
Revisiting Previously Searched Locations in VisualNeglect Role of Right Parietal and Frontal Lesions
in Misjudging Old Locations as New
Sabira K Mannan1 Dominic J Mort1 Tim L Hodgson1Jon Driver2 Christopher Kennard1 and Masud Husain12
Abstract
amp Right-hemisphere patients with left neglect often demon-strate abnormal visual search re-examining stimuli to the rightwhile ignoring those to the left But re-fixations alone do notreveal if patients misjudge whether they have searched alocation before Here we not only tracked the eye movementsof 16 neglect patients during search but also asked them toclick a response button only when they judged they werefixating a target for the very first time lsquolsquoRe-clickingrsquorsquo on pre-viously found targets would indicate that patients erroneouslyrespond to these as new discoveries Lesions were mappedwith high-resolution MRI Neglect patients with damageinvolving the right intraparietal sulcus or right inferior frontallobe lsquolsquore-clickedrsquorsquo on previously found targets on the right at apathological rate whereas those with medial occipito-temporal
lesions did not For the intraparietal sulcus patients the prob-ability of erroneous re-clicks on an old target increased withtime since first discovering it whereas for frontal patients itwas independent of search time suggesting different under-lying mechanisms in these two types of patient Re-click deficitscorrelated with degree of leftward neglect mainly due to bothbeing severe in intraparietal cases These results demonstratethat misjudging previously searched locations for new onescan contribute to pathological search in neglect with poten-tially different mechanisms being involved in intraparietalversus inferior frontal patients When combined with a spatialbias to the right such deficits might explain why many neglectpatients often re-examine rightward locations at the expenseof items to their left amp
INTRODUCTION
Patients with neglect often remain unaware of stimuli ontheir contralesional side Such neglect is thought to in-volve pathological biases of spatial attention towards theipsilesional side of space (Bisiach amp Vallar 2000 Smaniaet al 1998 Kinsbourne 1993) However biases in spa-tial attention may not be the only underlying impair-ment Increasingly the neglect syndrome is consideredto consist of a number of component deficits (Husain ampRorden 2003 Driver amp Husain 2002 Bisiach amp Vallar2000 Mesulam 1999 Rafal 1994) each of which mayexacerbate the others with the exact combination ofdeficits varying from patient to patient Some of the com-ponent deficits might not in isolation cause neglect butmay nevertheless contribute to the syndrome when com-bined with other deficits
One aspect of neglect that has often been observedbut which until recently had received relatively littleattention is the phenomenon of lsquolsquorevisitingrsquorsquo previouslysearched locations on the ipsilesional (usually right)side of space For example on cancellation tests where
patients are asked to mark each target on a sheet usingone mark per target some patients re-mark targetsthat they have visibly cancelled before Such re-markinghas sometimes been termed lsquolsquoperseverativersquorsquo becauseit might be due to lack of control in preventing re-cancellation and consistent with this view re-markinghas been shown to be associated with frontal lesions(Rusconi Maravita Bottini amp Vallar 2002 Na et al 1999)
When eye movements are tracked during visualsearch many patients with left neglect have also beenobserved to re-fixate items to the right recursivelysearching through items towards that side while con-tinuing to neglect leftward items (Sprenger Kompf ampHeide 2002 Behrmann Watt Black amp Barton 1997Chedru Leblanc amp Lhermitte 1973) In principle suchre-fixations could arise for several reasons and it shouldbe noted that re-fixations may not be equivalent to thelsquolsquore-markingrsquorsquo phenomena mentioned above given thatno visible mark is made by merely looking at a target
Zihl and Hebel (1997) first suggested that the largenumber of re-fixations made by parietal patients (with-out neglect in their study) might be indicative of a deficitin updating or maintaining locations across saccadesBut it could be argued that neglect patients re-fixate1Imperial College London 2University College London
D 2005 Massachusetts Institute of Technology Journal of Cognitive Neuroscience 172 pp 340ndash354
rightward items because for them there is nowhereelse to look That is they might in principle be perfectlyaware that they are re-fixating old previously discoveredlocations on the right Alternatively they may mistakenlyjudge that they are discovering new targets when actu-ally re-fixating previously found right targets A criticalissue for distinguishing these possibilities is whether ornot the patients can discriminate if they are fixating newor old locations during their search
A recent study introduced a new paradigm preciselyin order to assess whether a neglect patient misjudgedpreviously searched rightward locations for novel un-searched locations (Husain Mannan et al 2001) In acomputerized search task target Ts had to be foundamong Ls (Figure 1) while eye movements were trackedCritically the patient was also asked to click on a re-sponse button only when fixating a target that was judgedto be a new discovery not when re-fixating an old targetthat had previously been found and clicked upon Notethat unlike standard pen-and-paper cancellation testsbut more analogous to search in the real world the clickresponse did not leave any visible trace so in this taskobservers have to remember whether the currentlyfixated target locations had previously been inspected
Husain Mannan et al (2001) studied just a singleright-hemisphere patient with left neglect in this para-digm and found that he lsquolsquore-clickedrsquorsquo at a pathologicallyhigh rate on rightward targets when re-fixating thosethat had previously been found and clicked upon It wassuggested that this tendency to make misjudgmentsin responses to previously searched locations mightcontribute to neglect When combined with any right-ward spatial bias in attention it might exacerbate neglectfor leftward items as the patient would not realize hewas repeatedly covering the same ground on the right(Husain Mannan et al 2001)
However this proposal was based on only a single-case to date To investigate it further the present study
applied the new paradigm to 16 neglect patients withmiddle cerebral artery (MCA) or posterior cerebral artery(PCA) stroke In addition to assessing how prevalentthe lsquolsquore-click deficitrsquorsquo might be this study has two impor-tant features First we performed time-series analysesof search performance to assess whether any misjudg-ments in responses to previously found targets de-pended on the time since a given target had previouslybeen fixated and clicked upon1 Second we mapped thelesions of our patients using high-resolution MRI (Mortet al 2003) and reconstructed areas of lesion overlapassociated with re-clicking Taken together these ap-proaches allowed us to distinguish between two dif-ferent types of pathological re-clicking Patients withdamage to the right intraparietal sulcus showed re-clickbehavior that depended on the time since a previouslyfound target had initially been fixated whereas rightinferior frontal patients re-clicked at a pathological rateregardless of the time intervening between initial dis-covery and re-fixation These results show that manybut not all neglect patients misjudge previously foundtargets as if they were new discoveries with the defi-cit and its nature depending on which brain areas aredamaged
RESULTS
Neglect in the Search Task and Re-fixations
As expected neglect patients missed significantly moretargets on the left half of the screen (mean 38) thanthe right [18 t(15) = 27 p lt 05 for an example seeFigure 1] Healthy age-matched volunteers showedequal detection of targets on the left and right sidesmissing 16 on the left and 14 on the right [t(14) =099 ns] Re-fixation rate for each participant is definedas total number of re-fixationstotal number of targetsfixated expressed as a percentage Both patient and
Figure 1 An example of a
search array on which aresuperimposed fixations
(dots) and the interpolated
scanpath of one neglect
patient (Case 15) Noteneglect of stimuli on the left
and many re-fixations on
stimuli to the right
Mannan et al 341
control subjects were more likely to re-fixate targets thandistractors (28 times and 27 times for patients and con-trols respectively)
As a group neglect patients had a significantly higherre-fixation rates (mean = 142) than healthy controls[40 t(29) = 35 p lt 01 see Figure 2A] Ten ofthe 16 patients had re-fixation rates that fell abovethe 95 percentile for controls The critical issue forthis study is whether re-fixations are associated withmisjudging previously searched locations for novelones This is indexed by the lsquolsquore-click ratersquorsquo whichindicates any tendency for subjects to erroneously re-spond to previously found targets as if they were anew discovery
Re-click Rates
The re-click rate for each participant is given by totalnumber of re-clickstotal number of targets clicked uponexpressed as a percentage The mean re-click rate for
neglect patients (30) was significantly higher than forcontrols [6 t(29) = 31 p lt 01] who rarely consid-ered a re-fixated target to be a new discovery (Figure 2B)For the neglect group the probability of re-clicking washighest for targets to the right (Figure 3) In total 10 ofthe 16 patientsmdashhereafter termed high re-click groupmdashhad re-click rates beyond the upper 95 confidenceinterval for controls (Figure 2B) The remaining 6 pa-tients (low re-click group) had re-click rates within thenormal range demonstrating the re-clicking deficit isnot present in all neglect patients Moreover this resultalso shows that brain damage per se does not lead to apathological re-click rate Later (in the section on Ana-tomical Correlates) we show that the deficit cannot alsobe attributed to lesion size but rather is associated withdamage to specific brain regions
Importantly although 7 patients had both a high re-click and a high re-fixation rate 3 patients had an ab-normally high re-fixation rate and a normal re-click rateThus abnormal re-fixation rate alone does not neces-sarily identify patients who fail to distinguish betweennovel targets and previously inspected ones Hence theimportance of measuring re-click rates Neverthelessacross the group of patients there was a significant cor-relation between re-click and re-fixation rates (Figure 4)[r(15) = 78 p lt 0001] There was no such correla-tion between re-fixation and re-click rates for healthycontrols [r(14) = 05 ns]
Interestingly high re-clicking neglect patients notonly re-clicked more often on targets they had alreadyclicked upon but they also more frequently failedto click upon a fixated target they had not previouslyclicked upon The high re-click neglect group omittedto click on significantly more newly fixated targets
Figure 3 The distribution over space of re-clicks made by neglect
patients The total number of re-clicks made within each virtual column
of the screen is expressed as a percentage of the total number ofre-clicks made by patients
Figure 2 (A) Re-fixation rates for neglect patients and healthyindividuals with individual mean re-fixation rates for each subject
shown as circles Ten of 16 patients had re-fixation rates which fall
above the 95th percentile for healthy volunteers denoted by the
dashed line (B) Re-click rates for neglect patients and healthyvolunteers Ten patients had re-click rates above the 95th percentile
for healthy volunteers
342 Journal of Cognitive Neuroscience Volume 17 Number 2
than the low re-click neglect group [mean = 30omissions SE 4 vs 19 and 3 t(14) = 231 p lt05]2 Such an association between these lsquolsquoclick omis-sionsrsquorsquo and high re-click rate would not be expected ifre-clicking was entirely due to a lsquolsquoperseverativersquorsquo failureto inhibit responses to rightward targets because lsquolsquoclickomissionsrsquorsquo represent failures to respond Nor wouldthey be explained by items on the right being per-ceived to be more salient and thus automatically cap-turing responses
Anatomical Correlates of High Re-click Rates
Although 10 neglect patients demonstrated significant-ly elevated re-click rates 6 did not To determinewhether key anatomical locations underlie the distinc-tion between these groups we sought to isolate brainregions that were damaged in neglect patients withhigh re-click rates and equally importantly spared inpatients without the deficit Critically there was nocorrelation between lesion volume and re-click rate[r(13) = 31 ns] or between lesion age and re-clickrate [r(13) = 30 ns] Each patient was assigned toone of two subgroups according to whether they hadfrontal involvement or not Neglect patients withlsquolsquoposterior onlyrsquorsquo lesions had damage posterior to thecentral sulcus with no damage anterior to the pre-central sulcus (n = 9) Neglect patients with lsquolsquoposte-rior and anteriorrsquorsquo involvement had cortical damageanterior to the central sulcus (n = 5) all patients inthis subgroup also had some posterior damage
Parietal Correlate of High Re-click Rate
Among the 9 patients in the lsquolsquoposterior onlyrsquorsquo subgroup4 had an abnormally high re-click rate 3 with MCA ter-
ritory damage (Cases 5 6 and 8) and only 1 with PCAdamage (Figure 5A) This single PCA patient (Case 2)had damage to white matter deep to the occipito-temporo-parietal junction in contrast to the other 3PCA patients whose lesions did not extend so faranteriorly or dorsally The area of damage commonto all 3 MCA patients with abnormally high re-clickratesmdashwhile also being intact in the 5 posterior sub-group patients with normal re-click ratesmdashwas locatednear the anterior end of the intraparietal sulcus closeto the lateral bank (Figure 5C Talairach 46 40 28for centroid volume 058 cm3) This area was not sim-ply the most common region of overlap for all 8 MCApatients that was located more dorsally (Talairach37 55 38)
In order to assign statistical significance to the areaof overlap we performed a voxel-by-voxel analysis (Bateset al 2003) For every voxel that was damaged in atleast 3 subjects we compared the re-click rate of pa-tients with damage to that voxel with those of patientswithout damage to that particular voxel producing at statistic for a given voxel The area shown in yellow(Figure 5C) yields a value of t(7) = 491 indicatingsignificantly higher re-click rates associated with dam-age to this region ( p lt 002) This difference survivesBonferroni correction for multiple comparisons ( p lt05) where the number of comparisons was 6 (therewere only 6 regions in which the lesions of 3 or morepatients overlapped)
An alternative analytical approach is to focus only onpatients with relatively small lesions Seven patients(Cases 4ndash9 and 15) had focal parietal lesions lt31 cm3all had posterior lesions except Case 15 who also hadinvolvement anterior to the central sulcus The maxi-mum area of overlap common to all 4 patients with highre-click rates (Cases 5 6 8 15) which also cruciallyremained undamaged in the 3 patients with normal re-click rates (Cases 4 7 9) again centered on the lateralintraparietal sulcus (Figure 6) slightly more dorsally(Talairach 37 39 37 volume 011 cm3) and involvinga subset of the critical voxels identified in Figure 5 Notethat this overlap region will probably also involve whitematter fibers including those that may connect the pa-rietal cortex to the frontal lobe
Frontal Correlate of High Re-click Rate
Four of the five patients in the lsquolsquoposterior plus anteriorrsquorsquoneglect subgroup had an abnormally high re-click rateThe area of damage common to all four was in theinferior frontal lobe (Talairach 38 18 16 for centroid178 cm3) in a region corresponding to Brodmannrsquosarea 44 part of the premotor cortex (Figure 7A) Threeof the four posterior plus anterior neglect patients witha pathologically high re-click rate also had damage ex-tending into or bordering the ventrolateral prefrontalcortex The remaining patient who had the highest re-
Figure 4 Mean re-click rates plotted against mean re-fixation rates
for individual neglect patients There was a significant correlation[r(15) = 78 p lt 0001]
Mannan et al 343
click rate of all patients (Case 15) had damage involvingthe dorsolateral prefrontal cortex Only 1 of the 5 pos-terior plus anterior patients had a normal re-click rate(Figure 7B) Her lesion was posterior to the region ofoverlap seen in the four frontal patients with abnor-mally high re-click rates It involved the posterior premo-tor cortex and motor cortex and extended caudally intothe inferior parietal lobe Thus the subtraction of herlesion from the remaining posterior plus anterior patientswith high re-click rates (Figure 7C) does not alter theirmaximum overlap region (displayed in Figure 7A)
Anatomical Correlates of the lsquolsquoRe-fixationrsquorsquo Deficit
Ten neglect patients had a high re-fixation rate but 6patients did not There was no correlation betweenlesion volume and re-fixation rate [r(13) = 22 ns]or between age of lesion and re-click rate [r(13) =36 ns] Although some patients in either the poste-rior (Cases 1 2 4ndash8) or frontal group (Cases 11 13 15)had high re-fixation rates other patients with highlysimilar lesion locations had normal re-fixation ratesAs a consequence voxel-by-voxel analysis (Bates et al
Figure 5 The
neuroanatomical correlates of
the deficit in retaining spatiallocations for neglect patients
with posterior only (nonfrontal)
lesions (n = 9) (A) Individuallesions of patients with high
re-click rate (patients with MCA
damage Cases 8 5 6 patient
with PCA damage Case 2) andoverlap map for these
4 patients where maximum
overlap occurs for a small region
(shown in green) around theright anterior intraparietal
sulcus damaged in the 3 MCA
patients (B) Individual lesionsof patients with normal re-click
rate (patients with MCA
damage Cases 9 and 7 patients
with PCA damage Cases 1 3 4)and overlap map for these
5 patients with maximum
overlap occurring in the
occipital cortex Importantlynone of the 5 patients with
normal re-click rates have
damage to the common areaof overlap for patients with high
re-click rates shown in A
(C) Contrast map highlighting
intraparietal locus which isdamaged in 3 out of 4 patients
with a high re-click rate and
intact in the 5 patients with
normal re-click rates (shown inyellow) Regions that are
damaged in patients with high
re-click rates and spared in
those patients with normalre-click rates are shown in red to
yellow whereas regions in blue
denote areas that are commonto patients with
normal re-click rates but
spared in patients with high
re-click rates
344 Journal of Cognitive Neuroscience Volume 17 Number 2
2003) revealed no anatomical specificity associated withhigh re-fixation rates [t(7) = 085 ns] unlike thatassociated with high re-click rates even when onlyfocal lesions were considered [t(5) = 165 ns]
Time Interval between Clicking and Re-clicking ona Target
We next examined the probability of lsquolsquore-clickingrsquorsquo on atarget when re-fixating it as a function of the time lag ornumber of intervening saccades between the first clickon that target and the subsequent revisit An increasedprobability of re-clicking with increasing delay (or inter-vening number of saccades) would be consistent a failureto keep track of spatial locations during search whereasa flat or decreasing function might arise from impairedresponse inhibition (lsquolsquoperseverativersquorsquo behavior) or even
a failure to encode target location This time-series anal-ysis for the 10 patients with high re-click rates revealedtwo distinct patterns of behavior Half the patients de-monstrated a significantly increasing probability of mis-judging an already found item to be a novel one withincreasing delay (Figure 8A) or intervening saccades(Figure 8C) whereas the other 5 patients had a constantor even decreasing probability (Figure 8B and D)
Importantly most (45) patients in the group withincreasing re-click functions had damage to the focalparietal location previously identified in the analysisof patients with focal lesions and high re-click rates(Figure 6C) The other patient was Case 2 who had alarge PCA lesion extending to the white matter deep tothe occipito-temporo-parietal junction By contrast noneof the patients with flatdecreasing functions had dam-age to the key parietal region Instead the latter group
Figure 6 The
neuroanatomical correlates of
the deficit in retaining spatial
locations for neglect patientswith focal lesions (n = 7)
(A) Individual lesions of
patients with high re-click rate
(all 4 patients have MCAdamage) and overlap map for
these 4 patients where
maximum overlap occurs for asmall region around the right
intraparietal sulcus (shown in
red) damaged all 4 MCA
patients (B) Individual lesionsof patients with normal re-click
rate (all patients with PCA
damage) and overlap map for
these 3 patients with maximumoverlap occurring in the
occipital cortex Importantly
none of these 3 patients withnormal re-click rates have
damage to the common area of
overlap for patients with high
re-click rates shown in A(C) Contrast map highlighting
intraparietal locus which is
damaged in 4 out of 4 patients
with a high re-click rate andintact in the 3 patients with
normal re-click rates (shown in
yellow) Regions that are
damaged in patients with highre-click rates and spared in
those patients with normal
re-click rates are shown in redto yellow whereas regions in
blue denote areas that are
common to patients with
normal re-click rates butspared in patients with high
re-click rates
Mannan et al 345
had frontal cortical or subcortical involvement Thusincreasing re-clicking with time interval was associatedwith parietal damage whereas flat re-click functionswere associated with frontalsubcortical lesions
For neglect patients as a whole the intervals betweenan initial click on a target and a subsequent re-click werelong (mean = 40 saccades) but much shorter betweenan initial fixation and a later re-fixation (mean = 5) withthis difference being significant [t(15) = 315 p lt01] This again emphasizes the dissociation betweenre-fixations and re-clicks and also demonstrates that re-clicking does not occur on most re-fixations
Relationship of Re-click Rate to Neglect Severity
We assessed three separate measures of neglect Thefirst was the difference between miss rates for left ver-sus right targets within the experimental search taskitself the other two measures of neglect were based onthe clinical pen-and-paper measures of Mesulam shape-cancellation (Mesulam 1985) and line bisection fromthe BIT (Wilson Cockburn amp Halligan 1987)
On the experimental search task there was a moderatebut significant correlation between re-click rate and ne-glect in search across the group [r(15) = 47 p lt 05
Figure 9A] Thus the greater the re-clicking on right-ward items the greater the severity of leftward neglectThere was also a moderate but again significant corre-lation between re-click rates and the severity of individualpatientsrsquo clinical neglect as indexed by the cancellationmeasure [r(15) = 46 p lt 05 Figure 9B] Thesecorrelations were driven largely by three of the patientsidentified in our focal lesion analysis (Figure 6A) withinvolvement of the key parietal region and high re-clicking behavior Their data are shown in filled circlesto the right in the graphs displayed in Figure 93 Thecorrelation between deviation on line bisection andre-click rate was not significant [r(15) = 13 ns] How-ever one would not predict that a deficit in retaininglocations already visited (or response inhibition) wouldnecessarily impair performance on a task such as linebisection
Effect of Varying Number of Targets
In 7 patients (Cases 5 8 10 11 14ndash16) we also exam-ined the effect of varying the number of targets in thesearch task Patients viewed lsquolsquotarget onlyrsquorsquo displays con-taining either 2 6 or 19 targets randomly distributedon the screen without any L-shaped distractors In-
Figure 7 Theneuroanatomical correlates of
the re-click deficit in neglect
patients with posterior plusanterior lesions (n = 5)
(A) Individual lesions of
patients with high re-click rate
(all 4 patients have MCAdamage) and overlap map for
these 4 patients where
maximum overlap occurs for
a region in the right inferiorfrontal lobe (shown in red)
damaged all 4 MCA patients
(B) Individual lesions of a single
patient with normal re-clickrate (C) Contrast map
highlighting the inferior frontal
locus (shown in yellow) whichis damaged in 4 out of
4 patients with a high re-click
rate and intact in the patient
with normal re-click rateRegions that are damaged in
patients with high re-click rates
and spared in the patient
with normal re-click rate areshown in red to yellow
whereas regions in blue denote
areas that are damaged in thepatient with normal re-click
rates but spared in patients with
high re-click rates
346 Journal of Cognitive Neuroscience Volume 17 Number 2
creasing the number of targets led to significantly higherre-click rates ( Jonckheere statistic = 133 p lt 01Figure 10) as well as more severe neglect (Jonckheerestatistic = 129 p lt 01) consistent with the proposalthat the mechanisms underlying pathological re-clickingmight contribute to neglect behavior Note that a lsquolsquoper-severativersquorsquo failure of response inhibition would not onits own easily explain this finding For the same absolutenumber of click responses one would predict a higherre-click rate with smaller set-sizes than with larger ones(see also Husain Mannan et al 2001 for a discussion)Although the number of subjects were small there wasno difference in the performance pattern between the3 patients who had increasing re-click rate functions inthe time-series analysis and the 4 patients with flat ordecreasing re-click rate functions
Varying Perceptual Load during Search
Can re-click behavior be explained simply by impairedvisual perception of targets from discriminators Neglectpatients did find it more difficult than control subjectsto select targets from distractors fixating on average19 distractors for every target found [cf 14 for controlsubjects t(29) = 485 p lt 0001] Patients also some-times clicked on distractors (64) but no more often
than controls (61) and it is possible that this rate iseffectively inflated by our strict criterion of considering aclick to be a response on the item that is being fixated atthe time of the click (It is likely that sometimes ob-servers click in response to targets they fixated one ormore saccades previously) Re-clicks on distractors wereextremely rare (06 cf 30 for targets)
To examine the effect of reduced perceptual load in8 patients (Cases 5 7 8ndash10 13 1 4 16) 6 of whom werehigh re-clickers we used search displays with salientcross (+) targets rather than Ts L distractors wereunchanged Patients fixated significantly fewer distrac-tors when searching for cross (+) targets [distractortarget ratio of 134 cf 181 for T targets t(7) = 60 p lt001] suggesting they did indeed find targetndashdistractordiscrimination easier Importantly however there wasno significant difference in re-click rate [26 for +targets cf 27 for T targets t(14) = 041 ns] or evenin re-fixation rate [100 for + targets cf 119 fort(14) = 041 ns] Thus these changes in perceptualload did not influence re-click performance
DISCUSSION
On this multitarget visual search task neglect patientsmissed targets to their left and also re-fixated targets
Figure 8 The probability of
re-clicking or re-fixating on
target as a function of delay
between the original click andsubsequent re-fixation (for
patients with high re-click
rates) (A) Probability functions
for patients with significantpositive slopes Four patients of
these patients have damage to
the focal intraparietal locusidentified in Figure 6C whereas
the single patient with a PCA
lesion and no damage to the
critical parietal locationindicated by symbol has only
a very shallow positive slope
(B) Probability functions for
patients with no significantpositive slope None of these
5 patients have damage to the
critical intraparietal region (C)As for A but for delay measured
in terms of intervening
fixations (D) As for B but for
delay measured in terms ofintervening fixations
Mannan et al 347
to the right much more often than healthy individuals(Figure 2A) Critically 10 of the 16 neglect patients re-clicked on rightward targets at a significantly higher ratethan healthy control subjects (Figure 2B) For 5 of thesepatients the probability of re-clicking increased withtime (Figure 8A) whereas the other 5 neglect patientswith high re-click rates demonstrated a constant or de-creasing probability of re-clicking over time (Figure 8C)The remaining 6 neglect patients had re-click rateswithin the normal range demonstrating that neitherneglect nor brain damage per se leads to a pathologicalre-click rate
High-resolution MRI mapping of lesions revealed im-portant anatomical dissociations within the neglect pop-ulation Patients with pathological re-click rates hadlesions affecting either the right intraparietal sulcus orthe inferior frontal lobe the only exception (Case 2) hada large occipito-temporal lesion that extended deep intothe parietal white matter Neglect patients with normalre-click rates did not have damage to either of thecritical locations in the intraparietal sulcus or inferiorfrontal lobe (Figures 5 6 and 7) Instead they had moredorsal lesions (for MCA territory patients) or occipitaland medial-temporal damage (for PCA territory lesions)which can also result in neglect (Mort et al 2003Cals Devuyst Afsar Karapanayiotides amp Bogousslavsky2002) Importantly lesion location rather than the sizeor age of lesion was critical to the presence of the re-clicking impairment Across the group of neglect pa-tients there was a moderate but significant correlationbetween re-click rate and degree of neglect (Figure 9)driven largely by patients with parietal damage who hadboth a high re-click rate and also severe neglect consis-tent with the proposal that mechanisms underlying thisbehavior in parietal patients might have a functionallyimportant role in determining the severity of neglect
Role of Right Intraparietal Sulcus
Notably for all patients with damage to the critical locusin the right intraparietal sulcus (which may also involveparieto-frontal white matter fibers) the probability of re-clicking on a target increased with time (or interveningsaccades) since initial discovery (Figure 8) Because theyrarely re-clicked at short intervalsmdasheven though themajority of re-fixations occurred during this periodmdashthis pattern of behavior suggests that for these patientstarget locations were actually veridically perceived and
Figure 9 (A) Neglect on experimental search task (19 targets and44 distractors) plotted against mean re-click rate for individual neglect
patients Percentage neglect was calculated as the difference between
the number of targets missed on the left versus right expressed as apercentage of the total number of targets missed There was a
significant correlation between the degree of neglect and the re-click
rate [r(15) = 465 p lt 05] Patients with high re-click rates are
highlighted by black circles (for patients with damage to the focalparietal location identified in Figure 6C) and light gray circles (for
patients with frontal damage) while the single patient with PCA
damage and a high re-click rate is indicated by dark gray (B) The
number of targets missed on the Mesulam shape-cancellation taskplotted against mean re-click rate for individual neglect patients Again
there was a significant correlation between the number of targets
missed and the mean re-click rate [r(15) = 456 p lt 05] Patients withhigh re-click rates are denoted as in A
Figure 10 Mean re-click rate (filled circles) and degree of neglect
(unfilled squares) increased systematically with increasing numbers of
targets in target-only displays
348 Journal of Cognitive Neuroscience Volume 17 Number 2
well encoded into working memory Furthermore de-tailed single-case studies of two parietal patients withhigh re-click rates (Malhotra Mannan Driver amp Husain2004 Husain Mannan et al 2001) demonstrate usingdifferent psychophysical methods that these patientscan accurately encode spatial locations4 Increasing re-clicks with time (or intervening saccades) would how-ever be consistent with a deficit in keeping track oftarget locations during extended search
Several lines of evidence support the proposal thatthe critical intraparietal sulcus region we have identifiedmay have an important role in the spatial transforma-tions required to keep track of spatial locations acrosssaccades In rhesus monkeys neurons within the lateralbank of the intraparietal sulcus (area LIP) encode andlsquolsquomaintainrsquorsquo the remembered locations of saccadic tar-gets over time (Chafee amp Goldman-Rakic 1998 MazzoniBracewell Barash amp Andersen 1996) Some visuallyresponsive neurons in the LIP also appear to shift theirretinotopic representation at the onset of an intendedsaccade as if lsquolsquore-mappingrsquorsquo representations of objects inspace to update them across shifts of gaze (DuhamelColby amp Goldberg 1992)
In humans this computation has been investigatedusing the double-step saccade paradigm where twoflashes of light appear in quick succession such thatthe second target disappears before a saccade can bemade to the first target Patients with lesions involvingthe right parietal cortex are impaired at performing thistask suggesting that they are unable to update spatialrepresentations accurately after even one saccade (Heideamp Kompf 1998 Duhamel et al 1992) Similarly trans-cranial magnetic stimulation of the posterior parietalcortex prior to the second saccade reduces its accuracy(Donkelaar amp Muri 2002) and a recent study of patientswith parietal lesions also implicates this area in thespatial updating of inhibition of return after a saccade(Sapir Hayes Henik Danzinger amp Rafal 2004) Func-tional imaging studies of double-step (Tobler et al2001) and triple-step memory-guided saccades (HeideBinkofski et al 2001) have identified a region withinthe intraparietal sulcus (Talairach coordinates 40 4044 for Tobler et al and 44 48 36 Heide et al) This isclose to the critical region of lesion overlap we haveidentified in the posterior neglect patients with high re-click rates (Talairach 37 39 37)
We suggest that in our patients with lesions involvingthe right intraparietal sulcus uncertainty about locationsof previously found targets increases with each saccadeThis uncertainty is further exacerbated by interferencefrom newer target locations leading to increased prob-ability of misjudging old target locations for new assearch progresses Such progressively worsening uncer-tainty might be due to an accumulating impairment inlsquolsquore-mapping target locationsrsquorsquo across saccades or im-paired lsquolsquomemoryrsquorsquo of previously inspected target loca-tions across saccades As noted above there is evidence
for both types of signalmdash re-mapping and maintainingsaccadic spatial memorymdashin the parietal cortex and it ispossible that disruption to both these functions contrib-utes to the re-click deficit we have identified Certainlythe two potential types of deficit are not mutuallyincompatible
A slightly different account of the re-clicking behaviormight be framed in terms of the intriguing hypothesisproposed by Manly Woldt Watson and Warburton(2002) to explain visible re-marking on a standard can-cellation task They argue that such behavior is due topatients erroneously transposing leftward stimuli to theright resulting in apparent re-cancellation These au-thors found that removing targets from the left re-duced both neglect and re-marking However they didnot monitor eye movements so it is possible that sometargets that are logged as neglected (because they werenot cancelled) were nevertheless fixated and thereforeform part of the load on memory during search Fur-thermore in our experimental study subjects with ahigh re-click rate also failed to click on a higher pro-portion of fixated targets Manly et alrsquos proposal wouldnot easily account for such lsquolsquoclick omissionsrsquorsquo becausethese refer to fixated targets that were not actuallyresponded to
Finally it might be argued that increasing re-clickingwith search duration reflects a deficit in spatial workingmemory (SWM) that is independent of saccadic eyemovements (ie a decay in memory that is not trans-saccadic) Because time and number of interveningsaccades were not independently manipulated in thisunrestricted visual search paradigm we are not ablecategorically to distinguish between these possibilitiesWe note however that functional imaging studies ofSWM tasks that do not require eye movements oftendemonstrate activity related to the maintenance ofspatial information which is generally more posteriorand dorsal than the parietal locations isolated in thisstudy (DrsquoEsposito et al 1998) Moreover recent behav-ioral data indicate that SWM is more vulnerable tosaccadic eye movements than shifts in attention or limbmovements (Pearson amp Sahraie 2003)
Frontal Lesion Site Associated withHigh Re-click Rates
Four out of 5 neglect patients with lesions involvingposterior plus anterior damage also showed a high re-click rate and the area of damage common to thesepatients centered on the right inferior frontal lobe cor-responding to Brodmannrsquos area 44 (Figure 7C) Thisregion has previously been associated with frontalneglect (Husain amp Kennard 1996) and is consideredto be part of the ventral premotor cortex and thehomologue of Brocarsquos areas in the left hemisphere(Rizzolatti amp Arbib 1998 Jackson amp Husain 1996) Inthese frontal patients the probability of re-clicking did
Mannan et al 349
not increase with time interval between the first clickand re-fixation but rather it stayed constant or declinedwith increasing delays
Such behavior might be consistent with a failure toinhibit responses to rightward locationsmdasha type oflsquolsquoperseverativersquorsquo behavior Note though that this is notnecessarily the same type of perseverative responseas seen when some patients with neglect re-markpreviously found items on standard cancellation tests(Rusconi et al 2002 Na et al 1999) In those casestargets that are visibly cancelled are often marked againwhereas in our computerized search tasks no marks arevisibly left on targets once they have been clicked uponMoreover only one of our neglect patients (Case 1) de-monstrated re-cancellation of visibly cancelled targetson pen-and-paper tasks and then only on one occasionImportantly this patient actually had a normal re-clickrate (67) on our task and his stroke was in the PCAterritory sparing both the intraparietal sulcus and fron-tal lobe
In support of a role for the right frontal lobe in in-hibiting responses to previously found targets recentimaging studies of response inhibition have highlightedregions of the right inferior frontal gyrus (IFG) very closeto the centroid of damage for our frontal patients (38 1618 cf 43 22 21 Konishi Kyoichi Uchida Sekihara ampMiyashita 1998 and 41 16 19 Konishi Kyoichi UchidaKikyo et al 1999) Alternative explanations for the highprobability of re-clicking directly after clicking might bethat these patients have impaired perceptual discrimina-tion of targets impaired perceptual localization of targetsor that they encounter difficulty in encoding spatiallocations into working memory Our data are not sugges-tive of a perceptual discrimination problem as changingthe perceptual load did not inf luence re-click ratesMoreover the patients still had high re-click rates in thetarget only conditions (Figure 10) where no perceptualdiscrimination of targets and distractors was necessaryPoor perceptual localization would seem unlikely giventhat these patients have frontal lesions Furthermoreusing a vertical SWM task similar to the Corsi blocktapping task (Malhotra et al 2004) we have found thatpatients with neglect can localize and perceptually en-code single target locations as well as controls and aresignificantly impaired at remembering two or more loca-tions shown in sequence consistent with the presentobservations Some recent functional imaging data sug-gest that activation within the prefrontal cortex corre-lates well with successful encoding of material intoworking memory (Rypma amp DrsquoEsposito 2003) Furtherresearch is required to distinguish between these pos-sible explanations for the mechanisms underlying there-click deficit in frontal patients
The only patient with frontal damage who did re-clickmore frequently with increasing time had damage cen-tered on the dorsolateral prefrontal cortex (althoughalso encroaching on the IFG) Neurons in the dorsolat-
eral prefrontal cortex like those in the LIP encode theremembered locations of saccadic targets (Goldman-Rakic 1996) This patient (Case 15) who had the high-est re-click rate of all also had damage to the criticalparietal location described above (in contrast to all otherfrontal patients) Thus she may have suffered from acombination of underlying deficits
Re-fixations and Misjudging PreviouslyInspected Locations during Search
In this search task re-fixation and re-click rates weresignificantly correlated for neglect patients Howeverthere were important dissociations within individuals 2patients with PCA lesions had re-fixation rates of 220and 80 respectively but both had a re-click rate ofonly 7 These are clearly examples of individuals whodo not show pathological re-clicking despite having highre-fixation rates High re-fixation rates may result froma lack of integration of spatial information due to V1damage (Zihl amp von Cramon 1985) in our PCA patientsor possibly because of reduced inhibition of return(Bartolomeo Sieroff Decaix amp Chokron 2001) What-ever the exact cause for dissociations between re-fixa-tions and re-click rates our data suggest re-fixation doesnot necessarily index the same underlying deficits asrevealed by our re-click measure High re-fixation ratesunlike high re-click rates were also not significantlyassociated with damage to any specific cortical locationagain demonstrating a dissociation between re-fixationsand pathological re-clicking
The relationship between re-click rates and neglect isunlikely to be a simple one In our study there was amoderate but significant correlation between the twomeasures suggesting that this association is unlikely tohave arisen by chance However non-neglect patientscan also exhibit a high re-click rate (unpublished obser-vations from our laboratory) Taken together our resultssuggest that difficulties in keeping track of previouslyinspected targets (because of impaired trans-saccadic re-mapping andor memory) may be one contributingcomponent to visual neglect in patients with lesions in-volving the right intraparietal sulcus As mentioned thesearch task we used differed in one striking aspect fromstandard cancellation tests subjects do not visibly markthe targets they visit Whereas in cancellation tests pa-tients mark each target they find which might appear todispense with any need to remember which locationshave been searched before However it should be ap-preciated that in most standard cancellation tests theindividual marks made by patients may not be clearlyvisible in peripheral vision particularly if neglect pa-tients have a local spatial bias (Rafal 1994 RobertsonLamb amp Knight 1988) or a restricted field of view(Russell Malhotra amp Husain 2004) Thus when com-bined with a rightward attentional bias keeping track ofsearched locations may be critical to prevent gaze from
350 Journal of Cognitive Neuroscience Volume 17 Number 2
returning to rightward locations even if they have beenmarked before as the marks may not be visible untilgaze returns to them Consistent with this hypothesisneglect is greater when cancellation is performed usinginvisible marks compared to large and salient red marksthat can be seen peripherally (Wojciulik Husain Clarkeamp Driver 2001)
A task like the present search-and-click paradigm maymore closely resemble search in the real world than dotraditional cancellation tests When we examine a visualscene we do not leave marks on each object we ex-amine Instead just as in the search task used in thisstudy we have to keep track of the locations we visit ifwe are to avoid inefficient re-examination of thoselocations Neglect patients with damage to the rightintraparietal sulcus appear to be impaired at keepingtrack of searched locations in this way When combinedwith their rightward attentional bias we propose thatsuch an impairment leads to recursive search of stimulito the right in experimental search tasks as used hereand also in daily life where neglect is so apparentdespite unlimited viewing time
The paradigm we have used has the important virtuesof allowing patients to explore freelymdashas in real-worldsearch or clinical cancellation tasksmdashand relating theirre-click behavior directly to the severity of neglect in thesame task But it might be argued that it would be usefulalso to probe SWM in a more controlled manner by re-stricting observersrsquo eye movements to a specific set oflocations and probing memory of previously visitedplaces A recent study using a gaze-contingent para-digm has elegantly probed the capacity of spatial mem-ory during search in healthy observers and foundevidence for memory for up to four previous locations(McCarley Wang Kramer Irwin amp Peterson 2003)Variants of this paradigm may in the future be usefulto probe the capacity of SWM across saccades in patientswith neglect and also to probe further whether thereare differences between neglect patients with parietaland frontal lesions
METHODS
Subjects Imaging and Lesion Mapping
Sixteen patients with left neglect following right hemi-sphere infarction or hemorrhage were recruited Con-sent was obtained according to the Declaration ofHelsinki as approved by the hospital ethical commit-tees Presence of neglect was determined by clinicalobservation plus bedside tests including the Mesulamshape-cancellation task (Mesulam 1985) and line bisec-tion task (Wilson et al 1987) (see Table 1) Twelvepatients had MCA territory strokes the rest involvedonly the PCA territory which can also result in neglect(Cals et al 2002) Only one neglect patient (Case 1 whodid not have frontal involvement) demonstrated re-
cancellation of visibly cancelled targets on pen-and-paper tasks and then only on one occasion Fifteenage-matched healthy volunteers were controls (meanage = 63 SD = 14 vs 63 SD = 13 for patients)
All patients had clinical CT scans In addition dedi-cated high-resolution structural MRI scans were alsoobtained for 1416 neglect patients MRI scans wereunobtainable for the other two (Cases 11 and 16)because of claustrophobia or loss to follow up In theMCA neglect group median time between stroke andscan was 63 days with the shortest interval being35 days for the PCA neglect group one patient had tobe scanned relatively early at 9 days but the mediantime was 140 days MRI scans comprised one 256-sliceT1-weighted acquisition (MPRAGE TR 97 msec TE4 msec TA 12 min) performed in the coronal planealigned perpendicular to the anteriorndashposterior commis-sural line on a Siemens Vision 15-T scanner Within-and between-slice resolution was 1 mm For each pa-tient the area of damage was determined by detailedvisual inspection of the digital image for every singleslice Lesion delineation was performed by neurologists(authors D M and M H) by tracing by hand theboundary of the lesion onto every single 1-mm-thickaxial slice using a graphics tablet (WACOM Intuos A6)Combining all the slices then led to a 3-D region of in-terest (ROI) for each patient For one patient (Case 15)there were two areas of discrete focal damage one inthe frontal cortex and the other in the parietal cortex
Normalization of each MRI including the 3-D ROI tothe T1-template was performed with SPM99 (wwwfilionuclacukspm) The ROI was used as a mask for thelesion during determination of alignment parameters tominimize the contribution of abnormal brain to thisprocess (Rorden amp Brett 2000) The normalized ROIof each neglect patient was superimposed upon the T1template to find areas of maximal lesion overlap in eachgroup (see Results) as well as lesion volume usingMRIcro shareware (Rorden amp Brett 2000)
Experimental Procedure
Stimuli were presented on an SVGA monitor (29 22 in)viewed at 57 cm distance Each search display com-prised a virtual 9 7 grid each cell containing a single18 elementmdasheither a letter T (target) or a letter L (dis-tractor) (see Figure 1) The position of each elementwithin each cell was jittered with 19 targets and 44distractors randomly distributed over the virtual gridThe number of targets on the right and left halves ofthe display was balanced across trials Seven neglectpatients (Cases 5 8 10 11 14ndash16) were further testedon lsquolsquoTarget onlyrsquorsquo displays which had varying numbersof Ts (2 6 or 19) now with no L-shaped distractorsThe presentations of these lsquolsquoTarget onlyrsquorsquo screens wereinterspersed with those of the original search screens inrandom order in the same experimental session
Mannan et al 351
Eye movements were recorded using a video-basedpupil tracker with a temporal resolution of 4 msec andspatial accuracy of lt058 (EyeLink Sensorimotoric Sys-tems Berlin Germany) Subjects were asked to keeptheir head still The experimenter also monitored gazeposition on-line At the beginning of each session (andafter breaks in testing or following any unintended headmovement that disrupted gaze recordings) eye-trackingwas re-calibrated Each trial was initiated by the exper-imenter and began with a central fixation cross for 1 secthen the search display
Subjects were instructed to search for Ts (among anyLs) Critically while looking at any target they had toclick a response button held in the right hand onlyif they considered this target to be a new discovery(ie a target they had not clicked on before) Clickingthe response button did not change the display Theresponse button was held by the patient in a comfort-able position to the right of the midline Subjectswere instructed and reminded throughout testing toclick on each target found once only Any later re-click on
a previously found target thus indicates a failure to retainthe fact that this target had already been discoveredThe experimenter terminated each search when thesubject indicated verbally that they felt all targets in thedisplay had now been found Each subject searched aminimum of 5 displays (mean = 7 range 5ndash14) Eyemovements were analyzed off-line using custom soft-ware An automatic algorithm was used to extract sac-cades (saccade onset defined as eye speed rising above258 sec1 for two or more samples and a criterion ofminimum fixation duration of 60 msec) Blinks wereeliminated from the trace by visual inspection and sac-cade and fixation delineation were checked visually inthe record Next fixations were automatically assignedto items in the display if the mean position fell within18 of that item Two or more consecutive fixations onthe same item were re-scored as a single extendedfixation A re-fixation was scored only when at leastone fixation elsewhere on another item intervened
Most click responses were made during a fixation andthe few button-clicks occurring during a saccade were
Table 1 Patient Details
CaseAge
(years) PathologyVolume(cm3)
Age ofLesion (days)
Visual FieldDefect
Mean Deviationfrom Center on
Line Bisection Task(cm) (rightward +)
Mesulum Score(Maximum Possible
Score = 60) Lesion Type
1 68 Infarct 1282 64 + 867 17 PCA
2 76 Infarct 1528 227 + 003 41 PCA
3 68 Infarct 79 207 + 297 36 PCA
4 77 Infarct 251 158 + 150 54 PCA
5 63 Infarct 28 29 033 55 MCA
6 59 Hemorrhage 172 7 + 32 6 MCA
7 66 Hemorrhage 273 325 + 213 31 MCA
8 68 Infarct 309 3 25 17 MCA
9 29 Infarct 115 17 113 55 MCA
10 70 Infarct 1339 324 153 57 MCA
11 42 Infarct ndash 11 097 24 MCA
12 75 Infarct 757 38 013 4 MCA
13 51 Infarct 694 141 083 31 MCA
14 59 Infarct 1144 356 + 223 37 MCA
15 80 Infarct 261 44 35 6 MCA
16 64 Infarct ndash 30 033 55 MCA
MEAN 63 66 124 188 33
SE 4 13 32 5
Cases 1ndash9 had damage to the nonfrontal (posterior only) cortex whereas Cases 10ndash15 had both frontal and posterior (posterior and anterior) corticalinvolvement Case 16 is a patient with a subcortical lesion All but 3 of the patients with right MCA infarction had intact visual fields on confrontationTwo of these (Cases 7 and 14) were diagnosed with left homonymous hemianopia whereas one (Case 6) had a left inferior quandrantic field defectAll patients with PCA territory damage had partial or complete contralesional visual field loss No patient suffered from dysphasia
352 Journal of Cognitive Neuroscience Volume 17 Number 2
assigned to the previous fixation Clicks were assigned toa particular item if eye position at the time of the clickwas within 28 of that item or in the cases where morethan one item was within 28 of current eye position theclick response was simply attributed to the item closestto that eye position
Button-clicks were assigned to a particular item if eyeposition at the time of the click was within 28 of thatitem A re-click on an item required at least one fixationon another item between successive clicks (in practiceseveral fixations elsewhere typically intervened betweeninitial click and later re-click see Results)
Statistical analyses were performed using SPSS andSigmaplot (SPSS Science Chicago USA) Jonckheere sta-tistic was derived by hand Statistical significance was setas p lt 05
Acknowledgments
We thank the reviewers of this manuscript and Dr AndrewParton for their extremely helpful comments We thank thepatients who participated Dr J Wade Dr A Rudd and thestaff of the stroke units at Charing Cross and St ThomasrsquoHospitals This research was supported by grants from theWellcome Trust to M H C K and J D J D holds a RoyalSociety Wolfson Research Merit Award
Reprint requests should be sent to Drs Sabira Mannan andMasud Husain Division of Neuroscience and PsychologicalMedicine Imperial College London Charing Cross CampusLondon W6 8RF UK or via e-mail smannanimperialacukor mhusainimperialacuk
Notes
1 We are grateful to one of the reviewers for suggesting thistime-series analysis2 There was no significant difference between the number oftargets fixated between the two groups so differences in lsquolsquoclickomissionsrsquorsquo cannot be attributed to the number of fixatedtargets that had to be kept track of during search3 The correlation between these four points alone inFigure 9A is high [r(3) = 81 cf r(15) = 47 for the wholegroup]4 We have also tested 10 neglect patients and 10 controlstroke patients on a vertical version of the Corsi blocks task(clinically used to measure spatial working memory capacity)Neglect patients had significantly impaired capacity but wereable to localize single locations well suggesting that mislocal-ization is unlikely to account for their deficit on this spatialworking memory task
REFERENCES
Bartolomeo P Sieroff E Decaix C amp Chokron S (2001)Modulating the attentional bias in unilateral neglect theeffects of the strategic set Experimental Brain Research137 432ndash444
Bates E Wilson S M Saygin A P Dick F Sereno M IKnight R T amp Dronkers N F (2003) Voxel-basedlesion-symptom mapping Nature Neuroscience 6 448ndash450
Behrmann M Watt S Black S E amp Barton J J (1997)Impaired visual search in patients with unilateral neglect
An oculographic analysis Neuropsychologia 351445ndash1458
Bisiach E amp Vallar G (2000) Unilateral neglect in humans InG Rizzolatti (Ed) Handbook of neuropsychology (2nd edvol 1 pp 459ndash502) Amsterdam Elsevier
Cals N Devuyst G Afsar N Karapanayiotides T ampBogousslavsky J (2002) Pure superficial posterior cerebralartery territory infarction in The Lausanne Stroke RegistryJournal of Neurology 249 855ndash861
Chafee M amp Goldman-Rakic P (1998) Matching patterns ofactivity in primate prefrontal area 8a and parietal area 7ipneurons during a spatial working memory task Journal ofNeurophysiology 79 2919ndash2940
Chedru F Leblanc M amp Lhermitte F (1973) Visualsearching in normal and brain-damaged subjectsContributions to the study of unilateral subjectsCortex 9 94ndash111
DrsquoEsposito M Aguirre G K Zarahn E Ballard DShin R K amp Lease J (1998) Functional MRI studies ofspatial and nonspatial working memory Cognitive BrainResearch 7 1ndash13
Donkelaar P amp Muri R (2002) Craniotopic updating ofvisual space across saccades in the human posteriorparietal cortex Proceedings of the Royal Society London B269 735ndash739
Driver J amp Husain M (2002) The role of spatial workingmemory deficits in pathological search by neglect patientsIn the cognitive and neural bases of spatial neglect InH-O Karnath A D Milner amp G Vallar (Eds) The cognitiveand neural bases of spatial neglect Oxford OxfordUniversity Press
Duhamel J R Colby C amp Goldberg M E (1992) Theupdating of the representation of visual space in parietalcortex by intended eye movements Science 25590ndash92
Goldman-Rakic P S (1996) The prefrontal landscapemdashImplications of functional architecture for understandinghuman mentation and the central executive PhilosophicalTransactions of The Royal Society of London Series BBiological Sciences 351 1445ndash1453
Heide W Binkofski F Seitz R J Posse S Nitschke M FFreund H J amp Kompf D (2001) Activation offrontoparietal cortices during memorized triple-stepsequences of saccadic eye movements An fMRI studyEuropean Journal of Neuroscience 13 1177ndash1189
Heide W amp Kompf D (1998) Combined deficits ofsaccades and visuo-spatial orientation after cortical lesionsExperimental Brain Research 123 164ndash171
Husain M amp Kennard C (1996) Visual neglect associatedwith frontal lobe infarction Journal of Neurology 243652ndash657
Husain M Mannan S Hodgson T Wojciulik E Driver Jamp Kennard C (2001) Impaired spatial working memoryacross saccades contributes to abnormal search in parietalneglect Brain 121 941ndash952
Husain M amp Rorden C (2003) Non-spatially lateralizedmechanisms in hemispatial neglect Nature ReviewsNeuroscience 4 23ndash36
Jackson S R amp Husain M (1996) Visuomotor functionsof the lateral pre-motor cortex Current Opinion inNeurobiology 6 788ndash795
Kinsbourne M (1993) Orientational bias model of unilateralneglect Evidence from attentional gradients withinhemispace In I H Robertson amp J C Marshall (Eds)Unilateral neglect Clinical and experimental studies(pp 63ndash86) Hove Erlbaum
Konishi S Kyoichi N Uchida I Kikyo H Kameyama Mamp Miyauchi S (1999) Common inhibitory mechanism in
Mannan et al 353
human inferior prefrontal cortex revealed by event-relatedfunctional MRI Brain 122 981ndash991
Konishi S Kyoichi N Uchida I Sekihara K amp Miyashita Y(1998) No-go dominant brain activity in human inferiorprefrontal cortex revealed by functional magneticresonance imaging European Journal of Neuroscience10 1209ndash1213
Malhotra P Mannan S Driver J amp Husain M (2004)Impaired spatial working memory One component of thevisual neglect syndrome Cortex 40 667ndash676
Manly T Woldt K Watson P amp Warburton E (2002) Ismotor perseveration in unilateral neglect lsquolsquodrivenrsquorsquo by thepresence of neglected left-sided stimuli Neuropsychologia40 1794ndash1803
Mazzoni P Bracewell R Barash S amp Andersen R (1996)Spatially tuned auditory responses in area LIP of macaquesperforming delayed memory saccades to acoustic targetsJournal of Neurophysiology 75 1233ndash1241
McCarley J Wang R Kramer A Irwin D amp Peterson M(2003) How much memory does oculomotor search havePsychological Science 14 422ndash426
Mesulam M-M (1985) Principles of behavioural neurologyTests of directed attention and memory Philadelphia Davis
Mesulam M M (1999) Spatial attention and neglectParietal frontal and cingulate contributions to the mentalrepresentation and attentional targeting of salientextrapersonal events Philosophical Transactions of theRoyal Society of London Series B Biological Sciences354 1325ndash1346
Mort D Malhotra P Mannan S Rorden C Pambakian AKennard C amp Husain M (2003) The anatomy of visualneglect Brain 126 1986ndash1997
Na D L Adair J C Kang Y Chung C S Lee K H ampHeilman K M (1999) Motor perseverative behaviour ona line cancellation task Neurology 52 1569ndash1576
Pearson D G amp Sahraie A (2003) Oculomotor controland the maintenance of spatially and temporally distributedevents in visuo-spatial working memory QuarterlyJournal of Experimental Psychology 56A 1089ndash1111
Rafal R (1994) Neglect Current Opinion in Neurobiology4 231ndash236
Rizzolatti G amp Arbib M A (1998) Language within our graspTrends in Neurosciences 21 188ndash194
Robertson L C Lamb M R amp Knight R T (1988) Effectsof lesions of temporalndashparietal junction on perceptual andattentional processing in humans Journal of Neuroscience8 3757ndash3769
Rorden C amp Brett M (2000) Stereotaxic display of brainlesions Behavioural Neurology 12 191ndash200
Rusconi M L Maravita A Bottini G amp Vallar G (2002)Is the intact side really intact Perseverative responses inpatients with unilateral neglect A productive manifestationNeuropsychologia 40 594ndash604
Russell C Malhotra P amp Husain M (2004) Attentionmodulates the visual field in healthy observers and parietalpatients NeuroReport 15 189ndash193
Rypma B amp DrsquoEsposito M (2003) A subsequent-memoryeffect in dorsolateral prefrontal cortex Cognitive BrainResearch 16 162ndash166
Sapir A Hayes A Henik A Danzinger S amp Rafal R(2004) Parietal lobe lesions disrupt saccadic remapping ofinhibitory location tagging Journal of CognitiveNeuroscience 16 503ndash509
Smania N Martini M C Gambina G A T Palamara ANatale E amp Marzi C A (1998) The spatial distributionof visual attention in hemineglect and extinction patientsBrain 121 1759ndash1770
Sprenger A Kompf D amp Heide W (2002) Visual searchin patients with left visual hemineglect Progress in BrainResearch 140 395ndash416
Tobler P Felblinger J Burki M Nirkko A Ozdoba C ampMuri R (2001) Functional organisation of the saccadicreference system processing extraretinal signals in humansVision Research 41 1351ndash1358
Wilson B Cockburn J amp Halligan P W (1987)Behavioural inattention test Bury St EdmundsThames Valley
Wojciulik E Husain M Clarke K amp Driver J (2001)Spatial working memory deficit in unilateral neglectNeuropsychologia 39 390ndash396
Zihl J amp Hebel N (1997) Patterns of oculomotor scanningin patients with unilateral posterior parietal and frontallobe damage Neuropsychologia 35 893ndash906
Zihl J amp von Cramon D (1985) Visual field recovery fromscotoma in patients with postgeniculate damage A reviewof 55 cases Brain 108 335ndash365
354 Journal of Cognitive Neuroscience Volume 17 Number 2

rightward items because for them there is nowhereelse to look That is they might in principle be perfectlyaware that they are re-fixating old previously discoveredlocations on the right Alternatively they may mistakenlyjudge that they are discovering new targets when actu-ally re-fixating previously found right targets A criticalissue for distinguishing these possibilities is whether ornot the patients can discriminate if they are fixating newor old locations during their search
A recent study introduced a new paradigm preciselyin order to assess whether a neglect patient misjudgedpreviously searched rightward locations for novel un-searched locations (Husain Mannan et al 2001) In acomputerized search task target Ts had to be foundamong Ls (Figure 1) while eye movements were trackedCritically the patient was also asked to click on a re-sponse button only when fixating a target that was judgedto be a new discovery not when re-fixating an old targetthat had previously been found and clicked upon Notethat unlike standard pen-and-paper cancellation testsbut more analogous to search in the real world the clickresponse did not leave any visible trace so in this taskobservers have to remember whether the currentlyfixated target locations had previously been inspected
Husain Mannan et al (2001) studied just a singleright-hemisphere patient with left neglect in this para-digm and found that he lsquolsquore-clickedrsquorsquo at a pathologicallyhigh rate on rightward targets when re-fixating thosethat had previously been found and clicked upon It wassuggested that this tendency to make misjudgmentsin responses to previously searched locations mightcontribute to neglect When combined with any right-ward spatial bias in attention it might exacerbate neglectfor leftward items as the patient would not realize hewas repeatedly covering the same ground on the right(Husain Mannan et al 2001)
However this proposal was based on only a single-case to date To investigate it further the present study
applied the new paradigm to 16 neglect patients withmiddle cerebral artery (MCA) or posterior cerebral artery(PCA) stroke In addition to assessing how prevalentthe lsquolsquore-click deficitrsquorsquo might be this study has two impor-tant features First we performed time-series analysesof search performance to assess whether any misjudg-ments in responses to previously found targets de-pended on the time since a given target had previouslybeen fixated and clicked upon1 Second we mapped thelesions of our patients using high-resolution MRI (Mortet al 2003) and reconstructed areas of lesion overlapassociated with re-clicking Taken together these ap-proaches allowed us to distinguish between two dif-ferent types of pathological re-clicking Patients withdamage to the right intraparietal sulcus showed re-clickbehavior that depended on the time since a previouslyfound target had initially been fixated whereas rightinferior frontal patients re-clicked at a pathological rateregardless of the time intervening between initial dis-covery and re-fixation These results show that manybut not all neglect patients misjudge previously foundtargets as if they were new discoveries with the defi-cit and its nature depending on which brain areas aredamaged
RESULTS
Neglect in the Search Task and Re-fixations
As expected neglect patients missed significantly moretargets on the left half of the screen (mean 38) thanthe right [18 t(15) = 27 p lt 05 for an example seeFigure 1] Healthy age-matched volunteers showedequal detection of targets on the left and right sidesmissing 16 on the left and 14 on the right [t(14) =099 ns] Re-fixation rate for each participant is definedas total number of re-fixationstotal number of targetsfixated expressed as a percentage Both patient and
Figure 1 An example of a
search array on which aresuperimposed fixations
(dots) and the interpolated
scanpath of one neglect
patient (Case 15) Noteneglect of stimuli on the left
and many re-fixations on
stimuli to the right
Mannan et al 341
control subjects were more likely to re-fixate targets thandistractors (28 times and 27 times for patients and con-trols respectively)
As a group neglect patients had a significantly higherre-fixation rates (mean = 142) than healthy controls[40 t(29) = 35 p lt 01 see Figure 2A] Ten ofthe 16 patients had re-fixation rates that fell abovethe 95 percentile for controls The critical issue forthis study is whether re-fixations are associated withmisjudging previously searched locations for novelones This is indexed by the lsquolsquore-click ratersquorsquo whichindicates any tendency for subjects to erroneously re-spond to previously found targets as if they were anew discovery
Re-click Rates
The re-click rate for each participant is given by totalnumber of re-clickstotal number of targets clicked uponexpressed as a percentage The mean re-click rate for
neglect patients (30) was significantly higher than forcontrols [6 t(29) = 31 p lt 01] who rarely consid-ered a re-fixated target to be a new discovery (Figure 2B)For the neglect group the probability of re-clicking washighest for targets to the right (Figure 3) In total 10 ofthe 16 patientsmdashhereafter termed high re-click groupmdashhad re-click rates beyond the upper 95 confidenceinterval for controls (Figure 2B) The remaining 6 pa-tients (low re-click group) had re-click rates within thenormal range demonstrating the re-clicking deficit isnot present in all neglect patients Moreover this resultalso shows that brain damage per se does not lead to apathological re-click rate Later (in the section on Ana-tomical Correlates) we show that the deficit cannot alsobe attributed to lesion size but rather is associated withdamage to specific brain regions
Importantly although 7 patients had both a high re-click and a high re-fixation rate 3 patients had an ab-normally high re-fixation rate and a normal re-click rateThus abnormal re-fixation rate alone does not neces-sarily identify patients who fail to distinguish betweennovel targets and previously inspected ones Hence theimportance of measuring re-click rates Neverthelessacross the group of patients there was a significant cor-relation between re-click and re-fixation rates (Figure 4)[r(15) = 78 p lt 0001] There was no such correla-tion between re-fixation and re-click rates for healthycontrols [r(14) = 05 ns]
Interestingly high re-clicking neglect patients notonly re-clicked more often on targets they had alreadyclicked upon but they also more frequently failedto click upon a fixated target they had not previouslyclicked upon The high re-click neglect group omittedto click on significantly more newly fixated targets
Figure 3 The distribution over space of re-clicks made by neglect
patients The total number of re-clicks made within each virtual column
of the screen is expressed as a percentage of the total number ofre-clicks made by patients
Figure 2 (A) Re-fixation rates for neglect patients and healthyindividuals with individual mean re-fixation rates for each subject
shown as circles Ten of 16 patients had re-fixation rates which fall
above the 95th percentile for healthy volunteers denoted by the
dashed line (B) Re-click rates for neglect patients and healthyvolunteers Ten patients had re-click rates above the 95th percentile
for healthy volunteers
342 Journal of Cognitive Neuroscience Volume 17 Number 2
than the low re-click neglect group [mean = 30omissions SE 4 vs 19 and 3 t(14) = 231 p lt05]2 Such an association between these lsquolsquoclick omis-sionsrsquorsquo and high re-click rate would not be expected ifre-clicking was entirely due to a lsquolsquoperseverativersquorsquo failureto inhibit responses to rightward targets because lsquolsquoclickomissionsrsquorsquo represent failures to respond Nor wouldthey be explained by items on the right being per-ceived to be more salient and thus automatically cap-turing responses
Anatomical Correlates of High Re-click Rates
Although 10 neglect patients demonstrated significant-ly elevated re-click rates 6 did not To determinewhether key anatomical locations underlie the distinc-tion between these groups we sought to isolate brainregions that were damaged in neglect patients withhigh re-click rates and equally importantly spared inpatients without the deficit Critically there was nocorrelation between lesion volume and re-click rate[r(13) = 31 ns] or between lesion age and re-clickrate [r(13) = 30 ns] Each patient was assigned toone of two subgroups according to whether they hadfrontal involvement or not Neglect patients withlsquolsquoposterior onlyrsquorsquo lesions had damage posterior to thecentral sulcus with no damage anterior to the pre-central sulcus (n = 9) Neglect patients with lsquolsquoposte-rior and anteriorrsquorsquo involvement had cortical damageanterior to the central sulcus (n = 5) all patients inthis subgroup also had some posterior damage
Parietal Correlate of High Re-click Rate
Among the 9 patients in the lsquolsquoposterior onlyrsquorsquo subgroup4 had an abnormally high re-click rate 3 with MCA ter-
ritory damage (Cases 5 6 and 8) and only 1 with PCAdamage (Figure 5A) This single PCA patient (Case 2)had damage to white matter deep to the occipito-temporo-parietal junction in contrast to the other 3PCA patients whose lesions did not extend so faranteriorly or dorsally The area of damage commonto all 3 MCA patients with abnormally high re-clickratesmdashwhile also being intact in the 5 posterior sub-group patients with normal re-click ratesmdashwas locatednear the anterior end of the intraparietal sulcus closeto the lateral bank (Figure 5C Talairach 46 40 28for centroid volume 058 cm3) This area was not sim-ply the most common region of overlap for all 8 MCApatients that was located more dorsally (Talairach37 55 38)
In order to assign statistical significance to the areaof overlap we performed a voxel-by-voxel analysis (Bateset al 2003) For every voxel that was damaged in atleast 3 subjects we compared the re-click rate of pa-tients with damage to that voxel with those of patientswithout damage to that particular voxel producing at statistic for a given voxel The area shown in yellow(Figure 5C) yields a value of t(7) = 491 indicatingsignificantly higher re-click rates associated with dam-age to this region ( p lt 002) This difference survivesBonferroni correction for multiple comparisons ( p lt05) where the number of comparisons was 6 (therewere only 6 regions in which the lesions of 3 or morepatients overlapped)
An alternative analytical approach is to focus only onpatients with relatively small lesions Seven patients(Cases 4ndash9 and 15) had focal parietal lesions lt31 cm3all had posterior lesions except Case 15 who also hadinvolvement anterior to the central sulcus The maxi-mum area of overlap common to all 4 patients with highre-click rates (Cases 5 6 8 15) which also cruciallyremained undamaged in the 3 patients with normal re-click rates (Cases 4 7 9) again centered on the lateralintraparietal sulcus (Figure 6) slightly more dorsally(Talairach 37 39 37 volume 011 cm3) and involvinga subset of the critical voxels identified in Figure 5 Notethat this overlap region will probably also involve whitematter fibers including those that may connect the pa-rietal cortex to the frontal lobe
Frontal Correlate of High Re-click Rate
Four of the five patients in the lsquolsquoposterior plus anteriorrsquorsquoneglect subgroup had an abnormally high re-click rateThe area of damage common to all four was in theinferior frontal lobe (Talairach 38 18 16 for centroid178 cm3) in a region corresponding to Brodmannrsquosarea 44 part of the premotor cortex (Figure 7A) Threeof the four posterior plus anterior neglect patients witha pathologically high re-click rate also had damage ex-tending into or bordering the ventrolateral prefrontalcortex The remaining patient who had the highest re-
Figure 4 Mean re-click rates plotted against mean re-fixation rates
for individual neglect patients There was a significant correlation[r(15) = 78 p lt 0001]
Mannan et al 343
click rate of all patients (Case 15) had damage involvingthe dorsolateral prefrontal cortex Only 1 of the 5 pos-terior plus anterior patients had a normal re-click rate(Figure 7B) Her lesion was posterior to the region ofoverlap seen in the four frontal patients with abnor-mally high re-click rates It involved the posterior premo-tor cortex and motor cortex and extended caudally intothe inferior parietal lobe Thus the subtraction of herlesion from the remaining posterior plus anterior patientswith high re-click rates (Figure 7C) does not alter theirmaximum overlap region (displayed in Figure 7A)
Anatomical Correlates of the lsquolsquoRe-fixationrsquorsquo Deficit
Ten neglect patients had a high re-fixation rate but 6patients did not There was no correlation betweenlesion volume and re-fixation rate [r(13) = 22 ns]or between age of lesion and re-click rate [r(13) =36 ns] Although some patients in either the poste-rior (Cases 1 2 4ndash8) or frontal group (Cases 11 13 15)had high re-fixation rates other patients with highlysimilar lesion locations had normal re-fixation ratesAs a consequence voxel-by-voxel analysis (Bates et al
Figure 5 The
neuroanatomical correlates of
the deficit in retaining spatiallocations for neglect patients
with posterior only (nonfrontal)
lesions (n = 9) (A) Individuallesions of patients with high
re-click rate (patients with MCA
damage Cases 8 5 6 patient
with PCA damage Case 2) andoverlap map for these
4 patients where maximum
overlap occurs for a small region
(shown in green) around theright anterior intraparietal
sulcus damaged in the 3 MCA
patients (B) Individual lesionsof patients with normal re-click
rate (patients with MCA
damage Cases 9 and 7 patients
with PCA damage Cases 1 3 4)and overlap map for these
5 patients with maximum
overlap occurring in the
occipital cortex Importantlynone of the 5 patients with
normal re-click rates have
damage to the common areaof overlap for patients with high
re-click rates shown in A
(C) Contrast map highlighting
intraparietal locus which isdamaged in 3 out of 4 patients
with a high re-click rate and
intact in the 5 patients with
normal re-click rates (shown inyellow) Regions that are
damaged in patients with high
re-click rates and spared in
those patients with normalre-click rates are shown in red to
yellow whereas regions in blue
denote areas that are commonto patients with
normal re-click rates but
spared in patients with high
re-click rates
344 Journal of Cognitive Neuroscience Volume 17 Number 2
2003) revealed no anatomical specificity associated withhigh re-fixation rates [t(7) = 085 ns] unlike thatassociated with high re-click rates even when onlyfocal lesions were considered [t(5) = 165 ns]
Time Interval between Clicking and Re-clicking ona Target
We next examined the probability of lsquolsquore-clickingrsquorsquo on atarget when re-fixating it as a function of the time lag ornumber of intervening saccades between the first clickon that target and the subsequent revisit An increasedprobability of re-clicking with increasing delay (or inter-vening number of saccades) would be consistent a failureto keep track of spatial locations during search whereasa flat or decreasing function might arise from impairedresponse inhibition (lsquolsquoperseverativersquorsquo behavior) or even
a failure to encode target location This time-series anal-ysis for the 10 patients with high re-click rates revealedtwo distinct patterns of behavior Half the patients de-monstrated a significantly increasing probability of mis-judging an already found item to be a novel one withincreasing delay (Figure 8A) or intervening saccades(Figure 8C) whereas the other 5 patients had a constantor even decreasing probability (Figure 8B and D)
Importantly most (45) patients in the group withincreasing re-click functions had damage to the focalparietal location previously identified in the analysisof patients with focal lesions and high re-click rates(Figure 6C) The other patient was Case 2 who had alarge PCA lesion extending to the white matter deep tothe occipito-temporo-parietal junction By contrast noneof the patients with flatdecreasing functions had dam-age to the key parietal region Instead the latter group
Figure 6 The
neuroanatomical correlates of
the deficit in retaining spatial
locations for neglect patientswith focal lesions (n = 7)
(A) Individual lesions of
patients with high re-click rate
(all 4 patients have MCAdamage) and overlap map for
these 4 patients where
maximum overlap occurs for asmall region around the right
intraparietal sulcus (shown in
red) damaged all 4 MCA
patients (B) Individual lesionsof patients with normal re-click
rate (all patients with PCA
damage) and overlap map for
these 3 patients with maximumoverlap occurring in the
occipital cortex Importantly
none of these 3 patients withnormal re-click rates have
damage to the common area of
overlap for patients with high
re-click rates shown in A(C) Contrast map highlighting
intraparietal locus which is
damaged in 4 out of 4 patients
with a high re-click rate andintact in the 3 patients with
normal re-click rates (shown in
yellow) Regions that are
damaged in patients with highre-click rates and spared in
those patients with normal
re-click rates are shown in redto yellow whereas regions in
blue denote areas that are
common to patients with
normal re-click rates butspared in patients with high
re-click rates
Mannan et al 345
had frontal cortical or subcortical involvement Thusincreasing re-clicking with time interval was associatedwith parietal damage whereas flat re-click functionswere associated with frontalsubcortical lesions
For neglect patients as a whole the intervals betweenan initial click on a target and a subsequent re-click werelong (mean = 40 saccades) but much shorter betweenan initial fixation and a later re-fixation (mean = 5) withthis difference being significant [t(15) = 315 p lt01] This again emphasizes the dissociation betweenre-fixations and re-clicks and also demonstrates that re-clicking does not occur on most re-fixations
Relationship of Re-click Rate to Neglect Severity
We assessed three separate measures of neglect Thefirst was the difference between miss rates for left ver-sus right targets within the experimental search taskitself the other two measures of neglect were based onthe clinical pen-and-paper measures of Mesulam shape-cancellation (Mesulam 1985) and line bisection fromthe BIT (Wilson Cockburn amp Halligan 1987)
On the experimental search task there was a moderatebut significant correlation between re-click rate and ne-glect in search across the group [r(15) = 47 p lt 05
Figure 9A] Thus the greater the re-clicking on right-ward items the greater the severity of leftward neglectThere was also a moderate but again significant corre-lation between re-click rates and the severity of individualpatientsrsquo clinical neglect as indexed by the cancellationmeasure [r(15) = 46 p lt 05 Figure 9B] Thesecorrelations were driven largely by three of the patientsidentified in our focal lesion analysis (Figure 6A) withinvolvement of the key parietal region and high re-clicking behavior Their data are shown in filled circlesto the right in the graphs displayed in Figure 93 Thecorrelation between deviation on line bisection andre-click rate was not significant [r(15) = 13 ns] How-ever one would not predict that a deficit in retaininglocations already visited (or response inhibition) wouldnecessarily impair performance on a task such as linebisection
Effect of Varying Number of Targets
In 7 patients (Cases 5 8 10 11 14ndash16) we also exam-ined the effect of varying the number of targets in thesearch task Patients viewed lsquolsquotarget onlyrsquorsquo displays con-taining either 2 6 or 19 targets randomly distributedon the screen without any L-shaped distractors In-
Figure 7 Theneuroanatomical correlates of
the re-click deficit in neglect
patients with posterior plusanterior lesions (n = 5)
(A) Individual lesions of
patients with high re-click rate
(all 4 patients have MCAdamage) and overlap map for
these 4 patients where
maximum overlap occurs for
a region in the right inferiorfrontal lobe (shown in red)
damaged all 4 MCA patients
(B) Individual lesions of a single
patient with normal re-clickrate (C) Contrast map
highlighting the inferior frontal
locus (shown in yellow) whichis damaged in 4 out of
4 patients with a high re-click
rate and intact in the patient
with normal re-click rateRegions that are damaged in
patients with high re-click rates
and spared in the patient
with normal re-click rate areshown in red to yellow
whereas regions in blue denote
areas that are damaged in thepatient with normal re-click
rates but spared in patients with
high re-click rates
346 Journal of Cognitive Neuroscience Volume 17 Number 2
creasing the number of targets led to significantly higherre-click rates ( Jonckheere statistic = 133 p lt 01Figure 10) as well as more severe neglect (Jonckheerestatistic = 129 p lt 01) consistent with the proposalthat the mechanisms underlying pathological re-clickingmight contribute to neglect behavior Note that a lsquolsquoper-severativersquorsquo failure of response inhibition would not onits own easily explain this finding For the same absolutenumber of click responses one would predict a higherre-click rate with smaller set-sizes than with larger ones(see also Husain Mannan et al 2001 for a discussion)Although the number of subjects were small there wasno difference in the performance pattern between the3 patients who had increasing re-click rate functions inthe time-series analysis and the 4 patients with flat ordecreasing re-click rate functions
Varying Perceptual Load during Search
Can re-click behavior be explained simply by impairedvisual perception of targets from discriminators Neglectpatients did find it more difficult than control subjectsto select targets from distractors fixating on average19 distractors for every target found [cf 14 for controlsubjects t(29) = 485 p lt 0001] Patients also some-times clicked on distractors (64) but no more often
than controls (61) and it is possible that this rate iseffectively inflated by our strict criterion of considering aclick to be a response on the item that is being fixated atthe time of the click (It is likely that sometimes ob-servers click in response to targets they fixated one ormore saccades previously) Re-clicks on distractors wereextremely rare (06 cf 30 for targets)
To examine the effect of reduced perceptual load in8 patients (Cases 5 7 8ndash10 13 1 4 16) 6 of whom werehigh re-clickers we used search displays with salientcross (+) targets rather than Ts L distractors wereunchanged Patients fixated significantly fewer distrac-tors when searching for cross (+) targets [distractortarget ratio of 134 cf 181 for T targets t(7) = 60 p lt001] suggesting they did indeed find targetndashdistractordiscrimination easier Importantly however there wasno significant difference in re-click rate [26 for +targets cf 27 for T targets t(14) = 041 ns] or evenin re-fixation rate [100 for + targets cf 119 fort(14) = 041 ns] Thus these changes in perceptualload did not influence re-click performance
DISCUSSION
On this multitarget visual search task neglect patientsmissed targets to their left and also re-fixated targets
Figure 8 The probability of
re-clicking or re-fixating on
target as a function of delay
between the original click andsubsequent re-fixation (for
patients with high re-click
rates) (A) Probability functions
for patients with significantpositive slopes Four patients of
these patients have damage to
the focal intraparietal locusidentified in Figure 6C whereas
the single patient with a PCA
lesion and no damage to the
critical parietal locationindicated by symbol has only
a very shallow positive slope
(B) Probability functions for
patients with no significantpositive slope None of these
5 patients have damage to the
critical intraparietal region (C)As for A but for delay measured
in terms of intervening
fixations (D) As for B but for
delay measured in terms ofintervening fixations
Mannan et al 347
to the right much more often than healthy individuals(Figure 2A) Critically 10 of the 16 neglect patients re-clicked on rightward targets at a significantly higher ratethan healthy control subjects (Figure 2B) For 5 of thesepatients the probability of re-clicking increased withtime (Figure 8A) whereas the other 5 neglect patientswith high re-click rates demonstrated a constant or de-creasing probability of re-clicking over time (Figure 8C)The remaining 6 neglect patients had re-click rateswithin the normal range demonstrating that neitherneglect nor brain damage per se leads to a pathologicalre-click rate
High-resolution MRI mapping of lesions revealed im-portant anatomical dissociations within the neglect pop-ulation Patients with pathological re-click rates hadlesions affecting either the right intraparietal sulcus orthe inferior frontal lobe the only exception (Case 2) hada large occipito-temporal lesion that extended deep intothe parietal white matter Neglect patients with normalre-click rates did not have damage to either of thecritical locations in the intraparietal sulcus or inferiorfrontal lobe (Figures 5 6 and 7) Instead they had moredorsal lesions (for MCA territory patients) or occipitaland medial-temporal damage (for PCA territory lesions)which can also result in neglect (Mort et al 2003Cals Devuyst Afsar Karapanayiotides amp Bogousslavsky2002) Importantly lesion location rather than the sizeor age of lesion was critical to the presence of the re-clicking impairment Across the group of neglect pa-tients there was a moderate but significant correlationbetween re-click rate and degree of neglect (Figure 9)driven largely by patients with parietal damage who hadboth a high re-click rate and also severe neglect consis-tent with the proposal that mechanisms underlying thisbehavior in parietal patients might have a functionallyimportant role in determining the severity of neglect
Role of Right Intraparietal Sulcus
Notably for all patients with damage to the critical locusin the right intraparietal sulcus (which may also involveparieto-frontal white matter fibers) the probability of re-clicking on a target increased with time (or interveningsaccades) since initial discovery (Figure 8) Because theyrarely re-clicked at short intervalsmdasheven though themajority of re-fixations occurred during this periodmdashthis pattern of behavior suggests that for these patientstarget locations were actually veridically perceived and
Figure 9 (A) Neglect on experimental search task (19 targets and44 distractors) plotted against mean re-click rate for individual neglect
patients Percentage neglect was calculated as the difference between
the number of targets missed on the left versus right expressed as apercentage of the total number of targets missed There was a
significant correlation between the degree of neglect and the re-click
rate [r(15) = 465 p lt 05] Patients with high re-click rates are
highlighted by black circles (for patients with damage to the focalparietal location identified in Figure 6C) and light gray circles (for
patients with frontal damage) while the single patient with PCA
damage and a high re-click rate is indicated by dark gray (B) The
number of targets missed on the Mesulam shape-cancellation taskplotted against mean re-click rate for individual neglect patients Again
there was a significant correlation between the number of targets
missed and the mean re-click rate [r(15) = 456 p lt 05] Patients withhigh re-click rates are denoted as in A
Figure 10 Mean re-click rate (filled circles) and degree of neglect
(unfilled squares) increased systematically with increasing numbers of
targets in target-only displays
348 Journal of Cognitive Neuroscience Volume 17 Number 2
well encoded into working memory Furthermore de-tailed single-case studies of two parietal patients withhigh re-click rates (Malhotra Mannan Driver amp Husain2004 Husain Mannan et al 2001) demonstrate usingdifferent psychophysical methods that these patientscan accurately encode spatial locations4 Increasing re-clicks with time (or intervening saccades) would how-ever be consistent with a deficit in keeping track oftarget locations during extended search
Several lines of evidence support the proposal thatthe critical intraparietal sulcus region we have identifiedmay have an important role in the spatial transforma-tions required to keep track of spatial locations acrosssaccades In rhesus monkeys neurons within the lateralbank of the intraparietal sulcus (area LIP) encode andlsquolsquomaintainrsquorsquo the remembered locations of saccadic tar-gets over time (Chafee amp Goldman-Rakic 1998 MazzoniBracewell Barash amp Andersen 1996) Some visuallyresponsive neurons in the LIP also appear to shift theirretinotopic representation at the onset of an intendedsaccade as if lsquolsquore-mappingrsquorsquo representations of objects inspace to update them across shifts of gaze (DuhamelColby amp Goldberg 1992)
In humans this computation has been investigatedusing the double-step saccade paradigm where twoflashes of light appear in quick succession such thatthe second target disappears before a saccade can bemade to the first target Patients with lesions involvingthe right parietal cortex are impaired at performing thistask suggesting that they are unable to update spatialrepresentations accurately after even one saccade (Heideamp Kompf 1998 Duhamel et al 1992) Similarly trans-cranial magnetic stimulation of the posterior parietalcortex prior to the second saccade reduces its accuracy(Donkelaar amp Muri 2002) and a recent study of patientswith parietal lesions also implicates this area in thespatial updating of inhibition of return after a saccade(Sapir Hayes Henik Danzinger amp Rafal 2004) Func-tional imaging studies of double-step (Tobler et al2001) and triple-step memory-guided saccades (HeideBinkofski et al 2001) have identified a region withinthe intraparietal sulcus (Talairach coordinates 40 4044 for Tobler et al and 44 48 36 Heide et al) This isclose to the critical region of lesion overlap we haveidentified in the posterior neglect patients with high re-click rates (Talairach 37 39 37)
We suggest that in our patients with lesions involvingthe right intraparietal sulcus uncertainty about locationsof previously found targets increases with each saccadeThis uncertainty is further exacerbated by interferencefrom newer target locations leading to increased prob-ability of misjudging old target locations for new assearch progresses Such progressively worsening uncer-tainty might be due to an accumulating impairment inlsquolsquore-mapping target locationsrsquorsquo across saccades or im-paired lsquolsquomemoryrsquorsquo of previously inspected target loca-tions across saccades As noted above there is evidence
for both types of signalmdash re-mapping and maintainingsaccadic spatial memorymdashin the parietal cortex and it ispossible that disruption to both these functions contrib-utes to the re-click deficit we have identified Certainlythe two potential types of deficit are not mutuallyincompatible
A slightly different account of the re-clicking behaviormight be framed in terms of the intriguing hypothesisproposed by Manly Woldt Watson and Warburton(2002) to explain visible re-marking on a standard can-cellation task They argue that such behavior is due topatients erroneously transposing leftward stimuli to theright resulting in apparent re-cancellation These au-thors found that removing targets from the left re-duced both neglect and re-marking However they didnot monitor eye movements so it is possible that sometargets that are logged as neglected (because they werenot cancelled) were nevertheless fixated and thereforeform part of the load on memory during search Fur-thermore in our experimental study subjects with ahigh re-click rate also failed to click on a higher pro-portion of fixated targets Manly et alrsquos proposal wouldnot easily account for such lsquolsquoclick omissionsrsquorsquo becausethese refer to fixated targets that were not actuallyresponded to
Finally it might be argued that increasing re-clickingwith search duration reflects a deficit in spatial workingmemory (SWM) that is independent of saccadic eyemovements (ie a decay in memory that is not trans-saccadic) Because time and number of interveningsaccades were not independently manipulated in thisunrestricted visual search paradigm we are not ablecategorically to distinguish between these possibilitiesWe note however that functional imaging studies ofSWM tasks that do not require eye movements oftendemonstrate activity related to the maintenance ofspatial information which is generally more posteriorand dorsal than the parietal locations isolated in thisstudy (DrsquoEsposito et al 1998) Moreover recent behav-ioral data indicate that SWM is more vulnerable tosaccadic eye movements than shifts in attention or limbmovements (Pearson amp Sahraie 2003)
Frontal Lesion Site Associated withHigh Re-click Rates
Four out of 5 neglect patients with lesions involvingposterior plus anterior damage also showed a high re-click rate and the area of damage common to thesepatients centered on the right inferior frontal lobe cor-responding to Brodmannrsquos area 44 (Figure 7C) Thisregion has previously been associated with frontalneglect (Husain amp Kennard 1996) and is consideredto be part of the ventral premotor cortex and thehomologue of Brocarsquos areas in the left hemisphere(Rizzolatti amp Arbib 1998 Jackson amp Husain 1996) Inthese frontal patients the probability of re-clicking did
Mannan et al 349
not increase with time interval between the first clickand re-fixation but rather it stayed constant or declinedwith increasing delays
Such behavior might be consistent with a failure toinhibit responses to rightward locationsmdasha type oflsquolsquoperseverativersquorsquo behavior Note though that this is notnecessarily the same type of perseverative responseas seen when some patients with neglect re-markpreviously found items on standard cancellation tests(Rusconi et al 2002 Na et al 1999) In those casestargets that are visibly cancelled are often marked againwhereas in our computerized search tasks no marks arevisibly left on targets once they have been clicked uponMoreover only one of our neglect patients (Case 1) de-monstrated re-cancellation of visibly cancelled targetson pen-and-paper tasks and then only on one occasionImportantly this patient actually had a normal re-clickrate (67) on our task and his stroke was in the PCAterritory sparing both the intraparietal sulcus and fron-tal lobe
In support of a role for the right frontal lobe in in-hibiting responses to previously found targets recentimaging studies of response inhibition have highlightedregions of the right inferior frontal gyrus (IFG) very closeto the centroid of damage for our frontal patients (38 1618 cf 43 22 21 Konishi Kyoichi Uchida Sekihara ampMiyashita 1998 and 41 16 19 Konishi Kyoichi UchidaKikyo et al 1999) Alternative explanations for the highprobability of re-clicking directly after clicking might bethat these patients have impaired perceptual discrimina-tion of targets impaired perceptual localization of targetsor that they encounter difficulty in encoding spatiallocations into working memory Our data are not sugges-tive of a perceptual discrimination problem as changingthe perceptual load did not inf luence re-click ratesMoreover the patients still had high re-click rates in thetarget only conditions (Figure 10) where no perceptualdiscrimination of targets and distractors was necessaryPoor perceptual localization would seem unlikely giventhat these patients have frontal lesions Furthermoreusing a vertical SWM task similar to the Corsi blocktapping task (Malhotra et al 2004) we have found thatpatients with neglect can localize and perceptually en-code single target locations as well as controls and aresignificantly impaired at remembering two or more loca-tions shown in sequence consistent with the presentobservations Some recent functional imaging data sug-gest that activation within the prefrontal cortex corre-lates well with successful encoding of material intoworking memory (Rypma amp DrsquoEsposito 2003) Furtherresearch is required to distinguish between these pos-sible explanations for the mechanisms underlying there-click deficit in frontal patients
The only patient with frontal damage who did re-clickmore frequently with increasing time had damage cen-tered on the dorsolateral prefrontal cortex (althoughalso encroaching on the IFG) Neurons in the dorsolat-
eral prefrontal cortex like those in the LIP encode theremembered locations of saccadic targets (Goldman-Rakic 1996) This patient (Case 15) who had the high-est re-click rate of all also had damage to the criticalparietal location described above (in contrast to all otherfrontal patients) Thus she may have suffered from acombination of underlying deficits
Re-fixations and Misjudging PreviouslyInspected Locations during Search
In this search task re-fixation and re-click rates weresignificantly correlated for neglect patients Howeverthere were important dissociations within individuals 2patients with PCA lesions had re-fixation rates of 220and 80 respectively but both had a re-click rate ofonly 7 These are clearly examples of individuals whodo not show pathological re-clicking despite having highre-fixation rates High re-fixation rates may result froma lack of integration of spatial information due to V1damage (Zihl amp von Cramon 1985) in our PCA patientsor possibly because of reduced inhibition of return(Bartolomeo Sieroff Decaix amp Chokron 2001) What-ever the exact cause for dissociations between re-fixa-tions and re-click rates our data suggest re-fixation doesnot necessarily index the same underlying deficits asrevealed by our re-click measure High re-fixation ratesunlike high re-click rates were also not significantlyassociated with damage to any specific cortical locationagain demonstrating a dissociation between re-fixationsand pathological re-clicking
The relationship between re-click rates and neglect isunlikely to be a simple one In our study there was amoderate but significant correlation between the twomeasures suggesting that this association is unlikely tohave arisen by chance However non-neglect patientscan also exhibit a high re-click rate (unpublished obser-vations from our laboratory) Taken together our resultssuggest that difficulties in keeping track of previouslyinspected targets (because of impaired trans-saccadic re-mapping andor memory) may be one contributingcomponent to visual neglect in patients with lesions in-volving the right intraparietal sulcus As mentioned thesearch task we used differed in one striking aspect fromstandard cancellation tests subjects do not visibly markthe targets they visit Whereas in cancellation tests pa-tients mark each target they find which might appear todispense with any need to remember which locationshave been searched before However it should be ap-preciated that in most standard cancellation tests theindividual marks made by patients may not be clearlyvisible in peripheral vision particularly if neglect pa-tients have a local spatial bias (Rafal 1994 RobertsonLamb amp Knight 1988) or a restricted field of view(Russell Malhotra amp Husain 2004) Thus when com-bined with a rightward attentional bias keeping track ofsearched locations may be critical to prevent gaze from
350 Journal of Cognitive Neuroscience Volume 17 Number 2
returning to rightward locations even if they have beenmarked before as the marks may not be visible untilgaze returns to them Consistent with this hypothesisneglect is greater when cancellation is performed usinginvisible marks compared to large and salient red marksthat can be seen peripherally (Wojciulik Husain Clarkeamp Driver 2001)
A task like the present search-and-click paradigm maymore closely resemble search in the real world than dotraditional cancellation tests When we examine a visualscene we do not leave marks on each object we ex-amine Instead just as in the search task used in thisstudy we have to keep track of the locations we visit ifwe are to avoid inefficient re-examination of thoselocations Neglect patients with damage to the rightintraparietal sulcus appear to be impaired at keepingtrack of searched locations in this way When combinedwith their rightward attentional bias we propose thatsuch an impairment leads to recursive search of stimulito the right in experimental search tasks as used hereand also in daily life where neglect is so apparentdespite unlimited viewing time
The paradigm we have used has the important virtuesof allowing patients to explore freelymdashas in real-worldsearch or clinical cancellation tasksmdashand relating theirre-click behavior directly to the severity of neglect in thesame task But it might be argued that it would be usefulalso to probe SWM in a more controlled manner by re-stricting observersrsquo eye movements to a specific set oflocations and probing memory of previously visitedplaces A recent study using a gaze-contingent para-digm has elegantly probed the capacity of spatial mem-ory during search in healthy observers and foundevidence for memory for up to four previous locations(McCarley Wang Kramer Irwin amp Peterson 2003)Variants of this paradigm may in the future be usefulto probe the capacity of SWM across saccades in patientswith neglect and also to probe further whether thereare differences between neglect patients with parietaland frontal lesions
METHODS
Subjects Imaging and Lesion Mapping
Sixteen patients with left neglect following right hemi-sphere infarction or hemorrhage were recruited Con-sent was obtained according to the Declaration ofHelsinki as approved by the hospital ethical commit-tees Presence of neglect was determined by clinicalobservation plus bedside tests including the Mesulamshape-cancellation task (Mesulam 1985) and line bisec-tion task (Wilson et al 1987) (see Table 1) Twelvepatients had MCA territory strokes the rest involvedonly the PCA territory which can also result in neglect(Cals et al 2002) Only one neglect patient (Case 1 whodid not have frontal involvement) demonstrated re-
cancellation of visibly cancelled targets on pen-and-paper tasks and then only on one occasion Fifteenage-matched healthy volunteers were controls (meanage = 63 SD = 14 vs 63 SD = 13 for patients)
All patients had clinical CT scans In addition dedi-cated high-resolution structural MRI scans were alsoobtained for 1416 neglect patients MRI scans wereunobtainable for the other two (Cases 11 and 16)because of claustrophobia or loss to follow up In theMCA neglect group median time between stroke andscan was 63 days with the shortest interval being35 days for the PCA neglect group one patient had tobe scanned relatively early at 9 days but the mediantime was 140 days MRI scans comprised one 256-sliceT1-weighted acquisition (MPRAGE TR 97 msec TE4 msec TA 12 min) performed in the coronal planealigned perpendicular to the anteriorndashposterior commis-sural line on a Siemens Vision 15-T scanner Within-and between-slice resolution was 1 mm For each pa-tient the area of damage was determined by detailedvisual inspection of the digital image for every singleslice Lesion delineation was performed by neurologists(authors D M and M H) by tracing by hand theboundary of the lesion onto every single 1-mm-thickaxial slice using a graphics tablet (WACOM Intuos A6)Combining all the slices then led to a 3-D region of in-terest (ROI) for each patient For one patient (Case 15)there were two areas of discrete focal damage one inthe frontal cortex and the other in the parietal cortex
Normalization of each MRI including the 3-D ROI tothe T1-template was performed with SPM99 (wwwfilionuclacukspm) The ROI was used as a mask for thelesion during determination of alignment parameters tominimize the contribution of abnormal brain to thisprocess (Rorden amp Brett 2000) The normalized ROIof each neglect patient was superimposed upon the T1template to find areas of maximal lesion overlap in eachgroup (see Results) as well as lesion volume usingMRIcro shareware (Rorden amp Brett 2000)
Experimental Procedure
Stimuli were presented on an SVGA monitor (29 22 in)viewed at 57 cm distance Each search display com-prised a virtual 9 7 grid each cell containing a single18 elementmdasheither a letter T (target) or a letter L (dis-tractor) (see Figure 1) The position of each elementwithin each cell was jittered with 19 targets and 44distractors randomly distributed over the virtual gridThe number of targets on the right and left halves ofthe display was balanced across trials Seven neglectpatients (Cases 5 8 10 11 14ndash16) were further testedon lsquolsquoTarget onlyrsquorsquo displays which had varying numbersof Ts (2 6 or 19) now with no L-shaped distractorsThe presentations of these lsquolsquoTarget onlyrsquorsquo screens wereinterspersed with those of the original search screens inrandom order in the same experimental session
Mannan et al 351
Eye movements were recorded using a video-basedpupil tracker with a temporal resolution of 4 msec andspatial accuracy of lt058 (EyeLink Sensorimotoric Sys-tems Berlin Germany) Subjects were asked to keeptheir head still The experimenter also monitored gazeposition on-line At the beginning of each session (andafter breaks in testing or following any unintended headmovement that disrupted gaze recordings) eye-trackingwas re-calibrated Each trial was initiated by the exper-imenter and began with a central fixation cross for 1 secthen the search display
Subjects were instructed to search for Ts (among anyLs) Critically while looking at any target they had toclick a response button held in the right hand onlyif they considered this target to be a new discovery(ie a target they had not clicked on before) Clickingthe response button did not change the display Theresponse button was held by the patient in a comfort-able position to the right of the midline Subjectswere instructed and reminded throughout testing toclick on each target found once only Any later re-click on
a previously found target thus indicates a failure to retainthe fact that this target had already been discoveredThe experimenter terminated each search when thesubject indicated verbally that they felt all targets in thedisplay had now been found Each subject searched aminimum of 5 displays (mean = 7 range 5ndash14) Eyemovements were analyzed off-line using custom soft-ware An automatic algorithm was used to extract sac-cades (saccade onset defined as eye speed rising above258 sec1 for two or more samples and a criterion ofminimum fixation duration of 60 msec) Blinks wereeliminated from the trace by visual inspection and sac-cade and fixation delineation were checked visually inthe record Next fixations were automatically assignedto items in the display if the mean position fell within18 of that item Two or more consecutive fixations onthe same item were re-scored as a single extendedfixation A re-fixation was scored only when at leastone fixation elsewhere on another item intervened
Most click responses were made during a fixation andthe few button-clicks occurring during a saccade were
Table 1 Patient Details
CaseAge
(years) PathologyVolume(cm3)
Age ofLesion (days)
Visual FieldDefect
Mean Deviationfrom Center on
Line Bisection Task(cm) (rightward +)
Mesulum Score(Maximum Possible
Score = 60) Lesion Type
1 68 Infarct 1282 64 + 867 17 PCA
2 76 Infarct 1528 227 + 003 41 PCA
3 68 Infarct 79 207 + 297 36 PCA
4 77 Infarct 251 158 + 150 54 PCA
5 63 Infarct 28 29 033 55 MCA
6 59 Hemorrhage 172 7 + 32 6 MCA
7 66 Hemorrhage 273 325 + 213 31 MCA
8 68 Infarct 309 3 25 17 MCA
9 29 Infarct 115 17 113 55 MCA
10 70 Infarct 1339 324 153 57 MCA
11 42 Infarct ndash 11 097 24 MCA
12 75 Infarct 757 38 013 4 MCA
13 51 Infarct 694 141 083 31 MCA
14 59 Infarct 1144 356 + 223 37 MCA
15 80 Infarct 261 44 35 6 MCA
16 64 Infarct ndash 30 033 55 MCA
MEAN 63 66 124 188 33
SE 4 13 32 5
Cases 1ndash9 had damage to the nonfrontal (posterior only) cortex whereas Cases 10ndash15 had both frontal and posterior (posterior and anterior) corticalinvolvement Case 16 is a patient with a subcortical lesion All but 3 of the patients with right MCA infarction had intact visual fields on confrontationTwo of these (Cases 7 and 14) were diagnosed with left homonymous hemianopia whereas one (Case 6) had a left inferior quandrantic field defectAll patients with PCA territory damage had partial or complete contralesional visual field loss No patient suffered from dysphasia
352 Journal of Cognitive Neuroscience Volume 17 Number 2
assigned to the previous fixation Clicks were assigned toa particular item if eye position at the time of the clickwas within 28 of that item or in the cases where morethan one item was within 28 of current eye position theclick response was simply attributed to the item closestto that eye position
Button-clicks were assigned to a particular item if eyeposition at the time of the click was within 28 of thatitem A re-click on an item required at least one fixationon another item between successive clicks (in practiceseveral fixations elsewhere typically intervened betweeninitial click and later re-click see Results)
Statistical analyses were performed using SPSS andSigmaplot (SPSS Science Chicago USA) Jonckheere sta-tistic was derived by hand Statistical significance was setas p lt 05
Acknowledgments
We thank the reviewers of this manuscript and Dr AndrewParton for their extremely helpful comments We thank thepatients who participated Dr J Wade Dr A Rudd and thestaff of the stroke units at Charing Cross and St ThomasrsquoHospitals This research was supported by grants from theWellcome Trust to M H C K and J D J D holds a RoyalSociety Wolfson Research Merit Award
Reprint requests should be sent to Drs Sabira Mannan andMasud Husain Division of Neuroscience and PsychologicalMedicine Imperial College London Charing Cross CampusLondon W6 8RF UK or via e-mail smannanimperialacukor mhusainimperialacuk
Notes
1 We are grateful to one of the reviewers for suggesting thistime-series analysis2 There was no significant difference between the number oftargets fixated between the two groups so differences in lsquolsquoclickomissionsrsquorsquo cannot be attributed to the number of fixatedtargets that had to be kept track of during search3 The correlation between these four points alone inFigure 9A is high [r(3) = 81 cf r(15) = 47 for the wholegroup]4 We have also tested 10 neglect patients and 10 controlstroke patients on a vertical version of the Corsi blocks task(clinically used to measure spatial working memory capacity)Neglect patients had significantly impaired capacity but wereable to localize single locations well suggesting that mislocal-ization is unlikely to account for their deficit on this spatialworking memory task
REFERENCES
Bartolomeo P Sieroff E Decaix C amp Chokron S (2001)Modulating the attentional bias in unilateral neglect theeffects of the strategic set Experimental Brain Research137 432ndash444
Bates E Wilson S M Saygin A P Dick F Sereno M IKnight R T amp Dronkers N F (2003) Voxel-basedlesion-symptom mapping Nature Neuroscience 6 448ndash450
Behrmann M Watt S Black S E amp Barton J J (1997)Impaired visual search in patients with unilateral neglect
An oculographic analysis Neuropsychologia 351445ndash1458
Bisiach E amp Vallar G (2000) Unilateral neglect in humans InG Rizzolatti (Ed) Handbook of neuropsychology (2nd edvol 1 pp 459ndash502) Amsterdam Elsevier
Cals N Devuyst G Afsar N Karapanayiotides T ampBogousslavsky J (2002) Pure superficial posterior cerebralartery territory infarction in The Lausanne Stroke RegistryJournal of Neurology 249 855ndash861
Chafee M amp Goldman-Rakic P (1998) Matching patterns ofactivity in primate prefrontal area 8a and parietal area 7ipneurons during a spatial working memory task Journal ofNeurophysiology 79 2919ndash2940
Chedru F Leblanc M amp Lhermitte F (1973) Visualsearching in normal and brain-damaged subjectsContributions to the study of unilateral subjectsCortex 9 94ndash111
DrsquoEsposito M Aguirre G K Zarahn E Ballard DShin R K amp Lease J (1998) Functional MRI studies ofspatial and nonspatial working memory Cognitive BrainResearch 7 1ndash13
Donkelaar P amp Muri R (2002) Craniotopic updating ofvisual space across saccades in the human posteriorparietal cortex Proceedings of the Royal Society London B269 735ndash739
Driver J amp Husain M (2002) The role of spatial workingmemory deficits in pathological search by neglect patientsIn the cognitive and neural bases of spatial neglect InH-O Karnath A D Milner amp G Vallar (Eds) The cognitiveand neural bases of spatial neglect Oxford OxfordUniversity Press
Duhamel J R Colby C amp Goldberg M E (1992) Theupdating of the representation of visual space in parietalcortex by intended eye movements Science 25590ndash92
Goldman-Rakic P S (1996) The prefrontal landscapemdashImplications of functional architecture for understandinghuman mentation and the central executive PhilosophicalTransactions of The Royal Society of London Series BBiological Sciences 351 1445ndash1453
Heide W Binkofski F Seitz R J Posse S Nitschke M FFreund H J amp Kompf D (2001) Activation offrontoparietal cortices during memorized triple-stepsequences of saccadic eye movements An fMRI studyEuropean Journal of Neuroscience 13 1177ndash1189
Heide W amp Kompf D (1998) Combined deficits ofsaccades and visuo-spatial orientation after cortical lesionsExperimental Brain Research 123 164ndash171
Husain M amp Kennard C (1996) Visual neglect associatedwith frontal lobe infarction Journal of Neurology 243652ndash657
Husain M Mannan S Hodgson T Wojciulik E Driver Jamp Kennard C (2001) Impaired spatial working memoryacross saccades contributes to abnormal search in parietalneglect Brain 121 941ndash952
Husain M amp Rorden C (2003) Non-spatially lateralizedmechanisms in hemispatial neglect Nature ReviewsNeuroscience 4 23ndash36
Jackson S R amp Husain M (1996) Visuomotor functionsof the lateral pre-motor cortex Current Opinion inNeurobiology 6 788ndash795
Kinsbourne M (1993) Orientational bias model of unilateralneglect Evidence from attentional gradients withinhemispace In I H Robertson amp J C Marshall (Eds)Unilateral neglect Clinical and experimental studies(pp 63ndash86) Hove Erlbaum
Konishi S Kyoichi N Uchida I Kikyo H Kameyama Mamp Miyauchi S (1999) Common inhibitory mechanism in
Mannan et al 353
human inferior prefrontal cortex revealed by event-relatedfunctional MRI Brain 122 981ndash991
Konishi S Kyoichi N Uchida I Sekihara K amp Miyashita Y(1998) No-go dominant brain activity in human inferiorprefrontal cortex revealed by functional magneticresonance imaging European Journal of Neuroscience10 1209ndash1213
Malhotra P Mannan S Driver J amp Husain M (2004)Impaired spatial working memory One component of thevisual neglect syndrome Cortex 40 667ndash676
Manly T Woldt K Watson P amp Warburton E (2002) Ismotor perseveration in unilateral neglect lsquolsquodrivenrsquorsquo by thepresence of neglected left-sided stimuli Neuropsychologia40 1794ndash1803
Mazzoni P Bracewell R Barash S amp Andersen R (1996)Spatially tuned auditory responses in area LIP of macaquesperforming delayed memory saccades to acoustic targetsJournal of Neurophysiology 75 1233ndash1241
McCarley J Wang R Kramer A Irwin D amp Peterson M(2003) How much memory does oculomotor search havePsychological Science 14 422ndash426
Mesulam M-M (1985) Principles of behavioural neurologyTests of directed attention and memory Philadelphia Davis
Mesulam M M (1999) Spatial attention and neglectParietal frontal and cingulate contributions to the mentalrepresentation and attentional targeting of salientextrapersonal events Philosophical Transactions of theRoyal Society of London Series B Biological Sciences354 1325ndash1346
Mort D Malhotra P Mannan S Rorden C Pambakian AKennard C amp Husain M (2003) The anatomy of visualneglect Brain 126 1986ndash1997
Na D L Adair J C Kang Y Chung C S Lee K H ampHeilman K M (1999) Motor perseverative behaviour ona line cancellation task Neurology 52 1569ndash1576
Pearson D G amp Sahraie A (2003) Oculomotor controland the maintenance of spatially and temporally distributedevents in visuo-spatial working memory QuarterlyJournal of Experimental Psychology 56A 1089ndash1111
Rafal R (1994) Neglect Current Opinion in Neurobiology4 231ndash236
Rizzolatti G amp Arbib M A (1998) Language within our graspTrends in Neurosciences 21 188ndash194
Robertson L C Lamb M R amp Knight R T (1988) Effectsof lesions of temporalndashparietal junction on perceptual andattentional processing in humans Journal of Neuroscience8 3757ndash3769
Rorden C amp Brett M (2000) Stereotaxic display of brainlesions Behavioural Neurology 12 191ndash200
Rusconi M L Maravita A Bottini G amp Vallar G (2002)Is the intact side really intact Perseverative responses inpatients with unilateral neglect A productive manifestationNeuropsychologia 40 594ndash604
Russell C Malhotra P amp Husain M (2004) Attentionmodulates the visual field in healthy observers and parietalpatients NeuroReport 15 189ndash193
Rypma B amp DrsquoEsposito M (2003) A subsequent-memoryeffect in dorsolateral prefrontal cortex Cognitive BrainResearch 16 162ndash166
Sapir A Hayes A Henik A Danzinger S amp Rafal R(2004) Parietal lobe lesions disrupt saccadic remapping ofinhibitory location tagging Journal of CognitiveNeuroscience 16 503ndash509
Smania N Martini M C Gambina G A T Palamara ANatale E amp Marzi C A (1998) The spatial distributionof visual attention in hemineglect and extinction patientsBrain 121 1759ndash1770
Sprenger A Kompf D amp Heide W (2002) Visual searchin patients with left visual hemineglect Progress in BrainResearch 140 395ndash416
Tobler P Felblinger J Burki M Nirkko A Ozdoba C ampMuri R (2001) Functional organisation of the saccadicreference system processing extraretinal signals in humansVision Research 41 1351ndash1358
Wilson B Cockburn J amp Halligan P W (1987)Behavioural inattention test Bury St EdmundsThames Valley
Wojciulik E Husain M Clarke K amp Driver J (2001)Spatial working memory deficit in unilateral neglectNeuropsychologia 39 390ndash396
Zihl J amp Hebel N (1997) Patterns of oculomotor scanningin patients with unilateral posterior parietal and frontallobe damage Neuropsychologia 35 893ndash906
Zihl J amp von Cramon D (1985) Visual field recovery fromscotoma in patients with postgeniculate damage A reviewof 55 cases Brain 108 335ndash365
354 Journal of Cognitive Neuroscience Volume 17 Number 2
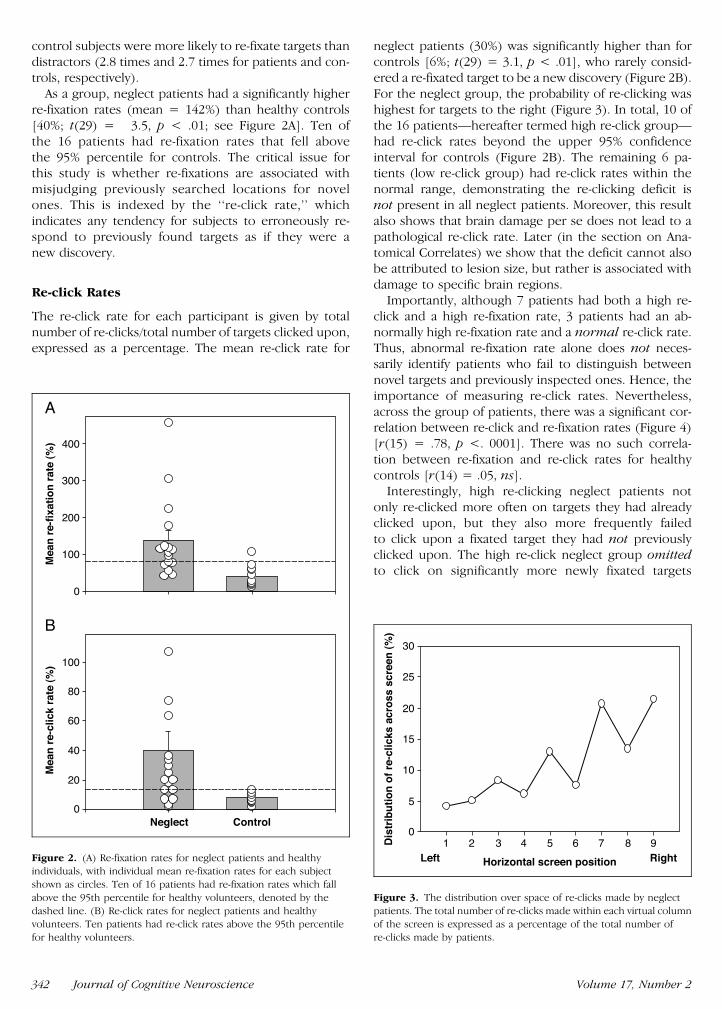
control subjects were more likely to re-fixate targets thandistractors (28 times and 27 times for patients and con-trols respectively)
As a group neglect patients had a significantly higherre-fixation rates (mean = 142) than healthy controls[40 t(29) = 35 p lt 01 see Figure 2A] Ten ofthe 16 patients had re-fixation rates that fell abovethe 95 percentile for controls The critical issue forthis study is whether re-fixations are associated withmisjudging previously searched locations for novelones This is indexed by the lsquolsquore-click ratersquorsquo whichindicates any tendency for subjects to erroneously re-spond to previously found targets as if they were anew discovery
Re-click Rates
The re-click rate for each participant is given by totalnumber of re-clickstotal number of targets clicked uponexpressed as a percentage The mean re-click rate for
neglect patients (30) was significantly higher than forcontrols [6 t(29) = 31 p lt 01] who rarely consid-ered a re-fixated target to be a new discovery (Figure 2B)For the neglect group the probability of re-clicking washighest for targets to the right (Figure 3) In total 10 ofthe 16 patientsmdashhereafter termed high re-click groupmdashhad re-click rates beyond the upper 95 confidenceinterval for controls (Figure 2B) The remaining 6 pa-tients (low re-click group) had re-click rates within thenormal range demonstrating the re-clicking deficit isnot present in all neglect patients Moreover this resultalso shows that brain damage per se does not lead to apathological re-click rate Later (in the section on Ana-tomical Correlates) we show that the deficit cannot alsobe attributed to lesion size but rather is associated withdamage to specific brain regions
Importantly although 7 patients had both a high re-click and a high re-fixation rate 3 patients had an ab-normally high re-fixation rate and a normal re-click rateThus abnormal re-fixation rate alone does not neces-sarily identify patients who fail to distinguish betweennovel targets and previously inspected ones Hence theimportance of measuring re-click rates Neverthelessacross the group of patients there was a significant cor-relation between re-click and re-fixation rates (Figure 4)[r(15) = 78 p lt 0001] There was no such correla-tion between re-fixation and re-click rates for healthycontrols [r(14) = 05 ns]
Interestingly high re-clicking neglect patients notonly re-clicked more often on targets they had alreadyclicked upon but they also more frequently failedto click upon a fixated target they had not previouslyclicked upon The high re-click neglect group omittedto click on significantly more newly fixated targets
Figure 3 The distribution over space of re-clicks made by neglect
patients The total number of re-clicks made within each virtual column
of the screen is expressed as a percentage of the total number ofre-clicks made by patients
Figure 2 (A) Re-fixation rates for neglect patients and healthyindividuals with individual mean re-fixation rates for each subject
shown as circles Ten of 16 patients had re-fixation rates which fall
above the 95th percentile for healthy volunteers denoted by the
dashed line (B) Re-click rates for neglect patients and healthyvolunteers Ten patients had re-click rates above the 95th percentile
for healthy volunteers
342 Journal of Cognitive Neuroscience Volume 17 Number 2
than the low re-click neglect group [mean = 30omissions SE 4 vs 19 and 3 t(14) = 231 p lt05]2 Such an association between these lsquolsquoclick omis-sionsrsquorsquo and high re-click rate would not be expected ifre-clicking was entirely due to a lsquolsquoperseverativersquorsquo failureto inhibit responses to rightward targets because lsquolsquoclickomissionsrsquorsquo represent failures to respond Nor wouldthey be explained by items on the right being per-ceived to be more salient and thus automatically cap-turing responses
Anatomical Correlates of High Re-click Rates
Although 10 neglect patients demonstrated significant-ly elevated re-click rates 6 did not To determinewhether key anatomical locations underlie the distinc-tion between these groups we sought to isolate brainregions that were damaged in neglect patients withhigh re-click rates and equally importantly spared inpatients without the deficit Critically there was nocorrelation between lesion volume and re-click rate[r(13) = 31 ns] or between lesion age and re-clickrate [r(13) = 30 ns] Each patient was assigned toone of two subgroups according to whether they hadfrontal involvement or not Neglect patients withlsquolsquoposterior onlyrsquorsquo lesions had damage posterior to thecentral sulcus with no damage anterior to the pre-central sulcus (n = 9) Neglect patients with lsquolsquoposte-rior and anteriorrsquorsquo involvement had cortical damageanterior to the central sulcus (n = 5) all patients inthis subgroup also had some posterior damage
Parietal Correlate of High Re-click Rate
Among the 9 patients in the lsquolsquoposterior onlyrsquorsquo subgroup4 had an abnormally high re-click rate 3 with MCA ter-
ritory damage (Cases 5 6 and 8) and only 1 with PCAdamage (Figure 5A) This single PCA patient (Case 2)had damage to white matter deep to the occipito-temporo-parietal junction in contrast to the other 3PCA patients whose lesions did not extend so faranteriorly or dorsally The area of damage commonto all 3 MCA patients with abnormally high re-clickratesmdashwhile also being intact in the 5 posterior sub-group patients with normal re-click ratesmdashwas locatednear the anterior end of the intraparietal sulcus closeto the lateral bank (Figure 5C Talairach 46 40 28for centroid volume 058 cm3) This area was not sim-ply the most common region of overlap for all 8 MCApatients that was located more dorsally (Talairach37 55 38)
In order to assign statistical significance to the areaof overlap we performed a voxel-by-voxel analysis (Bateset al 2003) For every voxel that was damaged in atleast 3 subjects we compared the re-click rate of pa-tients with damage to that voxel with those of patientswithout damage to that particular voxel producing at statistic for a given voxel The area shown in yellow(Figure 5C) yields a value of t(7) = 491 indicatingsignificantly higher re-click rates associated with dam-age to this region ( p lt 002) This difference survivesBonferroni correction for multiple comparisons ( p lt05) where the number of comparisons was 6 (therewere only 6 regions in which the lesions of 3 or morepatients overlapped)
An alternative analytical approach is to focus only onpatients with relatively small lesions Seven patients(Cases 4ndash9 and 15) had focal parietal lesions lt31 cm3all had posterior lesions except Case 15 who also hadinvolvement anterior to the central sulcus The maxi-mum area of overlap common to all 4 patients with highre-click rates (Cases 5 6 8 15) which also cruciallyremained undamaged in the 3 patients with normal re-click rates (Cases 4 7 9) again centered on the lateralintraparietal sulcus (Figure 6) slightly more dorsally(Talairach 37 39 37 volume 011 cm3) and involvinga subset of the critical voxels identified in Figure 5 Notethat this overlap region will probably also involve whitematter fibers including those that may connect the pa-rietal cortex to the frontal lobe
Frontal Correlate of High Re-click Rate
Four of the five patients in the lsquolsquoposterior plus anteriorrsquorsquoneglect subgroup had an abnormally high re-click rateThe area of damage common to all four was in theinferior frontal lobe (Talairach 38 18 16 for centroid178 cm3) in a region corresponding to Brodmannrsquosarea 44 part of the premotor cortex (Figure 7A) Threeof the four posterior plus anterior neglect patients witha pathologically high re-click rate also had damage ex-tending into or bordering the ventrolateral prefrontalcortex The remaining patient who had the highest re-
Figure 4 Mean re-click rates plotted against mean re-fixation rates
for individual neglect patients There was a significant correlation[r(15) = 78 p lt 0001]
Mannan et al 343
click rate of all patients (Case 15) had damage involvingthe dorsolateral prefrontal cortex Only 1 of the 5 pos-terior plus anterior patients had a normal re-click rate(Figure 7B) Her lesion was posterior to the region ofoverlap seen in the four frontal patients with abnor-mally high re-click rates It involved the posterior premo-tor cortex and motor cortex and extended caudally intothe inferior parietal lobe Thus the subtraction of herlesion from the remaining posterior plus anterior patientswith high re-click rates (Figure 7C) does not alter theirmaximum overlap region (displayed in Figure 7A)
Anatomical Correlates of the lsquolsquoRe-fixationrsquorsquo Deficit
Ten neglect patients had a high re-fixation rate but 6patients did not There was no correlation betweenlesion volume and re-fixation rate [r(13) = 22 ns]or between age of lesion and re-click rate [r(13) =36 ns] Although some patients in either the poste-rior (Cases 1 2 4ndash8) or frontal group (Cases 11 13 15)had high re-fixation rates other patients with highlysimilar lesion locations had normal re-fixation ratesAs a consequence voxel-by-voxel analysis (Bates et al
Figure 5 The
neuroanatomical correlates of
the deficit in retaining spatiallocations for neglect patients
with posterior only (nonfrontal)
lesions (n = 9) (A) Individuallesions of patients with high
re-click rate (patients with MCA
damage Cases 8 5 6 patient
with PCA damage Case 2) andoverlap map for these
4 patients where maximum
overlap occurs for a small region
(shown in green) around theright anterior intraparietal
sulcus damaged in the 3 MCA
patients (B) Individual lesionsof patients with normal re-click
rate (patients with MCA
damage Cases 9 and 7 patients
with PCA damage Cases 1 3 4)and overlap map for these
5 patients with maximum
overlap occurring in the
occipital cortex Importantlynone of the 5 patients with
normal re-click rates have
damage to the common areaof overlap for patients with high
re-click rates shown in A
(C) Contrast map highlighting
intraparietal locus which isdamaged in 3 out of 4 patients
with a high re-click rate and
intact in the 5 patients with
normal re-click rates (shown inyellow) Regions that are
damaged in patients with high
re-click rates and spared in
those patients with normalre-click rates are shown in red to
yellow whereas regions in blue
denote areas that are commonto patients with
normal re-click rates but
spared in patients with high
re-click rates
344 Journal of Cognitive Neuroscience Volume 17 Number 2
2003) revealed no anatomical specificity associated withhigh re-fixation rates [t(7) = 085 ns] unlike thatassociated with high re-click rates even when onlyfocal lesions were considered [t(5) = 165 ns]
Time Interval between Clicking and Re-clicking ona Target
We next examined the probability of lsquolsquore-clickingrsquorsquo on atarget when re-fixating it as a function of the time lag ornumber of intervening saccades between the first clickon that target and the subsequent revisit An increasedprobability of re-clicking with increasing delay (or inter-vening number of saccades) would be consistent a failureto keep track of spatial locations during search whereasa flat or decreasing function might arise from impairedresponse inhibition (lsquolsquoperseverativersquorsquo behavior) or even
a failure to encode target location This time-series anal-ysis for the 10 patients with high re-click rates revealedtwo distinct patterns of behavior Half the patients de-monstrated a significantly increasing probability of mis-judging an already found item to be a novel one withincreasing delay (Figure 8A) or intervening saccades(Figure 8C) whereas the other 5 patients had a constantor even decreasing probability (Figure 8B and D)
Importantly most (45) patients in the group withincreasing re-click functions had damage to the focalparietal location previously identified in the analysisof patients with focal lesions and high re-click rates(Figure 6C) The other patient was Case 2 who had alarge PCA lesion extending to the white matter deep tothe occipito-temporo-parietal junction By contrast noneof the patients with flatdecreasing functions had dam-age to the key parietal region Instead the latter group
Figure 6 The
neuroanatomical correlates of
the deficit in retaining spatial
locations for neglect patientswith focal lesions (n = 7)
(A) Individual lesions of
patients with high re-click rate
(all 4 patients have MCAdamage) and overlap map for
these 4 patients where
maximum overlap occurs for asmall region around the right
intraparietal sulcus (shown in
red) damaged all 4 MCA
patients (B) Individual lesionsof patients with normal re-click
rate (all patients with PCA
damage) and overlap map for
these 3 patients with maximumoverlap occurring in the
occipital cortex Importantly
none of these 3 patients withnormal re-click rates have
damage to the common area of
overlap for patients with high
re-click rates shown in A(C) Contrast map highlighting
intraparietal locus which is
damaged in 4 out of 4 patients
with a high re-click rate andintact in the 3 patients with
normal re-click rates (shown in
yellow) Regions that are
damaged in patients with highre-click rates and spared in
those patients with normal
re-click rates are shown in redto yellow whereas regions in
blue denote areas that are
common to patients with
normal re-click rates butspared in patients with high
re-click rates
Mannan et al 345
had frontal cortical or subcortical involvement Thusincreasing re-clicking with time interval was associatedwith parietal damage whereas flat re-click functionswere associated with frontalsubcortical lesions
For neglect patients as a whole the intervals betweenan initial click on a target and a subsequent re-click werelong (mean = 40 saccades) but much shorter betweenan initial fixation and a later re-fixation (mean = 5) withthis difference being significant [t(15) = 315 p lt01] This again emphasizes the dissociation betweenre-fixations and re-clicks and also demonstrates that re-clicking does not occur on most re-fixations
Relationship of Re-click Rate to Neglect Severity
We assessed three separate measures of neglect Thefirst was the difference between miss rates for left ver-sus right targets within the experimental search taskitself the other two measures of neglect were based onthe clinical pen-and-paper measures of Mesulam shape-cancellation (Mesulam 1985) and line bisection fromthe BIT (Wilson Cockburn amp Halligan 1987)
On the experimental search task there was a moderatebut significant correlation between re-click rate and ne-glect in search across the group [r(15) = 47 p lt 05
Figure 9A] Thus the greater the re-clicking on right-ward items the greater the severity of leftward neglectThere was also a moderate but again significant corre-lation between re-click rates and the severity of individualpatientsrsquo clinical neglect as indexed by the cancellationmeasure [r(15) = 46 p lt 05 Figure 9B] Thesecorrelations were driven largely by three of the patientsidentified in our focal lesion analysis (Figure 6A) withinvolvement of the key parietal region and high re-clicking behavior Their data are shown in filled circlesto the right in the graphs displayed in Figure 93 Thecorrelation between deviation on line bisection andre-click rate was not significant [r(15) = 13 ns] How-ever one would not predict that a deficit in retaininglocations already visited (or response inhibition) wouldnecessarily impair performance on a task such as linebisection
Effect of Varying Number of Targets
In 7 patients (Cases 5 8 10 11 14ndash16) we also exam-ined the effect of varying the number of targets in thesearch task Patients viewed lsquolsquotarget onlyrsquorsquo displays con-taining either 2 6 or 19 targets randomly distributedon the screen without any L-shaped distractors In-
Figure 7 Theneuroanatomical correlates of
the re-click deficit in neglect
patients with posterior plusanterior lesions (n = 5)
(A) Individual lesions of
patients with high re-click rate
(all 4 patients have MCAdamage) and overlap map for
these 4 patients where
maximum overlap occurs for
a region in the right inferiorfrontal lobe (shown in red)
damaged all 4 MCA patients
(B) Individual lesions of a single
patient with normal re-clickrate (C) Contrast map
highlighting the inferior frontal
locus (shown in yellow) whichis damaged in 4 out of
4 patients with a high re-click
rate and intact in the patient
with normal re-click rateRegions that are damaged in
patients with high re-click rates
and spared in the patient
with normal re-click rate areshown in red to yellow
whereas regions in blue denote
areas that are damaged in thepatient with normal re-click
rates but spared in patients with
high re-click rates
346 Journal of Cognitive Neuroscience Volume 17 Number 2
creasing the number of targets led to significantly higherre-click rates ( Jonckheere statistic = 133 p lt 01Figure 10) as well as more severe neglect (Jonckheerestatistic = 129 p lt 01) consistent with the proposalthat the mechanisms underlying pathological re-clickingmight contribute to neglect behavior Note that a lsquolsquoper-severativersquorsquo failure of response inhibition would not onits own easily explain this finding For the same absolutenumber of click responses one would predict a higherre-click rate with smaller set-sizes than with larger ones(see also Husain Mannan et al 2001 for a discussion)Although the number of subjects were small there wasno difference in the performance pattern between the3 patients who had increasing re-click rate functions inthe time-series analysis and the 4 patients with flat ordecreasing re-click rate functions
Varying Perceptual Load during Search
Can re-click behavior be explained simply by impairedvisual perception of targets from discriminators Neglectpatients did find it more difficult than control subjectsto select targets from distractors fixating on average19 distractors for every target found [cf 14 for controlsubjects t(29) = 485 p lt 0001] Patients also some-times clicked on distractors (64) but no more often
than controls (61) and it is possible that this rate iseffectively inflated by our strict criterion of considering aclick to be a response on the item that is being fixated atthe time of the click (It is likely that sometimes ob-servers click in response to targets they fixated one ormore saccades previously) Re-clicks on distractors wereextremely rare (06 cf 30 for targets)
To examine the effect of reduced perceptual load in8 patients (Cases 5 7 8ndash10 13 1 4 16) 6 of whom werehigh re-clickers we used search displays with salientcross (+) targets rather than Ts L distractors wereunchanged Patients fixated significantly fewer distrac-tors when searching for cross (+) targets [distractortarget ratio of 134 cf 181 for T targets t(7) = 60 p lt001] suggesting they did indeed find targetndashdistractordiscrimination easier Importantly however there wasno significant difference in re-click rate [26 for +targets cf 27 for T targets t(14) = 041 ns] or evenin re-fixation rate [100 for + targets cf 119 fort(14) = 041 ns] Thus these changes in perceptualload did not influence re-click performance
DISCUSSION
On this multitarget visual search task neglect patientsmissed targets to their left and also re-fixated targets
Figure 8 The probability of
re-clicking or re-fixating on
target as a function of delay
between the original click andsubsequent re-fixation (for
patients with high re-click
rates) (A) Probability functions
for patients with significantpositive slopes Four patients of
these patients have damage to
the focal intraparietal locusidentified in Figure 6C whereas
the single patient with a PCA
lesion and no damage to the
critical parietal locationindicated by symbol has only
a very shallow positive slope
(B) Probability functions for
patients with no significantpositive slope None of these
5 patients have damage to the
critical intraparietal region (C)As for A but for delay measured
in terms of intervening
fixations (D) As for B but for
delay measured in terms ofintervening fixations
Mannan et al 347
to the right much more often than healthy individuals(Figure 2A) Critically 10 of the 16 neglect patients re-clicked on rightward targets at a significantly higher ratethan healthy control subjects (Figure 2B) For 5 of thesepatients the probability of re-clicking increased withtime (Figure 8A) whereas the other 5 neglect patientswith high re-click rates demonstrated a constant or de-creasing probability of re-clicking over time (Figure 8C)The remaining 6 neglect patients had re-click rateswithin the normal range demonstrating that neitherneglect nor brain damage per se leads to a pathologicalre-click rate
High-resolution MRI mapping of lesions revealed im-portant anatomical dissociations within the neglect pop-ulation Patients with pathological re-click rates hadlesions affecting either the right intraparietal sulcus orthe inferior frontal lobe the only exception (Case 2) hada large occipito-temporal lesion that extended deep intothe parietal white matter Neglect patients with normalre-click rates did not have damage to either of thecritical locations in the intraparietal sulcus or inferiorfrontal lobe (Figures 5 6 and 7) Instead they had moredorsal lesions (for MCA territory patients) or occipitaland medial-temporal damage (for PCA territory lesions)which can also result in neglect (Mort et al 2003Cals Devuyst Afsar Karapanayiotides amp Bogousslavsky2002) Importantly lesion location rather than the sizeor age of lesion was critical to the presence of the re-clicking impairment Across the group of neglect pa-tients there was a moderate but significant correlationbetween re-click rate and degree of neglect (Figure 9)driven largely by patients with parietal damage who hadboth a high re-click rate and also severe neglect consis-tent with the proposal that mechanisms underlying thisbehavior in parietal patients might have a functionallyimportant role in determining the severity of neglect
Role of Right Intraparietal Sulcus
Notably for all patients with damage to the critical locusin the right intraparietal sulcus (which may also involveparieto-frontal white matter fibers) the probability of re-clicking on a target increased with time (or interveningsaccades) since initial discovery (Figure 8) Because theyrarely re-clicked at short intervalsmdasheven though themajority of re-fixations occurred during this periodmdashthis pattern of behavior suggests that for these patientstarget locations were actually veridically perceived and
Figure 9 (A) Neglect on experimental search task (19 targets and44 distractors) plotted against mean re-click rate for individual neglect
patients Percentage neglect was calculated as the difference between
the number of targets missed on the left versus right expressed as apercentage of the total number of targets missed There was a
significant correlation between the degree of neglect and the re-click
rate [r(15) = 465 p lt 05] Patients with high re-click rates are
highlighted by black circles (for patients with damage to the focalparietal location identified in Figure 6C) and light gray circles (for
patients with frontal damage) while the single patient with PCA
damage and a high re-click rate is indicated by dark gray (B) The
number of targets missed on the Mesulam shape-cancellation taskplotted against mean re-click rate for individual neglect patients Again
there was a significant correlation between the number of targets
missed and the mean re-click rate [r(15) = 456 p lt 05] Patients withhigh re-click rates are denoted as in A
Figure 10 Mean re-click rate (filled circles) and degree of neglect
(unfilled squares) increased systematically with increasing numbers of
targets in target-only displays
348 Journal of Cognitive Neuroscience Volume 17 Number 2
well encoded into working memory Furthermore de-tailed single-case studies of two parietal patients withhigh re-click rates (Malhotra Mannan Driver amp Husain2004 Husain Mannan et al 2001) demonstrate usingdifferent psychophysical methods that these patientscan accurately encode spatial locations4 Increasing re-clicks with time (or intervening saccades) would how-ever be consistent with a deficit in keeping track oftarget locations during extended search
Several lines of evidence support the proposal thatthe critical intraparietal sulcus region we have identifiedmay have an important role in the spatial transforma-tions required to keep track of spatial locations acrosssaccades In rhesus monkeys neurons within the lateralbank of the intraparietal sulcus (area LIP) encode andlsquolsquomaintainrsquorsquo the remembered locations of saccadic tar-gets over time (Chafee amp Goldman-Rakic 1998 MazzoniBracewell Barash amp Andersen 1996) Some visuallyresponsive neurons in the LIP also appear to shift theirretinotopic representation at the onset of an intendedsaccade as if lsquolsquore-mappingrsquorsquo representations of objects inspace to update them across shifts of gaze (DuhamelColby amp Goldberg 1992)
In humans this computation has been investigatedusing the double-step saccade paradigm where twoflashes of light appear in quick succession such thatthe second target disappears before a saccade can bemade to the first target Patients with lesions involvingthe right parietal cortex are impaired at performing thistask suggesting that they are unable to update spatialrepresentations accurately after even one saccade (Heideamp Kompf 1998 Duhamel et al 1992) Similarly trans-cranial magnetic stimulation of the posterior parietalcortex prior to the second saccade reduces its accuracy(Donkelaar amp Muri 2002) and a recent study of patientswith parietal lesions also implicates this area in thespatial updating of inhibition of return after a saccade(Sapir Hayes Henik Danzinger amp Rafal 2004) Func-tional imaging studies of double-step (Tobler et al2001) and triple-step memory-guided saccades (HeideBinkofski et al 2001) have identified a region withinthe intraparietal sulcus (Talairach coordinates 40 4044 for Tobler et al and 44 48 36 Heide et al) This isclose to the critical region of lesion overlap we haveidentified in the posterior neglect patients with high re-click rates (Talairach 37 39 37)
We suggest that in our patients with lesions involvingthe right intraparietal sulcus uncertainty about locationsof previously found targets increases with each saccadeThis uncertainty is further exacerbated by interferencefrom newer target locations leading to increased prob-ability of misjudging old target locations for new assearch progresses Such progressively worsening uncer-tainty might be due to an accumulating impairment inlsquolsquore-mapping target locationsrsquorsquo across saccades or im-paired lsquolsquomemoryrsquorsquo of previously inspected target loca-tions across saccades As noted above there is evidence
for both types of signalmdash re-mapping and maintainingsaccadic spatial memorymdashin the parietal cortex and it ispossible that disruption to both these functions contrib-utes to the re-click deficit we have identified Certainlythe two potential types of deficit are not mutuallyincompatible
A slightly different account of the re-clicking behaviormight be framed in terms of the intriguing hypothesisproposed by Manly Woldt Watson and Warburton(2002) to explain visible re-marking on a standard can-cellation task They argue that such behavior is due topatients erroneously transposing leftward stimuli to theright resulting in apparent re-cancellation These au-thors found that removing targets from the left re-duced both neglect and re-marking However they didnot monitor eye movements so it is possible that sometargets that are logged as neglected (because they werenot cancelled) were nevertheless fixated and thereforeform part of the load on memory during search Fur-thermore in our experimental study subjects with ahigh re-click rate also failed to click on a higher pro-portion of fixated targets Manly et alrsquos proposal wouldnot easily account for such lsquolsquoclick omissionsrsquorsquo becausethese refer to fixated targets that were not actuallyresponded to
Finally it might be argued that increasing re-clickingwith search duration reflects a deficit in spatial workingmemory (SWM) that is independent of saccadic eyemovements (ie a decay in memory that is not trans-saccadic) Because time and number of interveningsaccades were not independently manipulated in thisunrestricted visual search paradigm we are not ablecategorically to distinguish between these possibilitiesWe note however that functional imaging studies ofSWM tasks that do not require eye movements oftendemonstrate activity related to the maintenance ofspatial information which is generally more posteriorand dorsal than the parietal locations isolated in thisstudy (DrsquoEsposito et al 1998) Moreover recent behav-ioral data indicate that SWM is more vulnerable tosaccadic eye movements than shifts in attention or limbmovements (Pearson amp Sahraie 2003)
Frontal Lesion Site Associated withHigh Re-click Rates
Four out of 5 neglect patients with lesions involvingposterior plus anterior damage also showed a high re-click rate and the area of damage common to thesepatients centered on the right inferior frontal lobe cor-responding to Brodmannrsquos area 44 (Figure 7C) Thisregion has previously been associated with frontalneglect (Husain amp Kennard 1996) and is consideredto be part of the ventral premotor cortex and thehomologue of Brocarsquos areas in the left hemisphere(Rizzolatti amp Arbib 1998 Jackson amp Husain 1996) Inthese frontal patients the probability of re-clicking did
Mannan et al 349
not increase with time interval between the first clickand re-fixation but rather it stayed constant or declinedwith increasing delays
Such behavior might be consistent with a failure toinhibit responses to rightward locationsmdasha type oflsquolsquoperseverativersquorsquo behavior Note though that this is notnecessarily the same type of perseverative responseas seen when some patients with neglect re-markpreviously found items on standard cancellation tests(Rusconi et al 2002 Na et al 1999) In those casestargets that are visibly cancelled are often marked againwhereas in our computerized search tasks no marks arevisibly left on targets once they have been clicked uponMoreover only one of our neglect patients (Case 1) de-monstrated re-cancellation of visibly cancelled targetson pen-and-paper tasks and then only on one occasionImportantly this patient actually had a normal re-clickrate (67) on our task and his stroke was in the PCAterritory sparing both the intraparietal sulcus and fron-tal lobe
In support of a role for the right frontal lobe in in-hibiting responses to previously found targets recentimaging studies of response inhibition have highlightedregions of the right inferior frontal gyrus (IFG) very closeto the centroid of damage for our frontal patients (38 1618 cf 43 22 21 Konishi Kyoichi Uchida Sekihara ampMiyashita 1998 and 41 16 19 Konishi Kyoichi UchidaKikyo et al 1999) Alternative explanations for the highprobability of re-clicking directly after clicking might bethat these patients have impaired perceptual discrimina-tion of targets impaired perceptual localization of targetsor that they encounter difficulty in encoding spatiallocations into working memory Our data are not sugges-tive of a perceptual discrimination problem as changingthe perceptual load did not inf luence re-click ratesMoreover the patients still had high re-click rates in thetarget only conditions (Figure 10) where no perceptualdiscrimination of targets and distractors was necessaryPoor perceptual localization would seem unlikely giventhat these patients have frontal lesions Furthermoreusing a vertical SWM task similar to the Corsi blocktapping task (Malhotra et al 2004) we have found thatpatients with neglect can localize and perceptually en-code single target locations as well as controls and aresignificantly impaired at remembering two or more loca-tions shown in sequence consistent with the presentobservations Some recent functional imaging data sug-gest that activation within the prefrontal cortex corre-lates well with successful encoding of material intoworking memory (Rypma amp DrsquoEsposito 2003) Furtherresearch is required to distinguish between these pos-sible explanations for the mechanisms underlying there-click deficit in frontal patients
The only patient with frontal damage who did re-clickmore frequently with increasing time had damage cen-tered on the dorsolateral prefrontal cortex (althoughalso encroaching on the IFG) Neurons in the dorsolat-
eral prefrontal cortex like those in the LIP encode theremembered locations of saccadic targets (Goldman-Rakic 1996) This patient (Case 15) who had the high-est re-click rate of all also had damage to the criticalparietal location described above (in contrast to all otherfrontal patients) Thus she may have suffered from acombination of underlying deficits
Re-fixations and Misjudging PreviouslyInspected Locations during Search
In this search task re-fixation and re-click rates weresignificantly correlated for neglect patients Howeverthere were important dissociations within individuals 2patients with PCA lesions had re-fixation rates of 220and 80 respectively but both had a re-click rate ofonly 7 These are clearly examples of individuals whodo not show pathological re-clicking despite having highre-fixation rates High re-fixation rates may result froma lack of integration of spatial information due to V1damage (Zihl amp von Cramon 1985) in our PCA patientsor possibly because of reduced inhibition of return(Bartolomeo Sieroff Decaix amp Chokron 2001) What-ever the exact cause for dissociations between re-fixa-tions and re-click rates our data suggest re-fixation doesnot necessarily index the same underlying deficits asrevealed by our re-click measure High re-fixation ratesunlike high re-click rates were also not significantlyassociated with damage to any specific cortical locationagain demonstrating a dissociation between re-fixationsand pathological re-clicking
The relationship between re-click rates and neglect isunlikely to be a simple one In our study there was amoderate but significant correlation between the twomeasures suggesting that this association is unlikely tohave arisen by chance However non-neglect patientscan also exhibit a high re-click rate (unpublished obser-vations from our laboratory) Taken together our resultssuggest that difficulties in keeping track of previouslyinspected targets (because of impaired trans-saccadic re-mapping andor memory) may be one contributingcomponent to visual neglect in patients with lesions in-volving the right intraparietal sulcus As mentioned thesearch task we used differed in one striking aspect fromstandard cancellation tests subjects do not visibly markthe targets they visit Whereas in cancellation tests pa-tients mark each target they find which might appear todispense with any need to remember which locationshave been searched before However it should be ap-preciated that in most standard cancellation tests theindividual marks made by patients may not be clearlyvisible in peripheral vision particularly if neglect pa-tients have a local spatial bias (Rafal 1994 RobertsonLamb amp Knight 1988) or a restricted field of view(Russell Malhotra amp Husain 2004) Thus when com-bined with a rightward attentional bias keeping track ofsearched locations may be critical to prevent gaze from
350 Journal of Cognitive Neuroscience Volume 17 Number 2
returning to rightward locations even if they have beenmarked before as the marks may not be visible untilgaze returns to them Consistent with this hypothesisneglect is greater when cancellation is performed usinginvisible marks compared to large and salient red marksthat can be seen peripherally (Wojciulik Husain Clarkeamp Driver 2001)
A task like the present search-and-click paradigm maymore closely resemble search in the real world than dotraditional cancellation tests When we examine a visualscene we do not leave marks on each object we ex-amine Instead just as in the search task used in thisstudy we have to keep track of the locations we visit ifwe are to avoid inefficient re-examination of thoselocations Neglect patients with damage to the rightintraparietal sulcus appear to be impaired at keepingtrack of searched locations in this way When combinedwith their rightward attentional bias we propose thatsuch an impairment leads to recursive search of stimulito the right in experimental search tasks as used hereand also in daily life where neglect is so apparentdespite unlimited viewing time
The paradigm we have used has the important virtuesof allowing patients to explore freelymdashas in real-worldsearch or clinical cancellation tasksmdashand relating theirre-click behavior directly to the severity of neglect in thesame task But it might be argued that it would be usefulalso to probe SWM in a more controlled manner by re-stricting observersrsquo eye movements to a specific set oflocations and probing memory of previously visitedplaces A recent study using a gaze-contingent para-digm has elegantly probed the capacity of spatial mem-ory during search in healthy observers and foundevidence for memory for up to four previous locations(McCarley Wang Kramer Irwin amp Peterson 2003)Variants of this paradigm may in the future be usefulto probe the capacity of SWM across saccades in patientswith neglect and also to probe further whether thereare differences between neglect patients with parietaland frontal lesions
METHODS
Subjects Imaging and Lesion Mapping
Sixteen patients with left neglect following right hemi-sphere infarction or hemorrhage were recruited Con-sent was obtained according to the Declaration ofHelsinki as approved by the hospital ethical commit-tees Presence of neglect was determined by clinicalobservation plus bedside tests including the Mesulamshape-cancellation task (Mesulam 1985) and line bisec-tion task (Wilson et al 1987) (see Table 1) Twelvepatients had MCA territory strokes the rest involvedonly the PCA territory which can also result in neglect(Cals et al 2002) Only one neglect patient (Case 1 whodid not have frontal involvement) demonstrated re-
cancellation of visibly cancelled targets on pen-and-paper tasks and then only on one occasion Fifteenage-matched healthy volunteers were controls (meanage = 63 SD = 14 vs 63 SD = 13 for patients)
All patients had clinical CT scans In addition dedi-cated high-resolution structural MRI scans were alsoobtained for 1416 neglect patients MRI scans wereunobtainable for the other two (Cases 11 and 16)because of claustrophobia or loss to follow up In theMCA neglect group median time between stroke andscan was 63 days with the shortest interval being35 days for the PCA neglect group one patient had tobe scanned relatively early at 9 days but the mediantime was 140 days MRI scans comprised one 256-sliceT1-weighted acquisition (MPRAGE TR 97 msec TE4 msec TA 12 min) performed in the coronal planealigned perpendicular to the anteriorndashposterior commis-sural line on a Siemens Vision 15-T scanner Within-and between-slice resolution was 1 mm For each pa-tient the area of damage was determined by detailedvisual inspection of the digital image for every singleslice Lesion delineation was performed by neurologists(authors D M and M H) by tracing by hand theboundary of the lesion onto every single 1-mm-thickaxial slice using a graphics tablet (WACOM Intuos A6)Combining all the slices then led to a 3-D region of in-terest (ROI) for each patient For one patient (Case 15)there were two areas of discrete focal damage one inthe frontal cortex and the other in the parietal cortex
Normalization of each MRI including the 3-D ROI tothe T1-template was performed with SPM99 (wwwfilionuclacukspm) The ROI was used as a mask for thelesion during determination of alignment parameters tominimize the contribution of abnormal brain to thisprocess (Rorden amp Brett 2000) The normalized ROIof each neglect patient was superimposed upon the T1template to find areas of maximal lesion overlap in eachgroup (see Results) as well as lesion volume usingMRIcro shareware (Rorden amp Brett 2000)
Experimental Procedure
Stimuli were presented on an SVGA monitor (29 22 in)viewed at 57 cm distance Each search display com-prised a virtual 9 7 grid each cell containing a single18 elementmdasheither a letter T (target) or a letter L (dis-tractor) (see Figure 1) The position of each elementwithin each cell was jittered with 19 targets and 44distractors randomly distributed over the virtual gridThe number of targets on the right and left halves ofthe display was balanced across trials Seven neglectpatients (Cases 5 8 10 11 14ndash16) were further testedon lsquolsquoTarget onlyrsquorsquo displays which had varying numbersof Ts (2 6 or 19) now with no L-shaped distractorsThe presentations of these lsquolsquoTarget onlyrsquorsquo screens wereinterspersed with those of the original search screens inrandom order in the same experimental session
Mannan et al 351
Eye movements were recorded using a video-basedpupil tracker with a temporal resolution of 4 msec andspatial accuracy of lt058 (EyeLink Sensorimotoric Sys-tems Berlin Germany) Subjects were asked to keeptheir head still The experimenter also monitored gazeposition on-line At the beginning of each session (andafter breaks in testing or following any unintended headmovement that disrupted gaze recordings) eye-trackingwas re-calibrated Each trial was initiated by the exper-imenter and began with a central fixation cross for 1 secthen the search display
Subjects were instructed to search for Ts (among anyLs) Critically while looking at any target they had toclick a response button held in the right hand onlyif they considered this target to be a new discovery(ie a target they had not clicked on before) Clickingthe response button did not change the display Theresponse button was held by the patient in a comfort-able position to the right of the midline Subjectswere instructed and reminded throughout testing toclick on each target found once only Any later re-click on
a previously found target thus indicates a failure to retainthe fact that this target had already been discoveredThe experimenter terminated each search when thesubject indicated verbally that they felt all targets in thedisplay had now been found Each subject searched aminimum of 5 displays (mean = 7 range 5ndash14) Eyemovements were analyzed off-line using custom soft-ware An automatic algorithm was used to extract sac-cades (saccade onset defined as eye speed rising above258 sec1 for two or more samples and a criterion ofminimum fixation duration of 60 msec) Blinks wereeliminated from the trace by visual inspection and sac-cade and fixation delineation were checked visually inthe record Next fixations were automatically assignedto items in the display if the mean position fell within18 of that item Two or more consecutive fixations onthe same item were re-scored as a single extendedfixation A re-fixation was scored only when at leastone fixation elsewhere on another item intervened
Most click responses were made during a fixation andthe few button-clicks occurring during a saccade were
Table 1 Patient Details
CaseAge
(years) PathologyVolume(cm3)
Age ofLesion (days)
Visual FieldDefect
Mean Deviationfrom Center on
Line Bisection Task(cm) (rightward +)
Mesulum Score(Maximum Possible
Score = 60) Lesion Type
1 68 Infarct 1282 64 + 867 17 PCA
2 76 Infarct 1528 227 + 003 41 PCA
3 68 Infarct 79 207 + 297 36 PCA
4 77 Infarct 251 158 + 150 54 PCA
5 63 Infarct 28 29 033 55 MCA
6 59 Hemorrhage 172 7 + 32 6 MCA
7 66 Hemorrhage 273 325 + 213 31 MCA
8 68 Infarct 309 3 25 17 MCA
9 29 Infarct 115 17 113 55 MCA
10 70 Infarct 1339 324 153 57 MCA
11 42 Infarct ndash 11 097 24 MCA
12 75 Infarct 757 38 013 4 MCA
13 51 Infarct 694 141 083 31 MCA
14 59 Infarct 1144 356 + 223 37 MCA
15 80 Infarct 261 44 35 6 MCA
16 64 Infarct ndash 30 033 55 MCA
MEAN 63 66 124 188 33
SE 4 13 32 5
Cases 1ndash9 had damage to the nonfrontal (posterior only) cortex whereas Cases 10ndash15 had both frontal and posterior (posterior and anterior) corticalinvolvement Case 16 is a patient with a subcortical lesion All but 3 of the patients with right MCA infarction had intact visual fields on confrontationTwo of these (Cases 7 and 14) were diagnosed with left homonymous hemianopia whereas one (Case 6) had a left inferior quandrantic field defectAll patients with PCA territory damage had partial or complete contralesional visual field loss No patient suffered from dysphasia
352 Journal of Cognitive Neuroscience Volume 17 Number 2
assigned to the previous fixation Clicks were assigned toa particular item if eye position at the time of the clickwas within 28 of that item or in the cases where morethan one item was within 28 of current eye position theclick response was simply attributed to the item closestto that eye position
Button-clicks were assigned to a particular item if eyeposition at the time of the click was within 28 of thatitem A re-click on an item required at least one fixationon another item between successive clicks (in practiceseveral fixations elsewhere typically intervened betweeninitial click and later re-click see Results)
Statistical analyses were performed using SPSS andSigmaplot (SPSS Science Chicago USA) Jonckheere sta-tistic was derived by hand Statistical significance was setas p lt 05
Acknowledgments
We thank the reviewers of this manuscript and Dr AndrewParton for their extremely helpful comments We thank thepatients who participated Dr J Wade Dr A Rudd and thestaff of the stroke units at Charing Cross and St ThomasrsquoHospitals This research was supported by grants from theWellcome Trust to M H C K and J D J D holds a RoyalSociety Wolfson Research Merit Award
Reprint requests should be sent to Drs Sabira Mannan andMasud Husain Division of Neuroscience and PsychologicalMedicine Imperial College London Charing Cross CampusLondon W6 8RF UK or via e-mail smannanimperialacukor mhusainimperialacuk
Notes
1 We are grateful to one of the reviewers for suggesting thistime-series analysis2 There was no significant difference between the number oftargets fixated between the two groups so differences in lsquolsquoclickomissionsrsquorsquo cannot be attributed to the number of fixatedtargets that had to be kept track of during search3 The correlation between these four points alone inFigure 9A is high [r(3) = 81 cf r(15) = 47 for the wholegroup]4 We have also tested 10 neglect patients and 10 controlstroke patients on a vertical version of the Corsi blocks task(clinically used to measure spatial working memory capacity)Neglect patients had significantly impaired capacity but wereable to localize single locations well suggesting that mislocal-ization is unlikely to account for their deficit on this spatialworking memory task
REFERENCES
Bartolomeo P Sieroff E Decaix C amp Chokron S (2001)Modulating the attentional bias in unilateral neglect theeffects of the strategic set Experimental Brain Research137 432ndash444
Bates E Wilson S M Saygin A P Dick F Sereno M IKnight R T amp Dronkers N F (2003) Voxel-basedlesion-symptom mapping Nature Neuroscience 6 448ndash450
Behrmann M Watt S Black S E amp Barton J J (1997)Impaired visual search in patients with unilateral neglect
An oculographic analysis Neuropsychologia 351445ndash1458
Bisiach E amp Vallar G (2000) Unilateral neglect in humans InG Rizzolatti (Ed) Handbook of neuropsychology (2nd edvol 1 pp 459ndash502) Amsterdam Elsevier
Cals N Devuyst G Afsar N Karapanayiotides T ampBogousslavsky J (2002) Pure superficial posterior cerebralartery territory infarction in The Lausanne Stroke RegistryJournal of Neurology 249 855ndash861
Chafee M amp Goldman-Rakic P (1998) Matching patterns ofactivity in primate prefrontal area 8a and parietal area 7ipneurons during a spatial working memory task Journal ofNeurophysiology 79 2919ndash2940
Chedru F Leblanc M amp Lhermitte F (1973) Visualsearching in normal and brain-damaged subjectsContributions to the study of unilateral subjectsCortex 9 94ndash111
DrsquoEsposito M Aguirre G K Zarahn E Ballard DShin R K amp Lease J (1998) Functional MRI studies ofspatial and nonspatial working memory Cognitive BrainResearch 7 1ndash13
Donkelaar P amp Muri R (2002) Craniotopic updating ofvisual space across saccades in the human posteriorparietal cortex Proceedings of the Royal Society London B269 735ndash739
Driver J amp Husain M (2002) The role of spatial workingmemory deficits in pathological search by neglect patientsIn the cognitive and neural bases of spatial neglect InH-O Karnath A D Milner amp G Vallar (Eds) The cognitiveand neural bases of spatial neglect Oxford OxfordUniversity Press
Duhamel J R Colby C amp Goldberg M E (1992) Theupdating of the representation of visual space in parietalcortex by intended eye movements Science 25590ndash92
Goldman-Rakic P S (1996) The prefrontal landscapemdashImplications of functional architecture for understandinghuman mentation and the central executive PhilosophicalTransactions of The Royal Society of London Series BBiological Sciences 351 1445ndash1453
Heide W Binkofski F Seitz R J Posse S Nitschke M FFreund H J amp Kompf D (2001) Activation offrontoparietal cortices during memorized triple-stepsequences of saccadic eye movements An fMRI studyEuropean Journal of Neuroscience 13 1177ndash1189
Heide W amp Kompf D (1998) Combined deficits ofsaccades and visuo-spatial orientation after cortical lesionsExperimental Brain Research 123 164ndash171
Husain M amp Kennard C (1996) Visual neglect associatedwith frontal lobe infarction Journal of Neurology 243652ndash657
Husain M Mannan S Hodgson T Wojciulik E Driver Jamp Kennard C (2001) Impaired spatial working memoryacross saccades contributes to abnormal search in parietalneglect Brain 121 941ndash952
Husain M amp Rorden C (2003) Non-spatially lateralizedmechanisms in hemispatial neglect Nature ReviewsNeuroscience 4 23ndash36
Jackson S R amp Husain M (1996) Visuomotor functionsof the lateral pre-motor cortex Current Opinion inNeurobiology 6 788ndash795
Kinsbourne M (1993) Orientational bias model of unilateralneglect Evidence from attentional gradients withinhemispace In I H Robertson amp J C Marshall (Eds)Unilateral neglect Clinical and experimental studies(pp 63ndash86) Hove Erlbaum
Konishi S Kyoichi N Uchida I Kikyo H Kameyama Mamp Miyauchi S (1999) Common inhibitory mechanism in
Mannan et al 353
human inferior prefrontal cortex revealed by event-relatedfunctional MRI Brain 122 981ndash991
Konishi S Kyoichi N Uchida I Sekihara K amp Miyashita Y(1998) No-go dominant brain activity in human inferiorprefrontal cortex revealed by functional magneticresonance imaging European Journal of Neuroscience10 1209ndash1213
Malhotra P Mannan S Driver J amp Husain M (2004)Impaired spatial working memory One component of thevisual neglect syndrome Cortex 40 667ndash676
Manly T Woldt K Watson P amp Warburton E (2002) Ismotor perseveration in unilateral neglect lsquolsquodrivenrsquorsquo by thepresence of neglected left-sided stimuli Neuropsychologia40 1794ndash1803
Mazzoni P Bracewell R Barash S amp Andersen R (1996)Spatially tuned auditory responses in area LIP of macaquesperforming delayed memory saccades to acoustic targetsJournal of Neurophysiology 75 1233ndash1241
McCarley J Wang R Kramer A Irwin D amp Peterson M(2003) How much memory does oculomotor search havePsychological Science 14 422ndash426
Mesulam M-M (1985) Principles of behavioural neurologyTests of directed attention and memory Philadelphia Davis
Mesulam M M (1999) Spatial attention and neglectParietal frontal and cingulate contributions to the mentalrepresentation and attentional targeting of salientextrapersonal events Philosophical Transactions of theRoyal Society of London Series B Biological Sciences354 1325ndash1346
Mort D Malhotra P Mannan S Rorden C Pambakian AKennard C amp Husain M (2003) The anatomy of visualneglect Brain 126 1986ndash1997
Na D L Adair J C Kang Y Chung C S Lee K H ampHeilman K M (1999) Motor perseverative behaviour ona line cancellation task Neurology 52 1569ndash1576
Pearson D G amp Sahraie A (2003) Oculomotor controland the maintenance of spatially and temporally distributedevents in visuo-spatial working memory QuarterlyJournal of Experimental Psychology 56A 1089ndash1111
Rafal R (1994) Neglect Current Opinion in Neurobiology4 231ndash236
Rizzolatti G amp Arbib M A (1998) Language within our graspTrends in Neurosciences 21 188ndash194
Robertson L C Lamb M R amp Knight R T (1988) Effectsof lesions of temporalndashparietal junction on perceptual andattentional processing in humans Journal of Neuroscience8 3757ndash3769
Rorden C amp Brett M (2000) Stereotaxic display of brainlesions Behavioural Neurology 12 191ndash200
Rusconi M L Maravita A Bottini G amp Vallar G (2002)Is the intact side really intact Perseverative responses inpatients with unilateral neglect A productive manifestationNeuropsychologia 40 594ndash604
Russell C Malhotra P amp Husain M (2004) Attentionmodulates the visual field in healthy observers and parietalpatients NeuroReport 15 189ndash193
Rypma B amp DrsquoEsposito M (2003) A subsequent-memoryeffect in dorsolateral prefrontal cortex Cognitive BrainResearch 16 162ndash166
Sapir A Hayes A Henik A Danzinger S amp Rafal R(2004) Parietal lobe lesions disrupt saccadic remapping ofinhibitory location tagging Journal of CognitiveNeuroscience 16 503ndash509
Smania N Martini M C Gambina G A T Palamara ANatale E amp Marzi C A (1998) The spatial distributionof visual attention in hemineglect and extinction patientsBrain 121 1759ndash1770
Sprenger A Kompf D amp Heide W (2002) Visual searchin patients with left visual hemineglect Progress in BrainResearch 140 395ndash416
Tobler P Felblinger J Burki M Nirkko A Ozdoba C ampMuri R (2001) Functional organisation of the saccadicreference system processing extraretinal signals in humansVision Research 41 1351ndash1358
Wilson B Cockburn J amp Halligan P W (1987)Behavioural inattention test Bury St EdmundsThames Valley
Wojciulik E Husain M Clarke K amp Driver J (2001)Spatial working memory deficit in unilateral neglectNeuropsychologia 39 390ndash396
Zihl J amp Hebel N (1997) Patterns of oculomotor scanningin patients with unilateral posterior parietal and frontallobe damage Neuropsychologia 35 893ndash906
Zihl J amp von Cramon D (1985) Visual field recovery fromscotoma in patients with postgeniculate damage A reviewof 55 cases Brain 108 335ndash365
354 Journal of Cognitive Neuroscience Volume 17 Number 2
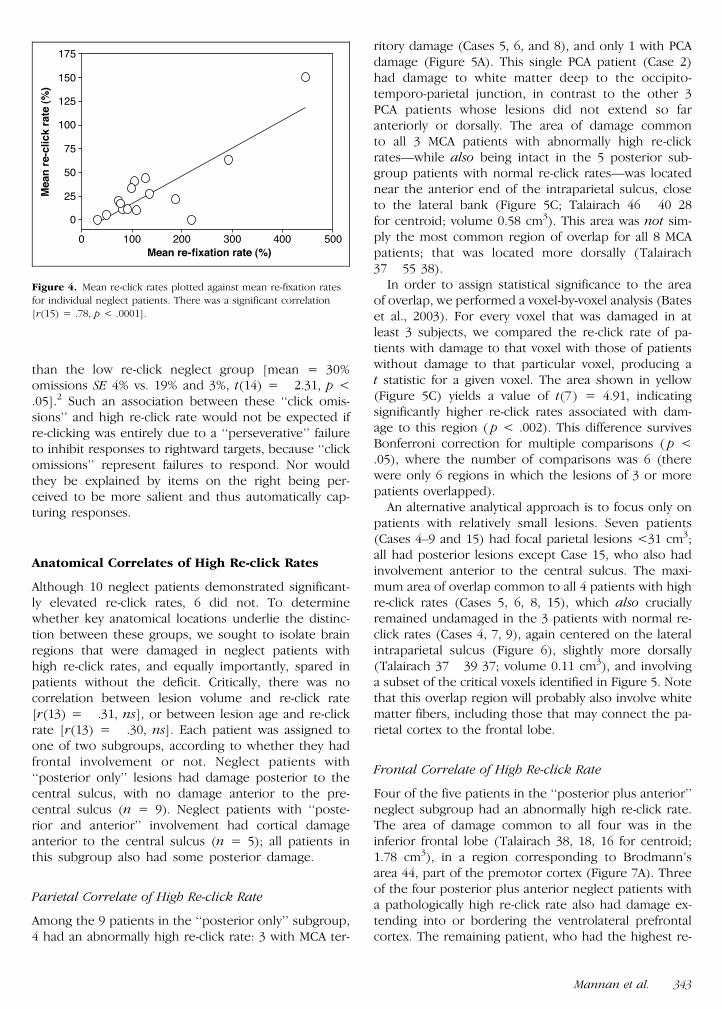
than the low re-click neglect group [mean = 30omissions SE 4 vs 19 and 3 t(14) = 231 p lt05]2 Such an association between these lsquolsquoclick omis-sionsrsquorsquo and high re-click rate would not be expected ifre-clicking was entirely due to a lsquolsquoperseverativersquorsquo failureto inhibit responses to rightward targets because lsquolsquoclickomissionsrsquorsquo represent failures to respond Nor wouldthey be explained by items on the right being per-ceived to be more salient and thus automatically cap-turing responses
Anatomical Correlates of High Re-click Rates
Although 10 neglect patients demonstrated significant-ly elevated re-click rates 6 did not To determinewhether key anatomical locations underlie the distinc-tion between these groups we sought to isolate brainregions that were damaged in neglect patients withhigh re-click rates and equally importantly spared inpatients without the deficit Critically there was nocorrelation between lesion volume and re-click rate[r(13) = 31 ns] or between lesion age and re-clickrate [r(13) = 30 ns] Each patient was assigned toone of two subgroups according to whether they hadfrontal involvement or not Neglect patients withlsquolsquoposterior onlyrsquorsquo lesions had damage posterior to thecentral sulcus with no damage anterior to the pre-central sulcus (n = 9) Neglect patients with lsquolsquoposte-rior and anteriorrsquorsquo involvement had cortical damageanterior to the central sulcus (n = 5) all patients inthis subgroup also had some posterior damage
Parietal Correlate of High Re-click Rate
Among the 9 patients in the lsquolsquoposterior onlyrsquorsquo subgroup4 had an abnormally high re-click rate 3 with MCA ter-
ritory damage (Cases 5 6 and 8) and only 1 with PCAdamage (Figure 5A) This single PCA patient (Case 2)had damage to white matter deep to the occipito-temporo-parietal junction in contrast to the other 3PCA patients whose lesions did not extend so faranteriorly or dorsally The area of damage commonto all 3 MCA patients with abnormally high re-clickratesmdashwhile also being intact in the 5 posterior sub-group patients with normal re-click ratesmdashwas locatednear the anterior end of the intraparietal sulcus closeto the lateral bank (Figure 5C Talairach 46 40 28for centroid volume 058 cm3) This area was not sim-ply the most common region of overlap for all 8 MCApatients that was located more dorsally (Talairach37 55 38)
In order to assign statistical significance to the areaof overlap we performed a voxel-by-voxel analysis (Bateset al 2003) For every voxel that was damaged in atleast 3 subjects we compared the re-click rate of pa-tients with damage to that voxel with those of patientswithout damage to that particular voxel producing at statistic for a given voxel The area shown in yellow(Figure 5C) yields a value of t(7) = 491 indicatingsignificantly higher re-click rates associated with dam-age to this region ( p lt 002) This difference survivesBonferroni correction for multiple comparisons ( p lt05) where the number of comparisons was 6 (therewere only 6 regions in which the lesions of 3 or morepatients overlapped)
An alternative analytical approach is to focus only onpatients with relatively small lesions Seven patients(Cases 4ndash9 and 15) had focal parietal lesions lt31 cm3all had posterior lesions except Case 15 who also hadinvolvement anterior to the central sulcus The maxi-mum area of overlap common to all 4 patients with highre-click rates (Cases 5 6 8 15) which also cruciallyremained undamaged in the 3 patients with normal re-click rates (Cases 4 7 9) again centered on the lateralintraparietal sulcus (Figure 6) slightly more dorsally(Talairach 37 39 37 volume 011 cm3) and involvinga subset of the critical voxels identified in Figure 5 Notethat this overlap region will probably also involve whitematter fibers including those that may connect the pa-rietal cortex to the frontal lobe
Frontal Correlate of High Re-click Rate
Four of the five patients in the lsquolsquoposterior plus anteriorrsquorsquoneglect subgroup had an abnormally high re-click rateThe area of damage common to all four was in theinferior frontal lobe (Talairach 38 18 16 for centroid178 cm3) in a region corresponding to Brodmannrsquosarea 44 part of the premotor cortex (Figure 7A) Threeof the four posterior plus anterior neglect patients witha pathologically high re-click rate also had damage ex-tending into or bordering the ventrolateral prefrontalcortex The remaining patient who had the highest re-
Figure 4 Mean re-click rates plotted against mean re-fixation rates
for individual neglect patients There was a significant correlation[r(15) = 78 p lt 0001]
Mannan et al 343
click rate of all patients (Case 15) had damage involvingthe dorsolateral prefrontal cortex Only 1 of the 5 pos-terior plus anterior patients had a normal re-click rate(Figure 7B) Her lesion was posterior to the region ofoverlap seen in the four frontal patients with abnor-mally high re-click rates It involved the posterior premo-tor cortex and motor cortex and extended caudally intothe inferior parietal lobe Thus the subtraction of herlesion from the remaining posterior plus anterior patientswith high re-click rates (Figure 7C) does not alter theirmaximum overlap region (displayed in Figure 7A)
Anatomical Correlates of the lsquolsquoRe-fixationrsquorsquo Deficit
Ten neglect patients had a high re-fixation rate but 6patients did not There was no correlation betweenlesion volume and re-fixation rate [r(13) = 22 ns]or between age of lesion and re-click rate [r(13) =36 ns] Although some patients in either the poste-rior (Cases 1 2 4ndash8) or frontal group (Cases 11 13 15)had high re-fixation rates other patients with highlysimilar lesion locations had normal re-fixation ratesAs a consequence voxel-by-voxel analysis (Bates et al
Figure 5 The
neuroanatomical correlates of
the deficit in retaining spatiallocations for neglect patients
with posterior only (nonfrontal)
lesions (n = 9) (A) Individuallesions of patients with high
re-click rate (patients with MCA
damage Cases 8 5 6 patient
with PCA damage Case 2) andoverlap map for these
4 patients where maximum
overlap occurs for a small region
(shown in green) around theright anterior intraparietal
sulcus damaged in the 3 MCA
patients (B) Individual lesionsof patients with normal re-click
rate (patients with MCA
damage Cases 9 and 7 patients
with PCA damage Cases 1 3 4)and overlap map for these
5 patients with maximum
overlap occurring in the
occipital cortex Importantlynone of the 5 patients with
normal re-click rates have
damage to the common areaof overlap for patients with high
re-click rates shown in A
(C) Contrast map highlighting
intraparietal locus which isdamaged in 3 out of 4 patients
with a high re-click rate and
intact in the 5 patients with
normal re-click rates (shown inyellow) Regions that are
damaged in patients with high
re-click rates and spared in
those patients with normalre-click rates are shown in red to
yellow whereas regions in blue
denote areas that are commonto patients with
normal re-click rates but
spared in patients with high
re-click rates
344 Journal of Cognitive Neuroscience Volume 17 Number 2
2003) revealed no anatomical specificity associated withhigh re-fixation rates [t(7) = 085 ns] unlike thatassociated with high re-click rates even when onlyfocal lesions were considered [t(5) = 165 ns]
Time Interval between Clicking and Re-clicking ona Target
We next examined the probability of lsquolsquore-clickingrsquorsquo on atarget when re-fixating it as a function of the time lag ornumber of intervening saccades between the first clickon that target and the subsequent revisit An increasedprobability of re-clicking with increasing delay (or inter-vening number of saccades) would be consistent a failureto keep track of spatial locations during search whereasa flat or decreasing function might arise from impairedresponse inhibition (lsquolsquoperseverativersquorsquo behavior) or even
a failure to encode target location This time-series anal-ysis for the 10 patients with high re-click rates revealedtwo distinct patterns of behavior Half the patients de-monstrated a significantly increasing probability of mis-judging an already found item to be a novel one withincreasing delay (Figure 8A) or intervening saccades(Figure 8C) whereas the other 5 patients had a constantor even decreasing probability (Figure 8B and D)
Importantly most (45) patients in the group withincreasing re-click functions had damage to the focalparietal location previously identified in the analysisof patients with focal lesions and high re-click rates(Figure 6C) The other patient was Case 2 who had alarge PCA lesion extending to the white matter deep tothe occipito-temporo-parietal junction By contrast noneof the patients with flatdecreasing functions had dam-age to the key parietal region Instead the latter group
Figure 6 The
neuroanatomical correlates of
the deficit in retaining spatial
locations for neglect patientswith focal lesions (n = 7)
(A) Individual lesions of
patients with high re-click rate
(all 4 patients have MCAdamage) and overlap map for
these 4 patients where
maximum overlap occurs for asmall region around the right
intraparietal sulcus (shown in
red) damaged all 4 MCA
patients (B) Individual lesionsof patients with normal re-click
rate (all patients with PCA
damage) and overlap map for
these 3 patients with maximumoverlap occurring in the
occipital cortex Importantly
none of these 3 patients withnormal re-click rates have
damage to the common area of
overlap for patients with high
re-click rates shown in A(C) Contrast map highlighting
intraparietal locus which is
damaged in 4 out of 4 patients
with a high re-click rate andintact in the 3 patients with
normal re-click rates (shown in
yellow) Regions that are
damaged in patients with highre-click rates and spared in
those patients with normal
re-click rates are shown in redto yellow whereas regions in
blue denote areas that are
common to patients with
normal re-click rates butspared in patients with high
re-click rates
Mannan et al 345
had frontal cortical or subcortical involvement Thusincreasing re-clicking with time interval was associatedwith parietal damage whereas flat re-click functionswere associated with frontalsubcortical lesions
For neglect patients as a whole the intervals betweenan initial click on a target and a subsequent re-click werelong (mean = 40 saccades) but much shorter betweenan initial fixation and a later re-fixation (mean = 5) withthis difference being significant [t(15) = 315 p lt01] This again emphasizes the dissociation betweenre-fixations and re-clicks and also demonstrates that re-clicking does not occur on most re-fixations
Relationship of Re-click Rate to Neglect Severity
We assessed three separate measures of neglect Thefirst was the difference between miss rates for left ver-sus right targets within the experimental search taskitself the other two measures of neglect were based onthe clinical pen-and-paper measures of Mesulam shape-cancellation (Mesulam 1985) and line bisection fromthe BIT (Wilson Cockburn amp Halligan 1987)
On the experimental search task there was a moderatebut significant correlation between re-click rate and ne-glect in search across the group [r(15) = 47 p lt 05
Figure 9A] Thus the greater the re-clicking on right-ward items the greater the severity of leftward neglectThere was also a moderate but again significant corre-lation between re-click rates and the severity of individualpatientsrsquo clinical neglect as indexed by the cancellationmeasure [r(15) = 46 p lt 05 Figure 9B] Thesecorrelations were driven largely by three of the patientsidentified in our focal lesion analysis (Figure 6A) withinvolvement of the key parietal region and high re-clicking behavior Their data are shown in filled circlesto the right in the graphs displayed in Figure 93 Thecorrelation between deviation on line bisection andre-click rate was not significant [r(15) = 13 ns] How-ever one would not predict that a deficit in retaininglocations already visited (or response inhibition) wouldnecessarily impair performance on a task such as linebisection
Effect of Varying Number of Targets
In 7 patients (Cases 5 8 10 11 14ndash16) we also exam-ined the effect of varying the number of targets in thesearch task Patients viewed lsquolsquotarget onlyrsquorsquo displays con-taining either 2 6 or 19 targets randomly distributedon the screen without any L-shaped distractors In-
Figure 7 Theneuroanatomical correlates of
the re-click deficit in neglect
patients with posterior plusanterior lesions (n = 5)
(A) Individual lesions of
patients with high re-click rate
(all 4 patients have MCAdamage) and overlap map for
these 4 patients where
maximum overlap occurs for
a region in the right inferiorfrontal lobe (shown in red)
damaged all 4 MCA patients
(B) Individual lesions of a single
patient with normal re-clickrate (C) Contrast map
highlighting the inferior frontal
locus (shown in yellow) whichis damaged in 4 out of
4 patients with a high re-click
rate and intact in the patient
with normal re-click rateRegions that are damaged in
patients with high re-click rates
and spared in the patient
with normal re-click rate areshown in red to yellow
whereas regions in blue denote
areas that are damaged in thepatient with normal re-click
rates but spared in patients with
high re-click rates
346 Journal of Cognitive Neuroscience Volume 17 Number 2
creasing the number of targets led to significantly higherre-click rates ( Jonckheere statistic = 133 p lt 01Figure 10) as well as more severe neglect (Jonckheerestatistic = 129 p lt 01) consistent with the proposalthat the mechanisms underlying pathological re-clickingmight contribute to neglect behavior Note that a lsquolsquoper-severativersquorsquo failure of response inhibition would not onits own easily explain this finding For the same absolutenumber of click responses one would predict a higherre-click rate with smaller set-sizes than with larger ones(see also Husain Mannan et al 2001 for a discussion)Although the number of subjects were small there wasno difference in the performance pattern between the3 patients who had increasing re-click rate functions inthe time-series analysis and the 4 patients with flat ordecreasing re-click rate functions
Varying Perceptual Load during Search
Can re-click behavior be explained simply by impairedvisual perception of targets from discriminators Neglectpatients did find it more difficult than control subjectsto select targets from distractors fixating on average19 distractors for every target found [cf 14 for controlsubjects t(29) = 485 p lt 0001] Patients also some-times clicked on distractors (64) but no more often
than controls (61) and it is possible that this rate iseffectively inflated by our strict criterion of considering aclick to be a response on the item that is being fixated atthe time of the click (It is likely that sometimes ob-servers click in response to targets they fixated one ormore saccades previously) Re-clicks on distractors wereextremely rare (06 cf 30 for targets)
To examine the effect of reduced perceptual load in8 patients (Cases 5 7 8ndash10 13 1 4 16) 6 of whom werehigh re-clickers we used search displays with salientcross (+) targets rather than Ts L distractors wereunchanged Patients fixated significantly fewer distrac-tors when searching for cross (+) targets [distractortarget ratio of 134 cf 181 for T targets t(7) = 60 p lt001] suggesting they did indeed find targetndashdistractordiscrimination easier Importantly however there wasno significant difference in re-click rate [26 for +targets cf 27 for T targets t(14) = 041 ns] or evenin re-fixation rate [100 for + targets cf 119 fort(14) = 041 ns] Thus these changes in perceptualload did not influence re-click performance
DISCUSSION
On this multitarget visual search task neglect patientsmissed targets to their left and also re-fixated targets
Figure 8 The probability of
re-clicking or re-fixating on
target as a function of delay
between the original click andsubsequent re-fixation (for
patients with high re-click
rates) (A) Probability functions
for patients with significantpositive slopes Four patients of
these patients have damage to
the focal intraparietal locusidentified in Figure 6C whereas
the single patient with a PCA
lesion and no damage to the
critical parietal locationindicated by symbol has only
a very shallow positive slope
(B) Probability functions for
patients with no significantpositive slope None of these
5 patients have damage to the
critical intraparietal region (C)As for A but for delay measured
in terms of intervening
fixations (D) As for B but for
delay measured in terms ofintervening fixations
Mannan et al 347
to the right much more often than healthy individuals(Figure 2A) Critically 10 of the 16 neglect patients re-clicked on rightward targets at a significantly higher ratethan healthy control subjects (Figure 2B) For 5 of thesepatients the probability of re-clicking increased withtime (Figure 8A) whereas the other 5 neglect patientswith high re-click rates demonstrated a constant or de-creasing probability of re-clicking over time (Figure 8C)The remaining 6 neglect patients had re-click rateswithin the normal range demonstrating that neitherneglect nor brain damage per se leads to a pathologicalre-click rate
High-resolution MRI mapping of lesions revealed im-portant anatomical dissociations within the neglect pop-ulation Patients with pathological re-click rates hadlesions affecting either the right intraparietal sulcus orthe inferior frontal lobe the only exception (Case 2) hada large occipito-temporal lesion that extended deep intothe parietal white matter Neglect patients with normalre-click rates did not have damage to either of thecritical locations in the intraparietal sulcus or inferiorfrontal lobe (Figures 5 6 and 7) Instead they had moredorsal lesions (for MCA territory patients) or occipitaland medial-temporal damage (for PCA territory lesions)which can also result in neglect (Mort et al 2003Cals Devuyst Afsar Karapanayiotides amp Bogousslavsky2002) Importantly lesion location rather than the sizeor age of lesion was critical to the presence of the re-clicking impairment Across the group of neglect pa-tients there was a moderate but significant correlationbetween re-click rate and degree of neglect (Figure 9)driven largely by patients with parietal damage who hadboth a high re-click rate and also severe neglect consis-tent with the proposal that mechanisms underlying thisbehavior in parietal patients might have a functionallyimportant role in determining the severity of neglect
Role of Right Intraparietal Sulcus
Notably for all patients with damage to the critical locusin the right intraparietal sulcus (which may also involveparieto-frontal white matter fibers) the probability of re-clicking on a target increased with time (or interveningsaccades) since initial discovery (Figure 8) Because theyrarely re-clicked at short intervalsmdasheven though themajority of re-fixations occurred during this periodmdashthis pattern of behavior suggests that for these patientstarget locations were actually veridically perceived and
Figure 9 (A) Neglect on experimental search task (19 targets and44 distractors) plotted against mean re-click rate for individual neglect
patients Percentage neglect was calculated as the difference between
the number of targets missed on the left versus right expressed as apercentage of the total number of targets missed There was a
significant correlation between the degree of neglect and the re-click
rate [r(15) = 465 p lt 05] Patients with high re-click rates are
highlighted by black circles (for patients with damage to the focalparietal location identified in Figure 6C) and light gray circles (for
patients with frontal damage) while the single patient with PCA
damage and a high re-click rate is indicated by dark gray (B) The
number of targets missed on the Mesulam shape-cancellation taskplotted against mean re-click rate for individual neglect patients Again
there was a significant correlation between the number of targets
missed and the mean re-click rate [r(15) = 456 p lt 05] Patients withhigh re-click rates are denoted as in A
Figure 10 Mean re-click rate (filled circles) and degree of neglect
(unfilled squares) increased systematically with increasing numbers of
targets in target-only displays
348 Journal of Cognitive Neuroscience Volume 17 Number 2
well encoded into working memory Furthermore de-tailed single-case studies of two parietal patients withhigh re-click rates (Malhotra Mannan Driver amp Husain2004 Husain Mannan et al 2001) demonstrate usingdifferent psychophysical methods that these patientscan accurately encode spatial locations4 Increasing re-clicks with time (or intervening saccades) would how-ever be consistent with a deficit in keeping track oftarget locations during extended search
Several lines of evidence support the proposal thatthe critical intraparietal sulcus region we have identifiedmay have an important role in the spatial transforma-tions required to keep track of spatial locations acrosssaccades In rhesus monkeys neurons within the lateralbank of the intraparietal sulcus (area LIP) encode andlsquolsquomaintainrsquorsquo the remembered locations of saccadic tar-gets over time (Chafee amp Goldman-Rakic 1998 MazzoniBracewell Barash amp Andersen 1996) Some visuallyresponsive neurons in the LIP also appear to shift theirretinotopic representation at the onset of an intendedsaccade as if lsquolsquore-mappingrsquorsquo representations of objects inspace to update them across shifts of gaze (DuhamelColby amp Goldberg 1992)
In humans this computation has been investigatedusing the double-step saccade paradigm where twoflashes of light appear in quick succession such thatthe second target disappears before a saccade can bemade to the first target Patients with lesions involvingthe right parietal cortex are impaired at performing thistask suggesting that they are unable to update spatialrepresentations accurately after even one saccade (Heideamp Kompf 1998 Duhamel et al 1992) Similarly trans-cranial magnetic stimulation of the posterior parietalcortex prior to the second saccade reduces its accuracy(Donkelaar amp Muri 2002) and a recent study of patientswith parietal lesions also implicates this area in thespatial updating of inhibition of return after a saccade(Sapir Hayes Henik Danzinger amp Rafal 2004) Func-tional imaging studies of double-step (Tobler et al2001) and triple-step memory-guided saccades (HeideBinkofski et al 2001) have identified a region withinthe intraparietal sulcus (Talairach coordinates 40 4044 for Tobler et al and 44 48 36 Heide et al) This isclose to the critical region of lesion overlap we haveidentified in the posterior neglect patients with high re-click rates (Talairach 37 39 37)
We suggest that in our patients with lesions involvingthe right intraparietal sulcus uncertainty about locationsof previously found targets increases with each saccadeThis uncertainty is further exacerbated by interferencefrom newer target locations leading to increased prob-ability of misjudging old target locations for new assearch progresses Such progressively worsening uncer-tainty might be due to an accumulating impairment inlsquolsquore-mapping target locationsrsquorsquo across saccades or im-paired lsquolsquomemoryrsquorsquo of previously inspected target loca-tions across saccades As noted above there is evidence
for both types of signalmdash re-mapping and maintainingsaccadic spatial memorymdashin the parietal cortex and it ispossible that disruption to both these functions contrib-utes to the re-click deficit we have identified Certainlythe two potential types of deficit are not mutuallyincompatible
A slightly different account of the re-clicking behaviormight be framed in terms of the intriguing hypothesisproposed by Manly Woldt Watson and Warburton(2002) to explain visible re-marking on a standard can-cellation task They argue that such behavior is due topatients erroneously transposing leftward stimuli to theright resulting in apparent re-cancellation These au-thors found that removing targets from the left re-duced both neglect and re-marking However they didnot monitor eye movements so it is possible that sometargets that are logged as neglected (because they werenot cancelled) were nevertheless fixated and thereforeform part of the load on memory during search Fur-thermore in our experimental study subjects with ahigh re-click rate also failed to click on a higher pro-portion of fixated targets Manly et alrsquos proposal wouldnot easily account for such lsquolsquoclick omissionsrsquorsquo becausethese refer to fixated targets that were not actuallyresponded to
Finally it might be argued that increasing re-clickingwith search duration reflects a deficit in spatial workingmemory (SWM) that is independent of saccadic eyemovements (ie a decay in memory that is not trans-saccadic) Because time and number of interveningsaccades were not independently manipulated in thisunrestricted visual search paradigm we are not ablecategorically to distinguish between these possibilitiesWe note however that functional imaging studies ofSWM tasks that do not require eye movements oftendemonstrate activity related to the maintenance ofspatial information which is generally more posteriorand dorsal than the parietal locations isolated in thisstudy (DrsquoEsposito et al 1998) Moreover recent behav-ioral data indicate that SWM is more vulnerable tosaccadic eye movements than shifts in attention or limbmovements (Pearson amp Sahraie 2003)
Frontal Lesion Site Associated withHigh Re-click Rates
Four out of 5 neglect patients with lesions involvingposterior plus anterior damage also showed a high re-click rate and the area of damage common to thesepatients centered on the right inferior frontal lobe cor-responding to Brodmannrsquos area 44 (Figure 7C) Thisregion has previously been associated with frontalneglect (Husain amp Kennard 1996) and is consideredto be part of the ventral premotor cortex and thehomologue of Brocarsquos areas in the left hemisphere(Rizzolatti amp Arbib 1998 Jackson amp Husain 1996) Inthese frontal patients the probability of re-clicking did
Mannan et al 349
not increase with time interval between the first clickand re-fixation but rather it stayed constant or declinedwith increasing delays
Such behavior might be consistent with a failure toinhibit responses to rightward locationsmdasha type oflsquolsquoperseverativersquorsquo behavior Note though that this is notnecessarily the same type of perseverative responseas seen when some patients with neglect re-markpreviously found items on standard cancellation tests(Rusconi et al 2002 Na et al 1999) In those casestargets that are visibly cancelled are often marked againwhereas in our computerized search tasks no marks arevisibly left on targets once they have been clicked uponMoreover only one of our neglect patients (Case 1) de-monstrated re-cancellation of visibly cancelled targetson pen-and-paper tasks and then only on one occasionImportantly this patient actually had a normal re-clickrate (67) on our task and his stroke was in the PCAterritory sparing both the intraparietal sulcus and fron-tal lobe
In support of a role for the right frontal lobe in in-hibiting responses to previously found targets recentimaging studies of response inhibition have highlightedregions of the right inferior frontal gyrus (IFG) very closeto the centroid of damage for our frontal patients (38 1618 cf 43 22 21 Konishi Kyoichi Uchida Sekihara ampMiyashita 1998 and 41 16 19 Konishi Kyoichi UchidaKikyo et al 1999) Alternative explanations for the highprobability of re-clicking directly after clicking might bethat these patients have impaired perceptual discrimina-tion of targets impaired perceptual localization of targetsor that they encounter difficulty in encoding spatiallocations into working memory Our data are not sugges-tive of a perceptual discrimination problem as changingthe perceptual load did not inf luence re-click ratesMoreover the patients still had high re-click rates in thetarget only conditions (Figure 10) where no perceptualdiscrimination of targets and distractors was necessaryPoor perceptual localization would seem unlikely giventhat these patients have frontal lesions Furthermoreusing a vertical SWM task similar to the Corsi blocktapping task (Malhotra et al 2004) we have found thatpatients with neglect can localize and perceptually en-code single target locations as well as controls and aresignificantly impaired at remembering two or more loca-tions shown in sequence consistent with the presentobservations Some recent functional imaging data sug-gest that activation within the prefrontal cortex corre-lates well with successful encoding of material intoworking memory (Rypma amp DrsquoEsposito 2003) Furtherresearch is required to distinguish between these pos-sible explanations for the mechanisms underlying there-click deficit in frontal patients
The only patient with frontal damage who did re-clickmore frequently with increasing time had damage cen-tered on the dorsolateral prefrontal cortex (althoughalso encroaching on the IFG) Neurons in the dorsolat-
eral prefrontal cortex like those in the LIP encode theremembered locations of saccadic targets (Goldman-Rakic 1996) This patient (Case 15) who had the high-est re-click rate of all also had damage to the criticalparietal location described above (in contrast to all otherfrontal patients) Thus she may have suffered from acombination of underlying deficits
Re-fixations and Misjudging PreviouslyInspected Locations during Search
In this search task re-fixation and re-click rates weresignificantly correlated for neglect patients Howeverthere were important dissociations within individuals 2patients with PCA lesions had re-fixation rates of 220and 80 respectively but both had a re-click rate ofonly 7 These are clearly examples of individuals whodo not show pathological re-clicking despite having highre-fixation rates High re-fixation rates may result froma lack of integration of spatial information due to V1damage (Zihl amp von Cramon 1985) in our PCA patientsor possibly because of reduced inhibition of return(Bartolomeo Sieroff Decaix amp Chokron 2001) What-ever the exact cause for dissociations between re-fixa-tions and re-click rates our data suggest re-fixation doesnot necessarily index the same underlying deficits asrevealed by our re-click measure High re-fixation ratesunlike high re-click rates were also not significantlyassociated with damage to any specific cortical locationagain demonstrating a dissociation between re-fixationsand pathological re-clicking
The relationship between re-click rates and neglect isunlikely to be a simple one In our study there was amoderate but significant correlation between the twomeasures suggesting that this association is unlikely tohave arisen by chance However non-neglect patientscan also exhibit a high re-click rate (unpublished obser-vations from our laboratory) Taken together our resultssuggest that difficulties in keeping track of previouslyinspected targets (because of impaired trans-saccadic re-mapping andor memory) may be one contributingcomponent to visual neglect in patients with lesions in-volving the right intraparietal sulcus As mentioned thesearch task we used differed in one striking aspect fromstandard cancellation tests subjects do not visibly markthe targets they visit Whereas in cancellation tests pa-tients mark each target they find which might appear todispense with any need to remember which locationshave been searched before However it should be ap-preciated that in most standard cancellation tests theindividual marks made by patients may not be clearlyvisible in peripheral vision particularly if neglect pa-tients have a local spatial bias (Rafal 1994 RobertsonLamb amp Knight 1988) or a restricted field of view(Russell Malhotra amp Husain 2004) Thus when com-bined with a rightward attentional bias keeping track ofsearched locations may be critical to prevent gaze from
350 Journal of Cognitive Neuroscience Volume 17 Number 2
returning to rightward locations even if they have beenmarked before as the marks may not be visible untilgaze returns to them Consistent with this hypothesisneglect is greater when cancellation is performed usinginvisible marks compared to large and salient red marksthat can be seen peripherally (Wojciulik Husain Clarkeamp Driver 2001)
A task like the present search-and-click paradigm maymore closely resemble search in the real world than dotraditional cancellation tests When we examine a visualscene we do not leave marks on each object we ex-amine Instead just as in the search task used in thisstudy we have to keep track of the locations we visit ifwe are to avoid inefficient re-examination of thoselocations Neglect patients with damage to the rightintraparietal sulcus appear to be impaired at keepingtrack of searched locations in this way When combinedwith their rightward attentional bias we propose thatsuch an impairment leads to recursive search of stimulito the right in experimental search tasks as used hereand also in daily life where neglect is so apparentdespite unlimited viewing time
The paradigm we have used has the important virtuesof allowing patients to explore freelymdashas in real-worldsearch or clinical cancellation tasksmdashand relating theirre-click behavior directly to the severity of neglect in thesame task But it might be argued that it would be usefulalso to probe SWM in a more controlled manner by re-stricting observersrsquo eye movements to a specific set oflocations and probing memory of previously visitedplaces A recent study using a gaze-contingent para-digm has elegantly probed the capacity of spatial mem-ory during search in healthy observers and foundevidence for memory for up to four previous locations(McCarley Wang Kramer Irwin amp Peterson 2003)Variants of this paradigm may in the future be usefulto probe the capacity of SWM across saccades in patientswith neglect and also to probe further whether thereare differences between neglect patients with parietaland frontal lesions
METHODS
Subjects Imaging and Lesion Mapping
Sixteen patients with left neglect following right hemi-sphere infarction or hemorrhage were recruited Con-sent was obtained according to the Declaration ofHelsinki as approved by the hospital ethical commit-tees Presence of neglect was determined by clinicalobservation plus bedside tests including the Mesulamshape-cancellation task (Mesulam 1985) and line bisec-tion task (Wilson et al 1987) (see Table 1) Twelvepatients had MCA territory strokes the rest involvedonly the PCA territory which can also result in neglect(Cals et al 2002) Only one neglect patient (Case 1 whodid not have frontal involvement) demonstrated re-
cancellation of visibly cancelled targets on pen-and-paper tasks and then only on one occasion Fifteenage-matched healthy volunteers were controls (meanage = 63 SD = 14 vs 63 SD = 13 for patients)
All patients had clinical CT scans In addition dedi-cated high-resolution structural MRI scans were alsoobtained for 1416 neglect patients MRI scans wereunobtainable for the other two (Cases 11 and 16)because of claustrophobia or loss to follow up In theMCA neglect group median time between stroke andscan was 63 days with the shortest interval being35 days for the PCA neglect group one patient had tobe scanned relatively early at 9 days but the mediantime was 140 days MRI scans comprised one 256-sliceT1-weighted acquisition (MPRAGE TR 97 msec TE4 msec TA 12 min) performed in the coronal planealigned perpendicular to the anteriorndashposterior commis-sural line on a Siemens Vision 15-T scanner Within-and between-slice resolution was 1 mm For each pa-tient the area of damage was determined by detailedvisual inspection of the digital image for every singleslice Lesion delineation was performed by neurologists(authors D M and M H) by tracing by hand theboundary of the lesion onto every single 1-mm-thickaxial slice using a graphics tablet (WACOM Intuos A6)Combining all the slices then led to a 3-D region of in-terest (ROI) for each patient For one patient (Case 15)there were two areas of discrete focal damage one inthe frontal cortex and the other in the parietal cortex
Normalization of each MRI including the 3-D ROI tothe T1-template was performed with SPM99 (wwwfilionuclacukspm) The ROI was used as a mask for thelesion during determination of alignment parameters tominimize the contribution of abnormal brain to thisprocess (Rorden amp Brett 2000) The normalized ROIof each neglect patient was superimposed upon the T1template to find areas of maximal lesion overlap in eachgroup (see Results) as well as lesion volume usingMRIcro shareware (Rorden amp Brett 2000)
Experimental Procedure
Stimuli were presented on an SVGA monitor (29 22 in)viewed at 57 cm distance Each search display com-prised a virtual 9 7 grid each cell containing a single18 elementmdasheither a letter T (target) or a letter L (dis-tractor) (see Figure 1) The position of each elementwithin each cell was jittered with 19 targets and 44distractors randomly distributed over the virtual gridThe number of targets on the right and left halves ofthe display was balanced across trials Seven neglectpatients (Cases 5 8 10 11 14ndash16) were further testedon lsquolsquoTarget onlyrsquorsquo displays which had varying numbersof Ts (2 6 or 19) now with no L-shaped distractorsThe presentations of these lsquolsquoTarget onlyrsquorsquo screens wereinterspersed with those of the original search screens inrandom order in the same experimental session
Mannan et al 351
Eye movements were recorded using a video-basedpupil tracker with a temporal resolution of 4 msec andspatial accuracy of lt058 (EyeLink Sensorimotoric Sys-tems Berlin Germany) Subjects were asked to keeptheir head still The experimenter also monitored gazeposition on-line At the beginning of each session (andafter breaks in testing or following any unintended headmovement that disrupted gaze recordings) eye-trackingwas re-calibrated Each trial was initiated by the exper-imenter and began with a central fixation cross for 1 secthen the search display
Subjects were instructed to search for Ts (among anyLs) Critically while looking at any target they had toclick a response button held in the right hand onlyif they considered this target to be a new discovery(ie a target they had not clicked on before) Clickingthe response button did not change the display Theresponse button was held by the patient in a comfort-able position to the right of the midline Subjectswere instructed and reminded throughout testing toclick on each target found once only Any later re-click on
a previously found target thus indicates a failure to retainthe fact that this target had already been discoveredThe experimenter terminated each search when thesubject indicated verbally that they felt all targets in thedisplay had now been found Each subject searched aminimum of 5 displays (mean = 7 range 5ndash14) Eyemovements were analyzed off-line using custom soft-ware An automatic algorithm was used to extract sac-cades (saccade onset defined as eye speed rising above258 sec1 for two or more samples and a criterion ofminimum fixation duration of 60 msec) Blinks wereeliminated from the trace by visual inspection and sac-cade and fixation delineation were checked visually inthe record Next fixations were automatically assignedto items in the display if the mean position fell within18 of that item Two or more consecutive fixations onthe same item were re-scored as a single extendedfixation A re-fixation was scored only when at leastone fixation elsewhere on another item intervened
Most click responses were made during a fixation andthe few button-clicks occurring during a saccade were
Table 1 Patient Details
CaseAge
(years) PathologyVolume(cm3)
Age ofLesion (days)
Visual FieldDefect
Mean Deviationfrom Center on
Line Bisection Task(cm) (rightward +)
Mesulum Score(Maximum Possible
Score = 60) Lesion Type
1 68 Infarct 1282 64 + 867 17 PCA
2 76 Infarct 1528 227 + 003 41 PCA
3 68 Infarct 79 207 + 297 36 PCA
4 77 Infarct 251 158 + 150 54 PCA
5 63 Infarct 28 29 033 55 MCA
6 59 Hemorrhage 172 7 + 32 6 MCA
7 66 Hemorrhage 273 325 + 213 31 MCA
8 68 Infarct 309 3 25 17 MCA
9 29 Infarct 115 17 113 55 MCA
10 70 Infarct 1339 324 153 57 MCA
11 42 Infarct ndash 11 097 24 MCA
12 75 Infarct 757 38 013 4 MCA
13 51 Infarct 694 141 083 31 MCA
14 59 Infarct 1144 356 + 223 37 MCA
15 80 Infarct 261 44 35 6 MCA
16 64 Infarct ndash 30 033 55 MCA
MEAN 63 66 124 188 33
SE 4 13 32 5
Cases 1ndash9 had damage to the nonfrontal (posterior only) cortex whereas Cases 10ndash15 had both frontal and posterior (posterior and anterior) corticalinvolvement Case 16 is a patient with a subcortical lesion All but 3 of the patients with right MCA infarction had intact visual fields on confrontationTwo of these (Cases 7 and 14) were diagnosed with left homonymous hemianopia whereas one (Case 6) had a left inferior quandrantic field defectAll patients with PCA territory damage had partial or complete contralesional visual field loss No patient suffered from dysphasia
352 Journal of Cognitive Neuroscience Volume 17 Number 2
assigned to the previous fixation Clicks were assigned toa particular item if eye position at the time of the clickwas within 28 of that item or in the cases where morethan one item was within 28 of current eye position theclick response was simply attributed to the item closestto that eye position
Button-clicks were assigned to a particular item if eyeposition at the time of the click was within 28 of thatitem A re-click on an item required at least one fixationon another item between successive clicks (in practiceseveral fixations elsewhere typically intervened betweeninitial click and later re-click see Results)
Statistical analyses were performed using SPSS andSigmaplot (SPSS Science Chicago USA) Jonckheere sta-tistic was derived by hand Statistical significance was setas p lt 05
Acknowledgments
We thank the reviewers of this manuscript and Dr AndrewParton for their extremely helpful comments We thank thepatients who participated Dr J Wade Dr A Rudd and thestaff of the stroke units at Charing Cross and St ThomasrsquoHospitals This research was supported by grants from theWellcome Trust to M H C K and J D J D holds a RoyalSociety Wolfson Research Merit Award
Reprint requests should be sent to Drs Sabira Mannan andMasud Husain Division of Neuroscience and PsychologicalMedicine Imperial College London Charing Cross CampusLondon W6 8RF UK or via e-mail smannanimperialacukor mhusainimperialacuk
Notes
1 We are grateful to one of the reviewers for suggesting thistime-series analysis2 There was no significant difference between the number oftargets fixated between the two groups so differences in lsquolsquoclickomissionsrsquorsquo cannot be attributed to the number of fixatedtargets that had to be kept track of during search3 The correlation between these four points alone inFigure 9A is high [r(3) = 81 cf r(15) = 47 for the wholegroup]4 We have also tested 10 neglect patients and 10 controlstroke patients on a vertical version of the Corsi blocks task(clinically used to measure spatial working memory capacity)Neglect patients had significantly impaired capacity but wereable to localize single locations well suggesting that mislocal-ization is unlikely to account for their deficit on this spatialworking memory task
REFERENCES
Bartolomeo P Sieroff E Decaix C amp Chokron S (2001)Modulating the attentional bias in unilateral neglect theeffects of the strategic set Experimental Brain Research137 432ndash444
Bates E Wilson S M Saygin A P Dick F Sereno M IKnight R T amp Dronkers N F (2003) Voxel-basedlesion-symptom mapping Nature Neuroscience 6 448ndash450
Behrmann M Watt S Black S E amp Barton J J (1997)Impaired visual search in patients with unilateral neglect
An oculographic analysis Neuropsychologia 351445ndash1458
Bisiach E amp Vallar G (2000) Unilateral neglect in humans InG Rizzolatti (Ed) Handbook of neuropsychology (2nd edvol 1 pp 459ndash502) Amsterdam Elsevier
Cals N Devuyst G Afsar N Karapanayiotides T ampBogousslavsky J (2002) Pure superficial posterior cerebralartery territory infarction in The Lausanne Stroke RegistryJournal of Neurology 249 855ndash861
Chafee M amp Goldman-Rakic P (1998) Matching patterns ofactivity in primate prefrontal area 8a and parietal area 7ipneurons during a spatial working memory task Journal ofNeurophysiology 79 2919ndash2940
Chedru F Leblanc M amp Lhermitte F (1973) Visualsearching in normal and brain-damaged subjectsContributions to the study of unilateral subjectsCortex 9 94ndash111
DrsquoEsposito M Aguirre G K Zarahn E Ballard DShin R K amp Lease J (1998) Functional MRI studies ofspatial and nonspatial working memory Cognitive BrainResearch 7 1ndash13
Donkelaar P amp Muri R (2002) Craniotopic updating ofvisual space across saccades in the human posteriorparietal cortex Proceedings of the Royal Society London B269 735ndash739
Driver J amp Husain M (2002) The role of spatial workingmemory deficits in pathological search by neglect patientsIn the cognitive and neural bases of spatial neglect InH-O Karnath A D Milner amp G Vallar (Eds) The cognitiveand neural bases of spatial neglect Oxford OxfordUniversity Press
Duhamel J R Colby C amp Goldberg M E (1992) Theupdating of the representation of visual space in parietalcortex by intended eye movements Science 25590ndash92
Goldman-Rakic P S (1996) The prefrontal landscapemdashImplications of functional architecture for understandinghuman mentation and the central executive PhilosophicalTransactions of The Royal Society of London Series BBiological Sciences 351 1445ndash1453
Heide W Binkofski F Seitz R J Posse S Nitschke M FFreund H J amp Kompf D (2001) Activation offrontoparietal cortices during memorized triple-stepsequences of saccadic eye movements An fMRI studyEuropean Journal of Neuroscience 13 1177ndash1189
Heide W amp Kompf D (1998) Combined deficits ofsaccades and visuo-spatial orientation after cortical lesionsExperimental Brain Research 123 164ndash171
Husain M amp Kennard C (1996) Visual neglect associatedwith frontal lobe infarction Journal of Neurology 243652ndash657
Husain M Mannan S Hodgson T Wojciulik E Driver Jamp Kennard C (2001) Impaired spatial working memoryacross saccades contributes to abnormal search in parietalneglect Brain 121 941ndash952
Husain M amp Rorden C (2003) Non-spatially lateralizedmechanisms in hemispatial neglect Nature ReviewsNeuroscience 4 23ndash36
Jackson S R amp Husain M (1996) Visuomotor functionsof the lateral pre-motor cortex Current Opinion inNeurobiology 6 788ndash795
Kinsbourne M (1993) Orientational bias model of unilateralneglect Evidence from attentional gradients withinhemispace In I H Robertson amp J C Marshall (Eds)Unilateral neglect Clinical and experimental studies(pp 63ndash86) Hove Erlbaum
Konishi S Kyoichi N Uchida I Kikyo H Kameyama Mamp Miyauchi S (1999) Common inhibitory mechanism in
Mannan et al 353
human inferior prefrontal cortex revealed by event-relatedfunctional MRI Brain 122 981ndash991
Konishi S Kyoichi N Uchida I Sekihara K amp Miyashita Y(1998) No-go dominant brain activity in human inferiorprefrontal cortex revealed by functional magneticresonance imaging European Journal of Neuroscience10 1209ndash1213
Malhotra P Mannan S Driver J amp Husain M (2004)Impaired spatial working memory One component of thevisual neglect syndrome Cortex 40 667ndash676
Manly T Woldt K Watson P amp Warburton E (2002) Ismotor perseveration in unilateral neglect lsquolsquodrivenrsquorsquo by thepresence of neglected left-sided stimuli Neuropsychologia40 1794ndash1803
Mazzoni P Bracewell R Barash S amp Andersen R (1996)Spatially tuned auditory responses in area LIP of macaquesperforming delayed memory saccades to acoustic targetsJournal of Neurophysiology 75 1233ndash1241
McCarley J Wang R Kramer A Irwin D amp Peterson M(2003) How much memory does oculomotor search havePsychological Science 14 422ndash426
Mesulam M-M (1985) Principles of behavioural neurologyTests of directed attention and memory Philadelphia Davis
Mesulam M M (1999) Spatial attention and neglectParietal frontal and cingulate contributions to the mentalrepresentation and attentional targeting of salientextrapersonal events Philosophical Transactions of theRoyal Society of London Series B Biological Sciences354 1325ndash1346
Mort D Malhotra P Mannan S Rorden C Pambakian AKennard C amp Husain M (2003) The anatomy of visualneglect Brain 126 1986ndash1997
Na D L Adair J C Kang Y Chung C S Lee K H ampHeilman K M (1999) Motor perseverative behaviour ona line cancellation task Neurology 52 1569ndash1576
Pearson D G amp Sahraie A (2003) Oculomotor controland the maintenance of spatially and temporally distributedevents in visuo-spatial working memory QuarterlyJournal of Experimental Psychology 56A 1089ndash1111
Rafal R (1994) Neglect Current Opinion in Neurobiology4 231ndash236
Rizzolatti G amp Arbib M A (1998) Language within our graspTrends in Neurosciences 21 188ndash194
Robertson L C Lamb M R amp Knight R T (1988) Effectsof lesions of temporalndashparietal junction on perceptual andattentional processing in humans Journal of Neuroscience8 3757ndash3769
Rorden C amp Brett M (2000) Stereotaxic display of brainlesions Behavioural Neurology 12 191ndash200
Rusconi M L Maravita A Bottini G amp Vallar G (2002)Is the intact side really intact Perseverative responses inpatients with unilateral neglect A productive manifestationNeuropsychologia 40 594ndash604
Russell C Malhotra P amp Husain M (2004) Attentionmodulates the visual field in healthy observers and parietalpatients NeuroReport 15 189ndash193
Rypma B amp DrsquoEsposito M (2003) A subsequent-memoryeffect in dorsolateral prefrontal cortex Cognitive BrainResearch 16 162ndash166
Sapir A Hayes A Henik A Danzinger S amp Rafal R(2004) Parietal lobe lesions disrupt saccadic remapping ofinhibitory location tagging Journal of CognitiveNeuroscience 16 503ndash509
Smania N Martini M C Gambina G A T Palamara ANatale E amp Marzi C A (1998) The spatial distributionof visual attention in hemineglect and extinction patientsBrain 121 1759ndash1770
Sprenger A Kompf D amp Heide W (2002) Visual searchin patients with left visual hemineglect Progress in BrainResearch 140 395ndash416
Tobler P Felblinger J Burki M Nirkko A Ozdoba C ampMuri R (2001) Functional organisation of the saccadicreference system processing extraretinal signals in humansVision Research 41 1351ndash1358
Wilson B Cockburn J amp Halligan P W (1987)Behavioural inattention test Bury St EdmundsThames Valley
Wojciulik E Husain M Clarke K amp Driver J (2001)Spatial working memory deficit in unilateral neglectNeuropsychologia 39 390ndash396
Zihl J amp Hebel N (1997) Patterns of oculomotor scanningin patients with unilateral posterior parietal and frontallobe damage Neuropsychologia 35 893ndash906
Zihl J amp von Cramon D (1985) Visual field recovery fromscotoma in patients with postgeniculate damage A reviewof 55 cases Brain 108 335ndash365
354 Journal of Cognitive Neuroscience Volume 17 Number 2
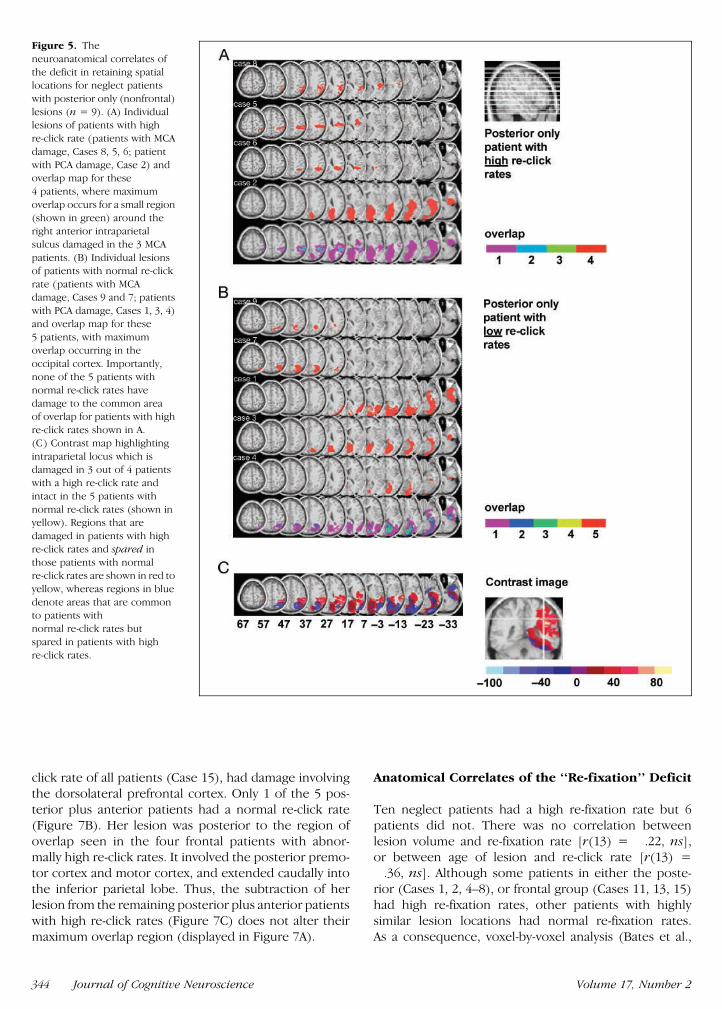
click rate of all patients (Case 15) had damage involvingthe dorsolateral prefrontal cortex Only 1 of the 5 pos-terior plus anterior patients had a normal re-click rate(Figure 7B) Her lesion was posterior to the region ofoverlap seen in the four frontal patients with abnor-mally high re-click rates It involved the posterior premo-tor cortex and motor cortex and extended caudally intothe inferior parietal lobe Thus the subtraction of herlesion from the remaining posterior plus anterior patientswith high re-click rates (Figure 7C) does not alter theirmaximum overlap region (displayed in Figure 7A)
Anatomical Correlates of the lsquolsquoRe-fixationrsquorsquo Deficit
Ten neglect patients had a high re-fixation rate but 6patients did not There was no correlation betweenlesion volume and re-fixation rate [r(13) = 22 ns]or between age of lesion and re-click rate [r(13) =36 ns] Although some patients in either the poste-rior (Cases 1 2 4ndash8) or frontal group (Cases 11 13 15)had high re-fixation rates other patients with highlysimilar lesion locations had normal re-fixation ratesAs a consequence voxel-by-voxel analysis (Bates et al
Figure 5 The
neuroanatomical correlates of
the deficit in retaining spatiallocations for neglect patients
with posterior only (nonfrontal)
lesions (n = 9) (A) Individuallesions of patients with high
re-click rate (patients with MCA
damage Cases 8 5 6 patient
with PCA damage Case 2) andoverlap map for these
4 patients where maximum
overlap occurs for a small region
(shown in green) around theright anterior intraparietal
sulcus damaged in the 3 MCA
patients (B) Individual lesionsof patients with normal re-click
rate (patients with MCA
damage Cases 9 and 7 patients
with PCA damage Cases 1 3 4)and overlap map for these
5 patients with maximum
overlap occurring in the
occipital cortex Importantlynone of the 5 patients with
normal re-click rates have
damage to the common areaof overlap for patients with high
re-click rates shown in A
(C) Contrast map highlighting
intraparietal locus which isdamaged in 3 out of 4 patients
with a high re-click rate and
intact in the 5 patients with
normal re-click rates (shown inyellow) Regions that are
damaged in patients with high
re-click rates and spared in
those patients with normalre-click rates are shown in red to
yellow whereas regions in blue
denote areas that are commonto patients with
normal re-click rates but
spared in patients with high
re-click rates
344 Journal of Cognitive Neuroscience Volume 17 Number 2
2003) revealed no anatomical specificity associated withhigh re-fixation rates [t(7) = 085 ns] unlike thatassociated with high re-click rates even when onlyfocal lesions were considered [t(5) = 165 ns]
Time Interval between Clicking and Re-clicking ona Target
We next examined the probability of lsquolsquore-clickingrsquorsquo on atarget when re-fixating it as a function of the time lag ornumber of intervening saccades between the first clickon that target and the subsequent revisit An increasedprobability of re-clicking with increasing delay (or inter-vening number of saccades) would be consistent a failureto keep track of spatial locations during search whereasa flat or decreasing function might arise from impairedresponse inhibition (lsquolsquoperseverativersquorsquo behavior) or even
a failure to encode target location This time-series anal-ysis for the 10 patients with high re-click rates revealedtwo distinct patterns of behavior Half the patients de-monstrated a significantly increasing probability of mis-judging an already found item to be a novel one withincreasing delay (Figure 8A) or intervening saccades(Figure 8C) whereas the other 5 patients had a constantor even decreasing probability (Figure 8B and D)
Importantly most (45) patients in the group withincreasing re-click functions had damage to the focalparietal location previously identified in the analysisof patients with focal lesions and high re-click rates(Figure 6C) The other patient was Case 2 who had alarge PCA lesion extending to the white matter deep tothe occipito-temporo-parietal junction By contrast noneof the patients with flatdecreasing functions had dam-age to the key parietal region Instead the latter group
Figure 6 The
neuroanatomical correlates of
the deficit in retaining spatial
locations for neglect patientswith focal lesions (n = 7)
(A) Individual lesions of
patients with high re-click rate
(all 4 patients have MCAdamage) and overlap map for
these 4 patients where
maximum overlap occurs for asmall region around the right
intraparietal sulcus (shown in
red) damaged all 4 MCA
patients (B) Individual lesionsof patients with normal re-click
rate (all patients with PCA
damage) and overlap map for
these 3 patients with maximumoverlap occurring in the
occipital cortex Importantly
none of these 3 patients withnormal re-click rates have
damage to the common area of
overlap for patients with high
re-click rates shown in A(C) Contrast map highlighting
intraparietal locus which is
damaged in 4 out of 4 patients
with a high re-click rate andintact in the 3 patients with
normal re-click rates (shown in
yellow) Regions that are
damaged in patients with highre-click rates and spared in
those patients with normal
re-click rates are shown in redto yellow whereas regions in
blue denote areas that are
common to patients with
normal re-click rates butspared in patients with high
re-click rates
Mannan et al 345
had frontal cortical or subcortical involvement Thusincreasing re-clicking with time interval was associatedwith parietal damage whereas flat re-click functionswere associated with frontalsubcortical lesions
For neglect patients as a whole the intervals betweenan initial click on a target and a subsequent re-click werelong (mean = 40 saccades) but much shorter betweenan initial fixation and a later re-fixation (mean = 5) withthis difference being significant [t(15) = 315 p lt01] This again emphasizes the dissociation betweenre-fixations and re-clicks and also demonstrates that re-clicking does not occur on most re-fixations
Relationship of Re-click Rate to Neglect Severity
We assessed three separate measures of neglect Thefirst was the difference between miss rates for left ver-sus right targets within the experimental search taskitself the other two measures of neglect were based onthe clinical pen-and-paper measures of Mesulam shape-cancellation (Mesulam 1985) and line bisection fromthe BIT (Wilson Cockburn amp Halligan 1987)
On the experimental search task there was a moderatebut significant correlation between re-click rate and ne-glect in search across the group [r(15) = 47 p lt 05
Figure 9A] Thus the greater the re-clicking on right-ward items the greater the severity of leftward neglectThere was also a moderate but again significant corre-lation between re-click rates and the severity of individualpatientsrsquo clinical neglect as indexed by the cancellationmeasure [r(15) = 46 p lt 05 Figure 9B] Thesecorrelations were driven largely by three of the patientsidentified in our focal lesion analysis (Figure 6A) withinvolvement of the key parietal region and high re-clicking behavior Their data are shown in filled circlesto the right in the graphs displayed in Figure 93 Thecorrelation between deviation on line bisection andre-click rate was not significant [r(15) = 13 ns] How-ever one would not predict that a deficit in retaininglocations already visited (or response inhibition) wouldnecessarily impair performance on a task such as linebisection
Effect of Varying Number of Targets
In 7 patients (Cases 5 8 10 11 14ndash16) we also exam-ined the effect of varying the number of targets in thesearch task Patients viewed lsquolsquotarget onlyrsquorsquo displays con-taining either 2 6 or 19 targets randomly distributedon the screen without any L-shaped distractors In-
Figure 7 Theneuroanatomical correlates of
the re-click deficit in neglect
patients with posterior plusanterior lesions (n = 5)
(A) Individual lesions of
patients with high re-click rate
(all 4 patients have MCAdamage) and overlap map for
these 4 patients where
maximum overlap occurs for
a region in the right inferiorfrontal lobe (shown in red)
damaged all 4 MCA patients
(B) Individual lesions of a single
patient with normal re-clickrate (C) Contrast map
highlighting the inferior frontal
locus (shown in yellow) whichis damaged in 4 out of
4 patients with a high re-click
rate and intact in the patient
with normal re-click rateRegions that are damaged in
patients with high re-click rates
and spared in the patient
with normal re-click rate areshown in red to yellow
whereas regions in blue denote
areas that are damaged in thepatient with normal re-click
rates but spared in patients with
high re-click rates
346 Journal of Cognitive Neuroscience Volume 17 Number 2
creasing the number of targets led to significantly higherre-click rates ( Jonckheere statistic = 133 p lt 01Figure 10) as well as more severe neglect (Jonckheerestatistic = 129 p lt 01) consistent with the proposalthat the mechanisms underlying pathological re-clickingmight contribute to neglect behavior Note that a lsquolsquoper-severativersquorsquo failure of response inhibition would not onits own easily explain this finding For the same absolutenumber of click responses one would predict a higherre-click rate with smaller set-sizes than with larger ones(see also Husain Mannan et al 2001 for a discussion)Although the number of subjects were small there wasno difference in the performance pattern between the3 patients who had increasing re-click rate functions inthe time-series analysis and the 4 patients with flat ordecreasing re-click rate functions
Varying Perceptual Load during Search
Can re-click behavior be explained simply by impairedvisual perception of targets from discriminators Neglectpatients did find it more difficult than control subjectsto select targets from distractors fixating on average19 distractors for every target found [cf 14 for controlsubjects t(29) = 485 p lt 0001] Patients also some-times clicked on distractors (64) but no more often
than controls (61) and it is possible that this rate iseffectively inflated by our strict criterion of considering aclick to be a response on the item that is being fixated atthe time of the click (It is likely that sometimes ob-servers click in response to targets they fixated one ormore saccades previously) Re-clicks on distractors wereextremely rare (06 cf 30 for targets)
To examine the effect of reduced perceptual load in8 patients (Cases 5 7 8ndash10 13 1 4 16) 6 of whom werehigh re-clickers we used search displays with salientcross (+) targets rather than Ts L distractors wereunchanged Patients fixated significantly fewer distrac-tors when searching for cross (+) targets [distractortarget ratio of 134 cf 181 for T targets t(7) = 60 p lt001] suggesting they did indeed find targetndashdistractordiscrimination easier Importantly however there wasno significant difference in re-click rate [26 for +targets cf 27 for T targets t(14) = 041 ns] or evenin re-fixation rate [100 for + targets cf 119 fort(14) = 041 ns] Thus these changes in perceptualload did not influence re-click performance
DISCUSSION
On this multitarget visual search task neglect patientsmissed targets to their left and also re-fixated targets
Figure 8 The probability of
re-clicking or re-fixating on
target as a function of delay
between the original click andsubsequent re-fixation (for
patients with high re-click
rates) (A) Probability functions
for patients with significantpositive slopes Four patients of
these patients have damage to
the focal intraparietal locusidentified in Figure 6C whereas
the single patient with a PCA
lesion and no damage to the
critical parietal locationindicated by symbol has only
a very shallow positive slope
(B) Probability functions for
patients with no significantpositive slope None of these
5 patients have damage to the
critical intraparietal region (C)As for A but for delay measured
in terms of intervening
fixations (D) As for B but for
delay measured in terms ofintervening fixations
Mannan et al 347
to the right much more often than healthy individuals(Figure 2A) Critically 10 of the 16 neglect patients re-clicked on rightward targets at a significantly higher ratethan healthy control subjects (Figure 2B) For 5 of thesepatients the probability of re-clicking increased withtime (Figure 8A) whereas the other 5 neglect patientswith high re-click rates demonstrated a constant or de-creasing probability of re-clicking over time (Figure 8C)The remaining 6 neglect patients had re-click rateswithin the normal range demonstrating that neitherneglect nor brain damage per se leads to a pathologicalre-click rate
High-resolution MRI mapping of lesions revealed im-portant anatomical dissociations within the neglect pop-ulation Patients with pathological re-click rates hadlesions affecting either the right intraparietal sulcus orthe inferior frontal lobe the only exception (Case 2) hada large occipito-temporal lesion that extended deep intothe parietal white matter Neglect patients with normalre-click rates did not have damage to either of thecritical locations in the intraparietal sulcus or inferiorfrontal lobe (Figures 5 6 and 7) Instead they had moredorsal lesions (for MCA territory patients) or occipitaland medial-temporal damage (for PCA territory lesions)which can also result in neglect (Mort et al 2003Cals Devuyst Afsar Karapanayiotides amp Bogousslavsky2002) Importantly lesion location rather than the sizeor age of lesion was critical to the presence of the re-clicking impairment Across the group of neglect pa-tients there was a moderate but significant correlationbetween re-click rate and degree of neglect (Figure 9)driven largely by patients with parietal damage who hadboth a high re-click rate and also severe neglect consis-tent with the proposal that mechanisms underlying thisbehavior in parietal patients might have a functionallyimportant role in determining the severity of neglect
Role of Right Intraparietal Sulcus
Notably for all patients with damage to the critical locusin the right intraparietal sulcus (which may also involveparieto-frontal white matter fibers) the probability of re-clicking on a target increased with time (or interveningsaccades) since initial discovery (Figure 8) Because theyrarely re-clicked at short intervalsmdasheven though themajority of re-fixations occurred during this periodmdashthis pattern of behavior suggests that for these patientstarget locations were actually veridically perceived and
Figure 9 (A) Neglect on experimental search task (19 targets and44 distractors) plotted against mean re-click rate for individual neglect
patients Percentage neglect was calculated as the difference between
the number of targets missed on the left versus right expressed as apercentage of the total number of targets missed There was a
significant correlation between the degree of neglect and the re-click
rate [r(15) = 465 p lt 05] Patients with high re-click rates are
highlighted by black circles (for patients with damage to the focalparietal location identified in Figure 6C) and light gray circles (for
patients with frontal damage) while the single patient with PCA
damage and a high re-click rate is indicated by dark gray (B) The
number of targets missed on the Mesulam shape-cancellation taskplotted against mean re-click rate for individual neglect patients Again
there was a significant correlation between the number of targets
missed and the mean re-click rate [r(15) = 456 p lt 05] Patients withhigh re-click rates are denoted as in A
Figure 10 Mean re-click rate (filled circles) and degree of neglect
(unfilled squares) increased systematically with increasing numbers of
targets in target-only displays
348 Journal of Cognitive Neuroscience Volume 17 Number 2
well encoded into working memory Furthermore de-tailed single-case studies of two parietal patients withhigh re-click rates (Malhotra Mannan Driver amp Husain2004 Husain Mannan et al 2001) demonstrate usingdifferent psychophysical methods that these patientscan accurately encode spatial locations4 Increasing re-clicks with time (or intervening saccades) would how-ever be consistent with a deficit in keeping track oftarget locations during extended search
Several lines of evidence support the proposal thatthe critical intraparietal sulcus region we have identifiedmay have an important role in the spatial transforma-tions required to keep track of spatial locations acrosssaccades In rhesus monkeys neurons within the lateralbank of the intraparietal sulcus (area LIP) encode andlsquolsquomaintainrsquorsquo the remembered locations of saccadic tar-gets over time (Chafee amp Goldman-Rakic 1998 MazzoniBracewell Barash amp Andersen 1996) Some visuallyresponsive neurons in the LIP also appear to shift theirretinotopic representation at the onset of an intendedsaccade as if lsquolsquore-mappingrsquorsquo representations of objects inspace to update them across shifts of gaze (DuhamelColby amp Goldberg 1992)
In humans this computation has been investigatedusing the double-step saccade paradigm where twoflashes of light appear in quick succession such thatthe second target disappears before a saccade can bemade to the first target Patients with lesions involvingthe right parietal cortex are impaired at performing thistask suggesting that they are unable to update spatialrepresentations accurately after even one saccade (Heideamp Kompf 1998 Duhamel et al 1992) Similarly trans-cranial magnetic stimulation of the posterior parietalcortex prior to the second saccade reduces its accuracy(Donkelaar amp Muri 2002) and a recent study of patientswith parietal lesions also implicates this area in thespatial updating of inhibition of return after a saccade(Sapir Hayes Henik Danzinger amp Rafal 2004) Func-tional imaging studies of double-step (Tobler et al2001) and triple-step memory-guided saccades (HeideBinkofski et al 2001) have identified a region withinthe intraparietal sulcus (Talairach coordinates 40 4044 for Tobler et al and 44 48 36 Heide et al) This isclose to the critical region of lesion overlap we haveidentified in the posterior neglect patients with high re-click rates (Talairach 37 39 37)
We suggest that in our patients with lesions involvingthe right intraparietal sulcus uncertainty about locationsof previously found targets increases with each saccadeThis uncertainty is further exacerbated by interferencefrom newer target locations leading to increased prob-ability of misjudging old target locations for new assearch progresses Such progressively worsening uncer-tainty might be due to an accumulating impairment inlsquolsquore-mapping target locationsrsquorsquo across saccades or im-paired lsquolsquomemoryrsquorsquo of previously inspected target loca-tions across saccades As noted above there is evidence
for both types of signalmdash re-mapping and maintainingsaccadic spatial memorymdashin the parietal cortex and it ispossible that disruption to both these functions contrib-utes to the re-click deficit we have identified Certainlythe two potential types of deficit are not mutuallyincompatible
A slightly different account of the re-clicking behaviormight be framed in terms of the intriguing hypothesisproposed by Manly Woldt Watson and Warburton(2002) to explain visible re-marking on a standard can-cellation task They argue that such behavior is due topatients erroneously transposing leftward stimuli to theright resulting in apparent re-cancellation These au-thors found that removing targets from the left re-duced both neglect and re-marking However they didnot monitor eye movements so it is possible that sometargets that are logged as neglected (because they werenot cancelled) were nevertheless fixated and thereforeform part of the load on memory during search Fur-thermore in our experimental study subjects with ahigh re-click rate also failed to click on a higher pro-portion of fixated targets Manly et alrsquos proposal wouldnot easily account for such lsquolsquoclick omissionsrsquorsquo becausethese refer to fixated targets that were not actuallyresponded to
Finally it might be argued that increasing re-clickingwith search duration reflects a deficit in spatial workingmemory (SWM) that is independent of saccadic eyemovements (ie a decay in memory that is not trans-saccadic) Because time and number of interveningsaccades were not independently manipulated in thisunrestricted visual search paradigm we are not ablecategorically to distinguish between these possibilitiesWe note however that functional imaging studies ofSWM tasks that do not require eye movements oftendemonstrate activity related to the maintenance ofspatial information which is generally more posteriorand dorsal than the parietal locations isolated in thisstudy (DrsquoEsposito et al 1998) Moreover recent behav-ioral data indicate that SWM is more vulnerable tosaccadic eye movements than shifts in attention or limbmovements (Pearson amp Sahraie 2003)
Frontal Lesion Site Associated withHigh Re-click Rates
Four out of 5 neglect patients with lesions involvingposterior plus anterior damage also showed a high re-click rate and the area of damage common to thesepatients centered on the right inferior frontal lobe cor-responding to Brodmannrsquos area 44 (Figure 7C) Thisregion has previously been associated with frontalneglect (Husain amp Kennard 1996) and is consideredto be part of the ventral premotor cortex and thehomologue of Brocarsquos areas in the left hemisphere(Rizzolatti amp Arbib 1998 Jackson amp Husain 1996) Inthese frontal patients the probability of re-clicking did
Mannan et al 349
not increase with time interval between the first clickand re-fixation but rather it stayed constant or declinedwith increasing delays
Such behavior might be consistent with a failure toinhibit responses to rightward locationsmdasha type oflsquolsquoperseverativersquorsquo behavior Note though that this is notnecessarily the same type of perseverative responseas seen when some patients with neglect re-markpreviously found items on standard cancellation tests(Rusconi et al 2002 Na et al 1999) In those casestargets that are visibly cancelled are often marked againwhereas in our computerized search tasks no marks arevisibly left on targets once they have been clicked uponMoreover only one of our neglect patients (Case 1) de-monstrated re-cancellation of visibly cancelled targetson pen-and-paper tasks and then only on one occasionImportantly this patient actually had a normal re-clickrate (67) on our task and his stroke was in the PCAterritory sparing both the intraparietal sulcus and fron-tal lobe
In support of a role for the right frontal lobe in in-hibiting responses to previously found targets recentimaging studies of response inhibition have highlightedregions of the right inferior frontal gyrus (IFG) very closeto the centroid of damage for our frontal patients (38 1618 cf 43 22 21 Konishi Kyoichi Uchida Sekihara ampMiyashita 1998 and 41 16 19 Konishi Kyoichi UchidaKikyo et al 1999) Alternative explanations for the highprobability of re-clicking directly after clicking might bethat these patients have impaired perceptual discrimina-tion of targets impaired perceptual localization of targetsor that they encounter difficulty in encoding spatiallocations into working memory Our data are not sugges-tive of a perceptual discrimination problem as changingthe perceptual load did not inf luence re-click ratesMoreover the patients still had high re-click rates in thetarget only conditions (Figure 10) where no perceptualdiscrimination of targets and distractors was necessaryPoor perceptual localization would seem unlikely giventhat these patients have frontal lesions Furthermoreusing a vertical SWM task similar to the Corsi blocktapping task (Malhotra et al 2004) we have found thatpatients with neglect can localize and perceptually en-code single target locations as well as controls and aresignificantly impaired at remembering two or more loca-tions shown in sequence consistent with the presentobservations Some recent functional imaging data sug-gest that activation within the prefrontal cortex corre-lates well with successful encoding of material intoworking memory (Rypma amp DrsquoEsposito 2003) Furtherresearch is required to distinguish between these pos-sible explanations for the mechanisms underlying there-click deficit in frontal patients
The only patient with frontal damage who did re-clickmore frequently with increasing time had damage cen-tered on the dorsolateral prefrontal cortex (althoughalso encroaching on the IFG) Neurons in the dorsolat-
eral prefrontal cortex like those in the LIP encode theremembered locations of saccadic targets (Goldman-Rakic 1996) This patient (Case 15) who had the high-est re-click rate of all also had damage to the criticalparietal location described above (in contrast to all otherfrontal patients) Thus she may have suffered from acombination of underlying deficits
Re-fixations and Misjudging PreviouslyInspected Locations during Search
In this search task re-fixation and re-click rates weresignificantly correlated for neglect patients Howeverthere were important dissociations within individuals 2patients with PCA lesions had re-fixation rates of 220and 80 respectively but both had a re-click rate ofonly 7 These are clearly examples of individuals whodo not show pathological re-clicking despite having highre-fixation rates High re-fixation rates may result froma lack of integration of spatial information due to V1damage (Zihl amp von Cramon 1985) in our PCA patientsor possibly because of reduced inhibition of return(Bartolomeo Sieroff Decaix amp Chokron 2001) What-ever the exact cause for dissociations between re-fixa-tions and re-click rates our data suggest re-fixation doesnot necessarily index the same underlying deficits asrevealed by our re-click measure High re-fixation ratesunlike high re-click rates were also not significantlyassociated with damage to any specific cortical locationagain demonstrating a dissociation between re-fixationsand pathological re-clicking
The relationship between re-click rates and neglect isunlikely to be a simple one In our study there was amoderate but significant correlation between the twomeasures suggesting that this association is unlikely tohave arisen by chance However non-neglect patientscan also exhibit a high re-click rate (unpublished obser-vations from our laboratory) Taken together our resultssuggest that difficulties in keeping track of previouslyinspected targets (because of impaired trans-saccadic re-mapping andor memory) may be one contributingcomponent to visual neglect in patients with lesions in-volving the right intraparietal sulcus As mentioned thesearch task we used differed in one striking aspect fromstandard cancellation tests subjects do not visibly markthe targets they visit Whereas in cancellation tests pa-tients mark each target they find which might appear todispense with any need to remember which locationshave been searched before However it should be ap-preciated that in most standard cancellation tests theindividual marks made by patients may not be clearlyvisible in peripheral vision particularly if neglect pa-tients have a local spatial bias (Rafal 1994 RobertsonLamb amp Knight 1988) or a restricted field of view(Russell Malhotra amp Husain 2004) Thus when com-bined with a rightward attentional bias keeping track ofsearched locations may be critical to prevent gaze from
350 Journal of Cognitive Neuroscience Volume 17 Number 2
returning to rightward locations even if they have beenmarked before as the marks may not be visible untilgaze returns to them Consistent with this hypothesisneglect is greater when cancellation is performed usinginvisible marks compared to large and salient red marksthat can be seen peripherally (Wojciulik Husain Clarkeamp Driver 2001)
A task like the present search-and-click paradigm maymore closely resemble search in the real world than dotraditional cancellation tests When we examine a visualscene we do not leave marks on each object we ex-amine Instead just as in the search task used in thisstudy we have to keep track of the locations we visit ifwe are to avoid inefficient re-examination of thoselocations Neglect patients with damage to the rightintraparietal sulcus appear to be impaired at keepingtrack of searched locations in this way When combinedwith their rightward attentional bias we propose thatsuch an impairment leads to recursive search of stimulito the right in experimental search tasks as used hereand also in daily life where neglect is so apparentdespite unlimited viewing time
The paradigm we have used has the important virtuesof allowing patients to explore freelymdashas in real-worldsearch or clinical cancellation tasksmdashand relating theirre-click behavior directly to the severity of neglect in thesame task But it might be argued that it would be usefulalso to probe SWM in a more controlled manner by re-stricting observersrsquo eye movements to a specific set oflocations and probing memory of previously visitedplaces A recent study using a gaze-contingent para-digm has elegantly probed the capacity of spatial mem-ory during search in healthy observers and foundevidence for memory for up to four previous locations(McCarley Wang Kramer Irwin amp Peterson 2003)Variants of this paradigm may in the future be usefulto probe the capacity of SWM across saccades in patientswith neglect and also to probe further whether thereare differences between neglect patients with parietaland frontal lesions
METHODS
Subjects Imaging and Lesion Mapping
Sixteen patients with left neglect following right hemi-sphere infarction or hemorrhage were recruited Con-sent was obtained according to the Declaration ofHelsinki as approved by the hospital ethical commit-tees Presence of neglect was determined by clinicalobservation plus bedside tests including the Mesulamshape-cancellation task (Mesulam 1985) and line bisec-tion task (Wilson et al 1987) (see Table 1) Twelvepatients had MCA territory strokes the rest involvedonly the PCA territory which can also result in neglect(Cals et al 2002) Only one neglect patient (Case 1 whodid not have frontal involvement) demonstrated re-
cancellation of visibly cancelled targets on pen-and-paper tasks and then only on one occasion Fifteenage-matched healthy volunteers were controls (meanage = 63 SD = 14 vs 63 SD = 13 for patients)
All patients had clinical CT scans In addition dedi-cated high-resolution structural MRI scans were alsoobtained for 1416 neglect patients MRI scans wereunobtainable for the other two (Cases 11 and 16)because of claustrophobia or loss to follow up In theMCA neglect group median time between stroke andscan was 63 days with the shortest interval being35 days for the PCA neglect group one patient had tobe scanned relatively early at 9 days but the mediantime was 140 days MRI scans comprised one 256-sliceT1-weighted acquisition (MPRAGE TR 97 msec TE4 msec TA 12 min) performed in the coronal planealigned perpendicular to the anteriorndashposterior commis-sural line on a Siemens Vision 15-T scanner Within-and between-slice resolution was 1 mm For each pa-tient the area of damage was determined by detailedvisual inspection of the digital image for every singleslice Lesion delineation was performed by neurologists(authors D M and M H) by tracing by hand theboundary of the lesion onto every single 1-mm-thickaxial slice using a graphics tablet (WACOM Intuos A6)Combining all the slices then led to a 3-D region of in-terest (ROI) for each patient For one patient (Case 15)there were two areas of discrete focal damage one inthe frontal cortex and the other in the parietal cortex
Normalization of each MRI including the 3-D ROI tothe T1-template was performed with SPM99 (wwwfilionuclacukspm) The ROI was used as a mask for thelesion during determination of alignment parameters tominimize the contribution of abnormal brain to thisprocess (Rorden amp Brett 2000) The normalized ROIof each neglect patient was superimposed upon the T1template to find areas of maximal lesion overlap in eachgroup (see Results) as well as lesion volume usingMRIcro shareware (Rorden amp Brett 2000)
Experimental Procedure
Stimuli were presented on an SVGA monitor (29 22 in)viewed at 57 cm distance Each search display com-prised a virtual 9 7 grid each cell containing a single18 elementmdasheither a letter T (target) or a letter L (dis-tractor) (see Figure 1) The position of each elementwithin each cell was jittered with 19 targets and 44distractors randomly distributed over the virtual gridThe number of targets on the right and left halves ofthe display was balanced across trials Seven neglectpatients (Cases 5 8 10 11 14ndash16) were further testedon lsquolsquoTarget onlyrsquorsquo displays which had varying numbersof Ts (2 6 or 19) now with no L-shaped distractorsThe presentations of these lsquolsquoTarget onlyrsquorsquo screens wereinterspersed with those of the original search screens inrandom order in the same experimental session
Mannan et al 351
Eye movements were recorded using a video-basedpupil tracker with a temporal resolution of 4 msec andspatial accuracy of lt058 (EyeLink Sensorimotoric Sys-tems Berlin Germany) Subjects were asked to keeptheir head still The experimenter also monitored gazeposition on-line At the beginning of each session (andafter breaks in testing or following any unintended headmovement that disrupted gaze recordings) eye-trackingwas re-calibrated Each trial was initiated by the exper-imenter and began with a central fixation cross for 1 secthen the search display
Subjects were instructed to search for Ts (among anyLs) Critically while looking at any target they had toclick a response button held in the right hand onlyif they considered this target to be a new discovery(ie a target they had not clicked on before) Clickingthe response button did not change the display Theresponse button was held by the patient in a comfort-able position to the right of the midline Subjectswere instructed and reminded throughout testing toclick on each target found once only Any later re-click on
a previously found target thus indicates a failure to retainthe fact that this target had already been discoveredThe experimenter terminated each search when thesubject indicated verbally that they felt all targets in thedisplay had now been found Each subject searched aminimum of 5 displays (mean = 7 range 5ndash14) Eyemovements were analyzed off-line using custom soft-ware An automatic algorithm was used to extract sac-cades (saccade onset defined as eye speed rising above258 sec1 for two or more samples and a criterion ofminimum fixation duration of 60 msec) Blinks wereeliminated from the trace by visual inspection and sac-cade and fixation delineation were checked visually inthe record Next fixations were automatically assignedto items in the display if the mean position fell within18 of that item Two or more consecutive fixations onthe same item were re-scored as a single extendedfixation A re-fixation was scored only when at leastone fixation elsewhere on another item intervened
Most click responses were made during a fixation andthe few button-clicks occurring during a saccade were
Table 1 Patient Details
CaseAge
(years) PathologyVolume(cm3)
Age ofLesion (days)
Visual FieldDefect
Mean Deviationfrom Center on
Line Bisection Task(cm) (rightward +)
Mesulum Score(Maximum Possible
Score = 60) Lesion Type
1 68 Infarct 1282 64 + 867 17 PCA
2 76 Infarct 1528 227 + 003 41 PCA
3 68 Infarct 79 207 + 297 36 PCA
4 77 Infarct 251 158 + 150 54 PCA
5 63 Infarct 28 29 033 55 MCA
6 59 Hemorrhage 172 7 + 32 6 MCA
7 66 Hemorrhage 273 325 + 213 31 MCA
8 68 Infarct 309 3 25 17 MCA
9 29 Infarct 115 17 113 55 MCA
10 70 Infarct 1339 324 153 57 MCA
11 42 Infarct ndash 11 097 24 MCA
12 75 Infarct 757 38 013 4 MCA
13 51 Infarct 694 141 083 31 MCA
14 59 Infarct 1144 356 + 223 37 MCA
15 80 Infarct 261 44 35 6 MCA
16 64 Infarct ndash 30 033 55 MCA
MEAN 63 66 124 188 33
SE 4 13 32 5
Cases 1ndash9 had damage to the nonfrontal (posterior only) cortex whereas Cases 10ndash15 had both frontal and posterior (posterior and anterior) corticalinvolvement Case 16 is a patient with a subcortical lesion All but 3 of the patients with right MCA infarction had intact visual fields on confrontationTwo of these (Cases 7 and 14) were diagnosed with left homonymous hemianopia whereas one (Case 6) had a left inferior quandrantic field defectAll patients with PCA territory damage had partial or complete contralesional visual field loss No patient suffered from dysphasia
352 Journal of Cognitive Neuroscience Volume 17 Number 2
assigned to the previous fixation Clicks were assigned toa particular item if eye position at the time of the clickwas within 28 of that item or in the cases where morethan one item was within 28 of current eye position theclick response was simply attributed to the item closestto that eye position
Button-clicks were assigned to a particular item if eyeposition at the time of the click was within 28 of thatitem A re-click on an item required at least one fixationon another item between successive clicks (in practiceseveral fixations elsewhere typically intervened betweeninitial click and later re-click see Results)
Statistical analyses were performed using SPSS andSigmaplot (SPSS Science Chicago USA) Jonckheere sta-tistic was derived by hand Statistical significance was setas p lt 05
Acknowledgments
We thank the reviewers of this manuscript and Dr AndrewParton for their extremely helpful comments We thank thepatients who participated Dr J Wade Dr A Rudd and thestaff of the stroke units at Charing Cross and St ThomasrsquoHospitals This research was supported by grants from theWellcome Trust to M H C K and J D J D holds a RoyalSociety Wolfson Research Merit Award
Reprint requests should be sent to Drs Sabira Mannan andMasud Husain Division of Neuroscience and PsychologicalMedicine Imperial College London Charing Cross CampusLondon W6 8RF UK or via e-mail smannanimperialacukor mhusainimperialacuk
Notes
1 We are grateful to one of the reviewers for suggesting thistime-series analysis2 There was no significant difference between the number oftargets fixated between the two groups so differences in lsquolsquoclickomissionsrsquorsquo cannot be attributed to the number of fixatedtargets that had to be kept track of during search3 The correlation between these four points alone inFigure 9A is high [r(3) = 81 cf r(15) = 47 for the wholegroup]4 We have also tested 10 neglect patients and 10 controlstroke patients on a vertical version of the Corsi blocks task(clinically used to measure spatial working memory capacity)Neglect patients had significantly impaired capacity but wereable to localize single locations well suggesting that mislocal-ization is unlikely to account for their deficit on this spatialworking memory task
REFERENCES
Bartolomeo P Sieroff E Decaix C amp Chokron S (2001)Modulating the attentional bias in unilateral neglect theeffects of the strategic set Experimental Brain Research137 432ndash444
Bates E Wilson S M Saygin A P Dick F Sereno M IKnight R T amp Dronkers N F (2003) Voxel-basedlesion-symptom mapping Nature Neuroscience 6 448ndash450
Behrmann M Watt S Black S E amp Barton J J (1997)Impaired visual search in patients with unilateral neglect
An oculographic analysis Neuropsychologia 351445ndash1458
Bisiach E amp Vallar G (2000) Unilateral neglect in humans InG Rizzolatti (Ed) Handbook of neuropsychology (2nd edvol 1 pp 459ndash502) Amsterdam Elsevier
Cals N Devuyst G Afsar N Karapanayiotides T ampBogousslavsky J (2002) Pure superficial posterior cerebralartery territory infarction in The Lausanne Stroke RegistryJournal of Neurology 249 855ndash861
Chafee M amp Goldman-Rakic P (1998) Matching patterns ofactivity in primate prefrontal area 8a and parietal area 7ipneurons during a spatial working memory task Journal ofNeurophysiology 79 2919ndash2940
Chedru F Leblanc M amp Lhermitte F (1973) Visualsearching in normal and brain-damaged subjectsContributions to the study of unilateral subjectsCortex 9 94ndash111
DrsquoEsposito M Aguirre G K Zarahn E Ballard DShin R K amp Lease J (1998) Functional MRI studies ofspatial and nonspatial working memory Cognitive BrainResearch 7 1ndash13
Donkelaar P amp Muri R (2002) Craniotopic updating ofvisual space across saccades in the human posteriorparietal cortex Proceedings of the Royal Society London B269 735ndash739
Driver J amp Husain M (2002) The role of spatial workingmemory deficits in pathological search by neglect patientsIn the cognitive and neural bases of spatial neglect InH-O Karnath A D Milner amp G Vallar (Eds) The cognitiveand neural bases of spatial neglect Oxford OxfordUniversity Press
Duhamel J R Colby C amp Goldberg M E (1992) Theupdating of the representation of visual space in parietalcortex by intended eye movements Science 25590ndash92
Goldman-Rakic P S (1996) The prefrontal landscapemdashImplications of functional architecture for understandinghuman mentation and the central executive PhilosophicalTransactions of The Royal Society of London Series BBiological Sciences 351 1445ndash1453
Heide W Binkofski F Seitz R J Posse S Nitschke M FFreund H J amp Kompf D (2001) Activation offrontoparietal cortices during memorized triple-stepsequences of saccadic eye movements An fMRI studyEuropean Journal of Neuroscience 13 1177ndash1189
Heide W amp Kompf D (1998) Combined deficits ofsaccades and visuo-spatial orientation after cortical lesionsExperimental Brain Research 123 164ndash171
Husain M amp Kennard C (1996) Visual neglect associatedwith frontal lobe infarction Journal of Neurology 243652ndash657
Husain M Mannan S Hodgson T Wojciulik E Driver Jamp Kennard C (2001) Impaired spatial working memoryacross saccades contributes to abnormal search in parietalneglect Brain 121 941ndash952
Husain M amp Rorden C (2003) Non-spatially lateralizedmechanisms in hemispatial neglect Nature ReviewsNeuroscience 4 23ndash36
Jackson S R amp Husain M (1996) Visuomotor functionsof the lateral pre-motor cortex Current Opinion inNeurobiology 6 788ndash795
Kinsbourne M (1993) Orientational bias model of unilateralneglect Evidence from attentional gradients withinhemispace In I H Robertson amp J C Marshall (Eds)Unilateral neglect Clinical and experimental studies(pp 63ndash86) Hove Erlbaum
Konishi S Kyoichi N Uchida I Kikyo H Kameyama Mamp Miyauchi S (1999) Common inhibitory mechanism in
Mannan et al 353
human inferior prefrontal cortex revealed by event-relatedfunctional MRI Brain 122 981ndash991
Konishi S Kyoichi N Uchida I Sekihara K amp Miyashita Y(1998) No-go dominant brain activity in human inferiorprefrontal cortex revealed by functional magneticresonance imaging European Journal of Neuroscience10 1209ndash1213
Malhotra P Mannan S Driver J amp Husain M (2004)Impaired spatial working memory One component of thevisual neglect syndrome Cortex 40 667ndash676
Manly T Woldt K Watson P amp Warburton E (2002) Ismotor perseveration in unilateral neglect lsquolsquodrivenrsquorsquo by thepresence of neglected left-sided stimuli Neuropsychologia40 1794ndash1803
Mazzoni P Bracewell R Barash S amp Andersen R (1996)Spatially tuned auditory responses in area LIP of macaquesperforming delayed memory saccades to acoustic targetsJournal of Neurophysiology 75 1233ndash1241
McCarley J Wang R Kramer A Irwin D amp Peterson M(2003) How much memory does oculomotor search havePsychological Science 14 422ndash426
Mesulam M-M (1985) Principles of behavioural neurologyTests of directed attention and memory Philadelphia Davis
Mesulam M M (1999) Spatial attention and neglectParietal frontal and cingulate contributions to the mentalrepresentation and attentional targeting of salientextrapersonal events Philosophical Transactions of theRoyal Society of London Series B Biological Sciences354 1325ndash1346
Mort D Malhotra P Mannan S Rorden C Pambakian AKennard C amp Husain M (2003) The anatomy of visualneglect Brain 126 1986ndash1997
Na D L Adair J C Kang Y Chung C S Lee K H ampHeilman K M (1999) Motor perseverative behaviour ona line cancellation task Neurology 52 1569ndash1576
Pearson D G amp Sahraie A (2003) Oculomotor controland the maintenance of spatially and temporally distributedevents in visuo-spatial working memory QuarterlyJournal of Experimental Psychology 56A 1089ndash1111
Rafal R (1994) Neglect Current Opinion in Neurobiology4 231ndash236
Rizzolatti G amp Arbib M A (1998) Language within our graspTrends in Neurosciences 21 188ndash194
Robertson L C Lamb M R amp Knight R T (1988) Effectsof lesions of temporalndashparietal junction on perceptual andattentional processing in humans Journal of Neuroscience8 3757ndash3769
Rorden C amp Brett M (2000) Stereotaxic display of brainlesions Behavioural Neurology 12 191ndash200
Rusconi M L Maravita A Bottini G amp Vallar G (2002)Is the intact side really intact Perseverative responses inpatients with unilateral neglect A productive manifestationNeuropsychologia 40 594ndash604
Russell C Malhotra P amp Husain M (2004) Attentionmodulates the visual field in healthy observers and parietalpatients NeuroReport 15 189ndash193
Rypma B amp DrsquoEsposito M (2003) A subsequent-memoryeffect in dorsolateral prefrontal cortex Cognitive BrainResearch 16 162ndash166
Sapir A Hayes A Henik A Danzinger S amp Rafal R(2004) Parietal lobe lesions disrupt saccadic remapping ofinhibitory location tagging Journal of CognitiveNeuroscience 16 503ndash509
Smania N Martini M C Gambina G A T Palamara ANatale E amp Marzi C A (1998) The spatial distributionof visual attention in hemineglect and extinction patientsBrain 121 1759ndash1770
Sprenger A Kompf D amp Heide W (2002) Visual searchin patients with left visual hemineglect Progress in BrainResearch 140 395ndash416
Tobler P Felblinger J Burki M Nirkko A Ozdoba C ampMuri R (2001) Functional organisation of the saccadicreference system processing extraretinal signals in humansVision Research 41 1351ndash1358
Wilson B Cockburn J amp Halligan P W (1987)Behavioural inattention test Bury St EdmundsThames Valley
Wojciulik E Husain M Clarke K amp Driver J (2001)Spatial working memory deficit in unilateral neglectNeuropsychologia 39 390ndash396
Zihl J amp Hebel N (1997) Patterns of oculomotor scanningin patients with unilateral posterior parietal and frontallobe damage Neuropsychologia 35 893ndash906
Zihl J amp von Cramon D (1985) Visual field recovery fromscotoma in patients with postgeniculate damage A reviewof 55 cases Brain 108 335ndash365
354 Journal of Cognitive Neuroscience Volume 17 Number 2
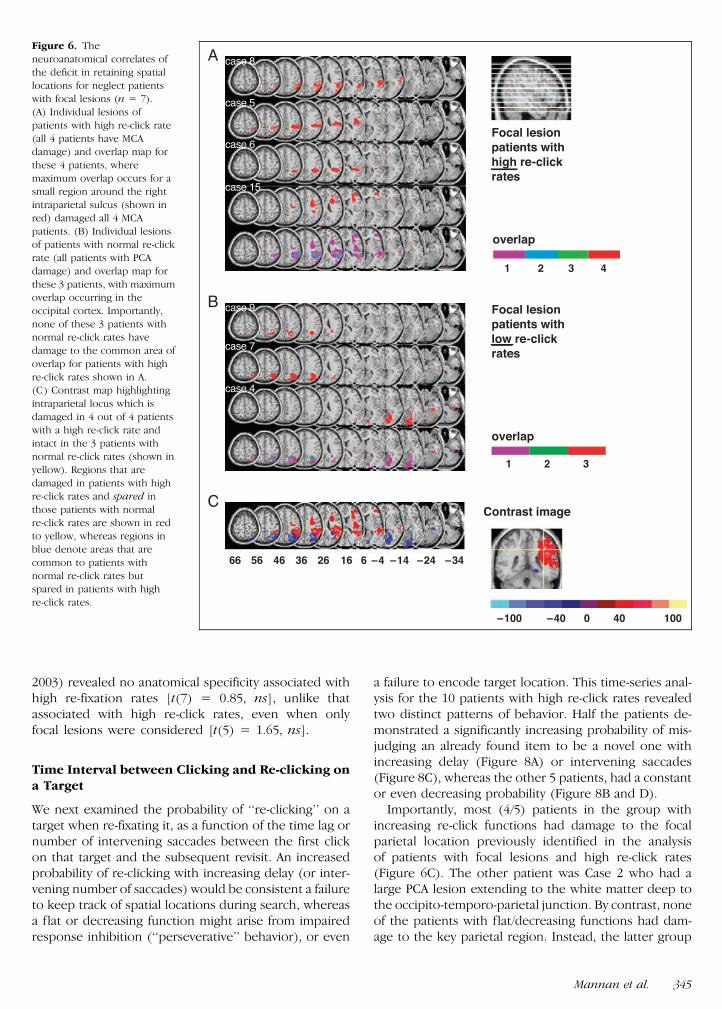
2003) revealed no anatomical specificity associated withhigh re-fixation rates [t(7) = 085 ns] unlike thatassociated with high re-click rates even when onlyfocal lesions were considered [t(5) = 165 ns]
Time Interval between Clicking and Re-clicking ona Target
We next examined the probability of lsquolsquore-clickingrsquorsquo on atarget when re-fixating it as a function of the time lag ornumber of intervening saccades between the first clickon that target and the subsequent revisit An increasedprobability of re-clicking with increasing delay (or inter-vening number of saccades) would be consistent a failureto keep track of spatial locations during search whereasa flat or decreasing function might arise from impairedresponse inhibition (lsquolsquoperseverativersquorsquo behavior) or even
a failure to encode target location This time-series anal-ysis for the 10 patients with high re-click rates revealedtwo distinct patterns of behavior Half the patients de-monstrated a significantly increasing probability of mis-judging an already found item to be a novel one withincreasing delay (Figure 8A) or intervening saccades(Figure 8C) whereas the other 5 patients had a constantor even decreasing probability (Figure 8B and D)
Importantly most (45) patients in the group withincreasing re-click functions had damage to the focalparietal location previously identified in the analysisof patients with focal lesions and high re-click rates(Figure 6C) The other patient was Case 2 who had alarge PCA lesion extending to the white matter deep tothe occipito-temporo-parietal junction By contrast noneof the patients with flatdecreasing functions had dam-age to the key parietal region Instead the latter group
Figure 6 The
neuroanatomical correlates of
the deficit in retaining spatial
locations for neglect patientswith focal lesions (n = 7)
(A) Individual lesions of
patients with high re-click rate
(all 4 patients have MCAdamage) and overlap map for
these 4 patients where
maximum overlap occurs for asmall region around the right
intraparietal sulcus (shown in
red) damaged all 4 MCA
patients (B) Individual lesionsof patients with normal re-click
rate (all patients with PCA
damage) and overlap map for
these 3 patients with maximumoverlap occurring in the
occipital cortex Importantly
none of these 3 patients withnormal re-click rates have
damage to the common area of
overlap for patients with high
re-click rates shown in A(C) Contrast map highlighting
intraparietal locus which is
damaged in 4 out of 4 patients
with a high re-click rate andintact in the 3 patients with
normal re-click rates (shown in
yellow) Regions that are
damaged in patients with highre-click rates and spared in
those patients with normal
re-click rates are shown in redto yellow whereas regions in
blue denote areas that are
common to patients with
normal re-click rates butspared in patients with high
re-click rates
Mannan et al 345
had frontal cortical or subcortical involvement Thusincreasing re-clicking with time interval was associatedwith parietal damage whereas flat re-click functionswere associated with frontalsubcortical lesions
For neglect patients as a whole the intervals betweenan initial click on a target and a subsequent re-click werelong (mean = 40 saccades) but much shorter betweenan initial fixation and a later re-fixation (mean = 5) withthis difference being significant [t(15) = 315 p lt01] This again emphasizes the dissociation betweenre-fixations and re-clicks and also demonstrates that re-clicking does not occur on most re-fixations
Relationship of Re-click Rate to Neglect Severity
We assessed three separate measures of neglect Thefirst was the difference between miss rates for left ver-sus right targets within the experimental search taskitself the other two measures of neglect were based onthe clinical pen-and-paper measures of Mesulam shape-cancellation (Mesulam 1985) and line bisection fromthe BIT (Wilson Cockburn amp Halligan 1987)
On the experimental search task there was a moderatebut significant correlation between re-click rate and ne-glect in search across the group [r(15) = 47 p lt 05
Figure 9A] Thus the greater the re-clicking on right-ward items the greater the severity of leftward neglectThere was also a moderate but again significant corre-lation between re-click rates and the severity of individualpatientsrsquo clinical neglect as indexed by the cancellationmeasure [r(15) = 46 p lt 05 Figure 9B] Thesecorrelations were driven largely by three of the patientsidentified in our focal lesion analysis (Figure 6A) withinvolvement of the key parietal region and high re-clicking behavior Their data are shown in filled circlesto the right in the graphs displayed in Figure 93 Thecorrelation between deviation on line bisection andre-click rate was not significant [r(15) = 13 ns] How-ever one would not predict that a deficit in retaininglocations already visited (or response inhibition) wouldnecessarily impair performance on a task such as linebisection
Effect of Varying Number of Targets
In 7 patients (Cases 5 8 10 11 14ndash16) we also exam-ined the effect of varying the number of targets in thesearch task Patients viewed lsquolsquotarget onlyrsquorsquo displays con-taining either 2 6 or 19 targets randomly distributedon the screen without any L-shaped distractors In-
Figure 7 Theneuroanatomical correlates of
the re-click deficit in neglect
patients with posterior plusanterior lesions (n = 5)
(A) Individual lesions of
patients with high re-click rate
(all 4 patients have MCAdamage) and overlap map for
these 4 patients where
maximum overlap occurs for
a region in the right inferiorfrontal lobe (shown in red)
damaged all 4 MCA patients
(B) Individual lesions of a single
patient with normal re-clickrate (C) Contrast map
highlighting the inferior frontal
locus (shown in yellow) whichis damaged in 4 out of
4 patients with a high re-click
rate and intact in the patient
with normal re-click rateRegions that are damaged in
patients with high re-click rates
and spared in the patient
with normal re-click rate areshown in red to yellow
whereas regions in blue denote
areas that are damaged in thepatient with normal re-click
rates but spared in patients with
high re-click rates
346 Journal of Cognitive Neuroscience Volume 17 Number 2
creasing the number of targets led to significantly higherre-click rates ( Jonckheere statistic = 133 p lt 01Figure 10) as well as more severe neglect (Jonckheerestatistic = 129 p lt 01) consistent with the proposalthat the mechanisms underlying pathological re-clickingmight contribute to neglect behavior Note that a lsquolsquoper-severativersquorsquo failure of response inhibition would not onits own easily explain this finding For the same absolutenumber of click responses one would predict a higherre-click rate with smaller set-sizes than with larger ones(see also Husain Mannan et al 2001 for a discussion)Although the number of subjects were small there wasno difference in the performance pattern between the3 patients who had increasing re-click rate functions inthe time-series analysis and the 4 patients with flat ordecreasing re-click rate functions
Varying Perceptual Load during Search
Can re-click behavior be explained simply by impairedvisual perception of targets from discriminators Neglectpatients did find it more difficult than control subjectsto select targets from distractors fixating on average19 distractors for every target found [cf 14 for controlsubjects t(29) = 485 p lt 0001] Patients also some-times clicked on distractors (64) but no more often
than controls (61) and it is possible that this rate iseffectively inflated by our strict criterion of considering aclick to be a response on the item that is being fixated atthe time of the click (It is likely that sometimes ob-servers click in response to targets they fixated one ormore saccades previously) Re-clicks on distractors wereextremely rare (06 cf 30 for targets)
To examine the effect of reduced perceptual load in8 patients (Cases 5 7 8ndash10 13 1 4 16) 6 of whom werehigh re-clickers we used search displays with salientcross (+) targets rather than Ts L distractors wereunchanged Patients fixated significantly fewer distrac-tors when searching for cross (+) targets [distractortarget ratio of 134 cf 181 for T targets t(7) = 60 p lt001] suggesting they did indeed find targetndashdistractordiscrimination easier Importantly however there wasno significant difference in re-click rate [26 for +targets cf 27 for T targets t(14) = 041 ns] or evenin re-fixation rate [100 for + targets cf 119 fort(14) = 041 ns] Thus these changes in perceptualload did not influence re-click performance
DISCUSSION
On this multitarget visual search task neglect patientsmissed targets to their left and also re-fixated targets
Figure 8 The probability of
re-clicking or re-fixating on
target as a function of delay
between the original click andsubsequent re-fixation (for
patients with high re-click
rates) (A) Probability functions
for patients with significantpositive slopes Four patients of
these patients have damage to
the focal intraparietal locusidentified in Figure 6C whereas
the single patient with a PCA
lesion and no damage to the
critical parietal locationindicated by symbol has only
a very shallow positive slope
(B) Probability functions for
patients with no significantpositive slope None of these
5 patients have damage to the
critical intraparietal region (C)As for A but for delay measured
in terms of intervening
fixations (D) As for B but for
delay measured in terms ofintervening fixations
Mannan et al 347
to the right much more often than healthy individuals(Figure 2A) Critically 10 of the 16 neglect patients re-clicked on rightward targets at a significantly higher ratethan healthy control subjects (Figure 2B) For 5 of thesepatients the probability of re-clicking increased withtime (Figure 8A) whereas the other 5 neglect patientswith high re-click rates demonstrated a constant or de-creasing probability of re-clicking over time (Figure 8C)The remaining 6 neglect patients had re-click rateswithin the normal range demonstrating that neitherneglect nor brain damage per se leads to a pathologicalre-click rate
High-resolution MRI mapping of lesions revealed im-portant anatomical dissociations within the neglect pop-ulation Patients with pathological re-click rates hadlesions affecting either the right intraparietal sulcus orthe inferior frontal lobe the only exception (Case 2) hada large occipito-temporal lesion that extended deep intothe parietal white matter Neglect patients with normalre-click rates did not have damage to either of thecritical locations in the intraparietal sulcus or inferiorfrontal lobe (Figures 5 6 and 7) Instead they had moredorsal lesions (for MCA territory patients) or occipitaland medial-temporal damage (for PCA territory lesions)which can also result in neglect (Mort et al 2003Cals Devuyst Afsar Karapanayiotides amp Bogousslavsky2002) Importantly lesion location rather than the sizeor age of lesion was critical to the presence of the re-clicking impairment Across the group of neglect pa-tients there was a moderate but significant correlationbetween re-click rate and degree of neglect (Figure 9)driven largely by patients with parietal damage who hadboth a high re-click rate and also severe neglect consis-tent with the proposal that mechanisms underlying thisbehavior in parietal patients might have a functionallyimportant role in determining the severity of neglect
Role of Right Intraparietal Sulcus
Notably for all patients with damage to the critical locusin the right intraparietal sulcus (which may also involveparieto-frontal white matter fibers) the probability of re-clicking on a target increased with time (or interveningsaccades) since initial discovery (Figure 8) Because theyrarely re-clicked at short intervalsmdasheven though themajority of re-fixations occurred during this periodmdashthis pattern of behavior suggests that for these patientstarget locations were actually veridically perceived and
Figure 9 (A) Neglect on experimental search task (19 targets and44 distractors) plotted against mean re-click rate for individual neglect
patients Percentage neglect was calculated as the difference between
the number of targets missed on the left versus right expressed as apercentage of the total number of targets missed There was a
significant correlation between the degree of neglect and the re-click
rate [r(15) = 465 p lt 05] Patients with high re-click rates are
highlighted by black circles (for patients with damage to the focalparietal location identified in Figure 6C) and light gray circles (for
patients with frontal damage) while the single patient with PCA
damage and a high re-click rate is indicated by dark gray (B) The
number of targets missed on the Mesulam shape-cancellation taskplotted against mean re-click rate for individual neglect patients Again
there was a significant correlation between the number of targets
missed and the mean re-click rate [r(15) = 456 p lt 05] Patients withhigh re-click rates are denoted as in A
Figure 10 Mean re-click rate (filled circles) and degree of neglect
(unfilled squares) increased systematically with increasing numbers of
targets in target-only displays
348 Journal of Cognitive Neuroscience Volume 17 Number 2
well encoded into working memory Furthermore de-tailed single-case studies of two parietal patients withhigh re-click rates (Malhotra Mannan Driver amp Husain2004 Husain Mannan et al 2001) demonstrate usingdifferent psychophysical methods that these patientscan accurately encode spatial locations4 Increasing re-clicks with time (or intervening saccades) would how-ever be consistent with a deficit in keeping track oftarget locations during extended search
Several lines of evidence support the proposal thatthe critical intraparietal sulcus region we have identifiedmay have an important role in the spatial transforma-tions required to keep track of spatial locations acrosssaccades In rhesus monkeys neurons within the lateralbank of the intraparietal sulcus (area LIP) encode andlsquolsquomaintainrsquorsquo the remembered locations of saccadic tar-gets over time (Chafee amp Goldman-Rakic 1998 MazzoniBracewell Barash amp Andersen 1996) Some visuallyresponsive neurons in the LIP also appear to shift theirretinotopic representation at the onset of an intendedsaccade as if lsquolsquore-mappingrsquorsquo representations of objects inspace to update them across shifts of gaze (DuhamelColby amp Goldberg 1992)
In humans this computation has been investigatedusing the double-step saccade paradigm where twoflashes of light appear in quick succession such thatthe second target disappears before a saccade can bemade to the first target Patients with lesions involvingthe right parietal cortex are impaired at performing thistask suggesting that they are unable to update spatialrepresentations accurately after even one saccade (Heideamp Kompf 1998 Duhamel et al 1992) Similarly trans-cranial magnetic stimulation of the posterior parietalcortex prior to the second saccade reduces its accuracy(Donkelaar amp Muri 2002) and a recent study of patientswith parietal lesions also implicates this area in thespatial updating of inhibition of return after a saccade(Sapir Hayes Henik Danzinger amp Rafal 2004) Func-tional imaging studies of double-step (Tobler et al2001) and triple-step memory-guided saccades (HeideBinkofski et al 2001) have identified a region withinthe intraparietal sulcus (Talairach coordinates 40 4044 for Tobler et al and 44 48 36 Heide et al) This isclose to the critical region of lesion overlap we haveidentified in the posterior neglect patients with high re-click rates (Talairach 37 39 37)
We suggest that in our patients with lesions involvingthe right intraparietal sulcus uncertainty about locationsof previously found targets increases with each saccadeThis uncertainty is further exacerbated by interferencefrom newer target locations leading to increased prob-ability of misjudging old target locations for new assearch progresses Such progressively worsening uncer-tainty might be due to an accumulating impairment inlsquolsquore-mapping target locationsrsquorsquo across saccades or im-paired lsquolsquomemoryrsquorsquo of previously inspected target loca-tions across saccades As noted above there is evidence
for both types of signalmdash re-mapping and maintainingsaccadic spatial memorymdashin the parietal cortex and it ispossible that disruption to both these functions contrib-utes to the re-click deficit we have identified Certainlythe two potential types of deficit are not mutuallyincompatible
A slightly different account of the re-clicking behaviormight be framed in terms of the intriguing hypothesisproposed by Manly Woldt Watson and Warburton(2002) to explain visible re-marking on a standard can-cellation task They argue that such behavior is due topatients erroneously transposing leftward stimuli to theright resulting in apparent re-cancellation These au-thors found that removing targets from the left re-duced both neglect and re-marking However they didnot monitor eye movements so it is possible that sometargets that are logged as neglected (because they werenot cancelled) were nevertheless fixated and thereforeform part of the load on memory during search Fur-thermore in our experimental study subjects with ahigh re-click rate also failed to click on a higher pro-portion of fixated targets Manly et alrsquos proposal wouldnot easily account for such lsquolsquoclick omissionsrsquorsquo becausethese refer to fixated targets that were not actuallyresponded to
Finally it might be argued that increasing re-clickingwith search duration reflects a deficit in spatial workingmemory (SWM) that is independent of saccadic eyemovements (ie a decay in memory that is not trans-saccadic) Because time and number of interveningsaccades were not independently manipulated in thisunrestricted visual search paradigm we are not ablecategorically to distinguish between these possibilitiesWe note however that functional imaging studies ofSWM tasks that do not require eye movements oftendemonstrate activity related to the maintenance ofspatial information which is generally more posteriorand dorsal than the parietal locations isolated in thisstudy (DrsquoEsposito et al 1998) Moreover recent behav-ioral data indicate that SWM is more vulnerable tosaccadic eye movements than shifts in attention or limbmovements (Pearson amp Sahraie 2003)
Frontal Lesion Site Associated withHigh Re-click Rates
Four out of 5 neglect patients with lesions involvingposterior plus anterior damage also showed a high re-click rate and the area of damage common to thesepatients centered on the right inferior frontal lobe cor-responding to Brodmannrsquos area 44 (Figure 7C) Thisregion has previously been associated with frontalneglect (Husain amp Kennard 1996) and is consideredto be part of the ventral premotor cortex and thehomologue of Brocarsquos areas in the left hemisphere(Rizzolatti amp Arbib 1998 Jackson amp Husain 1996) Inthese frontal patients the probability of re-clicking did
Mannan et al 349
not increase with time interval between the first clickand re-fixation but rather it stayed constant or declinedwith increasing delays
Such behavior might be consistent with a failure toinhibit responses to rightward locationsmdasha type oflsquolsquoperseverativersquorsquo behavior Note though that this is notnecessarily the same type of perseverative responseas seen when some patients with neglect re-markpreviously found items on standard cancellation tests(Rusconi et al 2002 Na et al 1999) In those casestargets that are visibly cancelled are often marked againwhereas in our computerized search tasks no marks arevisibly left on targets once they have been clicked uponMoreover only one of our neglect patients (Case 1) de-monstrated re-cancellation of visibly cancelled targetson pen-and-paper tasks and then only on one occasionImportantly this patient actually had a normal re-clickrate (67) on our task and his stroke was in the PCAterritory sparing both the intraparietal sulcus and fron-tal lobe
In support of a role for the right frontal lobe in in-hibiting responses to previously found targets recentimaging studies of response inhibition have highlightedregions of the right inferior frontal gyrus (IFG) very closeto the centroid of damage for our frontal patients (38 1618 cf 43 22 21 Konishi Kyoichi Uchida Sekihara ampMiyashita 1998 and 41 16 19 Konishi Kyoichi UchidaKikyo et al 1999) Alternative explanations for the highprobability of re-clicking directly after clicking might bethat these patients have impaired perceptual discrimina-tion of targets impaired perceptual localization of targetsor that they encounter difficulty in encoding spatiallocations into working memory Our data are not sugges-tive of a perceptual discrimination problem as changingthe perceptual load did not inf luence re-click ratesMoreover the patients still had high re-click rates in thetarget only conditions (Figure 10) where no perceptualdiscrimination of targets and distractors was necessaryPoor perceptual localization would seem unlikely giventhat these patients have frontal lesions Furthermoreusing a vertical SWM task similar to the Corsi blocktapping task (Malhotra et al 2004) we have found thatpatients with neglect can localize and perceptually en-code single target locations as well as controls and aresignificantly impaired at remembering two or more loca-tions shown in sequence consistent with the presentobservations Some recent functional imaging data sug-gest that activation within the prefrontal cortex corre-lates well with successful encoding of material intoworking memory (Rypma amp DrsquoEsposito 2003) Furtherresearch is required to distinguish between these pos-sible explanations for the mechanisms underlying there-click deficit in frontal patients
The only patient with frontal damage who did re-clickmore frequently with increasing time had damage cen-tered on the dorsolateral prefrontal cortex (althoughalso encroaching on the IFG) Neurons in the dorsolat-
eral prefrontal cortex like those in the LIP encode theremembered locations of saccadic targets (Goldman-Rakic 1996) This patient (Case 15) who had the high-est re-click rate of all also had damage to the criticalparietal location described above (in contrast to all otherfrontal patients) Thus she may have suffered from acombination of underlying deficits
Re-fixations and Misjudging PreviouslyInspected Locations during Search
In this search task re-fixation and re-click rates weresignificantly correlated for neglect patients Howeverthere were important dissociations within individuals 2patients with PCA lesions had re-fixation rates of 220and 80 respectively but both had a re-click rate ofonly 7 These are clearly examples of individuals whodo not show pathological re-clicking despite having highre-fixation rates High re-fixation rates may result froma lack of integration of spatial information due to V1damage (Zihl amp von Cramon 1985) in our PCA patientsor possibly because of reduced inhibition of return(Bartolomeo Sieroff Decaix amp Chokron 2001) What-ever the exact cause for dissociations between re-fixa-tions and re-click rates our data suggest re-fixation doesnot necessarily index the same underlying deficits asrevealed by our re-click measure High re-fixation ratesunlike high re-click rates were also not significantlyassociated with damage to any specific cortical locationagain demonstrating a dissociation between re-fixationsand pathological re-clicking
The relationship between re-click rates and neglect isunlikely to be a simple one In our study there was amoderate but significant correlation between the twomeasures suggesting that this association is unlikely tohave arisen by chance However non-neglect patientscan also exhibit a high re-click rate (unpublished obser-vations from our laboratory) Taken together our resultssuggest that difficulties in keeping track of previouslyinspected targets (because of impaired trans-saccadic re-mapping andor memory) may be one contributingcomponent to visual neglect in patients with lesions in-volving the right intraparietal sulcus As mentioned thesearch task we used differed in one striking aspect fromstandard cancellation tests subjects do not visibly markthe targets they visit Whereas in cancellation tests pa-tients mark each target they find which might appear todispense with any need to remember which locationshave been searched before However it should be ap-preciated that in most standard cancellation tests theindividual marks made by patients may not be clearlyvisible in peripheral vision particularly if neglect pa-tients have a local spatial bias (Rafal 1994 RobertsonLamb amp Knight 1988) or a restricted field of view(Russell Malhotra amp Husain 2004) Thus when com-bined with a rightward attentional bias keeping track ofsearched locations may be critical to prevent gaze from
350 Journal of Cognitive Neuroscience Volume 17 Number 2
returning to rightward locations even if they have beenmarked before as the marks may not be visible untilgaze returns to them Consistent with this hypothesisneglect is greater when cancellation is performed usinginvisible marks compared to large and salient red marksthat can be seen peripherally (Wojciulik Husain Clarkeamp Driver 2001)
A task like the present search-and-click paradigm maymore closely resemble search in the real world than dotraditional cancellation tests When we examine a visualscene we do not leave marks on each object we ex-amine Instead just as in the search task used in thisstudy we have to keep track of the locations we visit ifwe are to avoid inefficient re-examination of thoselocations Neglect patients with damage to the rightintraparietal sulcus appear to be impaired at keepingtrack of searched locations in this way When combinedwith their rightward attentional bias we propose thatsuch an impairment leads to recursive search of stimulito the right in experimental search tasks as used hereand also in daily life where neglect is so apparentdespite unlimited viewing time
The paradigm we have used has the important virtuesof allowing patients to explore freelymdashas in real-worldsearch or clinical cancellation tasksmdashand relating theirre-click behavior directly to the severity of neglect in thesame task But it might be argued that it would be usefulalso to probe SWM in a more controlled manner by re-stricting observersrsquo eye movements to a specific set oflocations and probing memory of previously visitedplaces A recent study using a gaze-contingent para-digm has elegantly probed the capacity of spatial mem-ory during search in healthy observers and foundevidence for memory for up to four previous locations(McCarley Wang Kramer Irwin amp Peterson 2003)Variants of this paradigm may in the future be usefulto probe the capacity of SWM across saccades in patientswith neglect and also to probe further whether thereare differences between neglect patients with parietaland frontal lesions
METHODS
Subjects Imaging and Lesion Mapping
Sixteen patients with left neglect following right hemi-sphere infarction or hemorrhage were recruited Con-sent was obtained according to the Declaration ofHelsinki as approved by the hospital ethical commit-tees Presence of neglect was determined by clinicalobservation plus bedside tests including the Mesulamshape-cancellation task (Mesulam 1985) and line bisec-tion task (Wilson et al 1987) (see Table 1) Twelvepatients had MCA territory strokes the rest involvedonly the PCA territory which can also result in neglect(Cals et al 2002) Only one neglect patient (Case 1 whodid not have frontal involvement) demonstrated re-
cancellation of visibly cancelled targets on pen-and-paper tasks and then only on one occasion Fifteenage-matched healthy volunteers were controls (meanage = 63 SD = 14 vs 63 SD = 13 for patients)
All patients had clinical CT scans In addition dedi-cated high-resolution structural MRI scans were alsoobtained for 1416 neglect patients MRI scans wereunobtainable for the other two (Cases 11 and 16)because of claustrophobia or loss to follow up In theMCA neglect group median time between stroke andscan was 63 days with the shortest interval being35 days for the PCA neglect group one patient had tobe scanned relatively early at 9 days but the mediantime was 140 days MRI scans comprised one 256-sliceT1-weighted acquisition (MPRAGE TR 97 msec TE4 msec TA 12 min) performed in the coronal planealigned perpendicular to the anteriorndashposterior commis-sural line on a Siemens Vision 15-T scanner Within-and between-slice resolution was 1 mm For each pa-tient the area of damage was determined by detailedvisual inspection of the digital image for every singleslice Lesion delineation was performed by neurologists(authors D M and M H) by tracing by hand theboundary of the lesion onto every single 1-mm-thickaxial slice using a graphics tablet (WACOM Intuos A6)Combining all the slices then led to a 3-D region of in-terest (ROI) for each patient For one patient (Case 15)there were two areas of discrete focal damage one inthe frontal cortex and the other in the parietal cortex
Normalization of each MRI including the 3-D ROI tothe T1-template was performed with SPM99 (wwwfilionuclacukspm) The ROI was used as a mask for thelesion during determination of alignment parameters tominimize the contribution of abnormal brain to thisprocess (Rorden amp Brett 2000) The normalized ROIof each neglect patient was superimposed upon the T1template to find areas of maximal lesion overlap in eachgroup (see Results) as well as lesion volume usingMRIcro shareware (Rorden amp Brett 2000)
Experimental Procedure
Stimuli were presented on an SVGA monitor (29 22 in)viewed at 57 cm distance Each search display com-prised a virtual 9 7 grid each cell containing a single18 elementmdasheither a letter T (target) or a letter L (dis-tractor) (see Figure 1) The position of each elementwithin each cell was jittered with 19 targets and 44distractors randomly distributed over the virtual gridThe number of targets on the right and left halves ofthe display was balanced across trials Seven neglectpatients (Cases 5 8 10 11 14ndash16) were further testedon lsquolsquoTarget onlyrsquorsquo displays which had varying numbersof Ts (2 6 or 19) now with no L-shaped distractorsThe presentations of these lsquolsquoTarget onlyrsquorsquo screens wereinterspersed with those of the original search screens inrandom order in the same experimental session
Mannan et al 351
Eye movements were recorded using a video-basedpupil tracker with a temporal resolution of 4 msec andspatial accuracy of lt058 (EyeLink Sensorimotoric Sys-tems Berlin Germany) Subjects were asked to keeptheir head still The experimenter also monitored gazeposition on-line At the beginning of each session (andafter breaks in testing or following any unintended headmovement that disrupted gaze recordings) eye-trackingwas re-calibrated Each trial was initiated by the exper-imenter and began with a central fixation cross for 1 secthen the search display
Subjects were instructed to search for Ts (among anyLs) Critically while looking at any target they had toclick a response button held in the right hand onlyif they considered this target to be a new discovery(ie a target they had not clicked on before) Clickingthe response button did not change the display Theresponse button was held by the patient in a comfort-able position to the right of the midline Subjectswere instructed and reminded throughout testing toclick on each target found once only Any later re-click on
a previously found target thus indicates a failure to retainthe fact that this target had already been discoveredThe experimenter terminated each search when thesubject indicated verbally that they felt all targets in thedisplay had now been found Each subject searched aminimum of 5 displays (mean = 7 range 5ndash14) Eyemovements were analyzed off-line using custom soft-ware An automatic algorithm was used to extract sac-cades (saccade onset defined as eye speed rising above258 sec1 for two or more samples and a criterion ofminimum fixation duration of 60 msec) Blinks wereeliminated from the trace by visual inspection and sac-cade and fixation delineation were checked visually inthe record Next fixations were automatically assignedto items in the display if the mean position fell within18 of that item Two or more consecutive fixations onthe same item were re-scored as a single extendedfixation A re-fixation was scored only when at leastone fixation elsewhere on another item intervened
Most click responses were made during a fixation andthe few button-clicks occurring during a saccade were
Table 1 Patient Details
CaseAge
(years) PathologyVolume(cm3)
Age ofLesion (days)
Visual FieldDefect
Mean Deviationfrom Center on
Line Bisection Task(cm) (rightward +)
Mesulum Score(Maximum Possible
Score = 60) Lesion Type
1 68 Infarct 1282 64 + 867 17 PCA
2 76 Infarct 1528 227 + 003 41 PCA
3 68 Infarct 79 207 + 297 36 PCA
4 77 Infarct 251 158 + 150 54 PCA
5 63 Infarct 28 29 033 55 MCA
6 59 Hemorrhage 172 7 + 32 6 MCA
7 66 Hemorrhage 273 325 + 213 31 MCA
8 68 Infarct 309 3 25 17 MCA
9 29 Infarct 115 17 113 55 MCA
10 70 Infarct 1339 324 153 57 MCA
11 42 Infarct ndash 11 097 24 MCA
12 75 Infarct 757 38 013 4 MCA
13 51 Infarct 694 141 083 31 MCA
14 59 Infarct 1144 356 + 223 37 MCA
15 80 Infarct 261 44 35 6 MCA
16 64 Infarct ndash 30 033 55 MCA
MEAN 63 66 124 188 33
SE 4 13 32 5
Cases 1ndash9 had damage to the nonfrontal (posterior only) cortex whereas Cases 10ndash15 had both frontal and posterior (posterior and anterior) corticalinvolvement Case 16 is a patient with a subcortical lesion All but 3 of the patients with right MCA infarction had intact visual fields on confrontationTwo of these (Cases 7 and 14) were diagnosed with left homonymous hemianopia whereas one (Case 6) had a left inferior quandrantic field defectAll patients with PCA territory damage had partial or complete contralesional visual field loss No patient suffered from dysphasia
352 Journal of Cognitive Neuroscience Volume 17 Number 2
assigned to the previous fixation Clicks were assigned toa particular item if eye position at the time of the clickwas within 28 of that item or in the cases where morethan one item was within 28 of current eye position theclick response was simply attributed to the item closestto that eye position
Button-clicks were assigned to a particular item if eyeposition at the time of the click was within 28 of thatitem A re-click on an item required at least one fixationon another item between successive clicks (in practiceseveral fixations elsewhere typically intervened betweeninitial click and later re-click see Results)
Statistical analyses were performed using SPSS andSigmaplot (SPSS Science Chicago USA) Jonckheere sta-tistic was derived by hand Statistical significance was setas p lt 05
Acknowledgments
We thank the reviewers of this manuscript and Dr AndrewParton for their extremely helpful comments We thank thepatients who participated Dr J Wade Dr A Rudd and thestaff of the stroke units at Charing Cross and St ThomasrsquoHospitals This research was supported by grants from theWellcome Trust to M H C K and J D J D holds a RoyalSociety Wolfson Research Merit Award
Reprint requests should be sent to Drs Sabira Mannan andMasud Husain Division of Neuroscience and PsychologicalMedicine Imperial College London Charing Cross CampusLondon W6 8RF UK or via e-mail smannanimperialacukor mhusainimperialacuk
Notes
1 We are grateful to one of the reviewers for suggesting thistime-series analysis2 There was no significant difference between the number oftargets fixated between the two groups so differences in lsquolsquoclickomissionsrsquorsquo cannot be attributed to the number of fixatedtargets that had to be kept track of during search3 The correlation between these four points alone inFigure 9A is high [r(3) = 81 cf r(15) = 47 for the wholegroup]4 We have also tested 10 neglect patients and 10 controlstroke patients on a vertical version of the Corsi blocks task(clinically used to measure spatial working memory capacity)Neglect patients had significantly impaired capacity but wereable to localize single locations well suggesting that mislocal-ization is unlikely to account for their deficit on this spatialworking memory task
REFERENCES
Bartolomeo P Sieroff E Decaix C amp Chokron S (2001)Modulating the attentional bias in unilateral neglect theeffects of the strategic set Experimental Brain Research137 432ndash444
Bates E Wilson S M Saygin A P Dick F Sereno M IKnight R T amp Dronkers N F (2003) Voxel-basedlesion-symptom mapping Nature Neuroscience 6 448ndash450
Behrmann M Watt S Black S E amp Barton J J (1997)Impaired visual search in patients with unilateral neglect
An oculographic analysis Neuropsychologia 351445ndash1458
Bisiach E amp Vallar G (2000) Unilateral neglect in humans InG Rizzolatti (Ed) Handbook of neuropsychology (2nd edvol 1 pp 459ndash502) Amsterdam Elsevier
Cals N Devuyst G Afsar N Karapanayiotides T ampBogousslavsky J (2002) Pure superficial posterior cerebralartery territory infarction in The Lausanne Stroke RegistryJournal of Neurology 249 855ndash861
Chafee M amp Goldman-Rakic P (1998) Matching patterns ofactivity in primate prefrontal area 8a and parietal area 7ipneurons during a spatial working memory task Journal ofNeurophysiology 79 2919ndash2940
Chedru F Leblanc M amp Lhermitte F (1973) Visualsearching in normal and brain-damaged subjectsContributions to the study of unilateral subjectsCortex 9 94ndash111
DrsquoEsposito M Aguirre G K Zarahn E Ballard DShin R K amp Lease J (1998) Functional MRI studies ofspatial and nonspatial working memory Cognitive BrainResearch 7 1ndash13
Donkelaar P amp Muri R (2002) Craniotopic updating ofvisual space across saccades in the human posteriorparietal cortex Proceedings of the Royal Society London B269 735ndash739
Driver J amp Husain M (2002) The role of spatial workingmemory deficits in pathological search by neglect patientsIn the cognitive and neural bases of spatial neglect InH-O Karnath A D Milner amp G Vallar (Eds) The cognitiveand neural bases of spatial neglect Oxford OxfordUniversity Press
Duhamel J R Colby C amp Goldberg M E (1992) Theupdating of the representation of visual space in parietalcortex by intended eye movements Science 25590ndash92
Goldman-Rakic P S (1996) The prefrontal landscapemdashImplications of functional architecture for understandinghuman mentation and the central executive PhilosophicalTransactions of The Royal Society of London Series BBiological Sciences 351 1445ndash1453
Heide W Binkofski F Seitz R J Posse S Nitschke M FFreund H J amp Kompf D (2001) Activation offrontoparietal cortices during memorized triple-stepsequences of saccadic eye movements An fMRI studyEuropean Journal of Neuroscience 13 1177ndash1189
Heide W amp Kompf D (1998) Combined deficits ofsaccades and visuo-spatial orientation after cortical lesionsExperimental Brain Research 123 164ndash171
Husain M amp Kennard C (1996) Visual neglect associatedwith frontal lobe infarction Journal of Neurology 243652ndash657
Husain M Mannan S Hodgson T Wojciulik E Driver Jamp Kennard C (2001) Impaired spatial working memoryacross saccades contributes to abnormal search in parietalneglect Brain 121 941ndash952
Husain M amp Rorden C (2003) Non-spatially lateralizedmechanisms in hemispatial neglect Nature ReviewsNeuroscience 4 23ndash36
Jackson S R amp Husain M (1996) Visuomotor functionsof the lateral pre-motor cortex Current Opinion inNeurobiology 6 788ndash795
Kinsbourne M (1993) Orientational bias model of unilateralneglect Evidence from attentional gradients withinhemispace In I H Robertson amp J C Marshall (Eds)Unilateral neglect Clinical and experimental studies(pp 63ndash86) Hove Erlbaum
Konishi S Kyoichi N Uchida I Kikyo H Kameyama Mamp Miyauchi S (1999) Common inhibitory mechanism in
Mannan et al 353
human inferior prefrontal cortex revealed by event-relatedfunctional MRI Brain 122 981ndash991
Konishi S Kyoichi N Uchida I Sekihara K amp Miyashita Y(1998) No-go dominant brain activity in human inferiorprefrontal cortex revealed by functional magneticresonance imaging European Journal of Neuroscience10 1209ndash1213
Malhotra P Mannan S Driver J amp Husain M (2004)Impaired spatial working memory One component of thevisual neglect syndrome Cortex 40 667ndash676
Manly T Woldt K Watson P amp Warburton E (2002) Ismotor perseveration in unilateral neglect lsquolsquodrivenrsquorsquo by thepresence of neglected left-sided stimuli Neuropsychologia40 1794ndash1803
Mazzoni P Bracewell R Barash S amp Andersen R (1996)Spatially tuned auditory responses in area LIP of macaquesperforming delayed memory saccades to acoustic targetsJournal of Neurophysiology 75 1233ndash1241
McCarley J Wang R Kramer A Irwin D amp Peterson M(2003) How much memory does oculomotor search havePsychological Science 14 422ndash426
Mesulam M-M (1985) Principles of behavioural neurologyTests of directed attention and memory Philadelphia Davis
Mesulam M M (1999) Spatial attention and neglectParietal frontal and cingulate contributions to the mentalrepresentation and attentional targeting of salientextrapersonal events Philosophical Transactions of theRoyal Society of London Series B Biological Sciences354 1325ndash1346
Mort D Malhotra P Mannan S Rorden C Pambakian AKennard C amp Husain M (2003) The anatomy of visualneglect Brain 126 1986ndash1997
Na D L Adair J C Kang Y Chung C S Lee K H ampHeilman K M (1999) Motor perseverative behaviour ona line cancellation task Neurology 52 1569ndash1576
Pearson D G amp Sahraie A (2003) Oculomotor controland the maintenance of spatially and temporally distributedevents in visuo-spatial working memory QuarterlyJournal of Experimental Psychology 56A 1089ndash1111
Rafal R (1994) Neglect Current Opinion in Neurobiology4 231ndash236
Rizzolatti G amp Arbib M A (1998) Language within our graspTrends in Neurosciences 21 188ndash194
Robertson L C Lamb M R amp Knight R T (1988) Effectsof lesions of temporalndashparietal junction on perceptual andattentional processing in humans Journal of Neuroscience8 3757ndash3769
Rorden C amp Brett M (2000) Stereotaxic display of brainlesions Behavioural Neurology 12 191ndash200
Rusconi M L Maravita A Bottini G amp Vallar G (2002)Is the intact side really intact Perseverative responses inpatients with unilateral neglect A productive manifestationNeuropsychologia 40 594ndash604
Russell C Malhotra P amp Husain M (2004) Attentionmodulates the visual field in healthy observers and parietalpatients NeuroReport 15 189ndash193
Rypma B amp DrsquoEsposito M (2003) A subsequent-memoryeffect in dorsolateral prefrontal cortex Cognitive BrainResearch 16 162ndash166
Sapir A Hayes A Henik A Danzinger S amp Rafal R(2004) Parietal lobe lesions disrupt saccadic remapping ofinhibitory location tagging Journal of CognitiveNeuroscience 16 503ndash509
Smania N Martini M C Gambina G A T Palamara ANatale E amp Marzi C A (1998) The spatial distributionof visual attention in hemineglect and extinction patientsBrain 121 1759ndash1770
Sprenger A Kompf D amp Heide W (2002) Visual searchin patients with left visual hemineglect Progress in BrainResearch 140 395ndash416
Tobler P Felblinger J Burki M Nirkko A Ozdoba C ampMuri R (2001) Functional organisation of the saccadicreference system processing extraretinal signals in humansVision Research 41 1351ndash1358
Wilson B Cockburn J amp Halligan P W (1987)Behavioural inattention test Bury St EdmundsThames Valley
Wojciulik E Husain M Clarke K amp Driver J (2001)Spatial working memory deficit in unilateral neglectNeuropsychologia 39 390ndash396
Zihl J amp Hebel N (1997) Patterns of oculomotor scanningin patients with unilateral posterior parietal and frontallobe damage Neuropsychologia 35 893ndash906
Zihl J amp von Cramon D (1985) Visual field recovery fromscotoma in patients with postgeniculate damage A reviewof 55 cases Brain 108 335ndash365
354 Journal of Cognitive Neuroscience Volume 17 Number 2
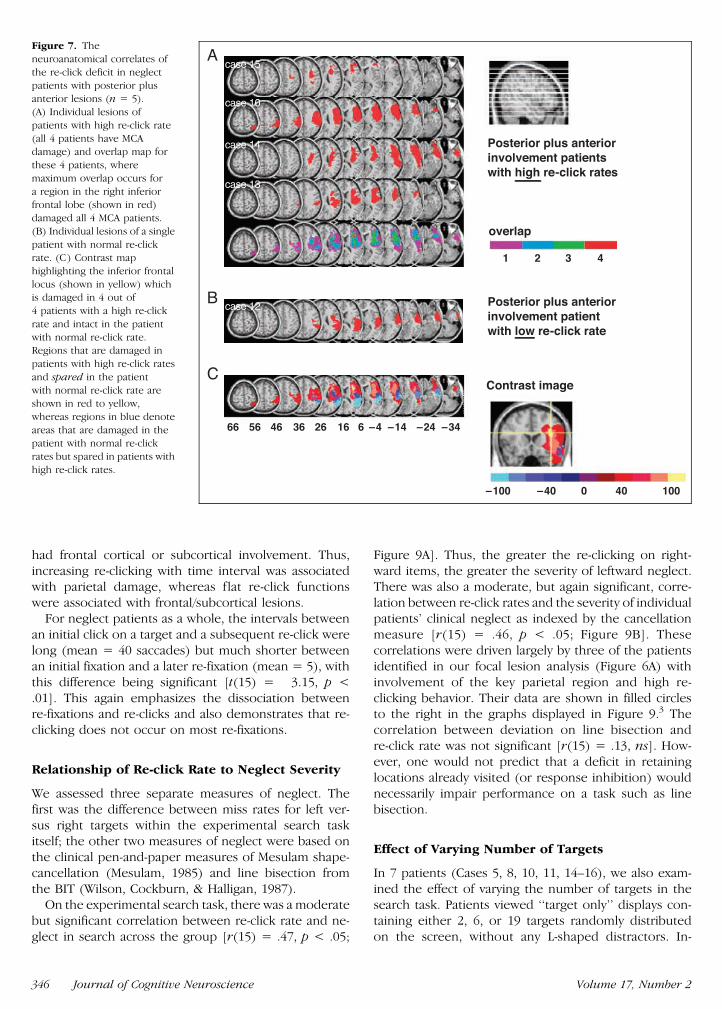
had frontal cortical or subcortical involvement Thusincreasing re-clicking with time interval was associatedwith parietal damage whereas flat re-click functionswere associated with frontalsubcortical lesions
For neglect patients as a whole the intervals betweenan initial click on a target and a subsequent re-click werelong (mean = 40 saccades) but much shorter betweenan initial fixation and a later re-fixation (mean = 5) withthis difference being significant [t(15) = 315 p lt01] This again emphasizes the dissociation betweenre-fixations and re-clicks and also demonstrates that re-clicking does not occur on most re-fixations
Relationship of Re-click Rate to Neglect Severity
We assessed three separate measures of neglect Thefirst was the difference between miss rates for left ver-sus right targets within the experimental search taskitself the other two measures of neglect were based onthe clinical pen-and-paper measures of Mesulam shape-cancellation (Mesulam 1985) and line bisection fromthe BIT (Wilson Cockburn amp Halligan 1987)
On the experimental search task there was a moderatebut significant correlation between re-click rate and ne-glect in search across the group [r(15) = 47 p lt 05
Figure 9A] Thus the greater the re-clicking on right-ward items the greater the severity of leftward neglectThere was also a moderate but again significant corre-lation between re-click rates and the severity of individualpatientsrsquo clinical neglect as indexed by the cancellationmeasure [r(15) = 46 p lt 05 Figure 9B] Thesecorrelations were driven largely by three of the patientsidentified in our focal lesion analysis (Figure 6A) withinvolvement of the key parietal region and high re-clicking behavior Their data are shown in filled circlesto the right in the graphs displayed in Figure 93 Thecorrelation between deviation on line bisection andre-click rate was not significant [r(15) = 13 ns] How-ever one would not predict that a deficit in retaininglocations already visited (or response inhibition) wouldnecessarily impair performance on a task such as linebisection
Effect of Varying Number of Targets
In 7 patients (Cases 5 8 10 11 14ndash16) we also exam-ined the effect of varying the number of targets in thesearch task Patients viewed lsquolsquotarget onlyrsquorsquo displays con-taining either 2 6 or 19 targets randomly distributedon the screen without any L-shaped distractors In-
Figure 7 Theneuroanatomical correlates of
the re-click deficit in neglect
patients with posterior plusanterior lesions (n = 5)
(A) Individual lesions of
patients with high re-click rate
(all 4 patients have MCAdamage) and overlap map for
these 4 patients where
maximum overlap occurs for
a region in the right inferiorfrontal lobe (shown in red)
damaged all 4 MCA patients
(B) Individual lesions of a single
patient with normal re-clickrate (C) Contrast map
highlighting the inferior frontal
locus (shown in yellow) whichis damaged in 4 out of
4 patients with a high re-click
rate and intact in the patient
with normal re-click rateRegions that are damaged in
patients with high re-click rates
and spared in the patient
with normal re-click rate areshown in red to yellow
whereas regions in blue denote
areas that are damaged in thepatient with normal re-click
rates but spared in patients with
high re-click rates
346 Journal of Cognitive Neuroscience Volume 17 Number 2
creasing the number of targets led to significantly higherre-click rates ( Jonckheere statistic = 133 p lt 01Figure 10) as well as more severe neglect (Jonckheerestatistic = 129 p lt 01) consistent with the proposalthat the mechanisms underlying pathological re-clickingmight contribute to neglect behavior Note that a lsquolsquoper-severativersquorsquo failure of response inhibition would not onits own easily explain this finding For the same absolutenumber of click responses one would predict a higherre-click rate with smaller set-sizes than with larger ones(see also Husain Mannan et al 2001 for a discussion)Although the number of subjects were small there wasno difference in the performance pattern between the3 patients who had increasing re-click rate functions inthe time-series analysis and the 4 patients with flat ordecreasing re-click rate functions
Varying Perceptual Load during Search
Can re-click behavior be explained simply by impairedvisual perception of targets from discriminators Neglectpatients did find it more difficult than control subjectsto select targets from distractors fixating on average19 distractors for every target found [cf 14 for controlsubjects t(29) = 485 p lt 0001] Patients also some-times clicked on distractors (64) but no more often
than controls (61) and it is possible that this rate iseffectively inflated by our strict criterion of considering aclick to be a response on the item that is being fixated atthe time of the click (It is likely that sometimes ob-servers click in response to targets they fixated one ormore saccades previously) Re-clicks on distractors wereextremely rare (06 cf 30 for targets)
To examine the effect of reduced perceptual load in8 patients (Cases 5 7 8ndash10 13 1 4 16) 6 of whom werehigh re-clickers we used search displays with salientcross (+) targets rather than Ts L distractors wereunchanged Patients fixated significantly fewer distrac-tors when searching for cross (+) targets [distractortarget ratio of 134 cf 181 for T targets t(7) = 60 p lt001] suggesting they did indeed find targetndashdistractordiscrimination easier Importantly however there wasno significant difference in re-click rate [26 for +targets cf 27 for T targets t(14) = 041 ns] or evenin re-fixation rate [100 for + targets cf 119 fort(14) = 041 ns] Thus these changes in perceptualload did not influence re-click performance
DISCUSSION
On this multitarget visual search task neglect patientsmissed targets to their left and also re-fixated targets
Figure 8 The probability of
re-clicking or re-fixating on
target as a function of delay
between the original click andsubsequent re-fixation (for
patients with high re-click
rates) (A) Probability functions
for patients with significantpositive slopes Four patients of
these patients have damage to
the focal intraparietal locusidentified in Figure 6C whereas
the single patient with a PCA
lesion and no damage to the
critical parietal locationindicated by symbol has only
a very shallow positive slope
(B) Probability functions for
patients with no significantpositive slope None of these
5 patients have damage to the
critical intraparietal region (C)As for A but for delay measured
in terms of intervening
fixations (D) As for B but for
delay measured in terms ofintervening fixations
Mannan et al 347
to the right much more often than healthy individuals(Figure 2A) Critically 10 of the 16 neglect patients re-clicked on rightward targets at a significantly higher ratethan healthy control subjects (Figure 2B) For 5 of thesepatients the probability of re-clicking increased withtime (Figure 8A) whereas the other 5 neglect patientswith high re-click rates demonstrated a constant or de-creasing probability of re-clicking over time (Figure 8C)The remaining 6 neglect patients had re-click rateswithin the normal range demonstrating that neitherneglect nor brain damage per se leads to a pathologicalre-click rate
High-resolution MRI mapping of lesions revealed im-portant anatomical dissociations within the neglect pop-ulation Patients with pathological re-click rates hadlesions affecting either the right intraparietal sulcus orthe inferior frontal lobe the only exception (Case 2) hada large occipito-temporal lesion that extended deep intothe parietal white matter Neglect patients with normalre-click rates did not have damage to either of thecritical locations in the intraparietal sulcus or inferiorfrontal lobe (Figures 5 6 and 7) Instead they had moredorsal lesions (for MCA territory patients) or occipitaland medial-temporal damage (for PCA territory lesions)which can also result in neglect (Mort et al 2003Cals Devuyst Afsar Karapanayiotides amp Bogousslavsky2002) Importantly lesion location rather than the sizeor age of lesion was critical to the presence of the re-clicking impairment Across the group of neglect pa-tients there was a moderate but significant correlationbetween re-click rate and degree of neglect (Figure 9)driven largely by patients with parietal damage who hadboth a high re-click rate and also severe neglect consis-tent with the proposal that mechanisms underlying thisbehavior in parietal patients might have a functionallyimportant role in determining the severity of neglect
Role of Right Intraparietal Sulcus
Notably for all patients with damage to the critical locusin the right intraparietal sulcus (which may also involveparieto-frontal white matter fibers) the probability of re-clicking on a target increased with time (or interveningsaccades) since initial discovery (Figure 8) Because theyrarely re-clicked at short intervalsmdasheven though themajority of re-fixations occurred during this periodmdashthis pattern of behavior suggests that for these patientstarget locations were actually veridically perceived and
Figure 9 (A) Neglect on experimental search task (19 targets and44 distractors) plotted against mean re-click rate for individual neglect
patients Percentage neglect was calculated as the difference between
the number of targets missed on the left versus right expressed as apercentage of the total number of targets missed There was a
significant correlation between the degree of neglect and the re-click
rate [r(15) = 465 p lt 05] Patients with high re-click rates are
highlighted by black circles (for patients with damage to the focalparietal location identified in Figure 6C) and light gray circles (for
patients with frontal damage) while the single patient with PCA
damage and a high re-click rate is indicated by dark gray (B) The
number of targets missed on the Mesulam shape-cancellation taskplotted against mean re-click rate for individual neglect patients Again
there was a significant correlation between the number of targets
missed and the mean re-click rate [r(15) = 456 p lt 05] Patients withhigh re-click rates are denoted as in A
Figure 10 Mean re-click rate (filled circles) and degree of neglect
(unfilled squares) increased systematically with increasing numbers of
targets in target-only displays
348 Journal of Cognitive Neuroscience Volume 17 Number 2
well encoded into working memory Furthermore de-tailed single-case studies of two parietal patients withhigh re-click rates (Malhotra Mannan Driver amp Husain2004 Husain Mannan et al 2001) demonstrate usingdifferent psychophysical methods that these patientscan accurately encode spatial locations4 Increasing re-clicks with time (or intervening saccades) would how-ever be consistent with a deficit in keeping track oftarget locations during extended search
Several lines of evidence support the proposal thatthe critical intraparietal sulcus region we have identifiedmay have an important role in the spatial transforma-tions required to keep track of spatial locations acrosssaccades In rhesus monkeys neurons within the lateralbank of the intraparietal sulcus (area LIP) encode andlsquolsquomaintainrsquorsquo the remembered locations of saccadic tar-gets over time (Chafee amp Goldman-Rakic 1998 MazzoniBracewell Barash amp Andersen 1996) Some visuallyresponsive neurons in the LIP also appear to shift theirretinotopic representation at the onset of an intendedsaccade as if lsquolsquore-mappingrsquorsquo representations of objects inspace to update them across shifts of gaze (DuhamelColby amp Goldberg 1992)
In humans this computation has been investigatedusing the double-step saccade paradigm where twoflashes of light appear in quick succession such thatthe second target disappears before a saccade can bemade to the first target Patients with lesions involvingthe right parietal cortex are impaired at performing thistask suggesting that they are unable to update spatialrepresentations accurately after even one saccade (Heideamp Kompf 1998 Duhamel et al 1992) Similarly trans-cranial magnetic stimulation of the posterior parietalcortex prior to the second saccade reduces its accuracy(Donkelaar amp Muri 2002) and a recent study of patientswith parietal lesions also implicates this area in thespatial updating of inhibition of return after a saccade(Sapir Hayes Henik Danzinger amp Rafal 2004) Func-tional imaging studies of double-step (Tobler et al2001) and triple-step memory-guided saccades (HeideBinkofski et al 2001) have identified a region withinthe intraparietal sulcus (Talairach coordinates 40 4044 for Tobler et al and 44 48 36 Heide et al) This isclose to the critical region of lesion overlap we haveidentified in the posterior neglect patients with high re-click rates (Talairach 37 39 37)
We suggest that in our patients with lesions involvingthe right intraparietal sulcus uncertainty about locationsof previously found targets increases with each saccadeThis uncertainty is further exacerbated by interferencefrom newer target locations leading to increased prob-ability of misjudging old target locations for new assearch progresses Such progressively worsening uncer-tainty might be due to an accumulating impairment inlsquolsquore-mapping target locationsrsquorsquo across saccades or im-paired lsquolsquomemoryrsquorsquo of previously inspected target loca-tions across saccades As noted above there is evidence
for both types of signalmdash re-mapping and maintainingsaccadic spatial memorymdashin the parietal cortex and it ispossible that disruption to both these functions contrib-utes to the re-click deficit we have identified Certainlythe two potential types of deficit are not mutuallyincompatible
A slightly different account of the re-clicking behaviormight be framed in terms of the intriguing hypothesisproposed by Manly Woldt Watson and Warburton(2002) to explain visible re-marking on a standard can-cellation task They argue that such behavior is due topatients erroneously transposing leftward stimuli to theright resulting in apparent re-cancellation These au-thors found that removing targets from the left re-duced both neglect and re-marking However they didnot monitor eye movements so it is possible that sometargets that are logged as neglected (because they werenot cancelled) were nevertheless fixated and thereforeform part of the load on memory during search Fur-thermore in our experimental study subjects with ahigh re-click rate also failed to click on a higher pro-portion of fixated targets Manly et alrsquos proposal wouldnot easily account for such lsquolsquoclick omissionsrsquorsquo becausethese refer to fixated targets that were not actuallyresponded to
Finally it might be argued that increasing re-clickingwith search duration reflects a deficit in spatial workingmemory (SWM) that is independent of saccadic eyemovements (ie a decay in memory that is not trans-saccadic) Because time and number of interveningsaccades were not independently manipulated in thisunrestricted visual search paradigm we are not ablecategorically to distinguish between these possibilitiesWe note however that functional imaging studies ofSWM tasks that do not require eye movements oftendemonstrate activity related to the maintenance ofspatial information which is generally more posteriorand dorsal than the parietal locations isolated in thisstudy (DrsquoEsposito et al 1998) Moreover recent behav-ioral data indicate that SWM is more vulnerable tosaccadic eye movements than shifts in attention or limbmovements (Pearson amp Sahraie 2003)
Frontal Lesion Site Associated withHigh Re-click Rates
Four out of 5 neglect patients with lesions involvingposterior plus anterior damage also showed a high re-click rate and the area of damage common to thesepatients centered on the right inferior frontal lobe cor-responding to Brodmannrsquos area 44 (Figure 7C) Thisregion has previously been associated with frontalneglect (Husain amp Kennard 1996) and is consideredto be part of the ventral premotor cortex and thehomologue of Brocarsquos areas in the left hemisphere(Rizzolatti amp Arbib 1998 Jackson amp Husain 1996) Inthese frontal patients the probability of re-clicking did
Mannan et al 349
not increase with time interval between the first clickand re-fixation but rather it stayed constant or declinedwith increasing delays
Such behavior might be consistent with a failure toinhibit responses to rightward locationsmdasha type oflsquolsquoperseverativersquorsquo behavior Note though that this is notnecessarily the same type of perseverative responseas seen when some patients with neglect re-markpreviously found items on standard cancellation tests(Rusconi et al 2002 Na et al 1999) In those casestargets that are visibly cancelled are often marked againwhereas in our computerized search tasks no marks arevisibly left on targets once they have been clicked uponMoreover only one of our neglect patients (Case 1) de-monstrated re-cancellation of visibly cancelled targetson pen-and-paper tasks and then only on one occasionImportantly this patient actually had a normal re-clickrate (67) on our task and his stroke was in the PCAterritory sparing both the intraparietal sulcus and fron-tal lobe
In support of a role for the right frontal lobe in in-hibiting responses to previously found targets recentimaging studies of response inhibition have highlightedregions of the right inferior frontal gyrus (IFG) very closeto the centroid of damage for our frontal patients (38 1618 cf 43 22 21 Konishi Kyoichi Uchida Sekihara ampMiyashita 1998 and 41 16 19 Konishi Kyoichi UchidaKikyo et al 1999) Alternative explanations for the highprobability of re-clicking directly after clicking might bethat these patients have impaired perceptual discrimina-tion of targets impaired perceptual localization of targetsor that they encounter difficulty in encoding spatiallocations into working memory Our data are not sugges-tive of a perceptual discrimination problem as changingthe perceptual load did not inf luence re-click ratesMoreover the patients still had high re-click rates in thetarget only conditions (Figure 10) where no perceptualdiscrimination of targets and distractors was necessaryPoor perceptual localization would seem unlikely giventhat these patients have frontal lesions Furthermoreusing a vertical SWM task similar to the Corsi blocktapping task (Malhotra et al 2004) we have found thatpatients with neglect can localize and perceptually en-code single target locations as well as controls and aresignificantly impaired at remembering two or more loca-tions shown in sequence consistent with the presentobservations Some recent functional imaging data sug-gest that activation within the prefrontal cortex corre-lates well with successful encoding of material intoworking memory (Rypma amp DrsquoEsposito 2003) Furtherresearch is required to distinguish between these pos-sible explanations for the mechanisms underlying there-click deficit in frontal patients
The only patient with frontal damage who did re-clickmore frequently with increasing time had damage cen-tered on the dorsolateral prefrontal cortex (althoughalso encroaching on the IFG) Neurons in the dorsolat-
eral prefrontal cortex like those in the LIP encode theremembered locations of saccadic targets (Goldman-Rakic 1996) This patient (Case 15) who had the high-est re-click rate of all also had damage to the criticalparietal location described above (in contrast to all otherfrontal patients) Thus she may have suffered from acombination of underlying deficits
Re-fixations and Misjudging PreviouslyInspected Locations during Search
In this search task re-fixation and re-click rates weresignificantly correlated for neglect patients Howeverthere were important dissociations within individuals 2patients with PCA lesions had re-fixation rates of 220and 80 respectively but both had a re-click rate ofonly 7 These are clearly examples of individuals whodo not show pathological re-clicking despite having highre-fixation rates High re-fixation rates may result froma lack of integration of spatial information due to V1damage (Zihl amp von Cramon 1985) in our PCA patientsor possibly because of reduced inhibition of return(Bartolomeo Sieroff Decaix amp Chokron 2001) What-ever the exact cause for dissociations between re-fixa-tions and re-click rates our data suggest re-fixation doesnot necessarily index the same underlying deficits asrevealed by our re-click measure High re-fixation ratesunlike high re-click rates were also not significantlyassociated with damage to any specific cortical locationagain demonstrating a dissociation between re-fixationsand pathological re-clicking
The relationship between re-click rates and neglect isunlikely to be a simple one In our study there was amoderate but significant correlation between the twomeasures suggesting that this association is unlikely tohave arisen by chance However non-neglect patientscan also exhibit a high re-click rate (unpublished obser-vations from our laboratory) Taken together our resultssuggest that difficulties in keeping track of previouslyinspected targets (because of impaired trans-saccadic re-mapping andor memory) may be one contributingcomponent to visual neglect in patients with lesions in-volving the right intraparietal sulcus As mentioned thesearch task we used differed in one striking aspect fromstandard cancellation tests subjects do not visibly markthe targets they visit Whereas in cancellation tests pa-tients mark each target they find which might appear todispense with any need to remember which locationshave been searched before However it should be ap-preciated that in most standard cancellation tests theindividual marks made by patients may not be clearlyvisible in peripheral vision particularly if neglect pa-tients have a local spatial bias (Rafal 1994 RobertsonLamb amp Knight 1988) or a restricted field of view(Russell Malhotra amp Husain 2004) Thus when com-bined with a rightward attentional bias keeping track ofsearched locations may be critical to prevent gaze from
350 Journal of Cognitive Neuroscience Volume 17 Number 2
returning to rightward locations even if they have beenmarked before as the marks may not be visible untilgaze returns to them Consistent with this hypothesisneglect is greater when cancellation is performed usinginvisible marks compared to large and salient red marksthat can be seen peripherally (Wojciulik Husain Clarkeamp Driver 2001)
A task like the present search-and-click paradigm maymore closely resemble search in the real world than dotraditional cancellation tests When we examine a visualscene we do not leave marks on each object we ex-amine Instead just as in the search task used in thisstudy we have to keep track of the locations we visit ifwe are to avoid inefficient re-examination of thoselocations Neglect patients with damage to the rightintraparietal sulcus appear to be impaired at keepingtrack of searched locations in this way When combinedwith their rightward attentional bias we propose thatsuch an impairment leads to recursive search of stimulito the right in experimental search tasks as used hereand also in daily life where neglect is so apparentdespite unlimited viewing time
The paradigm we have used has the important virtuesof allowing patients to explore freelymdashas in real-worldsearch or clinical cancellation tasksmdashand relating theirre-click behavior directly to the severity of neglect in thesame task But it might be argued that it would be usefulalso to probe SWM in a more controlled manner by re-stricting observersrsquo eye movements to a specific set oflocations and probing memory of previously visitedplaces A recent study using a gaze-contingent para-digm has elegantly probed the capacity of spatial mem-ory during search in healthy observers and foundevidence for memory for up to four previous locations(McCarley Wang Kramer Irwin amp Peterson 2003)Variants of this paradigm may in the future be usefulto probe the capacity of SWM across saccades in patientswith neglect and also to probe further whether thereare differences between neglect patients with parietaland frontal lesions
METHODS
Subjects Imaging and Lesion Mapping
Sixteen patients with left neglect following right hemi-sphere infarction or hemorrhage were recruited Con-sent was obtained according to the Declaration ofHelsinki as approved by the hospital ethical commit-tees Presence of neglect was determined by clinicalobservation plus bedside tests including the Mesulamshape-cancellation task (Mesulam 1985) and line bisec-tion task (Wilson et al 1987) (see Table 1) Twelvepatients had MCA territory strokes the rest involvedonly the PCA territory which can also result in neglect(Cals et al 2002) Only one neglect patient (Case 1 whodid not have frontal involvement) demonstrated re-
cancellation of visibly cancelled targets on pen-and-paper tasks and then only on one occasion Fifteenage-matched healthy volunteers were controls (meanage = 63 SD = 14 vs 63 SD = 13 for patients)
All patients had clinical CT scans In addition dedi-cated high-resolution structural MRI scans were alsoobtained for 1416 neglect patients MRI scans wereunobtainable for the other two (Cases 11 and 16)because of claustrophobia or loss to follow up In theMCA neglect group median time between stroke andscan was 63 days with the shortest interval being35 days for the PCA neglect group one patient had tobe scanned relatively early at 9 days but the mediantime was 140 days MRI scans comprised one 256-sliceT1-weighted acquisition (MPRAGE TR 97 msec TE4 msec TA 12 min) performed in the coronal planealigned perpendicular to the anteriorndashposterior commis-sural line on a Siemens Vision 15-T scanner Within-and between-slice resolution was 1 mm For each pa-tient the area of damage was determined by detailedvisual inspection of the digital image for every singleslice Lesion delineation was performed by neurologists(authors D M and M H) by tracing by hand theboundary of the lesion onto every single 1-mm-thickaxial slice using a graphics tablet (WACOM Intuos A6)Combining all the slices then led to a 3-D region of in-terest (ROI) for each patient For one patient (Case 15)there were two areas of discrete focal damage one inthe frontal cortex and the other in the parietal cortex
Normalization of each MRI including the 3-D ROI tothe T1-template was performed with SPM99 (wwwfilionuclacukspm) The ROI was used as a mask for thelesion during determination of alignment parameters tominimize the contribution of abnormal brain to thisprocess (Rorden amp Brett 2000) The normalized ROIof each neglect patient was superimposed upon the T1template to find areas of maximal lesion overlap in eachgroup (see Results) as well as lesion volume usingMRIcro shareware (Rorden amp Brett 2000)
Experimental Procedure
Stimuli were presented on an SVGA monitor (29 22 in)viewed at 57 cm distance Each search display com-prised a virtual 9 7 grid each cell containing a single18 elementmdasheither a letter T (target) or a letter L (dis-tractor) (see Figure 1) The position of each elementwithin each cell was jittered with 19 targets and 44distractors randomly distributed over the virtual gridThe number of targets on the right and left halves ofthe display was balanced across trials Seven neglectpatients (Cases 5 8 10 11 14ndash16) were further testedon lsquolsquoTarget onlyrsquorsquo displays which had varying numbersof Ts (2 6 or 19) now with no L-shaped distractorsThe presentations of these lsquolsquoTarget onlyrsquorsquo screens wereinterspersed with those of the original search screens inrandom order in the same experimental session
Mannan et al 351
Eye movements were recorded using a video-basedpupil tracker with a temporal resolution of 4 msec andspatial accuracy of lt058 (EyeLink Sensorimotoric Sys-tems Berlin Germany) Subjects were asked to keeptheir head still The experimenter also monitored gazeposition on-line At the beginning of each session (andafter breaks in testing or following any unintended headmovement that disrupted gaze recordings) eye-trackingwas re-calibrated Each trial was initiated by the exper-imenter and began with a central fixation cross for 1 secthen the search display
Subjects were instructed to search for Ts (among anyLs) Critically while looking at any target they had toclick a response button held in the right hand onlyif they considered this target to be a new discovery(ie a target they had not clicked on before) Clickingthe response button did not change the display Theresponse button was held by the patient in a comfort-able position to the right of the midline Subjectswere instructed and reminded throughout testing toclick on each target found once only Any later re-click on
a previously found target thus indicates a failure to retainthe fact that this target had already been discoveredThe experimenter terminated each search when thesubject indicated verbally that they felt all targets in thedisplay had now been found Each subject searched aminimum of 5 displays (mean = 7 range 5ndash14) Eyemovements were analyzed off-line using custom soft-ware An automatic algorithm was used to extract sac-cades (saccade onset defined as eye speed rising above258 sec1 for two or more samples and a criterion ofminimum fixation duration of 60 msec) Blinks wereeliminated from the trace by visual inspection and sac-cade and fixation delineation were checked visually inthe record Next fixations were automatically assignedto items in the display if the mean position fell within18 of that item Two or more consecutive fixations onthe same item were re-scored as a single extendedfixation A re-fixation was scored only when at leastone fixation elsewhere on another item intervened
Most click responses were made during a fixation andthe few button-clicks occurring during a saccade were
Table 1 Patient Details
CaseAge
(years) PathologyVolume(cm3)
Age ofLesion (days)
Visual FieldDefect
Mean Deviationfrom Center on
Line Bisection Task(cm) (rightward +)
Mesulum Score(Maximum Possible
Score = 60) Lesion Type
1 68 Infarct 1282 64 + 867 17 PCA
2 76 Infarct 1528 227 + 003 41 PCA
3 68 Infarct 79 207 + 297 36 PCA
4 77 Infarct 251 158 + 150 54 PCA
5 63 Infarct 28 29 033 55 MCA
6 59 Hemorrhage 172 7 + 32 6 MCA
7 66 Hemorrhage 273 325 + 213 31 MCA
8 68 Infarct 309 3 25 17 MCA
9 29 Infarct 115 17 113 55 MCA
10 70 Infarct 1339 324 153 57 MCA
11 42 Infarct ndash 11 097 24 MCA
12 75 Infarct 757 38 013 4 MCA
13 51 Infarct 694 141 083 31 MCA
14 59 Infarct 1144 356 + 223 37 MCA
15 80 Infarct 261 44 35 6 MCA
16 64 Infarct ndash 30 033 55 MCA
MEAN 63 66 124 188 33
SE 4 13 32 5
Cases 1ndash9 had damage to the nonfrontal (posterior only) cortex whereas Cases 10ndash15 had both frontal and posterior (posterior and anterior) corticalinvolvement Case 16 is a patient with a subcortical lesion All but 3 of the patients with right MCA infarction had intact visual fields on confrontationTwo of these (Cases 7 and 14) were diagnosed with left homonymous hemianopia whereas one (Case 6) had a left inferior quandrantic field defectAll patients with PCA territory damage had partial or complete contralesional visual field loss No patient suffered from dysphasia
352 Journal of Cognitive Neuroscience Volume 17 Number 2
assigned to the previous fixation Clicks were assigned toa particular item if eye position at the time of the clickwas within 28 of that item or in the cases where morethan one item was within 28 of current eye position theclick response was simply attributed to the item closestto that eye position
Button-clicks were assigned to a particular item if eyeposition at the time of the click was within 28 of thatitem A re-click on an item required at least one fixationon another item between successive clicks (in practiceseveral fixations elsewhere typically intervened betweeninitial click and later re-click see Results)
Statistical analyses were performed using SPSS andSigmaplot (SPSS Science Chicago USA) Jonckheere sta-tistic was derived by hand Statistical significance was setas p lt 05
Acknowledgments
We thank the reviewers of this manuscript and Dr AndrewParton for their extremely helpful comments We thank thepatients who participated Dr J Wade Dr A Rudd and thestaff of the stroke units at Charing Cross and St ThomasrsquoHospitals This research was supported by grants from theWellcome Trust to M H C K and J D J D holds a RoyalSociety Wolfson Research Merit Award
Reprint requests should be sent to Drs Sabira Mannan andMasud Husain Division of Neuroscience and PsychologicalMedicine Imperial College London Charing Cross CampusLondon W6 8RF UK or via e-mail smannanimperialacukor mhusainimperialacuk
Notes
1 We are grateful to one of the reviewers for suggesting thistime-series analysis2 There was no significant difference between the number oftargets fixated between the two groups so differences in lsquolsquoclickomissionsrsquorsquo cannot be attributed to the number of fixatedtargets that had to be kept track of during search3 The correlation between these four points alone inFigure 9A is high [r(3) = 81 cf r(15) = 47 for the wholegroup]4 We have also tested 10 neglect patients and 10 controlstroke patients on a vertical version of the Corsi blocks task(clinically used to measure spatial working memory capacity)Neglect patients had significantly impaired capacity but wereable to localize single locations well suggesting that mislocal-ization is unlikely to account for their deficit on this spatialworking memory task
REFERENCES
Bartolomeo P Sieroff E Decaix C amp Chokron S (2001)Modulating the attentional bias in unilateral neglect theeffects of the strategic set Experimental Brain Research137 432ndash444
Bates E Wilson S M Saygin A P Dick F Sereno M IKnight R T amp Dronkers N F (2003) Voxel-basedlesion-symptom mapping Nature Neuroscience 6 448ndash450
Behrmann M Watt S Black S E amp Barton J J (1997)Impaired visual search in patients with unilateral neglect
An oculographic analysis Neuropsychologia 351445ndash1458
Bisiach E amp Vallar G (2000) Unilateral neglect in humans InG Rizzolatti (Ed) Handbook of neuropsychology (2nd edvol 1 pp 459ndash502) Amsterdam Elsevier
Cals N Devuyst G Afsar N Karapanayiotides T ampBogousslavsky J (2002) Pure superficial posterior cerebralartery territory infarction in The Lausanne Stroke RegistryJournal of Neurology 249 855ndash861
Chafee M amp Goldman-Rakic P (1998) Matching patterns ofactivity in primate prefrontal area 8a and parietal area 7ipneurons during a spatial working memory task Journal ofNeurophysiology 79 2919ndash2940
Chedru F Leblanc M amp Lhermitte F (1973) Visualsearching in normal and brain-damaged subjectsContributions to the study of unilateral subjectsCortex 9 94ndash111
DrsquoEsposito M Aguirre G K Zarahn E Ballard DShin R K amp Lease J (1998) Functional MRI studies ofspatial and nonspatial working memory Cognitive BrainResearch 7 1ndash13
Donkelaar P amp Muri R (2002) Craniotopic updating ofvisual space across saccades in the human posteriorparietal cortex Proceedings of the Royal Society London B269 735ndash739
Driver J amp Husain M (2002) The role of spatial workingmemory deficits in pathological search by neglect patientsIn the cognitive and neural bases of spatial neglect InH-O Karnath A D Milner amp G Vallar (Eds) The cognitiveand neural bases of spatial neglect Oxford OxfordUniversity Press
Duhamel J R Colby C amp Goldberg M E (1992) Theupdating of the representation of visual space in parietalcortex by intended eye movements Science 25590ndash92
Goldman-Rakic P S (1996) The prefrontal landscapemdashImplications of functional architecture for understandinghuman mentation and the central executive PhilosophicalTransactions of The Royal Society of London Series BBiological Sciences 351 1445ndash1453
Heide W Binkofski F Seitz R J Posse S Nitschke M FFreund H J amp Kompf D (2001) Activation offrontoparietal cortices during memorized triple-stepsequences of saccadic eye movements An fMRI studyEuropean Journal of Neuroscience 13 1177ndash1189
Heide W amp Kompf D (1998) Combined deficits ofsaccades and visuo-spatial orientation after cortical lesionsExperimental Brain Research 123 164ndash171
Husain M amp Kennard C (1996) Visual neglect associatedwith frontal lobe infarction Journal of Neurology 243652ndash657
Husain M Mannan S Hodgson T Wojciulik E Driver Jamp Kennard C (2001) Impaired spatial working memoryacross saccades contributes to abnormal search in parietalneglect Brain 121 941ndash952
Husain M amp Rorden C (2003) Non-spatially lateralizedmechanisms in hemispatial neglect Nature ReviewsNeuroscience 4 23ndash36
Jackson S R amp Husain M (1996) Visuomotor functionsof the lateral pre-motor cortex Current Opinion inNeurobiology 6 788ndash795
Kinsbourne M (1993) Orientational bias model of unilateralneglect Evidence from attentional gradients withinhemispace In I H Robertson amp J C Marshall (Eds)Unilateral neglect Clinical and experimental studies(pp 63ndash86) Hove Erlbaum
Konishi S Kyoichi N Uchida I Kikyo H Kameyama Mamp Miyauchi S (1999) Common inhibitory mechanism in
Mannan et al 353
human inferior prefrontal cortex revealed by event-relatedfunctional MRI Brain 122 981ndash991
Konishi S Kyoichi N Uchida I Sekihara K amp Miyashita Y(1998) No-go dominant brain activity in human inferiorprefrontal cortex revealed by functional magneticresonance imaging European Journal of Neuroscience10 1209ndash1213
Malhotra P Mannan S Driver J amp Husain M (2004)Impaired spatial working memory One component of thevisual neglect syndrome Cortex 40 667ndash676
Manly T Woldt K Watson P amp Warburton E (2002) Ismotor perseveration in unilateral neglect lsquolsquodrivenrsquorsquo by thepresence of neglected left-sided stimuli Neuropsychologia40 1794ndash1803
Mazzoni P Bracewell R Barash S amp Andersen R (1996)Spatially tuned auditory responses in area LIP of macaquesperforming delayed memory saccades to acoustic targetsJournal of Neurophysiology 75 1233ndash1241
McCarley J Wang R Kramer A Irwin D amp Peterson M(2003) How much memory does oculomotor search havePsychological Science 14 422ndash426
Mesulam M-M (1985) Principles of behavioural neurologyTests of directed attention and memory Philadelphia Davis
Mesulam M M (1999) Spatial attention and neglectParietal frontal and cingulate contributions to the mentalrepresentation and attentional targeting of salientextrapersonal events Philosophical Transactions of theRoyal Society of London Series B Biological Sciences354 1325ndash1346
Mort D Malhotra P Mannan S Rorden C Pambakian AKennard C amp Husain M (2003) The anatomy of visualneglect Brain 126 1986ndash1997
Na D L Adair J C Kang Y Chung C S Lee K H ampHeilman K M (1999) Motor perseverative behaviour ona line cancellation task Neurology 52 1569ndash1576
Pearson D G amp Sahraie A (2003) Oculomotor controland the maintenance of spatially and temporally distributedevents in visuo-spatial working memory QuarterlyJournal of Experimental Psychology 56A 1089ndash1111
Rafal R (1994) Neglect Current Opinion in Neurobiology4 231ndash236
Rizzolatti G amp Arbib M A (1998) Language within our graspTrends in Neurosciences 21 188ndash194
Robertson L C Lamb M R amp Knight R T (1988) Effectsof lesions of temporalndashparietal junction on perceptual andattentional processing in humans Journal of Neuroscience8 3757ndash3769
Rorden C amp Brett M (2000) Stereotaxic display of brainlesions Behavioural Neurology 12 191ndash200
Rusconi M L Maravita A Bottini G amp Vallar G (2002)Is the intact side really intact Perseverative responses inpatients with unilateral neglect A productive manifestationNeuropsychologia 40 594ndash604
Russell C Malhotra P amp Husain M (2004) Attentionmodulates the visual field in healthy observers and parietalpatients NeuroReport 15 189ndash193
Rypma B amp DrsquoEsposito M (2003) A subsequent-memoryeffect in dorsolateral prefrontal cortex Cognitive BrainResearch 16 162ndash166
Sapir A Hayes A Henik A Danzinger S amp Rafal R(2004) Parietal lobe lesions disrupt saccadic remapping ofinhibitory location tagging Journal of CognitiveNeuroscience 16 503ndash509
Smania N Martini M C Gambina G A T Palamara ANatale E amp Marzi C A (1998) The spatial distributionof visual attention in hemineglect and extinction patientsBrain 121 1759ndash1770
Sprenger A Kompf D amp Heide W (2002) Visual searchin patients with left visual hemineglect Progress in BrainResearch 140 395ndash416
Tobler P Felblinger J Burki M Nirkko A Ozdoba C ampMuri R (2001) Functional organisation of the saccadicreference system processing extraretinal signals in humansVision Research 41 1351ndash1358
Wilson B Cockburn J amp Halligan P W (1987)Behavioural inattention test Bury St EdmundsThames Valley
Wojciulik E Husain M Clarke K amp Driver J (2001)Spatial working memory deficit in unilateral neglectNeuropsychologia 39 390ndash396
Zihl J amp Hebel N (1997) Patterns of oculomotor scanningin patients with unilateral posterior parietal and frontallobe damage Neuropsychologia 35 893ndash906
Zihl J amp von Cramon D (1985) Visual field recovery fromscotoma in patients with postgeniculate damage A reviewof 55 cases Brain 108 335ndash365
354 Journal of Cognitive Neuroscience Volume 17 Number 2
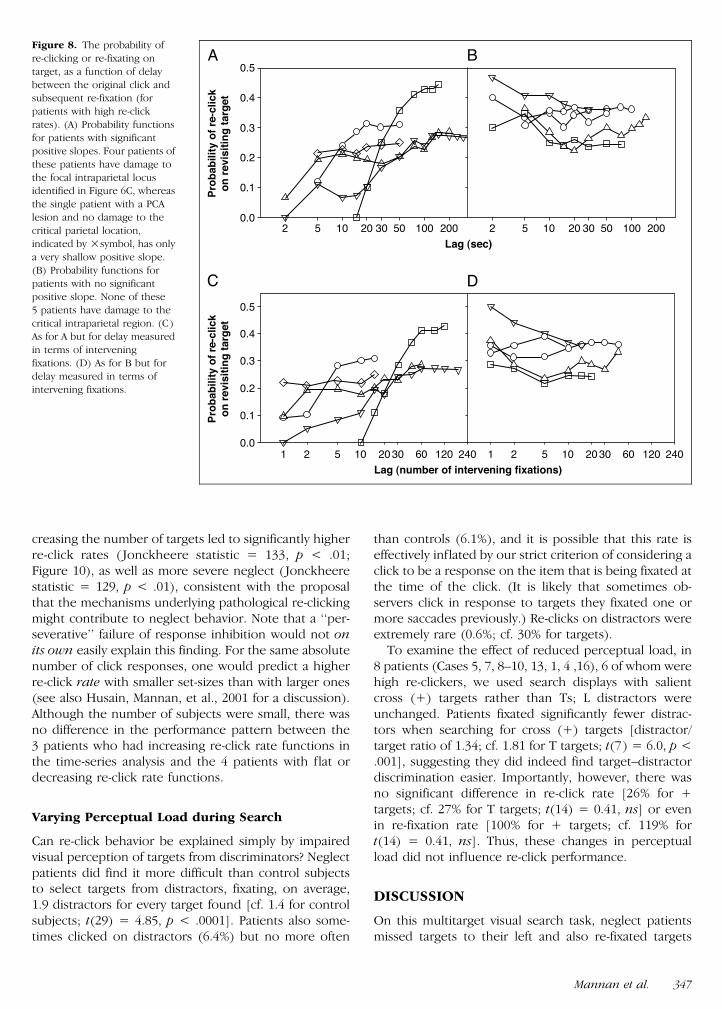
creasing the number of targets led to significantly higherre-click rates ( Jonckheere statistic = 133 p lt 01Figure 10) as well as more severe neglect (Jonckheerestatistic = 129 p lt 01) consistent with the proposalthat the mechanisms underlying pathological re-clickingmight contribute to neglect behavior Note that a lsquolsquoper-severativersquorsquo failure of response inhibition would not onits own easily explain this finding For the same absolutenumber of click responses one would predict a higherre-click rate with smaller set-sizes than with larger ones(see also Husain Mannan et al 2001 for a discussion)Although the number of subjects were small there wasno difference in the performance pattern between the3 patients who had increasing re-click rate functions inthe time-series analysis and the 4 patients with flat ordecreasing re-click rate functions
Varying Perceptual Load during Search
Can re-click behavior be explained simply by impairedvisual perception of targets from discriminators Neglectpatients did find it more difficult than control subjectsto select targets from distractors fixating on average19 distractors for every target found [cf 14 for controlsubjects t(29) = 485 p lt 0001] Patients also some-times clicked on distractors (64) but no more often
than controls (61) and it is possible that this rate iseffectively inflated by our strict criterion of considering aclick to be a response on the item that is being fixated atthe time of the click (It is likely that sometimes ob-servers click in response to targets they fixated one ormore saccades previously) Re-clicks on distractors wereextremely rare (06 cf 30 for targets)
To examine the effect of reduced perceptual load in8 patients (Cases 5 7 8ndash10 13 1 4 16) 6 of whom werehigh re-clickers we used search displays with salientcross (+) targets rather than Ts L distractors wereunchanged Patients fixated significantly fewer distrac-tors when searching for cross (+) targets [distractortarget ratio of 134 cf 181 for T targets t(7) = 60 p lt001] suggesting they did indeed find targetndashdistractordiscrimination easier Importantly however there wasno significant difference in re-click rate [26 for +targets cf 27 for T targets t(14) = 041 ns] or evenin re-fixation rate [100 for + targets cf 119 fort(14) = 041 ns] Thus these changes in perceptualload did not influence re-click performance
DISCUSSION
On this multitarget visual search task neglect patientsmissed targets to their left and also re-fixated targets
Figure 8 The probability of
re-clicking or re-fixating on
target as a function of delay
between the original click andsubsequent re-fixation (for
patients with high re-click
rates) (A) Probability functions
for patients with significantpositive slopes Four patients of
these patients have damage to
the focal intraparietal locusidentified in Figure 6C whereas
the single patient with a PCA
lesion and no damage to the
critical parietal locationindicated by symbol has only
a very shallow positive slope
(B) Probability functions for
patients with no significantpositive slope None of these
5 patients have damage to the
critical intraparietal region (C)As for A but for delay measured
in terms of intervening
fixations (D) As for B but for
delay measured in terms ofintervening fixations
Mannan et al 347
to the right much more often than healthy individuals(Figure 2A) Critically 10 of the 16 neglect patients re-clicked on rightward targets at a significantly higher ratethan healthy control subjects (Figure 2B) For 5 of thesepatients the probability of re-clicking increased withtime (Figure 8A) whereas the other 5 neglect patientswith high re-click rates demonstrated a constant or de-creasing probability of re-clicking over time (Figure 8C)The remaining 6 neglect patients had re-click rateswithin the normal range demonstrating that neitherneglect nor brain damage per se leads to a pathologicalre-click rate
High-resolution MRI mapping of lesions revealed im-portant anatomical dissociations within the neglect pop-ulation Patients with pathological re-click rates hadlesions affecting either the right intraparietal sulcus orthe inferior frontal lobe the only exception (Case 2) hada large occipito-temporal lesion that extended deep intothe parietal white matter Neglect patients with normalre-click rates did not have damage to either of thecritical locations in the intraparietal sulcus or inferiorfrontal lobe (Figures 5 6 and 7) Instead they had moredorsal lesions (for MCA territory patients) or occipitaland medial-temporal damage (for PCA territory lesions)which can also result in neglect (Mort et al 2003Cals Devuyst Afsar Karapanayiotides amp Bogousslavsky2002) Importantly lesion location rather than the sizeor age of lesion was critical to the presence of the re-clicking impairment Across the group of neglect pa-tients there was a moderate but significant correlationbetween re-click rate and degree of neglect (Figure 9)driven largely by patients with parietal damage who hadboth a high re-click rate and also severe neglect consis-tent with the proposal that mechanisms underlying thisbehavior in parietal patients might have a functionallyimportant role in determining the severity of neglect
Role of Right Intraparietal Sulcus
Notably for all patients with damage to the critical locusin the right intraparietal sulcus (which may also involveparieto-frontal white matter fibers) the probability of re-clicking on a target increased with time (or interveningsaccades) since initial discovery (Figure 8) Because theyrarely re-clicked at short intervalsmdasheven though themajority of re-fixations occurred during this periodmdashthis pattern of behavior suggests that for these patientstarget locations were actually veridically perceived and
Figure 9 (A) Neglect on experimental search task (19 targets and44 distractors) plotted against mean re-click rate for individual neglect
patients Percentage neglect was calculated as the difference between
the number of targets missed on the left versus right expressed as apercentage of the total number of targets missed There was a
significant correlation between the degree of neglect and the re-click
rate [r(15) = 465 p lt 05] Patients with high re-click rates are
highlighted by black circles (for patients with damage to the focalparietal location identified in Figure 6C) and light gray circles (for
patients with frontal damage) while the single patient with PCA
damage and a high re-click rate is indicated by dark gray (B) The
number of targets missed on the Mesulam shape-cancellation taskplotted against mean re-click rate for individual neglect patients Again
there was a significant correlation between the number of targets
missed and the mean re-click rate [r(15) = 456 p lt 05] Patients withhigh re-click rates are denoted as in A
Figure 10 Mean re-click rate (filled circles) and degree of neglect
(unfilled squares) increased systematically with increasing numbers of
targets in target-only displays
348 Journal of Cognitive Neuroscience Volume 17 Number 2
well encoded into working memory Furthermore de-tailed single-case studies of two parietal patients withhigh re-click rates (Malhotra Mannan Driver amp Husain2004 Husain Mannan et al 2001) demonstrate usingdifferent psychophysical methods that these patientscan accurately encode spatial locations4 Increasing re-clicks with time (or intervening saccades) would how-ever be consistent with a deficit in keeping track oftarget locations during extended search
Several lines of evidence support the proposal thatthe critical intraparietal sulcus region we have identifiedmay have an important role in the spatial transforma-tions required to keep track of spatial locations acrosssaccades In rhesus monkeys neurons within the lateralbank of the intraparietal sulcus (area LIP) encode andlsquolsquomaintainrsquorsquo the remembered locations of saccadic tar-gets over time (Chafee amp Goldman-Rakic 1998 MazzoniBracewell Barash amp Andersen 1996) Some visuallyresponsive neurons in the LIP also appear to shift theirretinotopic representation at the onset of an intendedsaccade as if lsquolsquore-mappingrsquorsquo representations of objects inspace to update them across shifts of gaze (DuhamelColby amp Goldberg 1992)
In humans this computation has been investigatedusing the double-step saccade paradigm where twoflashes of light appear in quick succession such thatthe second target disappears before a saccade can bemade to the first target Patients with lesions involvingthe right parietal cortex are impaired at performing thistask suggesting that they are unable to update spatialrepresentations accurately after even one saccade (Heideamp Kompf 1998 Duhamel et al 1992) Similarly trans-cranial magnetic stimulation of the posterior parietalcortex prior to the second saccade reduces its accuracy(Donkelaar amp Muri 2002) and a recent study of patientswith parietal lesions also implicates this area in thespatial updating of inhibition of return after a saccade(Sapir Hayes Henik Danzinger amp Rafal 2004) Func-tional imaging studies of double-step (Tobler et al2001) and triple-step memory-guided saccades (HeideBinkofski et al 2001) have identified a region withinthe intraparietal sulcus (Talairach coordinates 40 4044 for Tobler et al and 44 48 36 Heide et al) This isclose to the critical region of lesion overlap we haveidentified in the posterior neglect patients with high re-click rates (Talairach 37 39 37)
We suggest that in our patients with lesions involvingthe right intraparietal sulcus uncertainty about locationsof previously found targets increases with each saccadeThis uncertainty is further exacerbated by interferencefrom newer target locations leading to increased prob-ability of misjudging old target locations for new assearch progresses Such progressively worsening uncer-tainty might be due to an accumulating impairment inlsquolsquore-mapping target locationsrsquorsquo across saccades or im-paired lsquolsquomemoryrsquorsquo of previously inspected target loca-tions across saccades As noted above there is evidence
for both types of signalmdash re-mapping and maintainingsaccadic spatial memorymdashin the parietal cortex and it ispossible that disruption to both these functions contrib-utes to the re-click deficit we have identified Certainlythe two potential types of deficit are not mutuallyincompatible
A slightly different account of the re-clicking behaviormight be framed in terms of the intriguing hypothesisproposed by Manly Woldt Watson and Warburton(2002) to explain visible re-marking on a standard can-cellation task They argue that such behavior is due topatients erroneously transposing leftward stimuli to theright resulting in apparent re-cancellation These au-thors found that removing targets from the left re-duced both neglect and re-marking However they didnot monitor eye movements so it is possible that sometargets that are logged as neglected (because they werenot cancelled) were nevertheless fixated and thereforeform part of the load on memory during search Fur-thermore in our experimental study subjects with ahigh re-click rate also failed to click on a higher pro-portion of fixated targets Manly et alrsquos proposal wouldnot easily account for such lsquolsquoclick omissionsrsquorsquo becausethese refer to fixated targets that were not actuallyresponded to
Finally it might be argued that increasing re-clickingwith search duration reflects a deficit in spatial workingmemory (SWM) that is independent of saccadic eyemovements (ie a decay in memory that is not trans-saccadic) Because time and number of interveningsaccades were not independently manipulated in thisunrestricted visual search paradigm we are not ablecategorically to distinguish between these possibilitiesWe note however that functional imaging studies ofSWM tasks that do not require eye movements oftendemonstrate activity related to the maintenance ofspatial information which is generally more posteriorand dorsal than the parietal locations isolated in thisstudy (DrsquoEsposito et al 1998) Moreover recent behav-ioral data indicate that SWM is more vulnerable tosaccadic eye movements than shifts in attention or limbmovements (Pearson amp Sahraie 2003)
Frontal Lesion Site Associated withHigh Re-click Rates
Four out of 5 neglect patients with lesions involvingposterior plus anterior damage also showed a high re-click rate and the area of damage common to thesepatients centered on the right inferior frontal lobe cor-responding to Brodmannrsquos area 44 (Figure 7C) Thisregion has previously been associated with frontalneglect (Husain amp Kennard 1996) and is consideredto be part of the ventral premotor cortex and thehomologue of Brocarsquos areas in the left hemisphere(Rizzolatti amp Arbib 1998 Jackson amp Husain 1996) Inthese frontal patients the probability of re-clicking did
Mannan et al 349
not increase with time interval between the first clickand re-fixation but rather it stayed constant or declinedwith increasing delays
Such behavior might be consistent with a failure toinhibit responses to rightward locationsmdasha type oflsquolsquoperseverativersquorsquo behavior Note though that this is notnecessarily the same type of perseverative responseas seen when some patients with neglect re-markpreviously found items on standard cancellation tests(Rusconi et al 2002 Na et al 1999) In those casestargets that are visibly cancelled are often marked againwhereas in our computerized search tasks no marks arevisibly left on targets once they have been clicked uponMoreover only one of our neglect patients (Case 1) de-monstrated re-cancellation of visibly cancelled targetson pen-and-paper tasks and then only on one occasionImportantly this patient actually had a normal re-clickrate (67) on our task and his stroke was in the PCAterritory sparing both the intraparietal sulcus and fron-tal lobe
In support of a role for the right frontal lobe in in-hibiting responses to previously found targets recentimaging studies of response inhibition have highlightedregions of the right inferior frontal gyrus (IFG) very closeto the centroid of damage for our frontal patients (38 1618 cf 43 22 21 Konishi Kyoichi Uchida Sekihara ampMiyashita 1998 and 41 16 19 Konishi Kyoichi UchidaKikyo et al 1999) Alternative explanations for the highprobability of re-clicking directly after clicking might bethat these patients have impaired perceptual discrimina-tion of targets impaired perceptual localization of targetsor that they encounter difficulty in encoding spatiallocations into working memory Our data are not sugges-tive of a perceptual discrimination problem as changingthe perceptual load did not inf luence re-click ratesMoreover the patients still had high re-click rates in thetarget only conditions (Figure 10) where no perceptualdiscrimination of targets and distractors was necessaryPoor perceptual localization would seem unlikely giventhat these patients have frontal lesions Furthermoreusing a vertical SWM task similar to the Corsi blocktapping task (Malhotra et al 2004) we have found thatpatients with neglect can localize and perceptually en-code single target locations as well as controls and aresignificantly impaired at remembering two or more loca-tions shown in sequence consistent with the presentobservations Some recent functional imaging data sug-gest that activation within the prefrontal cortex corre-lates well with successful encoding of material intoworking memory (Rypma amp DrsquoEsposito 2003) Furtherresearch is required to distinguish between these pos-sible explanations for the mechanisms underlying there-click deficit in frontal patients
The only patient with frontal damage who did re-clickmore frequently with increasing time had damage cen-tered on the dorsolateral prefrontal cortex (althoughalso encroaching on the IFG) Neurons in the dorsolat-
eral prefrontal cortex like those in the LIP encode theremembered locations of saccadic targets (Goldman-Rakic 1996) This patient (Case 15) who had the high-est re-click rate of all also had damage to the criticalparietal location described above (in contrast to all otherfrontal patients) Thus she may have suffered from acombination of underlying deficits
Re-fixations and Misjudging PreviouslyInspected Locations during Search
In this search task re-fixation and re-click rates weresignificantly correlated for neglect patients Howeverthere were important dissociations within individuals 2patients with PCA lesions had re-fixation rates of 220and 80 respectively but both had a re-click rate ofonly 7 These are clearly examples of individuals whodo not show pathological re-clicking despite having highre-fixation rates High re-fixation rates may result froma lack of integration of spatial information due to V1damage (Zihl amp von Cramon 1985) in our PCA patientsor possibly because of reduced inhibition of return(Bartolomeo Sieroff Decaix amp Chokron 2001) What-ever the exact cause for dissociations between re-fixa-tions and re-click rates our data suggest re-fixation doesnot necessarily index the same underlying deficits asrevealed by our re-click measure High re-fixation ratesunlike high re-click rates were also not significantlyassociated with damage to any specific cortical locationagain demonstrating a dissociation between re-fixationsand pathological re-clicking
The relationship between re-click rates and neglect isunlikely to be a simple one In our study there was amoderate but significant correlation between the twomeasures suggesting that this association is unlikely tohave arisen by chance However non-neglect patientscan also exhibit a high re-click rate (unpublished obser-vations from our laboratory) Taken together our resultssuggest that difficulties in keeping track of previouslyinspected targets (because of impaired trans-saccadic re-mapping andor memory) may be one contributingcomponent to visual neglect in patients with lesions in-volving the right intraparietal sulcus As mentioned thesearch task we used differed in one striking aspect fromstandard cancellation tests subjects do not visibly markthe targets they visit Whereas in cancellation tests pa-tients mark each target they find which might appear todispense with any need to remember which locationshave been searched before However it should be ap-preciated that in most standard cancellation tests theindividual marks made by patients may not be clearlyvisible in peripheral vision particularly if neglect pa-tients have a local spatial bias (Rafal 1994 RobertsonLamb amp Knight 1988) or a restricted field of view(Russell Malhotra amp Husain 2004) Thus when com-bined with a rightward attentional bias keeping track ofsearched locations may be critical to prevent gaze from
350 Journal of Cognitive Neuroscience Volume 17 Number 2
returning to rightward locations even if they have beenmarked before as the marks may not be visible untilgaze returns to them Consistent with this hypothesisneglect is greater when cancellation is performed usinginvisible marks compared to large and salient red marksthat can be seen peripherally (Wojciulik Husain Clarkeamp Driver 2001)
A task like the present search-and-click paradigm maymore closely resemble search in the real world than dotraditional cancellation tests When we examine a visualscene we do not leave marks on each object we ex-amine Instead just as in the search task used in thisstudy we have to keep track of the locations we visit ifwe are to avoid inefficient re-examination of thoselocations Neglect patients with damage to the rightintraparietal sulcus appear to be impaired at keepingtrack of searched locations in this way When combinedwith their rightward attentional bias we propose thatsuch an impairment leads to recursive search of stimulito the right in experimental search tasks as used hereand also in daily life where neglect is so apparentdespite unlimited viewing time
The paradigm we have used has the important virtuesof allowing patients to explore freelymdashas in real-worldsearch or clinical cancellation tasksmdashand relating theirre-click behavior directly to the severity of neglect in thesame task But it might be argued that it would be usefulalso to probe SWM in a more controlled manner by re-stricting observersrsquo eye movements to a specific set oflocations and probing memory of previously visitedplaces A recent study using a gaze-contingent para-digm has elegantly probed the capacity of spatial mem-ory during search in healthy observers and foundevidence for memory for up to four previous locations(McCarley Wang Kramer Irwin amp Peterson 2003)Variants of this paradigm may in the future be usefulto probe the capacity of SWM across saccades in patientswith neglect and also to probe further whether thereare differences between neglect patients with parietaland frontal lesions
METHODS
Subjects Imaging and Lesion Mapping
Sixteen patients with left neglect following right hemi-sphere infarction or hemorrhage were recruited Con-sent was obtained according to the Declaration ofHelsinki as approved by the hospital ethical commit-tees Presence of neglect was determined by clinicalobservation plus bedside tests including the Mesulamshape-cancellation task (Mesulam 1985) and line bisec-tion task (Wilson et al 1987) (see Table 1) Twelvepatients had MCA territory strokes the rest involvedonly the PCA territory which can also result in neglect(Cals et al 2002) Only one neglect patient (Case 1 whodid not have frontal involvement) demonstrated re-
cancellation of visibly cancelled targets on pen-and-paper tasks and then only on one occasion Fifteenage-matched healthy volunteers were controls (meanage = 63 SD = 14 vs 63 SD = 13 for patients)
All patients had clinical CT scans In addition dedi-cated high-resolution structural MRI scans were alsoobtained for 1416 neglect patients MRI scans wereunobtainable for the other two (Cases 11 and 16)because of claustrophobia or loss to follow up In theMCA neglect group median time between stroke andscan was 63 days with the shortest interval being35 days for the PCA neglect group one patient had tobe scanned relatively early at 9 days but the mediantime was 140 days MRI scans comprised one 256-sliceT1-weighted acquisition (MPRAGE TR 97 msec TE4 msec TA 12 min) performed in the coronal planealigned perpendicular to the anteriorndashposterior commis-sural line on a Siemens Vision 15-T scanner Within-and between-slice resolution was 1 mm For each pa-tient the area of damage was determined by detailedvisual inspection of the digital image for every singleslice Lesion delineation was performed by neurologists(authors D M and M H) by tracing by hand theboundary of the lesion onto every single 1-mm-thickaxial slice using a graphics tablet (WACOM Intuos A6)Combining all the slices then led to a 3-D region of in-terest (ROI) for each patient For one patient (Case 15)there were two areas of discrete focal damage one inthe frontal cortex and the other in the parietal cortex
Normalization of each MRI including the 3-D ROI tothe T1-template was performed with SPM99 (wwwfilionuclacukspm) The ROI was used as a mask for thelesion during determination of alignment parameters tominimize the contribution of abnormal brain to thisprocess (Rorden amp Brett 2000) The normalized ROIof each neglect patient was superimposed upon the T1template to find areas of maximal lesion overlap in eachgroup (see Results) as well as lesion volume usingMRIcro shareware (Rorden amp Brett 2000)
Experimental Procedure
Stimuli were presented on an SVGA monitor (29 22 in)viewed at 57 cm distance Each search display com-prised a virtual 9 7 grid each cell containing a single18 elementmdasheither a letter T (target) or a letter L (dis-tractor) (see Figure 1) The position of each elementwithin each cell was jittered with 19 targets and 44distractors randomly distributed over the virtual gridThe number of targets on the right and left halves ofthe display was balanced across trials Seven neglectpatients (Cases 5 8 10 11 14ndash16) were further testedon lsquolsquoTarget onlyrsquorsquo displays which had varying numbersof Ts (2 6 or 19) now with no L-shaped distractorsThe presentations of these lsquolsquoTarget onlyrsquorsquo screens wereinterspersed with those of the original search screens inrandom order in the same experimental session
Mannan et al 351
Eye movements were recorded using a video-basedpupil tracker with a temporal resolution of 4 msec andspatial accuracy of lt058 (EyeLink Sensorimotoric Sys-tems Berlin Germany) Subjects were asked to keeptheir head still The experimenter also monitored gazeposition on-line At the beginning of each session (andafter breaks in testing or following any unintended headmovement that disrupted gaze recordings) eye-trackingwas re-calibrated Each trial was initiated by the exper-imenter and began with a central fixation cross for 1 secthen the search display
Subjects were instructed to search for Ts (among anyLs) Critically while looking at any target they had toclick a response button held in the right hand onlyif they considered this target to be a new discovery(ie a target they had not clicked on before) Clickingthe response button did not change the display Theresponse button was held by the patient in a comfort-able position to the right of the midline Subjectswere instructed and reminded throughout testing toclick on each target found once only Any later re-click on
a previously found target thus indicates a failure to retainthe fact that this target had already been discoveredThe experimenter terminated each search when thesubject indicated verbally that they felt all targets in thedisplay had now been found Each subject searched aminimum of 5 displays (mean = 7 range 5ndash14) Eyemovements were analyzed off-line using custom soft-ware An automatic algorithm was used to extract sac-cades (saccade onset defined as eye speed rising above258 sec1 for two or more samples and a criterion ofminimum fixation duration of 60 msec) Blinks wereeliminated from the trace by visual inspection and sac-cade and fixation delineation were checked visually inthe record Next fixations were automatically assignedto items in the display if the mean position fell within18 of that item Two or more consecutive fixations onthe same item were re-scored as a single extendedfixation A re-fixation was scored only when at leastone fixation elsewhere on another item intervened
Most click responses were made during a fixation andthe few button-clicks occurring during a saccade were
Table 1 Patient Details
CaseAge
(years) PathologyVolume(cm3)
Age ofLesion (days)
Visual FieldDefect
Mean Deviationfrom Center on
Line Bisection Task(cm) (rightward +)
Mesulum Score(Maximum Possible
Score = 60) Lesion Type
1 68 Infarct 1282 64 + 867 17 PCA
2 76 Infarct 1528 227 + 003 41 PCA
3 68 Infarct 79 207 + 297 36 PCA
4 77 Infarct 251 158 + 150 54 PCA
5 63 Infarct 28 29 033 55 MCA
6 59 Hemorrhage 172 7 + 32 6 MCA
7 66 Hemorrhage 273 325 + 213 31 MCA
8 68 Infarct 309 3 25 17 MCA
9 29 Infarct 115 17 113 55 MCA
10 70 Infarct 1339 324 153 57 MCA
11 42 Infarct ndash 11 097 24 MCA
12 75 Infarct 757 38 013 4 MCA
13 51 Infarct 694 141 083 31 MCA
14 59 Infarct 1144 356 + 223 37 MCA
15 80 Infarct 261 44 35 6 MCA
16 64 Infarct ndash 30 033 55 MCA
MEAN 63 66 124 188 33
SE 4 13 32 5
Cases 1ndash9 had damage to the nonfrontal (posterior only) cortex whereas Cases 10ndash15 had both frontal and posterior (posterior and anterior) corticalinvolvement Case 16 is a patient with a subcortical lesion All but 3 of the patients with right MCA infarction had intact visual fields on confrontationTwo of these (Cases 7 and 14) were diagnosed with left homonymous hemianopia whereas one (Case 6) had a left inferior quandrantic field defectAll patients with PCA territory damage had partial or complete contralesional visual field loss No patient suffered from dysphasia
352 Journal of Cognitive Neuroscience Volume 17 Number 2
assigned to the previous fixation Clicks were assigned toa particular item if eye position at the time of the clickwas within 28 of that item or in the cases where morethan one item was within 28 of current eye position theclick response was simply attributed to the item closestto that eye position
Button-clicks were assigned to a particular item if eyeposition at the time of the click was within 28 of thatitem A re-click on an item required at least one fixationon another item between successive clicks (in practiceseveral fixations elsewhere typically intervened betweeninitial click and later re-click see Results)
Statistical analyses were performed using SPSS andSigmaplot (SPSS Science Chicago USA) Jonckheere sta-tistic was derived by hand Statistical significance was setas p lt 05
Acknowledgments
We thank the reviewers of this manuscript and Dr AndrewParton for their extremely helpful comments We thank thepatients who participated Dr J Wade Dr A Rudd and thestaff of the stroke units at Charing Cross and St ThomasrsquoHospitals This research was supported by grants from theWellcome Trust to M H C K and J D J D holds a RoyalSociety Wolfson Research Merit Award
Reprint requests should be sent to Drs Sabira Mannan andMasud Husain Division of Neuroscience and PsychologicalMedicine Imperial College London Charing Cross CampusLondon W6 8RF UK or via e-mail smannanimperialacukor mhusainimperialacuk
Notes
1 We are grateful to one of the reviewers for suggesting thistime-series analysis2 There was no significant difference between the number oftargets fixated between the two groups so differences in lsquolsquoclickomissionsrsquorsquo cannot be attributed to the number of fixatedtargets that had to be kept track of during search3 The correlation between these four points alone inFigure 9A is high [r(3) = 81 cf r(15) = 47 for the wholegroup]4 We have also tested 10 neglect patients and 10 controlstroke patients on a vertical version of the Corsi blocks task(clinically used to measure spatial working memory capacity)Neglect patients had significantly impaired capacity but wereable to localize single locations well suggesting that mislocal-ization is unlikely to account for their deficit on this spatialworking memory task
REFERENCES
Bartolomeo P Sieroff E Decaix C amp Chokron S (2001)Modulating the attentional bias in unilateral neglect theeffects of the strategic set Experimental Brain Research137 432ndash444
Bates E Wilson S M Saygin A P Dick F Sereno M IKnight R T amp Dronkers N F (2003) Voxel-basedlesion-symptom mapping Nature Neuroscience 6 448ndash450
Behrmann M Watt S Black S E amp Barton J J (1997)Impaired visual search in patients with unilateral neglect
An oculographic analysis Neuropsychologia 351445ndash1458
Bisiach E amp Vallar G (2000) Unilateral neglect in humans InG Rizzolatti (Ed) Handbook of neuropsychology (2nd edvol 1 pp 459ndash502) Amsterdam Elsevier
Cals N Devuyst G Afsar N Karapanayiotides T ampBogousslavsky J (2002) Pure superficial posterior cerebralartery territory infarction in The Lausanne Stroke RegistryJournal of Neurology 249 855ndash861
Chafee M amp Goldman-Rakic P (1998) Matching patterns ofactivity in primate prefrontal area 8a and parietal area 7ipneurons during a spatial working memory task Journal ofNeurophysiology 79 2919ndash2940
Chedru F Leblanc M amp Lhermitte F (1973) Visualsearching in normal and brain-damaged subjectsContributions to the study of unilateral subjectsCortex 9 94ndash111
DrsquoEsposito M Aguirre G K Zarahn E Ballard DShin R K amp Lease J (1998) Functional MRI studies ofspatial and nonspatial working memory Cognitive BrainResearch 7 1ndash13
Donkelaar P amp Muri R (2002) Craniotopic updating ofvisual space across saccades in the human posteriorparietal cortex Proceedings of the Royal Society London B269 735ndash739
Driver J amp Husain M (2002) The role of spatial workingmemory deficits in pathological search by neglect patientsIn the cognitive and neural bases of spatial neglect InH-O Karnath A D Milner amp G Vallar (Eds) The cognitiveand neural bases of spatial neglect Oxford OxfordUniversity Press
Duhamel J R Colby C amp Goldberg M E (1992) Theupdating of the representation of visual space in parietalcortex by intended eye movements Science 25590ndash92
Goldman-Rakic P S (1996) The prefrontal landscapemdashImplications of functional architecture for understandinghuman mentation and the central executive PhilosophicalTransactions of The Royal Society of London Series BBiological Sciences 351 1445ndash1453
Heide W Binkofski F Seitz R J Posse S Nitschke M FFreund H J amp Kompf D (2001) Activation offrontoparietal cortices during memorized triple-stepsequences of saccadic eye movements An fMRI studyEuropean Journal of Neuroscience 13 1177ndash1189
Heide W amp Kompf D (1998) Combined deficits ofsaccades and visuo-spatial orientation after cortical lesionsExperimental Brain Research 123 164ndash171
Husain M amp Kennard C (1996) Visual neglect associatedwith frontal lobe infarction Journal of Neurology 243652ndash657
Husain M Mannan S Hodgson T Wojciulik E Driver Jamp Kennard C (2001) Impaired spatial working memoryacross saccades contributes to abnormal search in parietalneglect Brain 121 941ndash952
Husain M amp Rorden C (2003) Non-spatially lateralizedmechanisms in hemispatial neglect Nature ReviewsNeuroscience 4 23ndash36
Jackson S R amp Husain M (1996) Visuomotor functionsof the lateral pre-motor cortex Current Opinion inNeurobiology 6 788ndash795
Kinsbourne M (1993) Orientational bias model of unilateralneglect Evidence from attentional gradients withinhemispace In I H Robertson amp J C Marshall (Eds)Unilateral neglect Clinical and experimental studies(pp 63ndash86) Hove Erlbaum
Konishi S Kyoichi N Uchida I Kikyo H Kameyama Mamp Miyauchi S (1999) Common inhibitory mechanism in
Mannan et al 353
human inferior prefrontal cortex revealed by event-relatedfunctional MRI Brain 122 981ndash991
Konishi S Kyoichi N Uchida I Sekihara K amp Miyashita Y(1998) No-go dominant brain activity in human inferiorprefrontal cortex revealed by functional magneticresonance imaging European Journal of Neuroscience10 1209ndash1213
Malhotra P Mannan S Driver J amp Husain M (2004)Impaired spatial working memory One component of thevisual neglect syndrome Cortex 40 667ndash676
Manly T Woldt K Watson P amp Warburton E (2002) Ismotor perseveration in unilateral neglect lsquolsquodrivenrsquorsquo by thepresence of neglected left-sided stimuli Neuropsychologia40 1794ndash1803
Mazzoni P Bracewell R Barash S amp Andersen R (1996)Spatially tuned auditory responses in area LIP of macaquesperforming delayed memory saccades to acoustic targetsJournal of Neurophysiology 75 1233ndash1241
McCarley J Wang R Kramer A Irwin D amp Peterson M(2003) How much memory does oculomotor search havePsychological Science 14 422ndash426
Mesulam M-M (1985) Principles of behavioural neurologyTests of directed attention and memory Philadelphia Davis
Mesulam M M (1999) Spatial attention and neglectParietal frontal and cingulate contributions to the mentalrepresentation and attentional targeting of salientextrapersonal events Philosophical Transactions of theRoyal Society of London Series B Biological Sciences354 1325ndash1346
Mort D Malhotra P Mannan S Rorden C Pambakian AKennard C amp Husain M (2003) The anatomy of visualneglect Brain 126 1986ndash1997
Na D L Adair J C Kang Y Chung C S Lee K H ampHeilman K M (1999) Motor perseverative behaviour ona line cancellation task Neurology 52 1569ndash1576
Pearson D G amp Sahraie A (2003) Oculomotor controland the maintenance of spatially and temporally distributedevents in visuo-spatial working memory QuarterlyJournal of Experimental Psychology 56A 1089ndash1111
Rafal R (1994) Neglect Current Opinion in Neurobiology4 231ndash236
Rizzolatti G amp Arbib M A (1998) Language within our graspTrends in Neurosciences 21 188ndash194
Robertson L C Lamb M R amp Knight R T (1988) Effectsof lesions of temporalndashparietal junction on perceptual andattentional processing in humans Journal of Neuroscience8 3757ndash3769
Rorden C amp Brett M (2000) Stereotaxic display of brainlesions Behavioural Neurology 12 191ndash200
Rusconi M L Maravita A Bottini G amp Vallar G (2002)Is the intact side really intact Perseverative responses inpatients with unilateral neglect A productive manifestationNeuropsychologia 40 594ndash604
Russell C Malhotra P amp Husain M (2004) Attentionmodulates the visual field in healthy observers and parietalpatients NeuroReport 15 189ndash193
Rypma B amp DrsquoEsposito M (2003) A subsequent-memoryeffect in dorsolateral prefrontal cortex Cognitive BrainResearch 16 162ndash166
Sapir A Hayes A Henik A Danzinger S amp Rafal R(2004) Parietal lobe lesions disrupt saccadic remapping ofinhibitory location tagging Journal of CognitiveNeuroscience 16 503ndash509
Smania N Martini M C Gambina G A T Palamara ANatale E amp Marzi C A (1998) The spatial distributionof visual attention in hemineglect and extinction patientsBrain 121 1759ndash1770
Sprenger A Kompf D amp Heide W (2002) Visual searchin patients with left visual hemineglect Progress in BrainResearch 140 395ndash416
Tobler P Felblinger J Burki M Nirkko A Ozdoba C ampMuri R (2001) Functional organisation of the saccadicreference system processing extraretinal signals in humansVision Research 41 1351ndash1358
Wilson B Cockburn J amp Halligan P W (1987)Behavioural inattention test Bury St EdmundsThames Valley
Wojciulik E Husain M Clarke K amp Driver J (2001)Spatial working memory deficit in unilateral neglectNeuropsychologia 39 390ndash396
Zihl J amp Hebel N (1997) Patterns of oculomotor scanningin patients with unilateral posterior parietal and frontallobe damage Neuropsychologia 35 893ndash906
Zihl J amp von Cramon D (1985) Visual field recovery fromscotoma in patients with postgeniculate damage A reviewof 55 cases Brain 108 335ndash365
354 Journal of Cognitive Neuroscience Volume 17 Number 2
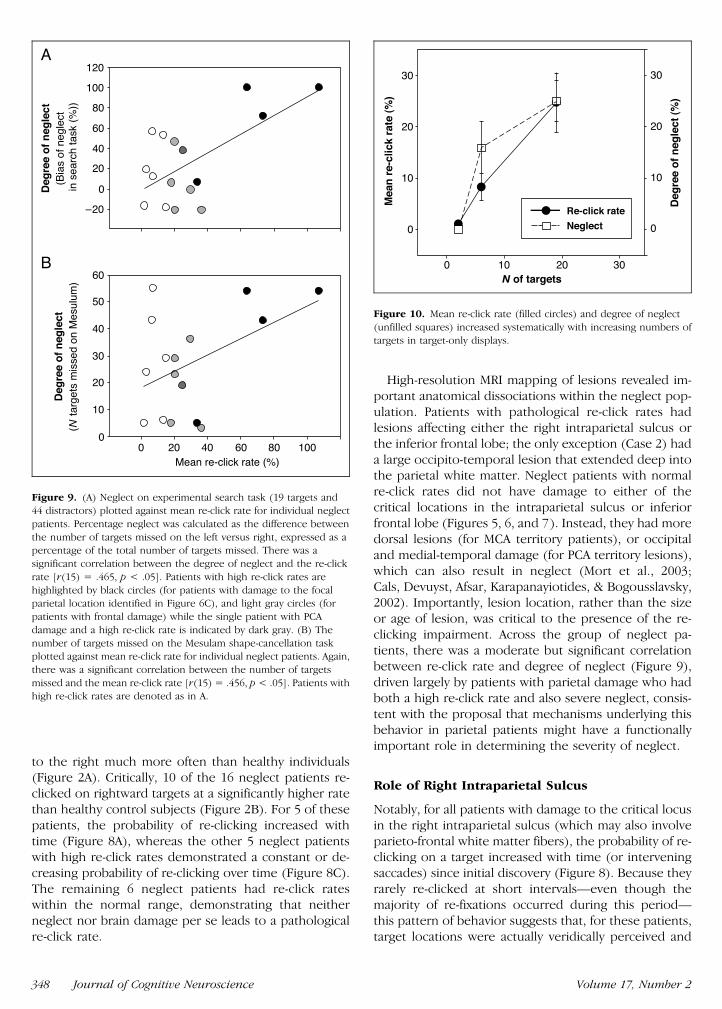
to the right much more often than healthy individuals(Figure 2A) Critically 10 of the 16 neglect patients re-clicked on rightward targets at a significantly higher ratethan healthy control subjects (Figure 2B) For 5 of thesepatients the probability of re-clicking increased withtime (Figure 8A) whereas the other 5 neglect patientswith high re-click rates demonstrated a constant or de-creasing probability of re-clicking over time (Figure 8C)The remaining 6 neglect patients had re-click rateswithin the normal range demonstrating that neitherneglect nor brain damage per se leads to a pathologicalre-click rate
High-resolution MRI mapping of lesions revealed im-portant anatomical dissociations within the neglect pop-ulation Patients with pathological re-click rates hadlesions affecting either the right intraparietal sulcus orthe inferior frontal lobe the only exception (Case 2) hada large occipito-temporal lesion that extended deep intothe parietal white matter Neglect patients with normalre-click rates did not have damage to either of thecritical locations in the intraparietal sulcus or inferiorfrontal lobe (Figures 5 6 and 7) Instead they had moredorsal lesions (for MCA territory patients) or occipitaland medial-temporal damage (for PCA territory lesions)which can also result in neglect (Mort et al 2003Cals Devuyst Afsar Karapanayiotides amp Bogousslavsky2002) Importantly lesion location rather than the sizeor age of lesion was critical to the presence of the re-clicking impairment Across the group of neglect pa-tients there was a moderate but significant correlationbetween re-click rate and degree of neglect (Figure 9)driven largely by patients with parietal damage who hadboth a high re-click rate and also severe neglect consis-tent with the proposal that mechanisms underlying thisbehavior in parietal patients might have a functionallyimportant role in determining the severity of neglect
Role of Right Intraparietal Sulcus
Notably for all patients with damage to the critical locusin the right intraparietal sulcus (which may also involveparieto-frontal white matter fibers) the probability of re-clicking on a target increased with time (or interveningsaccades) since initial discovery (Figure 8) Because theyrarely re-clicked at short intervalsmdasheven though themajority of re-fixations occurred during this periodmdashthis pattern of behavior suggests that for these patientstarget locations were actually veridically perceived and
Figure 9 (A) Neglect on experimental search task (19 targets and44 distractors) plotted against mean re-click rate for individual neglect
patients Percentage neglect was calculated as the difference between
the number of targets missed on the left versus right expressed as apercentage of the total number of targets missed There was a
significant correlation between the degree of neglect and the re-click
rate [r(15) = 465 p lt 05] Patients with high re-click rates are
highlighted by black circles (for patients with damage to the focalparietal location identified in Figure 6C) and light gray circles (for
patients with frontal damage) while the single patient with PCA
damage and a high re-click rate is indicated by dark gray (B) The
number of targets missed on the Mesulam shape-cancellation taskplotted against mean re-click rate for individual neglect patients Again
there was a significant correlation between the number of targets
missed and the mean re-click rate [r(15) = 456 p lt 05] Patients withhigh re-click rates are denoted as in A
Figure 10 Mean re-click rate (filled circles) and degree of neglect
(unfilled squares) increased systematically with increasing numbers of
targets in target-only displays
348 Journal of Cognitive Neuroscience Volume 17 Number 2
well encoded into working memory Furthermore de-tailed single-case studies of two parietal patients withhigh re-click rates (Malhotra Mannan Driver amp Husain2004 Husain Mannan et al 2001) demonstrate usingdifferent psychophysical methods that these patientscan accurately encode spatial locations4 Increasing re-clicks with time (or intervening saccades) would how-ever be consistent with a deficit in keeping track oftarget locations during extended search
Several lines of evidence support the proposal thatthe critical intraparietal sulcus region we have identifiedmay have an important role in the spatial transforma-tions required to keep track of spatial locations acrosssaccades In rhesus monkeys neurons within the lateralbank of the intraparietal sulcus (area LIP) encode andlsquolsquomaintainrsquorsquo the remembered locations of saccadic tar-gets over time (Chafee amp Goldman-Rakic 1998 MazzoniBracewell Barash amp Andersen 1996) Some visuallyresponsive neurons in the LIP also appear to shift theirretinotopic representation at the onset of an intendedsaccade as if lsquolsquore-mappingrsquorsquo representations of objects inspace to update them across shifts of gaze (DuhamelColby amp Goldberg 1992)
In humans this computation has been investigatedusing the double-step saccade paradigm where twoflashes of light appear in quick succession such thatthe second target disappears before a saccade can bemade to the first target Patients with lesions involvingthe right parietal cortex are impaired at performing thistask suggesting that they are unable to update spatialrepresentations accurately after even one saccade (Heideamp Kompf 1998 Duhamel et al 1992) Similarly trans-cranial magnetic stimulation of the posterior parietalcortex prior to the second saccade reduces its accuracy(Donkelaar amp Muri 2002) and a recent study of patientswith parietal lesions also implicates this area in thespatial updating of inhibition of return after a saccade(Sapir Hayes Henik Danzinger amp Rafal 2004) Func-tional imaging studies of double-step (Tobler et al2001) and triple-step memory-guided saccades (HeideBinkofski et al 2001) have identified a region withinthe intraparietal sulcus (Talairach coordinates 40 4044 for Tobler et al and 44 48 36 Heide et al) This isclose to the critical region of lesion overlap we haveidentified in the posterior neglect patients with high re-click rates (Talairach 37 39 37)
We suggest that in our patients with lesions involvingthe right intraparietal sulcus uncertainty about locationsof previously found targets increases with each saccadeThis uncertainty is further exacerbated by interferencefrom newer target locations leading to increased prob-ability of misjudging old target locations for new assearch progresses Such progressively worsening uncer-tainty might be due to an accumulating impairment inlsquolsquore-mapping target locationsrsquorsquo across saccades or im-paired lsquolsquomemoryrsquorsquo of previously inspected target loca-tions across saccades As noted above there is evidence
for both types of signalmdash re-mapping and maintainingsaccadic spatial memorymdashin the parietal cortex and it ispossible that disruption to both these functions contrib-utes to the re-click deficit we have identified Certainlythe two potential types of deficit are not mutuallyincompatible
A slightly different account of the re-clicking behaviormight be framed in terms of the intriguing hypothesisproposed by Manly Woldt Watson and Warburton(2002) to explain visible re-marking on a standard can-cellation task They argue that such behavior is due topatients erroneously transposing leftward stimuli to theright resulting in apparent re-cancellation These au-thors found that removing targets from the left re-duced both neglect and re-marking However they didnot monitor eye movements so it is possible that sometargets that are logged as neglected (because they werenot cancelled) were nevertheless fixated and thereforeform part of the load on memory during search Fur-thermore in our experimental study subjects with ahigh re-click rate also failed to click on a higher pro-portion of fixated targets Manly et alrsquos proposal wouldnot easily account for such lsquolsquoclick omissionsrsquorsquo becausethese refer to fixated targets that were not actuallyresponded to
Finally it might be argued that increasing re-clickingwith search duration reflects a deficit in spatial workingmemory (SWM) that is independent of saccadic eyemovements (ie a decay in memory that is not trans-saccadic) Because time and number of interveningsaccades were not independently manipulated in thisunrestricted visual search paradigm we are not ablecategorically to distinguish between these possibilitiesWe note however that functional imaging studies ofSWM tasks that do not require eye movements oftendemonstrate activity related to the maintenance ofspatial information which is generally more posteriorand dorsal than the parietal locations isolated in thisstudy (DrsquoEsposito et al 1998) Moreover recent behav-ioral data indicate that SWM is more vulnerable tosaccadic eye movements than shifts in attention or limbmovements (Pearson amp Sahraie 2003)
Frontal Lesion Site Associated withHigh Re-click Rates
Four out of 5 neglect patients with lesions involvingposterior plus anterior damage also showed a high re-click rate and the area of damage common to thesepatients centered on the right inferior frontal lobe cor-responding to Brodmannrsquos area 44 (Figure 7C) Thisregion has previously been associated with frontalneglect (Husain amp Kennard 1996) and is consideredto be part of the ventral premotor cortex and thehomologue of Brocarsquos areas in the left hemisphere(Rizzolatti amp Arbib 1998 Jackson amp Husain 1996) Inthese frontal patients the probability of re-clicking did
Mannan et al 349
not increase with time interval between the first clickand re-fixation but rather it stayed constant or declinedwith increasing delays
Such behavior might be consistent with a failure toinhibit responses to rightward locationsmdasha type oflsquolsquoperseverativersquorsquo behavior Note though that this is notnecessarily the same type of perseverative responseas seen when some patients with neglect re-markpreviously found items on standard cancellation tests(Rusconi et al 2002 Na et al 1999) In those casestargets that are visibly cancelled are often marked againwhereas in our computerized search tasks no marks arevisibly left on targets once they have been clicked uponMoreover only one of our neglect patients (Case 1) de-monstrated re-cancellation of visibly cancelled targetson pen-and-paper tasks and then only on one occasionImportantly this patient actually had a normal re-clickrate (67) on our task and his stroke was in the PCAterritory sparing both the intraparietal sulcus and fron-tal lobe
In support of a role for the right frontal lobe in in-hibiting responses to previously found targets recentimaging studies of response inhibition have highlightedregions of the right inferior frontal gyrus (IFG) very closeto the centroid of damage for our frontal patients (38 1618 cf 43 22 21 Konishi Kyoichi Uchida Sekihara ampMiyashita 1998 and 41 16 19 Konishi Kyoichi UchidaKikyo et al 1999) Alternative explanations for the highprobability of re-clicking directly after clicking might bethat these patients have impaired perceptual discrimina-tion of targets impaired perceptual localization of targetsor that they encounter difficulty in encoding spatiallocations into working memory Our data are not sugges-tive of a perceptual discrimination problem as changingthe perceptual load did not inf luence re-click ratesMoreover the patients still had high re-click rates in thetarget only conditions (Figure 10) where no perceptualdiscrimination of targets and distractors was necessaryPoor perceptual localization would seem unlikely giventhat these patients have frontal lesions Furthermoreusing a vertical SWM task similar to the Corsi blocktapping task (Malhotra et al 2004) we have found thatpatients with neglect can localize and perceptually en-code single target locations as well as controls and aresignificantly impaired at remembering two or more loca-tions shown in sequence consistent with the presentobservations Some recent functional imaging data sug-gest that activation within the prefrontal cortex corre-lates well with successful encoding of material intoworking memory (Rypma amp DrsquoEsposito 2003) Furtherresearch is required to distinguish between these pos-sible explanations for the mechanisms underlying there-click deficit in frontal patients
The only patient with frontal damage who did re-clickmore frequently with increasing time had damage cen-tered on the dorsolateral prefrontal cortex (althoughalso encroaching on the IFG) Neurons in the dorsolat-
eral prefrontal cortex like those in the LIP encode theremembered locations of saccadic targets (Goldman-Rakic 1996) This patient (Case 15) who had the high-est re-click rate of all also had damage to the criticalparietal location described above (in contrast to all otherfrontal patients) Thus she may have suffered from acombination of underlying deficits
Re-fixations and Misjudging PreviouslyInspected Locations during Search
In this search task re-fixation and re-click rates weresignificantly correlated for neglect patients Howeverthere were important dissociations within individuals 2patients with PCA lesions had re-fixation rates of 220and 80 respectively but both had a re-click rate ofonly 7 These are clearly examples of individuals whodo not show pathological re-clicking despite having highre-fixation rates High re-fixation rates may result froma lack of integration of spatial information due to V1damage (Zihl amp von Cramon 1985) in our PCA patientsor possibly because of reduced inhibition of return(Bartolomeo Sieroff Decaix amp Chokron 2001) What-ever the exact cause for dissociations between re-fixa-tions and re-click rates our data suggest re-fixation doesnot necessarily index the same underlying deficits asrevealed by our re-click measure High re-fixation ratesunlike high re-click rates were also not significantlyassociated with damage to any specific cortical locationagain demonstrating a dissociation between re-fixationsand pathological re-clicking
The relationship between re-click rates and neglect isunlikely to be a simple one In our study there was amoderate but significant correlation between the twomeasures suggesting that this association is unlikely tohave arisen by chance However non-neglect patientscan also exhibit a high re-click rate (unpublished obser-vations from our laboratory) Taken together our resultssuggest that difficulties in keeping track of previouslyinspected targets (because of impaired trans-saccadic re-mapping andor memory) may be one contributingcomponent to visual neglect in patients with lesions in-volving the right intraparietal sulcus As mentioned thesearch task we used differed in one striking aspect fromstandard cancellation tests subjects do not visibly markthe targets they visit Whereas in cancellation tests pa-tients mark each target they find which might appear todispense with any need to remember which locationshave been searched before However it should be ap-preciated that in most standard cancellation tests theindividual marks made by patients may not be clearlyvisible in peripheral vision particularly if neglect pa-tients have a local spatial bias (Rafal 1994 RobertsonLamb amp Knight 1988) or a restricted field of view(Russell Malhotra amp Husain 2004) Thus when com-bined with a rightward attentional bias keeping track ofsearched locations may be critical to prevent gaze from
350 Journal of Cognitive Neuroscience Volume 17 Number 2
returning to rightward locations even if they have beenmarked before as the marks may not be visible untilgaze returns to them Consistent with this hypothesisneglect is greater when cancellation is performed usinginvisible marks compared to large and salient red marksthat can be seen peripherally (Wojciulik Husain Clarkeamp Driver 2001)
A task like the present search-and-click paradigm maymore closely resemble search in the real world than dotraditional cancellation tests When we examine a visualscene we do not leave marks on each object we ex-amine Instead just as in the search task used in thisstudy we have to keep track of the locations we visit ifwe are to avoid inefficient re-examination of thoselocations Neglect patients with damage to the rightintraparietal sulcus appear to be impaired at keepingtrack of searched locations in this way When combinedwith their rightward attentional bias we propose thatsuch an impairment leads to recursive search of stimulito the right in experimental search tasks as used hereand also in daily life where neglect is so apparentdespite unlimited viewing time
The paradigm we have used has the important virtuesof allowing patients to explore freelymdashas in real-worldsearch or clinical cancellation tasksmdashand relating theirre-click behavior directly to the severity of neglect in thesame task But it might be argued that it would be usefulalso to probe SWM in a more controlled manner by re-stricting observersrsquo eye movements to a specific set oflocations and probing memory of previously visitedplaces A recent study using a gaze-contingent para-digm has elegantly probed the capacity of spatial mem-ory during search in healthy observers and foundevidence for memory for up to four previous locations(McCarley Wang Kramer Irwin amp Peterson 2003)Variants of this paradigm may in the future be usefulto probe the capacity of SWM across saccades in patientswith neglect and also to probe further whether thereare differences between neglect patients with parietaland frontal lesions
METHODS
Subjects Imaging and Lesion Mapping
Sixteen patients with left neglect following right hemi-sphere infarction or hemorrhage were recruited Con-sent was obtained according to the Declaration ofHelsinki as approved by the hospital ethical commit-tees Presence of neglect was determined by clinicalobservation plus bedside tests including the Mesulamshape-cancellation task (Mesulam 1985) and line bisec-tion task (Wilson et al 1987) (see Table 1) Twelvepatients had MCA territory strokes the rest involvedonly the PCA territory which can also result in neglect(Cals et al 2002) Only one neglect patient (Case 1 whodid not have frontal involvement) demonstrated re-
cancellation of visibly cancelled targets on pen-and-paper tasks and then only on one occasion Fifteenage-matched healthy volunteers were controls (meanage = 63 SD = 14 vs 63 SD = 13 for patients)
All patients had clinical CT scans In addition dedi-cated high-resolution structural MRI scans were alsoobtained for 1416 neglect patients MRI scans wereunobtainable for the other two (Cases 11 and 16)because of claustrophobia or loss to follow up In theMCA neglect group median time between stroke andscan was 63 days with the shortest interval being35 days for the PCA neglect group one patient had tobe scanned relatively early at 9 days but the mediantime was 140 days MRI scans comprised one 256-sliceT1-weighted acquisition (MPRAGE TR 97 msec TE4 msec TA 12 min) performed in the coronal planealigned perpendicular to the anteriorndashposterior commis-sural line on a Siemens Vision 15-T scanner Within-and between-slice resolution was 1 mm For each pa-tient the area of damage was determined by detailedvisual inspection of the digital image for every singleslice Lesion delineation was performed by neurologists(authors D M and M H) by tracing by hand theboundary of the lesion onto every single 1-mm-thickaxial slice using a graphics tablet (WACOM Intuos A6)Combining all the slices then led to a 3-D region of in-terest (ROI) for each patient For one patient (Case 15)there were two areas of discrete focal damage one inthe frontal cortex and the other in the parietal cortex
Normalization of each MRI including the 3-D ROI tothe T1-template was performed with SPM99 (wwwfilionuclacukspm) The ROI was used as a mask for thelesion during determination of alignment parameters tominimize the contribution of abnormal brain to thisprocess (Rorden amp Brett 2000) The normalized ROIof each neglect patient was superimposed upon the T1template to find areas of maximal lesion overlap in eachgroup (see Results) as well as lesion volume usingMRIcro shareware (Rorden amp Brett 2000)
Experimental Procedure
Stimuli were presented on an SVGA monitor (29 22 in)viewed at 57 cm distance Each search display com-prised a virtual 9 7 grid each cell containing a single18 elementmdasheither a letter T (target) or a letter L (dis-tractor) (see Figure 1) The position of each elementwithin each cell was jittered with 19 targets and 44distractors randomly distributed over the virtual gridThe number of targets on the right and left halves ofthe display was balanced across trials Seven neglectpatients (Cases 5 8 10 11 14ndash16) were further testedon lsquolsquoTarget onlyrsquorsquo displays which had varying numbersof Ts (2 6 or 19) now with no L-shaped distractorsThe presentations of these lsquolsquoTarget onlyrsquorsquo screens wereinterspersed with those of the original search screens inrandom order in the same experimental session
Mannan et al 351
Eye movements were recorded using a video-basedpupil tracker with a temporal resolution of 4 msec andspatial accuracy of lt058 (EyeLink Sensorimotoric Sys-tems Berlin Germany) Subjects were asked to keeptheir head still The experimenter also monitored gazeposition on-line At the beginning of each session (andafter breaks in testing or following any unintended headmovement that disrupted gaze recordings) eye-trackingwas re-calibrated Each trial was initiated by the exper-imenter and began with a central fixation cross for 1 secthen the search display
Subjects were instructed to search for Ts (among anyLs) Critically while looking at any target they had toclick a response button held in the right hand onlyif they considered this target to be a new discovery(ie a target they had not clicked on before) Clickingthe response button did not change the display Theresponse button was held by the patient in a comfort-able position to the right of the midline Subjectswere instructed and reminded throughout testing toclick on each target found once only Any later re-click on
a previously found target thus indicates a failure to retainthe fact that this target had already been discoveredThe experimenter terminated each search when thesubject indicated verbally that they felt all targets in thedisplay had now been found Each subject searched aminimum of 5 displays (mean = 7 range 5ndash14) Eyemovements were analyzed off-line using custom soft-ware An automatic algorithm was used to extract sac-cades (saccade onset defined as eye speed rising above258 sec1 for two or more samples and a criterion ofminimum fixation duration of 60 msec) Blinks wereeliminated from the trace by visual inspection and sac-cade and fixation delineation were checked visually inthe record Next fixations were automatically assignedto items in the display if the mean position fell within18 of that item Two or more consecutive fixations onthe same item were re-scored as a single extendedfixation A re-fixation was scored only when at leastone fixation elsewhere on another item intervened
Most click responses were made during a fixation andthe few button-clicks occurring during a saccade were
Table 1 Patient Details
CaseAge
(years) PathologyVolume(cm3)
Age ofLesion (days)
Visual FieldDefect
Mean Deviationfrom Center on
Line Bisection Task(cm) (rightward +)
Mesulum Score(Maximum Possible
Score = 60) Lesion Type
1 68 Infarct 1282 64 + 867 17 PCA
2 76 Infarct 1528 227 + 003 41 PCA
3 68 Infarct 79 207 + 297 36 PCA
4 77 Infarct 251 158 + 150 54 PCA
5 63 Infarct 28 29 033 55 MCA
6 59 Hemorrhage 172 7 + 32 6 MCA
7 66 Hemorrhage 273 325 + 213 31 MCA
8 68 Infarct 309 3 25 17 MCA
9 29 Infarct 115 17 113 55 MCA
10 70 Infarct 1339 324 153 57 MCA
11 42 Infarct ndash 11 097 24 MCA
12 75 Infarct 757 38 013 4 MCA
13 51 Infarct 694 141 083 31 MCA
14 59 Infarct 1144 356 + 223 37 MCA
15 80 Infarct 261 44 35 6 MCA
16 64 Infarct ndash 30 033 55 MCA
MEAN 63 66 124 188 33
SE 4 13 32 5
Cases 1ndash9 had damage to the nonfrontal (posterior only) cortex whereas Cases 10ndash15 had both frontal and posterior (posterior and anterior) corticalinvolvement Case 16 is a patient with a subcortical lesion All but 3 of the patients with right MCA infarction had intact visual fields on confrontationTwo of these (Cases 7 and 14) were diagnosed with left homonymous hemianopia whereas one (Case 6) had a left inferior quandrantic field defectAll patients with PCA territory damage had partial or complete contralesional visual field loss No patient suffered from dysphasia
352 Journal of Cognitive Neuroscience Volume 17 Number 2
assigned to the previous fixation Clicks were assigned toa particular item if eye position at the time of the clickwas within 28 of that item or in the cases where morethan one item was within 28 of current eye position theclick response was simply attributed to the item closestto that eye position
Button-clicks were assigned to a particular item if eyeposition at the time of the click was within 28 of thatitem A re-click on an item required at least one fixationon another item between successive clicks (in practiceseveral fixations elsewhere typically intervened betweeninitial click and later re-click see Results)
Statistical analyses were performed using SPSS andSigmaplot (SPSS Science Chicago USA) Jonckheere sta-tistic was derived by hand Statistical significance was setas p lt 05
Acknowledgments
We thank the reviewers of this manuscript and Dr AndrewParton for their extremely helpful comments We thank thepatients who participated Dr J Wade Dr A Rudd and thestaff of the stroke units at Charing Cross and St ThomasrsquoHospitals This research was supported by grants from theWellcome Trust to M H C K and J D J D holds a RoyalSociety Wolfson Research Merit Award
Reprint requests should be sent to Drs Sabira Mannan andMasud Husain Division of Neuroscience and PsychologicalMedicine Imperial College London Charing Cross CampusLondon W6 8RF UK or via e-mail smannanimperialacukor mhusainimperialacuk
Notes
1 We are grateful to one of the reviewers for suggesting thistime-series analysis2 There was no significant difference between the number oftargets fixated between the two groups so differences in lsquolsquoclickomissionsrsquorsquo cannot be attributed to the number of fixatedtargets that had to be kept track of during search3 The correlation between these four points alone inFigure 9A is high [r(3) = 81 cf r(15) = 47 for the wholegroup]4 We have also tested 10 neglect patients and 10 controlstroke patients on a vertical version of the Corsi blocks task(clinically used to measure spatial working memory capacity)Neglect patients had significantly impaired capacity but wereable to localize single locations well suggesting that mislocal-ization is unlikely to account for their deficit on this spatialworking memory task
REFERENCES
Bartolomeo P Sieroff E Decaix C amp Chokron S (2001)Modulating the attentional bias in unilateral neglect theeffects of the strategic set Experimental Brain Research137 432ndash444
Bates E Wilson S M Saygin A P Dick F Sereno M IKnight R T amp Dronkers N F (2003) Voxel-basedlesion-symptom mapping Nature Neuroscience 6 448ndash450
Behrmann M Watt S Black S E amp Barton J J (1997)Impaired visual search in patients with unilateral neglect
An oculographic analysis Neuropsychologia 351445ndash1458
Bisiach E amp Vallar G (2000) Unilateral neglect in humans InG Rizzolatti (Ed) Handbook of neuropsychology (2nd edvol 1 pp 459ndash502) Amsterdam Elsevier
Cals N Devuyst G Afsar N Karapanayiotides T ampBogousslavsky J (2002) Pure superficial posterior cerebralartery territory infarction in The Lausanne Stroke RegistryJournal of Neurology 249 855ndash861
Chafee M amp Goldman-Rakic P (1998) Matching patterns ofactivity in primate prefrontal area 8a and parietal area 7ipneurons during a spatial working memory task Journal ofNeurophysiology 79 2919ndash2940
Chedru F Leblanc M amp Lhermitte F (1973) Visualsearching in normal and brain-damaged subjectsContributions to the study of unilateral subjectsCortex 9 94ndash111
DrsquoEsposito M Aguirre G K Zarahn E Ballard DShin R K amp Lease J (1998) Functional MRI studies ofspatial and nonspatial working memory Cognitive BrainResearch 7 1ndash13
Donkelaar P amp Muri R (2002) Craniotopic updating ofvisual space across saccades in the human posteriorparietal cortex Proceedings of the Royal Society London B269 735ndash739
Driver J amp Husain M (2002) The role of spatial workingmemory deficits in pathological search by neglect patientsIn the cognitive and neural bases of spatial neglect InH-O Karnath A D Milner amp G Vallar (Eds) The cognitiveand neural bases of spatial neglect Oxford OxfordUniversity Press
Duhamel J R Colby C amp Goldberg M E (1992) Theupdating of the representation of visual space in parietalcortex by intended eye movements Science 25590ndash92
Goldman-Rakic P S (1996) The prefrontal landscapemdashImplications of functional architecture for understandinghuman mentation and the central executive PhilosophicalTransactions of The Royal Society of London Series BBiological Sciences 351 1445ndash1453
Heide W Binkofski F Seitz R J Posse S Nitschke M FFreund H J amp Kompf D (2001) Activation offrontoparietal cortices during memorized triple-stepsequences of saccadic eye movements An fMRI studyEuropean Journal of Neuroscience 13 1177ndash1189
Heide W amp Kompf D (1998) Combined deficits ofsaccades and visuo-spatial orientation after cortical lesionsExperimental Brain Research 123 164ndash171
Husain M amp Kennard C (1996) Visual neglect associatedwith frontal lobe infarction Journal of Neurology 243652ndash657
Husain M Mannan S Hodgson T Wojciulik E Driver Jamp Kennard C (2001) Impaired spatial working memoryacross saccades contributes to abnormal search in parietalneglect Brain 121 941ndash952
Husain M amp Rorden C (2003) Non-spatially lateralizedmechanisms in hemispatial neglect Nature ReviewsNeuroscience 4 23ndash36
Jackson S R amp Husain M (1996) Visuomotor functionsof the lateral pre-motor cortex Current Opinion inNeurobiology 6 788ndash795
Kinsbourne M (1993) Orientational bias model of unilateralneglect Evidence from attentional gradients withinhemispace In I H Robertson amp J C Marshall (Eds)Unilateral neglect Clinical and experimental studies(pp 63ndash86) Hove Erlbaum
Konishi S Kyoichi N Uchida I Kikyo H Kameyama Mamp Miyauchi S (1999) Common inhibitory mechanism in
Mannan et al 353
human inferior prefrontal cortex revealed by event-relatedfunctional MRI Brain 122 981ndash991
Konishi S Kyoichi N Uchida I Sekihara K amp Miyashita Y(1998) No-go dominant brain activity in human inferiorprefrontal cortex revealed by functional magneticresonance imaging European Journal of Neuroscience10 1209ndash1213
Malhotra P Mannan S Driver J amp Husain M (2004)Impaired spatial working memory One component of thevisual neglect syndrome Cortex 40 667ndash676
Manly T Woldt K Watson P amp Warburton E (2002) Ismotor perseveration in unilateral neglect lsquolsquodrivenrsquorsquo by thepresence of neglected left-sided stimuli Neuropsychologia40 1794ndash1803
Mazzoni P Bracewell R Barash S amp Andersen R (1996)Spatially tuned auditory responses in area LIP of macaquesperforming delayed memory saccades to acoustic targetsJournal of Neurophysiology 75 1233ndash1241
McCarley J Wang R Kramer A Irwin D amp Peterson M(2003) How much memory does oculomotor search havePsychological Science 14 422ndash426
Mesulam M-M (1985) Principles of behavioural neurologyTests of directed attention and memory Philadelphia Davis
Mesulam M M (1999) Spatial attention and neglectParietal frontal and cingulate contributions to the mentalrepresentation and attentional targeting of salientextrapersonal events Philosophical Transactions of theRoyal Society of London Series B Biological Sciences354 1325ndash1346
Mort D Malhotra P Mannan S Rorden C Pambakian AKennard C amp Husain M (2003) The anatomy of visualneglect Brain 126 1986ndash1997
Na D L Adair J C Kang Y Chung C S Lee K H ampHeilman K M (1999) Motor perseverative behaviour ona line cancellation task Neurology 52 1569ndash1576
Pearson D G amp Sahraie A (2003) Oculomotor controland the maintenance of spatially and temporally distributedevents in visuo-spatial working memory QuarterlyJournal of Experimental Psychology 56A 1089ndash1111
Rafal R (1994) Neglect Current Opinion in Neurobiology4 231ndash236
Rizzolatti G amp Arbib M A (1998) Language within our graspTrends in Neurosciences 21 188ndash194
Robertson L C Lamb M R amp Knight R T (1988) Effectsof lesions of temporalndashparietal junction on perceptual andattentional processing in humans Journal of Neuroscience8 3757ndash3769
Rorden C amp Brett M (2000) Stereotaxic display of brainlesions Behavioural Neurology 12 191ndash200
Rusconi M L Maravita A Bottini G amp Vallar G (2002)Is the intact side really intact Perseverative responses inpatients with unilateral neglect A productive manifestationNeuropsychologia 40 594ndash604
Russell C Malhotra P amp Husain M (2004) Attentionmodulates the visual field in healthy observers and parietalpatients NeuroReport 15 189ndash193
Rypma B amp DrsquoEsposito M (2003) A subsequent-memoryeffect in dorsolateral prefrontal cortex Cognitive BrainResearch 16 162ndash166
Sapir A Hayes A Henik A Danzinger S amp Rafal R(2004) Parietal lobe lesions disrupt saccadic remapping ofinhibitory location tagging Journal of CognitiveNeuroscience 16 503ndash509
Smania N Martini M C Gambina G A T Palamara ANatale E amp Marzi C A (1998) The spatial distributionof visual attention in hemineglect and extinction patientsBrain 121 1759ndash1770
Sprenger A Kompf D amp Heide W (2002) Visual searchin patients with left visual hemineglect Progress in BrainResearch 140 395ndash416
Tobler P Felblinger J Burki M Nirkko A Ozdoba C ampMuri R (2001) Functional organisation of the saccadicreference system processing extraretinal signals in humansVision Research 41 1351ndash1358
Wilson B Cockburn J amp Halligan P W (1987)Behavioural inattention test Bury St EdmundsThames Valley
Wojciulik E Husain M Clarke K amp Driver J (2001)Spatial working memory deficit in unilateral neglectNeuropsychologia 39 390ndash396
Zihl J amp Hebel N (1997) Patterns of oculomotor scanningin patients with unilateral posterior parietal and frontallobe damage Neuropsychologia 35 893ndash906
Zihl J amp von Cramon D (1985) Visual field recovery fromscotoma in patients with postgeniculate damage A reviewof 55 cases Brain 108 335ndash365
354 Journal of Cognitive Neuroscience Volume 17 Number 2

well encoded into working memory Furthermore de-tailed single-case studies of two parietal patients withhigh re-click rates (Malhotra Mannan Driver amp Husain2004 Husain Mannan et al 2001) demonstrate usingdifferent psychophysical methods that these patientscan accurately encode spatial locations4 Increasing re-clicks with time (or intervening saccades) would how-ever be consistent with a deficit in keeping track oftarget locations during extended search
Several lines of evidence support the proposal thatthe critical intraparietal sulcus region we have identifiedmay have an important role in the spatial transforma-tions required to keep track of spatial locations acrosssaccades In rhesus monkeys neurons within the lateralbank of the intraparietal sulcus (area LIP) encode andlsquolsquomaintainrsquorsquo the remembered locations of saccadic tar-gets over time (Chafee amp Goldman-Rakic 1998 MazzoniBracewell Barash amp Andersen 1996) Some visuallyresponsive neurons in the LIP also appear to shift theirretinotopic representation at the onset of an intendedsaccade as if lsquolsquore-mappingrsquorsquo representations of objects inspace to update them across shifts of gaze (DuhamelColby amp Goldberg 1992)
In humans this computation has been investigatedusing the double-step saccade paradigm where twoflashes of light appear in quick succession such thatthe second target disappears before a saccade can bemade to the first target Patients with lesions involvingthe right parietal cortex are impaired at performing thistask suggesting that they are unable to update spatialrepresentations accurately after even one saccade (Heideamp Kompf 1998 Duhamel et al 1992) Similarly trans-cranial magnetic stimulation of the posterior parietalcortex prior to the second saccade reduces its accuracy(Donkelaar amp Muri 2002) and a recent study of patientswith parietal lesions also implicates this area in thespatial updating of inhibition of return after a saccade(Sapir Hayes Henik Danzinger amp Rafal 2004) Func-tional imaging studies of double-step (Tobler et al2001) and triple-step memory-guided saccades (HeideBinkofski et al 2001) have identified a region withinthe intraparietal sulcus (Talairach coordinates 40 4044 for Tobler et al and 44 48 36 Heide et al) This isclose to the critical region of lesion overlap we haveidentified in the posterior neglect patients with high re-click rates (Talairach 37 39 37)
We suggest that in our patients with lesions involvingthe right intraparietal sulcus uncertainty about locationsof previously found targets increases with each saccadeThis uncertainty is further exacerbated by interferencefrom newer target locations leading to increased prob-ability of misjudging old target locations for new assearch progresses Such progressively worsening uncer-tainty might be due to an accumulating impairment inlsquolsquore-mapping target locationsrsquorsquo across saccades or im-paired lsquolsquomemoryrsquorsquo of previously inspected target loca-tions across saccades As noted above there is evidence
for both types of signalmdash re-mapping and maintainingsaccadic spatial memorymdashin the parietal cortex and it ispossible that disruption to both these functions contrib-utes to the re-click deficit we have identified Certainlythe two potential types of deficit are not mutuallyincompatible
A slightly different account of the re-clicking behaviormight be framed in terms of the intriguing hypothesisproposed by Manly Woldt Watson and Warburton(2002) to explain visible re-marking on a standard can-cellation task They argue that such behavior is due topatients erroneously transposing leftward stimuli to theright resulting in apparent re-cancellation These au-thors found that removing targets from the left re-duced both neglect and re-marking However they didnot monitor eye movements so it is possible that sometargets that are logged as neglected (because they werenot cancelled) were nevertheless fixated and thereforeform part of the load on memory during search Fur-thermore in our experimental study subjects with ahigh re-click rate also failed to click on a higher pro-portion of fixated targets Manly et alrsquos proposal wouldnot easily account for such lsquolsquoclick omissionsrsquorsquo becausethese refer to fixated targets that were not actuallyresponded to
Finally it might be argued that increasing re-clickingwith search duration reflects a deficit in spatial workingmemory (SWM) that is independent of saccadic eyemovements (ie a decay in memory that is not trans-saccadic) Because time and number of interveningsaccades were not independently manipulated in thisunrestricted visual search paradigm we are not ablecategorically to distinguish between these possibilitiesWe note however that functional imaging studies ofSWM tasks that do not require eye movements oftendemonstrate activity related to the maintenance ofspatial information which is generally more posteriorand dorsal than the parietal locations isolated in thisstudy (DrsquoEsposito et al 1998) Moreover recent behav-ioral data indicate that SWM is more vulnerable tosaccadic eye movements than shifts in attention or limbmovements (Pearson amp Sahraie 2003)
Frontal Lesion Site Associated withHigh Re-click Rates
Four out of 5 neglect patients with lesions involvingposterior plus anterior damage also showed a high re-click rate and the area of damage common to thesepatients centered on the right inferior frontal lobe cor-responding to Brodmannrsquos area 44 (Figure 7C) Thisregion has previously been associated with frontalneglect (Husain amp Kennard 1996) and is consideredto be part of the ventral premotor cortex and thehomologue of Brocarsquos areas in the left hemisphere(Rizzolatti amp Arbib 1998 Jackson amp Husain 1996) Inthese frontal patients the probability of re-clicking did
Mannan et al 349
not increase with time interval between the first clickand re-fixation but rather it stayed constant or declinedwith increasing delays
Such behavior might be consistent with a failure toinhibit responses to rightward locationsmdasha type oflsquolsquoperseverativersquorsquo behavior Note though that this is notnecessarily the same type of perseverative responseas seen when some patients with neglect re-markpreviously found items on standard cancellation tests(Rusconi et al 2002 Na et al 1999) In those casestargets that are visibly cancelled are often marked againwhereas in our computerized search tasks no marks arevisibly left on targets once they have been clicked uponMoreover only one of our neglect patients (Case 1) de-monstrated re-cancellation of visibly cancelled targetson pen-and-paper tasks and then only on one occasionImportantly this patient actually had a normal re-clickrate (67) on our task and his stroke was in the PCAterritory sparing both the intraparietal sulcus and fron-tal lobe
In support of a role for the right frontal lobe in in-hibiting responses to previously found targets recentimaging studies of response inhibition have highlightedregions of the right inferior frontal gyrus (IFG) very closeto the centroid of damage for our frontal patients (38 1618 cf 43 22 21 Konishi Kyoichi Uchida Sekihara ampMiyashita 1998 and 41 16 19 Konishi Kyoichi UchidaKikyo et al 1999) Alternative explanations for the highprobability of re-clicking directly after clicking might bethat these patients have impaired perceptual discrimina-tion of targets impaired perceptual localization of targetsor that they encounter difficulty in encoding spatiallocations into working memory Our data are not sugges-tive of a perceptual discrimination problem as changingthe perceptual load did not inf luence re-click ratesMoreover the patients still had high re-click rates in thetarget only conditions (Figure 10) where no perceptualdiscrimination of targets and distractors was necessaryPoor perceptual localization would seem unlikely giventhat these patients have frontal lesions Furthermoreusing a vertical SWM task similar to the Corsi blocktapping task (Malhotra et al 2004) we have found thatpatients with neglect can localize and perceptually en-code single target locations as well as controls and aresignificantly impaired at remembering two or more loca-tions shown in sequence consistent with the presentobservations Some recent functional imaging data sug-gest that activation within the prefrontal cortex corre-lates well with successful encoding of material intoworking memory (Rypma amp DrsquoEsposito 2003) Furtherresearch is required to distinguish between these pos-sible explanations for the mechanisms underlying there-click deficit in frontal patients
The only patient with frontal damage who did re-clickmore frequently with increasing time had damage cen-tered on the dorsolateral prefrontal cortex (althoughalso encroaching on the IFG) Neurons in the dorsolat-
eral prefrontal cortex like those in the LIP encode theremembered locations of saccadic targets (Goldman-Rakic 1996) This patient (Case 15) who had the high-est re-click rate of all also had damage to the criticalparietal location described above (in contrast to all otherfrontal patients) Thus she may have suffered from acombination of underlying deficits
Re-fixations and Misjudging PreviouslyInspected Locations during Search
In this search task re-fixation and re-click rates weresignificantly correlated for neglect patients Howeverthere were important dissociations within individuals 2patients with PCA lesions had re-fixation rates of 220and 80 respectively but both had a re-click rate ofonly 7 These are clearly examples of individuals whodo not show pathological re-clicking despite having highre-fixation rates High re-fixation rates may result froma lack of integration of spatial information due to V1damage (Zihl amp von Cramon 1985) in our PCA patientsor possibly because of reduced inhibition of return(Bartolomeo Sieroff Decaix amp Chokron 2001) What-ever the exact cause for dissociations between re-fixa-tions and re-click rates our data suggest re-fixation doesnot necessarily index the same underlying deficits asrevealed by our re-click measure High re-fixation ratesunlike high re-click rates were also not significantlyassociated with damage to any specific cortical locationagain demonstrating a dissociation between re-fixationsand pathological re-clicking
The relationship between re-click rates and neglect isunlikely to be a simple one In our study there was amoderate but significant correlation between the twomeasures suggesting that this association is unlikely tohave arisen by chance However non-neglect patientscan also exhibit a high re-click rate (unpublished obser-vations from our laboratory) Taken together our resultssuggest that difficulties in keeping track of previouslyinspected targets (because of impaired trans-saccadic re-mapping andor memory) may be one contributingcomponent to visual neglect in patients with lesions in-volving the right intraparietal sulcus As mentioned thesearch task we used differed in one striking aspect fromstandard cancellation tests subjects do not visibly markthe targets they visit Whereas in cancellation tests pa-tients mark each target they find which might appear todispense with any need to remember which locationshave been searched before However it should be ap-preciated that in most standard cancellation tests theindividual marks made by patients may not be clearlyvisible in peripheral vision particularly if neglect pa-tients have a local spatial bias (Rafal 1994 RobertsonLamb amp Knight 1988) or a restricted field of view(Russell Malhotra amp Husain 2004) Thus when com-bined with a rightward attentional bias keeping track ofsearched locations may be critical to prevent gaze from
350 Journal of Cognitive Neuroscience Volume 17 Number 2
returning to rightward locations even if they have beenmarked before as the marks may not be visible untilgaze returns to them Consistent with this hypothesisneglect is greater when cancellation is performed usinginvisible marks compared to large and salient red marksthat can be seen peripherally (Wojciulik Husain Clarkeamp Driver 2001)
A task like the present search-and-click paradigm maymore closely resemble search in the real world than dotraditional cancellation tests When we examine a visualscene we do not leave marks on each object we ex-amine Instead just as in the search task used in thisstudy we have to keep track of the locations we visit ifwe are to avoid inefficient re-examination of thoselocations Neglect patients with damage to the rightintraparietal sulcus appear to be impaired at keepingtrack of searched locations in this way When combinedwith their rightward attentional bias we propose thatsuch an impairment leads to recursive search of stimulito the right in experimental search tasks as used hereand also in daily life where neglect is so apparentdespite unlimited viewing time
The paradigm we have used has the important virtuesof allowing patients to explore freelymdashas in real-worldsearch or clinical cancellation tasksmdashand relating theirre-click behavior directly to the severity of neglect in thesame task But it might be argued that it would be usefulalso to probe SWM in a more controlled manner by re-stricting observersrsquo eye movements to a specific set oflocations and probing memory of previously visitedplaces A recent study using a gaze-contingent para-digm has elegantly probed the capacity of spatial mem-ory during search in healthy observers and foundevidence for memory for up to four previous locations(McCarley Wang Kramer Irwin amp Peterson 2003)Variants of this paradigm may in the future be usefulto probe the capacity of SWM across saccades in patientswith neglect and also to probe further whether thereare differences between neglect patients with parietaland frontal lesions
METHODS
Subjects Imaging and Lesion Mapping
Sixteen patients with left neglect following right hemi-sphere infarction or hemorrhage were recruited Con-sent was obtained according to the Declaration ofHelsinki as approved by the hospital ethical commit-tees Presence of neglect was determined by clinicalobservation plus bedside tests including the Mesulamshape-cancellation task (Mesulam 1985) and line bisec-tion task (Wilson et al 1987) (see Table 1) Twelvepatients had MCA territory strokes the rest involvedonly the PCA territory which can also result in neglect(Cals et al 2002) Only one neglect patient (Case 1 whodid not have frontal involvement) demonstrated re-
cancellation of visibly cancelled targets on pen-and-paper tasks and then only on one occasion Fifteenage-matched healthy volunteers were controls (meanage = 63 SD = 14 vs 63 SD = 13 for patients)
All patients had clinical CT scans In addition dedi-cated high-resolution structural MRI scans were alsoobtained for 1416 neglect patients MRI scans wereunobtainable for the other two (Cases 11 and 16)because of claustrophobia or loss to follow up In theMCA neglect group median time between stroke andscan was 63 days with the shortest interval being35 days for the PCA neglect group one patient had tobe scanned relatively early at 9 days but the mediantime was 140 days MRI scans comprised one 256-sliceT1-weighted acquisition (MPRAGE TR 97 msec TE4 msec TA 12 min) performed in the coronal planealigned perpendicular to the anteriorndashposterior commis-sural line on a Siemens Vision 15-T scanner Within-and between-slice resolution was 1 mm For each pa-tient the area of damage was determined by detailedvisual inspection of the digital image for every singleslice Lesion delineation was performed by neurologists(authors D M and M H) by tracing by hand theboundary of the lesion onto every single 1-mm-thickaxial slice using a graphics tablet (WACOM Intuos A6)Combining all the slices then led to a 3-D region of in-terest (ROI) for each patient For one patient (Case 15)there were two areas of discrete focal damage one inthe frontal cortex and the other in the parietal cortex
Normalization of each MRI including the 3-D ROI tothe T1-template was performed with SPM99 (wwwfilionuclacukspm) The ROI was used as a mask for thelesion during determination of alignment parameters tominimize the contribution of abnormal brain to thisprocess (Rorden amp Brett 2000) The normalized ROIof each neglect patient was superimposed upon the T1template to find areas of maximal lesion overlap in eachgroup (see Results) as well as lesion volume usingMRIcro shareware (Rorden amp Brett 2000)
Experimental Procedure
Stimuli were presented on an SVGA monitor (29 22 in)viewed at 57 cm distance Each search display com-prised a virtual 9 7 grid each cell containing a single18 elementmdasheither a letter T (target) or a letter L (dis-tractor) (see Figure 1) The position of each elementwithin each cell was jittered with 19 targets and 44distractors randomly distributed over the virtual gridThe number of targets on the right and left halves ofthe display was balanced across trials Seven neglectpatients (Cases 5 8 10 11 14ndash16) were further testedon lsquolsquoTarget onlyrsquorsquo displays which had varying numbersof Ts (2 6 or 19) now with no L-shaped distractorsThe presentations of these lsquolsquoTarget onlyrsquorsquo screens wereinterspersed with those of the original search screens inrandom order in the same experimental session
Mannan et al 351
Eye movements were recorded using a video-basedpupil tracker with a temporal resolution of 4 msec andspatial accuracy of lt058 (EyeLink Sensorimotoric Sys-tems Berlin Germany) Subjects were asked to keeptheir head still The experimenter also monitored gazeposition on-line At the beginning of each session (andafter breaks in testing or following any unintended headmovement that disrupted gaze recordings) eye-trackingwas re-calibrated Each trial was initiated by the exper-imenter and began with a central fixation cross for 1 secthen the search display
Subjects were instructed to search for Ts (among anyLs) Critically while looking at any target they had toclick a response button held in the right hand onlyif they considered this target to be a new discovery(ie a target they had not clicked on before) Clickingthe response button did not change the display Theresponse button was held by the patient in a comfort-able position to the right of the midline Subjectswere instructed and reminded throughout testing toclick on each target found once only Any later re-click on
a previously found target thus indicates a failure to retainthe fact that this target had already been discoveredThe experimenter terminated each search when thesubject indicated verbally that they felt all targets in thedisplay had now been found Each subject searched aminimum of 5 displays (mean = 7 range 5ndash14) Eyemovements were analyzed off-line using custom soft-ware An automatic algorithm was used to extract sac-cades (saccade onset defined as eye speed rising above258 sec1 for two or more samples and a criterion ofminimum fixation duration of 60 msec) Blinks wereeliminated from the trace by visual inspection and sac-cade and fixation delineation were checked visually inthe record Next fixations were automatically assignedto items in the display if the mean position fell within18 of that item Two or more consecutive fixations onthe same item were re-scored as a single extendedfixation A re-fixation was scored only when at leastone fixation elsewhere on another item intervened
Most click responses were made during a fixation andthe few button-clicks occurring during a saccade were
Table 1 Patient Details
CaseAge
(years) PathologyVolume(cm3)
Age ofLesion (days)
Visual FieldDefect
Mean Deviationfrom Center on
Line Bisection Task(cm) (rightward +)
Mesulum Score(Maximum Possible
Score = 60) Lesion Type
1 68 Infarct 1282 64 + 867 17 PCA
2 76 Infarct 1528 227 + 003 41 PCA
3 68 Infarct 79 207 + 297 36 PCA
4 77 Infarct 251 158 + 150 54 PCA
5 63 Infarct 28 29 033 55 MCA
6 59 Hemorrhage 172 7 + 32 6 MCA
7 66 Hemorrhage 273 325 + 213 31 MCA
8 68 Infarct 309 3 25 17 MCA
9 29 Infarct 115 17 113 55 MCA
10 70 Infarct 1339 324 153 57 MCA
11 42 Infarct ndash 11 097 24 MCA
12 75 Infarct 757 38 013 4 MCA
13 51 Infarct 694 141 083 31 MCA
14 59 Infarct 1144 356 + 223 37 MCA
15 80 Infarct 261 44 35 6 MCA
16 64 Infarct ndash 30 033 55 MCA
MEAN 63 66 124 188 33
SE 4 13 32 5
Cases 1ndash9 had damage to the nonfrontal (posterior only) cortex whereas Cases 10ndash15 had both frontal and posterior (posterior and anterior) corticalinvolvement Case 16 is a patient with a subcortical lesion All but 3 of the patients with right MCA infarction had intact visual fields on confrontationTwo of these (Cases 7 and 14) were diagnosed with left homonymous hemianopia whereas one (Case 6) had a left inferior quandrantic field defectAll patients with PCA territory damage had partial or complete contralesional visual field loss No patient suffered from dysphasia
352 Journal of Cognitive Neuroscience Volume 17 Number 2
assigned to the previous fixation Clicks were assigned toa particular item if eye position at the time of the clickwas within 28 of that item or in the cases where morethan one item was within 28 of current eye position theclick response was simply attributed to the item closestto that eye position
Button-clicks were assigned to a particular item if eyeposition at the time of the click was within 28 of thatitem A re-click on an item required at least one fixationon another item between successive clicks (in practiceseveral fixations elsewhere typically intervened betweeninitial click and later re-click see Results)
Statistical analyses were performed using SPSS andSigmaplot (SPSS Science Chicago USA) Jonckheere sta-tistic was derived by hand Statistical significance was setas p lt 05
Acknowledgments
We thank the reviewers of this manuscript and Dr AndrewParton for their extremely helpful comments We thank thepatients who participated Dr J Wade Dr A Rudd and thestaff of the stroke units at Charing Cross and St ThomasrsquoHospitals This research was supported by grants from theWellcome Trust to M H C K and J D J D holds a RoyalSociety Wolfson Research Merit Award
Reprint requests should be sent to Drs Sabira Mannan andMasud Husain Division of Neuroscience and PsychologicalMedicine Imperial College London Charing Cross CampusLondon W6 8RF UK or via e-mail smannanimperialacukor mhusainimperialacuk
Notes
1 We are grateful to one of the reviewers for suggesting thistime-series analysis2 There was no significant difference between the number oftargets fixated between the two groups so differences in lsquolsquoclickomissionsrsquorsquo cannot be attributed to the number of fixatedtargets that had to be kept track of during search3 The correlation between these four points alone inFigure 9A is high [r(3) = 81 cf r(15) = 47 for the wholegroup]4 We have also tested 10 neglect patients and 10 controlstroke patients on a vertical version of the Corsi blocks task(clinically used to measure spatial working memory capacity)Neglect patients had significantly impaired capacity but wereable to localize single locations well suggesting that mislocal-ization is unlikely to account for their deficit on this spatialworking memory task
REFERENCES
Bartolomeo P Sieroff E Decaix C amp Chokron S (2001)Modulating the attentional bias in unilateral neglect theeffects of the strategic set Experimental Brain Research137 432ndash444
Bates E Wilson S M Saygin A P Dick F Sereno M IKnight R T amp Dronkers N F (2003) Voxel-basedlesion-symptom mapping Nature Neuroscience 6 448ndash450
Behrmann M Watt S Black S E amp Barton J J (1997)Impaired visual search in patients with unilateral neglect
An oculographic analysis Neuropsychologia 351445ndash1458
Bisiach E amp Vallar G (2000) Unilateral neglect in humans InG Rizzolatti (Ed) Handbook of neuropsychology (2nd edvol 1 pp 459ndash502) Amsterdam Elsevier
Cals N Devuyst G Afsar N Karapanayiotides T ampBogousslavsky J (2002) Pure superficial posterior cerebralartery territory infarction in The Lausanne Stroke RegistryJournal of Neurology 249 855ndash861
Chafee M amp Goldman-Rakic P (1998) Matching patterns ofactivity in primate prefrontal area 8a and parietal area 7ipneurons during a spatial working memory task Journal ofNeurophysiology 79 2919ndash2940
Chedru F Leblanc M amp Lhermitte F (1973) Visualsearching in normal and brain-damaged subjectsContributions to the study of unilateral subjectsCortex 9 94ndash111
DrsquoEsposito M Aguirre G K Zarahn E Ballard DShin R K amp Lease J (1998) Functional MRI studies ofspatial and nonspatial working memory Cognitive BrainResearch 7 1ndash13
Donkelaar P amp Muri R (2002) Craniotopic updating ofvisual space across saccades in the human posteriorparietal cortex Proceedings of the Royal Society London B269 735ndash739
Driver J amp Husain M (2002) The role of spatial workingmemory deficits in pathological search by neglect patientsIn the cognitive and neural bases of spatial neglect InH-O Karnath A D Milner amp G Vallar (Eds) The cognitiveand neural bases of spatial neglect Oxford OxfordUniversity Press
Duhamel J R Colby C amp Goldberg M E (1992) Theupdating of the representation of visual space in parietalcortex by intended eye movements Science 25590ndash92
Goldman-Rakic P S (1996) The prefrontal landscapemdashImplications of functional architecture for understandinghuman mentation and the central executive PhilosophicalTransactions of The Royal Society of London Series BBiological Sciences 351 1445ndash1453
Heide W Binkofski F Seitz R J Posse S Nitschke M FFreund H J amp Kompf D (2001) Activation offrontoparietal cortices during memorized triple-stepsequences of saccadic eye movements An fMRI studyEuropean Journal of Neuroscience 13 1177ndash1189
Heide W amp Kompf D (1998) Combined deficits ofsaccades and visuo-spatial orientation after cortical lesionsExperimental Brain Research 123 164ndash171
Husain M amp Kennard C (1996) Visual neglect associatedwith frontal lobe infarction Journal of Neurology 243652ndash657
Husain M Mannan S Hodgson T Wojciulik E Driver Jamp Kennard C (2001) Impaired spatial working memoryacross saccades contributes to abnormal search in parietalneglect Brain 121 941ndash952
Husain M amp Rorden C (2003) Non-spatially lateralizedmechanisms in hemispatial neglect Nature ReviewsNeuroscience 4 23ndash36
Jackson S R amp Husain M (1996) Visuomotor functionsof the lateral pre-motor cortex Current Opinion inNeurobiology 6 788ndash795
Kinsbourne M (1993) Orientational bias model of unilateralneglect Evidence from attentional gradients withinhemispace In I H Robertson amp J C Marshall (Eds)Unilateral neglect Clinical and experimental studies(pp 63ndash86) Hove Erlbaum
Konishi S Kyoichi N Uchida I Kikyo H Kameyama Mamp Miyauchi S (1999) Common inhibitory mechanism in
Mannan et al 353
human inferior prefrontal cortex revealed by event-relatedfunctional MRI Brain 122 981ndash991
Konishi S Kyoichi N Uchida I Sekihara K amp Miyashita Y(1998) No-go dominant brain activity in human inferiorprefrontal cortex revealed by functional magneticresonance imaging European Journal of Neuroscience10 1209ndash1213
Malhotra P Mannan S Driver J amp Husain M (2004)Impaired spatial working memory One component of thevisual neglect syndrome Cortex 40 667ndash676
Manly T Woldt K Watson P amp Warburton E (2002) Ismotor perseveration in unilateral neglect lsquolsquodrivenrsquorsquo by thepresence of neglected left-sided stimuli Neuropsychologia40 1794ndash1803
Mazzoni P Bracewell R Barash S amp Andersen R (1996)Spatially tuned auditory responses in area LIP of macaquesperforming delayed memory saccades to acoustic targetsJournal of Neurophysiology 75 1233ndash1241
McCarley J Wang R Kramer A Irwin D amp Peterson M(2003) How much memory does oculomotor search havePsychological Science 14 422ndash426
Mesulam M-M (1985) Principles of behavioural neurologyTests of directed attention and memory Philadelphia Davis
Mesulam M M (1999) Spatial attention and neglectParietal frontal and cingulate contributions to the mentalrepresentation and attentional targeting of salientextrapersonal events Philosophical Transactions of theRoyal Society of London Series B Biological Sciences354 1325ndash1346
Mort D Malhotra P Mannan S Rorden C Pambakian AKennard C amp Husain M (2003) The anatomy of visualneglect Brain 126 1986ndash1997
Na D L Adair J C Kang Y Chung C S Lee K H ampHeilman K M (1999) Motor perseverative behaviour ona line cancellation task Neurology 52 1569ndash1576
Pearson D G amp Sahraie A (2003) Oculomotor controland the maintenance of spatially and temporally distributedevents in visuo-spatial working memory QuarterlyJournal of Experimental Psychology 56A 1089ndash1111
Rafal R (1994) Neglect Current Opinion in Neurobiology4 231ndash236
Rizzolatti G amp Arbib M A (1998) Language within our graspTrends in Neurosciences 21 188ndash194
Robertson L C Lamb M R amp Knight R T (1988) Effectsof lesions of temporalndashparietal junction on perceptual andattentional processing in humans Journal of Neuroscience8 3757ndash3769
Rorden C amp Brett M (2000) Stereotaxic display of brainlesions Behavioural Neurology 12 191ndash200
Rusconi M L Maravita A Bottini G amp Vallar G (2002)Is the intact side really intact Perseverative responses inpatients with unilateral neglect A productive manifestationNeuropsychologia 40 594ndash604
Russell C Malhotra P amp Husain M (2004) Attentionmodulates the visual field in healthy observers and parietalpatients NeuroReport 15 189ndash193
Rypma B amp DrsquoEsposito M (2003) A subsequent-memoryeffect in dorsolateral prefrontal cortex Cognitive BrainResearch 16 162ndash166
Sapir A Hayes A Henik A Danzinger S amp Rafal R(2004) Parietal lobe lesions disrupt saccadic remapping ofinhibitory location tagging Journal of CognitiveNeuroscience 16 503ndash509
Smania N Martini M C Gambina G A T Palamara ANatale E amp Marzi C A (1998) The spatial distributionof visual attention in hemineglect and extinction patientsBrain 121 1759ndash1770
Sprenger A Kompf D amp Heide W (2002) Visual searchin patients with left visual hemineglect Progress in BrainResearch 140 395ndash416
Tobler P Felblinger J Burki M Nirkko A Ozdoba C ampMuri R (2001) Functional organisation of the saccadicreference system processing extraretinal signals in humansVision Research 41 1351ndash1358
Wilson B Cockburn J amp Halligan P W (1987)Behavioural inattention test Bury St EdmundsThames Valley
Wojciulik E Husain M Clarke K amp Driver J (2001)Spatial working memory deficit in unilateral neglectNeuropsychologia 39 390ndash396
Zihl J amp Hebel N (1997) Patterns of oculomotor scanningin patients with unilateral posterior parietal and frontallobe damage Neuropsychologia 35 893ndash906
Zihl J amp von Cramon D (1985) Visual field recovery fromscotoma in patients with postgeniculate damage A reviewof 55 cases Brain 108 335ndash365
354 Journal of Cognitive Neuroscience Volume 17 Number 2

not increase with time interval between the first clickand re-fixation but rather it stayed constant or declinedwith increasing delays
Such behavior might be consistent with a failure toinhibit responses to rightward locationsmdasha type oflsquolsquoperseverativersquorsquo behavior Note though that this is notnecessarily the same type of perseverative responseas seen when some patients with neglect re-markpreviously found items on standard cancellation tests(Rusconi et al 2002 Na et al 1999) In those casestargets that are visibly cancelled are often marked againwhereas in our computerized search tasks no marks arevisibly left on targets once they have been clicked uponMoreover only one of our neglect patients (Case 1) de-monstrated re-cancellation of visibly cancelled targetson pen-and-paper tasks and then only on one occasionImportantly this patient actually had a normal re-clickrate (67) on our task and his stroke was in the PCAterritory sparing both the intraparietal sulcus and fron-tal lobe
In support of a role for the right frontal lobe in in-hibiting responses to previously found targets recentimaging studies of response inhibition have highlightedregions of the right inferior frontal gyrus (IFG) very closeto the centroid of damage for our frontal patients (38 1618 cf 43 22 21 Konishi Kyoichi Uchida Sekihara ampMiyashita 1998 and 41 16 19 Konishi Kyoichi UchidaKikyo et al 1999) Alternative explanations for the highprobability of re-clicking directly after clicking might bethat these patients have impaired perceptual discrimina-tion of targets impaired perceptual localization of targetsor that they encounter difficulty in encoding spatiallocations into working memory Our data are not sugges-tive of a perceptual discrimination problem as changingthe perceptual load did not inf luence re-click ratesMoreover the patients still had high re-click rates in thetarget only conditions (Figure 10) where no perceptualdiscrimination of targets and distractors was necessaryPoor perceptual localization would seem unlikely giventhat these patients have frontal lesions Furthermoreusing a vertical SWM task similar to the Corsi blocktapping task (Malhotra et al 2004) we have found thatpatients with neglect can localize and perceptually en-code single target locations as well as controls and aresignificantly impaired at remembering two or more loca-tions shown in sequence consistent with the presentobservations Some recent functional imaging data sug-gest that activation within the prefrontal cortex corre-lates well with successful encoding of material intoworking memory (Rypma amp DrsquoEsposito 2003) Furtherresearch is required to distinguish between these pos-sible explanations for the mechanisms underlying there-click deficit in frontal patients
The only patient with frontal damage who did re-clickmore frequently with increasing time had damage cen-tered on the dorsolateral prefrontal cortex (althoughalso encroaching on the IFG) Neurons in the dorsolat-
eral prefrontal cortex like those in the LIP encode theremembered locations of saccadic targets (Goldman-Rakic 1996) This patient (Case 15) who had the high-est re-click rate of all also had damage to the criticalparietal location described above (in contrast to all otherfrontal patients) Thus she may have suffered from acombination of underlying deficits
Re-fixations and Misjudging PreviouslyInspected Locations during Search
In this search task re-fixation and re-click rates weresignificantly correlated for neglect patients Howeverthere were important dissociations within individuals 2patients with PCA lesions had re-fixation rates of 220and 80 respectively but both had a re-click rate ofonly 7 These are clearly examples of individuals whodo not show pathological re-clicking despite having highre-fixation rates High re-fixation rates may result froma lack of integration of spatial information due to V1damage (Zihl amp von Cramon 1985) in our PCA patientsor possibly because of reduced inhibition of return(Bartolomeo Sieroff Decaix amp Chokron 2001) What-ever the exact cause for dissociations between re-fixa-tions and re-click rates our data suggest re-fixation doesnot necessarily index the same underlying deficits asrevealed by our re-click measure High re-fixation ratesunlike high re-click rates were also not significantlyassociated with damage to any specific cortical locationagain demonstrating a dissociation between re-fixationsand pathological re-clicking
The relationship between re-click rates and neglect isunlikely to be a simple one In our study there was amoderate but significant correlation between the twomeasures suggesting that this association is unlikely tohave arisen by chance However non-neglect patientscan also exhibit a high re-click rate (unpublished obser-vations from our laboratory) Taken together our resultssuggest that difficulties in keeping track of previouslyinspected targets (because of impaired trans-saccadic re-mapping andor memory) may be one contributingcomponent to visual neglect in patients with lesions in-volving the right intraparietal sulcus As mentioned thesearch task we used differed in one striking aspect fromstandard cancellation tests subjects do not visibly markthe targets they visit Whereas in cancellation tests pa-tients mark each target they find which might appear todispense with any need to remember which locationshave been searched before However it should be ap-preciated that in most standard cancellation tests theindividual marks made by patients may not be clearlyvisible in peripheral vision particularly if neglect pa-tients have a local spatial bias (Rafal 1994 RobertsonLamb amp Knight 1988) or a restricted field of view(Russell Malhotra amp Husain 2004) Thus when com-bined with a rightward attentional bias keeping track ofsearched locations may be critical to prevent gaze from
350 Journal of Cognitive Neuroscience Volume 17 Number 2
returning to rightward locations even if they have beenmarked before as the marks may not be visible untilgaze returns to them Consistent with this hypothesisneglect is greater when cancellation is performed usinginvisible marks compared to large and salient red marksthat can be seen peripherally (Wojciulik Husain Clarkeamp Driver 2001)
A task like the present search-and-click paradigm maymore closely resemble search in the real world than dotraditional cancellation tests When we examine a visualscene we do not leave marks on each object we ex-amine Instead just as in the search task used in thisstudy we have to keep track of the locations we visit ifwe are to avoid inefficient re-examination of thoselocations Neglect patients with damage to the rightintraparietal sulcus appear to be impaired at keepingtrack of searched locations in this way When combinedwith their rightward attentional bias we propose thatsuch an impairment leads to recursive search of stimulito the right in experimental search tasks as used hereand also in daily life where neglect is so apparentdespite unlimited viewing time
The paradigm we have used has the important virtuesof allowing patients to explore freelymdashas in real-worldsearch or clinical cancellation tasksmdashand relating theirre-click behavior directly to the severity of neglect in thesame task But it might be argued that it would be usefulalso to probe SWM in a more controlled manner by re-stricting observersrsquo eye movements to a specific set oflocations and probing memory of previously visitedplaces A recent study using a gaze-contingent para-digm has elegantly probed the capacity of spatial mem-ory during search in healthy observers and foundevidence for memory for up to four previous locations(McCarley Wang Kramer Irwin amp Peterson 2003)Variants of this paradigm may in the future be usefulto probe the capacity of SWM across saccades in patientswith neglect and also to probe further whether thereare differences between neglect patients with parietaland frontal lesions
METHODS
Subjects Imaging and Lesion Mapping
Sixteen patients with left neglect following right hemi-sphere infarction or hemorrhage were recruited Con-sent was obtained according to the Declaration ofHelsinki as approved by the hospital ethical commit-tees Presence of neglect was determined by clinicalobservation plus bedside tests including the Mesulamshape-cancellation task (Mesulam 1985) and line bisec-tion task (Wilson et al 1987) (see Table 1) Twelvepatients had MCA territory strokes the rest involvedonly the PCA territory which can also result in neglect(Cals et al 2002) Only one neglect patient (Case 1 whodid not have frontal involvement) demonstrated re-
cancellation of visibly cancelled targets on pen-and-paper tasks and then only on one occasion Fifteenage-matched healthy volunteers were controls (meanage = 63 SD = 14 vs 63 SD = 13 for patients)
All patients had clinical CT scans In addition dedi-cated high-resolution structural MRI scans were alsoobtained for 1416 neglect patients MRI scans wereunobtainable for the other two (Cases 11 and 16)because of claustrophobia or loss to follow up In theMCA neglect group median time between stroke andscan was 63 days with the shortest interval being35 days for the PCA neglect group one patient had tobe scanned relatively early at 9 days but the mediantime was 140 days MRI scans comprised one 256-sliceT1-weighted acquisition (MPRAGE TR 97 msec TE4 msec TA 12 min) performed in the coronal planealigned perpendicular to the anteriorndashposterior commis-sural line on a Siemens Vision 15-T scanner Within-and between-slice resolution was 1 mm For each pa-tient the area of damage was determined by detailedvisual inspection of the digital image for every singleslice Lesion delineation was performed by neurologists(authors D M and M H) by tracing by hand theboundary of the lesion onto every single 1-mm-thickaxial slice using a graphics tablet (WACOM Intuos A6)Combining all the slices then led to a 3-D region of in-terest (ROI) for each patient For one patient (Case 15)there were two areas of discrete focal damage one inthe frontal cortex and the other in the parietal cortex
Normalization of each MRI including the 3-D ROI tothe T1-template was performed with SPM99 (wwwfilionuclacukspm) The ROI was used as a mask for thelesion during determination of alignment parameters tominimize the contribution of abnormal brain to thisprocess (Rorden amp Brett 2000) The normalized ROIof each neglect patient was superimposed upon the T1template to find areas of maximal lesion overlap in eachgroup (see Results) as well as lesion volume usingMRIcro shareware (Rorden amp Brett 2000)
Experimental Procedure
Stimuli were presented on an SVGA monitor (29 22 in)viewed at 57 cm distance Each search display com-prised a virtual 9 7 grid each cell containing a single18 elementmdasheither a letter T (target) or a letter L (dis-tractor) (see Figure 1) The position of each elementwithin each cell was jittered with 19 targets and 44distractors randomly distributed over the virtual gridThe number of targets on the right and left halves ofthe display was balanced across trials Seven neglectpatients (Cases 5 8 10 11 14ndash16) were further testedon lsquolsquoTarget onlyrsquorsquo displays which had varying numbersof Ts (2 6 or 19) now with no L-shaped distractorsThe presentations of these lsquolsquoTarget onlyrsquorsquo screens wereinterspersed with those of the original search screens inrandom order in the same experimental session
Mannan et al 351
Eye movements were recorded using a video-basedpupil tracker with a temporal resolution of 4 msec andspatial accuracy of lt058 (EyeLink Sensorimotoric Sys-tems Berlin Germany) Subjects were asked to keeptheir head still The experimenter also monitored gazeposition on-line At the beginning of each session (andafter breaks in testing or following any unintended headmovement that disrupted gaze recordings) eye-trackingwas re-calibrated Each trial was initiated by the exper-imenter and began with a central fixation cross for 1 secthen the search display
Subjects were instructed to search for Ts (among anyLs) Critically while looking at any target they had toclick a response button held in the right hand onlyif they considered this target to be a new discovery(ie a target they had not clicked on before) Clickingthe response button did not change the display Theresponse button was held by the patient in a comfort-able position to the right of the midline Subjectswere instructed and reminded throughout testing toclick on each target found once only Any later re-click on
a previously found target thus indicates a failure to retainthe fact that this target had already been discoveredThe experimenter terminated each search when thesubject indicated verbally that they felt all targets in thedisplay had now been found Each subject searched aminimum of 5 displays (mean = 7 range 5ndash14) Eyemovements were analyzed off-line using custom soft-ware An automatic algorithm was used to extract sac-cades (saccade onset defined as eye speed rising above258 sec1 for two or more samples and a criterion ofminimum fixation duration of 60 msec) Blinks wereeliminated from the trace by visual inspection and sac-cade and fixation delineation were checked visually inthe record Next fixations were automatically assignedto items in the display if the mean position fell within18 of that item Two or more consecutive fixations onthe same item were re-scored as a single extendedfixation A re-fixation was scored only when at leastone fixation elsewhere on another item intervened
Most click responses were made during a fixation andthe few button-clicks occurring during a saccade were
Table 1 Patient Details
CaseAge
(years) PathologyVolume(cm3)
Age ofLesion (days)
Visual FieldDefect
Mean Deviationfrom Center on
Line Bisection Task(cm) (rightward +)
Mesulum Score(Maximum Possible
Score = 60) Lesion Type
1 68 Infarct 1282 64 + 867 17 PCA
2 76 Infarct 1528 227 + 003 41 PCA
3 68 Infarct 79 207 + 297 36 PCA
4 77 Infarct 251 158 + 150 54 PCA
5 63 Infarct 28 29 033 55 MCA
6 59 Hemorrhage 172 7 + 32 6 MCA
7 66 Hemorrhage 273 325 + 213 31 MCA
8 68 Infarct 309 3 25 17 MCA
9 29 Infarct 115 17 113 55 MCA
10 70 Infarct 1339 324 153 57 MCA
11 42 Infarct ndash 11 097 24 MCA
12 75 Infarct 757 38 013 4 MCA
13 51 Infarct 694 141 083 31 MCA
14 59 Infarct 1144 356 + 223 37 MCA
15 80 Infarct 261 44 35 6 MCA
16 64 Infarct ndash 30 033 55 MCA
MEAN 63 66 124 188 33
SE 4 13 32 5
Cases 1ndash9 had damage to the nonfrontal (posterior only) cortex whereas Cases 10ndash15 had both frontal and posterior (posterior and anterior) corticalinvolvement Case 16 is a patient with a subcortical lesion All but 3 of the patients with right MCA infarction had intact visual fields on confrontationTwo of these (Cases 7 and 14) were diagnosed with left homonymous hemianopia whereas one (Case 6) had a left inferior quandrantic field defectAll patients with PCA territory damage had partial or complete contralesional visual field loss No patient suffered from dysphasia
352 Journal of Cognitive Neuroscience Volume 17 Number 2
assigned to the previous fixation Clicks were assigned toa particular item if eye position at the time of the clickwas within 28 of that item or in the cases where morethan one item was within 28 of current eye position theclick response was simply attributed to the item closestto that eye position
Button-clicks were assigned to a particular item if eyeposition at the time of the click was within 28 of thatitem A re-click on an item required at least one fixationon another item between successive clicks (in practiceseveral fixations elsewhere typically intervened betweeninitial click and later re-click see Results)
Statistical analyses were performed using SPSS andSigmaplot (SPSS Science Chicago USA) Jonckheere sta-tistic was derived by hand Statistical significance was setas p lt 05
Acknowledgments
We thank the reviewers of this manuscript and Dr AndrewParton for their extremely helpful comments We thank thepatients who participated Dr J Wade Dr A Rudd and thestaff of the stroke units at Charing Cross and St ThomasrsquoHospitals This research was supported by grants from theWellcome Trust to M H C K and J D J D holds a RoyalSociety Wolfson Research Merit Award
Reprint requests should be sent to Drs Sabira Mannan andMasud Husain Division of Neuroscience and PsychologicalMedicine Imperial College London Charing Cross CampusLondon W6 8RF UK or via e-mail smannanimperialacukor mhusainimperialacuk
Notes
1 We are grateful to one of the reviewers for suggesting thistime-series analysis2 There was no significant difference between the number oftargets fixated between the two groups so differences in lsquolsquoclickomissionsrsquorsquo cannot be attributed to the number of fixatedtargets that had to be kept track of during search3 The correlation between these four points alone inFigure 9A is high [r(3) = 81 cf r(15) = 47 for the wholegroup]4 We have also tested 10 neglect patients and 10 controlstroke patients on a vertical version of the Corsi blocks task(clinically used to measure spatial working memory capacity)Neglect patients had significantly impaired capacity but wereable to localize single locations well suggesting that mislocal-ization is unlikely to account for their deficit on this spatialworking memory task
REFERENCES
Bartolomeo P Sieroff E Decaix C amp Chokron S (2001)Modulating the attentional bias in unilateral neglect theeffects of the strategic set Experimental Brain Research137 432ndash444
Bates E Wilson S M Saygin A P Dick F Sereno M IKnight R T amp Dronkers N F (2003) Voxel-basedlesion-symptom mapping Nature Neuroscience 6 448ndash450
Behrmann M Watt S Black S E amp Barton J J (1997)Impaired visual search in patients with unilateral neglect
An oculographic analysis Neuropsychologia 351445ndash1458
Bisiach E amp Vallar G (2000) Unilateral neglect in humans InG Rizzolatti (Ed) Handbook of neuropsychology (2nd edvol 1 pp 459ndash502) Amsterdam Elsevier
Cals N Devuyst G Afsar N Karapanayiotides T ampBogousslavsky J (2002) Pure superficial posterior cerebralartery territory infarction in The Lausanne Stroke RegistryJournal of Neurology 249 855ndash861
Chafee M amp Goldman-Rakic P (1998) Matching patterns ofactivity in primate prefrontal area 8a and parietal area 7ipneurons during a spatial working memory task Journal ofNeurophysiology 79 2919ndash2940
Chedru F Leblanc M amp Lhermitte F (1973) Visualsearching in normal and brain-damaged subjectsContributions to the study of unilateral subjectsCortex 9 94ndash111
DrsquoEsposito M Aguirre G K Zarahn E Ballard DShin R K amp Lease J (1998) Functional MRI studies ofspatial and nonspatial working memory Cognitive BrainResearch 7 1ndash13
Donkelaar P amp Muri R (2002) Craniotopic updating ofvisual space across saccades in the human posteriorparietal cortex Proceedings of the Royal Society London B269 735ndash739
Driver J amp Husain M (2002) The role of spatial workingmemory deficits in pathological search by neglect patientsIn the cognitive and neural bases of spatial neglect InH-O Karnath A D Milner amp G Vallar (Eds) The cognitiveand neural bases of spatial neglect Oxford OxfordUniversity Press
Duhamel J R Colby C amp Goldberg M E (1992) Theupdating of the representation of visual space in parietalcortex by intended eye movements Science 25590ndash92
Goldman-Rakic P S (1996) The prefrontal landscapemdashImplications of functional architecture for understandinghuman mentation and the central executive PhilosophicalTransactions of The Royal Society of London Series BBiological Sciences 351 1445ndash1453
Heide W Binkofski F Seitz R J Posse S Nitschke M FFreund H J amp Kompf D (2001) Activation offrontoparietal cortices during memorized triple-stepsequences of saccadic eye movements An fMRI studyEuropean Journal of Neuroscience 13 1177ndash1189
Heide W amp Kompf D (1998) Combined deficits ofsaccades and visuo-spatial orientation after cortical lesionsExperimental Brain Research 123 164ndash171
Husain M amp Kennard C (1996) Visual neglect associatedwith frontal lobe infarction Journal of Neurology 243652ndash657
Husain M Mannan S Hodgson T Wojciulik E Driver Jamp Kennard C (2001) Impaired spatial working memoryacross saccades contributes to abnormal search in parietalneglect Brain 121 941ndash952
Husain M amp Rorden C (2003) Non-spatially lateralizedmechanisms in hemispatial neglect Nature ReviewsNeuroscience 4 23ndash36
Jackson S R amp Husain M (1996) Visuomotor functionsof the lateral pre-motor cortex Current Opinion inNeurobiology 6 788ndash795
Kinsbourne M (1993) Orientational bias model of unilateralneglect Evidence from attentional gradients withinhemispace In I H Robertson amp J C Marshall (Eds)Unilateral neglect Clinical and experimental studies(pp 63ndash86) Hove Erlbaum
Konishi S Kyoichi N Uchida I Kikyo H Kameyama Mamp Miyauchi S (1999) Common inhibitory mechanism in
Mannan et al 353
human inferior prefrontal cortex revealed by event-relatedfunctional MRI Brain 122 981ndash991
Konishi S Kyoichi N Uchida I Sekihara K amp Miyashita Y(1998) No-go dominant brain activity in human inferiorprefrontal cortex revealed by functional magneticresonance imaging European Journal of Neuroscience10 1209ndash1213
Malhotra P Mannan S Driver J amp Husain M (2004)Impaired spatial working memory One component of thevisual neglect syndrome Cortex 40 667ndash676
Manly T Woldt K Watson P amp Warburton E (2002) Ismotor perseveration in unilateral neglect lsquolsquodrivenrsquorsquo by thepresence of neglected left-sided stimuli Neuropsychologia40 1794ndash1803
Mazzoni P Bracewell R Barash S amp Andersen R (1996)Spatially tuned auditory responses in area LIP of macaquesperforming delayed memory saccades to acoustic targetsJournal of Neurophysiology 75 1233ndash1241
McCarley J Wang R Kramer A Irwin D amp Peterson M(2003) How much memory does oculomotor search havePsychological Science 14 422ndash426
Mesulam M-M (1985) Principles of behavioural neurologyTests of directed attention and memory Philadelphia Davis
Mesulam M M (1999) Spatial attention and neglectParietal frontal and cingulate contributions to the mentalrepresentation and attentional targeting of salientextrapersonal events Philosophical Transactions of theRoyal Society of London Series B Biological Sciences354 1325ndash1346
Mort D Malhotra P Mannan S Rorden C Pambakian AKennard C amp Husain M (2003) The anatomy of visualneglect Brain 126 1986ndash1997
Na D L Adair J C Kang Y Chung C S Lee K H ampHeilman K M (1999) Motor perseverative behaviour ona line cancellation task Neurology 52 1569ndash1576
Pearson D G amp Sahraie A (2003) Oculomotor controland the maintenance of spatially and temporally distributedevents in visuo-spatial working memory QuarterlyJournal of Experimental Psychology 56A 1089ndash1111
Rafal R (1994) Neglect Current Opinion in Neurobiology4 231ndash236
Rizzolatti G amp Arbib M A (1998) Language within our graspTrends in Neurosciences 21 188ndash194
Robertson L C Lamb M R amp Knight R T (1988) Effectsof lesions of temporalndashparietal junction on perceptual andattentional processing in humans Journal of Neuroscience8 3757ndash3769
Rorden C amp Brett M (2000) Stereotaxic display of brainlesions Behavioural Neurology 12 191ndash200
Rusconi M L Maravita A Bottini G amp Vallar G (2002)Is the intact side really intact Perseverative responses inpatients with unilateral neglect A productive manifestationNeuropsychologia 40 594ndash604
Russell C Malhotra P amp Husain M (2004) Attentionmodulates the visual field in healthy observers and parietalpatients NeuroReport 15 189ndash193
Rypma B amp DrsquoEsposito M (2003) A subsequent-memoryeffect in dorsolateral prefrontal cortex Cognitive BrainResearch 16 162ndash166
Sapir A Hayes A Henik A Danzinger S amp Rafal R(2004) Parietal lobe lesions disrupt saccadic remapping ofinhibitory location tagging Journal of CognitiveNeuroscience 16 503ndash509
Smania N Martini M C Gambina G A T Palamara ANatale E amp Marzi C A (1998) The spatial distributionof visual attention in hemineglect and extinction patientsBrain 121 1759ndash1770
Sprenger A Kompf D amp Heide W (2002) Visual searchin patients with left visual hemineglect Progress in BrainResearch 140 395ndash416
Tobler P Felblinger J Burki M Nirkko A Ozdoba C ampMuri R (2001) Functional organisation of the saccadicreference system processing extraretinal signals in humansVision Research 41 1351ndash1358
Wilson B Cockburn J amp Halligan P W (1987)Behavioural inattention test Bury St EdmundsThames Valley
Wojciulik E Husain M Clarke K amp Driver J (2001)Spatial working memory deficit in unilateral neglectNeuropsychologia 39 390ndash396
Zihl J amp Hebel N (1997) Patterns of oculomotor scanningin patients with unilateral posterior parietal and frontallobe damage Neuropsychologia 35 893ndash906
Zihl J amp von Cramon D (1985) Visual field recovery fromscotoma in patients with postgeniculate damage A reviewof 55 cases Brain 108 335ndash365
354 Journal of Cognitive Neuroscience Volume 17 Number 2

returning to rightward locations even if they have beenmarked before as the marks may not be visible untilgaze returns to them Consistent with this hypothesisneglect is greater when cancellation is performed usinginvisible marks compared to large and salient red marksthat can be seen peripherally (Wojciulik Husain Clarkeamp Driver 2001)
A task like the present search-and-click paradigm maymore closely resemble search in the real world than dotraditional cancellation tests When we examine a visualscene we do not leave marks on each object we ex-amine Instead just as in the search task used in thisstudy we have to keep track of the locations we visit ifwe are to avoid inefficient re-examination of thoselocations Neglect patients with damage to the rightintraparietal sulcus appear to be impaired at keepingtrack of searched locations in this way When combinedwith their rightward attentional bias we propose thatsuch an impairment leads to recursive search of stimulito the right in experimental search tasks as used hereand also in daily life where neglect is so apparentdespite unlimited viewing time
The paradigm we have used has the important virtuesof allowing patients to explore freelymdashas in real-worldsearch or clinical cancellation tasksmdashand relating theirre-click behavior directly to the severity of neglect in thesame task But it might be argued that it would be usefulalso to probe SWM in a more controlled manner by re-stricting observersrsquo eye movements to a specific set oflocations and probing memory of previously visitedplaces A recent study using a gaze-contingent para-digm has elegantly probed the capacity of spatial mem-ory during search in healthy observers and foundevidence for memory for up to four previous locations(McCarley Wang Kramer Irwin amp Peterson 2003)Variants of this paradigm may in the future be usefulto probe the capacity of SWM across saccades in patientswith neglect and also to probe further whether thereare differences between neglect patients with parietaland frontal lesions
METHODS
Subjects Imaging and Lesion Mapping
Sixteen patients with left neglect following right hemi-sphere infarction or hemorrhage were recruited Con-sent was obtained according to the Declaration ofHelsinki as approved by the hospital ethical commit-tees Presence of neglect was determined by clinicalobservation plus bedside tests including the Mesulamshape-cancellation task (Mesulam 1985) and line bisec-tion task (Wilson et al 1987) (see Table 1) Twelvepatients had MCA territory strokes the rest involvedonly the PCA territory which can also result in neglect(Cals et al 2002) Only one neglect patient (Case 1 whodid not have frontal involvement) demonstrated re-
cancellation of visibly cancelled targets on pen-and-paper tasks and then only on one occasion Fifteenage-matched healthy volunteers were controls (meanage = 63 SD = 14 vs 63 SD = 13 for patients)
All patients had clinical CT scans In addition dedi-cated high-resolution structural MRI scans were alsoobtained for 1416 neglect patients MRI scans wereunobtainable for the other two (Cases 11 and 16)because of claustrophobia or loss to follow up In theMCA neglect group median time between stroke andscan was 63 days with the shortest interval being35 days for the PCA neglect group one patient had tobe scanned relatively early at 9 days but the mediantime was 140 days MRI scans comprised one 256-sliceT1-weighted acquisition (MPRAGE TR 97 msec TE4 msec TA 12 min) performed in the coronal planealigned perpendicular to the anteriorndashposterior commis-sural line on a Siemens Vision 15-T scanner Within-and between-slice resolution was 1 mm For each pa-tient the area of damage was determined by detailedvisual inspection of the digital image for every singleslice Lesion delineation was performed by neurologists(authors D M and M H) by tracing by hand theboundary of the lesion onto every single 1-mm-thickaxial slice using a graphics tablet (WACOM Intuos A6)Combining all the slices then led to a 3-D region of in-terest (ROI) for each patient For one patient (Case 15)there were two areas of discrete focal damage one inthe frontal cortex and the other in the parietal cortex
Normalization of each MRI including the 3-D ROI tothe T1-template was performed with SPM99 (wwwfilionuclacukspm) The ROI was used as a mask for thelesion during determination of alignment parameters tominimize the contribution of abnormal brain to thisprocess (Rorden amp Brett 2000) The normalized ROIof each neglect patient was superimposed upon the T1template to find areas of maximal lesion overlap in eachgroup (see Results) as well as lesion volume usingMRIcro shareware (Rorden amp Brett 2000)
Experimental Procedure
Stimuli were presented on an SVGA monitor (29 22 in)viewed at 57 cm distance Each search display com-prised a virtual 9 7 grid each cell containing a single18 elementmdasheither a letter T (target) or a letter L (dis-tractor) (see Figure 1) The position of each elementwithin each cell was jittered with 19 targets and 44distractors randomly distributed over the virtual gridThe number of targets on the right and left halves ofthe display was balanced across trials Seven neglectpatients (Cases 5 8 10 11 14ndash16) were further testedon lsquolsquoTarget onlyrsquorsquo displays which had varying numbersof Ts (2 6 or 19) now with no L-shaped distractorsThe presentations of these lsquolsquoTarget onlyrsquorsquo screens wereinterspersed with those of the original search screens inrandom order in the same experimental session
Mannan et al 351
Eye movements were recorded using a video-basedpupil tracker with a temporal resolution of 4 msec andspatial accuracy of lt058 (EyeLink Sensorimotoric Sys-tems Berlin Germany) Subjects were asked to keeptheir head still The experimenter also monitored gazeposition on-line At the beginning of each session (andafter breaks in testing or following any unintended headmovement that disrupted gaze recordings) eye-trackingwas re-calibrated Each trial was initiated by the exper-imenter and began with a central fixation cross for 1 secthen the search display
Subjects were instructed to search for Ts (among anyLs) Critically while looking at any target they had toclick a response button held in the right hand onlyif they considered this target to be a new discovery(ie a target they had not clicked on before) Clickingthe response button did not change the display Theresponse button was held by the patient in a comfort-able position to the right of the midline Subjectswere instructed and reminded throughout testing toclick on each target found once only Any later re-click on
a previously found target thus indicates a failure to retainthe fact that this target had already been discoveredThe experimenter terminated each search when thesubject indicated verbally that they felt all targets in thedisplay had now been found Each subject searched aminimum of 5 displays (mean = 7 range 5ndash14) Eyemovements were analyzed off-line using custom soft-ware An automatic algorithm was used to extract sac-cades (saccade onset defined as eye speed rising above258 sec1 for two or more samples and a criterion ofminimum fixation duration of 60 msec) Blinks wereeliminated from the trace by visual inspection and sac-cade and fixation delineation were checked visually inthe record Next fixations were automatically assignedto items in the display if the mean position fell within18 of that item Two or more consecutive fixations onthe same item were re-scored as a single extendedfixation A re-fixation was scored only when at leastone fixation elsewhere on another item intervened
Most click responses were made during a fixation andthe few button-clicks occurring during a saccade were
Table 1 Patient Details
CaseAge
(years) PathologyVolume(cm3)
Age ofLesion (days)
Visual FieldDefect
Mean Deviationfrom Center on
Line Bisection Task(cm) (rightward +)
Mesulum Score(Maximum Possible
Score = 60) Lesion Type
1 68 Infarct 1282 64 + 867 17 PCA
2 76 Infarct 1528 227 + 003 41 PCA
3 68 Infarct 79 207 + 297 36 PCA
4 77 Infarct 251 158 + 150 54 PCA
5 63 Infarct 28 29 033 55 MCA
6 59 Hemorrhage 172 7 + 32 6 MCA
7 66 Hemorrhage 273 325 + 213 31 MCA
8 68 Infarct 309 3 25 17 MCA
9 29 Infarct 115 17 113 55 MCA
10 70 Infarct 1339 324 153 57 MCA
11 42 Infarct ndash 11 097 24 MCA
12 75 Infarct 757 38 013 4 MCA
13 51 Infarct 694 141 083 31 MCA
14 59 Infarct 1144 356 + 223 37 MCA
15 80 Infarct 261 44 35 6 MCA
16 64 Infarct ndash 30 033 55 MCA
MEAN 63 66 124 188 33
SE 4 13 32 5
Cases 1ndash9 had damage to the nonfrontal (posterior only) cortex whereas Cases 10ndash15 had both frontal and posterior (posterior and anterior) corticalinvolvement Case 16 is a patient with a subcortical lesion All but 3 of the patients with right MCA infarction had intact visual fields on confrontationTwo of these (Cases 7 and 14) were diagnosed with left homonymous hemianopia whereas one (Case 6) had a left inferior quandrantic field defectAll patients with PCA territory damage had partial or complete contralesional visual field loss No patient suffered from dysphasia
352 Journal of Cognitive Neuroscience Volume 17 Number 2
assigned to the previous fixation Clicks were assigned toa particular item if eye position at the time of the clickwas within 28 of that item or in the cases where morethan one item was within 28 of current eye position theclick response was simply attributed to the item closestto that eye position
Button-clicks were assigned to a particular item if eyeposition at the time of the click was within 28 of thatitem A re-click on an item required at least one fixationon another item between successive clicks (in practiceseveral fixations elsewhere typically intervened betweeninitial click and later re-click see Results)
Statistical analyses were performed using SPSS andSigmaplot (SPSS Science Chicago USA) Jonckheere sta-tistic was derived by hand Statistical significance was setas p lt 05
Acknowledgments
We thank the reviewers of this manuscript and Dr AndrewParton for their extremely helpful comments We thank thepatients who participated Dr J Wade Dr A Rudd and thestaff of the stroke units at Charing Cross and St ThomasrsquoHospitals This research was supported by grants from theWellcome Trust to M H C K and J D J D holds a RoyalSociety Wolfson Research Merit Award
Reprint requests should be sent to Drs Sabira Mannan andMasud Husain Division of Neuroscience and PsychologicalMedicine Imperial College London Charing Cross CampusLondon W6 8RF UK or via e-mail smannanimperialacukor mhusainimperialacuk
Notes
1 We are grateful to one of the reviewers for suggesting thistime-series analysis2 There was no significant difference between the number oftargets fixated between the two groups so differences in lsquolsquoclickomissionsrsquorsquo cannot be attributed to the number of fixatedtargets that had to be kept track of during search3 The correlation between these four points alone inFigure 9A is high [r(3) = 81 cf r(15) = 47 for the wholegroup]4 We have also tested 10 neglect patients and 10 controlstroke patients on a vertical version of the Corsi blocks task(clinically used to measure spatial working memory capacity)Neglect patients had significantly impaired capacity but wereable to localize single locations well suggesting that mislocal-ization is unlikely to account for their deficit on this spatialworking memory task
REFERENCES
Bartolomeo P Sieroff E Decaix C amp Chokron S (2001)Modulating the attentional bias in unilateral neglect theeffects of the strategic set Experimental Brain Research137 432ndash444
Bates E Wilson S M Saygin A P Dick F Sereno M IKnight R T amp Dronkers N F (2003) Voxel-basedlesion-symptom mapping Nature Neuroscience 6 448ndash450
Behrmann M Watt S Black S E amp Barton J J (1997)Impaired visual search in patients with unilateral neglect
An oculographic analysis Neuropsychologia 351445ndash1458
Bisiach E amp Vallar G (2000) Unilateral neglect in humans InG Rizzolatti (Ed) Handbook of neuropsychology (2nd edvol 1 pp 459ndash502) Amsterdam Elsevier
Cals N Devuyst G Afsar N Karapanayiotides T ampBogousslavsky J (2002) Pure superficial posterior cerebralartery territory infarction in The Lausanne Stroke RegistryJournal of Neurology 249 855ndash861
Chafee M amp Goldman-Rakic P (1998) Matching patterns ofactivity in primate prefrontal area 8a and parietal area 7ipneurons during a spatial working memory task Journal ofNeurophysiology 79 2919ndash2940
Chedru F Leblanc M amp Lhermitte F (1973) Visualsearching in normal and brain-damaged subjectsContributions to the study of unilateral subjectsCortex 9 94ndash111
DrsquoEsposito M Aguirre G K Zarahn E Ballard DShin R K amp Lease J (1998) Functional MRI studies ofspatial and nonspatial working memory Cognitive BrainResearch 7 1ndash13
Donkelaar P amp Muri R (2002) Craniotopic updating ofvisual space across saccades in the human posteriorparietal cortex Proceedings of the Royal Society London B269 735ndash739
Driver J amp Husain M (2002) The role of spatial workingmemory deficits in pathological search by neglect patientsIn the cognitive and neural bases of spatial neglect InH-O Karnath A D Milner amp G Vallar (Eds) The cognitiveand neural bases of spatial neglect Oxford OxfordUniversity Press
Duhamel J R Colby C amp Goldberg M E (1992) Theupdating of the representation of visual space in parietalcortex by intended eye movements Science 25590ndash92
Goldman-Rakic P S (1996) The prefrontal landscapemdashImplications of functional architecture for understandinghuman mentation and the central executive PhilosophicalTransactions of The Royal Society of London Series BBiological Sciences 351 1445ndash1453
Heide W Binkofski F Seitz R J Posse S Nitschke M FFreund H J amp Kompf D (2001) Activation offrontoparietal cortices during memorized triple-stepsequences of saccadic eye movements An fMRI studyEuropean Journal of Neuroscience 13 1177ndash1189
Heide W amp Kompf D (1998) Combined deficits ofsaccades and visuo-spatial orientation after cortical lesionsExperimental Brain Research 123 164ndash171
Husain M amp Kennard C (1996) Visual neglect associatedwith frontal lobe infarction Journal of Neurology 243652ndash657
Husain M Mannan S Hodgson T Wojciulik E Driver Jamp Kennard C (2001) Impaired spatial working memoryacross saccades contributes to abnormal search in parietalneglect Brain 121 941ndash952
Husain M amp Rorden C (2003) Non-spatially lateralizedmechanisms in hemispatial neglect Nature ReviewsNeuroscience 4 23ndash36
Jackson S R amp Husain M (1996) Visuomotor functionsof the lateral pre-motor cortex Current Opinion inNeurobiology 6 788ndash795
Kinsbourne M (1993) Orientational bias model of unilateralneglect Evidence from attentional gradients withinhemispace In I H Robertson amp J C Marshall (Eds)Unilateral neglect Clinical and experimental studies(pp 63ndash86) Hove Erlbaum
Konishi S Kyoichi N Uchida I Kikyo H Kameyama Mamp Miyauchi S (1999) Common inhibitory mechanism in
Mannan et al 353
human inferior prefrontal cortex revealed by event-relatedfunctional MRI Brain 122 981ndash991
Konishi S Kyoichi N Uchida I Sekihara K amp Miyashita Y(1998) No-go dominant brain activity in human inferiorprefrontal cortex revealed by functional magneticresonance imaging European Journal of Neuroscience10 1209ndash1213
Malhotra P Mannan S Driver J amp Husain M (2004)Impaired spatial working memory One component of thevisual neglect syndrome Cortex 40 667ndash676
Manly T Woldt K Watson P amp Warburton E (2002) Ismotor perseveration in unilateral neglect lsquolsquodrivenrsquorsquo by thepresence of neglected left-sided stimuli Neuropsychologia40 1794ndash1803
Mazzoni P Bracewell R Barash S amp Andersen R (1996)Spatially tuned auditory responses in area LIP of macaquesperforming delayed memory saccades to acoustic targetsJournal of Neurophysiology 75 1233ndash1241
McCarley J Wang R Kramer A Irwin D amp Peterson M(2003) How much memory does oculomotor search havePsychological Science 14 422ndash426
Mesulam M-M (1985) Principles of behavioural neurologyTests of directed attention and memory Philadelphia Davis
Mesulam M M (1999) Spatial attention and neglectParietal frontal and cingulate contributions to the mentalrepresentation and attentional targeting of salientextrapersonal events Philosophical Transactions of theRoyal Society of London Series B Biological Sciences354 1325ndash1346
Mort D Malhotra P Mannan S Rorden C Pambakian AKennard C amp Husain M (2003) The anatomy of visualneglect Brain 126 1986ndash1997
Na D L Adair J C Kang Y Chung C S Lee K H ampHeilman K M (1999) Motor perseverative behaviour ona line cancellation task Neurology 52 1569ndash1576
Pearson D G amp Sahraie A (2003) Oculomotor controland the maintenance of spatially and temporally distributedevents in visuo-spatial working memory QuarterlyJournal of Experimental Psychology 56A 1089ndash1111
Rafal R (1994) Neglect Current Opinion in Neurobiology4 231ndash236
Rizzolatti G amp Arbib M A (1998) Language within our graspTrends in Neurosciences 21 188ndash194
Robertson L C Lamb M R amp Knight R T (1988) Effectsof lesions of temporalndashparietal junction on perceptual andattentional processing in humans Journal of Neuroscience8 3757ndash3769
Rorden C amp Brett M (2000) Stereotaxic display of brainlesions Behavioural Neurology 12 191ndash200
Rusconi M L Maravita A Bottini G amp Vallar G (2002)Is the intact side really intact Perseverative responses inpatients with unilateral neglect A productive manifestationNeuropsychologia 40 594ndash604
Russell C Malhotra P amp Husain M (2004) Attentionmodulates the visual field in healthy observers and parietalpatients NeuroReport 15 189ndash193
Rypma B amp DrsquoEsposito M (2003) A subsequent-memoryeffect in dorsolateral prefrontal cortex Cognitive BrainResearch 16 162ndash166
Sapir A Hayes A Henik A Danzinger S amp Rafal R(2004) Parietal lobe lesions disrupt saccadic remapping ofinhibitory location tagging Journal of CognitiveNeuroscience 16 503ndash509
Smania N Martini M C Gambina G A T Palamara ANatale E amp Marzi C A (1998) The spatial distributionof visual attention in hemineglect and extinction patientsBrain 121 1759ndash1770
Sprenger A Kompf D amp Heide W (2002) Visual searchin patients with left visual hemineglect Progress in BrainResearch 140 395ndash416
Tobler P Felblinger J Burki M Nirkko A Ozdoba C ampMuri R (2001) Functional organisation of the saccadicreference system processing extraretinal signals in humansVision Research 41 1351ndash1358
Wilson B Cockburn J amp Halligan P W (1987)Behavioural inattention test Bury St EdmundsThames Valley
Wojciulik E Husain M Clarke K amp Driver J (2001)Spatial working memory deficit in unilateral neglectNeuropsychologia 39 390ndash396
Zihl J amp Hebel N (1997) Patterns of oculomotor scanningin patients with unilateral posterior parietal and frontallobe damage Neuropsychologia 35 893ndash906
Zihl J amp von Cramon D (1985) Visual field recovery fromscotoma in patients with postgeniculate damage A reviewof 55 cases Brain 108 335ndash365
354 Journal of Cognitive Neuroscience Volume 17 Number 2
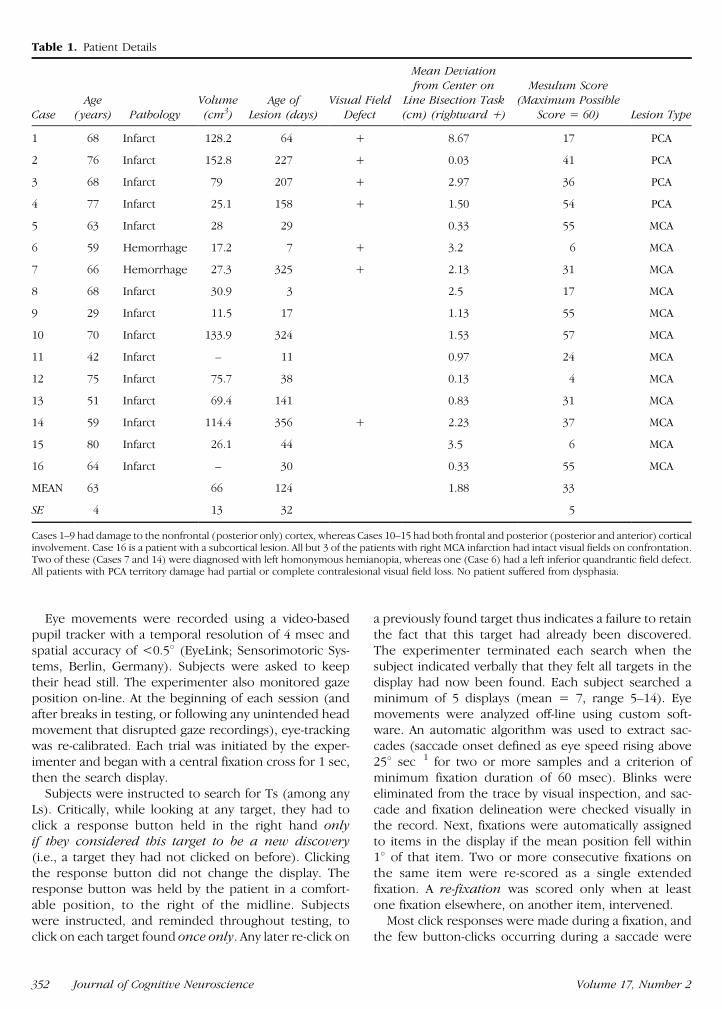
Eye movements were recorded using a video-basedpupil tracker with a temporal resolution of 4 msec andspatial accuracy of lt058 (EyeLink Sensorimotoric Sys-tems Berlin Germany) Subjects were asked to keeptheir head still The experimenter also monitored gazeposition on-line At the beginning of each session (andafter breaks in testing or following any unintended headmovement that disrupted gaze recordings) eye-trackingwas re-calibrated Each trial was initiated by the exper-imenter and began with a central fixation cross for 1 secthen the search display
Subjects were instructed to search for Ts (among anyLs) Critically while looking at any target they had toclick a response button held in the right hand onlyif they considered this target to be a new discovery(ie a target they had not clicked on before) Clickingthe response button did not change the display Theresponse button was held by the patient in a comfort-able position to the right of the midline Subjectswere instructed and reminded throughout testing toclick on each target found once only Any later re-click on
a previously found target thus indicates a failure to retainthe fact that this target had already been discoveredThe experimenter terminated each search when thesubject indicated verbally that they felt all targets in thedisplay had now been found Each subject searched aminimum of 5 displays (mean = 7 range 5ndash14) Eyemovements were analyzed off-line using custom soft-ware An automatic algorithm was used to extract sac-cades (saccade onset defined as eye speed rising above258 sec1 for two or more samples and a criterion ofminimum fixation duration of 60 msec) Blinks wereeliminated from the trace by visual inspection and sac-cade and fixation delineation were checked visually inthe record Next fixations were automatically assignedto items in the display if the mean position fell within18 of that item Two or more consecutive fixations onthe same item were re-scored as a single extendedfixation A re-fixation was scored only when at leastone fixation elsewhere on another item intervened
Most click responses were made during a fixation andthe few button-clicks occurring during a saccade were
Table 1 Patient Details
CaseAge
(years) PathologyVolume(cm3)
Age ofLesion (days)
Visual FieldDefect
Mean Deviationfrom Center on
Line Bisection Task(cm) (rightward +)
Mesulum Score(Maximum Possible
Score = 60) Lesion Type
1 68 Infarct 1282 64 + 867 17 PCA
2 76 Infarct 1528 227 + 003 41 PCA
3 68 Infarct 79 207 + 297 36 PCA
4 77 Infarct 251 158 + 150 54 PCA
5 63 Infarct 28 29 033 55 MCA
6 59 Hemorrhage 172 7 + 32 6 MCA
7 66 Hemorrhage 273 325 + 213 31 MCA
8 68 Infarct 309 3 25 17 MCA
9 29 Infarct 115 17 113 55 MCA
10 70 Infarct 1339 324 153 57 MCA
11 42 Infarct ndash 11 097 24 MCA
12 75 Infarct 757 38 013 4 MCA
13 51 Infarct 694 141 083 31 MCA
14 59 Infarct 1144 356 + 223 37 MCA
15 80 Infarct 261 44 35 6 MCA
16 64 Infarct ndash 30 033 55 MCA
MEAN 63 66 124 188 33
SE 4 13 32 5
Cases 1ndash9 had damage to the nonfrontal (posterior only) cortex whereas Cases 10ndash15 had both frontal and posterior (posterior and anterior) corticalinvolvement Case 16 is a patient with a subcortical lesion All but 3 of the patients with right MCA infarction had intact visual fields on confrontationTwo of these (Cases 7 and 14) were diagnosed with left homonymous hemianopia whereas one (Case 6) had a left inferior quandrantic field defectAll patients with PCA territory damage had partial or complete contralesional visual field loss No patient suffered from dysphasia
352 Journal of Cognitive Neuroscience Volume 17 Number 2
assigned to the previous fixation Clicks were assigned toa particular item if eye position at the time of the clickwas within 28 of that item or in the cases where morethan one item was within 28 of current eye position theclick response was simply attributed to the item closestto that eye position
Button-clicks were assigned to a particular item if eyeposition at the time of the click was within 28 of thatitem A re-click on an item required at least one fixationon another item between successive clicks (in practiceseveral fixations elsewhere typically intervened betweeninitial click and later re-click see Results)
Statistical analyses were performed using SPSS andSigmaplot (SPSS Science Chicago USA) Jonckheere sta-tistic was derived by hand Statistical significance was setas p lt 05
Acknowledgments
We thank the reviewers of this manuscript and Dr AndrewParton for their extremely helpful comments We thank thepatients who participated Dr J Wade Dr A Rudd and thestaff of the stroke units at Charing Cross and St ThomasrsquoHospitals This research was supported by grants from theWellcome Trust to M H C K and J D J D holds a RoyalSociety Wolfson Research Merit Award
Reprint requests should be sent to Drs Sabira Mannan andMasud Husain Division of Neuroscience and PsychologicalMedicine Imperial College London Charing Cross CampusLondon W6 8RF UK or via e-mail smannanimperialacukor mhusainimperialacuk
Notes
1 We are grateful to one of the reviewers for suggesting thistime-series analysis2 There was no significant difference between the number oftargets fixated between the two groups so differences in lsquolsquoclickomissionsrsquorsquo cannot be attributed to the number of fixatedtargets that had to be kept track of during search3 The correlation between these four points alone inFigure 9A is high [r(3) = 81 cf r(15) = 47 for the wholegroup]4 We have also tested 10 neglect patients and 10 controlstroke patients on a vertical version of the Corsi blocks task(clinically used to measure spatial working memory capacity)Neglect patients had significantly impaired capacity but wereable to localize single locations well suggesting that mislocal-ization is unlikely to account for their deficit on this spatialworking memory task
REFERENCES
Bartolomeo P Sieroff E Decaix C amp Chokron S (2001)Modulating the attentional bias in unilateral neglect theeffects of the strategic set Experimental Brain Research137 432ndash444
Bates E Wilson S M Saygin A P Dick F Sereno M IKnight R T amp Dronkers N F (2003) Voxel-basedlesion-symptom mapping Nature Neuroscience 6 448ndash450
Behrmann M Watt S Black S E amp Barton J J (1997)Impaired visual search in patients with unilateral neglect
An oculographic analysis Neuropsychologia 351445ndash1458
Bisiach E amp Vallar G (2000) Unilateral neglect in humans InG Rizzolatti (Ed) Handbook of neuropsychology (2nd edvol 1 pp 459ndash502) Amsterdam Elsevier
Cals N Devuyst G Afsar N Karapanayiotides T ampBogousslavsky J (2002) Pure superficial posterior cerebralartery territory infarction in The Lausanne Stroke RegistryJournal of Neurology 249 855ndash861
Chafee M amp Goldman-Rakic P (1998) Matching patterns ofactivity in primate prefrontal area 8a and parietal area 7ipneurons during a spatial working memory task Journal ofNeurophysiology 79 2919ndash2940
Chedru F Leblanc M amp Lhermitte F (1973) Visualsearching in normal and brain-damaged subjectsContributions to the study of unilateral subjectsCortex 9 94ndash111
DrsquoEsposito M Aguirre G K Zarahn E Ballard DShin R K amp Lease J (1998) Functional MRI studies ofspatial and nonspatial working memory Cognitive BrainResearch 7 1ndash13
Donkelaar P amp Muri R (2002) Craniotopic updating ofvisual space across saccades in the human posteriorparietal cortex Proceedings of the Royal Society London B269 735ndash739
Driver J amp Husain M (2002) The role of spatial workingmemory deficits in pathological search by neglect patientsIn the cognitive and neural bases of spatial neglect InH-O Karnath A D Milner amp G Vallar (Eds) The cognitiveand neural bases of spatial neglect Oxford OxfordUniversity Press
Duhamel J R Colby C amp Goldberg M E (1992) Theupdating of the representation of visual space in parietalcortex by intended eye movements Science 25590ndash92
Goldman-Rakic P S (1996) The prefrontal landscapemdashImplications of functional architecture for understandinghuman mentation and the central executive PhilosophicalTransactions of The Royal Society of London Series BBiological Sciences 351 1445ndash1453
Heide W Binkofski F Seitz R J Posse S Nitschke M FFreund H J amp Kompf D (2001) Activation offrontoparietal cortices during memorized triple-stepsequences of saccadic eye movements An fMRI studyEuropean Journal of Neuroscience 13 1177ndash1189
Heide W amp Kompf D (1998) Combined deficits ofsaccades and visuo-spatial orientation after cortical lesionsExperimental Brain Research 123 164ndash171
Husain M amp Kennard C (1996) Visual neglect associatedwith frontal lobe infarction Journal of Neurology 243652ndash657
Husain M Mannan S Hodgson T Wojciulik E Driver Jamp Kennard C (2001) Impaired spatial working memoryacross saccades contributes to abnormal search in parietalneglect Brain 121 941ndash952
Husain M amp Rorden C (2003) Non-spatially lateralizedmechanisms in hemispatial neglect Nature ReviewsNeuroscience 4 23ndash36
Jackson S R amp Husain M (1996) Visuomotor functionsof the lateral pre-motor cortex Current Opinion inNeurobiology 6 788ndash795
Kinsbourne M (1993) Orientational bias model of unilateralneglect Evidence from attentional gradients withinhemispace In I H Robertson amp J C Marshall (Eds)Unilateral neglect Clinical and experimental studies(pp 63ndash86) Hove Erlbaum
Konishi S Kyoichi N Uchida I Kikyo H Kameyama Mamp Miyauchi S (1999) Common inhibitory mechanism in
Mannan et al 353
human inferior prefrontal cortex revealed by event-relatedfunctional MRI Brain 122 981ndash991
Konishi S Kyoichi N Uchida I Sekihara K amp Miyashita Y(1998) No-go dominant brain activity in human inferiorprefrontal cortex revealed by functional magneticresonance imaging European Journal of Neuroscience10 1209ndash1213
Malhotra P Mannan S Driver J amp Husain M (2004)Impaired spatial working memory One component of thevisual neglect syndrome Cortex 40 667ndash676
Manly T Woldt K Watson P amp Warburton E (2002) Ismotor perseveration in unilateral neglect lsquolsquodrivenrsquorsquo by thepresence of neglected left-sided stimuli Neuropsychologia40 1794ndash1803
Mazzoni P Bracewell R Barash S amp Andersen R (1996)Spatially tuned auditory responses in area LIP of macaquesperforming delayed memory saccades to acoustic targetsJournal of Neurophysiology 75 1233ndash1241
McCarley J Wang R Kramer A Irwin D amp Peterson M(2003) How much memory does oculomotor search havePsychological Science 14 422ndash426
Mesulam M-M (1985) Principles of behavioural neurologyTests of directed attention and memory Philadelphia Davis
Mesulam M M (1999) Spatial attention and neglectParietal frontal and cingulate contributions to the mentalrepresentation and attentional targeting of salientextrapersonal events Philosophical Transactions of theRoyal Society of London Series B Biological Sciences354 1325ndash1346
Mort D Malhotra P Mannan S Rorden C Pambakian AKennard C amp Husain M (2003) The anatomy of visualneglect Brain 126 1986ndash1997
Na D L Adair J C Kang Y Chung C S Lee K H ampHeilman K M (1999) Motor perseverative behaviour ona line cancellation task Neurology 52 1569ndash1576
Pearson D G amp Sahraie A (2003) Oculomotor controland the maintenance of spatially and temporally distributedevents in visuo-spatial working memory QuarterlyJournal of Experimental Psychology 56A 1089ndash1111
Rafal R (1994) Neglect Current Opinion in Neurobiology4 231ndash236
Rizzolatti G amp Arbib M A (1998) Language within our graspTrends in Neurosciences 21 188ndash194
Robertson L C Lamb M R amp Knight R T (1988) Effectsof lesions of temporalndashparietal junction on perceptual andattentional processing in humans Journal of Neuroscience8 3757ndash3769
Rorden C amp Brett M (2000) Stereotaxic display of brainlesions Behavioural Neurology 12 191ndash200
Rusconi M L Maravita A Bottini G amp Vallar G (2002)Is the intact side really intact Perseverative responses inpatients with unilateral neglect A productive manifestationNeuropsychologia 40 594ndash604
Russell C Malhotra P amp Husain M (2004) Attentionmodulates the visual field in healthy observers and parietalpatients NeuroReport 15 189ndash193
Rypma B amp DrsquoEsposito M (2003) A subsequent-memoryeffect in dorsolateral prefrontal cortex Cognitive BrainResearch 16 162ndash166
Sapir A Hayes A Henik A Danzinger S amp Rafal R(2004) Parietal lobe lesions disrupt saccadic remapping ofinhibitory location tagging Journal of CognitiveNeuroscience 16 503ndash509
Smania N Martini M C Gambina G A T Palamara ANatale E amp Marzi C A (1998) The spatial distributionof visual attention in hemineglect and extinction patientsBrain 121 1759ndash1770
Sprenger A Kompf D amp Heide W (2002) Visual searchin patients with left visual hemineglect Progress in BrainResearch 140 395ndash416
Tobler P Felblinger J Burki M Nirkko A Ozdoba C ampMuri R (2001) Functional organisation of the saccadicreference system processing extraretinal signals in humansVision Research 41 1351ndash1358
Wilson B Cockburn J amp Halligan P W (1987)Behavioural inattention test Bury St EdmundsThames Valley
Wojciulik E Husain M Clarke K amp Driver J (2001)Spatial working memory deficit in unilateral neglectNeuropsychologia 39 390ndash396
Zihl J amp Hebel N (1997) Patterns of oculomotor scanningin patients with unilateral posterior parietal and frontallobe damage Neuropsychologia 35 893ndash906
Zihl J amp von Cramon D (1985) Visual field recovery fromscotoma in patients with postgeniculate damage A reviewof 55 cases Brain 108 335ndash365
354 Journal of Cognitive Neuroscience Volume 17 Number 2

assigned to the previous fixation Clicks were assigned toa particular item if eye position at the time of the clickwas within 28 of that item or in the cases where morethan one item was within 28 of current eye position theclick response was simply attributed to the item closestto that eye position
Button-clicks were assigned to a particular item if eyeposition at the time of the click was within 28 of thatitem A re-click on an item required at least one fixationon another item between successive clicks (in practiceseveral fixations elsewhere typically intervened betweeninitial click and later re-click see Results)
Statistical analyses were performed using SPSS andSigmaplot (SPSS Science Chicago USA) Jonckheere sta-tistic was derived by hand Statistical significance was setas p lt 05
Acknowledgments
We thank the reviewers of this manuscript and Dr AndrewParton for their extremely helpful comments We thank thepatients who participated Dr J Wade Dr A Rudd and thestaff of the stroke units at Charing Cross and St ThomasrsquoHospitals This research was supported by grants from theWellcome Trust to M H C K and J D J D holds a RoyalSociety Wolfson Research Merit Award
Reprint requests should be sent to Drs Sabira Mannan andMasud Husain Division of Neuroscience and PsychologicalMedicine Imperial College London Charing Cross CampusLondon W6 8RF UK or via e-mail smannanimperialacukor mhusainimperialacuk
Notes
1 We are grateful to one of the reviewers for suggesting thistime-series analysis2 There was no significant difference between the number oftargets fixated between the two groups so differences in lsquolsquoclickomissionsrsquorsquo cannot be attributed to the number of fixatedtargets that had to be kept track of during search3 The correlation between these four points alone inFigure 9A is high [r(3) = 81 cf r(15) = 47 for the wholegroup]4 We have also tested 10 neglect patients and 10 controlstroke patients on a vertical version of the Corsi blocks task(clinically used to measure spatial working memory capacity)Neglect patients had significantly impaired capacity but wereable to localize single locations well suggesting that mislocal-ization is unlikely to account for their deficit on this spatialworking memory task
REFERENCES
Bartolomeo P Sieroff E Decaix C amp Chokron S (2001)Modulating the attentional bias in unilateral neglect theeffects of the strategic set Experimental Brain Research137 432ndash444
Bates E Wilson S M Saygin A P Dick F Sereno M IKnight R T amp Dronkers N F (2003) Voxel-basedlesion-symptom mapping Nature Neuroscience 6 448ndash450
Behrmann M Watt S Black S E amp Barton J J (1997)Impaired visual search in patients with unilateral neglect
An oculographic analysis Neuropsychologia 351445ndash1458
Bisiach E amp Vallar G (2000) Unilateral neglect in humans InG Rizzolatti (Ed) Handbook of neuropsychology (2nd edvol 1 pp 459ndash502) Amsterdam Elsevier
Cals N Devuyst G Afsar N Karapanayiotides T ampBogousslavsky J (2002) Pure superficial posterior cerebralartery territory infarction in The Lausanne Stroke RegistryJournal of Neurology 249 855ndash861
Chafee M amp Goldman-Rakic P (1998) Matching patterns ofactivity in primate prefrontal area 8a and parietal area 7ipneurons during a spatial working memory task Journal ofNeurophysiology 79 2919ndash2940
Chedru F Leblanc M amp Lhermitte F (1973) Visualsearching in normal and brain-damaged subjectsContributions to the study of unilateral subjectsCortex 9 94ndash111
DrsquoEsposito M Aguirre G K Zarahn E Ballard DShin R K amp Lease J (1998) Functional MRI studies ofspatial and nonspatial working memory Cognitive BrainResearch 7 1ndash13
Donkelaar P amp Muri R (2002) Craniotopic updating ofvisual space across saccades in the human posteriorparietal cortex Proceedings of the Royal Society London B269 735ndash739
Driver J amp Husain M (2002) The role of spatial workingmemory deficits in pathological search by neglect patientsIn the cognitive and neural bases of spatial neglect InH-O Karnath A D Milner amp G Vallar (Eds) The cognitiveand neural bases of spatial neglect Oxford OxfordUniversity Press
Duhamel J R Colby C amp Goldberg M E (1992) Theupdating of the representation of visual space in parietalcortex by intended eye movements Science 25590ndash92
Goldman-Rakic P S (1996) The prefrontal landscapemdashImplications of functional architecture for understandinghuman mentation and the central executive PhilosophicalTransactions of The Royal Society of London Series BBiological Sciences 351 1445ndash1453
Heide W Binkofski F Seitz R J Posse S Nitschke M FFreund H J amp Kompf D (2001) Activation offrontoparietal cortices during memorized triple-stepsequences of saccadic eye movements An fMRI studyEuropean Journal of Neuroscience 13 1177ndash1189
Heide W amp Kompf D (1998) Combined deficits ofsaccades and visuo-spatial orientation after cortical lesionsExperimental Brain Research 123 164ndash171
Husain M amp Kennard C (1996) Visual neglect associatedwith frontal lobe infarction Journal of Neurology 243652ndash657
Husain M Mannan S Hodgson T Wojciulik E Driver Jamp Kennard C (2001) Impaired spatial working memoryacross saccades contributes to abnormal search in parietalneglect Brain 121 941ndash952
Husain M amp Rorden C (2003) Non-spatially lateralizedmechanisms in hemispatial neglect Nature ReviewsNeuroscience 4 23ndash36
Jackson S R amp Husain M (1996) Visuomotor functionsof the lateral pre-motor cortex Current Opinion inNeurobiology 6 788ndash795
Kinsbourne M (1993) Orientational bias model of unilateralneglect Evidence from attentional gradients withinhemispace In I H Robertson amp J C Marshall (Eds)Unilateral neglect Clinical and experimental studies(pp 63ndash86) Hove Erlbaum
Konishi S Kyoichi N Uchida I Kikyo H Kameyama Mamp Miyauchi S (1999) Common inhibitory mechanism in
Mannan et al 353
human inferior prefrontal cortex revealed by event-relatedfunctional MRI Brain 122 981ndash991
Konishi S Kyoichi N Uchida I Sekihara K amp Miyashita Y(1998) No-go dominant brain activity in human inferiorprefrontal cortex revealed by functional magneticresonance imaging European Journal of Neuroscience10 1209ndash1213
Malhotra P Mannan S Driver J amp Husain M (2004)Impaired spatial working memory One component of thevisual neglect syndrome Cortex 40 667ndash676
Manly T Woldt K Watson P amp Warburton E (2002) Ismotor perseveration in unilateral neglect lsquolsquodrivenrsquorsquo by thepresence of neglected left-sided stimuli Neuropsychologia40 1794ndash1803
Mazzoni P Bracewell R Barash S amp Andersen R (1996)Spatially tuned auditory responses in area LIP of macaquesperforming delayed memory saccades to acoustic targetsJournal of Neurophysiology 75 1233ndash1241
McCarley J Wang R Kramer A Irwin D amp Peterson M(2003) How much memory does oculomotor search havePsychological Science 14 422ndash426
Mesulam M-M (1985) Principles of behavioural neurologyTests of directed attention and memory Philadelphia Davis
Mesulam M M (1999) Spatial attention and neglectParietal frontal and cingulate contributions to the mentalrepresentation and attentional targeting of salientextrapersonal events Philosophical Transactions of theRoyal Society of London Series B Biological Sciences354 1325ndash1346
Mort D Malhotra P Mannan S Rorden C Pambakian AKennard C amp Husain M (2003) The anatomy of visualneglect Brain 126 1986ndash1997
Na D L Adair J C Kang Y Chung C S Lee K H ampHeilman K M (1999) Motor perseverative behaviour ona line cancellation task Neurology 52 1569ndash1576
Pearson D G amp Sahraie A (2003) Oculomotor controland the maintenance of spatially and temporally distributedevents in visuo-spatial working memory QuarterlyJournal of Experimental Psychology 56A 1089ndash1111
Rafal R (1994) Neglect Current Opinion in Neurobiology4 231ndash236
Rizzolatti G amp Arbib M A (1998) Language within our graspTrends in Neurosciences 21 188ndash194
Robertson L C Lamb M R amp Knight R T (1988) Effectsof lesions of temporalndashparietal junction on perceptual andattentional processing in humans Journal of Neuroscience8 3757ndash3769
Rorden C amp Brett M (2000) Stereotaxic display of brainlesions Behavioural Neurology 12 191ndash200
Rusconi M L Maravita A Bottini G amp Vallar G (2002)Is the intact side really intact Perseverative responses inpatients with unilateral neglect A productive manifestationNeuropsychologia 40 594ndash604
Russell C Malhotra P amp Husain M (2004) Attentionmodulates the visual field in healthy observers and parietalpatients NeuroReport 15 189ndash193
Rypma B amp DrsquoEsposito M (2003) A subsequent-memoryeffect in dorsolateral prefrontal cortex Cognitive BrainResearch 16 162ndash166
Sapir A Hayes A Henik A Danzinger S amp Rafal R(2004) Parietal lobe lesions disrupt saccadic remapping ofinhibitory location tagging Journal of CognitiveNeuroscience 16 503ndash509
Smania N Martini M C Gambina G A T Palamara ANatale E amp Marzi C A (1998) The spatial distributionof visual attention in hemineglect and extinction patientsBrain 121 1759ndash1770
Sprenger A Kompf D amp Heide W (2002) Visual searchin patients with left visual hemineglect Progress in BrainResearch 140 395ndash416
Tobler P Felblinger J Burki M Nirkko A Ozdoba C ampMuri R (2001) Functional organisation of the saccadicreference system processing extraretinal signals in humansVision Research 41 1351ndash1358
Wilson B Cockburn J amp Halligan P W (1987)Behavioural inattention test Bury St EdmundsThames Valley
Wojciulik E Husain M Clarke K amp Driver J (2001)Spatial working memory deficit in unilateral neglectNeuropsychologia 39 390ndash396
Zihl J amp Hebel N (1997) Patterns of oculomotor scanningin patients with unilateral posterior parietal and frontallobe damage Neuropsychologia 35 893ndash906
Zihl J amp von Cramon D (1985) Visual field recovery fromscotoma in patients with postgeniculate damage A reviewof 55 cases Brain 108 335ndash365
354 Journal of Cognitive Neuroscience Volume 17 Number 2

human inferior prefrontal cortex revealed by event-relatedfunctional MRI Brain 122 981ndash991
Konishi S Kyoichi N Uchida I Sekihara K amp Miyashita Y(1998) No-go dominant brain activity in human inferiorprefrontal cortex revealed by functional magneticresonance imaging European Journal of Neuroscience10 1209ndash1213
Malhotra P Mannan S Driver J amp Husain M (2004)Impaired spatial working memory One component of thevisual neglect syndrome Cortex 40 667ndash676
Manly T Woldt K Watson P amp Warburton E (2002) Ismotor perseveration in unilateral neglect lsquolsquodrivenrsquorsquo by thepresence of neglected left-sided stimuli Neuropsychologia40 1794ndash1803
Mazzoni P Bracewell R Barash S amp Andersen R (1996)Spatially tuned auditory responses in area LIP of macaquesperforming delayed memory saccades to acoustic targetsJournal of Neurophysiology 75 1233ndash1241
McCarley J Wang R Kramer A Irwin D amp Peterson M(2003) How much memory does oculomotor search havePsychological Science 14 422ndash426
Mesulam M-M (1985) Principles of behavioural neurologyTests of directed attention and memory Philadelphia Davis
Mesulam M M (1999) Spatial attention and neglectParietal frontal and cingulate contributions to the mentalrepresentation and attentional targeting of salientextrapersonal events Philosophical Transactions of theRoyal Society of London Series B Biological Sciences354 1325ndash1346
Mort D Malhotra P Mannan S Rorden C Pambakian AKennard C amp Husain M (2003) The anatomy of visualneglect Brain 126 1986ndash1997
Na D L Adair J C Kang Y Chung C S Lee K H ampHeilman K M (1999) Motor perseverative behaviour ona line cancellation task Neurology 52 1569ndash1576
Pearson D G amp Sahraie A (2003) Oculomotor controland the maintenance of spatially and temporally distributedevents in visuo-spatial working memory QuarterlyJournal of Experimental Psychology 56A 1089ndash1111
Rafal R (1994) Neglect Current Opinion in Neurobiology4 231ndash236
Rizzolatti G amp Arbib M A (1998) Language within our graspTrends in Neurosciences 21 188ndash194
Robertson L C Lamb M R amp Knight R T (1988) Effectsof lesions of temporalndashparietal junction on perceptual andattentional processing in humans Journal of Neuroscience8 3757ndash3769
Rorden C amp Brett M (2000) Stereotaxic display of brainlesions Behavioural Neurology 12 191ndash200
Rusconi M L Maravita A Bottini G amp Vallar G (2002)Is the intact side really intact Perseverative responses inpatients with unilateral neglect A productive manifestationNeuropsychologia 40 594ndash604
Russell C Malhotra P amp Husain M (2004) Attentionmodulates the visual field in healthy observers and parietalpatients NeuroReport 15 189ndash193
Rypma B amp DrsquoEsposito M (2003) A subsequent-memoryeffect in dorsolateral prefrontal cortex Cognitive BrainResearch 16 162ndash166
Sapir A Hayes A Henik A Danzinger S amp Rafal R(2004) Parietal lobe lesions disrupt saccadic remapping ofinhibitory location tagging Journal of CognitiveNeuroscience 16 503ndash509
Smania N Martini M C Gambina G A T Palamara ANatale E amp Marzi C A (1998) The spatial distributionof visual attention in hemineglect and extinction patientsBrain 121 1759ndash1770
Sprenger A Kompf D amp Heide W (2002) Visual searchin patients with left visual hemineglect Progress in BrainResearch 140 395ndash416
Tobler P Felblinger J Burki M Nirkko A Ozdoba C ampMuri R (2001) Functional organisation of the saccadicreference system processing extraretinal signals in humansVision Research 41 1351ndash1358
Wilson B Cockburn J amp Halligan P W (1987)Behavioural inattention test Bury St EdmundsThames Valley
Wojciulik E Husain M Clarke K amp Driver J (2001)Spatial working memory deficit in unilateral neglectNeuropsychologia 39 390ndash396
Zihl J amp Hebel N (1997) Patterns of oculomotor scanningin patients with unilateral posterior parietal and frontallobe damage Neuropsychologia 35 893ndash906
Zihl J amp von Cramon D (1985) Visual field recovery fromscotoma in patients with postgeniculate damage A reviewof 55 cases Brain 108 335ndash365
354 Journal of Cognitive Neuroscience Volume 17 Number 2
Recommended

















