) KFDK 1, 2, 3
27 424.3 424.26 b-Lg f(146–148) HIR ALPMHIR 42.6 Mullally et al. (1997) 1, 2
28 677.3 677.34 b-Lg f(149–154) LSFNPT 1, 2, 3, 4, 5
29 477.2 477.22 b-Lg f(151–154) FNPT 1, 2, 3, 4, 5
30 617.4 618.29 b-Lg f(154–158) TQLEE 1, 2, 3
31 259.1 259.15 b-Lg f(155–156) QL 1, 2, 3, 4
32 645.3 645.30 b-Lg f(155–159) QLEEQ 1, 2, 3, 4, 5
33 549.4 549.29 a-La f(7–10) EVFR 1, 2
34 525.3 525.27 a-La f(104–107) WLAH 5
35 1022.4 1022.51 BSA f(69–77) LFGDELCKV 5
36 655.4 655.35 BSA f(109–114) SPDLPK 5
37 444.0 443.24 BSA f(223–226) PKAE 1, 2, 3, 4, 5
38 446.3 446.20 BSA f(290–293) AEVE 1, 2, 3, 4, 5
39 627.5 627.41 BSA f(454–458) LILNR 1, 2
40 747.4 746.40 BSA f(567–573) FAVEGPK 3, 4, 5
aObserved mass (Da).bCalculated monoisotopic mass (Da).cPreviously described ACE-inhibitory peptides that share at least three C-terminal residues with those found in this study.dProtein concentration needed to inhibit 50% the original ACE activity.e (1) undigested product; (2) digestion with pepsin for 90min; (3) digestion with pepsin for 90min and Corolase PP for 30min; (4) digestion with
pepsin for 90min and Corolase PP for 120min; (5) digestion with pepsin for 90min and Corolase PP for 240min.
B. Hern !andez-Ledesma et al. / International Dairy Journal 14 (2004) 889–898894
A total of 40 peptide fragments were identified in thepermeates of the digested and undigested IF-7 (Table 1).Most of them derived from b-lactoglobulin (b-Lg), andonly 8 of the total identified peptides corresponded to a-lactalbumin and bovine serum albumin fragments. Ofthe 40 peptides, 32 sequences were already found in thepermeate of the undigested product although theconcentration of some of them increased markedlyduring digestion. Only 7 peptides could not be foundin the undigested sample and these fragments were all
formed during digestion with Corolase PP (that ispeptides 2, 15, 20, 34, 35, 36 and 40 in Table 1). Theseresults demonstrate that most of the peptide fragmentspresent in the extensively hydrolysed whey formulasurvived simulated gastrointestinal digestion and onlysome new peptides were formed during hydrolysis withthe pancreatic extract.Because the UV- and the total ion current chromato-
grams of the undigested IF-1 permeate showed alow peptide content, peptide identification was first
ARTICLE IN PRESS
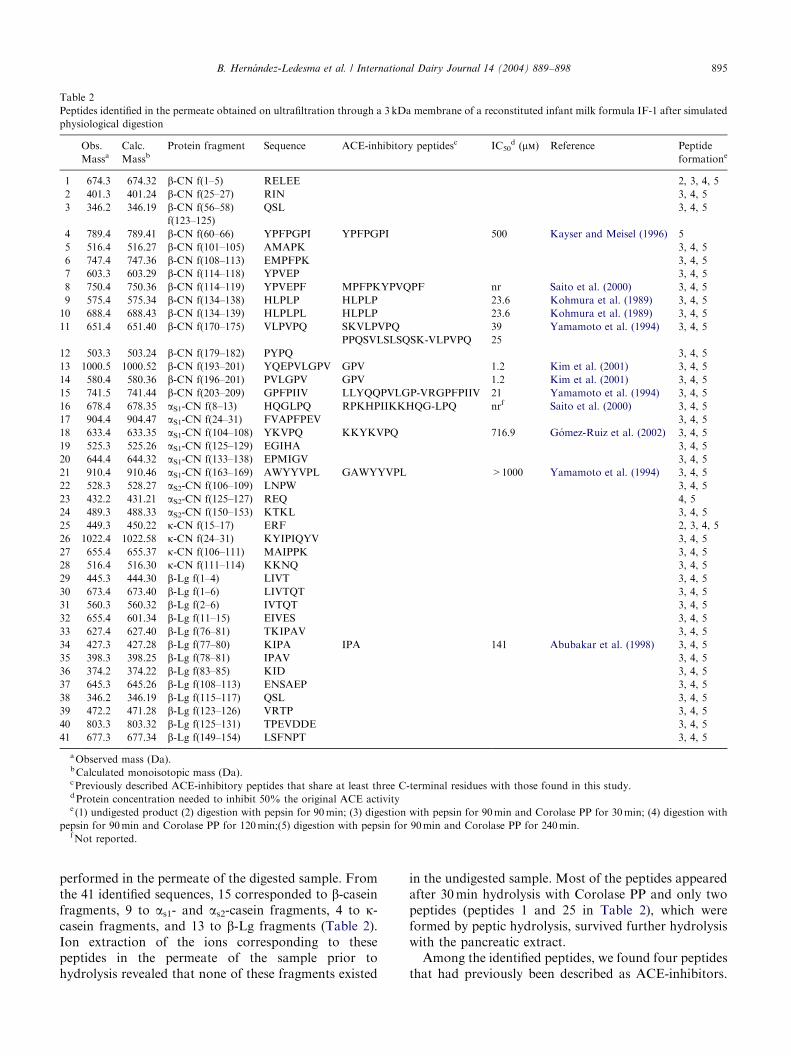
Table 2
Peptides identified in the permeate obtained on ultrafiltration through a 3 kDa membrane of a reconstituted infant milk formula IF-1 after simulated
physiological digestion
Obs.
MassaCalc.
MassbProtein fragment Sequence ACE-inhibitory peptidesc IC50
d (mm) Reference Peptide
formatione
1 674.3 674.32 b-CN f(1–5) RELEE 2, 3, 4, 5
2 401.3 401.24 b-CN f(25–27) RIN 3, 4, 5
3 346.2 346.19 b-CN f(56–58) QSL 3, 4, 5
f(123–125)
4 789.4 789.41 b-CN f(60–66) YPFPGPI YPFPGPI 500 Kayser and Meisel (1996) 5
5 516.4 516.27 b-CN f(101–105) AMAPK 3, 4, 5
6 747.4 747.36 b-CN f(108–113) EMPFPK 3, 4, 5
7 603.3 603.29 b-CN f(114–118) YPVEP 3, 4, 5
8 750.4 750.36 b-CN f(114–119) YPVEPF MPFPKYPVQPF nr Saito et al. (2000) 3, 4, 5
9 575.4 575.34 b-CN f(134–138) HLPLP HLPLP 23.6 Kohmura et al. (1989) 3, 4, 5
10 688.4 688.43 b-CN f(134–139) HLPLPL HLPLP 23.6 Kohmura et al. (1989) 3, 4, 5
11 651.4 651.40 b-CN f(170–175) VLPVPQ SKVLPVPQ 39 Yamamoto et al. (1994) 3, 4, 5
PPQSVLSLSQSK-VLPVPQ 25
12 503.3 503.24 b-CN f(179–182) PYPQ 3, 4, 5
13 1000.5 1000.52 b-CN f(193–201) YQEPVLGPV GPV 1.2 Kim et al. (2001) 3, 4, 5
14 580.4 580.36 b-CN f(196–201) PVLGPV GPV 1.2 Kim et al. (2001) 3, 4, 5
15 741.5 741.44 b-CN f(203–209) GPFPIIV LLYQQPVLGP-VRGPFPIIV 21 Yamamoto et al. (1994) 3, 4, 5
16 678.4 678.35 aS1-CN f(8–13) HQGLPQ RPKHPIIKKHQG-LPQ nrf Saito et al. (2000) 3, 4, 5
17 904.4 904.47 aS1-CN f(24–31) FVAPFPEV 3, 4, 5
18 633.4 633.35 aS1-CN f(104–108) YKVPQ KKYKVPQ 716.9 G !omez-Ruiz et al. (2002) 3, 4, 5
19 525.3 525.26 aS1-CN f(125–129) EGIHA 3, 4, 5
20 644.4 644.32 aS1-CN f(133–138) EPMIGV 3, 4, 5
21 910.4 910.46 aS1-CN f(163–169) AWYYVPL GAWYYVPL >1000 Yamamoto et al. (1994) 3, 4, 5
22 528.3 528.27 aS2-CN f(106–109) LNPW 3, 4, 5
23 432.2 431.21 aS2-CN f(125–127) REQ 4, 5
24 489.3 488.33 aS2-CN f(150–153) KTKL 3, 4, 5
25 449.3 450.22 k-CN f(15–17) ERF 2, 3, 4, 5
26 1022.4 1022.58 k-CN f(24–31) KYIPIQYV 3, 4, 5
27 655.4 655.37 k-CN f(106–111) MAIPPK 3, 4, 5
28 516.4 516.30 k-CN f(111–114) KKNQ 3, 4, 5
29 445.3 444.30 b-Lg f(1–4) LIVT 3, 4, 5
30 673.4 673.40 b-Lg f(1–6) LIVTQT 3, 4, 5
31 560.3 560.32 b-Lg f(2–6) IVTQT 3, 4, 5
32 655.4 601.34 b-Lg f(11–15) EIVES 3, 4, 5
33 627.4 627.40 b-Lg f(76–81) TKIPAV 3, 4, 5
34 427.3 427.28 b-Lg f(77–80) KIPA IPA 141 Abubakar et al. (1998) 3, 4, 5
35 398.3 398.25 b-Lg f(78–81) IPAV 3, 4, 5
36 374.2 374.22 b-Lg f(83–85) KID 3, 4, 5
37 645.3 645.26 b-Lg f(108–113) ENSAEP 3, 4, 5
38 346.2 346.19 b-Lg f(115–117) QSL 3, 4, 5
39 472.2 471.28 b-Lg f(123–126) VRTP 3, 4, 5
40 803.3 803.32 b-Lg f(125–131) TPEVDDE 3, 4, 5
41 677.3 677.34 b-Lg f(149–154) LSFNPT 3, 4, 5
aObserved mass (Da).bCalculated monoisotopic mass (Da).cPreviously described ACE-inhibitory peptides that share at least three C-terminal residues with those found in this study.dProtein concentration needed to inhibit 50% the original ACE activitye (1) undigested product (2) digestion with pepsin for 90min; (3) digestion with pepsin for 90min and Corolase PP for 30min; (4) digestion with
pepsin for 90min and Corolase PP for 120min;(5) digestion with pepsin for 90min and Corolase PP for 240min.fNot reported.
B. Hern !andez-Ledesma et al. / International Dairy Journal 14 (2004) 889–898 895
performed in the permeate of the digested sample. Fromthe 41 identified sequences, 15 corresponded to b-caseinfragments, 9 to as1- and as2-casein fragments, 4 to k-casein fragments, and 13 to b-Lg fragments (Table 2).Ion extraction of the ions corresponding to thesepeptides in the permeate of the sample prior tohydrolysis revealed that none of these fragments existed
in the undigested sample. Most of the peptides appearedafter 30min hydrolysis with Corolase PP and only twopeptides (peptides 1 and 25 in Table 2), which wereformed by peptic hydrolysis, survived further hydrolysiswith the pancreatic extract.Among the identified peptides, we found four peptides
that had previously been described as ACE-inhibitors.

ARTICLE IN PRESSB. Hern !andez-Ledesma et al. / International Dairy Journal 14 (2004) 889–898896
The b-Lg peptides RVY and LVR (peptides 8 and 22 inTable 1) had demonstrated strong ACE-inhibitoryactivity with IC50 values as low as 205.6 and 14 mm,respectively (Matsufuji et al., 1994; Maruyama,Miyoshi, & Tanaka, 1989). Similary, b-casein peptides,YPFPGPI and HLPLP (peptide 4 and 9, in Table 2),have been found to exhibit ACE-inhibitory activitieswith IC50 values of 500 and 23.6 mm, respectively(Kayser & Meisel, 1996; Kohmura, Nio, Kubo, Min-oshima, & Munekata, 1989). Moreover, some of thepeptides identified in this study share some of structuralfeatures with ACE-inhibitory peptides previously de-scribed (Pihlanto-Lepp.al.a et al., 2000; Yamamoto,Akino, & Takano, 1994). ACE appears to prefersubstrates or competitive inhibitors containing hydro-phobic amino acids at the three C-terminal positions.Among the most favorable C-terminal amino acids arearomatic amino acids, as well as the imino acid proline.ACE binds only weakly to peptides that have terminaldicarboxylic amino acids (Cheung, Wang, Ondetti,Sabo, & Cushman, 1980). Some peptides included inTables 1 and 2 ended with proline or aromatic residues(for example, peptides 7, 9 and 22 in Table 2). Inaddition, some peptides share some C-terminal residueswith different ACE-inhibitory peptides previously de-scribed in the literature (Tables 1 and 2). For instance,peptides 10 and 13 (Table 1) and peptide 34 (Table 2)share the three C-terminal residues with peptide IPA.This ACE-inhibitory tri-peptide (IC50 value of 141 mm)had previously been found in a hydrolysate of wheyproteins with proteinase K (Abubakar, Saito, Kitazawa,Kawai, & Itoh, 1998). Structure–activity correlationsof many ACE-inhibitory peptides indicate that theirC-terminal tripeptide residues play a predominant rolein competitive binding to the active site of ACE.Although the activity of di- or tri-peptides with ACE-inhibitory activity can not always be strictly extrapo-lated to larger peptides (Cushman, Cheung, Sabo, &Ondetti, 1977), the structural similarity of the C-terminal region may allow the prediction similaractivity. Similarly, peptides 13 and 14 in Table 2 sharethe C-terminal tri-peptide with GPV, an active peptideisolated from bovine skin-gelatin hydrolysate (Kim,Byun, Park, & Shahidi, 2001). The presence of thesesequences in the hydrolysate can explain the ACE-inhibitory activities found in the digested IF-1, and inthe ultrafiltration permeates corresponding to theundigested and digested IF-7 formula.The structure of some of the peptide sequences found
in the digested permeates may suggest the presence ofother biological activities in this hydrolysate. Forinstance, b-Lg f(78–83) (peptide 15 in Table 1) has beenfound to exert antimicrobial activity (Pellegrini, Det-tling, Thomas, & Hunziker, 2001) and peptide b-Lgf(146–148) (peptide 27 in Table 1) is very similar to b-Lgf(146–149) which possesses opioid activity (Pihlanto-
Lepp.al.a, Paakkari, Rinta-Koski, & Antila, 1997).Similarly, b-casein f(60–66) (peptide 4 in Table 2) hasimmunomodulatory and opioid activities (Kayser &Meisel, 1996; Meisel, Frister, & Schlimme, 1989;Migliore-Samour, Floc’h, & Joll"es, 1989). The peptideb-casein f(170-175) (peptide 11 in Table 2) may possessantioxidant activity, as occurs with b-casein f(170-176)(Rival, Boeriu, & Wichers, 2001). The peptides b-caseinf(193-201) and b-casein f(106-111) (peptide 13 and 27)have the same sequence, with the exception of theC-terminal residue, as b-casein f(193–202) and b-caseinf(106–112) which exhibit immunomodulatory and anti-thrombotic activity, respectively (Meisel & Schlimme,1994; Bouhallab, Moll!e, & L!eonil, 1992).
4. Conclusion
The present results demonstrated the presence of lowin vitro ACE-inhibitory activity in most of thecommercial infant milk formulas considered. However,two formulas, which corresponded to extensivelyhydrolysed milk protein formulas, showed potentACE-inhibitory activity. The ACE-inhibitory activityof a non-hydrolysed milk protein-based formula (IF-1)increased after simulated gastrointestinal digestion, butthe activity of a hydrolysed-whey formula (IF-7) did notchange after hydrolysis with gastrointestinal enzymes.The use of tandem MS and extraction of the ion ofinterest allowed us to identify the peptides formed at theend of the simulated digestion, and to follow theirformation during simulated physiological digestion.Most of peptides found at the end of the simulatedgastrointestinal digestion of the milk protein-basedformula (IF-1) were formed during incubation with thepancreatic extract. However, the peptides identified inthe ultrafiltration permeates of both the undigested, anddigested hydrolysed-whey formula (IF-7) were similar,thereby demonstrating that most of the peptides inthe hydrolysed-whey formula survive physiologicaldigestion. Some of the peptides identified in thehydrolysates correspond to previously described ACE-inhibitors, and to peptides which had been previouslyreported as antimicrobials, immunomodulators andopioid fragments.
Acknowledgements
This work has received financial support from theProjects AGL 2000-1480 and CAL01-046-C2. Theauthors acknowledge R .ohm Enzyme Gmbh for provid-ing the enzymatic preparation (Corolase PP). B.H.-L.was the recipient of a fellowship from the Ministerio deEducaci !on y Cultura, Spain.

ARTICLE IN PRESSB. Hern !andez-Ledesma et al. / International Dairy Journal 14 (2004) 889–898 897
References
Abubakar, A., Saito, T., Kitazawa, H., Kawai, Y., & Itoh, T. (1998).
New derivation of the inhibitory activity against angiotensin
converting enzyme (ACE) from sweet cheese whey. Tohoku Journal
of Agricultural Research, 47, 1–8.
Alting, A. C., Meijer, R. J. G. M., & van Beresteijn, E. C. H. (1997).
Incomplete elimination of the ABBOS epitope of bovine serum
albumin under simulated gastointestinal conditions of infants.
Diabetes Care, 20, 875–880.
Biemann, K., & Scoble, H. A. (1987). Characterization by tandem
mass spectrometry of structural modifications in proteins. Science,
237, 992–998.
Bouhallab, S., Moll!e, D., & L!eonil, J. (1992). Tryptic hydrolysis of
caseinmacropeptide in membrane reactor: Preparation of bioactive
peptides. Biotechnology Letters, 14, 805–810.
Caffarelli, C., Plebani, A., Poiesi, C., Petroccione, T., Spattini, A., &
Cavagni, G. (2002). Determination of allergenicity to three cow’s
milk hydrolysates and an amino acid-derived formula in children
with cow’s milk allergy. Clinical and Experimental Allergy, 32,
74–79.
Cheung, H.-S., Wang, F.-L., Ondetti, M. A., Sabo, E. F., & Cushman,
D. W. (1980). Binding of peptide substrates and inhibitors of
angiotensin-converting enzyme. Importance of the COOH-terminal
dipeptide sequence. Journal of Biological Chemistry, 255, 401–407.
Cushman, D. W., & Cheung, H. S. (1971). Spectrophotometric assay
and properties of the angiotensin-converting enzyme of rabbit lung.
Biochemical Pharmacology, 20, 1637–1648.
Cushman, D. W., Cheung, H. S., Sabo, E. F., & Ondetti, M. A. (1977).
Design of potent competitive inhibitors of angiotensin-converting
enzyme. Carboxyalkanoyl and mercaptoalkanoyl amino acids.
Biochemistry, 16, 5484–5491.
Doi, E., Shibata, D., & Matoba, T. (1981,). Modified colorimetric
ninhydrin methods for peptidase assay. Analytical Biochemistry,
118, 173–184.
Gobetti, M., Ferranti, P., Smacchi, E., Goffredi, F., & Addeo, F.
(2000). Production of angiotensin-I-converting-enzyme-inhibitory
peptides in fermented milks started by Lactobacillus delbrueckii
subsp bulgaricus SS1 and Lactococcus lactis subsp cremoris FT4.
Applied Environmental Microbiology, 66, 3898–3904.
G !omez-Ruiz, J. A., Ramos, M., & Recio, I. (2002). Angiotensin-
converting enzyme-inhibitory peptides in Manchego cheeses
manufactured with different starters cultures. International Dairy
Journal, 12, 697–706.
Hern!andez-Ledesma, B., Recio, I., Ramos, M., & Amigo, L. (2002).
Preparation of ovine and caprine b-lactoglobulin hydrolysates withACE-inhibitory activity Identification of active peptides from
caprine b-lactoglobulin hydrolysed with thermolysin. International
Dairy Journal, 12, 805–812.
Kayser, H., & Meisel, H. (1996). Stimulation of human peripheral
blood lymphocytes by bioactive peptides derived from bovine milk
proteins. FEBS Letters, 383, 18–20.
Kim, S.-K., Byun, H.-G., Park, P.-J., & Shahidi, F. (2001).
Angiotensin I converting enzyme inhibitory peptides purified from
bovine skin gelatin hydrolysate. Journal of Agricultural and Food
Chemistry, 49, 2992–2997.
Kim, Y. K., Yoon, S., Yu, D. Y., L .onnerdal, B., & Chung, B. H.
(1999). Novel angiotensin-I—converting enzyme inhibitory pep-
tides derived from recombinant human as1-casein expressed in
Escherichia coli. Journal of Dairy Research, 66, 431–439.
Kohmura, M., Nio, N., Kubo, K., Minoshima, Y., & Munekata, E.
(1989). Inhibition of Angiotensin-converting enzyme by synthetic
peptides of human b-casein. Agricultural and Biological Chemistry,
53, 2107–2114.
Leclerc, P.-L., Gauthier, S. F., Bachelard, H., Santure, M., & Roy, D.
(2002). Antihypertensive activity of casein-enriched milk fermented
by Lactobacillus helveticus. International Dairy Journal, 12,
995–1004.
Maeno, M., Yamamoto, N., & Takano, T. (1996). Identification of an
antihypertensive peptide from casein hydrolysate produced by a
proteinase from Lactobacillus helveticus CP790. Journal of Dairy
Science, 79, 1316–1321.
Maruyama, S., Miyoshi, S., & Tanaka, H. (1989). Angiotensing I
converting enzyme inhibitors derived from Ficus carica. Agricultur-
al and Biological Chemistry, 53, 2763–2767.
Matsufuji, H., Matsui, T., Seki, E., Osajima, K., Nakashima, M.,
& Osajima, Y. (1994). Angiotensin-I-converting enzyme
inhibitory peptides in alkaline portease hydrolyzate derived from
sardine muscle. Bioscience, Biotechnology and Biochemistry, 58,
2244–2245.
McLuckey, S. A., Van Berkel, G. J., Glish, G. L., Huang, E. C., &
Henion, J. D. (1991). Ion spray liquid chromatography/ion trap
mass spectrometry determination of biomolecules. Analytical
Chemistry, 63, 375–383.
Meisel, H. (1997). Biochemical properties of regulatory peptides
derived from milk proteins. Biopolymers, 43, 119–128.
Meisel, H., Frister, H., & Schlimme, E. (1989). Biologically active
peptides in milk proteins. Zeitschrift f .ur Ern .ahrungswiss, 28,
267–278.
Meisel, H., Goepfert, A., & G .unther, S. (1997). ACE-inhibitory
activities in milk products. Milchwissenschaft, 52, 307–311.
Meisel, H., & Schlimme, E. (1994). Inhibitors of angiotensin-
converting-enzyme derived from bovine casein (casokinins). In V.
Brantl, & H. Teschemacher (Eds.), b-Casomorphins and related
peptides: recent developments (pp. 27–33). Weinheim: VCH.
Migliore-Samour, D., Floc’h, F., & Joll"es, P. (1989). Biologically
active casein peptides implicated in immunomodulation. Journal of
Dairy Research, 56, 357–362.
Mullally, M. M., Meisel, H., & Fitzgerald, R. J. (1997). Identification
of a novel angiotensin-I-converting enzyme inhibitory peptide
corresponding to tryptic fragment of bovine b-lactoglobulin. FEBS
Letters, 402, 99–101.
Nakamura, Y., Yamamoto, N., Saki, K., Okubo, A., Yamazaki, S., &
Takano, T. (1995). Purification and characterisation of angiotensin
I-converting enzyme inhibitors from sour milk. Journal of Dairy
Science, 78, 777–783.
Papayannopoulus, A. I. (1995). The interpretation of collision-induced
dissociation tandem mass spectra of peptide. Mass Spectrometry
Reviews, 14, 49–73.
Pellegrini, A., Dettling, C., Thomas, U., & Hunziker, P. (2001).
Isolation and characterization of four bactericidal domains in
the bovine b-lactoglobulin. Biochimica et Biophysica Acta, 1526,
131–140.
Pihlanto-Lepp.al.a, A., Koskinen, P., Piilola, K., Tupasela, T.,
& Korhonen, H. (2000). Angiotensin-I-converting enzyme inhibi-
tory properties of whey protein digests: Concentration and
characterization of active peptides. Journal of Dairy Research, 67,
53–64.
Pihlanto-Lepp.al.a, A., Paakkari, I., Rinta-Koski, M., & Antila, P.
(1997). Bioactive peptide derived from in vitro proteolysis of
bovine b-lactoglobulin and its effects on smooth muscle. The
Journal of Dairy Research, 64, 149–155.
Pihlanto-Lepp.al.a, A., Rokka, T., & Korhonen, H. (1998). Angiotensin
I converting enzyme inhibitory peptides derived from bovine milk
proteins. International Dairy Journal, 8, 325–331.
Rival, S. G., Boeriu, C. G., & Wichers, H. J. (2001). Caseins and
casein hydrolysates 2. Antioxidative properties and relevance
to lipoxygenase inhibition. Journal of Agricultural and Food
Chemistry, 49, 295–302.
Roepstorff, P., & Fohlman, J. (1984). Proposal for a common
nomenclature for sequence ions in mass spectra of peptides.
Biomedical Mass Spectrometry, 11, 601.

ARTICLE IN PRESSB. Hern !andez-Ledesma et al. / International Dairy Journal 14 (2004) 889–898898
Saito, T., Nakamura, T., Kitazawa, H., Kawai, Y., & Itoh, T. (2000).
Isolation and structural analysis of antihypertensive peptides that exist
naturally in Gouda Cheese. Journal of Dairy Science, 83, 1434–1440.
Vermeirssen, V., Van Camp, J., Decroos, K., Van Wijmelbeke, L., &
Verstraete, W. (2003). The impact of fermentation and in vitro
digesti!on on the formation of angiotensin-I—converting enzyme
inhibitory activity from pea and whey protein. Journal of Dairy
Science, 86, 429–438.
Yamamoto, N., Akino, A., & Takano, T. (1994). Antihypertensive
effect of the peptides derived from casein by an extracellular
proteinase from Lactobacillus helveticus CP790. Journal of Dairy
Science, 77, 917–922.
Recommended

















