
STUDIES OF HTLV-1 p12(I) IN CALCINEURIN BINDING, CALCIUM-
MEDIATED CELL SIGNALLING AND VIRAL TRANSMISSION
DISSERTATION
Presented in Partial Fulfillment of the Requirements for
the Degree Doctor of Philosophy in the Graduate
School of The Ohio State University
By
Seung-jae Kim, D.V.M., M.S.
* * * * *
The Ohio State University
2006
Dissertation Committee:
Professor Michael D. Lairmore, Adviser Approved by
Professor Patrick Green
Professor Kathleen Boris-Lawrie ________________________
Professor Natarajan Muthusamy Adviser
Veterinary Biosciences Graduate program

ii
ABSTRACT
Human T- lymphotropic virus type 1 (HTLV-1) causes a variety of
lymphoproliferative and neurodegenerative diseases. The mechanisms of early viral
infection, such as virus-mediated T cell activation, cell-to-cell transmission, and early
regulation of viral and cellular gene transcription are incompletely understood. The pX
ORF I encoded protein p12I is critically required for productive infection in a rabbit
model and viral infectivity in non-stimulated PBMC. HTLV-1 p12I regulates calcium-
mediated signaling in T cells, and induces activation of NFAT and enhancement of IL-2
production and p300 expression.
We identified a PSLPI/LTmotif in p12I, which is highly homologous to the
PxIxIT calcineurin-binding motif of NFAT. Full-length p12I and PSLPI/LT motif
containing mutants bound calcineurin in both immunoprecipitation and calmodulin bead
pull-down assays. In contrast, serial mutations of p12I that lacked the PSLPI/LT motif
or had selective alanine substitutions of the motif (p12I AxAxAA) exhibited abolished
or decreased binding affinity with calcineurin. We then tested if p12I binding to
calcineurin affected NFAT activity. p12I competed with NFAT for calcineurin binding
in calmodulin bead pull-down experiments. Furthermore, the p12I AxAxAA mutant
enhanced NFAT nuclear translocation compared to wild type p12I and increased NFAT
transcriptional activity two fold greater than wild type p12I. Thus the reduced

iii
binding of p12I to calcineurin allows enhanced nuclear translocation and transcription
mediated by NFAT, suggesting that HTLV-1 p12I modulates NFAT activation to
promote early virus infection of T lymphocytes.
We further tested the role of p12I on early HTLV-1 cell-to-cell transmission,
particularly, the role of p12I on LFA-1-mediated T cell adhesion. Our data indicated
that abrogation of pX ORF I mRNA expression in HTLV-1 infected cells (ACH.p12I)
resulted in reduction of LFA-1-mediated adhesion compared to wild-type HTLV-1
expressing cells (ACH). Furthermore, expression of p12I in Jurkat T-cells using
lentiviral vectors, enhanced LFA-1-mediated cell adhesion, which was inhibited by the
inhibitors of calcium-mediated signaling such as BAPTA-AM, SK&F 96365 and
calpeptin. Similar to the intracellular calcium mobilizer, thapsigargin, the expression of
p12I in Jurkat T-cells induced cell surface clustering of LFA-1 without changing the
level of integrin expression. Our data indicated that HTLV-1 p12I promotes cell-to-cell
spread by inducing LFA-1 clustering on T-cells via calcium-dependent signaling.
Lastly, we investigated the role of expression of HTLV-1 pX ORF I in early
HTLV-1 transmission. We compared the activation status between wild type ACH cells
and pX ORF I expression abrogated ACH.p12 cells. We then tested if the expression of
ORF I is required to induce bystander activation of uninfected target cells by
performing both proliferation assays and flow cytometric analysis of target cells.

iv
We found that abrogation of pX ORF I message decreased CD69 expression in HTLV-1
infected cells, but did not affect the HTLV-1- induced bystander target cell activation.
Furthermore, we performed real-time RT-PCR to detect pX ORF I and pX ORF III/IV
mRNA after coculture of both wild type ACH and ACH.p12 cells with pre-stimulated
or non-stimulated T cells. By measuring Tax/Rex mRNA in target cells, we were able to
compare viral infectivity of wild type ACH to ACH.p12 immortalized T cells and viral
mRNA expression in newly infected target cells. Our data indicated that expression of
pX ORF I is required for efficient HTLV-1 transmission to target cells and identified
differential expression patterns of Tax/Rex and pX ORF I mRNA during early cell-to-
cell transmission.
In conclusion, HTLV-1 p12I modulates calcium-mediated signaling and induces
enhancement of viral cell-to-cell transmission by inducing LFA-1 clustering and
facilitating the early establishment of infection by regulating viral gene expression,
providing further evidence that p12I is critically required for early HTLV-1 infection.

v
Dedicated to Rokyoun, Thomas, and my parents

vi
ACKNOWLEDGMENTS
First and foremost, I would like to thank my advisor, Dr. Michael Lairmore for
his valuable guidance and support throughout my graduate training. His offer to join his
laboratory provided me opportunity to start my carrer as a scientist. I really appreciate
his support and commitment to instructing me to allow me to fufill my academic goal.
He has always supported my imperfect ideas and helped me to develop the ability to
conduct scientific independent research that will unlimitedly benefit my future career in
every aspect.
I would like to thank my committee members Drs. Patrick Green, Kathleen
Boris-Lawrie and Natarajan Muthusamy, who have provided valuable guidance and
discussion on my thesis. I would like to express my gratitude to other members of the
Center for Retrovirus Research, in particular Dr. Larry Mathes, faculty members and
students, who have shared helpful ideas and discussion on my projects.
My studies could not have been succeful without the support of my calleagues
who worked together to aid in the completion of my research. I would like to thank all
my previous and current lab members, Wei Ding, Bjoern Albrecht, James Stanely,
Jesica Alcorn, Hajime Hiraragi, Lee Silverman, Amrithraj Nair, Bindhu Michael,

vii
Andrew Phipps, Antara Datta, Chris Premandandan, Rashade Haynes, Andy
Montgomery, John Nisbet, Laurie Millward,and Bevin Zimmerman. In particular, I
thank Wei Ding, Bjoern Albrecht, and Amrithraj Nair who helped me to perform many
experiments and shared their skills and ideas for my projects. I appreciate Min Li who
helped me to carry out real time PCR and shared her reagents. I am indebted to the
technical assistance provided by Rick Meister and Bryan McElwain, who helped flow
cytometrical anlalysis. I also thank Soledad Fernandez for statiscal analysis and Tim
Vojt for helping figure preparation.
Without continous prays and steadfast devotes of my wife, Rokyoun and my
parents, I would not be where I am today. They have always encouraged me to do my
best. Lastly, I appreciate my son, Thomas, who has always made me happy.

viii
VITA
June 19, 1972 ------------------------------ Born – Gwangju, South Korea 1995----------------------------------------- B.S. & D.V.M. Chonnam National University, Gwangju, South Korea 1995 - 1997 -------------------------------- Research Associate, Chonnam National University, Gwangju, South Korea 1997----------------------------------------- M.S. (Veterinary Pathobiology) Chonnam National University, Gwangju, South Korea 1998 - 1999 -------------------------------- Research Internship
Korean Science and Engineering Foundatoin Chonnam National University,
Gwangju, South Korea 1999 – present ----------------------------- Graduate Research Associate Department of Veterinary Biosciences The Ohio State University, Columbus, Ohio
PUBLICATIONS
Research Publications 1. Kim SJ, Nair AM, Fernandez S, Mathes L, and Lairmore MD. (2006) Enhancement of LFA-1-mediated T cell adhesion by Human T lymphotropic virus type 1 p12I Journal of Immunology, 176(9):5463-5470. 2. Hiraragi H, Kim SJ, Phipps AJ, Silic-Benussi M, Ciminale V, Ratner L, Green PL, and Lairmore MD. (2005) Human T-lymphotropic virus type 1 mitochondria localizing protein p13II is required for viral Infectivity in vivo. Journal of Virology, 80(7):3469-76.

ix
3. Kim SJ, Ding W, Albrecht B, Green PL, and Lairmore MD. (2003) A conserved calcineurin-binding motif in human T lymphotropic virus type 1 p12I functions to modulate nuclear factor of activated T cell activation. Journal of Biological Chemistry, 278(18):15550-15557. 4. Ding W, Kim SJ, Nair AM, Michael B, Boris-Lawrie K, Tripp A, Feuer G, and Lairmore MD. (2003) Human T-cell lymphotropic virus type 1 p12I enhances interleukin-2 production during T-cell activation. Journal of Virology, 77(20):11027-11039. 5. Ding W, Albrecht B, Kelley RE, Muthusamy N, Kim SJ, Altschuld RA, and Lairmore MD. (2002). Human T-cell lymphotropic virus type 1 p12(I) expression increases cytoplasmic calcium to enhance the activation of nuclear factor of activated T cells. Journal of Virology, 76(20):10374-82 6. Kim SJ and Park NY. (1997) Application of In situ hybridization for diagnosis of porcine reproductive and respiratory syndrome. Korean Journal of Veterinary Research (in Korean) 37(4), 793-807.
FIELDS OF STUDY
Major Field: Veterinary Biosciences

x
TABLE OF CONTENTS
Page Abstract--------------------------------------------------------------------------------------------- ii
Dedication ----------------------------------------------------------------------------------------- v
Acknowledgments --------------------------------------------------------------------------------vi
Vita------------------------------------------------------------------------------------------------ viii
List of Figures ----------------------------------------------------------------------------------- xiii
Chapters:
1. Literature Review: Molecular mechanism of Human T-Lymphotropic Virus Type-1 Infection, T Cell Activation and Transmission ----------------------------- 1
1.1 HTLV-1 Epidemiology--------------------------------------------------------- 1 1.2 HTLV-1- Associated Diseases ------------------------------------------------ 3
1.2.1 Adult T cell Leukemia/Lymphoma ------------------------------------ 3 1.2.2 HTLV-1- Associated Myelopathy/Tropical Spastic Paraparesis --- 5 1.2.3 Other Diseases Associated with HTLV-1-Infection ----------------- 6
1.3 HTLV-1 Viral Structure and Genome Organization ----------------------- 7 1.4 Structural and Enzymatic proteins of HTLV-1------------------------------ 9 1.5 Replication Cycle of HTLV-1 ----------------------------------------------- 10 1.6 HTLV-1 Regulatory Proteins: Tax and Rex ------------------------------- 12
1.6.1 Tax ---------------------------------------------------------------------- 12 1.6.2 Rex ---------------------------------------------------------------------- 16
1.7 HTLV-1 Non-Structural Proteins ------------------------------------------- 18 1.7.1 Minus Strand Encoded HBZ ------------------------------------------- 20

xi
1.7.2 pX ORF II Protein: p13II and p30II ------------------------------------ 21
1.7.2.1 p13II – Role in Viral Replication and Cell Survival --------- 22 1.7.2.2 p30II - A Selective Repressor of Transcription --------------- 24 1.7.2 pX ORF I Protein: p12I ------------------------------------------------ 26 1.7.3.1 Structure of p12I-------------------------------------------------- 26 1.7.3.2 p12I Subcellular Localization and Protein Interaction ------ 28 1.7.3.3 Role of p12I in Viral Infectivity-------------------------------- 31 1.7.3.4 Role p12I in Calcium- Mediated T Cell Signaling Pathway 32
1.8 HTLV-1 Cell-to-cell Transmission ----------------------------------------- 36 1.9 Regulation of Integrin --------------------------------------------------------- 38 1.10 Bystander Cell Activation by HTLV-1-------------------------------------- 41 1.11 References----------------------------------------------------------------------- 43
2. A Conserved Calcineurin-binding Motif in Human T Lymphotropic Virus Type 1 p12I Functions to Modulate NFAT Activation. ----------------------------------- 78
2.1 Introduction --------------------------------------------------------------------- 78 2.2 Materials and Methods -------------------------------------------------------- 81 2.3 Results Discussion ------------------------------------------------------------- 87 2.4 Discussion----------------------------------------------------------------------- 94 2.5 References----------------------------------------------------------------------- 97
3. Enhancement of LFA-1-Mediated T-Cell Adhesion by Human T-Lymphotropic Virus type 1 p12I --------------------------------------------------------------------- 111
3.1 Introduction ------------------------------------------------------------------- 111 3.2 Materials and Methods ------------------------------------------------------ 114 3.3 Results ------------------------------------------------------------------------- 118 3.4 Discussion--------------------------------------------------------------------- 124 3.5 References--------------------------------------------------------------------- 126

xii
4. Expression of HTLV-1 pX openreading frame I enhances early viral infectivity during cell-to-cell transmission in T-lymphocytes-------------------------------- 138
4.1 Introduction ------------------------------------------------------------------- 138 4.2 Materials and Methods ------------------------------------------------------ 142 4.3 Results ------------------------------------------------------------------------- 146 4.4 Discussion--------------------------------------------------------------------- 154 4.5 References--------------------------------------------------------------------- 159
5. Synopsis and Future Directions ----------------------------------------------------- 177
5.1 Studies to test the role of p12I binding to calcineurin on HTLV-1-mediated T cell activation and in vivo and in vitro viral infectivity --- 178
5.2 Further investigation of the role of p12I in MTOC polarization and virological synapse formation ---------------------------------------------- 179
5.3 Studies to explore the mechanisms of LFA-1 affinity regulation influenced by p12I expression ---------------------------------------------- 180
5.4 Studies required to test the role of p12I in HTLV-1 envelope glycoprotein folding and cell surface expression ------------------------ 181
5.5 Possible role of p12I in regulating CD69 expression during HTLV-1 infection ----------------------------------------------------------------------- 182
5.6 The potential role of p12I in regulation of viral gene expression during HTLV-1 cell-to-cell transmission ------------------------------------------ 182
5.7 Studies to test the influence of pX OFR I or p12I in early expression of HTLV-1 regulatory proteins ------------------------------------------------ 184
5.8 References--------------------------------------------------------------------- 185
Bibliography ------------------------------------------------------------------------------------ 188

xiii
LIST OF FIGURES
Figure Page 1.1 Schematic illustration of HTLV-1 proviral genome, mRNA and protein
species ------------------------------------------------------------------------------------ 75
1.2 Schematic Illustration of HTLV-1 non-structural protein p12I-------------------- 76
1.3 A model of p12I role in calcium-mediated signaling ------------------------------- 77
2.1 HTLV-1 p12I contains a highly conserved putative calcineurin-binding motif in p12I --------------------------------------------------------------------------- 102
2.2 The C-terminal half of p12I containing the PSLPI/LT motif is required for calcineurin binding --------------------------------------------------------------- 103
2.3 Alanine substitution mutant of p12I (AxAxAA) decreases binding affinity for calcineurin---------------------------------------------------------------- 104
2.4 p12I and calcineurin binding is inhibited by calcium chelators but not inhibited by cyclosporin A ---------------------------------------------------------- 105
2.5 p12I binding to calcineurin decreases the amount of NFAT binding to calcineurin ----------------------------------------------------------------------------- 106
2.6 The substitution mutation in calcineurin binding sequence in p12I induces increased NFAT transcription activity and did not compete against wild type p12I for NFAT transcription activity ------------------------- 107
2.7 Reduced p12I binding to calcineurin induces increased NFAT nuclear translocation --------------------------------------------------------------------------- 108

xiv
2.8 Calcineurin phosphatase activity is not affected by p12I binding to calcineurin ----------------------------------------------------------------------------- 110
3.1 LFA-1 mediated adhesion is reduced in ACH.p12I cell lines ------------------- 132
3.2 ACH.p12I cells have decreased binding of sICAM-1 without alteration of LFA-1 expression -------------------------------------------------------------------- 133
3.3 p12I stable expression in Jurkat T-cells did not alter LFA-1 affinity ---------- 134
3.4 Expression of p12I in Jurkat T-cells induced LFA-1-mediated cell adhesion ------------------------------------------------------------------------------- 135
3.5 HTLV-1 p12I-mediated LFA-1 activation is inhibited by calcium signal inhibitors ------------------------------------------------------------------------------- 136
3.6 Expression of p12I in Jurkat T-cells modulated surface distribution of LFA-1 on the cell membrane -------------------------------------------------------- 137
4.1 Cell surface expression of early activation markers and adhesion molecules on wild type HTLV-1 infected cells and T cells lacking pX ORF I expression --------------------------------------------------------------------- 165
4.2 Deletion of pX ORF I expression in HTLV-1 infected cells did not affect bystander proliferation of uninfected target PBMC ------------------------------ 167
4.3 Abrogation of pX ORF I expression in HTLV-1 infected cells did not affect bystander activation of target PBMC----------------------------------------------- 168
4.4 HTLV-1 pX ORF I and pX ORF III/IV mRNAs were differentially expressed during the early HTLV-1 transmission --------------------------------------------- 171
4.5 Target PBMC were selectively sorted from HTLV-1 infected cells------------ 173

xv
4.6 Expression of pX ORF I enhanced HTLV-1 viral transmission to target PBMC-- 174
4.7 Expression of HTLV-1 pX ORF I and pX ORF III/IV mRNAs in de novo HTLV--1 infected target PBMC --------------------------------------------------------- 175

1
CHAPTER 1
LITERATURE REVIEW
MOLECULAR MECHANISM OF HTLV-1 INFECTION, T CELL
ACTIVATION, AND TRANSMISSION
1.1 HTLV-1 Epidemiology
The first identified human retrovirus, human T lymphotropic virus type 1
(HTLV-1) was detected as type C particles propagated from T cell lymphoblastoid cell
lines and peripheral blood lymphocytes of a cutaneous T cell lymphoma patient by
Poiesz, Gallo and their colleagues in 1980 1. Subsequent epidemiologic, immunologic,
and molecular biologic studies 2-5 during the early 1980’s demonstrated that HTLV-1
was the etiological agent of adult T-cell leukemia/lymphoma (ATLL) that was
previously reported in Japan in 1977 6,7. By the mid-1980’s, HTLV-1 was also
confirmed as a causative agent of a degenerative neurologic disease, HTLV-1-
associated myelopathy/tropical spastic paraparesis (HAM/TSP) 8,9.

2
HTLV-1 infections have been reported throughout the world and it is estimated
that 15 to 25 million people are infected 10,11. Infected individuals however are primarily
confined in highly endemic areas of southern Japan 3, the Caribbean 12, central Africa 13,
Central and South America 14,15, Melanesian Islands in the Pacific basin 16 and among
certain high risk groups within Europe and the United states of America (USA) 17.
Within these endemic areas, the seroprevalence rate among general population varies
between 0.1 % and 30 %, and the rate increases with age and is higher in females than
males 11,18.
Unlike human immunodeficiency virus 1 (HIV-1), HTLV-1 is poorly infectious
as cell-free virions 19. Cell-cell contact between virus infected and target cells is
required for efficient transmission 20. Breast feeding and perinatal contamination of the
infant with blood of an infected mother are major routes of transmission in endemic
areas 21,22, but transmission of HTLV-1 via trans placenta route is considered extremely
rare 23. The risk of HTLV-1 infection via these routes are correlated to maternal factors
such as high HTLV-1 antibody titer, prolonged ruptured membranes during delivery,
and low socioeconomic status 24. Exposure to infected blood or blood products is
another major route of HTLV-1 transmission 25. The most common cause of blood-to-
blood transmission occurs among intravenous drug users by sharing needles 26,27.
Because transmission through infected blood products has been a major public health
concern, all blood products are screened in Japan, the USA and Brazil 11,28-33. Sexual
transmission is a less efficient mode of HTLV-1 transmission 34. Male to female
transmission via semen is about four times as frequent as female to male transmission 34.

3
For diagnosis of HTLV-1 infection, enzyme linked immunosorbent assays
(ELISA) 35-37 in USA, and particle agglutination assays in Japan are widely used for
detection of anti-HTLV-1 antibodies. After these prototype screening assays,
polymerase chain reaction (PCR) or western blot assays are performed as confirmation
tests 38.
1.2 HTLV-1 Associated Diseases
Approximately 5% of HTLV-1 infected people develop HTLV-1- mediated
disease and the risk increases to 8-10 % when the patient has other illnesses 11. Most
HTLV-1 carriers remain asymptomatic throughout their lives and only develop disease
after months to years of infection 39. HTLV-1 infection is strongly associated with
ATLL and HAM/HSP, and has been implicated to cause or complicate immune
mediated conditions.
1.2.1 Adult T Cell Leukemia/ Lymphoma (ATLL)
In 1977, ATLL was recognized from epidemiological studies in southwest
Japan 6,7. Geographically clustered patients with lymphoid neoplasms were just
identified because of their unique clinical features 6,7. Characteristics of ATLL include:
adult onset, acute or chronic leukemia with rapid progression, resistance to treatment,
peripheral, pleomorphic leukemic cells with markedly deformed nuclei, frequent
lymphadenopathy, hepatosplenomegaly and hypercalcemia, absence of mediastinal
tumors, and frequent skin lesions 2. Similar to non-Hodgkin’s lymphoma, affected

4
patients present with malaise, fever, jaundice, drowsiness, weight loss, and
opportunistic infections 2. The diagnosis of ATLL is made based on specific parameters
including sero-positivity to HTLV-1, marked leukocytosis, morphology of neoplastic T
cells (“cerbriform” or “flower cell”), T cell immunophenotyping, hypercalcemia,
increased circulating levels of the IL-2 receptor α-chain (IL-2Rα/CD25) and elevated
serum lactate dehydrogenase (LDH) levels 40. The predominant phenotype of ATLL
neoplastic cells is characteristic of helper T cells: CD3+, CD4+, L-Selectin+, CD25+,
CD45RA+, HLA-DR+, CD29-, and CD45RO- in circulating peripheral blood, or CD3+,
CD4+, L-Selectin+, CD29+, CD45RO+, HLA-DR+, and CD45RA- in cutaneous and
lymphoma lesions 41,42.
ATLL can be classified into four subcategories (smoldering, chronic, acute, and
lymphoma) based on clinical and laboratory features including the percentage of
abnormal T cells in the peripheral blood, blood LDH and calcium levels, and malignant
tumors in various organs 40,43. Smoldering ATLL, which is characterized by the
presence of a few neoplastic cells (less than 5%) in the peripheral blood, has the best
prognosis among the four subtypes. Four-year survival rates vary from 5.0% for acute,
5.7 % for lymphoma, 26.9% for chronic and 62.8% for smoldering 40. Although various
therapeutic strategies 44 have been tried, treatment of ATLL is still unsatisfactory.
ATLL is characterized by clonal expansion of mature T cells, each harboring a
single copy or multiple copies of HTLV-1 sequences 45. Initial polyclonal expansion of
infected cells is followed by a progression to oligoclonal and then to monoclonal

5
proliferation in vivo 46-48. The principal mode of viral replication after initial viral spread
in vivo is by mitosis of infected cells. The low incidence and long clinical latency of
ATLL strongly suggest that accumulation of genetic mutations are required for T cell
transformation in addition to HTLV-1 infection 49 . Although molecular pathogenesis of
ATLL is not fully understood, evidences suggests that HTLV-1 oncoproteins like Tax
play a critical role in transformation of infected lymphocytes by modulating T cell
activation and death pathways 50.
1.2.2 HTLV-1 Associated Myelopathy/Tropical Spastic Paraparesis (HAM/TSP)
In 1985, Gessain and colleagues 8 first reported that a group of patients in
French Martinique with a slowly progressive neurologic disorder, called Tropical
Spastic Paraparesis (TSP), had antibodies directed against HTLV-1. Subsequently,
another neurologic condition observed among HTLV-1-infected individuals in southern
Japan, termed HTLV-1-Associated Myelopathy (HAM) was reported by Osame et al 9.
Studies the established that TSP and HAM were clinically are identical diseases with
the common viral etiology of HTLV-1.
Compared to ATLL, HAM/TSP has a relatively shorter latency period ranging
from months to decades 51. HTLV-1 seropositive women are more likely to develop
disease than men 52. HAM/TSP is a chronic progressive demyelinating disease
predominantly affecting the thoracic spinal cord 28,53-56. Symptoms of HAM/TSP are
related to myelopathy including low back pain, weakness and spasm of lower
extremities, and dysfunction of the urinary bladder 57. Occasionally, a cerebellar

6
syndrome with ataxia and intention tremor is observed 58. Also, a predominant
dysfunction of descending sympathetic pathway has been reported in some cases 59.
Severe cellular destruction and inflammation both in the spinal cord and brain
characterized by multiple foci of severe demyelination and mononuclear cell infiltrates
with perivascular cuffing, parenchymal invasion and gliosis within white matter are
histopathological features of HAM/TSP 60, 61.
Risk of development and progression to HAM/TSP is increased when HTLV-1
proviral copy number is high in infected individuals’ blood leukocytes 62-66, and is
clearly associated with transfusion of HTLV-1-contaminated blood products 67,68. Other
factors influencing HAM/TSP development are host genetic factors 69, host immune
response, and perhaps viral variation especially in tax 63,70-74. Although the mechanisms
of HAM/TSP development by HTLV-1 infection are not yet clear, important role for
proinflammatory cytokines and activated lymphocytes in the development of the disease
has been suggested 61. High levels of inflammatory cytokines, such as IFN-γ, TNF-α,
IL-1 and IL-6 75-78, and large numbers of activated lymphocytes are present in the
cerebrospinal fluid (CSF) of affected individuals. Several hypotheses of HAM/TSP
pathogenesis such as CD8+ T cells mediate CNS damage 79, or autoimmune response
mediated HTLV-1- associated CNS damage 80,81 are suggested 45.
1.2.3 Other Diseases Associated with HTLV-1-Infection
A variety of autoimmune disorders and chronic inflammatory conditions are
associated with HTLV-1 infection, even though linkage between these diseases and

7
HTLV-1 infection is not as strong as those for ATLL and HAM/TSP. These include
idiopathic uveitis 82, arthropathy 83, Sjögren’s syndrome 84, infective dermatitis 85,
polymyositis 86, lymphadenitis 87, chronic respiratory disease 88, and acute myeloid
leukemia 89. Also, HTLV-1 infection is associated with severe Strongyloides stercoralis
infection 90,91. However, the role of HTLV-1 infection in the pathogenesis of these
disorders is still controversial.
1.3 HTLV-1 Virus Structure and Genome Organization
HTLV-1 is classified as the genus Deltaretrovirus, along with bovine leukemia
virus (BLV) and simian T lymphotropic virus (STLV). The mature HTLV-1 virion is
spherical and has a diameter of 110 to 140 nm with type C retroviral morphology 92.
The outer envelope is composed of host cell-derived cell membrane that contains the
viral encoded envelope glycoprotein spikes. The core of the virion consists of a highly
dense, spherical ribonucleoprotein complex of two copies of the 9 kb genomic RNA and
the host cell origin, primer tRNA-Pro, with virus encoded enzymatic and structural
proteins including reverse transcriptase and integrase, protease, nucleocapsid, capsid
and matrix 93.
The HTLV-1 genome contains elements common to other retroviruses, as well
as genes unique to HTLV-1. The proviral DNA genome is 9032 nucleotides long 94 is
flanked by long terminal repeat (LTR), which is a hallmark of retroviral genomic
structure, at each ends. Each LTR, composed of a U3 (unique 3'), R (repeated), and U5
(unique 5') region , contains sequences essential for viral reverse transcription,

8
integration, transcription, polyadenylation, splicing and viral message transport (Fig.
1.1) 95-97. A unique feature of the HTLV-1 LTR is the presence of three imperfect
tandem 21-base-pair repeats in its U3 region, collectively called Tax-responsive
elements-1 (TRE-1) upstream of the transcription start site 98. These three cis acting
regulatory elements are required for Tax-mediated trans-activation 99-104. In the middle
of the genome, gag , pol, and env genes are located, which are common to all known
retroviruses. The structural Gag precursor (later cleaved into matrix [MA], capsid [CA],
and nucleocapsid [NC] proteins) and enzymatic proteins (protease [PR], reverse
transcriptase [RT] and integrase [IN]) are translated from an unspliced RNA, and the
envelope protein (transmembrane [TM] and surface glycoprotein [SU]) is translated
from a singly spliced RNA 105.
In the pX region, which is located between env and 3’ LTR, HTLV-1 genome
contains unique genes encoded for regulatory and accessory proteins. These proteins are
generated by alternative splicing and internal initiation from the four open reading
frames (ORF). Regulatory proteins Tax and Rex, which are essential for the viral life
cycle, encoded by ORF IV and ORF III, respectively 106,107. ORF I and ORF II encode
four accessory proteins, p12I, p27I, p30II and p13II 95-97,108 109(Fig. 1.1). More recently, a
novel HTLV-1 protein, HBZ which is encoded by complementary (minus) RNA was
identified 110.

9
1.4 Structural and Enzymatic Proteins of HTLV-1
After translation into a single precursor polyprotein p55 (Gag), HTLV-1 Gag
protein is targeted to the inner side of lipid plasma membrane through post-translational
myristylation at its N-terminal end 111 and cleaved into matrix (MA, 19 kDa), capsid
(CA, 24 kDa) and nucelocapsid (NC, 15 kDa) proteins by viral protease(Fig. 1.1) 105,112.
Functionally, three types of domains, the M (membrane binding), I (interaction between
Gag proteins), and L (late budding) domains, have been identified in the Gag sequences
of different retroviruses 113,114. The p19 MA protein contains L domain (PPPY motif),
which is critical in budding of HTLV-1 virus particles 115. This budding process
requires interaction between L domain and cellular proteins Nedd4.1 and Tsg101 116.
CA proteins interact with each other and form the shell of an inner core structure.
Negatively charged NC proteins interact with viral RNA genome within the encased
capsid structure 45.
HTLV-1 protease (PR), which is generated by ribosomal frame shifting near 3'
end of gag and 5' of pol, is self-cleaved to become the active form 117. The catalytic
activities of PR are required for whole HTLV-1 life cycle, because mature or active
forms of viral proteins are generated by PR 118. Reverse transcriptase (RT) and integrase
(IN), which are produced by cleavage of the Gag/Pol precursor by the PR 118, have
enzymatic activities; Mg2+-dependent reverse transcription and proviral DNA
integration respectively. RT also has RNase H activity which specifically degrades the
RNA in the RNA-DNA duplexes during the reverse transcription process 93,118.

10
HTLV-1 envelope (ENV) protein is synthesized as a 61 to 68 kDa glycoprotein
in the endoplasmic reticulum (ER) and subsequently transported to the Golgi apparatus
through the secretory pathway. In the Golgi apparatus precursor Env protein is cleaved
into the surface (SU, gp46) and the transmembrane (TM, gp21) protein 119. SU-TM
heterodimer is essential for receptor binding and the fusion events viral entry120.
Recently, an ubiquitous glucose transporter, GLUT-1 has been identified as a HTLV-1
receptor by Manel et al 121.
1.5 Replication Cycle of HTLV-1
Major events in the retroviral replication cycle include adsorption and entry,
reverse transcription, nuclear transport and integration, viral gene expression, and viral
protein synthesis, processing, and assembly. From entry to integration step, the process
accomplished by the viral structural proteins and several enzymatic proteins occur in the
virion without de novo viral gene expression 122. The process of viral gene expression
and assembly, which is a later stage of viral replication, depends on host cellular
transcription and protein synthesis machinery, as well as viral machinery.
Viral particle attachment and entry involve an interaction between envelope (SU) and
HTLV-1 receptors. Many cell surface molecules have been implicated as HTLV-1
receptors including adhesion molecules123, heat shock cognate proteins 124, lipids 125,
lipid rafts 126, heparan sulfate 127 and transferrin receptor 128. More recently, the first
definitive identification was reported by Manel et al 121, who identified GLUT-1, a
ubiquitous glucose transport protein, as a receptor for HTLV-1. Before identification,

11
they found that HTLV-1 candidate receptor is poorly expressed on resting T cells but is
up-regulated after T-cell activation 129,130. Expression of full-length HTLV envelope and
envelope driven peptide resulted in dramatic changes in glucose metabolism such as
reduced glucose uptake and consumption 121. Because of these findings, they suspected
GLUT-1 as a HTLV-1 receptor. Further characterization of GLUT-1 and SU interaction
is required for understanding of HTLV-1 pathogenesis, as well as for developing
strategies to prevent HTLV-1 early transmission 131.
After envelope mediated attachment and fusion, components of the virion such
as NC and CA enter into cytoplasm. In the cytoplasm, RT (RNA-dependent/DNA-
dependent polymerase) initiates the synthesis of double-stranded DNA (dsDNA) from
viral genomic RNA. Viral dsDNA associates with cellular and viral proteins to form a
pre-integration complex 132, which is then transported into the nucleus where the
integration of the proviral DNA into the host genome occur. HTLV-1 integration is
considered to take place randomly in the host genome by virally encoded IN. However,
retroviral selection of integration sites is influenced by properties of host genomic DNA
properties such as bendability or open structure, which are commonly observed in
chromatin region of actively transcribed genes 133.
After integration, HTLV-1 provirus behaves like host genomic DNA; therefore
it requires participation of cellular transcription, translation, and transport machinery,
as well as viral proteins especially Tax and Rex 134. Integrated provirus passively
spreads to daughter cells by host cell division, or actively produces progeny virions.
The HTLV-1 replication cycle is completed when virions are assembled and released by

12
budding. However, HTLV-1 infectious virions are not efficiently released from the
infected cells, therefore viral transfer by cell-to-cell contact is a more efficient way for
HTLV-1 transmission 20.
1.6 Regulatory Proteins: Tax and Rex
1.6.1 Tax
Tax (the transcriptional activator of pX region) regulatory proteins are unique to
deltaretroviruses and are important for productive viral replication and proliferation of
host cells. HTLV-1 Tax, a 40 kDa phosphoprotein, was identified as a trans-activator of
viral gene transcription 135-139. Tax ,which is translated from a doubly spliced
mRNA(Fig. 1.1) 135-139, mainly accumulates in the nuclear region of HTLV-1-infected
cells and shuttles into the cytoplasm using a nuclear export protein 140,141. HTLV-1 Tax,
which has a pleiotropic function, is associated with transcriptional activation or
repression of various viral or cellular genes and alteration of cell cycle and apoptosis in
infected cells 142. It activates transcription by recruiting or modifying the activity of
cellular transcription factors such as cyclic AMP-responsive element binding protein
(CREB), serum-responsive factor (SRF) and NF-κB 143-147. Tax mediated transcriptional
activity can be achieved by various mechanism. For example, Tax activates CREB and
SRF mediated transcription by stabilizing the complexes of the transcription machinery,
or it enhances NF-κB mediated transcription by destabilizing IκB indirectly 148.

13
HTLV-1 Tax activates expression of viral genes via the LTR. Three highly
conserved 21-bp repeat elements collectively referred to as Tax-response element 1
(TRE-1) 99-104 are located in the U3 region of 5’ LTR 101-104. Similar to cAMP-
responsive elements (CRE) of host genomic sequence, HTLV-1 TRE-1 elements are
binding sites for multiple cellular transcription factors including CREB 149,150, cAMP
response element modulator (CREM) 149, activating transcription factors (ATFs) 151,
Tax-responsive element binding proteins (TREB) 152, activator protein-1 (AP-1)
102,153,154, and activator protein-2 (AP-2) 155. Also, a second enhancer element called
Tax-responsive element 2 (TRE-2) located between the central and proximal TRE-1 is
also important for viral transcription 156,157. Ets family transcription factors (Ets-1, Ets-
2, Elf-1 and TIF-1) and c-Myb transcription factors bind to TRE-2 158-162. Tax binds
directly to GC-rich sequences flanking the TRE-1 elements 163-165, and interacts with
basic region of host cellular basic leucine zipper (bZIP) transcription factors resulting in
enhanced bZIP dimerization and DNA binding activity 166.
A bZIP transcription factor, CREB requires a protein kinase A (PKA)-
mediated phosphorylation to bind with the transcriptional cofactor CBP/p300 during the
cell activation signaling 167,168. By interacting with both CREB and CBP/p300, Tax
eliminates the requirement of PKA activation, CREB phosphorylation, and recruitment
of CBP/p300 to the transcriptional complex. Therefore, CREB-Tax-CBP/p300 complex
results in constitutive activation of this pathway in HTLV-1 infected cells 169. Histone
acetylation by coactivators CBP/ p300 has been shown to play a major role in activating
HTLV-1 transcription 170. Also, CBP/p300 associated factor (P/CAF) is required for
Tax- mediated constitutive transcriptional activation 171,172.

14
In addition to viral gene expression, Tax affects a variety of cellular genes. In
fact, a gene array study has demonstrated that Tax modulates expression of hundreds of
cellular genes173. These includes cytokine genes, IL-2 174, IL-6 175, IL-8 176, IL-2Rα 174,
IL-1 177, GM-CSF 178, TNFα 179 and TNFβ 180, genes of transcription factors c-myc 181,
c-fos 182, c-sis 183, erg-1 184, c-rel 185, and genes involved in apoptosis Bcl-xL 186,187 and
DNA repair (PCNA) 188. Tax modulates cellular gene expression via at least four
distinct cellular signaling pathways: CREB, NF-κB, AP-1, and SRF 146,154,184,189,190.
Activation of NF-κB signaling by Tax is thought be critical for ATLL
pathogenesis, because ATLL cells express numerous cytokines and their receptors,
which are known to be regulated by the NF-κB pathway190. Over-expression of IL-2Rα
191, which is one of key features of ATLL cells, is induced by Tax-mediated NF-κB
activation 192. However, constitutive NF-κB activity is observed within ATLL cells of
patients, even though expression of Tax is lacking in these cells 193. The mechanism
underlying this Tax-independent pathway of NF-B activation remains poorly
understood.
Activation of NF-κB by Tax occurs in a different manner both in nuclear and
cytoplasmic components. Tax binds to multiple NF-κB family proteins such as p50, p52,
p65, c-Rel 147,194 and lyt10 195. Within the nucleus, Tax has been shown to bind to the
p50 and p52 subunits of NF-κB 147,195. Because both NF-κB proteins and Tax bind to
CBP/p300 196-198, Tax may complex with NF-κB and CBP/p300 to enhance the stability
of transcription machinery. However, the predominant mode of Tax mediated NF-κB
activation occurs by interacting with cytoplasmic IκB proteins, the inhibitor of NF-κB

15
190. The NF-κB heterodimer is usually retained in the cytoplasm by interacting with IκB
proteins 199. When cell signal is activated, IκB proteins are phosphorylated by IκB
kinase (IKK) and subsequently ubiquitinated and degraded in the proteasome 190.
Interaction between Tax and IκBα destabilizes NF-κB and IκB complex and causes the
nuclear translocation of NF-κB 200-202. Moreover, Tax activates IKK which consists of
two catalytic subunits, IKKα and IKKβ and a regulatory IKKγ subunit (also known as
NEMO) 200,202. Tax interacts with IKKγ subunit and recruits MAP3Ks to this complex
190, and resulting in phosphorylation and degradation of IκB and subsequent NF-κB
activation 203.
HTLV-1 Tax was also demonstrated to inhibit expression of several cellular
genes, such as p18INK4c, p53 and Bax 204. Sequestration effect of Tax on p300/CBP is
thought be one mechanism of repression activity 205,206, because as a transcriptional co-
activator, p300/CBP is required for cellular gene expression by binding to multiple
transcription factors 207,208. For example, promoter of p18INK4c gene requires
CBP/p300 co-factor for activation of E-47 transcription factor 209.
In addition to the transcriptional deregulation, Tax binds to a number of protein
complexes regulating transformation, cell cycle and apoptosis. Proteins affecting cell
cycle include p16INK4a 210 and p15INK4b 211,212 that directly regulate the Rb protein
and the G1/S phase of the cell cycle. Tax also interacts with human mitotic checkpoint
protein MAD1 213. Tax can affect apoptosis by interacting with an inhibitor of cell death
protein, A20 214 which is induced by a variety of inflammatory stimuli. A variety of

16
small cytoplasmic GTPases including RhoA, Rac1 and Cdc42, which are components
of cytoskeletal regulation, are also associated with Tax 215.
Collectively, HTLV-1 Tax effects on a wide variety of cellular targets by
transactivating, repressing or interacting to promote cell proliferation and
leukemogenesis.
1.6.2 Rex
Unlike Tax, ORF III encoded protein, Rex, regulates viral gene expression post-
transcriptional. Rex is a 27 kDa RNA-binding protein that is essential for transport of
viral mRNAs. Rex facilitates transport of unspliced RNA (gag/pol/pro) and singly
spliced RNA (env) from nucleus to cytoplasm, while it may inhibit the splicing and
transport of doubly spliced RNAs that encode the regulatory and accessory proteins in
the pX region. Therefore, Rex regulates the balance between viral structural and
regulatory gene expression. HTLV-1 Rex is not required for cellular immortalization in
vitro, but it is required for viral spread and persistence in vivo106,216,217. Rex is also
reported to have a role in stabilizing unspliced transcripts in T cells 218.
Rex performs its function by interacting with a sequence called the Rex-
responsive element (RxRE) within the U3 and R regions of the 3' LTR 219. RxRE forms
a stable secondary RNA structure, consisting of four stem loops and a long stem
structure 219-224. The stem-loop structure is also critical for appropriate polyadenylation

17
of viral RNAs 225,226. Because RxRE is present in all mRNAs, i.e. unspliced, singly and
doubly spliced mRNAs, other cis-acting sequences may play a role in the determination
of Rex regulation of mRNA nuclear export. Inhibitory sequences within the viral RNA
have been identified, which are termed cis-acting repressive sequences (CRS) 227-229.
Rex has multiple domains, which are important for its function, including
RNA-binding domain, nuclear localization sequence (NLS), nuclear export sequence
(NES), and multimerization domain. A highly basic N-terminal RNA-binding domain is
located within aa 1−19 230,231. This domain also functions as an NLS and is necessary
for the transport of unspliced viral mRNAs to the cytoplasm 232,233. The NES sequence
(also known as activation domain) is located in the middle of the Rex protein. Rex
mediated RNA transport is reported to depend on CRM1/exportin pathway 234,235. The
NES is required for the shuttling of Rex between the nucleus and cytoplasm236 The
activity of Rex is affected through phosphorylation. Treatment of infected cells with
protein kinase C inhibitor resulted in accumulation of unspliced mRNA and decreased
gag protein synthesis 237.
In addition to the full length 27 kDa form of Rex, a truncated 21 kDa form of
Rex (p21Rex) has been detected in HTLV-1 infected cell lines (Fig. 1.1) 108,109,238.
Because p21Rex has a truncation in the N terminal NLS sequence, it inhibits the shuttling
function of the full-length form of Rex protein when over-expressed 239. However the
function of p21Rex in the HTLV-1 replication and pathogenesis is still unknown.

18
1.7 HTLV-1 Non-structural Proteins
In addition to the regulatory proteins Tax and Rex, which are encoded in ORF
III and IV respectively, the HTLV-1 pX region encodes four additional accessory
proteins, p12I, p27I, p13II, and p30II. These proteins are generated from alternatively
spliced mRNA encoded in ORF I and II (Fig. 1.1) 95,240-242. All of these spliced mRNA
species have a common first exon encoded from nucleotides (nt) 1-119 in the R region
of the viral 5’ LTR. Doubly spliced mRNAs encode second exons that start at either nt
4641 or 4658 and end at nt 4831. Various splice acceptor sites in the pX region
correspond to the third exon start point for doubly spliced messages or the second exon
start site for singly spliced mRNA. A splice acceptor at nt 6383 is used to generate the
pX ORF I proteins, p27I (doubly spliced) and p121 (singly spliced). Similar to pX ORF
I, pX ORF II proteins are also produced from two alternatively spliced mRNAs. A
splice acceptor site at nt 6478 is used for generating mRNA encoding the larger protein,
p30II (doubly spliced) and a site at nt 6875 creating mRNA used for the smaller protein
p13II. A splice acceptor site at nt 6950 is used to produce doubly spliced pX-tax/rex
mRNA that encodes Tax or Rex using ORF IV or ORF III 95-97,108, 109.
Although HTLV-1 accessory proteins were thought not to be required for viral
replication 243, recent findings indicated that HTLV-1 accessory proteins play a critical
role in viral infectivity, maintenance of high viral loads, host cell activation, and
regulation of gene transcription 244-257. Even though the proteins were not directly

19
detected in HTLV-1 infected cells, indirect evidences indicate that pX ORFs I and II
mRNAs and proteins are expressed both in vitro and in vivo. mRNAs of pX ORF I and
II were detected by reverse transcription-PCR (RT-PCR) assays or semi-quantitative
RNase protection assays in infected cell lines and freshly isolated cells from HTLV-1-
infected subjects 242,258 . Furthermore, humoral antibody 254,259 and cytotoxic T cell 260
responses against recombinant proteins or peptides of the pX ORF I and II proteins are
detected in HTLV-1 infected patients, and asymptomatic carriers. These findings
strongly indicate that ORF I/II encoding proteins are produced in vivo and play roles in
HTLV-1 infection and leukemogenesis.
Chronically infected HTLV-1 cell lines were found to have 100 to 1,000-fold
less ORF I mRNA expressed compared to ORF III/IV mRNA species, and ORF II
mRNA amounts were 500 to 2,500-fold lower than ORF III/IV mRNA, suggesting that
HTLV-1 proteins expression are differentially regulated by unknown mechanism 261.
However, these data were collected from chronically infected cell lines. Therefore, a
requirement of ORF I and ORF II encoded proteins in HTLV-1 pathogenesis can not be
ruled out, especially in the early phases of infection in vivo. Furthermore, the nucleotide
sequence and genomic region encoding these accessory proteins, particularly p12I, are
highly conserved among different HTLV-1 strains, HTLV-2 and the highly related
simian T lymphotropic virus type 1 (STLV-1) 240,262-264.
In addition to these regulatory and accessory proteins, a novel viral protein
HTLV-1 bZIP factor (HBZ), encoded from antisense RNA was recently identified 110.

20
1.7.1 Minus Strand Encoded HBZ
Several ORFs encoded from the complementary (minus) strand of HTLV-1
RNA genome were reported initially by Larocca et al 265. Through RNA blotting, 2.5
and 2.9 kb minus strand RNAs were detected in HTLV-1 infected T-cells, but not in an
uninfected control. A novel viral protein, HBZ encoded by these complementary strands
of the HTLV-1 RNA genome was identified recently 110. HBZ is a nuclear localizing
protein composed of 209 amino acids that contains an N-terminal transcriptional
activation domain and a leucine zipper motif in its C terminus 110, as well as three
nuclear localizing signals 266.
By interacting with bZIP transcription factor, CREB-2, HBZ abolishs the ability
of CREB-2 to activate Tax mediated viral transcription, when it exogeneously
overexpressed. As a result, HBZ was suggested to repress Tax-mediated viral
transcription from the HTLV-1 LTR 110.
Moreover, HBZ interacts with the activator protein-1 (AP-1) transcription
factors c-Jun , JunB and JunD 267, 268. While the interaction with c-Jun results in down
regulation of AP-1 transcriptional activity, transcription function of JunB and JunD was
enhanced 267, 268. HBZ suppresses c-Jun- mediated AP-1 transcription by preventing c-
Jun DNA-binding 267 and promoting c-Jun degradation through a proteasome-dependent
pathway 269.
Rescently Arnold et al 270 evalutated the role of HBZ in vitro cellular
immortalization, and in vivo viral infectivity and persistence by generating HBZ

21
deletion mutatants. Mutation of HBZ did not affect in vitro viral replication and cellular
immortalization. However, rabbits inoculated with HBZ mutant cells displayed a
decreased antibody response and reduced infectivity when compared to wild type
HTLV-1 infected rabbits. These findings indicated that role of HBZ was not related
with repressive effect on Tax and AP-1 - mediated in vitro immortalization, but was
related with in vivo viral infectivity and persistence.
The role of HBZ in the pathogenesis of HTLV-1 infection is unclear, but
studies to date susggest that HBZ may contribute to the dysregulation of viral or cellular
gene expession to alter viral replication and perhaps disease.
1.7.2 pX ORF II Proteins: p13II and p30II
A 241 amino acid protein, p30II is generated from the doubly spliced mRNA
pX-tax-orf II 95,96 while p13II is translated from the singly spliced message pX-orf II 95-
97. However, an internal start codon in p30II message can be used to produce the smaller
p13II protein, which represents the C-terminal 87 amino acids of p30II.
Earlier studies suggested that ORF II encoding proteins were dispensable for
viral replication or immortalization of primary cells, because a viral strain isolated from
leukemic cells was reported to contain a premature stop condon in pX ORF II 271.
However, this study did not consider the possible role of ORF II encoded proteins in the
early viral infection in vivo. Furthermore, rabbits inoculated with a HTLV-1 proviral
clone with selective mutations in ORF II (ACH.p30/p13) failed to develop productive
infections 248.

22
1.7.2.1 p13II – Role in Viral Replication and Cell Survival
HTLV-1 p13II localizes to both the nucleus 242 and to mitochondria 256,272.
Because p13II has no DNA binding motifs nor transcriptional activity 273, a role of p13II
in mitochondrial function has been emphasized. p13II localizes to inner mitochondrial
membranes 272 using an atypical mitochondrial targeting sequence (MTS) located in the
N terminus (amino acids 22 and 31) 256. Unique features of p13II MTS include: its
short length, lack of positive charge and its unusual property of remaining intact upon
mitochondria import 256,274.
Accumulation of p13II in mitochondria results in disruption of the mitochondrial
inner membrane potential (∆ψ) with altered conductance to Ca2+ and K+. It also induces
mitochondrial swelling and fragmentation, suggesting a possible role of p13II in
apoptosis 256. Although p13II causes these alterations in mitochondrial morphology, the
protein does not change the permeability transition pore (PTP) driven by cation fluxes
or release of cytochrome c, which are two key events in some apoptosis pathway 256.
The potential role of p13II in cell population was implicated by Silic-Benussi et
al. 275. Expression of p13II resulted in the growth suppression in both HeLa and Jurkat
T cells in vitro, and reduced tumorgenicity of Ras and Myc co-transfected rat
embryonal fibroblasts (REFs). Hiraragi et al. 276 recently found that Jurkat T cells
expressing p13II were more sensitive to apoptosis when treated with apoptosis inducing
agents, such as ceramide and Fas ligand (FasL). Furthermore this apoptosis was
inhibited by a farnesyl transferase inhibitor (FTI) treatment, which blocks the
posttranslational modification of Ras to its active form, suggesting a role for p13II in

23
Ras-mediated cell signaling. These data indicate that exogenously expressed p13II
affects HTLV-1-induced lymphocyte proliferation by altering the balance in Ras-
mediated lymphocyte survival and death.
Cellular proteins that are physically associated with p13II have been identified
by yeast two-hybrid screening. p13II associated with the products of two cDNA clones,
C44 and C254, encoding nucleoside monophosphate kinase superfamily and actin-
binding protein 280 (ABP280), respectively 277. The protein product of C44 has
structural similarities to archeal adenylate kinases that is an eukaryotic mitochondrial
protein involved in energy metabolism. Interestingly, this protein is expressed in Jurkat
T cells and proliferating PBMC, but not quiescent PBMC 277. ABP280 is part of the
cytoskeleton and functions in the insertion of adhesion molecules into the cell
membrane 277. Furthermore, similar to an accessory protein of BLV, G4 that localizes to
mitochondria, p13II binds to farnesyl pyrophosphate synthetase (FPPS) 278. FPPS, which
is involved in the mevalonate/squalene pathway to synthesize of FPP, is a substrate
required for prenylation of Ras oncoprotein 279. Interestingly, Hiraragi et al. 276 found
recently that p13II mediated apoptosis sensitization was dependent on Ras mediated
signal pathway.
Although the biological significance of this viral accessory protein in the
pathogenesis of HTLV-1 infection remains still unclear, recent in vivo rabbit inoculation
with selectively p13II expression abrogated ACH cell line (729.ACH.p13) resulted in
significant reduced HTLV-1 infection, indicating the critical requirement of p13II for
establishment of HTLV-1 infection in vivo 280 .

24
1.7.2.2 p30II - A Selective Repressor of Transcription
HTLV-1 p30II mainly localizes to nucleus, specifically the nucleolus through
highly conserved bipartite nuclear localization signals (NLS) 241,281. It also contains
serine/threonine-rich regions that share distant homology to the activation domain of
transcription factors of the POU family, such as POU-2, Oct-1/2 and Pit-1 96,241. In
addition, p30II co-localizes and physically interacts with the cellular transcriptional co-
adaptor p300 in the nucleus 247. Taken together, these findings suggest that p30II has a
role in regulation of viral and cellular gene expression.
Recent studies indicate a role for p30II as a transcriptional regulator or a
negative regulator of mRNA expression. As a transcriptional regulator, p30II alters the
basal level of CRE and TRE - mediated transcription 247,250. Zhang et al.250 reported
that low concentrations of p30II stimulated HTLV-1 TRE-driven reporter gene activity,
whereas higher concentrations repressed LTR (TRE) and CRE-driven reporter gene
activity. Furthermore, p30II activated transcription through its central core region
(amino acid 62 and 220) 250. In a subsequent study, p30II - mediated transcriptional
activity was enhanced by CBP/p300, a critical co-adaptor of cell transcription, via
binding highly conserved KIX region of CBP/p300 247. Through DNA binding assays,
p30II inhibited the CREB-Tax-CBP/p300 complexes on TRE oligonucleotides 247
suggesting that p30II differentially regulates viral transcription via the sequestration of
CBP/p300.

25
Nicot et al. 282 demonstrated that expression of p30II results in selective nuclear
retention of spliced Tax/Rex mRNA resulting in decreased viral gene expression 282.
The mechanism by which p30II regulates the Tax/Rex mRNA retention in the nucleus is
still unclear. Because p30II does not contain a conserved RNA-binding motif, it may
bind directly via unknown motif or indirectly via ribonuceloprotein complexes which
are important for viral nuclear export. Recent studies by Younis et al. 283 support these
findings by showing that p28II, a homologue of p30II, regulates HTLV-2 gene
expression by a similar posttranscriptional manner. This same group reported that p30II
and p28II actually travel with transcription compex using chromatin
immunoprecipitation assay 284.
Microarray gene expression analyses of Jurkat T cells stably expressing p30II
indicated that p30II selectively represses many cellular genes involed in T-cell
activation, adhesion and apoptosis 285. Although the precise mechanism that p30II
affects gene regulation is still unclear, the critical role of p30II in HTLV-1 infection is
implicated from rabbit infection studies 286. Inoculation of the ACH.30 cell line, which
produces a selectively truncated of p30II, resulted in significantly reduced HTLV-1
proviral load in rabbits. Interestingly, sequencing results from ACH.30-inoculated
rabbits revealed a reversion to wild-type sequence, suggesting HTLV-1 requires p30II to
survive in vivo 286 .

26
1.7.3 pX ORF-I Protein: p12I
HTLV-1 p12I can be translated from singly spliced or doubly spliced ORF I
mRNA via initiation at an internal methionine codon. Because full-length p27I cDNA
expression plasmid produced only p12I protein, p27I mRNA was thought to be
preferentially used over the p12I message 242. However, p27I can be generated using in
vitro transcription-translation systems 96 and CTL responses against p27I specific
peptides were demonstrated in asymptomatic and disease subjects 260.
1.7.3.1 Structure of p12I
p12I is a small 99 amino acids hydrophobic protein with a high percent of
leucine (31%) and proline (17%) residues 242. The amino acid sequence of p12I is
highly conserved among viral samples of HTLV-1-infected individuals 47,287 Further
analyses of p12I suggested potential secondary α helix structures at amino acids 12-32
and amino acids 48-67 that function as transmembrane regions, which overlap putative
leucine zipper motifs (Fig. 1.2) 97,288. These distinct secondary structures are thought to
be involved in protein membrane localization or homo-oligomerization of the protein.
Indeed, p12I was demonstrated to form dimers by Trovato et al 288. Furthermore, p12I
contains four proline-rich (PXXP) Src homology 3 (SH3)-binding domains (Fig. 1.2) 47,
a motif implicated in cell signalling, suggesting that p12I plays a role in modulating cell
signal transduction. Among the four PXXP motifs, the first (at amino acids 8-11) and
the third (at amino acids 70-74) motifs are highly conserved among different HTLV-1

27
strains. Interestingly, these PXXP motifs are often preceded by an arginine residue at +2
position 289. In addition, p12I has one dileucine motif (DXXXLL) at amino acid 26-31
(Fig. 1.2), which is known as a sorting motif in HIV Nef 290. Although, its functional
role in p12I has not been determined, the dileucine motif in Nef is required for down
regulation of cell surface molecules, such as CD4 and MHC class I through endocytosis
and trafficking to the ER-Golgi compartments by associating with adapter protein 1
(AP-1), AP-2 and AP-3 290. Furthermore, in studies reported in Chapter 2 in this thesis
we have identified a conserved PSLP(I/L)T sequence in p12I (Fig. 1.2), which has
homology to the calcineurin-binding PxIxIT motif of nuclear factor of activated T cells
(NFAT) 244. In Chapter 2 of this thesis, the role of this motif in modulation of NFAT
activation is addressed.
Sequence analysis of p12I reveals potential post-translational modifications
sites such as ubiquitylation, glycosylation, and phosphorylation. Trovato et al. 288 have
reported that p12I is ubiquitylated on lysine residue at amino acid position 88 (Fig. 1.2).
Substitution of this lysine with an arginine resulted in increased stability of the protein.
Interestingly, they reported that natural alleles of HAM/TSP patients exclusively
contain a lysine residue at amino acid 88, whereas arginine is found in HTLV-1 strains
isolated from most ATLL patients and asymptomatic carriers, suggesting ubiquitylation
of p12I may result in different disease outcomes. However, an analysis by Martins et
al.291 indicted that lysine residue at this position was neither related to disease outcome,
nor is able to be used as a marker of progression to HAM/TSP, because only one out of
37 HAM/TSP patients carried the lysine residue and also one of 40 asymptomatic
HTLV-1 carriers had this rare phenotype. Additionally, HTLV-1 p12I has a potential N-

28
linked glycosylation site at amino acid 51 (asparagine) and multiple potential O-linked
glycosylation sites (serine and threonine). However, a deglycosylation study revealed
that p12I is not a glycoprotein 292. Through sequence analysis of p12I, we have found
several potential phophorylation sites such as protein kinase C (LTMR) site at amino
acid 75. However, p12I does not appear to be phosphorylated as demonstrated by a
phosphate metabolic labeling assay in 293T cells transiently expressing p12I
(unpublished data).
1.7.3.2 p12I Subcellular Localization and Protein Interaction
HTLV-1 p12I localizes in cellular endomembranes 241, predominantly in the ER
and cis-Golgi apparatus which is evidenced by immunofluorescent confocal microscopy,
electron microscopy and subcellular fractionation studies in 293T and Hela-Tat cells 292.
p12I was retained in the ER and cis-Golgi after blocking de novo protein synthesis or
disrupting ER-to-Golgi protein transport 292, suggesting p12I is neither transported to the
cell membrane nor secreted extracellularly. Thus, p12I localization in the ER is required
for its calcium-mediated NFAT activation 293.
Interestingly, two ER localizing proteins, calreticulin and calnexin, appear to
directly bind with exogenously expressed p12I 292. These two proteins are involved in
multiple cellular functions including calcium homeostasis, protein folding, integrin
mediated signaling, and function as molecular chaperones 294,295. The biological
significance of p12I interaction with these proteins in viral pathogenesis remains to be
elucidated.

29
The second putative trans-membrane domain of HTLV-1 p12I shares
approximately 50% amino-acid sequence homology with the bovine papilloma virus
(BPV) E5 protein and Epstein-Barr virus (EBV) LMP-1 protein 296,297. Similar to E5
protein of BPV, p12I binds to the 16 kD subunit of the vacuolar H+-ATPase (16K) 297,298.
When tested in a focus-formation assay, p12I itself did not induce transformation of
mouse C127 fibroblasts, however it enhanced the ability of E5 in inducing
transformation 297, suggesting a role of p12I in the transformation activity by enhancing
Tax oncogenic functions. However, a complex p12I and 16K does not correlate with the
transforming ability of p12I, while the E5 binding to 16 K appears to be important in the
E5-mediated transformation.
p12I was demonstrated to interact with the immature form of the interleukin-2
receptor β and γ chain in a transient over-expression system (Fig. 1.2) 299. Through the
central proline-rich region (amino acid 37-47), p12I interacts with these IL-2 receptors
288. Further studies demonstrated that the p12I-binding on the cytoplasmic domain of the
IL-2 receptor β chain was involved in the recruitment of Janus-associated kinases (Jak)
1 and Jak3 after IL-2 ligand binding 300. As a result of this interaction, the DNA binding
activity and transcriptional activity of signal transducers and activators of transcription-
5 (STAT5) were increased, suggesting p12I expression may decrease the threshold
required for T-cell activation. However, ACH.p12I cells, which are selectively p12I
expression abrogated, have no significant differences in IL-2 receptor chain (α, β, γc)
expression, in IL-2-mediated proliferation, or in IL-2-induced phosphorylated forms of
Stat3, Stat5, Jak1, or Jak3 when compared with wild type cell line ACH 301. These

30
controversial findings can be explained if p12I affects IL-2 receptor -mediated cell
signal during the early stages of HTLV-1 infection, not in the immortalized cell stage.
Furthermore, HTLV-1 p12I was demonstrated to associate with immature forms
of the major histocompatibility complex class I (MHC I) and inhibits interactions of
MHC I with β2-microglobulin, when these proteins were coexpressed in Hela-Tat cells,
which resulted in a decrease of surface expression of MHC I by directing the newly
synthesized MHC-I to the proteasome for degradation 302-304 These data suggest a role
of p12I in escape from immune surveillance. However, cell surface MHC I and MHC II
expression were not significantly different between PBMC immortalized by transfection
of wild type (ACH) and pX-ORF I ablated proviral clones (ACH.p12I) 301, indicating
that effect of p12I on MHC I expression is subtle in immortalized cells. Furthermore,
the possible function of p12I on escaping immune surveillance during the early stage of
viral infection is also unlikely, because p12I message abrogated ACH.p12I cells , which
were inoculated into rabbits, did not elicit strong immune response, while wild type
ACH cells did 253. Therefore, the early loss of viral infectivity of ACH.p12I cells is
unlikely due to immune mediated cell death nor p12I-mediated down regulation of
MHC I.
On work presented herein, we discovered that HTLV-1 p12I binds calcineurin
via a highly conserved PxIxIT binding motif 244 which is found in variety of calcineurin
binding proteins found in yeast, mammalian cells, and viruses 305. The biological
significance of this binding is still undetermined, but it may modulate p12I-mediated
calcium dependent signaling in T cells. In Chapter 2 of this thesis, I discussed the role
of this motif in modulating specifically NFAT activation in T cells.

31
1.7.3.3 Role of p12I in Viral Infectivity
Although pX ORF I of HTLV-1 was initially reported to be dispensable for viral
infectivity and primary lymphocyte transformation in vitro 243,306, studies from our
laboratory indicated that p12I is critically required for establish HTLV-1 infection in
vivo in a rabbit model and in primary T cells 251,253. Rabbits inoculated with ACH.p12I
failed to establish persistent infection, which was indicated by dramatic decreases in
humoral responses against HTLV-1 antigens, absence of viral antigen p19 production
from ex vivo PBMC culture, and transient detection of provirus by PCR 253. These
results suggested that p12I has a role in T cell activation, since most circulating
lymphocytes in vivo are quiescent or non-activated. Because initial studies used
activated PBMC, which were stimulated by IL-2 and phytohemagglutinin (PHA), as
target cells, the function of p12I as a T cell activator might be not required for
productive HTLV-1 infectivity in typical cell culture assays.
Our data supported a hypothesis that p12I is critical for T cell activation during
the early stages of viral infection 251. When ACH.p12I cells were cocultured with naive,
quiescent PBMC in the absence of exogenous stimuli, which more accurately reflect the
virus-cell interactions in vivo, a dramatic reduction in the viral infectivity resulted.
Furthermore, when T cell stimulators were added to the coculture, ACH.p12I cells were
restored in their ability to infect quiescent target T cells 251. These data indicated that
HTLV-1 p12I is required for efficient HTLV-1 infection in quiescent PBMC and
suggest a role for p12I in target T cell activation. The function of p12I is similar to that

32
of HIV-1 Nef, which is required for viral infectivity in quiescent T lymphocytes 307-309.
Interestingly, p12I complemented Nef function for efficient HIV-1 infection of
macrophages 310.
1.7.3.4 Role of p12I in Calcium- Mediated T Cell Signaling.
As implicated from in vivo and in vitro infectivity assays to test p12I function
and its structural properties, the viral protein role in T cell activation has become more
clear 245 . p12I was demonstrated to activate a major T cell transcription factor, NFAT,
in Jurkat T cells, without alteration of AP-1 or NFκB-mediated transcription 245. This
specific NFAT activation was dependent upon the Ras/MAP kinase pathway stimulated
by the phorbol ester, PMA. Further inhibition assays were performed to determine how
p12I activates NFAT. Inhibition of phospholipase C- γ (PLC-γ) and LAT (linker for
activation of T cells), which are upstream of calcium-mediated pathway, did not affect
NFAT activity, whereas inhibition of calcium-dependent signals by calcineurin inhibitor
(cyclosporin A), intracellular calcium chelator (BAPTA-AM), and a dominant negative
mutant of NFAT, abolished NFAT-dependent transcription, indicating direct p12I
function on calcium homeostasis in ER. Furthermore, the functional substitution of p12I
with thapsigargin, which released intracellular calcium from ER stores, supports the
hypothesis that p12I activates NFAT in a calcium-dependent manner (Fig. 1.3).
We have directly tested this hypothesis by measuring cytoplasmic calcium
concentration in Jurkat T cells 246. Expression of HTLV-1 p12I increases the basal
cytoplasmic calcium concentration and concurrently diminishes calcium available from

33
ER stores. Inhibition assays using inhibitors of inositol 1,4,5-triphosphate receptor
(IP3R) or calcium release-activated calcium channels (CRAC) suggested that p12I-
mediated NFAT activation occurs through on IP3R mediated the calcium release from
ER stores and subsequent by extracellular calcium entry via CRAC. Moreover, we
found that p12I stable expression enhanced the production of interleukin-2 (IL-2) which
is a downstream gene of calcium-NFAT mediated pathway, in Jurkat T cells and
primary lymphocytes Fig. 1.3) 293.
In Chapter 2 of this thesis, I provide evidence that p12I can regulate NFAT
activity either positively via elevating cytosolic calcium from ER stores or negatively
by calcineurin binding through PxIxIT motif 244. Although the significance of this
opposed regulatory functions of p12I is still unclear, there are several similarities
between p12I and cellular proteins such as CAML (Ca2+-modulating cyclophilin ligand)
and Bcl-2. CAML which contains two putative transmembrane domains localizes in the
ER and induces calcium release from the ER and leads to NFAT activation like HTLV-
1 p12I. Also CAML indirectly binds with calcineurin through cyclophilin binding. An
anti-apoptotic protein, Bcl-2 also has similar functional properties with p12I: calcium
release from the ER and calcineurin binding 311,312. Bcl-2 localizes to the ER and
mitochondria and maintains calcium homeostasis, suggesting its functions as an ion
channel protein 313 On the other hand, Bcl-2 binds calcineurin via its BH4 domain, and
inhibits NFAT activity by sequestering calcineurin from NFAT binding without
affecting calcineurin catalytic activity 314,315. Therefore, p12I may affect apoptosis in
HTLV-1 infected T cells similar to Bcl-2.

34
Furthermore, in a subsequent study, our laboratory performed gene array
analysis in stably p12I expressing Jurkat T cells to test if p12I regulates the expression
of cellular genes in a calcium-dependent manner 316. Alteration of genes by p12I
involved cell proliferation, apoptosis, cell adhesion, immune response modulation and T
cell signaling predominantly in a calcium-dependent manner. Interestingly, p12I
expression induced increased expression of p300, which is a key cellular transcriptional
co-adaptor 316 and a co-activator for HTLV-1 LTR transcription 172. p300/CBP functions
as a co-adaptor of transcription at the HTLV-1 promoter and plays a critical role in the
regulation of Tax-dependent HTLV-1 transcription in infected T-cells 170. HTLV-1 Tax
trans-activates HTLV-1 and cellular gene transcription through its interaction with the
p300 and CBP co-activators 317. Therefore, p12I may enhance Tax-mediated HTLV-1
viral gene expression and infection by increasing p300 expression. In addition, p12I
may modulate clonal expansion, cell survival and transformation of HTLV-1 infected
cells through p300 and Tax mediated transactivation.
Calcium release from the ER by p12I and subsequent activation of calcium
mediated cell signal transduction including stimulation of NFAT 246, increased IL-2
production 293 and increased p300 expression316, might be critical during the early
stages of HTLV-1 infection. For HIV-1 active replication, activation of NFAT is
required in primary T cells. NFAT activation is sufficient to overcome a blockade at
reverse transcription and induces highly permissive state for HIV-1 replication 318.
Therefore, p12I may induce NFAT activation to induce a permissive state for active
HTLV-1 early replication. Moreover, by increasing production of cytokine IL-2, p12I

35
activates T lymphocytes division, which is a prerequisite for the retroviral provirus to
integrate into the host cell genome and permit the subsequent viral replication and
productive infection. p12I expression in HTLV-1-infected T lymphocytes may lower the
calcium threshold in order to respond to weak stimuli, which would normally not
activate T cells. These stimuli may come from direct contact via T cell receptor or cell
surface receptors or soluble factors such as cytokines and chemokines 319.
Interestingly, many viral proteins modulates the cellular Ca2+ homeostasis
during virus replication or pathogenesis 320. Hepatitis B virus X protein (HBx) 321,
rotavirus NSP4 322, and protein K7 of kaposi's sarcoma-associated herpesvirus (KSHV)
323 increase cytoplasmic calcium for viral replication. Also, HIV-1 Nef, which plays a
role in viral pathognesis, modulates calcium homeostasis through IP3R mediated
pathway to activate NFAT 307. Therefore, modulation of calcium homeostasis by p12I
may play a role in HTLV-1 viral replication and pathogenesis.
In the Chapter 3 of this thesis, I tested the influence of p12I in another calcium -
calmodulin mediated T cell signal pathway. The expression of p12I in Jurkat T cells
induced cell surface clustering of LFA-1, which was inhibited by calcium chelator
BAPTA-AM, calcium channel blocker SK&F 96365, and calpeptin, an inhibitor of the
calcium-dependent protease calpain. These data indicate that HTLV-1 p12I induces
LFA-1 clustering through calcium dependent signaling and may influence cell-to-cell
transmission of virus.

36
1.8 HTLV-1 Cell-to-Cell Transmission
Unlike HIV-1, naturally HTLV-1 infected lymphocytes produce limited cell-free
viral particles, and these are produced only during in vitro conditions such as
continuous culture of transfected or producer cell lines 64,324. Moreover, HTLV-1 is
poorly infectious as cell-free virions 19: only one out of 105 to 106 particles is infectious
325. Cell-to-cell contact between virus infected and target cells is required for efficient
transmission 20,324,326.
The mechanism of cell-to-cell spread of HTLV-1 is still not completely
understood. However, the interaction between HTLV-1 envelope proteins and its
receptor or between cellular adhesion molecules are thought to be required, as cell
fusion or syncytium formation is induced by cell surface expression of envelope protein
327-329, or cellular ICAM-1, ICAM-3, and VCAM adhesion molecules123,330.
A recent confocal observation by Igakura et al 331 indicated that the structure of
spontaneously formed cell conjugate between HTLV-1 infected cell and target
lymphocytes resembles an immunologic synapse, which is a specialized contact
structure made between a lymphocyte and another cell, such as an antigen-presenting
cell. They found that polarization of the adhesion molecule talin and the microtubule
organizing centre (MTOC) to the cell contact areas with accumulation of core protein
Gag and the viral genome at the cell-to-cell junction, followed by transfer of both the
Gag protein and the HTLV-1 genome to the uninfected cell. They termed this structure
the “virological synapse” and highlighted on important difference between the

37
virological and the immunological synapse: the MTOC is oriented toward the synapse
in an immunological synapse, but the MTOC is polarized inside the HTLV-1–infected
cell in a virological synapse. Therefore, they suggested that these events were not
caused by T cell receptor (TCR)-mediated recognition of HTLV-1 antigens, but
triggered by HTLV-1 infection and cell-to-cell contact 64.
In a subsequent study, Barnard et al 332 investigated T-cell surface molecules
involved in triggering the polarization of MTOC in the HTLV-1–associated virological
synapse. They carried out antibody-coated bead-cell conjugate formation assays using
antibodies against T-cell surface molecules such as CD2 and CD3, LFA-1 (CD11a and
CD18), CD28, ICAM-1 and CD25, which are involved in T cell activation or up-
regulated in HTLV-1 infected cells. They found that engagement of ICAM-1 and LFA-
1 molecules caused MTOC polarization in HTLV-1–infected cells. Moreover, they
exhibited that that ICAM-1 was more highly associated with MTOC polarization than
LFA-1 molecules by performing inhibitory assay using soluble inhibitory cyclic
peptides or LFA1 mutated cell lines. These authors strongly suggested that the ICAM-
1/LFA-1 interaction plays an important role in polarization of MTOC and formation of
virological synapse. The increased expression of ICAM-1 in HTLV-1 infection 333 may
facilitate more HTLV-1 viral cell-to-cell spread. Interestingly, in HIV-1 viral
transmission or syncytium formation, ICAM-1/LFA-1 interaction is important 334-336.
In addition to cellular factors, Nejmeddine et al 337 tested if HTLV-1 viral
protein Tax affected MTOC polarization and virological synapse formation. They found
that Tax protein is present in cis-Golgi compartment around the MTOC and in the cell-

38
to-cell contact area, as well as in the nucleus. MTOC and Tax were always polarized in
the same direction and this MTOC/Tax reorientation was dependent on the function of
the Rac family and Cdc42 small GTPase, which are important in cell movement and
polarization 338. Expression of Tax sufficiently induced polarization of the MTOC
toward the target cell, suggesting that Tax promotes cell-to-cell spread of HTLV-1
through virological synapse.
In Chapter 3 of this thesis, I have tested if HTLV-1 nonstructural protein p12I
affects LFA-1 mediated T cell adhesion believed to be important in MTOC
reorientation and virological synapse formation and in facilitating HTLV-1 cell-to-cell
spread. Interaction between LFA-1 and its ligand ICAM-1 is required in the formation
of the immunologic synapse 339 and plays role in reducing the threshold for T-cell
activation 340. Moreover, signal transduction through LFA-1 (outside in signal) which
regulates cytoskeletal rearrangement and cell movement 341, may play a critical role in
MTOC polarization and virological synapse formation. Therefore, HTLV-1 p12I may
enhance HTLV-1 cell-to-cell transfer via promoting LFA-1 mediated adhesion and
virological synapse formation.
1.9 Regulation of Integrin Activity
Integrins are heterodimeric adhesion receptors composed of α and β subunits. T
cells possess at least 12 of the 24 known integrin heterodimers, the pattern of expression
is changed depending on maturation status or subset of T cells. The β1 subunit integrins
(α1-6β), which are commonly found in many cell types, bind extracellular matrix. There

39
are four kinds of leukocyte-specific ß2 integrins αLβ2, αMβ2, αXβ2 and αDβ2) on T
cells, αLβ2 (leukocyte function-associated antigen-1; LFA-1) is most the abundant T
cell integrin 341-343.
Integrins are required for all of T cell function, such as migration to peripheral
lymph nodes and inflammatory sites, and formation of the immunological synapse in
antigen presentation and cytotoxicity. Because the ligands for integrins are widely
expressed in tissue and cells, the activity of integrins are strictly regulated. Integrins are
adhesive to their ligands only when cells were exposed to stimuli such as engagement of
T cell receptors or cytokines and chemokines stimulation. This process termed ‘inside-
out signaling’ starts from intracellular signals that trigger rapid integrin activation. In
addition, ligand binding to the integrin itself transfers intracellular signal transduction
(outside–in signaling) to regulate cytoskeletal polarization and T cell movement and
migration 341,343.
Inside-out signaling causes both clustering of integrins on the cell surface, or an
increase in the affinity for ligand through conformational change. Experimentally
addition of the divalent cations Mn2+ or Mg2+ with EGTA or specific antibodies that
bind to the extracellular portion of LFA-1 cause conformational and affinity alteration
344-346. However, identification of physiological affinity modulation is difficult, because
the increase of LFA-1 affinity is rapid and transient. Therefore, the mechanism of
inside out signaling to activate LFA-1 affinity is not well understood compared to that
of LFA-1 clustering 341. However recent studies exhibited that a few microliters of
chemokine could induce affinity increase, whereas lasting stimulation induced LFA-1
clustering 347. Moreover, talin, Rap1 and RapL proteins are reported to be required for

40
affinity regulation of LFA-1 343,348. The cytoskeletal protein talin is associated with the
integrin cytoplasmic domain, modulating affinity for β1, β2 and β3 integrins 349.
Feigelson et al. 350 showed that functional Src-family kinase p56 (Lck) is required for
maintaining high affinity status of integrin α4β1 on T cells, which leads to rapid binding
of circulating T cell to endothelium.
Release of integrins from the cytoskeletal network is thought to permit increased
lateral movement and clustering of integrins. From TCR to LFA-1 clusteirng, key
signaling proteins such as Vav-1 (guanine exchange factor), ADAP, SKAP-55 (adaptor
protein), Rap1 (small GTPase) and RAPL (Rap1 binding protein) were identified
through studies of transgenic and knockout mice models 341,351-354. For example, LFA-
1clustering is induced by activation of GTPase Rac, which is stimulated by signal
cascade from TCR activaiton, phosphorylation of SKAP-55, a subsequent binding with
ADAP and SLP-76, and complex formation with Vav-1, a GEF for GTPase Rac 349,355.
Furthermore, protein kinase C (PKC) activation by phorbol esters triggers integrin
clustering by interacting with β2 subunit and by phosphorylating residuces in its
cytoplasmic tail 354,356. Moreover, elevation of cytoplasmic calcium by TCR activation
or by treatment with calcium mobilizers such as ionomycin and thapsigargin 345 can
induce clustering of LFA-1. Phorbol esters and calcium mediated LFA-1 activation
also requires the calcium-dependent protease calpain, which is suggested to play a role
in releasing integrin from cytoskeleton 345. Interestingly, the function of HTLV-1 p12I
is similar to that of thapsigargin i.e. inducing calcium release from the ER stores to
activate T cells 246.

41
Signal transduction through activated LFA-1 (outside-in signaling) can cause T
cell attachment and lamellipodial movement at the leading edge by activating myosin
light chain kinase (MLCK), whereas it leads T cell detachment at trailing edge through
RhoA and Rho kinase (ROCK) mediated pathway 341,357,358. Interestingly, LFA-1
interacts with the microtubule network, which are required for HTLV-1 cell-to-cell
transmission 331. LFA-1 engagement triggers translocation of Pyk-2, PKCβI and PKCδ,
which are known to important in T cell migration, to the MTOC 359,360. Therefore,
signaling from activated LFA-1 may enhance translocation of HTLV-1 viral or cellular
proteins, which are required for HTLV-1 cell-to-cell spread, to the MTOC.
Furthermore, these reports suggest that LFA-1 activation by HTLV-1 p12I also may
promote cell-to-cell transmission through LFA-1 mediated microtubular system
activation (MTOC polarization).
1.10 Bystander Cell Activation by HTLV-1
One of immunological hallmarks of HTLV-1 infection is spontaneous
proliferation of PBMC, which are isolated from HAM/TSP patients and asymptomatic
carriers, in vitro without exogenous antigens or stimulants 361-364. The magnitude of
spontaneous proliferation is more pronounced in HAM/TSP patients than in
asymptomatic carriers361-364. It is suggested that spontaneous proliferation of PBMC is
due to Tax mediated up regulation of IL2- IL2 receptor autocrine loop, because Tax
protein, which is not detected in fresh ex vivo PBMC, but can be found after several
hours of culture in vitro 365,366. Moreover, Wucherpfennig et al 367 and Kimata et al 368

42
demonstrated that direct interaction between HTLV-1 infected cells and uninfected
resting T cells induced spontaneous proliferation.
Proliferation or stimulation of uninfected PBMC was induced by direct
interaction with irradiated or fixed HTLV-1 infected cells, but not by isolated virus
particles nor soluble factors produced from infected cells 367,368. This activation process
was blocked by monoclonal antibodies that inhibit interaction of CD2/LFA-3 (CD58),
LFA-1/ICAM-1 (CD54) and IL2/ IL-2 receptor (CD25). The strong inhibition was
found in treatment of antibodies against CD2 and CD58. However, antibodies against
class I or class II MHC complex or against various regions of the HTLV-1 envelope
proteins gp46 and gp21 did not block activation of uninfected cells. These data indicate
that envelope proteins of HTLV-1 viral particles are not mitogenic. Instead, target
uninfected cell activation by HTLV-1 infected cells requires cell-to-cell contact via
interaction of cell surface molecules especially CD2 and LFA-3 (CD58).
Our previous data suggest that the role of p12I is related with T cell activation
during the early HTLV-1 infection, and demonstrated that p12I activates LFA-1
clustering in HTLV-1 infected cells, therefore, interaction between LFA-1 and ICAM-1
may activate target cell T cell activation. I present data in Chapter 4 of this thesis to
address the role of p12I in activation of uninfected target cells.
Overall, data presented in this thesis provided further evidence that a highly
conserved viral accessory protein, p12I is critically required for early HTLV-1 infection.
I demonstrated in this thesis that p12I regulates NFAT activation by interacting with

43
calcineurin, enhances cell-to-cell adhesion by inducing LFA-1 clustering and facilitates
early viral infectivity via regulating viral gene expression during early cell-to-cell
transmission.
1.11 REFERENCES
(1) Poiesz BJ, Ruscetti FW, Gazdar AF et al. Detection and isolation of type C retrovirus particles from fresh and cultured lymphocytes of a patient with cutaneous T-cell lymphoma. Proc Natl Acad Sci USA. 1980;77:7415-7419.
(2) Hinuma Y, Nagata K, Hanaoka M et al. Adult T Cell Leukemia: Antigens in an ATL cell line and detection of antibodies to antigen in human sera. Proc Natl Acad Sci U S A. 1981;78:6476-6480.
(3) Hinuma Y, Komoda H, Chosa T et al. Antibodies to adult T-cell leukemia virus-associated antigen (ATLA) in sera from patients with ATL and controls in Japan: a nation-wide seroepidemiological study. Int J Cancer. 1982;29:631.
(4) Miyoshi I, Kubonishi I, Yoshimoto S et al. Type C virus particles in a cord T-cell line derived by co-cultivating normal human cord leukocytes and human leukaemic T cells. Nature. 1981;294:770-771.
(5) Yoshida M, Miyoshi I, Hinuma Y. Isolation and characterization of retrovirus from cell lines of human T-cell leukemia and its implication in the disease. Proc Natl Acad Sci USA. 1982;79:2031-2035.
(6) Takatsuki K, Uchiyama T, Sagawa K, Yodoi J. Adult T-cell leukemia in Japan. In: Seno S, Takaku F, Irino S, eds. Topics in Hematology. Amsterdam: Excerpta Medica; 1977:73-77.
(7) Uchiyama T, Yodoi J, Sagawa K, Takatsuki K, Uchino H. Adult T-cell leukemia: clinical and hematologic features of 16 cases. Blood. 1977;50:481-492.
(8) Gessain A, Barin F, Vernant J et al. Antibodies to human T lymphotropic virus
type 1 in patients with tropical spastic paresis. Lancet. 1985;2:407-410.
(9) Osame M, Usuku K, Izumo S et al. HTLV-I associated myelopathy, a new clinical entity. Lancet. 1986;i:1031-1032.
(10) Gessain A, Mahieux R. [A virus called HTLV-1. Epidemiological aspects]. Presse Med. 2000;29:2233-2239.

44
(11) Edlich RF, Arnette JA, Williams FM. Global epidemic of human T-cell
lymphotropic virus type-I (HTLV-I). J Emerg Med. 2000;18:109-119.
(12) Blattner W, Kalyanaraman V, Robert-Guroff M et al. The human type-C retrovirus, HTLV, in blacks from the Caribbean region and relationchip to adult T-cell leukemia/lymphoma. Int J Cancer. 1982;30:257.
(13) Saxinger WC, Blattner W, Levine P et al. Human T-cell leukemia virus (HTLV-I) antibodies in Africa. Science. 1984;225:1473.
(14) Reeves W, Saxinger C, Brenes M et al. Human T-cell lymphotropic virus type I (HTLV-I) and risk factors in metropolitan Panama. Am J Epidemiol. 1988;127:539.
(15) Merino F, Robert-Guroff M, Clark J et al. Natural antibodies to human T-cell leukemia/lymphoma virus in healthy Venezuelan populations. Int J Cancer. 1984;34:501.
(16) Asher DM, Goudsmit J, Pomeroy K et al. Antibodies to HTLV-I in populations of the southwestern Pacific. J Med Virol. 1988;26:339.
(17) Blayney DW, Blattner WA, Robert-Guroff M et al. The human T-cell leukemia-lymphoma virus in the southeastern United States. JAMA. 1983;250:1048-1052.
(18) Gessain A, Mahieux R. [A virus called HTLV-1. Epidemiological aspects]
Presse Med. 2000;29:2233-2239. (19) Hollsberg P, Hafler D. Pathogenesis of diseases induced by human lymphotropic
virus type I infection. N Engl J Med. 1993;328:1173-1182.
(20) Yamamoto N, Okada M, Koyanagi Y, Kannagi M, Hinuma Y. Transformation of human Leukocytes by cocultivation with an adult T cell leukemia virus producer cell line. Science. 1982;217:737-739.
(21) Hino S, Yamaguchi K, Katamine S et al. Mother-to-child transmission of human T-cell leukemia virus type-I. Jpn J Cancer Res. 1985;76:474-480.
(22) Hino S. Milk-borne transmission of HTLV-I as a major route in the endemic
cycle. Acta Paediatr Jpn. 1989;31:428-435. (23) Fujino T, Fujiyoshi T, Yashiki S et al. HTLV-I transmission from mother to
fetus via placenta. Lancet. 1992;340:1157. (24) Wiktor SZ, Pate EJ, Murphy EL et al. Mother-to-child transmission of human T-
cell lymphotropic virus type I (HTLV-I) in Jamaica: association with antibodies

45
to envelope glycoprotein (gp46) epitopes. J Acquir Immune Defic Syndr. 1993;6:1162-1167.
(25) Manns A, Wilks RJ, Murphy EL et al. A prospective study of transmission by transfusion of HTLV-I and risk factors associated with seroconversion. Int J Cancer. 1992;51:886-891.
(26) Khabbaz RF, Douglas JM, Judson FN et al. Seroprevalence of human T-lymphotropic virus type I or II in sexually transmitted disease clinic patients in the USA. J Infect Dis. 1990;162:241-244.
(27) Khabbaz RF, Onorato IM, Cannon RO et al. Seroprevalence of HTLV-1 and HTLV-2 among intravenous drug users and persons in clinics for sexually transmitted diseases. N Engl J Med. 1992;326:375-380.
(28) Nagai M, Osame M. Human T-cell lymphotropic virus type I and neurological diseases. J Neurovirol. 2003;9:228-235.
(29) Courouce AM, Pillonel J, Lemaire JM, Saura C. HTLV testing in blood transfusion. Vox Sang. 1998;74 Suppl 2:165-169.
(30) Ferreira OC, Planelles V, Rosenblatt JD. Human T-cell leukemia viruses: Epidemiology, biology, and pathogenesis. Blood Rev. 1997;11:91-104.
(31) Murphy EL, Glynn SA, Fridey J et al. Increased incidence of infectious diseases during prospective follow-up of human T-lymphotropic virus type II- and I-infected blood donors. ARCH INTERN MED. 1999;159:1485-1491.
(32) Murphy EL, Busch MP, Tong M, Cornett P, Vyas GN. A prospective study of the risk of transfusion-acquired viral infections. Transfus Med. 1998;8:173-178.
(33) Schreiber GB, Murphy EL, Horton JA et al. Risk factors for human T-cell lymphotropic virus types I and II (HTLV-I and -II) in blood donors: the Retrovirus Epidemiology Donor Study. NHLBI Retrovirus Epidemiology Donor Study. J Acquir Immune Defic Syndr Hum Retrovirol. 1997;14:263-271.
(34) Stuver SO, Tachibana N, Okayama A et al. Heterosexual transmission of human T cell leukemia/lymphoma virus type I among married couples in southwestern Japan: an initial report from the Miyazaki Cohort Study. J Infect Dis. 1993;167:57-65.
(35) Anderson DW, Epstein JS, Lee TH et al. Serological confirmation of human T-lymphotropic virus type I infection in healthy blood and plasma donors. Blood. 1989;74:2585-2591.

46
(36) Lairmore MD, Lal R. Other Human Retrovirus Infections: HTLV-I and HTLV-II. In: Schocketman G, George JR, eds. AIDS Testing. Methodology and Management Issues. New York: Springer-Verlag; 1995:168-188.
(37) Toedter GP, Pearlman S, Hofheinz D et al. Development of a monoclonal antibody-based p24 capsid antigen detection assay for HTLV-I, HTLV-II, and STLV-I infection. AIDS Res Hum Retroviruses. 1992;8:527-532.
(38) Bangham CR. HTLV-1 infections. J Clin Pathol. 2000;53:581-586.
(39) Arisawa K, Soda M, Endo S et al. Evaluation of adult T-cell leukemia/lymphoma incidence and its impact on non-Hodgkin lymphoma incidence in southwestern Japan. Int J Cancer. 2000;85:319-324.
(40) Shimoyama M. Diagnostic criteria and classification of clinical subtypes of adult T- cell leukaemia-lymphoma. A report from the Lymphoma Study Group (1984- 87). Br J Haematol. 1991;79:428-437.
(41) Kamihira S, Sohda H, Atogami S et al. Phenotypic diversity and prognosis of
adult T-cell leukemia. Leuk Res. 1992;16:435-441.
(42) Nicot C. Current views in HTLV-I-associated adult T-cell leukemia/lymphoma. Am J Hematol. 2005;78:232-239.
(43) Kawano F, Yamaguchi K, Nishimura H, Tsuda H, Takatsuki K. Variation in the clinical courses of adult T-cell leukemia. Cancer. 1985;55:851-856.
(44) Bazarbachi A, Hermine O. Treatment of adult T-cell leukaemia/lymphoma: current strategy and future perspectives. Virus Res. 2001;78:79-92.
(45) Yao J, Wigdahl B. Human T cell lymphotropic virus type I genomic expression and impact on intracellular signaling pathways during neurodegenerative disease and leukemia. Front Biosci. 2000;5:D138-D168.
(46) Yoshida M, Seiki M, Yamaguchi K, Takatsuki K. Monoclonal integration of human T-cell leukemia provirus in all primary tumors of adult T-cell leukemia suggests causative role of human T-cell leukemia virus in the disease. Proc Natl Acad Sci U S A. 1984;81:2534-2537.
(47) Franchini G. Molecular mechanisms of human T-cell leukemia/lymphotropic virus type I infection. Blood. 1995;86:3619-3639.
(48) Hollsberg P, Wucherpfennig KW, Ausubel LJ et al. Characterization of HTLV-1 In Vivo Infected T Cell Clones IL-2-independent Growth of Nontransformed T Cells. J Immunol. 1992;148:3256-3263.

47
(49) Gatza ML, Watt JC, Marriott SJ. Cellular transformation by the HTLV-I Tax protein, a jack-of-all-trades. Oncogene. 2003;22:5141-5149.
(50) Franchini G, Nicot C, Johnson JM. Seizing of T cells by human T-cell leukemia/lymphoma virus type 1. Adv Cancer Res. 2003;89:69-132.
(51) Osame M, Janssen R, Kubota H et al. Nationwide survey of HTLV-I-associated Myelopathy in Japan: Association with blood transfusion. Ann Neurol. 1990;28:50-56.
(52) Uchiyama T. Human T cell leukemia virus type I (HTLV-I) and human diseases. [Review] [134 refs]. Annual Review of Immunology. 1997;15:15-37:15-37.
(53) Adedayo O, Grell G, Bellot P. Hospital admissions for human T-cell lymphotropic virus type-1 (HTLV-1) associated diseases in Dominica. Postgrad Med J. 2003;79:341-344.
(54) Kasahata N, Shiota J, Miyazawa Y, Nakano I, Murayama S. Acute human T-lymphotropic virus type 1-associated myelopathy: a clinicopathologic study. Arch Neurol. 2003;60:873-876.
(55) Furukawa Y, Kubota R, Eiraku N et al. Human T-cell lymphotropic virus type I (HTLV-I)-related clinical and laboratory findings for HTLV-I-infected blood donors. J Acquir Immune Defic Syndr. 2003;32:328-334.
(56) Kiwaki T, Umehara F, Arimura Y et al. The clinical and pathological features of peripheral neuropathy accompanied with HTLV-I associated myelopathy. J Neurol Sci. 2003;206:17-21.
(57) Molgaard CA, Eisenman PA, Ryden LA, Golbeck AL. Neuroepidemiology of human T-lymphotrophic virus type-I-associated tropical spastic paraparesis. Neuroepidemiology. 1989;8:109-123.
(58) Castillo LC, Gracia F, Roman GC et al. Spinocerebellar syndrome in patients infected with human T-lymphotropic virus types I and II (HTLV-I/HTLV-II): report of 3 cases from Panama. Acta Neurol Scand. 2000;101:405-412.
(59) Alamy AH, Menezes FB, Leite AC, Nascimento OM, Araujo AQ. Dysautonomia in human T-cell lymphotrophic virus type I-associated myelopathy/tropical spastic paraparesis. Ann Neurol. 2001;50:681-685.
(60) Osame M. Pathological mechanisms of human T-cell lymphotropic virus type I-associated myelopathy (HAM/TSP). J Neurovirol. 2002;8:359-364.
(61) Jacobson S. Immunopathogenesis of human T cell lymphotropic virus type I-associated neurologic disease. J Infect Dis. 2002;186 Suppl 2:S187-S192.

48
(62) Nitta T, Tanaka M, Sun B, Hanai S, Miwa M. The genetic background as a determinant of human T-cell leukemia virus type 1 proviral load. Biochem Biophys Res Commun. 2003;309:161-165.
(63) Bangham CR. Human T-lymphotropic virus type 1 (HTLV-1): persistence and immune control. Int J Hematol. 2003;78:297-303.
(64) Bangham CR. The immune control and cell-to-cell spread of human T-lymphotropic virus type 1. J Gen Virol. 2003;84:3177-3189.
(65) Yamano Y, Nagai M, Brennan M et al. Correlation of human T-cell lymphotropic virus type 1 (HTLV-1) mRNA with proviral DNA load, virus-specific CD8(+) T cells, and disease severity in HTLV-1-associated myelopathy (HAM/TSP). Blood. 2002;99:88-94.
(66) Kitze B, Usuku K. HTLV-1-mediated immunopathological CNS disease. Curr Top Microbiol Immunol. 2002;265:197-211.
(67) Osame M, Izumo S, Igata A et al. Blood transfusion and HTLV-I-associated myelopathy. Lancet. 1986;104.
(68) Osame M, Igata A, Usuku K, Rosales RL, Matsumoto M. Mother-to-child transmission in HTLV-I associated myelopathy. Lancet. 1987;1:106.
(69) Nagai M, Usuku K, Matsumoto W et al. Analysis of HTLV-I proviral load in 202 HAM/TSP patients and 243 asymptomatic HTLV-I carriers: high proviral load strongly predisposes to HAM/TSP. J Neurovirol. 1998;4:586-593.
(70) Daenke S, Nightingale S, Cruickshank JK, Bangham CRM. Sequence variants of human T-cell lymphotropic virus type I from patients with tropical spastic paraparesis and adult T-cell leukemia do not distinguish neurological from leukemic isolates. J Virol. 1990;64:1278-1282.
(71) Evangelista A, Maroushek S, Minnigan H et al. Nucleotide sequence analysis of a provirus derived from an individual with tropical spastic paraparesis. Microb Pathog. 1990;8:259-278.
(72) Gould KG, Bangham CR. Virus variation, escape from cytotoxic T lymphocytes and human retroviral persistence. Semin Cell Dev Biol. 1998;9:321-328.
(73) Mahieux R, Ibrahim F, Mauclere P et al. Molecular epidemiology of 58 new African human T-cell leukemia virus type 1 (HTLV-1) strains: Identification of a new and distinct HTLV-1 molecular subtype in central Africa and in pygmies. J Virol. 1997;71:1317-1333.

49
(74) Saito M, Furukawa Y, Kubota R et al. Frequent mutation in pX region of HTLV-1 is observed in HAM/TSP patients, but is not specifically associated with the central nervous system lesions. J Neurovirol. 1995;1:286-294.
(75) Kuroda Y, Matsui M. Cerebrospinal Fluid Interferon-Gamma Is Increased in HTLV- I- Associated Myelopathy. J Neuroimmunol. 1993;42:223-226.
(76) Matsui M, Nagumo F, Tadano J, Kuroda Y. Characterization of humoral and cellular immunity in the central nervous system of HAM/TSP. J Neurol Sci. 1995;130:183-189.
(77) Azimi N, Mariner J, Jacobson S, Waldmann TA. How does interleukin 15 contribute to the pathogenesis of HTLV type 1- associated Myelopathy/Tropical spastic paraparesis? AIDS Res Hum Retroviruses. 2000;16:1717-1722.
(78) Kawakami K, Miyazato A, Iwakura Y, Saito A. Induction of lymphocytic inflammatory changes in lung interstitium by human T lymphotropic virus type I. AMER J RESPIR CRIT CARE MED. 1999;160:995-1000.
(79) Levin MC, Jacobson S. HTLV-I associated myelopathy/tropical spastic paraparesis (HAM/TSP): A chronic progressive neurologic disease associated with immunologically mediated damage to the central nervous system. J Neurovirology. 1997;3:126-140.
(80) Levin MC, Lee SM, Kalume F et al. Autoimmunity due to molecular mimicry as a cause of neurological disease. Nat Med. 2002;8:509-513.
(81) Levin MC, Lee SM, Morcos Y, Brady J, Stuart J. Cross-reactivity between immunodominant human T lymphotropic virus type I tax and neurons: implications for molecular mimicry. J Infect Dis. 2002;186:1514-1517.
(82) Mochizuki M, Watanabe T, Yamaguchi K et al. HTLV-I Uveitis: A distinct clinical entity caused by HTLV-I. Jpn J Cancer Res. 1992;83:236-239.
(83) Nishioka K, Maruyama I, Sato K et al. Chronic inflammatory arthropathy associated with HTLV-I [letter]. Lancet. 1989;i:441.
(84) Terada K, Katamine S, Eguchi K et al. Prevalence of serum and salivary antibodies to HTLV-1 in Sjogren's syndrome. Lancet. 1994;344:1116-1119.
(85) Lagrenade L, Hanchard B, Fletcher V, Cranston B, Blattner W. Infective dermatitis of Jamaican children: a marker for HTLV-I infection. Lancet. 1990;336:1345-1347.

50
(86) Leonmonzon M, Illa I, Dalakas MC. Polymyositis in patients infected with human T-cell leukemia virus type I: The role of the virus in the cause of the disease. Ann Neurol. 1994;36:643-649.
(87) Ohshima K, Kikuchi M, Masuda Y et al. Human T-cell leukemia virus type I associated lymphadenitis. Cancer. 1992;69:239-248.
(88) Kimura I, Tsubota T, Tada S, Sogawa J. Presence of antibodies against adult T
cell leukemia antigen in the patients with chronic respiratory diseases. Acta Med Okayama. 1986;40:281-284.
(89) Xu RZ, Gao QK, Wang SJ et al. Human acute myeloid leukemias may be
etiologically associated with new human retroviral infection. Leuk Res. 1996;20:449-455.
(90) Yajima A, Kawada A, Aragane Y, Tezuka T. Detection of HTLV-I proviral DNA in sarcoidosis. Dermatology. 2001;203:53-56.
(91) Terashima A, Gotuzzo E, Alvarez H et al. [STRONGYLOIDES STERCORALIS: CLINICAL SEVERE FORMS ASSOCIATED TO HTLV-1 INFECTION]. Rev Gastroenterol Peru. 1999;19:35-40.
(92) Coffin JM. Structure and Classification of Retroviruses. The Retroviridae. 1992;1:19-49.
(93) Coffin JM. Retroviridae: The Viruses and Their Replication. 1767-1835. 1996. Phildaelphia, PA, Lippinocott-Raven Publishers.
(94) Seiki M, Hattori S, Hirayama Y, Yoshida M. Human adult T-cell leukemia
virus: Complete nucleotide sequence of the provirus genome integrated in leukemia cell DNA. Proc Natl Acad Sci. 1983;80:3618-3622.
(95) Berneman ZN, Gartenhaus RB, Reitz MS et al. Expression of alternatively spliced human T-lymphotropic virus type 1 pX mRNA in infected cell lines and in primary uncultured cells from patients with adult T-cell leukemia/lymphoma and healthy carriers. Proc Natl Acad Sci USA. 1992;89:3005-3009.
(96) Ciminale V, Pavlakis GN, Derse D, Cunningham CP, Felber BK. Complex splicing in the human T-cell leukemia virus (HTLV) family of retroviruses: Novel mRNAs and proteins produced by HTLV type I. J Virol. 1992;66:1737-1745.
(97) Koralnik IJ, Gessain A, Klotman ME et al. Protein isoforms encoded by the pX region of human T-cell leukemia/lymphotropic virus type 1. Proc Natl Acad Sci USA. 1992;89:8813-8817.

51
(98) Shimotohno K, Takano M, Teruuchi T, Miwa M. Requirement of multiple copies of a 21-nucleotide sequence in the U3 regions of human T-cell leukemia virus type I and type II long terminal repeats for trans-acting activation of transcription. Proc Natl Acad Sci U S A. 1986;83:8112-8116.
(99) Brady JN, Jeang KT, Duvall J, Khoury G. Identification of p40x-Responsive
Regulatory Sequences within the Human T-Cell Leukemia Virus Type 1 Long Terminal Repeat. J Virol. 1987;61:2175-2181.
(100) Kitado H, Chen IS, Shah NP et al. U3 sequences from HTLV-I and -II LTRs confer pX protein response to a murine leukemia virus LTR. Science. 1987;235:901-904.
(101) Tillmann M, Wessner R, Wigdahl B. Identification of human T-cell
lymphotropic virus type I 21-base- pair repeat-specific and glial cell-specific DNA- protein complexes. J Virol. 1994;68:4597-4608.
(102) Wessner R, Wigdahl B. AP-1 derived from mature monocytes and astrocytes preferentially interacts with the HTLV-I promoter central 21 bp repeat. Leukemia. 1997;11 Suppl 3:21-4.:21-24.
(103) Wessner R, Tillmann-Bogush M, Wigdahl B. Characterization of a glial cell-specific DNA-protein complex formed with the human T cell lymphotropic virus type I (HTLV-I) enhancer. J Neurovirol. 1995;1:62-77.
(104) Grant C, Barmak K, Alefantis T et al. Human T cell leukemia virus type I and neurologic disease: events in bone marrow, peripheral blood, and central nervous system during normal immune surveillance and neuroinflammation. J Cell Physiol. 2002;190:133-159.
(105) Oroszlan S. Chemical Analysis of HTLV-1 Structural proteins. In: Gallo RC, Essex ME, Gross L, eds. Human T-cell Leukemia Lymphoma Viruses. Cold Spring Harbor Laboratory Press; 1984:101-110.
(106) Kiyokawa T, Seiki M, Iwashita S et al. p27x-III and p21x-III, proteins encoded
by the pX sequence of human T-cell leukemia virus type 1. Proc Natl Acad Sci USA. 1985;82:8359-8363.
(107) Seiki M, Hikikoshi A, Taniguchi T, Yoshida M. Expression of the pX Gene of HTLV-1: General Splicing Mechanism in the HTLV Family. Science. 1985;228:1532-1534.
(108) Furukawa K, Furukawa K, Shiku H. Alternatively spliced mRNA of the pX region of human T lymphotropic virus type I proviral genome. FEBS Lett. 1991;295:141-145.

52
(109) Cereseto A, Berneman Z, Koralnik I et al. Differential expression of alternatively spliced pX mRNAs in HTLV-I-infected cell lines. Leukemia. 1997;11:866-870.
(110) Gaudray G, Gachon F, Basbous J et al. The complementary strand of the human T-cell leukemia virus type 1 RNA genome encodes a bZIP transcription factor that down-regulates viral transcription. J Virol. 2002;76:12813-12822.
(111) Ootsuyama Y, Shimotohno K, Miwa M, Oroszlan S, Sugimura T. Myristylation of gag protein in human T-cell leukemia virus type-I and type-II. Jpn J Cancer Res. 1985;76:1132-1135.
(112) Hattori S, Kiyokawa T, Imagawa K et al. Identification of gag and env gene products of human T-cell leukemia virus (HTLV). Virology. 1984;136:338-347.
(113) Craven RC, Parent LJ. Dynamic interactions of the gag polyprotein. In: Krausslich HG, ed. Morphogenesis and Maturation of Retroviruses. Springer-Verlag Berl; 1996:65-94.
(114) Garnier L, Bowzard JB, Wills JW. Recent advances and remaining problems in HIV assembly. AIDS. 1998;12 Suppl A:S5-16.
(115) Le B, I, Prevost MC, Dokhelar MC, Rosenberg AR. The PPPY motif of human T-cell leukemia virus type 1 Gag protein is required early in the budding process. J Virol. 2002;76:10024-10029.
(116) Blot V, Perugi F, Gay B et al. Nedd4.1-mediated ubiquitination and subsequent recruitment of Tsg101 ensure HTLV-1 Gag trafficking towards the multivesicular body pathway prior to virus budding. J Cell Sci. 2004;117:2357-2367.
(117) Green PL, Chen ISY. Molecular features of the human T-cell leukemia virus. Mechanisms of transformation and leukemogenicity. In: Levy JA, ed. The Retroviridae.3. New York: Plenum Press; 1994:277-311.
(118) Nam SH, Kidokoro M, Shida H, Hatanaka M. Processing of gag Precursor Polyprotein of Human T-Cell Leukemia Virus Type 1 by Virus-Encoded Protease. J Virol. 1988;62:3718-3728.
(119) Pique C, Pham D, Tursz T, Dokhelar MC. Human T-Cell Leukemia Virus Type I Envelope Protein Maturation Process: Requirements for Syncytium Formation. J Virol. 1992;66:906-913.
(120) Le B, I, Grange MP, Delamarre L et al. HTLV-1 structural proteins. Virus Res. 2001;78:5-16.

53
(121) Manel N, Kim FJ, Kinet S et al. The ubiquitous glucose transporter GLUT-1 is a receptor for HTLV. Cell. 2003;115:449-459.
(122) Green PL, Chen ISY. Regulation of human T cell leukemia virus expression. FASEB J. 1990;4:169-175.
(123) Hildreth JEK, Subramanium A, Hampton RA. Human T-cell lymphotropic virus type 1 (HTLV-1)-induced syncytium formation mediated by vascular cell adhesion molecule-1: Evidence for involvement of cell adhesion molecules in HTLV-1 biology. J Virol. 1997;71:1173-1180.
(124) Sagara Y, Ishida C, Inoue Y, Shiraki H, Maeda Y. 71-Kilodalton heat shock cognate protein acts as a cellular receptor for syncytium formation induced by human T-cell lymphotropic virus type 1. J.Virol. 72, 535-541. 1998.
(125) Sagara Y, Inoue Y, Kojima E et al. Phosphatidylglycerol Participates in
Syncytium Formation Induced by HTLV Type 1-Bearing Cells. AIDS Res Hum Retroviruses. 2001;17:125-135.
(126) Niyogi K, Hildreth JE. Characterization of new syncytium-inhibiting monoclonal antibodies implicates lipid rafts in human t-cell leukemia virus type 1 syncytium formation. J Virol. 2001;75:7351-7361.
(127) Pinon JD, Klasse PJ, Jassal SR et al. Human T-cell leukemia virus type 1 envelope glycoprotein gp46 interacts with cell surface heparan sulfate proteoglycans. J Virol. 2003;77:9922-9930.
(128) Petrini M, Pelosi-Testa E, Sposi NM et al. Constitutive expression and abnormal glycosylation of transferrin receptor in acute T-cell leukemia. Cancer Res. 1989;49:6989-6996.
(129) Manel N, Kinet S, Battini JL et al. The HTLV receptor is an early T-cell activation marker whose expression requires de novo protein synthesis. Blood. 2003;101:1913-1918.
(130) Nath MD, Ruscetti FW, Petrow-Sadowski C, Jones KS. Regulation of the cell-surface expression of an HTLV-I binding protein in human T cells during immune activation. Blood. 2003;101:3085-3092.
(131) Overbaugh J. HTLV-1 sweet-talks its way into cells. Nat Med. 2004;10:20-21.
(132) Bukrinsky MI, Sharova N, Dempsey MP et al. Active nuclear import of human immunodeficiency virus type 1 preintegration complexes. Proc Natl Acad Sci U S A. 1992;89:6580-6584.

54
(133) Cereseto A, Giacca M. Integration site selection by retroviruses. AIDS Rev. 2004;6:13-21.
(134) Cann AJ, Chen ISY. Human T-Cell Leulemia Virus Types I and II. 1849-1880. 1996. Philadelphia, PA, Lippincott-Raven Publishers.
(135) Cann AJ, Rosenblatt JD, Wachsman W, Shah NP, Chen IS. Identification of the
gene responsible for human T-cell leukaemia virus transcriptional regulation. Nature. 1985;318:571-574.
(136) Felber BK, Paskalis H, Kleinmanewing C. The pX Protein of HTLV-1 is a
Transcriptional Activator of its Long Terminal Repeats. Science. 1985;229:675-679.
(137) Fujisawa J, Seiki M, Kiyokawa T, Yoshida M. Functional activation of the long terminal repeat of human T-cell leukemia virus type 1 by a trans-acting factor. Proc Natl Acad Sci USA. 1985;82:2277-2281.
(138) Sodroski J, Rosen C, Goh WC, Haseltine W. A transcriptional activator protein encoded by the x-lor region of the human T-cell leukemia virus. Science. 1985;228:1430-1434.
(139) Sodroski JG, Rosen CA, Haseltine WA. Trans-Acting Transcriptional
Activation of the Long Terminal Repeat of Human T Lymphotropic Viruses in Infected Cells. Science. 1984;223:381-385.
(140) Burton M, Upadhyaya CD, Maier B, Hope TJ, Semmes OJ. Human T-cell leukemia virus type 1 tax shuttles between functionally discrete subcellular targets. J Virol. 2000;74:2351-2364.
(141) Semmes OJ, Jeang KT. Localization of human T-cell leukemia virus type 1 Tax to subnuclear compartments that overlap with interchromatin speckles. J Virol. 1996;70:6347-6357.
(142) Jeang KT, Giam CZ, Majone F, Aboud M. Life, death, and tax: role of HTLV-I oncoprotein in genetic instability and cellular transformation. J Biol Chem. 2004;279:31991-31994.
(143) Franklin AA, Kubik MF, Uittenbogaard MN et al. Transactivation by the human T-cell leukemia virus Tax protein is mediated through enhanced binding of activating transcription factor-2 (ATF-2) ATF-2 response and cAMP element-binding protein (CREB). J Biol Chem. 1993;268:21225-21231.
(144) Pise-Masison CA, Jeong SJ, Brady JN. Human T cell leukemia virus type 1: the role of Tax in leukemogenesis. Arch Immunol Ther Exp (Warsz ). 2005;53:283-296.

55
(145) Portis T, Harding JC, Ratner L. The contribution of NF-kappa B activity to spontaneous proliferation and resistance to apoptosis in human T-cell leukemia virus type 1 Tax- induced tumors. Blood. 2001;98:1200-1208.
(146) Sun SC, Ballard DW. Persistent activation of NF-kappa B by the Tax transforming protein of HTLV-1: hijacking cellular I kappa B kinases. Oncogene. 1999;18:6948-6958.
(147) Suzuki T, Hirai H, Fujisawa J, Fujita T, Yoshida M. A trans-activator Tax of human T-cell leukemia virus type 1 binds to NF- kappa B p50 and serum response factor (SRF) and associates with enhancer DNAs of the NF-kappa B site and CArG box. Oncogene. 1993;8:2391-2397.
(148) Yoshida M. HTLV-1 Tax: regulation of gene expression and disease. [Review].
Trends in Microbiology. 1993;1(4):131-135.
(149) Suzuki T, Fujisawa J, Toita M, Yoshida M. The trans-activator Tax of human T-cell leukemia virus type 1 (HTLV-1) interacts with cAMP-responsive element (CRE) binding and CRE modulator proteins that bind to the 21-base-pair enhancer of HTLV-1. Proc Natl Acad Sci U S A. 1993;90:610-614.
(150) Zhao LJ, Giam CZ. Human T-cell lymphotropic virus type I (HTLV-I) transcriptional activator, Tax, enhances CREB binding to HTLV-I 21-base-pair repeats by protein-protein interaction. Proceedings of the National Academy of Sciences of the United States of America. 1992;89:7070-7074.
(151) Hai TW, Liu F, Coukos WJ, Green MR. Transcription factor ATF cDNA clones: an extensive family of leucine zipper proteins able to selectively form DNA-binding heterodimers. Genes Dev. 1989;3:2083-2090.
(152) Beraud C, Lombard-Platet G, Michal Y, Jalinot P. Binding of the HTLV-I Tax1 transactivator to the inducible 21 bp enhancer is mediated by the cellular factor HEB1. EMBO J. 1991;10:3795-3803.
(153) Barnhart MK, Connor LM, Marriott SJ. Function of the human T-cell leukemia
virus type 1 21-base-pair repeats in basal transcription. J Virol. 1997;71:337-344.
(154) Jeang KT, Chiu R, Santos E, Kim SJ. Induction of the HTLV-I LTR by Jun occurs through the Tax-responsive 21- bp elements. Virology. 1991;181:218-227.
(155) Muchardt C, Seeler JS, Nirula A, Gong S, Gaynor R. Transcription factor AP-2
activates gene expression of HTLV-I. EMBO Journal. 1992;11:2573-2581.
(156) Marriott SJ, Lindholm PF, Brown KM et al. A 36-kilodalton cellular transcription factor mediates an indirect interaction of human T-cell

56
leukemia/lymphoma virus type 1 Tax1 with a responsive element in the viral long terminal repeat. Mol Cell Biol. 1990;10:4192-4201.
(157) Tanimura A, Teshima H, Fujisawa JI, Yoshida M. A New Regulatory Element that Augments the Tax-Dependent Enhancer of Human T-Cell Leukemia Virus Type 1 and Cloning of cDNAs Encoding Its Binding Proteins. J Virol. 1993;67:5375-5382.
(158) Bosselut R, Duvall JF, Gegonne A et al. The product of the c-ets-1 proto-oncogene and the related Ets2 protein act as transcriptional activators of the long terminal repeat of human T cell leukemia virus HTLV-1. EMBO J. 1990;9:3137-3144.
(159) Bosselut R, Lim F, Romond PC et al. Myb protein binds to multiple sites in the
human T cell lymphotropic virus type 1 long terminal repeat and transactivates LTR-mediated expression. Virology. 1992;186:764-769.
(160) Clark NM, Smith MJ, Hilfinger JM, Markovitz DM. Activation of the human T-
cell leukemia virus type I enhancer is mediated by binding sites for Elf-1 and the pets factor. J Virol. 1993;67:5522-5528.
(161) Colgin MA, Nyborg JK. The human T-cell leukemia virus type 1 oncoprotein Tax inhibits the transcriptional activity of c-Myb through competition for the CREB binding protein. J Virol. 1998;72:9396-9399.
(162) Torgeman A, Mor-Vaknin N, Aboud M. Sp1 is involved in a protein kinase C-independent activation of human T cell leukemia virus type I long terminal repeat by 12-O-tetradecanoylphorbol-13-acetate. Virology. 1999;254:279-287.
(163) Kimzey AL, Dynan WS. Specific regions of contact between human T-cell leukemia virus type I Tax protein and DNA identified by photocross- linking. J Biol Chem. 1998;273:13768-13775.
(164) Lenzmeier BA, Baird EE, Dervan PB, Nyborg JK. The Tax protein-DNA interaction is essential for HTLV-I transactivation in vitro. J Mol Biol. 1999;291:731-744.
(165) Lundblad JR, Kwok RP, Laurance ME et al. The human T-cell leukemia virus-1 transcriptional activator Tax enhances cAMP-responsive element-binding protein (CREB) binding activity through interactions with the DNA minor groove. J Biol Chem. 1998;273:19251-19259.
(166) Perini G, Wagner S, Green MR. Recognition of bZIP proteins by the human T-cell leukaemia virus transactivator Tax. Nature. 1995;376:602-605.

57
(167) Chrivia JC, Kwok RP, Lamb N et al. Phosphorylated CREB binds specifically to the nuclear protein CBP. Nature. 1993;365(6449):855-859.
(168) Kwok RP, Lundblad JR, Chrivia JC et al. Nuclear protein CBP is a coactivator for the transcription factor. Nature. 1994;370(6486):223-226.
(169) Bex F, Yin MJ, Burny A, Gaynor RB. Differential transcriptional activation by human T-cell leukemia virus type 1 Tax mutants is mediated by distinct interactions with CREB binding protein and p300. Mol Cell Biol. 1998;18:2392-2405.
(170) Lemasson I, Polakowski NJ, Laybourn PJ, Nyborg JK. Transcription factor binding and histone modifications on the integrated proviral promoter in human T-cell leukemia virus-I-infected T-cells. J Biol Chem. 2002;277:49459-49465.
(171) Harrod R, Kuo YL, Tang Y et al. p300 and p300/cAMP-responsive element-binding protein associated factor interact with human T-cell lymphotropic virus type-1 Tax in a multi- histone acetyltransferase/activator-enhancer complex. J Biol Chem. 2000;275:11852-11857.
(172) Jiang H, Lu H, Schiltz RL et al. PCAF interacts with tax and stimulates tax transactivation in a histone acetyltransferase-independent manner [published erratum appears in Mol Cell Biol 2000 Mar;20(5):1897]. Mol Cell Biol. 1999;19:8136-8145.
(173) Ng PW, Iha H, Iwanaga Y et al. Genome-wide expression changes induced by HTLV-1 Tax: evidence for MLK- 3 mixed lineage kinase involvement in Tax-mediated NF-kappaB activation. Oncogene. 2001;20:4484-4496.
(174) Maruyama M, Shibuya H, Harada H et al. Evidence for aberrant activation of the interleukin-2 autocrine loop by HTLV-1-encoded p40x and T3/Ti complex triggering. Cell. 1987;48:343-350.
(175) Muraoka O, Kaisho T, Tanabe M, Hirano T. Transcriptional activation of the
interleukin-6 gene by HTLV-1. Immunology Letters. 1993;37(2-3):159-165.
(176) Mori N, Mukaida N, Ballard DW, Matsushima K, Yamamoto N. Human T-cell leukemia virus type I Tax transactivates human interleukin 8 gene through acting concurrently on AP- 1 and nuclear factor-kappa B-like sites. Cancer Res. 1998;58:3993-4000.
(177) Mori N, Prager D. Transactivation of the interleukin-1 alpha promoter by human T- cell leukemia virus type I and type II tax proteins. Blood. 1996;87:3410-3417.
(178) Miyatake S, Seiki M, Malefijt RD et al. Activation of T cell-derived lymphokine genes in T cells and fibroblasts: effects of human T cell leukemia virus type I

58
p40x protein and bovine papilloma virus encoded E2 protein. Nucleic Acids Res. 1988;16:6547-6566.
(179) Cowan EP, Alexander RK, Daniel S, Kashanchi F, Brady JN. Induction of
tumor necrosis factor alpha in human neuronal cells by extracellular human T-cell lymphotropic virus type 1 Tax(1). J Virol. 1997;71:6982-6989.
(180) Lindholm PF, Reid RL, Brady JN. Extracellular Tax Protein Stimulates Tumor Necrosis Factor-B and Immunoglobulin Kappa Light Chain Expression in Lymphoid Cells. J Virol. 1992;66:1294-1302.
(181) Duyao MP, Kessler DJ, Spicer DB et al. Transactivation of the c-myc Promoter by Human T Cell Leukemia virus Type 1 tax is Mediated by NFkB. J Biol Chem. 1992;267:16288-16291.
(182) Fujii M, Sassone-Corsi P, Verma I. c-fos promotor trans-activation of cAMP on IL-2 stimulated gene expression. Proc Natl Acad Sci U S A. 1988;8526-8530.
(183) Ratner L. Regulation of expression of the c-sis proto-oncogene. Nucleic Acids Res. 1989;17:4101-4115.
(184) Fujii M, Tsuchiya H, Chuhjo T, Akizawa T, Seiki M. Interaction of HTLV-1
Tax1 with p67SRF causes the aberrant induction of cellular immediate early genes through CArG boxes. Genes Dev. 1992;6:2066-2076.
(185) Li CC, Ruscetti FW, Rice NR et al. Differential expression of Rel family
members in human T-cell leukemia virus type I-infected cells: transcriptional activation of c-rel by Tax protein. J Virol. 1993;67:4205-4213.
(186) Nicot C, Mahieux R, Takemoto S, Franchini G. Bcl-X(L) is up-regulated by HTLV-I and HTLV-II in vitro and in ex vivo ATLL samples. Blood. 2000;96:275-281.
(187) Tsukahara T, Kannagi M, Ohashi T et al. Induction of Bcl-x(L) expression by human T-cell leukemia virus type 1 tax through NF-kappa B in apoptosis-resistant T-cell transfectants with tax. J Virol. 1999;73:7981-7987.
(188) Ressler S, Morris GF, Marriott SJ. Human T-cell leukemia virus type 1 Tax transactivates the human proliferating cell nuclear antigen promoter. J Virol. 1997;71:1181-1190.
(189) Mesnard JM, Devaux C. Multiple control levels of cell proliferation by human T- cell leukemia virus type 1 tax protein. Virology. 1999;257:277-284.

59
(190) Jeang KT. Functional activities of the human T-cell leukemia virus type I Tax oncoprotein: cellular signaling through NF-kappa B. Cytokine Growth Factor Rev. 2001;12:207-217.
(191) Teshigawara K, Maeda M, Nishino K et al. Adult T leukemia cells produce a lymphokine that augments interleukin 2 receptor expression. J Mol Cell Immunol. 1985;2:17-26.
(192) Inoue J, Seiki M, Taniguchi T, Tsuru S, Yoshida M. Induction of interleukin 2 receptor gene expression by p40x encoded by human T-cell leukemia virus type 1. EMBO J. 1986;5:2883-2888.
(193) Mori N, Fujii M, Ikeda S et al. Constitutive activation of NF-kappa B in primary adult T- cell leukemia cells. Blood. 1999;93:2360-2368.
(194) Suzuki T, Hirai H, Yoshida M. Tax protein of HTLV-1 interacts with the Rel homology domain of NF-kappa B p65 and c-Rel proteins bound to the NF-kappa B binding site and activates transcription. Oncogene. 1994;9:3099-3105.
(195) Murakami T, Hirai H, Suzuki T, Fujisawa JI, Yoshida M. HTLV-1 tax enhances NF-kappa B2 expression and binds to the products p52 and p100, but does not suppress the inhibitory function of p100. Virology. 1995;206:1066-1074.
(196) Gerritsen ME, Williams AJ, Neish AS et al. CREB-binding protein/p300 are transcriptional coactivators of p65. Proc Natl Acad Sci U S A. 1997;94:2927-2932.
(197) Perkins ND, Felzien LK, Betts JC et al. Regulation of NF-kappaB by cyclin-dependent kinases associated with the p300 coactivator. Science. 1997;275:523-527.
(198) Zhong H, Voll RE, Ghosh S. Phosphorylation of NF-kappa B p65 by PKA stimulates transcriptional activity by promoting a novel bivalent interaction with the coactivator CBP/p300. Mol Cell. 1998;1:661-671.
(199) Baeuerle PA, Baltimore D. I kappa B: a specific inhibitor of the NF-kappa B transcription factor. Science. 1988;242:540-546.
(200) Hirai H, Suzuki T, Fujisawa J, Inoue J, Yoshida M. Tax Protein of Human T-Cell Leukemia Virus Type I Binds to the Ankyrin Motifs of Inhibitory Factor kappa b and Induces Nuclear Translocation of Transcription Factor NF- kappa b Proteins for Transcriptional Activation. Proc Natl Acad Sci USA. 1994;91:3584-3588.

60
(201) Maggirwar SB, Harhaj E, Sun SC. Activation of NF-kappa B/Rel by Tax involves degradation of I kappa B alpha and is blocked by a proteasome inhibitor. Oncogene. 1995;11:993-998.
(202) Suzuki T, Hirai H, Murakami T, Yoshida M. Tax protein of HTLV-1 destabilizes the complexes of NF- kappa B and I kappa B-alpha and induces nuclear translocation of NF-kappa B for transcriptional activation. Oncogene. 1995;10:1199-1207.
(203) Li XH, Murphy KM, Palka KT, Surabhi RM, Gaynor RB. The human T-cell leukemia virus type-1 Tax protein regulates the activity of the I kappa B kinase complex. J Biol Chem. 1999;274:34417-34424.
(204) Brauweiler A, Garrus JE, Reed JC, Nyborg JK. Repression of Bax gene expression by the HTLV-I tax protein: Implications for suppression of apoptosis in virally infected cells. Virology. 1997;231:135-140.
(205) Kwok RPS, Laurance ME, Lundblad JR et al. Control of cAMP-regulated enhancers by the viral transactivator Tax through CREB and the co-activator CBP. Nature. 1996;380:642-646.
(206) Suzuki T, Uchidatoita M, Yoshida M. Tax protein of HTLV-1 inhibits CBP/p300-mediated transcription by interfering with recruitment of CBP/p300 onto DNA element of E-box or p53 binding site. Oncogene. 1999;18:4137-4143.
(207) Eckner R, Yao TP, Oldread E, Livingston DM. Interaction and functional collaboration of p300/CBP and bHLH proteins in muscle and B-cell differentiation. Genes Dev. 1996;10:2478-2490.
(208) Lundblad JR, Kwok RP, Laurance ME, Harter ML, Goodman RH. Adenoviral E1A-associated protein p300 as a functional homologue of the transcriptional co-activator CBP. Nature. 1995;374:85-88.
(209) Yoshida M. Multiple viral strategies of htlv-1 for dysregulation of cell growth control. Annu Rev Immunol. 2001;19:475-496.
(210) Suzuki T, Kitao S, Matsushime H, Yoshida M. HTLV-1 Tax protein interacts with cyclin-dependent kinase inhibitor p16(INK4A) and counteracts its inhibitory activity towards CDK4. EMBO J. 1996;15:1607-1614.
(211) Kamb A, Gruis NA, Weaver-Feldhaus J et al. A cell cycle regulator potentially involved in genesis of many tumor types. Science. 1994;264:436-440.
(212) Suzuki T, Narita T, Uchidatoita M, Yoshida M. Down-regulation of the INK4 family of cyclin-dependent kinase inhibitors by tax protein of HTLV-1 through two distinct mechanisms. Virology. 1999;259:384-391.

61
(213) Jin DY, Spencer F, Jeang KT. Human T cell leukemia virus type 1 oncoprotein Tax targets the human mitotic checkpoint protein MAD1. Cell. 1998;93:81-91.
(214) Laherty CD, Perkins ND, Dixit VM. Human T-Cell Leukemia Virus Type-I Tax and Phorbol 12- Myristate 13-Acetate Induce Expression of the A20 Zinc Finger Protein by Distinct Mechanisms Involving Nuclear Factor-kappaB. J Biol Chem. 1993;268:5032-5039.
(215) Wu K, Bottazzi ME, de La FC et al. Protein profile of tax-associated complexes. J Biol Chem. 2004;279:495-508.
(216) Inoue J, Seiki M, Yoshida M. The second pX product p27 chi-III of HTLV-1 is required for gag gene expression. FEBS Lett. 1986;209:187-190.
(217) Inoue J, Yoshida M, Seiki M. Transcriptional (p40x) and post-transcriptional
(p27x-III) regulators are required for the expression and replication of human t-cell leukemia virus type I genes. Proc Natl Acad Sci USA. 1987;84:3653-3657.
(218) Grone M, Koch C, Grassmann R. The HTLV-1 Rex protein induces nuclear accumulation of unspliced viral RNA by avoiding intron excision and degradation. Virology. 1996;218:316-325.
(219) Seiki M, Inoue J, Hidaka M, Yoshida M. Two cis-acting elements responsible for posttranscriptional trans-regulation of gene expression of human T-cell leukemia virus type I. Proc Natl Acad Sci U S A. 1988;85:7124-7128.
(220) Ballaun C, Farrington GK, Dobrovnik M et al. Functional Analysis of Human T-Cell Leukemia Virus Type 1 rex-Response Element: Direct RNA Binding of Rex Protein Correlates with In Vivo Activity. J Virol. 1991;65:4408-4413.
(221) Barshira A, Panet A, Honigman A. An RNA Secondary Structure Juxtaposes Two Remote Genetic Signals for Human T-Cell Leukemia Virus Type 1 RNA 3'-End Processing. J Virol. 1991;65:5165-5173.
(222) Black AC, Ruland C, Yip MT et al. Human T-Cell Leukemia Virus Type II Rex Binding and Activity Require an Intact splice Donor Site and a Specific RNA Secondary Structure. J Virol. 1991;65:6645-6653.
(223) Black AC, Chen ISY, Arrigo S et al. Regulation of HTLV-II gene expretion by rex involves positive and negative cis-Acting elements in the 5 long terminal repeat. Virology. 1991;181:433-444.
(224) Kim JH, Kaufman PA, Hanly SM, Rimsky LT, Greene WC. Rex transregulation of human T-cell leukemia virus type II gene expression. J Virol. 1991;65:405-414.

62
(225) Ahmed YF, Gilmartin GM, Hanly SM, Nevins JR, Greene WC. The HTLV-1 Rex Response Element Mediates a Novel Form of mRNA Polyadenylation. Cell. 1991;64:727-737.
(226) Derse D. Bovine leukemia virus transcription is controlled by a virus-encoded trans-acting factor and by cis-acting response elements. J Virol. 1987;61:2462-2471.
(227) Black AC, Ruland CT, Luo J et al. Binding of Nuclear Proteins to HTLV-II Cis-Acting Repressive Sequence (CRS) RNA Correlates with CRS Function. Virology. 1994;200:29-41.
(228) King JA, Bridger JM, Lochelt M et al. Nucleocytoplasmic transport of HTLV-1 RNA is regulated by two independent LTR encoded nuclear retention elements. Oncogene. 1998;16:3309-3316.
(229) Seiki M, Hikikoshi A, Yoshida M. The U5 sequence is a cis-acting repressive element for genomic RNA expression of human T cell leukemia virus type I. Virology. 1990;176:81-86.
(230) Bogerd HP, Huckaby GL, Ahmed YF, Hanly SM, Greene WC. The type I human T-cell leukemia virus (HTLV-I) Rex trans-activator binds directly to the HTLV-I Rex and the type 1 human immunodeficiency virus Rev RNA response elements. Proc Natl Acad Sci U S A. 1991;88:5704-5708.
(231) Grassmann R, Berchtold S, Aepinus C et al. In Vitro Binding of Human T-Cell Leukemia Virus rex Proteins to the rex-Response Element of Viral Transcripts. J Virol. 1991;65:3721-3727.
(232) Nosaka T, Siomi H, Adachi Y et al. Nucleolar targeting signal of human T-cell leukemia virus type I rex- encoded protein is essential for cytoplasmic accumulation of unspliced viral mRNA. Proc Natl Acad Sci U S A. 1989;86:9798-9802.
(233) Siomi H, Shida H, Nam SH et al. Sequence requirements for nucleolar
localization of human T cell leukemia virus type I pX protein, which regulates viral RNA processing. Cell. 1988;55:197-209.
(234) Bogerd HP, Echarri A, Ross TM, Cullen BR. Inhibition of human
immunodeficiency virus Rev and human T- cell leukemia virus Rex function, but not Mason-Pfizer monkey virus constitutive transport element activity, by a mutant human nucleoporin targeted to Crm1. J Virol. 1998;72:8627-8635.
(235) Hakata Y, Umemoto T, Matsushita S, Shida H. Involvement of human CRM1 (exportin 1) in the export and multimerization of the Rex protein of human T-cell leukemia virus type 1. J Virol. 1998;72:6602-6607.

63
(236) Palmeri D, Malim MH. The human T-cell leukemia virus type 1 posttranscriptional trans- activator Rex contains a nuclear export signal. J Virol. 1996;70:6442-6445.
(237) Adachi Y, Copeland TD, Takahashi C et al. Phosphorylation of the Rex protein of human T-cell leukemia virus type I. Journal of Biological Chemistry. 1992;267:21977-21981.
(238) Orita S, Saiga A, Takagi S et al. A novel alternatively spliced viral mRNA transcribed in cells infected with human T cell leukemia virus type 1 is mainly responsible for expressing p21X protein. FEBS Lett. 1991;295:127-134.
(239) Kubota S, Hatanaka M, Pomerantz RJ. Nucleo-cytoplasmic redistribution of the
HTLV-I rex protein: Alterations by coexpression of the HTLV-I p21(x) protein. Virology. 1996;220:502-507.
(240) Ciminale V, D'Agostino D, Zotti L et al. Expression and characterization of proteins produced by mRNAs spliced into the X region of the human T-cell leukemia/lymphotropic virus type II. Virology. 1995;209:445-456.
(241) Koralnik IJ, Fullen J, Franchini G. The p12I, p13II, and p30II proteins encoded by human T-cell leukemia/lymphotropic virus type I open reading frames I and II are localized in three different cellular compartments. J Virol. 1993;67:2360-2366.
(242) Koralnik IJ, Gessain A, Klotman ME et al. Protein isoforms encoded by the pX region of human T-cell leukemia/lymphotropic virus type I. Proc Natl Acad Sci U S A. 1992;89:8813-8817.
(243) Derse D, Mikovits J, Ruscetti F. X-I and X-II open reading frames of HTLV-I are not required for virus replication or for immortalization of primary T-cells in vitro. Virology. 1997;237:123-128.
(244) Kim SJ, Ding W, Albrecht B, Green PL, Lairmore MD. A conserved calcineurin-binding motif in human T lymphotropic virus type 1 p12I functions to modulate nuclear factor of activated T cell activation. J Biol Chem. 2003;278:15550-15557.
(245) Albrecht B, D'Souza CD, Ding W et al. Activation of nuclear factor of activated T cells by human T- lymphotropic virus type 1 accessory protein p12(I). J Virol. 2002;76:3493-3501.
(246) Ding W, Albrecht B., Kelley RE et al. Human T lymphotropic virus type 1 p12(I) expression increases cytoplasmic calcium to enhance the activation of nuclear factor of activated T cells. J Virol. 2002;76:10374-10382.

64
(247) Zhang W, Nisbet JW, Albrecht B et al. Human T-lymphotropic virus type 1 p30(II) regulates gene transcription by binding CREB binding protein/p300. J Virol. 2001;75:9885-9895.
(248) Bartoe JT, Albrecht B, Collins ND et al. Functional role of pX open reading frame II of human T- lymphotropic virus type 1 in maintenance of viral loads in vivo. J Virol. 2000;74:1094-1100.
(249) Lairmore MD, Albrecht B, D'Souza C et al. In vitro and in vivo functional analysis of human T cell lymphotropic virus type 1 pX open reading frames I and II. AIDS Res Hum Retroviruses. 2000;16:1757-1764.
(250) Zhang W, Nisbet JW, Bartoe JT, Ding W, Lairmore MD. Human T-lymphotropic virus type 1 p30(II) functions as a transcription factor and differentially modulates CREB-responsive promoters. J Virol. 2000;74:11270-11277.
(251) Albrecht B, Collins ND, Burniston MT et al. Human T-lymphotropic virus type 1 open reading frame I p12(I) is required for efficient viral infectivity in primary lymphocytes. J Virol. 2000;74:9828-9835.
(252) Johnson JM, Franchini G. Retroviral proteins that target the major histocompatibility complex class I. Virus Res. 2002;88:119-127.
(253) Collins ND, Newbound GC, Albrecht B et al. Selective ablation of human T-cell lymphotropic virus type 1 p12I reduces viral infectivity in vivo. Blood. 1998;91:4701-4707.
(254) Dekaban GA, Peters AA, Mulloy JC et al. The HTLV-I orfI protein is recognized by serum antibodies from naturally infected humans and experimentally infected rabbits. Virology. 2000;274:86-93.
(255) D'Agostino DM, Zotti L, Ferro T et al. The p13II protein of HTLV type 1: comparison with mitochondrial proteins coded by other human viruses. AIDS Res Hum Retroviruses. 2000;16:1765-1770.
(256) Ciminale V, Zotti L, Dagostino DM et al. Mitochondrial targeting of the p13(II) protein coded by the x-II ORF of human T-cell leukemia/lymphotropic virus type I (HTLV-I). Oncogene. 1999;18:4505-4514.
(257) Trovato R, Mulloy JC, Johnson JM et al. A lysine-to-arginine change found in natural alleles of the human T-cell lymphotropic/leukemia virus type 1 p12(I) protein greatly influences its stability. J Virol. 1999;73:6460-6467.

65
(258) Cereseto A, Kislyakova T, Washington-Parks R, Franchini G. Differential response to genotoxic stress in immortalized or transformed HTLV-I-infected T-cells. J Gen Virol. 1999;80:1575-1581.
(259) Chen YA, Chen S, Fu C, Chen J, Osame M. Antibody reactivities to tumor-suppressor protein p53 and HTLV-I TOF, REX, and TAX in HTLV-I-infected people with differing clinical status. Int J Cancer. 1997;71:196-202.
(260) Pique C, Uretavidal A, Gessain A et al. Evidence for the chronic in vivo production of human T cell leukemia virus type I Rof and Tof proteins from cytotoxic T lymphocytes directed against viral peptides. J Exp Med. 2000;191:567-572.
(261) Princler GL, Julias JG, Hughes SH, Derse D. Roles of viral and cellular proteins in the expression of alternatively spliced HTLV-1 pX mRNAs. Virology. 2003;317:136-145.
(262) Ciminale V, D'Agostino DM, Zotti L, Chieco-Bianchi L. Coding potential of the X region of human T-cell leukemia/lymphotropic virus type II. J Acquir Immune Defic Syndr Hum Retrovirol. 1996;13 Suppl 1:S220-S227.
(263) Saksena NK, Srinivasan A, Ge YC et al. Simian T cell leukemia virus type I from naturally infected feral monkeys from Central and West Africa encodes a 91-amino acid p12 (ORF-I) protein as opposed to a 99-amino acid protein encoded by HTLV type I from humans. AIDS Res Hum Retroviruses. 1997;13:425-432.
(264) Shaw GM, Gonda MA, Flickinger GH et al. Genomes of evolutionarily divergent members of the human T-cell leukemia virus family (HTLV-I and HTLV-II) are highly conserved, especially in pX. Proc Natl Acad Sci USA. 1984;81:4544-4548.
(265) Larocca D, Chao LA, Seto MH, Brunck TK. Human T-cell leukemia virus minus strand transcription in infected T-cells. Biochem Biophys Res Commun. 1989;163:1006-1013.
(266) Hivin P, Frederic M, rpin-Andre C et al. Nuclear localization of HTLV-I bZIP factor (HBZ) is mediated by three distinct motifs. J Cell Sci. 2005;118:1355-1362.
(267) Thebault S, Basbous J, Hivin P, Devaux C, Mesnard JM. HBZ interacts with JunD and stimulates its transcriptional activity. FEBS Lett. 2004;562:165-170.
(268) Basbous J, Arpin C, Gaudray G et al. The HBZ factor of human T-cell leukemia virus type I dimerizes with transcription factors JunB and c-Jun and modulates their transcriptional activity. J Biol Chem. 2003;278:43620-43627.

66
(269) Matsumoto J, Ohshima T, Isono O, Shimotohno K. HTLV-1 HBZ suppresses AP-1 activity by impairing both the DNA-binding ability and the stability of c-Jun protein. Oncogene. 2005;24:1001-1010.
(270) Arnold J, Yamamoto B, Li M et al. Enhancement of infectivity and persistence in vivo by HBZ, a natural antisense coded protein of HTLV-1. Blood. 2006.
(271) Chou KS, Okayama A, Tachibana N, Lee TH, Essex M. Nucleotide sequence analysis of a full-length human T-cell leukemia virus type I from adult T-cell leukemia cells: A prematurely terminated pX open reading frame II. Int J Cancer. 1995;60:701-706.
(272) D'Agostino DM, Ranzato L, Arrigoni G et al. Mitochondrial alterations induced by the p13II protein of human T-cell leukemia virus type 1. Critical role of arginine residues. J Biol Chem. 2002;277:34424-34433.
(273) Roithmann S, Pique C, Le Cesne A et al. The open reading frame I (ORF I)/ORF II part of the human T-cell leukemia virus type 1 X region is dispensable for p40tax, p27rex, or envelope expression. J Virol. 1994;68:3448-3451.
(274) D'Agostino DM, Zotti L, Ferro T et al. Expression and functional properties of proteins encoded in the x-II ORF of HTLV-I. Virus Res. 2001;78:35-43.
(275) Silic-Benussi M, Cavallari I, Zorzan T et al. Suppression of tumor growth and cell proliferation by p13II, a mitochondrial protein of human T cell leukemia virus type 1. Proc Natl Acad Sci U S A. 2004;101:6629-6634.
(276) Hiraragi H, Michael B, Nair A et al. Human T-lymphotropic virus type 1 mitochondria localizing protein p13II sensitizes Jurkat T cells to Ras-mediated apoptosis. J Virol. 2005;In press.
(277) Hou X, Foley S, Cueto M, Robinson MA. The Human T-Cell Leukemia Virus Type I (HTLV-I) X Region Encoded Protein p13(II) Interacts with Cellular Proteins. Virology. 2000;277:127-135.
(278) Lefebvre L, Vanderplasschen A, Ciminale V et al. Oncoviral Bovine Leukemia Virus G4 and Human T-Cell Leukemia Virus Type 1 p13(II) Accessory Proteins Interact with Farnesyl Pyrophosphate Synthetase. J Virol. 2002;76:1400-1414.
(279) Lefebvre L, Ciminale V, Vanderplasschen A et al. Subcellular localization of the bovine leukemia virus R3 and G4 accessory proteins. J Virol. 2002;76:7843-7854.
(280) Hiraragi H, Kim SJ, Phipps AJ et al. Human T-Lymphotropic Virus Type 1 Mitochondrion-Localizing Protein p13II Is Required for Viral Infectivity In Vivo. J Virol. 2006;80:3469-3476.

67
(281) D'Agostino DM, Ciminale V, Zotti L, Rosato A, Chieco-Bianchi L. The human T-cell lymphotropic virus type 1 Tof protein contains a bipartite nuclear localization signal that is able to functionally replace the amino-terminal domain of Rex. J Virol. 1997;71:75-83.
(282) Nicot C, Dundr M, Johnson JM et al. HTLV-1-encoded p30II is a post-transcriptional negative regulator of viral replication. Nat Med. 2004;10:197-201.
(283) Younis I, Khair L, Dundr M et al. Repression of Human T-Cell Leukemia Virus Type 1 and Type 2 Replication by a Viral mRNA-Encoded Posttranscriptional Regulator. J Virol. 2004;78:11077-11083.
(284) Younis I, Boris-Lawrie K, Green PL. Human T-cell leukemia virus open reading frame II encodes a posttranscriptional repressor that is recruited at the level of transcription. J Virol. 2006;80:181-191.
(285) Michael B, Nair AM, Hiraragi H et al. Human T lymphotropic virus type-1 p30II alters cellular gene expression to selectively enhance signaling pathways that activate T lymphocytes. Retrovirology. 2004;1:39.
(286) Silverman LR, Phipps AJ, Montgomery A, Ratner L, Lairmore MD. Human T-cell lymphotropic virus type 1 open reading frame II-encoded p30II is required for in vivo replication: evidence of in vivo reversion. J Virol. 2004;78:3837-3845.
(287) Iniguez AM, Otsuki K, Magalhaes GP, Silva EA, Vicente AC. Genetic markers on the HTLV-1 p12I protein sequences from Brazilian HAM/TSP patients and asymptomatic HTLV-1 carrier isolates. AIDS Res Hum Retroviruses. 2005;21:580-582.
(288) Trovato R, Mulloy JC, Johnson JM et al. A lysine-to-arginine change found in natural alleles of the human T-cell lymphotropic leukemia virus type 1 p12 protein greatly influences its stability. J Virol. 1999;73:6460-6467.
(289) Pawson T. Protein modules and signalling networks. Nature. 1995;373:573-580. (290) Craig HM, Reddy TR, Riggs NL, Dao PP, Guatelli JC. Interactions of HIV-1 nef
with the mu subunits of adaptor protein complexes 1, 2, and 3: role of the dileucine-based sorting motif. Virology. 2000;271:9-17.
(291) Martins ML, Soares BC, Ribas JG et al. Frequency of p12K and p12R Alleles of HTLV Type 1 in HAM/TSP Patients and in Asymptomatic HTLV Type 1 Carriers. AIDS Res Hum Retroviruses. 2002;18:899-902.

68
(292) Ding W, Albrecht B, Luo R et al. Endoplasmic Reticulum and cis-Golgi Localization of Human T- Lymphotropic Virus Type 1 p12(I): Association with Calreticulin and Calnexin. J Virol. 2001;75:7672-7682.
(293) Ding W, Kim SJ, Nair AM et al. Human T-Cell Lymphotropic Virus Type 1 p12(I) Enhances Interleukin-2 Production during T-Cell Activation. J Virol. 2003;77:11027-11039.
(294) Krause KH, Michalak M. Calreticulin. Cell. 1997;88:439-443.
(295) Michalak M, Corbett EF, Mesaeli N, Nakamura K, Opas M. Calreticulin: one protein, one gene, many functions. Biochem J. 1999;344 Pt 2:281-292.
(296) Garry RF. Sequence similarities between latent membrane protein LMP- 1 of Epstein-Barr virus, integral membrane protein p12(I) of human T cell leukemia lymphotropic virus type 1, E5 transformation protein of bovine papillomavirus, and the transmembrane proteins of slowly transforming retroviruses. AIDS Res Hum Retroviruses. 1995;11:431-432.
(297) Franchini G, Mulloy JC, Koralnik IJ et al. The human T-cell leukemia/lymphotropic virus type I p12I protein cooperates with the E5 oncoprotein of bovine papillomavirus in cell transformation and binds the 16-kilodalton subunit of the vacuolar H+ ATPase. Journal of Virology. 1993;67:7701-7704.
(298) Koralnik I, Mulloy JC, Andresson T, Fullen J, Franchini G. Mapping of the intermolecular association of the human T-cell leukemia/lymphotropic virus type 1 p12I and the vacuolar H+ATPase 16 kDa subunit protein. J Gen Virol. 1995;76:1909-1916.
(299) Trovato R, Mulloy JC, Johnson JM et al. A lysine-to-arginine change found in natural alleles of the human T-cell lymphotropic leukemia virus type 1 p12 protein greatly influences its stability. J Virol. 1999;73:6460-6467.
(300) Nicot C, Mulloy JC, Ferrari MG et al. HTLV-1 p12(I) protein enhances STAT5 activation and decreases the interleukin-2 requirement for proliferation of primary human peripheral blood mononuclear cells. Blood. 2001;98:823-829.
(301) Collins ND, D'Souza C, Albrecht B et al. Proliferation response to interleukin-2 and Jak/Stat activation of T cells immortalized by human T-cell lymphotropic virus type 1 is independent of open reading frame I expression. J Virol. 1999;73:9642-9649.
(302) Johnson JM, Mulloy JC, Ciminale V et al. The MHC class I heavy chain is a common target of the small proteins encoded by the 3' end of HTLV type 1 and HTLV type 2. AIDS Res Hum Retroviruses. 2000;16:1777-1781.

69
(303) Johnson JM, Fullen J, Casareto L, Mulloy JC. p12(I) May Contribute TO HTLV-1 Escape from Immune Detection. Leukemia. 2000;14:558.
(304) Johnson JM, Nicot C, Fullen J et al. Free Major Histocompatibility Complex
Class I Heavy Chain Is Preferentially Targeted for Degradation by Human T-Cell Leukemia/Lymphotropic Virus Type 1 p12(I) Protein. J Virol. 2001;75:6086-6094.
(305) Boustany LM, Cyert MS. Calcineurin-dependent regulation of Crz1p nuclear export requires Msn5p and a conserved calcineurin docking site. Genes Dev. 2002;16:608-619.
(306) Robek MD, Wong FH, Ratner L. Human T-Cell leukemia virus type 1 pX-I and pX-II open reading frames are dispensable for the immortalization of primary lymphocytes. J Virol. 1998;72:4458-4462.
(307) Manninen A, Saksela K. HIV-1 Nef interacts with inositol trisphosphate receptor to activate calcium signaling in T cells. J Exp Med. 2002;195:1023-1032.
(308) Petit C, Buseyne F, Boccaccio C et al. Nef is required for efficient hiv-1 replication in cocultures of dendritic cells and lymphocytes. Virology. 2001;286:225-236.
(309) Wu Y, Marsh JW. Selective transcription and modulation of resting T cell activity by preintegrated HIV DNA. Science. 2001;293:1503-1506.
(310) Tsukahara T, Ratner L. Substitution of HIV Type 1 Nef with HTLV-1 p12. AIDS Res Hum Retroviruses. 2004;20:938-943.
(311) Foyouzi-Youssefi R, Arnaudeau S, Borner C et al. Bcl-2 decreases the free Ca2+ concentration within the endoplasmic reticulum. Proc Natl Acad Sci U S A. 2000;97:5723-5728.
(312) He H, Lam M, McCormick TS, Distelhorst CW. Maintenance of calcium homeostasis in the endoplasmic reticulum by Bcl-2. J Cell Biol. 1997;138:1219-1228.
(313) Foyouzi-Youssefi R, Arnaudeau S, Borner C et al. Bcl-2 decreases the free Ca2+ concentration within the endoplasmic reticulum. Proc Natl Acad Sci U S A. 2000;97:5723-5728.
(314) Shibasaki F, Kondo E, Akagi T, Mckeon F. Suppression of signalling through transcription factor NF-AT by interactions between calcineurin and Bcl-2. Nature. 1997;386:728-731.

70
(315) Srivastava RK, Sasaki CY, Hardwick JM, Longo DL. Bcl-2-mediated drug resistance: inhibition of apoptosis by blocking nuclear factor of activated T lymphocytes (NFAT)-induced Fas ligand transcription. J Exp Med. 1999;190:253-265.
(316) Nair A, Michael B, Hiraragi H et al. Human T lymphotropic virus type 1 accessory protein p12I modulates calcium-mediated cellular gene expression and enhances p300 expression in T lymphocytes. AIDS Res Hum Retroviruses. 2005;21:273-284.
(317) Bex F, Gaynor RB. Regulation of gene expression by HTLV-I Tax protein. Methods. 1998;16:83-94.
(318) Kinoshita S, Chen BK, Kaneshima H, Nolan GP. Host control of HIV-1 parasitism in T cells by the nuclear factor of activated T cells. Cell. 1998;95:595-604.
(319) Ballard DW. Molecular mechanisms in lymphocyte activation and growth. Immunol Res. 2001;23:157-166.
(320) Ganem D. Virology. The X files--one step closer to closure. Science. 2001;294:2299-2300.
(321) Bouchard MJ, Wang LH, Schneider RJ. Calcium signaling by HBx protein in hepatitis B virus DNA replication. Science. 2001;294:2376-2378.
(322) Tian P, Estes MK, Hu Y et al. The rotavirus nonstructural glycoprotein NSP4 mobilizes Ca2+ from the endoplasmic reticulum. J Virol. 1995;69:5763-5772.
(323) Feng P, Park J, Lee BS et al. Kaposi's sarcoma-associated herpesvirus mitochondrial K7 protein targets a cellular calcium-modulating cyclophilin ligand to modulate intracellular calcium concentration and inhibit apoptosis. J Virol. 2002;76:11491-11504.
(324) Popovic M, Sarin PS, Robert-Gurroff M et al. Isolation and transmission of human retrovirus (human t-cell leukemia virus). Science. 1983;219:856-859.
(325) Fan N, Gavalchin J, Paul B et al. Infection of Peripheral Blood Mononuclear Cells and Cell Lines by Cell-Free Human T-Cell Lymphoma/Leukemia Virus Type 1. J Clin Microbiol. 1992;30:905-910.
(326) Okochi K, Sato H, Hinuma Y. A retrospective study on transmission of adult T cell leukemia virus by blood transfusion: seroconversion in recipients. Vox Sang. 1984;46:245-253.

71
(327) Delamarre L, Rosenberg AR, Pique C, Pham D, Dokhelar MC. A novel human T-Leukemia virus type 1 cell-to-cell transmission assay permits definition of SU glycoprotein amino acids important for infectivity. J Virol. 1997;71:259-266.
(328) Jassal SR, Lairmore MD, Leigh-Brown AJ, Brighty DW. Soluble recombinant HTLV-1 surface glycoprotein competitively inhibits syncytia formation and viral infection of cells. Virus Res. 2001;78:17-34.
(329) Nagy K, Clapham P, Cheingsong-Popov R, Weiss R. Human T cell leukemia virus type 1: Induction of syncytia and inhibition by patient's sera. Int J Cancer. 1983;32:321-328.
(330) Daenke S, Mccracken SA, Booth S. Human T-cell leukaemia lymphoma virus type 1 syncytium formation is regulated in a cell-specific manner by ICAM-1, ICAM-3 and VCAM-1 and can be inhibited by antibodies to integrin beta(2) or beta(7). J Gen Virol. 1999;80:1429-1436.
(331) Igakura T, Stinchcombe JC, Goon PK et al. Spread of HTLV-I between lymphocytes by virus-induced polarization of the cytoskeleton. Science. 2003;299:1713-1716.
(332) Barnard AL, Igakura T, Tanaka Y, Taylor GP, Bangham CR. Engagement of specific T cell surface molecules regulates cytoskeletal polarization in HTLV-1-infected lymphocytes. Blood. 2005.
(333) Fukudome K, Furuse M, Fukuhara N et al. Strong induction of ICAM-1 in human T cells transformed by human T-cell-leukemia virus type 1 and depression of ICAM-1 or LFA-1 in adult T-cell-leukemia-derived cell lines. Int J Cancer. 1992;52:418-427.
(334) Bounou S, Giguere JF, Cantin R et al. The importance of virus-associated host ICAM-1 in human immunodeficiency virus type 1 dissemination depends on the cellular context. FASEB J. 2004;18:1294-1296.
(335) Fortin JF, Barbeau B, Hedman H, Lundgren E, Tremblay MJ. Role of the leukocyte function antigen-1 conformational state in the process of human immunodeficiency virus type 1-mediated syncytium formation and virus infection. Virology. 1999;257:228-238.
(336) Giguere JF, Tremblay MJ. Statin compounds reduce human immunodeficiency virus type 1 replication by preventing the interaction between virion-associated host intercellular adhesion molecule 1 and its natural cell surface ligand LFA-1. J Virol. 2004;78:12062-12065.

72
(337) Nejmeddine M, Barnard AL, Tanaka Y, Taylor GP, Bangham CR. HTLV-1 tax protein triggers microtubule reorientation in the virological synapse. J Biol Chem. 2005.
(338) Allen WE, Zicha D, Ridley AJ, Jones GE. A role for Cdc42 in macrophage chemotaxis. J Cell Biol. 1998;141:1147-1157.
(339) Sims TN, Dustin ML. The immunological synapse: integrins take the stage. Immunol Rev. 2002;186:100-117.
(340) Perez OD, Mitchell D, Jager GC et al. Leukocyte functional antigen 1 lowers T cell activation thresholds and signaling through cytohesin-1 and Jun-activating binding protein 1. Nat Immunol. 2003;4:1083-1092.
(341) Hogg N, Laschinger M, Giles K, Mcdowall A. T-cell integrins: more than just sticking points. J Cell Sci. 2003;116:4695-4705.
(342) von Andrian UH, Mackay CR. T-cell function and migration. Two sides of the same coin. N Engl J Med. 2000;343:1020-1034.
(343) Carman CV, Springer TA. Integrin avidity regulation: are changes in affinity and conformation underemphasized? Curr Opin Cell Biol. 2003;15:547-556.
(344) Stewart M, Hogg N. Regulation of leukocyte integrin function: affinity vs. avidity. J Cell Biochem. 1996;61:554-561.
(345) Stewart MP, Mcdowall A, Hogg N. LFA-1-mediated adhesion is regulated by cytoskeletal restraint and by a Ca2+-dependent protease, calpain. J Cell Biol. 1998;140:699-707.
(346) Kucik DF, Dustin ML, Miller JM, Brown EJ. Adhesion-activating phorbol ester increases the mobility of leukocyte integrin LFA-1 in cultured lymphocytes. J Clin Invest. 1996;97:2139-2144.
(347) Constantin G, Majeed M, Giagulli C et al. Chemokines trigger immediate beta2 integrin affinity and mobility changes: differential regulation and roles in lymphocyte arrest under flow. Immunity. 2000;13:759-769.
(348) Dustin ML, Bivona TG, Philips MR. Membranes as messengers in T cell adhesion signaling. Nat Immunol. 2004;5:363-372.
(349) Kim M, Carman CV, Springer TA. Bidirectional transmembrane signaling by cytoplasmic domain separation in integrins. Science. 2003;301:1720-1725.
(350) Feigelson SW, Grabovsky V, Winter E et al. The Src kinase p56(lck) up-regulates VLA-4 integrin affinity. Implications for rapid spontaneous and

73
chemokine-triggered T cell adhesion to VCAM-1 and fibronectin. J Biol Chem. 2001;276:13891-13901.
(351) Ardouin L, Bracke M, Mathiot A et al. Vav1 transduces TCR signals required for LFA-1 function and cell polarization at the immunological synapse. Eur J Immunol. 2003;33:790-797.
(352) Griffiths EK, Krawczyk C, Kong YY et al. Positive regulation of T cell activation and integrin adhesion by the adapter Fyb/Slap. Science. 2001;293:2260-2263.
(353) Wang H, Moon EY, Azouz A et al. SKAP-55 regulates integrin adhesion and formation of T cell-APC conjugates. Nat Immunol. 2003;4:366-374.
(354) Katagiri K, Hattori M, Minato N et al. Rap1 is a potent activation signal for leukocyte function-associated antigen 1 distinct from protein kinase C and phosphatidylinositol-3-OH kinase. Mol Cell Biol. 2000;20:1956-1969.
(355) D'Souza-Schorey C, Boettner B, Van AL. Rac regulates integrin-mediated spreading and increased adhesion of T lymphocytes. Molecular & Cellular Biology. 1998;18:3936-3946.
(356) Fagerholm S, Morrice N, Gahmberg CG, Cohen P. Phosphorylation of the cytoplasmic domain of the integrin CD18 chain by protein kinase C isoforms in leukocytes. J Biol Chem. 2002;277:1728-1738.
(357) Hogg N, Henderson R, Leitinger B et al. Mechanisms contributing to the activity of integrins on leukocytes. Immunol Rev. 2002;186:164-171.
(358) Smith A, Bracke M, Leitinger B, Porter JC, Hogg N. LFA-1-induced T cell migration on ICAM-1 involves regulation of MLCK-mediated attachment and ROCK-dependent detachment. J Cell Sci. 2003;116:3123-3133.
(359) Rodriguez-Fernandez JL, Gomez M, Luque A et al. The interaction of activated integrin lymphocyte function-associated antigen 1 with ligand intercellular adhesion molecule 1 induces activation and redistribution of focal adhesion kinase and proline-rich tyrosine kinase 2 in T lymphocytes. Mol Biol Cell. 1999;10:1891-1907.
(360) Volkov Y, Long A, Kelleher D. Inside the crawling T cell: leukocyte function-associated antigen-1 cross-linking is associated with microtubule-directed translocation of protein kinase C isoenzymes beta(I) and delta. J Immunol. 1998;161:6487-6495.

74
(361) Itoyama Y, Minato S, Kira J et al. Spontaneous proliferation of peripheral blood lymphocytes increased in patients with HTLV-I-associated myelopathy. Neurology. 1988;38:1302-1307.
(362) Jacobson S, Zaninovic V, Mora C et al. Immunological findings in neurological diseases associated with antibodies to HTLV-I: activated lymphocytes in tropical spastic paraparesis. Ann Neurol. 1988;23 Suppl:S196-S200.
(363) Matsumoto M, Sugimoto M, Nakashima H et al. Spontaneous T cell proliferation and release of soluble interleukin-2 receptors in patients with HTLV-I-associated myelopathy. Am J Trop Med Hyg. 1990;42:365-373.
(364) Usuku K, Sonoda S, Osame M et al. HLA haplotype-linked high immune responsiveness against HTLV-I in HTLV-I-associated myelopathy: comparison with adult T-cell leukemia/lymphoma. Ann Neurol. 1988;23 Suppl:S143-50.:S143-S150.
(365) Hanon E, Hall S, Taylor GP et al. Abundant Tax protein expression in CD4+T cells infected with human T-cell lymphotropic virus type I (HTLV-I) is prevented by cytotoxic T lymphocytes. Blood. 2000;95:1386-1392.
(366) Tendler CL, Greenberg SJ, Blattner WA et al. Transactivation of interleukin 2 and its receptor induces immune activation in human T-cell lymphotropic virus type I-associated myelopathy: pathogenic implications and a rationale for immunotherapy. Proceedings of the National Academy of Sciences of the United States of America. 1990;87:5218-5222.
(367) Wucherpfennig KW, Hollsberg P, Richardson JH, Benjamin D, Hafler DA. T-cell activation by autologous human T-cell leukemia virus type I- infected T-cell clones. Proc Natl Acad Sci U S A. 1992;89:2110-2114.
(368) Kimata JT, Palker TJ, Ratner L. The Mitogenic Activity of Human T-Cell
Leukemia Virus Type- I Is T-Cell Associated and Requires the CD2/LFA-3 Activation Pathway. J Virol. 1993;67:3134-3141.

75
Figure 1.1 Schematic illustration of HTLV-1 proviral genome, mRNA and protein
species Diagram of HTLV-1 proviral genome (top), mRNA (middle), and protein
species (bottom). Numbers correspond to nucleotide positions of each exon splice
acceptor and donor sites with respect to the full length HTLV-1 genome. (Modified
from Wycuff and Marriott, 2005 Frontiers in Biosciences)

76
Figure 1.2. Schematic Illustration of HTLV-1 non-structural protein p12I
Diagram of p12I with putative and identified functional motifs. Abbreviations: TM,
transmembrane region; LZip, leucine zipper motif; DxxxLL, dileucine motif; PxxP,
SH3 binding motif; K/R, lysine-to-arginine variant at position 88 (arginine at this
position increases stability of protein); PxIxIT, coserved calcineurin binding site.
Numbers below bars indicate amino acid position numbers.

77
Figure 1.3. A model of p12I role in calcium-mediated signaling
Abbreviations: TCR, T cell receptor ; PTKs, protein tyrosine kinases ; PLCγ,
phospholipase Cγ ; DAG, diacylglycerolotein ; PKC, protein kinase C ;MEK mitogen-
activated protein (MAP) or extracellular signal-regulated (Erk) kinase ; AP-1, activator
protein-1 ;IP-3, inositol trisphosphate ; IP3R, inositol trisphosphate receptor ; ER,
endoplasmic reticulum; MLCK, myosin light chain kinase ; CaMKII,
Ca2+/Calmodulin-Dependent Protein Kinase II ;NFAT-ppp, phophorylated nuclear
factor of activated T-cell nuclear factor; NFATre, NFAT responsive element ; IL-2,
Interleukin 2

78
CHAPTER 2
A CONSERVED CALCINEURIN-BINDING MOTIF IN HUMAN T-
LYMPHOTROPIC VIRUS TYPE 1 p12I FUNCTIONS TO MODULATE
NFAT ACTIVATION
2.1 INTRODUCTION
As a complex retrovirus, human T lymphotropic virus type 1 (HTLV-1) encodes
common retrovirus structural and enzymatic proteins, as well as regulatory (Tax and
Rex) and accessory (p12I, p27I, p13II and p30II) proteins from four open reading frames
(ORF I-IV) in its unique pX region 1-4. The understanding of the molecular pathogenesis
of HTLV-1 infection, immortalization and transformation has been primarily focused
on the role of Tax and Rex 5,6, however emerging evidence also indicates the important
roles of accessory proteins in establishment of HTLV-1 infection 7.
The ORF I encoded protein HTLV-1 p12I is highly conserved among different
isolates and its messenger RNA can be detected in infected cell lines and directly from
infected patient cells 2-4,8,9. Humoral antibody and cytotoxic T lymphocyte responses

79
against ORF I encoded viral proteins are elicited in infected carriers and diseased
patients indicating the expression of the protein during the natural infection 10,11. p12I
localizes in the endoplasmic reticulum and cis-Golgi compartment 12-14, and associates
with the H+ vacuolar ATPase 15, the interleukin-2 (IL-2) receptor β- and γ-chain 16 and
major histocompatibility complex class I heavy chain 17. A small hydrophobic protein,
p12I contains several potential functional motifs including two transmembrane domains,
leucine zipper motifs, and four SH3 domain-binding motifs (PxxP) 2,15,18. However,
defined motifs of p12I that may alter key T cell signaling pathways that thereby
influence cell activation have not been identified.
We have reported that p12I is required for early establishment of HTLV-1
infection 19,20. When compared with the wild type infectious molecular clone (ACH),
ORF I mutated ACH immortalized T cell lines (ACH·p12I) failed to establish infection
in rabbits 21,22. ACH·p12I also exhibited reduced infectivity in quiescent peripheral
blood mononuclear cells (PBMC), but not in activated PBMC, suggesting a role for
p12I in T cell activation during the early stages of infection 20. Recently we have
demonstrated that p12I localizes to the ER and associates with an ER luminal protein,
calreticulin 12 and selectively activates the nuclear factor of activated T cells (NFAT) in
a calcium- dependent manner 12,23. Importantly, p12I expression in lymphocytes directly
increases cytoplasmic calcium from ER calcium stores 12,24, which serves to explain
how the protein activates the NFAT signaling pathway.
The transcriptional activation of NFAT is regulated by the calcium/calmodulin
dependent serine/threonine phosphatase, calcineurin 25,26. Through protein-protein
interaction, calcineurin triggers the dephosphorylation and subsequent nuclear

80
translocation of NFAT, which results in transactivation of NFAT inducible cytokine
genes including IL-2 25,26. A conserved calcineurin-binding motif (PxIxIT) has been
identified in the N-terminal regulatory domain of NFAT, which serves as a major
binding site with both inactivated and activated calcineurin 27,28. When this motif is
mutated, NFAT is functionally impaired 27,28. In addition, secondary calcineurin binding
sites in NFAT have been identified that contribute to higher affinity binding with
calcineurin 29,30.
The NFAT SPRIEIT calcineurin binding sequence and synthetic peptides
containing the critical binding amino acids (PxIxIT) inhibit binding between calcineurin
and NFAT without affecting calcineurin phosphatase activity 27,28 . A number of
calcineurin binding proteins inhibit either calcineurin phosphatase activity or its
substrate NFAT transcriptional activity. These include the anti-apoptotic protein BCL-
2 31,32, calcineurin B homologous protein (CHP) 33, AKAP79 (A kinase anchoring
protein) 34, and myocyte –enriched calcineurin-interacting protein 1(MCIP1) 35.
Interestingly, the African swine fever virus immunosuppressive protein A238L inhibits
NFAT activation, and has calcineurin binding sequence resembling that of NFAT 36,37.
Herein, we identified a conserved sequence (PSLPI/LT) in p12I, which is highly
homologous to the PxIxIT calcineurin-binding motif of NFAT. Full-length p12I and the
PSLPI/LT motif containing mutants bound calcineurin in both immunoprecipitation and
calmodulin bead pull-down assays. In contrast, serial mutations of p12I that lacked
PSLPI/LT motif or had selective alanine substitutions of the motif (p12I AxAxAA)
exhibited abolished or decreased binding affinity with calcineurin. Furthermore, p12I
competed with NFAT for calcineurin binding in calmodulin bead pull-down

81
experiments. Interestingly, the p12I AxAxAA mutant induced more NFAT nuclear
translocation than wild type and increased NFAT transcriptional activity (~ 2 fold) in a
reporter gene assay when compared to wild type p12I. The existence of a calcineurin
binding motif in p12I suggests that there may be at least two regulatory actions for p12I
that modulate NFAT activation: 1) to cause calcium release from ER stores and 2)
through calcineurin binding. Collectively our data provide new insight into the role of a
retroviral accessory protein in mediating early events in T cell infection and activation.
2.2 MATERIALS AND METHODS
Cell lines. The 293T HEK (American Type Culture Collection, catalog number 1573)
cells and Hela-Tat cells (AIDS Research and Reference Reagent Program, National
Institutes of Health, HLtat, catalog number 1293) were maintained in Dulbecco's
modified Eagle medium (supplemented with 10% fetal bovine serum, 100 µg of
streptomycin plus penicillin/ml, and 2 mM L-glutamine [Life Technologies, Rockville,
MD]). Jurkat T cells (clone E6-1, American Type Culture Collection catalog number
TIB-152) were maintained in RPMI media (Life Technologies, Rockville, MD)
supplemented with 15% fetal bovine serum, 100 µg of streptomycin plus penicillin/ml,
and 2 mM L-glutamine, and 10 mM HEPES (Life Technologies, Rockville,
MD)(complete RPMI).

82
Plasmids and site directed mutagenesis. pME 18S and pMEp12I plasmids were kindly
provided by G. Franchini (National Institute of Health)14. pMEp12I expresses an
influenza hemagglutinin (HA1) tagged HTLV-1 p12I fusion protein. HA-NFAT1c-GFP
expression vector 27 (A. Rao, Harvard Medical School) was used for NFAT expression
in 293T and Hela-Tat cell lines. The serial deletion mutants of p12I (1-47, 48-99, 15-69,
and 15-86) were previously described 12. Point mutants of p12I were generated by PCR
based site directed mutagenesis using a QuickChangeTM site-directed mutagenesis kit
(Stratagene La Jolla, CA). Point mutant L74I was made by substituting the 74th leucine
residue to isoleucine within the 99-amino acid of p12I. Point mutant p12I AxAxAA was
made by multiple alanine substitution of the PxLxLT motif (e.g., 70th proline to alanine,
71st leucine to alanine, 73rd leucine to alanine and 75th threonine to alanine) motif. To
construct L74I mutant, 5'CCTCAGCCCGTCGCTGCCGATCACGATGCGTTTCCCC
3' and 5' GGGGAAACGCATCGTGATCGGCAGCGACGGGCTGAGG 3' primers
were used. To construct p12I AxAxAA mutant, 5'
CCTCTTCTCCTCAGCGCGTCGGCGCCGGCAGCGATGCGTTTCCCC 3' and 5'
GGGGAAACGCATCGCTGCCGGCGCCGACGCGCTGAGGAGAAGAGG 3'
primers were used. Sanger sequencing was used to confirm that the sequences of each
p12I mutant plasmids were correct and in-frame. An NFAT-luciferase construct
pNFAT-luc, which has a trimerized human distal IL-2 NFAT site inserted into the
minimal IL-2 promoter was used for the reporter gene assay 38.
Transient transfection and immunoprecipitation assay. 293T cells were seeded at
50% confluence in 10-cm-diameter tissue culture dishes 24 h before transfection. Cells

83
were transfected with 10 µg of pME-18S, wild type pMEp12I and mutants of p12I using
LipofectamineTM reagent 12 (Invitrogen, Carlsbad, CA). For Jurkat T cell transfection,
107 cells were electroporated in complete RPMI (350 V, 975 µF; Biorad Gene Pulser II)
with 30 µg pME 18S, wild type pMEp12I and mutant expression vectors. To test the
binding between p12I and calcineurin, 293T cells or Jurkat T cells were transfected with
wild type pMEp12I or mutated p12I-expression vectors. Transfected 293T cells (1-2 x
107) or Jurkat T cells (1 x 107) were lysed with Triton X-100 buffer [1% deoxycholic
acid, 0.1% sodium dodecyl sulfate (SDS), 1% Triton X-100, 150 mM NaCL, 50 mM
Tris-HCl (pH 7.5), with Complete® Protease Inhibitor (Roche, Indianapolis, IN)]. CaCl2
(2 mM), EDTA (5 mM) or EGTA (5 mM) were added to test the calcium chelation
effect in p12I and calcineurin binding. Cell lysates were incubated with 1:200 diluted
rabbit polyclonal calcineurin antibody (Chemicon International, Inc., Temecular, CA)
overnight. The immune complex mixture was then incubated with 50 µl of protein A/G
plus agarose beads (Santa Cruz Biotechnology, Santa Cruz, CA) for 4 h. Beads were
washed three times in Triton X-100 buffer, boiled in SDS sample buffer and
supernatants were analyzed by Western immunoblotting. To test pharmacological
inhibitors in p12I and calcineurin binding assay, cyclosporin A (Sigma, St. Louis, MD)
(10 µM) or BAPTA-AM [Glycine, N, N’-1, 2-ethanediylbis (oxy-2, 1-phenylene)-bis-
N-2- (acetyloxy) methoxy-2-oxoethyl]-, [bis(acetyloxy)methyl-ester] (Molecular Probes,
Eugene, OR) (1 µM) were added in culture 30 min before cell lysis.

84
Calmodulin agarose bead pull-down assay. To test p12I binding with calcineurin and
to test for competition between p12I and NFAT for calcineurin binding, a calmodulin
bead assay was performed as previously described 39. Briefly, 293T cells or Jurkat T
cells were transfected with wild type pMEp12I or mutated p12I expression vectors or a
NFAT expression vector. Transfected 293T (2 x 107) or Jurkat T cell (1 x 107) were
lysed in Triton X-100 buffer [1% deoxycholic acid, 0.1% sodium dodecyl sulfate (SDS),
1% Triton X-100, 150 mM NaCL, 50 mM Tris-HCl (pH 7.5), with Complete® protease
inhibitor (Roche, Indianapolis, IN)] with CaCl2 (2 mM). Calmodulin-agarose
(Phosphodiesterase 3’, 5’-Cyclic Nucleotide Activator from bovine brain) beads
(Sigma) were washed three times in 1 ml of preactivation buffer (10 mM Tris, pH 8.0,
150 mM NaCl, and 2 mM CaCl2) to activate calmodulin. Lysates were then incubated
with 60 µl of packed beads for 3 h at 4 ° C. To remove unbound proteins, the beads
were washed three times with washing buffer [20 mM Hepes, pH 7.4, 150 mM NaCl, 5
mM sodium pyrophosphate, 10% glycerol, 1% Triton X-100, and protease inhibitors
supplemented with CaCl2 (2 mM)]. The beads were boiled in SDS sample buffer and
supernatants were analyzed by Western immunoblotting.
Immunoblotting. Western immunoblotting was used to analyze immunoprecipiated
and calmodulin bead pull downed products. Protein concentrations in all lysates were
determined by BCA assay (micro BCA Protein Assay, Pierce, Rockford, IL). Cell
lysates were separated by SDS-polyacrylamide gel electrophoresis (SDS-PAGE)
followed by transfer to nitrocellulose membranes. Membranes were blocked with 5%
non-fat dry milk for 2 h, incubated overnight at 4° C with monoclonal anti-HA antibody

85
(1:1000) (clone 16B-12, Covance Research Products, CA), with polyclonal calcineurin
antibody (1:1000) (Chemicon International Inc., Temecular, CA) or with monoclonal
NFAT1c antibody (1:500)(Santa Cruz Biotechnology, Santa Cruz, CA) for detection of
p12I, calcineurin and NFAT2 23 respectively. The blots were developed using
horseradish peroxidase-labeled secondary antibody and enhanced chemiluminescence
(Cell Signaling Technologies, Beverly, MA). The density of each band was measured
using a commercial software package (Gel-pro Analyzer software, Media Cybernetic,
Inc., Silver Spring, MD).
NFAT Reporter gene assay. For analysis of NFAT transcriptional activity in wild type
pMEp12I and alanine substituted mutant (p12I AxAxAA) transfected Jurkat T cells,
NFAT driven luciferase activity assay was performed as previously described 23. Briefly
Jurkat T cells (107) were electroporated as described above with 30 µg expression
vectors (pME 18S, pMEp12I, or p12I AxAxAA mutant), 10 µg reporter plasmid
(NFAT-luc) and 1 µg pCMV•SPORT-β-gal to normalize for transfection efficiency.
The transfected cells were plated in 6-well plates at a density of 5x105/ml. Cells were
stimulated with 20 ng/ml phorbol myristate acetate (PMA, Sigma, St. Louis, MO) at 6 h
post-transfection. After 18 h stimulation, cells were lysed (Cell Culture Lysis Reagent,
Promega, Madison, WI) and analyzed for luciferase activity according to the
manufacturer’s protocol (Promega, Madison, WI). Values were normalized for
transfection efficiency based on β-gal activity. Data expressed in reporter gene assay

86
were the mean of at least two independent experiments conducted in triplicate.
Statistical analysis was performed using Student’s t-test.
NFAT-GFP localization. HeLa-Tat cells were seeded into chamber slides (Fisher
Scientific, Pittsburg, PA) and were cotransfected with 2 µg of HA-NFAT1c-GFP
expression vector 27 and 4 µg pME 18S, pMEp12I, or p12I AxAxAA mutant expression
vectors. Two days post-transfection cells were fixed for 15 min with 2%
paraformaldehyde, and were visualized by an Olympus BH-2 fluorescence microscope
or Zeiss LSM510 confocal microscope. To quantify the HA-NFAT1c-GFP nuclear
localization, cells that had sole or predominant nuclear fluorescence were counted as
positive cells. Values were obtained from two wells of a single representative
experiment (~600 cells). To measure GFP intensity in cytoplasmic and nuclear regions
MetaMorph® Imaging software (Universal Imaging Corporation, Downingtown PA)
was used. Values represent nuclear/cytoplasmic ratio of GFP intensity of up to 25 cells.
Calcineurin phosphatase activity. Endogenous calcineurin phosphatase activity of
Jurkat cells was measured by using the calcineurin assay kit (Biomol, Plymouth
Meeting, PA) according to the manufacturer's instructions. 107 Jurkat cells were
transfected with 30 µg expression vectors (pME 18S, pMEp12I, or p12I AxAxAA
mutant). At 24 h post-transfection, phosphatase activity was measured using
phosphopeptide substrate (RII peptide) in the presence or absence of EGTA. The
released free PO4 was measured colorimetrically by using the Biomol Green reagent.

87
2.3 RESULTS
The putative calcineurin-binding motif in p12I is highly conserved. The peptide
alignment of p12I sequences with the calcineurin binding peptide sequence from NFAT
(SPRIEIT) was carried out using a commercial software package (Align X software of
Vector NTI suite, InforMax Inc., Bethesda, MD). We compared p12I sequences from
pMEp12I plasmid, ACH molecular proviral clone, and 5 different p12I sequences from
the Entrez protein database (http://www.ncbi.nlm.nih.gov). We identified a highly
conserved 70PSLPI/LT75 sequence in p12I (Fig. 2.1). p12I sequences from various
HTLV-1 strains had nearly identical sequences compared with the consensus
calcineurin binding PxIxIT sequence of NFAT except the 72nd leucine residue. The
pMEp12I plasmid was the only clone that had leucine residue at amino acid 74. This
putative calcineurin-binding motif is also conserved in the closely related simian T cell
leukemia virus type 1 (STLV-1) p12I 40. However, the accessory proteins R3 and G4 of
bovine leukemia virus (BLV), also a member of the Deltaretrovirus group, did not share
these consensus sequences 41.
The C-terminal half (48-99) of p12I containing PSLPI/LT binds calcineurin. To test
if p12I binds calcineurin, we transiently transfected p12I into both 293T and Jurkat T
cells and performed both immunoprecipitation and calmodulin bead pull-down
experiments (Fig. 2.2). In calcium containing buffer, calmodulin beads precipitated both
calcineurin and its binding protein NFAT 39. p12I was consistently pulled down in
calmodulin agarose bead and immunoprecipitation assays (Fig. 2.2B). Thus, p12I, as a

88
full-length protein, had the capacity to bind endogenously expressed calcineurin in both
293T and Jurkat T cells. As expected using the pMEp12I vector, the detected p12I in
higher percentage SDS gels (15 %) showed typical doublet bands as described 18. The
two forms of p12I may be due to post-translational modification of p12I, such as
phosphorylation or ubiquitylation 18.
To define the regions in p12I that are responsible for binding with calcineurin,
we tested serially deleted p12I mutants for calcineurin binding (Fig. 2.2A). The mutants
p12I 48-99, p12I 15-86, and p12I 15-69 bound calcineurin in immunoprecipitation and
calmodulin bead pull-down assays (Fig. 2.2B). However, the mutant p12I 1-47 (the N
terminal half of p12I) failed to bind calcineurin in either assay (Fig. 2.2B). These data
indicated that amino acids 48- 99 of p12I, which contains the putative calcineurin-
binding (70PSLPI/LT75) motif was likely responsible for calcineurin binding. The
mutant p12I 15-69, which did not contain the putative calcineurin binding motif, weakly
bound to calcineurin (Fig. 2.2B). This finding was not unexpected, because secondary
calcineurin binding sites are predicted from the p12I amino acid sequence when
compared to homologous alignment of secondary binding sequences of NFAT (data not
shown).
PSLPI/LT motif is responsible for interaction between p12I and calcineurin. To
more specifically test if the calcineurin-binding region in p12I was due to the PSLPI/LT
motif, we constructed an alanine substitution mutant of p12I (p12I AxAxAA) and
carried out immunoprecipitation and calmodulin bead pull-down assays (Fig. 2.3).

89
These substitutions were designed to replace critical proline, leucine, isoleucine, and
threonine amino residues with alanine residues (Fig. 2.3A). The p12I AxAxAA mutant
had significantly reduced binding affinity for calcineurin (Fig. 2.3B). This result
indicated that the PSLPI/LT motif in p12I is largely responsible for calcineurin binding.
Our data, however, did not exclude the possibility secondary binding sites in p12I as
implicated from our serially deletion mutant results. In addition, we constructed a
mutant L74I that converted a leucine from pMEp12I to an isoleucine more common at
this position in p12I sequences from most HTLV-1 strains including the ACH molecular
clone (PSLPLT vs. PSLPIT) (Fig. 2.1). There was no significant difference in binding
affinity between these two sequences (Fig 2.3). The other single substitution mutants in
the putative calcineurin-binding (70PSLPI/LT75) motif, P70A, S71A, P73A, and T75A
did not result in significant reduction in binding affinity (data not shown).
p12I and calcineurin binding is calcium-dependent but is not affected by
cyclosporin A. Calcineurin is activated via cooperative interactions between calcium
and calmodulin 42. Previous reports indicated that calcium was required for binding of
NFAT to calcineurin as a complex with calmodulin sepharose beads 39. To test the
calcium requirement for the association between p12I and calcineurin, we treated cell
lysates with the calcium chelators, EDTA and EGTA or used BAPTA-AM treated cells
prior to conducting immunoprecipitation experiments. The strong calcium chelator
EGTA eliminated p12I binding to calcineurin (Fig. 2.4, lane 4) whereas the weaker
calcium chelator EDTA and BAPTA-AM reduced the binding affinity in

90
immunoprecipitation assays (Fig. 2.4, lanes, 3, and 5). p12I binding was decreased when
cell lysates were treated with decreased concentration of CaCl2 or when treated with
increasing concentration of EDTA (data not shown). These data indicated that the
binding of p12I to calcineurin required calcium, which is similar to the requirement of
calcium for effective NFAT binding to calcineurin 39. In parallel, because the binding of
NFAT to calcineurin was not affected by calcineurin inhibitor cyclosoporin A 39, we
tested the affect of cyclosporin A in the p12I binding to calcineurin. As expected, p12I
binding was not inhibited by cyclosporin A (Fig. 2.4, lane 6). This result suggested that
the cyclosporin A-cyclophilin complex site is distinct from sites required for the
calcium/calmodulin activation and NFAT and p12I binding 43. Collectively our data
indicated that the calcineurin binding properties of NFAT and p12I were similar in
requiring calcium for calmodulin binding and each did not compete with binding of the
cyclosporin A-cyclophilin complex. The results suggested both proteins might have
similar binding mechanism or share a same binding site in calcineurin.
p12I competes with NFAT for calcineurin binding. Because synthetic peptides or
proteins, which contain the calcineurin-binding motif, compete with NFAT for
calcineurin binding 27,28,37, we tested if the p12I and calcineurin binding inhibits NFAT
and calcineurin binding. We transiently cotransfected 293T cells with a constant amount
of NFAT expression plasmid with increasing amounts of the wild type pMEp12I
expression plasmid (Fig. 2.5A), as well as plasmids expressing p12I mutants (p12I 1-47,
48-99,15-69, or AxAxAA mutants) (Fig 2.5B). We then performed calmodulin bead
pull-down assay to detect the amount of bound NFAT and wild type p12I or its mutant

91
proteins to the calcineurin A subunit. As expected, wild type p12I and the calcineurin
binding mutants, p12I 48-99, p12I 15-69 and p12I AxAxAA were bound in
concentration–dependent manner, but the non-calcineurin binding mutant p12I 1-47
expressed in a parallel manner failed to bind calcineurin (Fig. 2.5B). The amount of
calcineurin (A subunit) from pull-down experiments remained constant in all assays
(Fig. 2.5A). Both wild type p12I (Fig. 2.5A) and p12I 48-99 (Fig. 2.5B) effectively
competed against NFAT for calcineurin binding. In the non-calcineurin binding mutant
p12I 1-47 transfection, the amount of pull-downed NFAT was relatively constant
regardless of the amount of mutant p12I 1-47 transfection (Fig 2.5B). In addition, the
weakly calcineurin binding p12I 15-69 mutant, which did not contain calcineurin
binding sequence, did not compete with NFAT for calcineurin binding (Fig 2.5B). As
expected, mutation in calcineurin binding sequence (p12I AxAxAA mutant) resulted in
reduction of competition between p12I and NFAT (Fig 5B). Together, these results
indicated PSLPI/LT calcineurin binding sequence is critical for p12I competition with
NFAT for calcineurin binding.
The PSLPI/LT calcineurin binding sequence in p12I is an inhibitory motif for
NFAT transcriptional activity. p12I activates NFAT transcriptional activity in T cells
in a calcium-dependent manner 25. In contrast, the NFAT SPRIEIT calcineurin binding
sequence and synthetic peptides based on this motif inhibit NFAT transcriptional
activation by competing with NFAT for calcineurin binding 27. To further investigate if
p12I binding to calcineurin affected NFAT transcriptional activity, we performed an
NFAT driven reporter gene assay using p12I alanine substitution mutant, p12I AxAxAA,

92
which exhibited lowered binding affinity for calcineurin in the immunoprecipitation and
calmodulin bead pull-down assay. Jurkat T cells were transiently transfected with wild
type pMEp12I or p12I AxAxAA expression plasmids and then measured NFAT
transcriptional activities. While the wild type pMEp12I showed typical approximate 20
fold induction in the presence of PMA stimulation 23 compared to the pME empty
vector, the p12I AxAxAA mutant induced an approximate 40 fold enhancement of
luciferase activity compared to the pME control vector (Fig. 2.6). These data suggested
that the lowered binding between calcineurin and the p12I AxAxAA mutant allows
more NFAT to be available for the reporter gene assay. Alternatively, the calcineurin-
binding motif in p12I may function as a negative modulator motif for NFAT activation.
In addition, co-transfection of wild type pMEp12I and p12IAxAxAA mutant resulted in
approximate 25 fold induction over the pME control vector, which indicated pME p12I
and p12I AxAxAA did not compete for NFAT activation.
Lowered p12I binding affinity to calcineurin induces more NFAT nuclear
translocation. To test if p12I binding to calcineurin affected NFAT nuclear
translocation, we performed NFAT-GFP fusion protein localization assay when co-
expressed with wild type p12I or p12I AxAxAA mutant in Hela-Tat cells. As expected,
both wild type p12I and p12I AxAxAA mutant expression induced NFAT-GFP nuclear
translocation, while NFAT-GFP was retained in the cytoplasm in Hela-Tat cells
transfected with pME vector control (Fig. 2.7 A, B, and C). Nuclear translocation of
NFAT-GFP was activated by ionomycin and was inhibited by calcineurin inhibitor
cyclosporin A (Fig. 2.7 D). When untreated, cells transfected wild type p12I or p12I

93
AxAxAA mutant had approximate 80 % nuclear NFAT-GFP translocation, which was
5-6 fold greater than in pME control transfected (Fig. 2.7 D). We then measured GFP
intensity in nuclear and cytoplasmic regions of NFAT-GFP expressing cells to quantify
NFAT nuclear translocation. As demonstrated in Fig. 2.7 E, The nuclear/cytoplasmic
ratio of NFAT-GFP was less than 1 in the control pME transfected cells, approximately
2 in pME p12I and 2.5 in p12I AxAxAA mutant transfection. Thus p12I AxAxAA, the
lowered binding mutant to calcineurin, induced more NFAT translocation than wild
type p12I.
p12I binding to calcineurin does not affect phosphatase activity. Because many
calcineurin binding proteins inhibit calcineurin phosphatase activity 42, we tested if p12I
binding to calcineurin affected calcineurin phosphatase activity. We measured
endogenous calcineurin phosphatase activity in Jurkat T cells expressing pME, pME
p12I and the p12I AxAxAA mutant. As expected wild type p12I stimulated calcineurin
phosphatase activity ~ 3 fold over pME empty vector (Fig. 2.8). However, the
difference between phosphatase activity induced by wild type p12I and the p12I
AxAxAA mutant was not statically significant (p > 0.5). Wild type p12I, which binds
calcineurin with higher affinity compared to the p12I AxAxAA mutant, did not show
significant reduction in phosphatase activity (Fig. 2.8). Similar to NFAT, our data
indicated that p12I binding to calcineurin via PSLPI/LT calcineurin binding sequence
did not inhibit calcineurin phosphatase activity. These data indicate that the increased
NFAT transcription and nuclear translocation induced by the p12I AxAxAA mutant was

94
independent of calcineurin binding and was more likely due to the known calcium
mobilization properties of the viral protein.
2.4 DISCUSSION
Herein, we are the first to demonstrate that the HTLV-1 accessory protein p12I
binds to calcineurin and modulates NFAT activation. We identified in p12I, a highly
conserved calcineurin-binding motif, which is homologous to a PxIxIT calcineurin-
binding motif of NFAT.
p12I was consistently co-precipitated with calcineurin in both
immunoprecipitation and calmodulin bead pull-down assays. We found that PSLPI/LT
motif in p12I was largely responsible for calcineurin binding through the use of serial
deletion and alanine substitution mutants. Like NFAT, p12I binding required calcium
but was not inhibited by cyclosporin A. Thus, these two proteins appear to bind
calcineurin via similar motifs.
The PxIxIT calcineurin binding motif or highly related sequences are found in a
variety of calcineurin binding proteins of yeast, mammalian cells, and viruses 44. These
proteins are either calcineurin substrates or inhibitors of calcineurin mediated signaling.
For example, the yeast transcription factor Crz1p has a PIISIQ motif that mediates
calcineurin interaction 35 and its pattern of regulation is similar to that of NFAT.
Therefore, the dephosphorylation, nuclear accumulation and transcriptional activity of
Crz1p depend on this calcineurin-binding motif 44. Like, NFAT or Crz1p, p12I itself
may be a substrate of calcineurin. We are currently investigating if p12I is regulated by

95
calcineurin-mediated dephosphorylation. Also, calcineurin binding motifs are found in
calcineurin inhibitor proteins. Cellular protein MCIP1 and cain/cabin1 (calcineurin
inhibitor) are endogenous calcineurin inhibitors. MCIP1, which contains a PKIIQT
calcineurin-binding motif, is expressed abundantly in striated muscle, and inhibits
calcineurin dependent signaling pathways when over-expressed 35. Cain/cabin1 has a
PEITVT motif in a 38-amino acid putative calcineurin-binding domain 45. A viral
protein A238L of African swine fever virus, which inhibits NFAT transcriptional
activity, interacts with calcineurin via the PKIIIT sequence 37.
Our data indicates that PSLPI/LT calcineurin binding sequence of p12I
modulates NFAT nuclear translocation and transcription activity. This sequence of p12I
is critical for competition with NFAT for calcineurin binding. This result is consistent
with our previous study that tested NFAT driven reporter gene assay using truncation
mutants and alanine substitution mutants (AxxA) of putative PxxP motifs (SH3 domain
binding motifs) 46. There were two positive (aa 33-47, aa 87-99) and two negative
regions (aa 1-14, aa 70-86) for NFAT transcription in p12I and the mutation in the 3rd
PxxP motif (P70xxP73) resulted in increased NFAT activation about 2 fold more than
wild type. The 70th –75th PSLPI/LT calcineurin binding site corresponds to the aa 70-86
negative region and 3rd PxxP motif. This NFAT inhibitory function of the PSLPI/LT
motif was not from the inhibition of calcineurin phosphatase activity. Calcineurin
binding proteins such as AKAP 79, CHP, cain/cabin1, and A238L inhibit calcineurin
phosphatase activity by presumably affecting calcineurin active sites 42. However,
PxIxIT motif-mediated binding itself does not inhibit the catalytic activity of
calcineurin, because NFAT activation requires enzymatic activity of calcineurin, as well

96
as binding via this motif. Synthetic peptides based on PxIxIT motif inhibit NFAT
activation without affecting calcineurin catalytic activity 27,28. Likewise, p12I binding to
calcineurin via a similar motif does not inhibit calcineurin catalytic activity, but instead
influences NFAT and calcineurin interaction by competing for binding with NFAT
similar to artificial peptides representing this motif 27,28.
It is unclear why p12I has two regulatory functions for NFAT transcriptional
activity; positive modulation by increasing cytosolic calcium concentration from ER
stores 12 and negative modulation by calcineurin binding. Interestingly, Bcl-2 has these
similar properties with p12I, and the functional relationship between calcium release
from the ER and calcineurin binding of Bcl-2 is also still unresolved. Bcl-2 maintains
calcium homeostasis and prevents apoptosis by localizing not only at the mitochondrial
membrane, but also in the ER membranes 47,48. When Bcl-2 is at the ER membrane, its
function appears to increase ER calcium permeability, like p12I 47. Although its function
at the ER membrane is still poorly understood, it has been suggested that Bcl-2
functions as ion channel protein 47. On the other hand, Bcl-2 binds calcineurin via its
BH4 domain, and inhibits NFAT activity by sequestering calcineurin from NFAT
binding without affecting calcineurin catalytic activity 31,32. By inhibiting NFAT
activation, Bcl-2 may prevent Fas ligand (FasL) expression and further apoptosis. Like
Bcl-2, p12I may affect apoptosis in HTLV-1 infected T cells. Another ER membrane
protein, calcium-modulator and cyclophilin ligand (CAML) also has parallel properties
to p12I. It activates NFAT by increasing calcium flux when overexpressed in Jurkat T
cells 49,50. Also, CAML binds with calcineurin indirectly, through its association with
cyclophilin 51.

97
In summary, we demonstrate that the HTLV-1 accessory protein p12I interacts
with calcineurin, an important regulator of NFAT signaling, via a highly conserved
PSLPI/LT motif. Thus, HTLV-1, a retrovirus associated with lymphoproliferative
disease, expresses a conserved accessory protein to further T cell activation, an
important antecedent to effective viral infection, via a calcium/calcineurin/NFAT
pathway.
2.5 REFERENCES
(1) Seiki M, Hikikoshi A, Taniguchi T, Yoshida M. Expression of the pX Gene of HTLV-1: General Splicing Mechanism in the HTLV Family. Science. 1985;228:1532-1534.
(2) Koralnik IJ, Gessain A, Klotman ME et al. Protein isoforms encoded by the pX region of human T-cell leukemia/lymphotropic virus type 1. Proc Natl Acad Sci USA. 1992;89:8813-8817.
(3) Furukawa K, Furukawa K, Shiku H. Alternatively spliced mRNA of the pX region of human T lymphotropic virus type I proviral genome. FEBS Lett. 1991;295:141-145.
(4) Berneman ZN, Gartenhaus RB, Reitz MS et al. Expression of alternatively spliced human T-lymphotropic virus type 1 pX mRNA in infected cell lines and in primary uncultured cells from patients with adult T-cell leukemia/lymphoma and healthy carriers. Proc Natl Acad Sci USA. 1992;89:3005-3009.
(5) Mesnard JM, Devaux C. Multiple control levels of cell proliferation by human T- cell leukemia virus type 1 tax protein. Virology. 1999;257:277-284.
(6) Yoshida M. Multiple viral strategies of htlv-1 for dysregulation of cell growth control. Annu Rev Immunol. 2001;19:475-496.
(7) Lairmore MD, Albrecht B, D'Souza C et al. In vitro and in vivo functional analysis of human T cell lymphotropic virus type 1 pX open reading frames I and II. AIDS Res Hum Retroviruses. 2000;16:1757-1764.

98
(8) Cereseto A, Berneman Z, Koralnik I et al. Differential expression of alternatively spliced pX mRNAs in HTLV-I-infected cell lines. Leukemia. 1997;11:866-870.
(9) Ciminale V, Pavlakis GN, Derse D, Cunningham CP, Felber BK. Complex splicing in the human T-cell leukemia virus (HTLV) family of retroviruses: Novel mRNAs and proteins produced by HTLV type I. J Virol. 1992;66:1737-1745.
(10) Dekaban GA, Peters AA, Mulloy JC et al. The HTLV-I orfI protein is recognized by serum antibodies from naturally infected humans and experimentally infected rabbits. Virology. 2000;274:86-93.
(11) Pique C, Uretavidal A, Gessain A et al. Evidence for the chronic in vivo production of human T cell leukemia virus type I Rof and Tof proteins from cytotoxic T lymphocytes directed against viral peptides. J Exp Med. 2000;191:567-572.
(12) Ding W, Albrecht B, Luo R et al. Endoplasmic Reticulum and cis-Golgi Localization of Human T- Lymphotropic Virus Type 1 p12(I): Association with Calreticulin and Calnexin. J Virol. 2001;75:7672-7682.
(13) Koralnik IJ, Mulloy JC, Andresson T, Fullen J, Franchini G. Mapping of the intermolecular association of the human T cell leukaemia/lymphotropic virus type I p12(I) and the vacuolar H+- ATPase 16 kDa subunit protein. J Gen Virol. 1995;76:1909-1916.
(14) Koralnik IJ, Fullen J, Franchini G. The p12, p13 and p30 proteins encoded by human T-cell leukemia/lymphotropic virus type-1 open reading frames I and II are localized in three different cellular compartments. J Virol. 1993;67:2360-2366.
(15) Franchini G. Molecular mechanisms of human T-cell leukemia/lymphotropic virus type I infection. Blood. 1995;86:3619-3639.
(16) Mulloy JC, Crowley RW, Fullen J, Leonard WJ, Franchini G. The human T-cell leukemia/lymphotropic virus type 1 p12(I) protein binds the interleukin-2 receptor beta and gamma(c) chains and affects their expression on the cell surface. J Virol. 1996;70:3599-3605.
(17) Johnson JM, Nicot C, Fullen J et al. Free Major Histocompatibility Complex Class I Heavy Chain Is Preferentially Targeted for Degradation by Human T-Cell Leukemia/Lymphotropic Virus Type 1 p12(I) Protein. J Virol. 2001;75:6086-6094.

99
(18) Trovato R, Mulloy JC, Johnson JM et al. A lysine-to-arginine change found in natural alleles of the human T-cell lymphotropic leukemia virus type 1 p12 protein greatly influences its stability. J Virol. 1999;73:6460-6467.
(19) Collins ND, Newbound GC, Albrecht B et al. Selective ablation of human T-cell lymphotropic virus type 1 p12(I) reduces viral infectivity in vivo. Blood. 1998;91:4701-4707.
(20) Albrecht B, Collins ND, Burniston MT et al. Human T-lymphotropic virus type 1 open reading frame I p12(I) is required for efficient viral infectivity in primary lymphocytes. J Virol. 2000;74:9828-9835.
(21) Collins ND, Newbound GC, Ratner L, Lairmore MD. In vitro CD4(+) lymphocyte transformation and infection in a rabbit model with a molecular clone of human T-cell lymphotropic virus type 1. J Virol. 1996;70:7241-7246.
(22) Kimata JT, Wong F, Wang J, Ratner L. Construction and characterization of infectious human T-cell leukemia virus type 1 molecular clones. Virology. 1994;204:656-664.
(23) Albrecht B, D'Souza CD, Ding W et al. Activation of nuclear factor of activated T cells by human T- lymphotropic virus type 1 accessory protein p12(I). J Virol. 2002;76:3493-3501.
(24) Ding W, Albrecht B., Kelley RE et al. Human T lymphotropic virus type 1 p12(I) expression increases cytoplasmic calcium to enhance the activation of nuclear factor of activated T cells. J Virol. 2002;76:10374-10382.
(25) Masuda ES, Imamura R, Amasaki Y, Arai K, Arai N. Signalling into the T-cell nucleus: NFAT regulation. Cell Signal. 1998;10:599-611.
(26) Rao A, Luo C, Hogan PG. Transcription factors of the NFAT family: regulation and function. Annu Rev Immunol. 1997;15:707-747.
(27) Aramburu J, Garcia-Cozar F, Raghavan A et al. Selective inhibition of NFAT activation by a peptide spanning the calcineurin targeting site of NFAT. Mol Cell. 1998;1:627-637.
(28) Aramburu J, Yaffe MB, Lopez-Rodriguez C et al. Affinity-driven peptide selection of an NFAT inhibitor more selective than cyclosporin A. Science. 1999;285:2129-2133.
(29) Garcia-Cozar FJ, Okamura H, Aramburu JF et al. Two-site interaction of nuclear factor of activated T cells with activated calcineurin. J Biol Chem. 1998;273:23877-23883.

100
(30) Park S, Uesugi M, Verdine GL. A second calcineurin binding site on the NFAT regulatory domain. Proc Natl Acad Sci U S A. 2000;97:7130-7135.
(31) Shibasaki F, Kondo E, Akagi T, Mckeon F. Suppression of signalling through transcription factor NF-AT by interactions between calcineurin and Bcl-2. Nature. 1997;386:728-731.
(32) Srivastava RK, Sasaki CY, Hardwick JM, Longo DL. Bcl-2-mediated drug resistance: inhibition of apoptosis by blocking nuclear factor of activated T lymphocytes (NFAT)-induced Fas ligand transcription. J Exp Med. 1999;190:253-265.
(33) Lin X, Sikkink RA, Rusnak F, Barber DL. Inhibition of calcineurin phosphatase activity by a calcineurin B homologous protein. J Biol Chem. 1999;274:36125-36131.
(34) Kashishian A, Howard M, Loh C et al. AKAP79 inhibits calcineurin through a site distinct from the immunophilin-binding region. J Biol Chem. 1998;273:27412-27419.
(35) Rothermel B, Vega RB, Yang J et al. A protein encoded within the Down syndrome critical region is enriched in striated muscles and inhibits calcineurin signaling. J Biol Chem. 2000;275:8719-8725.
(36) Miskin JE, Abrams CC, Goatley LC, Dixon LK. A viral mechanism for inhibition of the cellular phosphatase calcineurin. Science. 1998;281:562-565.
(37) Miskin JE, Abrams CC, Dixon LK. African swine fever virus protein A238L interacts with the cellular phosphatase calcineurin via a binding domain similar to that of NFAT. J Virol. 2000;74:9412-9420.
(38) Northrop JP, Ullman KS, Crabtree GR. Characterization of the nuclear and cytoplasmic components of the lymphoid-specific nuclear factor of activated T cells (NF-AT) complex. J Biol Chem. 1993;268:2917-2923.
(39) Loh C, Shaw KT, Carew J et al. Calcineurin binds the transcription factor NFAT1 and reversibly regulates its activity. J Biol Chem. 1996;271:10884-10891.
(40) Saksena NK, Srinivasan A, Ge YC et al. Simian T cell leukemia virus type I from naturally infected feral monkeys from Central and West Africa encodes a 91-amino acid p12 (ORF-I) protein as opposed to a 99-amino acid protein encoded by HTLV type I from humans. AIDS Res Hum Retroviruses. 1997;13:425-432.

101
(41) Lefebvre L, Ciminale V, Vanderplasschen A et al. Subcellular localization of the bovine leukemia virus R3 and G4 accessory proteins. J Virol. 2002;76:7843-7854.
(42) Rusnak F, Mertz P. Calcineurin: form and function. Physiol Rev. 2000;80:1483-1521.
(43) Griffith JP, Kim JL, Kim EE et al. X-ray structure of calcineurin inhibited by the immunophilin-immunosuppressant FKBP12-FK506 complex. Cell. 1995;82:507-522.
(44) Boustany LM, Cyert MS. Calcineurin-dependent regulation of Crz1p nuclear export requires Msn5p and a conserved calcineurin docking site. Genes Dev. 2002;16:608-619.
(45) Lai MM, Burnett PE, Wolosker H, Blackshaw S, Snyder SH. Cain, a novel physiologic protein inhibitor of calcineurin. J Biol Chem. 1998;273:18325-18331.
(46) Ding W, Kim SJ, Nair AM et al. Human T-Cell Lymphotropic Virus Type 1 p12(I) Enhances Interleukin-2 Production during T-Cell Activation. J Virol. 2003;77:11027-11039.
(47) Foyouzi-Youssefi R, Arnaudeau S, Borner C et al. Bcl-2 decreases the free Ca2+ concentration within the endoplasmic reticulum. Proc Natl Acad Sci U S A. 2000;97:5723-5728.
(48) He H, Lam M, McCormick TS, Distelhorst CW. Maintenance of calcium homeostasis in the endoplasmic reticulum by Bcl-2. J Cell Biol. 1997;138:1219-1228.
(49) Holloway MP, Bram RJ. A hydrophobic domain of Ca2+-modulating cyclophilin ligand modulates calcium influx signaling in T lymphocytes. J Biol Chem. 1996;271:8549-8552.
(50) von Bulow GU, Bram RJ. NF-AT activation induced by a CAML-interacting member of the tumor necrosis factor receptor superfamily. Science. 1997;278:138-141.
(51) Holloway MP, Bram RJ. Co-localization of calcium-modulating cyclophilin ligand with intracellular calcium pools. J Biol Chem. 1998;273:16346-16350.

102
Figure 2.1. HTLV-1 p12I contains a highly conserved putative calcineurin-binding
motif in p12I. Sequences from HTLV-1 and STLV-1 were aligned with calcineurin
binding sequence (SPRIEIT) of NFAT using a commercial software program (Align
X,Vector NTI version 7.0). ACH 22 pMEp12I 16, HTLV-1A (Entrez Protein AAA45390),
HTLV-1B (AAA85329), HTLV-1C (AAB23360), HTLV-1D (B46181), HTLV-1E
(C61547), and STLV-1 isolate number 14 40 were compared. The conserved SPSLPI/LT
sequence is indicated in shaded box and black boxes indicate amino acid residues that
have similar properties. Consensus motif is indicated in bottom row.

103
Figure 2.2. The C-terminal half of p12I containing the PSLPI/LT motif is required
for calcineurin binding. (A) Schematic representation of wild type pMEp12I and serial
p12I deletion mutants. Shaded boxes indicate PxxP motifs and bar represents the
location of the PSLPI/LT conserved sequence. (B) Co-precipitation of wild type p12I
and serial deletion p12I mutants with calcineurin in immunoprecipitation (IP) and
calmodulin agarose bead pull-down (CaM bead) assays. Jurkat T or 293T cells
transfected with pME empty vector wild type pMEp12I or mutant p12I plasmids were
lysed and analyzed by Western blot. Polyclonal calcineurin antibody was used for
immunoprecipitation assay. Monoclonal HA-1 antibody was used to detect wild type
and mutant p12I.

104
Figure 2.3. Alanine substitution mutant of p12I (AxAxAA) decreases binding
affinity for calcineurin. (A) Schematic representation of wild type pMEp12I, alanine
substitution mutant p12I AxAxAA, and L74I plasmid. Grey boxes are PxxP motifs and
bar represents the PSLPI/LT conserved sequence. (B) 293T cells were transfected with
wild type pMEp12I, p12I AxAxAA, and L74I plasmid. Binding was tested by
immunoprecipitation (IP) and calmodulin agarose bead pull-down (CaM bead) assays.

105
Figure 2.4. p12I and calcineurin binding is inhibited by calcium chelators but not
inhibited by cyclosporin A. CaCl2 (2 mM), Calcium chelators EDTA (5 mM) (lane 3),
or EGTA (5 mM) (lane 4) were added in Triton X-100 cell lysis buffer and calcium
chelator BAPTA-AM (1 µM) (lane 5), or calcineurin inhibitor cyclosporin A (10 µM)
(lane 6) were added in 293T cells 30 min before cell lysis and following
immunoprecipitation (IP) assay.
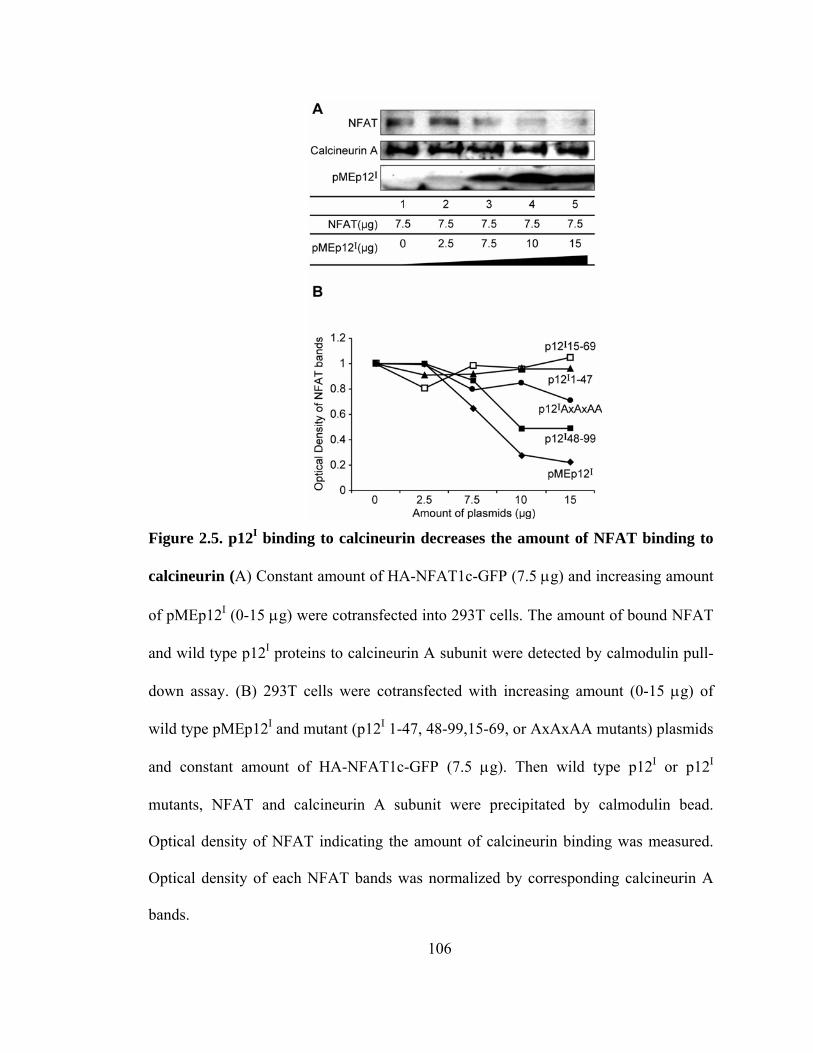
106
Figure 2.5. p12I binding to calcineurin decreases the amount of NFAT binding to
calcineurin (A) Constant amount of HA-NFAT1c-GFP (7.5 µg) and increasing amount
of pMEp12I (0-15 µg) were cotransfected into 293T cells. The amount of bound NFAT
and wild type p12I proteins to calcineurin A subunit were detected by calmodulin pull-
down assay. (B) 293T cells were cotransfected with increasing amount (0-15 µg) of
wild type pMEp12I and mutant (p12I 1-47, 48-99,15-69, or AxAxAA mutants) plasmids
and constant amount of HA-NFAT1c-GFP (7.5 µg). Then wild type p12I or p12I
mutants, NFAT and calcineurin A subunit were precipitated by calmodulin bead.
Optical density of NFAT indicating the amount of calcineurin binding was measured.
Optical density of each NFAT bands was normalized by corresponding calcineurin A
bands.

107
Figure 2.6. The substitution mutation in calcineurin binding sequence in p12I
induces increased NFAT transcription activity and did not compete against wild
type p12I for NFAT transcription activity. Jurkat T cells were co-transfected with 30
µg of effector plasmid [pME control vector, pMEp12I, p12I AxAxAA mutant, or both
pMEp12Iand p12I AxAxAA mutant (15 µg each)] and 10 µg of NFAT reporter plasmid
and then were left un-stimulated or stimulated with PMA. Each bar represent the fold
induction of NFAT-luciferase activity in wild type pMEp12I, p12I AxAxAA mutant, or
both plasmids transfected cells over luciferase activity of the pME empty vector
transfected cells. Values were the means of quadruplicate samples in two independent
experiments and were statistically significant (*, P < 0.01, pMEp12I compared to
p12IAxAxAA alone).
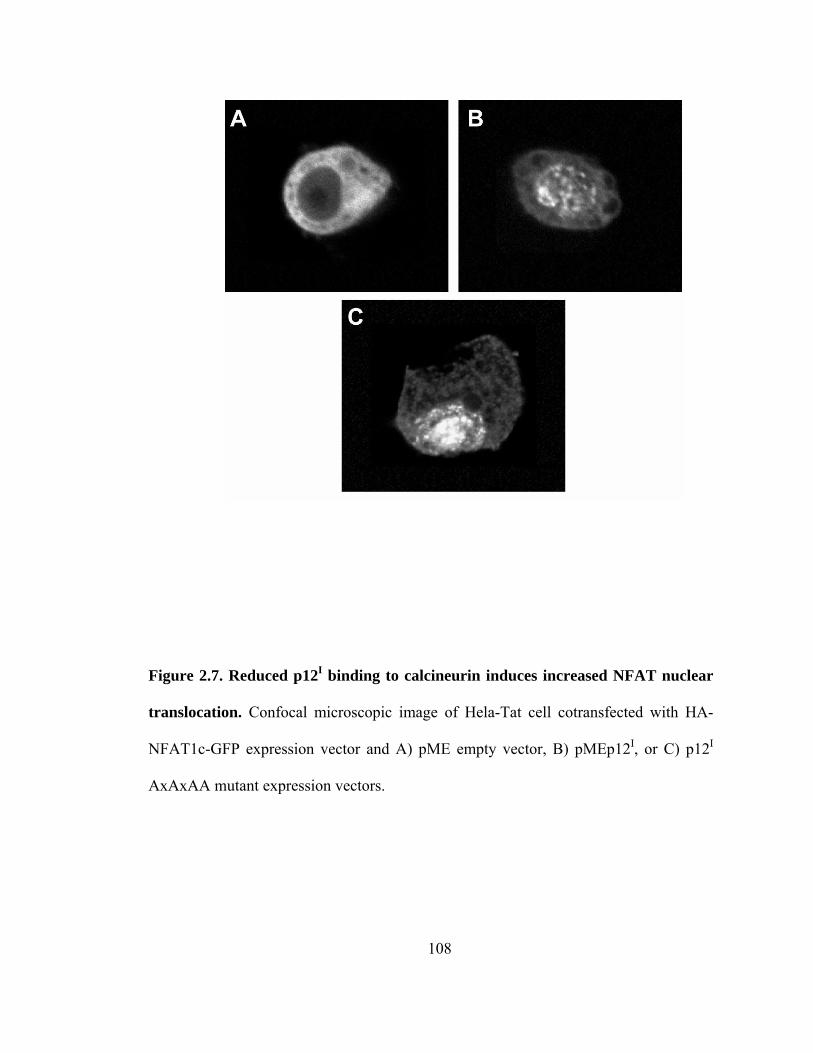
108
Figure 2.7. Reduced p12I binding to calcineurin induces increased NFAT nuclear
translocation. Confocal microscopic image of Hela-Tat cell cotransfected with HA-
NFAT1c-GFP expression vector and A) pME empty vector, B) pMEp12I, or C) p12I
AxAxAA mutant expression vectors.

109
D) The percent of cells that had nuclear predominant fluorescence. Cells were untreated,
stimulated with ionomycin (3 µM, 10 min before fixation), or inhibited with
cyclosporin A (1 µM, 16 h before fixation). E) Nuclear/cytoplasmic ratio of NFAT-GFP
intensity of cells transfected with pME, pMEp12I or p12I AxAxAA vector (*, P < 0.05,
pME vector control compared to pMEp12I. **, P < 0.05, p12I AxAxAA compared to
pMEp12I).

110
Figure 2.8. Calcineurin phosphatase activity is not affected by p12I binding to
calcineurin. Endogenous calcineurin activity from Jurkat T cells transfected with pME,
pMEp12I or p12I AxAxAA mutant was tested by measuring free phosphate (nmol)
released from phosphopeptide substrate (RII peptide) (*, P < 0.01 vs. pMEp12I and p12I
AxAxAA).

111
CHAPTER 3
ENHANCEMENT OF LFA-1-MEDIATED T-CELL ADHESION BY
HUMAN T-LYMPHOTROPIC VIRUS TYPE 1 p12I
3.1 INTRODUCTION
Human T-lymphotropic virus type 1 (HTLV-1), the first identified human
retrovirus, infects approximately 15 to 25 million people worldwide 1 and causes adult
T-cell leukemia/lymphoma (ATLL) and other lymphocyte-mediated diseases 2-5. The
primary targets for HTLV-1 infection are CD4+ T-cells, although CD8+ T-cells, B-cells,
and macrophages also can be infected 4,6. HTLV-1 is naturally transmitted in a highly
cell-associated manner via beast milk, semen and blood 2,7,8. Thus, cell-to-cell contact
is required for efficient HTLV-1 transmission both in vivo 9 and in vitro 10,11.
Furthermore, HTLV-1 transmission occurs directly through cell-to-cell junctions

112
(virological synapse) that mimic immunological synapses 12,13. HTLV-1- infected cells
have reorganized microtubule-organizing centers (MTOC) near points of cell contacts 14.
At these sites structural proteins such as Gag, along with the HTLV-1 genome,
aggregate and transfer from infected to target lymphocytes. Interestingly, microtubule
reorganization is triggered by antibodies to ICAM-1 (CD54) or the IL-2 receptor
(CD25) 14,15. While HTLV-1 Tax contributes to microtubule reorganization 12-15, it is
unclear whether other viral proteins, known to be important in the early events of
infection modulate cell-to-cell contact to accomplish viral transmission.
The HTLV-1 genome encodes structural and enzymatic (Gag, Pol, and Env),
regulatory (Tax and Rex) and nonstructural or “accessory” (p12I, p27I, p13II, and p30II)
proteins 16-20. HTLV-1 nonstructural proteins are important in viral replication and viral
spread in vivo 18,20-23. A highly conserved pX ORF I-encoded protein, p12I, is expressed
during the natural viral infection 24,25 and is critical for the establishment of HTLV-1
infection in non-activated T-cells in vitro and in animals models 26,27. p12I localizes to
the endoplasmic reticulum (ER) and Golgi apparatus, releases calcium from ER stores
to trigger nuclear factor of activated T-cells (NFAT)-mediated transcription leading to
IL-2 production and T-cell activation 21,28-30.
One of the key events of T-cell activation is the modulation of the expression of
adhesion molecules such as lymphocyte function-associated antigen 1 (LFA-1), which
mediates lymphocyte adherence to the vascular wall and lymphocyte migration into
tissues, and contributes to the immunological synapse 31. LFA-1 is a heterodimeric
transmembrane molecule composed of a unique α subunit (αL; CD11a) and a β2 subunit
(CD18) 32,33. Activation of LFA-1 is tightly controlled by “inside-out” signaling, which

113
is induced by ligation of the T-cell receptor (TCR) or certain chemokine receptors 32-35.
LFA-1-mediated T-cell adhesion results from either increasing the affinity for its ligand,
intercellular adhesion molecule-1 (ICAM-1), or clustering of LFA-1 (avidity) on the
cell membrane 32-35. Although recent studies suggested that Talin, Rap1 and RapL
proteins are required for regulation of LFA-1 expression 31,36, the detailed mechanism of
signaling leading to increased affinity is unclear 34. Under experimental conditions,
divalent cations such as Mn2+ or Mg2+ and activating antibodies can induce affinity
activation of LFA-1 34. The signaling pathways that result in increased integrin lateral
mobility and clustering include proteins such as Vav-1 (guanine exchange factor),
ADAP, SKAP-55 (adaptor protein), and Rap1 (small GTPase) 34,37-40. Protein kinase C
(PKC) activation by phorbol esters also can trigger integrin clustering by interacting
with the β2 subunit 34,40. Furthermore, integrin clustering can be induced by elevated
cytoplasmic calcium via TCR activation or treatment with calcium mobilizers such as
ionomycin and thapsigargin (TG) 41. Calcium-mediated LFA-1 activation also requires
the calcium-dependent protease calpain 41. Interestingly, the HTLV-1 accessory protein
p12I mimics the function of TG and induces calcium release from the ER stores to
activate T-cells 29.
Herein, we tested if the expression of HTLV-1 p12I could activate LFA-1-
mediated T-cell adhesion in a calcium-dependent manner. LFA-1-mediated adhesion of
wild type HTLV-1 (ACH) T-cells was greater than T-cells expressing a proviral clone
lacking p12I (ACH.p12). Expression of p12I in Jurkat T-cells also enhanced LFA-1-
mediated cell adhesion, which was inhibited by the calcium chelator BAPTA-AM, the
calcium channel blocker SK&F 96365 and calpeptin, an inhibitor of the calcium-

114
dependent protease calpain. Similar to the effects of TG, the expression of p12I in
Jurkat T-cells induced cell surface clustering of LFA-1 without changing LFA-1
expression. Our data indicate that p12I induces LFA-1 clustering on T-cells via
calcium-dependent signaling, which would promote spread of HTLV-1 to target T-cells.
3.2 MATERIALS AND METHODS
Cell lines
ACH.2 and ACH.p12.4 cell lines were generated from the outgrowth of
immortalized PBMC previously transfected with molecular clones of HTLV-1 (ACH
and ACH.p12I) 29,42. The ACH.p12I plasmid has a mutation resulting in complete
ablation of p12I mRNA expression without affecting the expression of other viral genes
26. ACH.2p and ACH.p12.4p cells were infected cell lines created by co-culturing naïve
human PBMC with lethally γ-irradiated (10,000 rads) ACH.2 and ACH.p12.4 cells,
respectively. Target PBMC were pre-activated for four days with human IL-2
(10 U/ml) and phytohemagglutinin (PHA) (2 µg/ml). ACH.2, ACH.p12.4, ACH.2p and
ACH.p12.4p cells were maintained in RPMI 1640 supplemented with 15% fetal bovine
serum, L-glutamine (0.3 mg/ml), penicillin (100 U/ml), streptomycin (100 µg/ml), and
recombinant IL-2 (10 U/ml). Viral p19 antigen production from the ACH.2, ACH.p12.4,
ACH.2p and ACH.p12.4p cell lines was measured in cell culture supernatants by
enzyme-linked immunosorbent assay (ELISA) in quadruplicate samples (Zeptomatrix)
according to the manufacturer's protocol. Jurkat T-cells (clone E6-1; American Type
Culture Collection catalog number TIB-152) were maintained in RPMI 1640

115
(Invitrogen) supplemented with 10 % fetal bovine serum, 100 µg of streptomycin-
penicillin/ml, and 2 mM L-glutamine. For lentivirus production, the 293T cell line
(American Type Culture Collection), which stably expresses the simian virus 40 (SV40)
T antigen, was maintained in Dulbecco’s modified Eagle medium (DMEM, Invitrogen)
supplemented with 10% fetal bovine serum (FBS), 100 µg/ml streptomycin/penicillin
plus 2 mM L-glutamine.
Recombinant lentivirus production and infection of Jurkat T-cells
Recombinant lentiviruses were prepared as previously described 21. pWPT-
IRES-GFP (empty plasmid) was generated by cloning an internal ribosome entry site
(IRES) sequence (pHR’CMV/Tax1/eGFP, G. Feuer, SUNY, Syracuse, NY) into
pWPT-GFP (D. Trono, University of Geneva, Geneva, Switzerland). pWPT-p12IHA-
IRES-GFP was generated by cloning p12IHA from pME-p12I (G. Franchini, National
Cancer Institute, National Institutes of Health) into pWPT-IRES-GFP. To generate
recombinant virus, 293T cells (5 x 106) were transfected with 2 µg of pHCMV-G, 10 µg
of pCMV∆R8.2 and 10 µg of pWPT-p12IHA-IRES-GFP or pWPT-IRES-GFP using
calcium phosphate. Supernatants were collected at 24, 48 and 72 h post transfection,
and filtered through a 0.2 µm filter. To obtain viral pellets, supernatants were
centrifuged at 6,500 xg for 16 h at 4° C, and then pellets were suspended in DMEM
medium overnight at 4° C. The concentrated virus was aliquotted and stored at -80°C.
After determination of viral titer, Jurkat T-cells were infected with recombinant virus at
a multiplicity of infection of 3 in the presence of 8 µg/ml polybrene (Sigma) and spin-
infected at 2700 rpm for 1 h at 25° C. Expression of eGFP and p12I was confirmed by

116
flow cytometry, western blot or reverse transcription- polymerase chain reaction (RT-
PCR) 21. For detection of p12I mRNA, cellular RNA was isolated using RNAqueous
(Ambion). RNA was converted to cDNA using the Reverse Transcription system
(Promega) and was amplified with AmpliTaq DNA polymerase (Perkin Elmer) using
CCTCTTTCTCCCGCTCTTTT (forward) and GGCCAAGCTAGCGTAATCTG
(reverse) primers. Transduced Jurkat T-cells (control and p12I expressing cells) were
used for experiments at 10 to 60 days post infection when expression of eGFP was
greater than 85% as determined by flow cytometric analysis.
Cell adhesion to immobilized ICAM-1
Cell-adhesion assays were performed as described elsewhere 41,43. Briefly,
recombinant human ICAM-1/Fc chimera (R&D system) in phosphate-buffered saline
(PBS) (0.5ug/well) was used for coating 96-well tissue culture plates overnight at 4° C.
Nonspecific binding was blocked with bovine serum albumin (BSA) in PBS for 1 h at
room temperature followed by three washes in PBS and once wash with RPMI 1640 or
HEPES buffer (20 mM HEPES, 140 mM NaCl, 2 mg/ml glucose, pH 7.4) for
MgCl2/EGTA stimulation. Cells (2 x 105 in 50 µl of RPMI 1640 or HEPES buffer)
were added to each well in the presence or absence of 50 µl of integrin stimulatory or
inhibitory reagents (2 x final concentrations) as indicated in results and figure legends.
Ionomycin (Sigma), TG (Calbiochem), phorbol 12-myristate 13-acetate (PMA, Sigma)
and MgCl2/EGTA were used as stimuli to test LFA-1-mediated adhesion. The
intracellular calcium chelator BAPTA-AM (Invitrogen), calcium release-activated
calcium (CRAC) channel blocker SK&F 96365 (Calbiochem) and calpain inhibitor

117
calpeptin (Calbiochem) were used for LFA-1 inhibition. The plates were incubated on
ice for 20 min, were centrifuged for one min at 30 ×g and incubated for 30 min at 37°
C. The plates were washed carefully with pre-warmed RPMI 1640 or HEPES buffer
three times. The percentage of attached cells was calculated by viable cell staining with
tetrazolium dye (CellTiter 96 Cell Proliferation Assay, Promega). Statistical differences
between groups were determined using the Wilcoxon- Mann Whitney test because we
assumed that differences between the two groups were not normally distributed.
P < 0.05 was considered significant.
Flow cytometry and soluble ICAM-1Fc binding assay
Flow cytometric analysis of the expression of cell surface markers CD11a and
CD18 was performed as described 42. Red-phycoerythrin (R-PE)-conjugated anti-
CD11a and FITC-conjugated anti-CD18 antibodies were used according to the
manufacturer's recommendations (Southern Biotechnology). For soluble ICAM-1 Fc
(sICAM-1) binding assay, cells were washed with RPMI 1640 or HEPES buffer and
resuspended into 50 µl of stimulator (2 x final concentrations) containing media or
HEPES buffer. Cells were mixed with 50 µl of recombinant human ICAM-1 Fc
chimera (R&D System) at a final concentration 5 µg/ml and incubated for 30 min at 37°
C. Subsequently, the cells were washed twice in ice-cold 0.2 % BSA and 0.2 % sodium
azide in PBS and incubated with R-PE-conjugated goat anti-human IgG Fc-specific
antibody (Jackson Immunoresearch Labs) for 20 min on ice. Cells were washed twice
in ice-cold 0.2 % BSA and 0.2 % sodium azide in PBS and fixed in 1 %

118
paraformaldehyde. Fluorescence of cells was measured by FACScan (Becton
Dickinson) analysis.
Confocal microscopy to detect LFA-1 distribution
For detection of LFA-1 distribution on the surface of T-cells by confocal
microscopy, Jurkat T-cells were attached to cover slips coated with a 0.01 % solution of
poly-L-lysine (Sigma). Cells were incubated for 30 min at 37° C with or without
stimulation and then were fixed with 0.5 % paraformaldehyde for 10 min. After
washing once with PBS and three times with RPMI 1640, cells were stained with R-PE-
conjugated CD11a antibody (Southern Biotechnology) for 30 min at 37° C, washed
once with RPMI 1640, mounted and analyzed using the Leica TCS SP2 AOBS confocal
system (Leica, Heidelberg, Germany). Leica Confocal Software (Leica Lite) was used
to quantify the fluorescence signal on cell membranes representing LFA-1 clustering as
described previously 41. Student’s t test was used for statistical analysis of differences
between two groups.
3.3 RESULTS
LFA-1-mediated adhesion in HTLV-1 immortalized T-cells lacking p12I expression
To examine the role of p12I in LFA-1-mediated cell adhesion, we first carried
out adhesion assays using wild type HTLV-1 (ACH cell line; ACH.2 and ACH.2p) and
T-cells lacking p12I expression (ACH. p12I; ACH.p12.4 and ACH.p12.4p). We
generated newly infected ACH.2p and ACH.p12.4p cells by co-culturing pre-activated

119
naïve human PBMC with lethally irradiated ACH.2 and ACH.p12.4, respectively, to
rule out the influence of specific clonal effects during in vitro culture. Virus replication
measured by viral p19 antigen ELISA, was similar among ACH.2, ACH.p12.4, ACH.2p
and ACH.p12.4p (Fig. 3.1A). These results are consistent with previous studies that
indicate that p12I is not required for HTLV-1 replication in activated T-cells or
immortalized T-cell lines 26,44.
LFA-1-mediated T-cell binding can result either from increased affinity for
ICAM-1 or clustering of LFA-1 on the cell surface 32-35. LFA-1 clustering can be
activated by TCR signaling, phorbol ester stimulation or increased intracellular calcium
levels 32-35. Thapsigargin and PMA-induced LFA-1-mediated adhesion of the wild type
ACH cell lines, ACH.2 and ACH.2p, was two-fold greater compared to unstimulated
cells (Fig. 3.1B). T-cells lacking p12I expression (ACH.p12.4 and ACH.p12.4p) did not
respond to the LFA-1 clustering reagents thapsigargin or PMA (Fig. 3.1B).
Mg2+/EGTA treatment, which resulted in an increase of LFA-1 affinity in vitro,
increased ACH.2 and ACH.2p cell adhesion up to 50-60%, but did not induce
significant adhesion in ACH.p12.4 and ACH.p12.4p cells (Fig. 3.1B). The failure of
ACH.p12.4 and ACH.p12.4p cells to respond to reagents that cause LFA-1 cluster and
enhance T-cell adhesion could be explained by these cells having either lowered LFA-1
expression or decreased binding affinity.
ACH.p12I cells express LFA-1, but have decreased binding of sICAM-1
To examine if lowered LFA-1 mediated adhesion in ACH.p12 cells was due to
changes in LFA-1 expression, we performed flow cytometric analysis using anti-CD11a

120
and anti-CD18 antibodies. There was no significant difference in integrin expression on
the cell surface among ACH.2, ACH.2p, ACH.p12.4 and ACH.p12.4p (Fig. 3.2A).
T-cell binding to sICAM-1 is increased only when LFA-1 affinity is enhanced
by agonists such as Mg2+-stimulation. In contrast, LFA-1 clustering on T-cells is
stimulated by reagents that stimulate calcium release from cell stores, such as TG or
reagents that induce TCR signaling, such as PMA 41,43. We therefore tested ACH wild
type and ACH.12 mutant T-cells for their sICAM-1 binding properties using flow
cytometry to measure LFA-1 affinity. As expected, none of the clustering stimulants
(TG and PMA) exhibited any induction of sICAM-1 binding in any of the cell lines (Fig.
3.2B). Mg2 -stimulated wild type ACH, ACH.2 and ACH.2p cells had enhanced
sICAM-1 binding, when compared to the ACH.p12I cell lines, ACH.p12.4 and
ACH.p12.4p (Fig. 3.2B). These data were consistent with our Mg2+/EGTA experiments
using immobilized ICAM-1 (Fig. 3.1B).
p12I expression in Jurkat T-cells did not affect LFA-1 affinity
To test whether HTLV-1 p12I expression specifically influenced LFA-1-
mediated T-cell adhesion, we stably expressed p12I in Jurkat T-cells using recombinant
lentiviruses (21). All empty control and p12I-expressing Jurkat T-cells were more than
85% GFP positive (Fig. 3.3A). Expression of HTLV-1 p12I also was confirmed by p12I
mRNA amplification by RT-PCR (Fig. 3.3B) and immunoprecipitation using polyclonal
HA-1 antibody (data not shown).

121
sICAM binding was tested using both empty control and p12I Jurkat T-cells. As
expected, TG and PMA, which activate LFA-1 clustering, did not induce sICAM-1
binding (Fig. 3.3A). Mg2+/EGTA treatment, which stimulates LFA-1 affinity, was used
to compare sICAM-1 binding in both empty control and p12I Jurkat T-cells. Our data
indicated no significant difference in LFA-1 affinity between the two cell types (Fig.
3.3A). These data demonstrated that p12I expression does not affect LFA-1 affinity.
Expression of p12I in Jurkat T cells induced LFA-1 clustering
Calcium mobilizers, such as ionomycin and TG enhance LFA-1-mediated T-cell
adhesion by affecting LFA-1 clustering 34,41. Since HTLV-1 p12I functionally mimics
TG by increasing intracellular calcium release from ER stores, we tested if p12I
expression might affect LFA-1 clustering. Jurkat T-cells that stably expressed p12I
exhibited enhanced cell adhesion to ICAM-1 molecules compared to empty vector-
transduced control Jurkat T-cells in all untreated, TG, ionomycin, PMA and
Mg2+/EGTA treatment groups (Fig. 3.4A). Since p12I did not affect LFA-1 affinity (Fig.
3.3), the enhancement observed in Mg2+/EGTA treated p12I Jurkat T-cell likely was due
to pre-clustered LFA-1 in p12I-expressing cells.
To clarify whether p12I expression affected clustering or conformational
changes of LFA-1, we measured the fold increase of cell adhesion over untreated cells
in the empty control and p12I Jurkat T-cells that were treated with increasing
concentrations of TG, ionomycin, PMA or Mg2+(Fig. 3.4B, C, D and E). As expected,
the calcium mobilizers TG and ionomycin induced increased cell adhesion in a dose-
dependent manner at lower concentrations and led to decreased cell adhesion at higher

122
concentrations in both cell types (Fig. 3.4B and C). The peak adhesion by TG and
ionomycin was observed between 2.5 to 5 µM and 0.5 to 1 µM, respectively, similar to
a previous report 41. Another LFA-1 clustering agent, PMA, also induced a dose-
dependent increase in adhesion to ICAM-1 at lower concentrations, but adhesion was
saturated at higher concentrations in both cell types (Fig. 3.4D). At the lower
concentrations, p12I Jurkat T-cell adhesion was higher than that of empty control Jurkat
T-cells after TG, ionomycin, or PMA stimulation (Fig. 3.4B, C, and D), suggesting that
p12I expression on Jurkat T-cells reduced the threshold required for the activation of
LFA-1 clustering.
As expected, Mg2+ also induced a dose-dependent increase and saturation of cell
adhesion in both cell types, however, the fold increase over untreated cells was not
significantly different between empty control and p12I Jurkat T-cells (Fig. 3.4E). The
affinity of both cell types was increased to a similar level by Mg2+, suggesting that
expression of p12I did not affect LFA-1 affinity. These data indicated that enhanced
LFA-1-mediated cell adhesion in p12I Jurkat T-cells was due to enhanced LFA-1
clustering rather than increased affinity.
HTLV-1 p12I - mediated LFA-1 activation is calcium dependent
LFA-1 clustering requires signaling pathways leading to the elevation of
calcium and activation of calpain, a calcium-dependent protease 41. To determine
whether LFA-1-activation induced by p12I was dependent on calcium-mediated
signaling pathways, we performed cell adhesion assays using inhibitors such as the
intracellular calcium chelator BAPTA-AM, the calcium channel blocker SK&F 96365,

123
and the calpain inhibitor calpeptin (Fig. 3.5). As expected, all inhibitors abolished TG
and PMA-induced cell adhesion in both cell types (Fig. 3.5). These results suggested
that LFA-1 clustering, due to p12I, was dependent on both intracellular and extracellular
calcium, as well as calpain. In contrast to TG and PMA stimulation, cell adhesion
induced by Mg2+/EGTA was not blocked completely by the inhibitors (Fig. 3.5). These
data were consistent with a previous report that Mg2+-induced affinity increases were
not altered by calcium channel blockers or calpain inhibitors 41.
HTLV-1 p12I modulated surface distribution of LFA-1
To confirm if the membrane distribution of LFA-1 was altered in p12I-
expressing Jurkat T-cells, we measured LFA-1 expression by confocal microscopy
using R-PE-conjugated CD11a. Untransduced mock Jurkat T-cells were either
untreated or treated with 1 µM of TG as a control experiment. A higher intensity of
LFA-1 fluorescence was observed frequently in TG-treated cells (Fig. 3.6A). Similar to
TG treatment, p12I expression in Jurkat T-cells had more than a two-fold enhanced
fluorescence indicating increased LFA-1 clustering on the cell membranes (Fig. 3.6B).
Increased LFA-1 fluorescence in p12I expressing Jurkat T cells was not due to increased
cell surface expression of LFA-1. Cell surface expression of CD11a was measured by
flow cytometry and was not significantly different between empty control and p12I
expressing Jurkat T cells (Fig. 3.6C). These data indicated that p12I expression altered
LFA-1 cell surface distribution favoring clustering of the adhesion molecule without
changing total LFA-1 cell surface expression and were consistent with our cell adhesion
and sICAM-1 binding assays.

124
3.4 DISCUSSION
In this study, we found that HTLV-1 p12I expression in T-cells altered LFA-1-
mediated T-cell adhesion to ICAM-1. Abrogation of p12I expression in HTLV-1
infected cells (ACH.p12I) resulted in decreased LFA-1-mediated cell adhesion, which
was due to a reduced affinity of LFA-1 compared wild type HTLV-1-infected cells. To
identify the specific role of p12I in LFA-1-mediated cell adhesion, we stably expressed
p12I in Jurkat T-cells using lentiviral vectors. Expression of p12I increased LFA-1-
mediated cell adhesion to ICAM-1 by inducing clustering of LFA-1 on Jurkat T-cells.
Our data are consistent with previous reports from our group and others 26-28,45 that
demonstrated the important role of HTLV-1 p12I in T-cell activation and cell-to-cell
transmission during the early stages of HTLV-1 infection.
An important role of LFA-1 and ICAM-1 in HTLV-1 cell-to-cell transmission
was implicated in a recent study by Barnard et al 14. By cross-linking with antibodies to
cell surface molecules, they identified that LFA-1 and ICAM-1 were involved in
triggering microtubule-organizing center (MTOC) polarization, which is characteristic
of HTLV-1-induced virological synapses. Our data suggested that p12I may synergize
to modify cell adhesion and contribute to MTOC polarization by increasing cell
adhesion through clustered LFA-1. Interestingly, transmission of other viruses is also
enhanced by the LFA-1/ ICAM-1 interaction. For example, measles virus infection
induced increased LFA-1 expression on monocytes resulting in enhanced viral
transmission to endothelial cells 14,46. Furthermore, LFA-1 activation of T-cells

125
increased their susceptibility to human immunodeficiency virus type 1 (HIV-1)
infection 47,48.
Our results presented herein, support our previous findings, which demonstrated
that T-cells expressing ACH. p12I have reduced viral infectivity when co-cultured with
quiescent T-cells 27. ACH.p12I cells do not exhibit efficient cell-to-cell contact and
therefore are disadvantaged for cell-to-cell transmission. This tenet would explain, in
part, data indicating that HTLV-1 p12I is required for in vivo viral infectivity in rabbits
26. The early block in infection observed in this animal study could be explained by
inadequate LFA-1 activation on HTLV-1-infected cells, which would inhibit the ability
of HTLV-1 to spread to non-activated target cells. Our findings are also consistent
with studies that indicate that HTLV-1 infection is established by early viral spread to
T-cells, but subsequently maintained by clonal expansion of T-cells49-51. If the virus
lacks the ability to rapidly spread from infected cells to its targets early in the infection,
it is likely that HTLV-1 infection is eliminated by a robust immune response52.
The observed affinity reduction in ACH.p12I cells might have resulted from
alteration of inside-out signaling. Although signaling pathways that regulate LFA-1
affinity are incompletely defined compared to that of LFA-1 clustering modulation 34,
transient affinity increases of LFA-1 and α1β4 are induced by chemokine stimulation
53,53,54. In addition, the lymphocyte specific protein tyrosine kinase, Lck, has been
identified as a critical enzyme to maintain the high affinity status of integrin α1β4 55.
Interestingly, expression of Lck gene was down-regulated by the HTLV-1 accessory
protein p30II 56. Thus, a balance of HTLV-1 regulatory and accessory protein
expression is likely required to ensure cell-to-cell transmission of the virus.

126
HTLV-1 p12I expression in Jurkat T-cells induced LFA-1-mediated cell
adhesion in a calcium-dependent manner. p12I mimics TG, which is an LFA-1
clustering agent, by releasing calcium from ER stores and increasing intracellular
calcium concentration without activation of the TCR signaling pathway 29. We cannot
rule out the possibility that p12I may activate LFA-1-mediated adhesion via chemokine
secretion, which can increase affinity and induce clustering. We have reported that p12I
enhanced IL-2 production through NFAT activation 28,30,45. IL-2 produced by HTLV-1
infected cells may activate LFA-1 on both infected and target cells in an autocrine or
paracrine manner in the local tissue microenvironment in vivo 30,45. Our data presented
herein, together with these previous studies suggests that p12I expression in HTLV-1-
infected T-lymphocytes lowers the threshold of calcium signaling events to promote T-
cell activation and cell to cell transmission.
In summary, our data indicate that HTLV-1 p12I expression induced
LFA-1-mediated T-cell adhesion to ICAM-1 by activating LFA-1 clustering in a
calcium-dependent manner. Our data demonstrate that p12I enhances HTLV-1 cell-to-
cell transmission during the early stages of infection by activating LFA-1-mediated
adhesion and by supporting the formation of the HTLV-1 virological synapse.
3.5 REFERENCES
(1) Proietti FA, Carneiro-Proietti AB, Catalan-Soares BC, Murphy EL. Global epidemiology of HTLV-I infection and associated diseases. Oncogene. 2005;24:6058-6068.
(2) Mahieux R, Gessain A. HTLV-1 and associated adult T-cell leukemia/lymphoma. Rev Clin Exp Hematol. 2003;7:336-361.

127
(3) Franchini G, Fukumoto R, Fullen JR. T-cell control by human T-cell leukemia/lymphoma virus type 1. Int J Hematol. 2003;78:280-296.
(4) Kehn K, Berro R, de La FC et al. Mechanisms of HTLV-1 transformation. Front Biosci. 2004;9:2347-2372.
(5) Hall WW, Fujii M. Deregulation of cell-signaling pathways in HTLV-1 infection. Oncogene. 2005;24:5965-5975.
(6) Manns A, Hisada M, Lagrenade L. Human T-lymphotropic virus type I infection. Lancet. 1999;353:1951-1958.
(7) Weiss RA. Milk-borne transmission of HTLV-1. [Review]. Japanese Journal of Cancer Research. 1993;84(3):inside front cover.
(8) Bangham CR. The immune control and cell-to-cell spread of human T-lymphotropic virus type 1. J Gen Virol. 2003;84:3177-3189.
(9) Okochi K, Sato H, Hinuma Y. A retrospective study on transmission of adult T cell leukemia virus by blood transfusion: seroconversion in recipients. Vox Sang. 1984;46:245-253.
(10) Yamamoto N, Okada M, Koyanagi Y, Kannagi M, Hinuma Y. Transformation of human Leukocytes by cocultivation with an adult T cell leukemia virus producer cell line. Science. 1982;217:737-739.
(11) Popovic M, Sarin PS, Robert-Gurroff M et al. Isolation and transmission of human retrovirus (human t-cell leukemia virus). Science. 1983;219:856-859.
(12) Dustin M. Viral spread through protoplasmic kiss. Nat Cell Biol. 2003;5:271-272.
(13) Igakura T, Stinchcombe JC, Goon PK et al. Spread of HTLV-I between lymphocytes by virus-induced polarization of the cytoskeleton. Science. 2003;299:1713-1716.
(14) Barnard AL, Igakura T, Tanaka Y, Taylor GP, Bangham CR. Engagement of specific T cell surface molecules regulates cytoskeletal polarization in HTLV-1-infected lymphocytes. Blood. 2005.
(15) Nejmeddine M, Barnard AL, Tanaka Y, Taylor GP, Bangham CR. Human T-lymphotropic Virus, Type 1, Tax Protein Triggers Microtubule Reorientation in the Virological Synapse. J Biol Chem. 2005;280:29653-29660.
(16) Pise-Masison CA, Jeong SJ, Brady JN. Human T cell leukemia virus type 1: the role of Tax in leukemogenesis. Arch Immunol Ther Exp (Warsz ). 2005;53:283-296.

128
(17) Manel N, Battini JL, Taylor N, Sitbon M. HTLV-1 tropism and envelope receptor. Oncogene. 2005;24:6016-6025.
(18) Bindhu M, Nair A, Lairmore MD. Role of accessory proteins of HTLV-1 in viral replication, T cell activation, and cellular gene expression. Front Biosci. 2005;10:17-37.
(19) Franchini G, Nicot C, Johnson JM. Seizing of T cells by human T-cell leukemia/lymphoma virus type 1. Adv Cancer Res. 2003;89:69-132.
(20) Nicot C, Harrod RL, Ciminale V, Franchini G. Human T-cell leukemia/lymphoma virus type 1 nonstructural genes and their functions. Oncogene. 2005;24:6026-6034.
(21) Nair A, Michael B, Hiraragi H et al. Human T lymphotropic virus type 1 accessory protein p12I modulates calcium-mediated cellular gene expression and enhances p300 expression in T lymphocytes. AIDS Res Hum Retroviruses. 2005;21:273-284.
(22) Hiraragi H, Michael B, Nair A et al. Human T-lymphotropic virus type 1 mitochondrion-localizing protein p13II sensitizes Jurkat T cells to Ras-mediated apoptosis. J Virol. 2005;79:9449-9457.
(23) D'Agostino DM, Silic-Benussi M, Hiraragi H, Lairmore MD, Ciminale V. The human T-cell leukemia virus type 1 p13(II) protein: effects on mitochondrial function and cell growth. Cell Death Differ. 2005.
(24) Dekaban GA, Peters AA, Mulloy JC et al. The HTLV-I orfI protein is recognized by serum antibodies from naturally infected humans and experimentally infected rabbits. Virology. 2000;274:86-93.
(25) Pique C, Uretavidal A, Gessain A et al. Evidence for the chronic in vivo production of human T cell leukemia virus type I Rof and Tof proteins from cytotoxic T lymphocytes directed against viral peptides. J Exp Med. 2000;191:567-572.
(26) Collins ND, Newbound GC, Albrecht B et al. Selective ablation of human T-cell lymphotropic virus type 1 p12I reduces viral infectivity in vivo. Blood. 1998;91:4701-4707.
(27) Albrecht B, Collins ND, Burniston MT et al. Human T-lymphotropic virus type 1 open reading frame I p12(I) is required for efficient viral infectivity in primary lymphocytes. J Virol. 2000;74:9828-9835.

129
(28) Albrecht B, D'Souza CD, Ding W et al. Activation of nuclear factor of activated T cells by human T- lymphotropic virus type 1 accessory protein p12(I). J Virol. 2002;76:3493-3501.
(29) Ding W, Albrecht B., Kelley RE et al. Human T lymphotropic virus type 1 p12(I) expression increases cytoplasmic calcium to enhance the activation of nuclear factor of activated T cells. J Virol. 2002;76:10374-10382.
(30) Ding W, Kim SJ, Nair AM et al. Human T-Cell Lymphotropic Virus Type 1 p12(I) Enhances Interleukin-2 Production during T-Cell Activation. J Virol. 2003;77:11027-11039.
(31) Dustin ML, Bivona TG, Philips MR. Membranes as messengers in T cell adhesion signaling. Nat Immunol. 2004;5:363-372.
(32) Hogg N, Henderson R, Leitinger B et al. Mechanisms contributing to the activity of integrins on leukocytes. Immunol Rev. 2002;186:164-171.
(33) van KY, Figdor CG. Activation and inactivation of adhesion molecules. Curr Top Microbiol Immunol. 1993;184:235-256.
(34) Hogg N, Laschinger M, Giles K, Mcdowall A. T-cell integrins: more than just sticking points. J Cell Sci. 2003;116:4695-4705.
(35) Hogg N, Smith A, Mcdowall A et al. How T cells use LFA-1 to attach and migrate. Immunol Lett. 2004;92:51-54.
(36) Carman CV, Springer TA. Integrin avidity regulation: are changes in affinity and conformation underemphasized? Curr Opin Cell Biol. 2003;15:547-556.
(37) Ardouin L, Bracke M, Mathiot A et al. Vav1 transduces TCR signals required for LFA-1 function and cell polarization at the immunological synapse. Eur J Immunol. 2003;33:790-797.
(38) Griffiths EK, Krawczyk C, Kong YY et al. Positive regulation of T cell activation and integrin adhesion by the adapter Fyb/Slap. Science. 2001;293:2260-2263.
(39) Wang H, Moon EY, Azouz A et al. SKAP-55 regulates integrin adhesion and formation of T cell-APC conjugates. Nat Immunol. 2003;4:366-374.
(40) Katagiri K, Hattori M, Minato N et al. Rap1 is a potent activation signal for leukocyte function-associated antigen 1 distinct from protein kinase C and phosphatidylinositol-3-OH kinase. Mol Cell Biol. 2000;20:1956-1969.

130
(41) Stewart MP, Mcdowall A, Hogg N. LFA-1-mediated adhesion is regulated by cytoskeletal restraint and by a Ca2+-dependent protease, calpain. J Cell Biol. 1998;140:699-707.
(42) Collins ND, D'Souza C, Albrecht B et al. Proliferation response to interleukin-2 and Jak/Stat activation of T cells immortalized by human T-cell lymphotropic virus type 1 is independent of open reading frame I expression. J Virol. 1999;73:9642-9649.
(43) Stewart MP, Cabanas C, Hogg N. T cell adhesion to intercellular adhesion molecule-1 (ICAM-1) is controlled by cell spreading and the activation of integrin LFA-1. J Immunol. 1996;156:1810-1817.
(44) Robek MD, Wong FH, Ratner L. Human T-Cell leukemia virus type 1 pX-I and pX-II open reading frames are dispensable for the immortalization of primary lymphocytes. J Virol. 1998;72:4458-4462.
(45) Nicot C, Mulloy JC, Ferrari MG et al. HTLV-1 p12(I) protein enhances STAT5 activation and decreases the interleukin-2 requirement for proliferation of primary human peripheral blood mononuclear cells. Blood. 2001;98:823-829.
(46) Hummel KB, Bellini WJ, Offermann MK. Strain-specific differences in LFA-1 induction on measles virus-infected monocytes and adhesion and viral transmission to endothelial cells. J Virol. 1998;72:8403-8407.
(47) Fortin JF, Cantin R, Tremblay MJ. T cells expressing activated LFA-1 are more susceptible to infection with human immunodeficiency virus type 1 particles bearing host-encoded ICAM-1. J Virol. 1998;72:2105-2112.
(48) Arias RA, Munoz LD, Munoz-Fernandez MA. Transmission of HIV-1 infection between trophoblast placental cells and T-cells take place via an LFA-1-mediated cell to cell contact. Virology. 2003;307:266-277.
(49) Wattel E, Vartanian JP, Pannetier C, Wainhobson S. Clonal expansion of human T-Cell leukemia virus type I- infected cells in asymptomatic and symptomatic carriers without malignancy. J Virol. 1995;69:2863-2868.
(50) Cavrois M, Leclercq I, Gout O et al. Persistent oligoclonal expansion of human T-cell leukemia virus type 1 infected circulating cells in patients with Tropical spastic paraparesis/HTLV-1 associated myelopathy. Oncogene. 1998;17:77-82.
(51) Mortreux F, Kazanji M, Gabet AS, de Thoisy B, Wattel E. Two-step nature of human T-cell leukemia virus type 1 replication in experimentally infected squirrel monkeys (Saimiri sciureus). J Virol. 2001;75:1083-1089.

131
(52) Mosley AJ, Asquith B, Bangham CR. Cell-mediated immune response to human T-lymphotropic virus type I. Viral Immunol. 2005;18:293-305.
(53) Constantin G, Majeed M, Giagulli C et al. Chemokines trigger immediate beta2 integrin affinity and mobility changes: differential regulation and roles in lymphocyte arrest under flow. Immunity. 2000;13:759-769.
(54) Chan JR, Hyduk SJ, Cybulsky MI. Chemoattractants induce a rapid and transient upregulation of monocyte alpha4 integrin affinity for vascular cell adhesion molecule 1 which mediates arrest: an early step in the process of emigration. J Exp Med. 2001;193:1149-1158.
(55) Feigelson SW, Grabovsky V, Winter E et al. The Src kinase p56(lck) up-regulates VLA-4 integrin affinity. Implications for rapid spontaneous and chemokine-triggered T cell adhesion to VCAM-1 and fibronectin. J Biol Chem. 2001;276:13891-13901.
(56) Michael B, Nair AM, Hiraragi H et al. Human T lymphotropic virus type-1 p30II alters cellular gene expression to selectively enhance signaling pathways that activate T lymphocytes. Retrovirology. 2004;1:39.

132
Figure 3.1. LFA-1 mediated adhesion is reduced in ACH.p12I cell lines.
(A) p19 antigen production from ACH.2, ACH.p12.4, ACH.2p and ACH.p12.4p cell
lines was measured using ELISA. Supernatants were collected 24 days post-infection.
Values are the means (± SEM) of triplicate samples and represent two independent
experiments. (B) Cells were treated with TG (5 µM), PMA(50 ng/ml) or
Mg2+/EGTA(5 mM/1 mM) and incubated on ICAM-1Fc-coated 96-well plates for
30 min at 37° C. The proportion of adhesion was measured as a percentage of total cells
added per well. Values are the means (± SEM) of triplicate sample and represent four
independent experiments. Significantly reduced adhesion was observed in TG, PMA
and Mg2+/EGTA treated ACH.p12.4 and ACH.p12.4p cell lines (Wilcoxon- Mann
Whitney test, P < 0.05).

133
Figure 3.2. ACH.p12I cells have decreased binding of sICAM-1 without alteration
of LFA-1 expression.
(A) Flow cytometry specific for LFA-1 molecules CD11a and CD18 was performed on
ACH.2, ACH.p12.4, ACH.2p and ACH.p12.4p (106cells per sample). (B) Soluble
ICAM-1Fc binding assay was performed on ACH.2, ACH.p12.4, ACH.2p and
ACH.p12.4p cell lines. Cells were treated with TG (5 uM), PMA(50 ng/ml) or
Mg2+/EGTA(5 mM/1 mM) and incubated with sICAM-1Fc for 30 min at 37° C. Bound
sICAM-1 was detected with R-PE-conjugated goat anti-human IgG Fc-specific
antibody and analyzed by flow cytometry. Data are expressed as mean fluorescence
intensity. Figure is representative of two experiments.

134
Figure 3.3. p12I stable expression in Jurkat T-cells did not alter LFA-1 affinity.
(A) Flow cytometric analysis for both GFP expression and sICAM-1 binding was
performed in empty control and p12I-expressing Jurkat T-cells. Both samples were
more than 85% GFP positive indicating high transduction efficiency. Soluble ICAM-
1Fc binding was measured concurrently. Cells were treated with TG (5 µM), PMA(50
ng/ml) or Mg2+/EGTA(5 mM/1 mM) and incubated with sICAM-1Fc for 30 min at 37°
C. Bound sICAM-1 was detected with R-PE-conjugated goat anti-human IgG Fc-
specific antibody and analyzed by flow cytometry. (B) RT-PCR was performed using
total cellular RNA isolated from mock, empty control and p12I-expressing Jurkat T-
cells at 30 and 60 days post-infection with lentiviral vectors.

135
Figure 3.4. Expression of p12I in Jurkat T-cells induced LFA-1-mediated cell
adhesion. Adhesion to immobilized ICAM-1 was performed using empty control and
p12I expressing Jurkat T-cells. (A) Cells were treated with TG (5 µM), Ionomycin (0.5
µM) PMA (50 ng/ml) or Mg2+/EGTA (5 mM/1 mM) and incubated on ICAM-1Fc-
coated for 30 min at 37° C. The adhesion was measured as a percentage of total cells
added per well. Values are the means (± SEM) of triplicate samples and represent three
independent experiments. P < 0.05 was by Wilann-Whitney test, (B-E) Cells were
treated with increasing concentrations of (B) TG, (C) Ionomycin, (D) PMA, and (E)
Mg2+ (with constant 1 mM EGTA concentration) and incubated for the adhesion assay.
The fold increase was calculated by dividing the percentages of adhered cells in the
presence of each treatment with that of untreated cell adhesion. Figure is representative
of two experiments.

136
Figure 3.5. HTLV-1 p12I-mediated LFA-1 activation is inhibited by calcium signal
inhibitors. Empty control and p12I-expressing Jurkat T-cells were pretreated with
BAPTA-AM (50 µM), SK&F 96365 (100 µM) or calpeptin (100 µg/ml) for 30 min at
37° C and adhesion assay was performed in presence or absence of Thapsigargin (5
µM), PMA (50 ng/ml) or Mg2+ /EGTA (5 mM/1 mM) stimulation.

137
Figure 3.6. Expression of p12I in Jurkat T-cells modulated surface distribution of
LFA-1 on the cell membrane. (A) Untransduced mock Jurkat T cells or (B)
transduced empty control and p12I expressing Jurkat T cells were stained with R-PE
conjugated LFA-1 mAb and analyzed by confocal microscopy. Fluorescent signal
exceeding 50 unit was expressed as bright blue color. Signal of LFA-1 was quantified
on the membrane of 30 cells. Mean fluorescence units (± SEM) were compared in right
bar graphs (A) between TG untreated and treated groups, or (B) between empty control
and p12I expressing Jurkat T cells. Significant (t test P < 0.01) increased fluorescence in
TG treated and p12I expressing Jurkat T cells was observed. C) Flow cytometric
analysis for LFA-1 cell surface expression in GFP expressing empty control and in
empty control and p12I expressing Jurkat T cells was performed.

138
CHAPTER 4
EXPRESSION OF HTLV-1 PX OPEN READING FRAME I ENHANCES
EARLY VIRAL INFECTIVITY DURING CELL-TO-CELL
TRANSMISSION IN T-LYMPHOCYTES
4.1 INTRODUCTION
Human T-cell lymphotropic virus type 1 (HTLV-1), which infects 15 to 25
million people worldwide 1, is the etiological agent of adult T cell leukemia/lymphoma
(ATLL), and is highly associated with HTLV-1-associated myelopathy/tropical spastic
paraparesis (HAM/TSP) and a variety immune-mediated disorders 2-4. In addition to
retroviral common structural and enzymatic proteins (Gag, Pol, and Env), HTLV-1
genome encodes, regulatory (Tax and Rex) and nonstructural accessory proteins (p12I,
p27I, p30II and p13II) 5-9, which are generated by alternative splicing and internal
initiation, from four open reading frames (ORF) located in pX region.

139
Both p27I (doubly spliced) and p121 (singly spliced) are produced from pX ORF
I. p12I can also be translated from the p27I message via initiation at an internal
methionine codon. It is thought that p12I is preferentially expressed from the p27I
message 10. Importantly, the expression of pX ORF I proteins during natural infection is
evidenced by detection of its mRNA in infected cells and elicitation of humoral and
cytotoxic T cell responses against pX ORF I-encoded proteins in infected carriers and
patients 11,12. Moreover, in chronically HTLV-1 infected cell lines, the expression of pX
ORF I mRNA was measured at 100 to 1,000-fold less than pX ORF III/IV mRNA by
real time reverse transcription-polymerase chain reaction (RT-PCR), suggesting that
expression of HTLV-1 pX genes are differentially regulated 13. However, regulation of
pX mRNA expression was not tested during early phase of HTLV-1 infection. The
expression of mRNA can be modulated by specific stimuli resulted from cell-to-cell
contact between HTLV-1 infected cells and target cells. Furthermore, events during de
novo HTLV-1 infection in target cells may affect expression pattern of pX gene
products.
HTLV-1 infection is naturally acquired following contact between target cells
and virus infected cells contained in breast milk, semen and blood 14,15. CD4 + T cells
are the major target of HTLV-1 infection, immortalization and transformation 16,17.
Mechanisms of HTLV-1 mediated immortalization and transformation have been
extensively investigated, however early events of HTLV-1 transmission and infection
are poorly understood. Activated T cells are more susceptible to HTLV-1 infection and
transformation compared to non-stimulated or quiescent/resting T cells 18, suggesting

140
that T cell activation is required for the virus to efficiently establish infection. HTLV-1
infection can further activate uninfected, resting T cells in an antigen-nonspecific
manner or stimulate antiviral T cells in an antigen-specific manner. Non-specific
proliferation and stimulation of uninfected PBMC can be induced by direct contact with
irradiated or fixed HTLV-1 infected cells through the combined interaction of adhesion
molecules CD2/LFA-3 (CD58), LFA-1/ICAM-1 (CD54), and IL-2/IL-2R autocrine
activity 4,19,20. CD2/CD58 interaction is particularly important for the activation of
uninfected T cells by HTLV-I-infected T cells 11.
Studies from our laboratory have implicated HTLV-1 pX ORF I expression in
HTLV-1 infection in a rabbit model and in T cell activation 21,22. Selective ablation of
pX ORF I mRNA expression from a HTLV-1 proviral clone (ACH.p12) resulted in
dramatic reduction of viral infectivity compared to wild type (ACH) in a rabbit model 23.
Furthermore, when cocultured with naïve, quiescent peripheral blood mononuclear cells
(PBMC), ACH.p12I cells exhibited reduced viral infectivity 24. However, in the
presence of the T cell stimulants, phytohemagglutinins (PHA) and IL-2, ACH.p12I
cells were restored in their ability to infect target T cells 24. These reports indicate that
pX ORF I expression is required for efficient HTLV-1 infection in PBMC and
suggesting a role for p12I in target T cell activation.
Ectopic expression of p12I causes calcium mediated T cell activation 25-27. The
small hydrophobic protein accumulates in the endoplasmic reticulum (ER) 28, where it
associates with calreticulin, calnexin and calcineurin 29, and leading to the release of
calcium from ER stores 26. As a result of HTLV-1 p12I expression, calcium-dependent

141
signaling causes activation and nuclear translocation of the nuclear factor of activated T
cells (NFAT) 25, and subsequent up-regulation of IL-2 production 27 and p300
expression 30. In addition, p12I expression leads to enhanced LFA-1 clustering on Jurkat
T cells, implying a role for pX ORF I in cell-to-cell transmission of HTLV-1 31.
Cell-to-cell HTLV-1 transmission occurs directly through virological synapse by
inducing polarization of microtubule-organizing centers (MTOC) to cell contact
junction 32-35. Engagement of adhesion molecules, ICAM-1, LFA-1, and IL-2 receptor
(CD25) triggered strong polarization of the MTOC, suggesting their critical role in
HTLV-1 transmission. HTLV-1 Tax is suggested as a viral factor contributing viral
cell-to-cell transmission, since Tax expression triggered MTOC polarization directly
and may synergize by up-regulating expression of ICAM-1 32 36.
In this study, we investigated the role of expression of HTLV-1 pX ORF I in
early HTLV-1 transmission by comparing the activation status between wild type
HTLV-1 infected cells (ACH) and T cells lacking pX ORF I expression (ACH.p12). We
then tested if the expression of ORF I is required to induce bystander activation of
uninfected target cells by performing both proliferation assays and flow cytometric
analysis of target cells We found that abrogation of Px ORF I message decreased CD69
expression in HTLV-1 infected cells, but did not affect the HTLV-1- induced bystander
target cell activation.
Finally, we performed real-time RT-PCR to detect pX ORF I and pX ORF III/IV
mRNA after coculture of both wild type ACH and ACH.p12 cells with pre-stimulated

142
or non-stimulated T cells. By measuring Tax/Rex (pX ORF III/IV) mRNA in target
cells, we were able to compare viral infectivity of wild type ACH to ACH.p12
immortalized T cells and viral mRNA expression in newly infected target cells. Our
data indicated that expression of pX ORF I is required for efficient HTLV-1
transmission to target cells and defined for the first time Tax/Rex and pX ORF I mRNA
expression in early cell-to-cell transmission.
4.2 MATERIALS AND METHODS
Cells
HTLV-1 infected cell lines, ACH.2, ACH.2p, ACH.p12.4 and ACH.p12.4p have
been previously described 23,31. ACH.2 and ACH.p12.4 cells were generated following
transfection of molecular clones of HTLV-1 (ACH and ACH.p12I) 23,37. ACH.p12I
plasmid contains a mutation in the splice acceptor of the third exon of the pX ORF I
DNA, resulting in complete ablation of pX ORF I expression without affecting the
expression of other viral genes 23,37. ACH.2p and ACH.p12.4p cells were created from
ACH.2 and ACH.p12.4 cells by coculturing PBMC isolated from same dono 31. ACH.2,
ACH.p12.4, ACH.2p and ACH.p12.4p cells were cultured in RPMI 1640 supplemented
with 15% fetal bovine serum, L-glutamine (0.3 mg/ml), penicillin (100 U/ml),
streptomycin (100 µg/ml), and recombinant IL-2 (20 U/ml). HTLV-1 infected human T
cell line, MT-2 cells were grown in RPMI 1640 supplemented with 10% fetal bovine
serum, L-glutamine (0.3 mg/ml), penicillin (100 U/ml), streptomycin (100 µg/ml).
Normal uninfected human PBMC were obtained by leukophoresis and maintained as

143
previously described 24. For coculture experiment, target PBMC were either
unstimulated or pre-activated for four days with human IL-2 (20 U/ml) and
phytohemagglutinin (PHA) (2 µg/ml).
Coculture of HTLV-1 infected cells and target PBMC
HTLV-1 infected effecter cells (ACH.2, ACH.2p, ACH.p12.4 and ACH.p12.4p)
were cocultivated as live or fixed cells with freshly isolated non-stimulated or pre-
activated PBMC. For fixation, cells were incubated 1% paraformaldehye (PFA) for 30
min and washed twice with media before coculture. Permeable transwell inserts with
diameter of 6.5 mm and pore size of 0.4 µm (Corning Inc.) were used to separately
culture live effecter cells from target PBMC and to test the role of soluble factors on
cell activation. Live, fixed or separated effecter cells were cocultured with non-
stimulated or activated PBMC at 1:2 or 1:1 ratio at 2 million cells/ml prior to
proliferation assay, flow cytometric analysis or RNA extraction for real time RT-PCR.
Proliferation assay
Tetrazolium dye (MTS) proliferation assay (CellTiter 96 Cell Proliferation
Assay, Promega) was used according to the manufacturer's instructions. To measure
proliferation of target PBMC cocultured with fixed HTLV-1 infected effecter cells, the
MTS assay was performed at the time of coculture (time 0) and at 3 days after coculture.
Data was expressed as fold proliferation of target cells, which was measured by optical
density in triplicate samples.

144
Flow cytometric analysis and cell sorting.
Flow cytometric analysis was performed as described 38 to measure expression
of cell surface markers. Red-phycoerythrin (R-PE) conjugated antibodies recognizes
CD25 (Southern Biotechnology), CD58 (Southern Biotechnology), and CD11a
(Southern Biotechnology) and FITC conjugated antibodies recognizes CD69 (BD
Bioscience), CD2 (Southern Biotechnology), and CD54 (Southern Biotechnology)
antibodies were used for cell surface staining according to the manufacturer's
recommendations. Fluorescence of cells was measured by FACScan (Becton
Dickinson) analysis.
For evaluating bystander target PBMC activation, target PBMC were pre-
labeled with 0.5 µM succinimidyl ester of carboxyfluorescein diacetate (CFDA-SE)
(Invitrogen) according to the manufacturer's instructions to differentiate from unlabeled
HTLV-1 infected effecter cells during flow cytometric analysis. After 24 and 36 h
coculture, cells were stained with R-PE conjugated antibodies recognizes CD25
(Southern Biotechnology) and CD69 (BD Bioscience). CD25 and CD69 percentage
positive were measured in the target PBMC (CMFD-SE positive cells).
For further real time RT-PCR analysis, cell sorting using a FACSAria flow
cytometer (Becton Dickinson) was performed to selectively collect target PBMC from
HTLV-1 infected cells (ACH.2p, and ACH.p12.4p) at 1 and 3 days after coculture.
Before cell sorting, we pre-labeled HTLV-1 infected cells with 0.5 µM 5-
chloromethylfluorescein diacetate (CMFDA) (Invitrogen) according to the
manufacturer's recommendation.

145
Real time RT-PCR
Total RNA was extracted from cells using RNAqueous Kit (Ambion) and
complementary DNA (cDNA) was generated from 1 µg of RNA using a Reverse
Transcription system kit (Promega) as described by manufacturer. Taqman real time
PCR was performed using a Prism 7000 sequence detector system (Applied
Biosystems). Primers and probes used for detection of HTLV-1 pX ORF III/IV and pX
ORF I mRNAs were previously described 13. Amplication of Tax/Rex cDNA were
performed using forward splice site-specific primer (ex2/6950) 5’-
ACCAACACCATGGCCCA-3’ and reverse specific primer (3’7170) 5’-
GAGTCGAGGGATAAGGAAC-3’. Taqman probe for Tax/Rex cDNA was (TMP-2)
5’FAM-ATC ACC TGG GAC CCC ATC- TAMRA-3’. For amplication of pX ORF I
cDNA (p12I and p27I messages), same reverse primer (3’6552) 5’-
GGAGAAAGCAGGAAGAGC-3’ and Taqman probe (TMP-1), 5’FAM-
TTCGCCTTCTCAGCCCCTTGTCT-TAMRA-3’ were used. The p27I forward splice
site-specific primer (ex2/6383) 5’-ACCAACACCATGGCAACT-3’ and p12I forward
specific primer (ex1/6383) 5’-GTCCGCCGTCTAGCAAC- 3’ were used. For
normalization purpose, we measured human GAPDH copy number at the same cDNA
sample. Forward primer 5’- CATCAATGACCCCTTCATTGAC- 3’, reverse primer 5’-
CGCCCCACTTGATTTTGGA-3’, and Taqman probe 5’FAM-
TGGCAAATTCCATGGCACCGTC-TAMRA-3’ were used to detect human GAPDH
cDNA. Standard curves for each real time PCR reaction was generated from cDNA
plasmids: SE356 for Tax/Rex cDNA (P. Green, Ohio State University); pMS0-1.8 for

146
p27 I (D. Derse, National Cancer Institute, Frederick, MD); pMT2-2.3 for p12I (D.
Derse, National Cancer Institute) and pBluescript hGAPDH (K. Boris-Lawrie, Ohio
State University). These cDNAs were diluted from 1 X 101 to 1 X 107 copies to create
standard curves. Twenty five µl volumes containing 12.5 µl of universal PCR master
mix (Applied Biosystems), 200 nM of specific probe, 600nM of each specific primers
and 2.5 µl of cDNA template were used for each reaction. Each assays were performed
in 96-well optical plates (Applied Biosystems) containing duplicate wells for standard,
non-template control and samples. Real time PCR reaction was 50 ° C for 2 min, 95 ° C
for 10 min for activation of DNA polymerase, and 40 cycles of 15 s at 94.5 ° C and 1
min at 60 ° C. The threshold cycle (Ct) values were used to plot standard curves. The
correlation values of standard curves for each assay were always more than 97 %. The
copy numbers of each cDNA samples were calculated from their measured Ct value
base on standard curves and normalized to 1 x 106 copies of human GAPDH.
4.3 RESULTS
Abrogation of pX ORF I expression influences activation status of HTLV-1
infected cells
To determine if the expression of HTLV-1 pX ORF I could activate HTLV-1
infected cells, we performed flow cytometric analysis of T-cell activation markers (CD
25/IL-2 receptor α and CD69) on wild type HTLV-1 infected cells (ACH cell line;
ACH.2 and ACH.2p) and T cells lacking pX ORF I expression (ACH.p12 cell line;

147
ACH.p12.4 and ACH.p12.4p). ACH.2p and ACH.p12.4p cells were newly generated
from PBMC donated from same donor by cocultivating with ACH.2 and ACH.p12.4
cells respectively. Expression of IL-2 receptor α (CD25) was approximately 99% in
both wild type ACH cells and ACH.p12 cells (Fig. 4.1A) and was not significantly
different between each cell line. On the other hand, the expression of CD69 was
consistently higher in wild type ACH cells compare to ACH.p12 cells. The enhanced
CD69 expression was evident comparing expression between ACH.2 (68.6 % positive)
and ACH.p12.4 (42.4 %) and between newly generated cell lines ACH.2p (70.9 %) and
ACH.p12.4p (49.6 %), These data suggest that abrogation of pX ORF I mRNA
expression may influence CD69 cell surface expression. These data are also consistent
with the known calcium regulatory property of pX ORF I encoded p12I, the known
responsiveness of CD69 expression to calcium signaling 39, and the activation status of
HTLV-1 infected cells.
Abrogation of pX ORF I expression in HTLV-1 infected cells did not affect
bystander proliferation of uninfected target PBMC.
Because bystander activation of uninfected target cells can be induced by
interaction between CD2/CD58, and LFA-1/ICAM-1 interaction 19,20, we measured
expression of CD2, CD58 (LFA-3), CD11a (LFA-1), and CD54 (ICAM-1) in ACH and
ACH.p12 cell lines to test whether the deletion of pX ORF I protein expression could
modulate cell surface expression of these adhesion molecules. Approximately 99 % of
all cell types (ACH.2, ACH.2p, ACH.p12.4 and ACH.p12.4p) were positive for CD2,
CD58, CD11a and CD54 (Fig. 4.1B and C). These results indicate that abrogation of

148
p12I did not affect cell surface expression of adhesion molecules, which are critical for
the cell contact-dependent bystander target cell activation.
We then tested whether the expression of pX ORF I in HTLV-1 infected cells
could induce proliferation of bystander uninfected target cells. As previous studies
indicated 19,20, fixed or irradiated HTLV-1 infected cells were able to induce non-
specific activation of uninfected target cells when they were cocultured. We measured
proliferation of target quiescent PBMC by cocultivating fixed effecter cells (ACH.2,
ACH.2p, ACH.p12.4 and ACH.p12.4p) using MTS colorimetric assays in the presence
or absence of IL-2 and PHA stimulation (Fig. 4.2). As expected, fixed HTLV-1 infected
cells induced significant proliferation of target PBMC after 3 days of coculture in the
absence of stimulation. However, the fold proliferation of target PBMC induced by wild
type ACH and ACH.p12 cells was not significantly different, indicating abrogation of
p12I in HTLV-1 infected cells did not affect the HTLV-1–mediated non-specific
bystander target cell proliferation.
Abrogation of pX ORF I expression in HTLV-1 infected cells did not affect
bystander activation of target PBMC.
To further test if pX ORF I expression could influence the activation of target
PBMC, we measured expression of activation markers (CD25 and CD69) on target
PBMC by flow cytometric analysis. HTLV-1 infected effecter cells (ACH.2, ACH.2p,
ACH.p12.4 and ACH.p12.4p) were cocultivated as live or fixed cells with freshly
prepared quiescent PBMC. Live effecter cells were also cocultured separately in
permeable transwell inserts to test the role of soluble factors in cell activation. Target

149
PBMC were pre-labeled with CFDA-SE to differentiate target PBMC from HTLV-1
infected effecter cells.
By flow cytometric analysis, we were able to differentiate CFDA-SE labeled
target PBMC from unlabeled HTLV-1 infected effecter cells (Fig. 4.3 A). CD25 and
CD69 percentage positive were measured in the target PBMC (CMFD-SE positive
cells) after 24 h (Fig. 4.3 B-D) or 36 h (Fig. 4.3 E-G) coculture. As expected, when
PBMC were stimulated with IL-2 and PHA, the expression of activation marker CD69
and CD25 was up-regulated at 24 h and 36 h after stimulation, with CD69 earlier
induction than CD25 (Fig. 4.3 B-G). The expression of CD69 and CD25 in target
PBMC, which were cocultured with live or fixed effecter cells (ACH.2, ACH.2p,
ACH.p12.4 and ACH.p12.4p), was significantly increased when compared that of
unstimulated PBMC control (Fig.4.3 B, C, E and F). However, effecter cells cultured in
transwell insert did not up-regulate CD69 and CD25 expression in target PBMC, which
was not significantly greater than that of unstimulated PBMC control (Fig.4.3 D and G).
These results are consistent with previous studies 19,20 that indicated that non-specific
stimulation of uninfected PBMC was induced by direct contact, but not by soluble
factors from infected cells.
However, the induction ability of activation markers up-regulation on target
PBMC was not significantly different between wild type ACH (ACH.2 and ACH.2p)
and ACH.p12 cell lines (ACH.p12.4 and ACH.p12.4p) (Fig.4.3 B-G). As consistent
with proliferation assay result, the data indicated abrogation of p12I in HTLV-1 infected
cells did not affect the non-specific target cell activation. These results suggest that

150
bystander target cell activation can be sufficiently induced without expression of pX
ORF I by other mechanism, such as Tax- mediated T cell activation 40.
HTLV-1 pX ORF I and pX ORF III/IV mRNAs were differentially expressed
during the early HTLV-1 transmission.
To examine role of pX ORF I messages during early HTLV-1 infection and
transmission, we carried out real time-RT-PCR. Before coculture assay, we measured
relative mRNA copy number (per one million human GAPDH mRNA) of Tax/Rex,
p27I, and p12I messages in HTLV-1 infected cells (MT-2, ACH.2p, and ACH.p12.4p),
as well as in uninfected PBMC. As consistent with previous report 13, Tax/Rex mRNA
was the most abundant, and p27I and p12I mRNA were detected at 1000-2000 fold less
than Tax/Rex mRNA in MT-2 cells (Fig. 4.4 A). In wild type ACH cell line, ACH.2p
cells, p27I and p12I mRNAs were detected approximately at 400-1000 fold less than
Tax/Rex mRNA (Fig. 4.4 A). As expected, both p27I and p12I mRNA were not detected
in ACH.p12 cells (Fig. 4.4 A), which are lacking pX ORF I mRNA expression due to an
introduction of a mutation in splice accepter site of the third exon 23,41.
We then profiled pX ORF I mRNA (p12I and p27I messages) and pX ORF
III/IV mRNA (Tax/Rex message) from 3 hours to 24 hours after cocultivating HTLV-1
infected cells (ACH.2p or ACH.p12.4p) with unstimulated quiescent or activated target
PBMC. Tax/Rex, p12I and p27I mRNAs from unsorted coculture samples of HTLV-1
infected and target cells were differentially expressed through out coculture (Fig. 4.4 B-
D). Up-regulation of Tax/Rex mRNA was induced from 3 hours and peaked 6 hours

151
after coculture when wild type ACH.2p cells were coculture with quiescent PBMC.
This up-regulation of Tax/Rex message was greater and faster than that observed in
coculture of ACH.p12.4p cells and quiescent PBMC (Fig. 4.4 B). Interestingly, when
cocultured with activated PBMC, expression of Tax/Rex messages were not highly up-
regulated compared to cocultured with quiescent PBMC. These results indicated that
expression pattern of Tax/Rex mRNA during the HTLV-1 early transmission is affected
by the pX ORF I proteins (p12I and p27I) expression and by activation status of target
cells. Also, these results suggest that stimuli induced by pX ORF I proteins or target
cells may modulate Tax/Rex mRNA expression during HTLV-1 transmission.
Furthermore, p27I and p12I mRNA were also differentially expressed during
early HTLV-1 transmission (Fig. 4.4 C and D). As expected, p12I and p27I messages
were not detected throughout the all time points, when cocultured with pX ORF I
expression abrogated ACH.p12.4p cells. The peak expression of both p27I and p12I
mRNA was detected at 12 hours after coculture. Expression of p12I mRNA was not
affected by activation status of target PBMC (Fig. 4.4 D). These results indicate that
expression of pX ORF I mRNA expressed early in cell-to-cell transmission of HTLV-1
and is tightly regulated during HTLV-1 transmission, suggesting a role of pX ORF I
proteins in HTLV-1 early infection.

152
Expression of pX ORF I proteins increased HTLV-1 viral transmission to target
PBMC.
To examine de novo expression of pX ORF I and pX ORF III/IV mRNA in
target PBMC, we selectively collected target PBMC by sorting out HTLV-1 infected
cells (ACH.2p, and ACH.p12.4p) at 1 and 3 days after coculture. Before coculture, we
pre-labeled HTLV-1 infected cells with CMFDA, which freely diffuses into cells and
reacts with intracellular thiols and generates green florescent by being cleaved by
cytoplasmic easterase. This long term cell tracer is retained in live cells through several
generations, without being transferred to adjacent cells 42. As expected, we could
differentiate CMFDA unlabeled target PBMC from stained ACH.2p and ACH.p12.4p
cells by flow cytometric analysis at 1 or 3 day after coculture (Fig. 4.5 A). Collected
target PBMC were consistently more than 95 % CMFDA negative (Fig. 4.5 B) and
labeled HTLV-1 cells were more than 90 % CMFDA positive (Fig. 4.5 C) after cell
sorting.
To test role of pX ORF I protein expression in HTLV-1 viral infectivity on
target PBMC, we then measured Tax/Rex mRNA both in CMFDA positive HTLV-1
producer cells and CMFDA negative target PBMC after cell sorting of 1 and 3 day
coculture. Because expression of Tax/Rex mRNA was significantly correlated with
HTLV-1 proviral DNA load 43, viral infectivity was evaluated by measuring percent of
Tax/Rex mRNA expressed in target PBMC at the point of cell sorting (Fig. 4.6). As
expected, a longer 3 day-coculture increased viral infectivity of both ACH.2p and
ACH.p12.4p cells. At 1 day-coculture, viral infectivity of ACH.p12.4p cells to the

153
quiescent PBMC was significantly decreased compared to that of wild type ACH.2p
cells. However, to the activated PBMC, viral infectivity between wild type ACH.2p and
ACH.p12.4p cells was not significantly different. Whereas, after 3 days coculture, viral
infectivity of wild type ACH.2p cells was consistently greater than that of pX ORF I
expression abrogated ACH.p12.4p cells both in quiescent and activated PBMC (Fig.
4.6). These results were consistent with previous in vitro infectivity data measured
HTLV-1 p19 antigen, suggesting the expression of pX ORF I mRNA is required for
efficient viral transmission to target cells.
Expression of pX ORF I proteins enhanced early establishment HTLV-1 de novo
infection in target PBMC.
To profile expression of Tax/Rex, p27I and p12I mRNA in newly infected target
PBMC, we selectively cultured target PBMC (CMFDA negative), which were sorted
from the 3 day-coculture cells and carried out real time RT-PCR. The relative copy
number of Tax/Rex mRNA was measured in quiescent or activated target PBMC, which
cocultivated with wild type ACH.2p or ACH.p12.4p cells (Fig. 4.7 A). The fold
increase of Tax/Rex mRNA expression was measured on day 5 (2 days after sorting)
and on day 7 (4 day after sorting) (Fig. 4.7 B). From day 3 to day 5, the fold increase of
Tax/Rex mRNA expression was greater in activated PBMC (8 to 10 fold) than in
quiescent PBMC (1 to 3 fold), and the expression Tax/Rex mRNA was not significantly
increased from day 5 to day 7, suggesting establishment of HTLV-1 infection in target
PBMC occurs in 5 days after coculture. Furthermore, the fold increase of Tax/Rex
mRNA expression was consistently higher in target PBMC when they were cocultivated

154
with wild type ACH.2p cells than when they were cultured with ACH.p12.4p cells (Fig.
4.7 B). These results indicate that expression of pX ORF I protein in newly infected
cells facilitated establishment of HTLV-1 infection.
Furthermore, we profiled pX ORF I mRNA expression in sorted target PBMC to
examine how expression of pX ORF I mRNAs are regulated in newly infected cells.
The expression of p27I and p12I mRNA was consistently detected in activated target
PBMC that were cocultivated with wild type ACH.2p cells (Fig. 4.7 C). Relative copy
number of pX ORF I mRNA was consistently increased after cell sorting. However,
ratio to Tax/Rex mRNA was decreased on day 5 and was slightly increased on day 7
(Fig. 4.7 D), indicating that expression of pX ORF I mRNAs were differentially
regulated during early establishment of HTLV-1 infection.
4.4 DISCUSSION
In this study, we demonstrated a role of expression of HTLV-1 pX ORF I
during early HTLV-1 transmission and infection. The abrogation of pX ORF I message
influenced the activation status of HTLV-1 infected cells, but did not effect HTLV-1-
induced bystander target cell activation. Our data indicates that the infectivity of T cell
lines expressing the full length infectious clone of HTLV-1 (wild type ACH) was
greater than that of ACH.p12 immortalized T cells as measured by Tax/Rex mRNA in
target cells. We also demonstrated that pX ORF I is an early transcript expressed cell-
to-cell transmission of HTLV-1. Our results were consistent with previous studies that

155
implicated the expression of pX ORF I plays a role in activation of HTLV-1 infected T
cell and is required for efficient HTLV-1 cell-to-cell transmission.
Expression of IL-2 receptor α (CD25) was consistently enhanced in both wild
type ACH cells and ACH.p12 cells lacking pX ORF I expression, indicating CD25
expression is not influenced by abrogation of pX ORF I expression. However, the early
activation marker CD69 expression was consistently higher in wild type ACH cells.
Interestingly, expression of CD69 is dependent on calcium mediated signaling 39.
Previous results from our laboratory indicated that pX ORF I encoded p12I is a
regulatory of calcium signaling leading the downstream gene expression including
NFAT activation 25-27. Our data links these two observations and offers a mechanism for
enhancement of CD69 expression on HTLV-1 infected cells. CD69 expression was up-
regulated in HTLV infected cells from HAM patients or asymptomatic carriers after 2
days ex vivo culture, but not was not significantly increased when they were freshly
isolated compared to uninfected control cells 44, indicating stimulation was generated
during ex vivo culture. Although CD69 is a well established and an earliest activation
marker of lymphocytes, the precise role in immunity is still elusive due to lack of
known ligands and a suitable in vivo model 45. Recent findings suggested that CD69
acts as immunoregulatory function by producing transforming growth factor-β (TGF-β)
45,46, which down-regulates antigen-presenting cell (APC) function 47, suggesting a
possible role of CD69 expression on HTLV-1 infected cell in escaping immune
surveillance. However, our previous rabbit model results do not support this hypothesis
because ACH.p12 cells did not elicit strong immune response compare to wild type
ACH cells, which have higher CD69 expression, when they were inoculated into rabbits

156
23. Further studies will be required to understand the role of CD69 in HTLV-1 cell-to-
cell transmission and the role of viral gene products in its expression.
Activation of target PBMC is advantageous for retroviral integration, subsequent
viral replication and productive infection. Bystander non-specific activation of PBMC
can be induced by direct contact with HTLV-1 infected cells via interaction of adhesion
molecule such as CD2 / LFA-3 (CD58) and LFA-1(CD11a) / ICAM (CD54)19,20.
Furthermore, signaling via CD2 and ICAM-1 receptor enhanced HTLV-1 replication
48,49, suggesting that the receptor- mediated signaling generated during HTLV-1 cell-to-
cell transmission is important for early viral infection. Because both expressions of
LFA-3 (CD58) and ICAM-1 (CD54) were constitutively enhanced in HTLV-1 infected
cells 50,51, target cells may be activated via CD2 or LFA-1 mediated signaling.
Although the cell surface expression of CD2, LFA-3 (CD58), LFA-1 (CD11a),
ICAM (CD54), and IL-2 receptor (CD25) was not significantly different between wild
type ACH and ACH.p12 cells in this study, LFA-1-mediated adhesion due to LFA-1
clustering in ACH cells was significantly enhanced compared to ACH.p12 cells in
previous study 31. Together our studies indicate that pX ORF I expression influences
LFA-1 clustering on infected cells to enhance cell-to-cell adhesion and stimuli derived
from cell contact with wild ACH cells may be greater than that from ACH.p12 cell
contact. We have previously demonstrated that ACH cells have increased IL-2
production in presence of T cell stimulation compared to ACH.2 cells 27. However,
bystander activation of target PBMC induced by these two cell types was not
significantly different, indicating that target cell activation can be sufficiently induced
in the absence of pX ORF I mRNA. Interestingly, HTLV-1 Tax induced transcriptional

157
activation ICAM-1 gene 40, suggesting that the ICAM-1 expression may be up-regulated
by Tax in HTLV-1 infected cells and may resulting in sufficient activation of target
PBMC by cell-to-cell contact. Furthermore, antibodies against LFA-1 and ICAM-1
triggered re-polarization of MTOC, which is characteristic of HTLV-1-induced
virological synapses 33, suggesting that pX ORF I protein may synergize to MTOC
polarization and contribute to HTLV-1 cell-to-cell transmission.
Accumulation of Gag and Env proteins was observed in cell contact junction
(virological synapse) with in 40 min after coculture of HTLV-1 infected cells with
target cells, and the transfer of Gag protein in target cells was detected within 2 hours 35.
Furthermore, newly synthesized unintegrated HTLV-1 DNA (de novo reverse
transcription) was detected within 4-8 hours after coculture 52, indicating that HTLV-1
cell-to-cell transmission occur in a short time frame. Early events during cell-to-cell
transmission may determine HTLV-1 vial infectivity to target cells by modulating
expression of HTLV-1 gene expression. Therefore, we profiled expression of pX ORF I
mRNA(p12I and p27I) and pX ORF III/IV (Tax/Rex) mRNA from 3 hours to 7 days
after cocultivating HTLV-1 infected cells. We were particularly interested in the role of
pX ORF I protein during early HTLV-1 transmission and therefore utilized ACH.p12
cells, which are lacking pX ORF I mRNA expression.
In this study, we demonstrated that expression of Tax/Rex mRNA was
differentially regulated during early HTLV-1 transmission. Furthermore, Tax/Rex
expression was enhanced by pX ORF I expression and activation status of target cells.
Our data suggested that stimuli and their corresponding signaling pathways, which were
generated from pX ORF I proteins or cell contacts with target cells, may modulate

158
Tax/Rex mRNA expression and subsequent Tax-mediated viral production during
HTLV-1 transmission. Interestingly, elevation of intracellular calcium resulted in
enhancement of structural protein synthesis and activation of long terminal repeat
(LTR) - mediated transcription in cells infected with bovine leukemia virus (BLV),
another member of the deltaretrovirus genus 53 A number of studies have been reported
a role of Tax in transcriptional regulation of expression of viral and cellular genes 54,
but the influence of other viral ORFs in Tax expression is unclear. Signaling via CD2
and ICAM-148,49, which enhanced viral replication and production, may affect
expression of Tax/Rex in HTLV-1 cells.
Our data indicate that the expression of pX ORF I (p12I and p27I) mRNA while
during early HTLV-1 transmission was always lower than that of Tax/Rex mRNA. Our
result combined with our previous studies of HTLV-1 infectivity in target PBMC verify
a critical role of pX ORF I expression in viral transmission. Our data indicate that an
early increase of Tax/Rex mRNA expression in sorted target cells, which reflects de
novo HTLV-1 gene expression. Tax/Rex mRNA expression was increased in cells that
also expressed pX ORF I, suggesting a another role of pX ORF I protein in establishing
HTLV-1 infection in target cells. While the mechanism of pX ORF I expression in viral
replication is beyond the scope of our current study, based on previous studies of pX
ORF I encoded p12I, the viral protein may influence the viral life cycle (e.g., reverse
transcription) directly or indirectly through its unknown cell activation properties and
calcium signaling properties. If pX ORF I encoded p12I enhances early events
following virus entry such as reverse transcription and integration, the protein would
need to be present in viral pre-integration complexes or expressed early following initial

159
integration events. Our data is the first to indicate that indeed pX ORF I is expressed
early (within 12 h) of cell-to-cell contact. Analogously, the human immunodeficiency
virus (HIV) accessory protein, Nef, which functions in a manner similar to HTLV-1
p12I, is found as a protein in HIV virions 55 and is selectively expressed before viral
integration 56. Similarly, our data would support a role for HTLV-1 pX ORF I protein in
the formation of the viral synapse or selectively expressed before or immediately
following integration to enhance de novo HTLV-1 infection in target cells.
In summary, our study further defines a role of pX ORF I expression in
activation of HTLV-1 infected cells. Our data indicated that expression of pX ORF I
enhanced early establishment HTLV-1 infection in target cells during cell-to-cell
transmission and that abrogation of pX ORF I expression did not influence HTLV-1
mediated bystander cell activation. Collectively, we have clarified the role of pX ORF I
in HTLV-1 replication and viral transmission during the earliest event of viral infection
in T lymphocytes.
4.5 References
(1) Gessain A, Mahieux R. [A virus called HTLV-1. Epidemiological aspects. Presse Med. 2000;29:2233-2239.
(2) Bangham CR. HTLV-1 infections. J Clin Pathol. 2000;53:581-586.
(3) Franchini G. Molecular mechanisms of human T-cell leukemia/lymphotropic virus type I infection. Blood. 1995;86:3619-3639.
(4) Hollsberg P. Mechanisms of T-cell activation by human T-cell lymphotropic virus type I. Microbiol Mol Biol Rev. 1999;63:308-333.
(5) Berneman ZN, Gartenhaus RB, Reitz MS et al. Expression of alternatively spliced human T-lymphotropic virus type 1 pX mRNA in infected cell lines and

160
in primary uncultured cells from patients with adult T-cell leukemia/lymphoma and healthy carriers. Proc Natl Acad Sci USA. 1992;89:3005-3009.
(6) Ciminale V, Pavlakis GN, Derse D, Cunningham CP, Felber BK. Complex splicing in the human T-cell leukemia virus (HTLV) family of retroviruses: Novel mRNAs and proteins produced by HTLV type I. J Virol. 1992;66:1737-1745.
(7) Furukawa K, Furukawa K, Shiku H. Alternatively spliced mRNA of the pX region of human T lymphotropic virus type I proviral genome. FEBS Lett. 1991;295:141-145.
(8) Koralnik IJ, Gessain A, Klotman ME et al. Protein isoforms encoded by the pX region of human T-cell leukemia/lymphotropic virus type 1. Proc Natl Acad Sci USA. 1992;89:8813-8817.
(9) Cereseto A, Berneman Z, Koralnik I et al. Differential expression of alternatively spliced pX mRNAs in HTLV-I-infected cell lines. Leukemia. 1997;11:866-870.
(10) Koralnik IJ, Gessain A, Klotman ME et al. Protein isoforms encoded by the pX region of human T-cell leukemia/lymphotropic virus type I. Proc Natl Acad Sci U S A. 1992;89:8813-8817.
(11) Dekaban GA, Peters AA, Mulloy JC et al. The HTLV-I orfI protein is recognized by serum antibodies from naturally infected humans and experimentally infected rabbits. Virology. 2000;274:86-93.
(12) Pique C, Uretavidal A, Gessain A et al. Evidence for the chronic in vivo production of human T cell leukemia virus type I Rof and Tof proteins from cytotoxic T lymphocytes directed against viral peptides. J Exp Med. 2000;191:567-572.
(13) Princler GL, Julias JG, Hughes SH, Derse D. Roles of viral and cellular proteins in the expression of alternatively spliced HTLV-1 pX mRNAs. Virology. 2003;317:136-145.
(14) Mueller N.E., Blattner W.A. Retroviruses-Human T-Cell Lymphotropic Virus. Fourth Edition, 785-814. 1997. New York, N.Y., Plenum Publishing Corp.
(15) Bangham CR. The immune control and cell-to-cell spread of human T-lymphotropic virus type 1. J Gen Virol. 2003;84:3177-3189.
(16) Manns A, Hisada M, Lagrenade L. Human T-lymphotropic virus type I infection. Lancet. 1999;353:1951-1958.

161
(17) Kehn K, Berro R, de La FC et al. Mechanisms of HTLV-1 transformation. Front Biosci. 2004;9:2347-2372.
(18) Merl S, Kloster B, Moore J et al. Efficient Transformation of Previously Activated and Dividing T Lymphocytes by Human T Cell Leukemia-Lymphoma Virus. Blood. 1984;64:967-974.
(19) Kimata JT, Palker TJ, Ratner L. The Mitogenic Activity of Human T-Cell Leukemia Virus Type- I Is T-Cell Associated and Requires the CD2/LFA-3 Activation Pathway. J Virol. 1993;67:3134-3141.
(20) Wucherpfennig KW, Hollsberg P, Richardson JH, Benjamin D, Hafler DA. T-cell activation by autologous human T-cell leukemia virus type I- infected T-cell clones. Proc Natl Acad Sci U S A. 1992;89:2110-2114.
(21) Albrecht B, Lairmore MD. Critical role of human T-lymphotropic virus type 1
accessory proteins in viral replication and pathogenesis. Microbiol Mol Biol Rev. 2002;66:396-406.
(22) Michael B, Nair A, Lairmore MD. Role of accessory proteins of HTLV-1 in viral replication, T cell activation, and cellular gene expression. Front Biosci. 2004;9:2556-2576.
(23) Collins ND, Newbound GC, Albrecht B et al. Selective ablation of human T-cell lymphotropic virus type 1 p12I reduces viral infectivity in vivo. Blood. 1998;91:4701-4707.
(24) Albrecht B, Collins ND, Burniston MT et al. Human T-lymphotropic virus type 1 open reading frame I p12(I) is required for efficient viral infectivity in primary lymphocytes. J Virol. 2000;74:9828-9835.
(25) Albrecht B, D'Souza CD, Ding W et al. Activation of nuclear factor of activated T cells by human T- lymphotropic virus type 1 accessory protein p12(I). J Virol. 2002;76:3493-3501.
(26) Ding W, Albrecht B., Kelley RE et al. Human T lymphotropic virus type 1 p12(I) expression increases cytoplasmic calcium to enhance the activation of nuclear factor of activated T cells. J Virol. 2002;76:10374-10382.
(27) Ding W, Kim SJ, Nair AM et al. Human T-Cell Lymphotropic Virus Type 1 p12(I) Enhances Interleukin-2 Production during T-Cell Activation. J Virol. 2003;77:11027-11039.
(28) Ding W, Albrecht B, Luo R et al. Endoplasmic Reticulum and cis-Golgi Localization of Human T- Lymphotropic Virus Type 1 p12(I): Association with Calreticulin and Calnexin. J Virol. 2001;75:7672-7682.

162
(29) Kim SJ, Ding W, Albrecht B, Green PL, Lairmore MD. A conserved calcineurin-binding motif in human T lymphotropic virus type 1 p12I functions to modulate nuclear factor of activated T cell activation. J Biol Chem. 2003;278:15550-15557.
(30) Nair A, Michael B, Hiraragi H et al. Human T lymphotropic virus type 1 accessory protein p12I modulates calcium-mediated cellular gene expression and enhances p300 expression in T lymphocytes. AIDS Res Hum Retroviruses. 2005;21:273-284.
(31) Kim SJ, Nair AM, Fernandez S, Mathes L, Lairmore MD. Enhancement of LFA-1-mediated T cell adhesion by human T lymphotropic virus type 1 p12I1. J Immunol. 2006;176:5463-5470.
(32) Nejmeddine M, Barnard AL, Tanaka Y, Taylor GP, Bangham CR. Human T-lymphotropic Virus, Type 1, Tax Protein Triggers Microtubule Reorientation in the Virological Synapse. J Biol Chem. 2005;280:29653-29660.
(33) Barnard AL, Igakura T, Tanaka Y, Taylor GP, Bangham CR. Engagement of specific T cell surface molecules regulates cytoskeletal polarization in HTLV-1-infected lymphocytes. Blood. 2005.
(34) Dustin M. Viral spread through protoplasmic kiss. Nat Cell Biol. 2003;5:271-272.
(35) Igakura T, Stinchcombe JC, Goon PK et al. Spread of HTLV-I between lymphocytes by virus-induced polarization of the cytoskeleton. Science. 2003;299:1713-1716.
(36) Mori N, Murakami S, Oda S, Eto S. Human T-cell leukemia virus type I tax induces intracellular adhesion molecule-1 expression in T cells. Blood. 1994;84:350-351.
(37) Robek MD, Wong FH, Ratner L. Human T-Cell leukemia virus type 1 pX-I and pX-II open reading frames are dispensable for the immortalization of primary lymphocytes. J Virol. 1998;72:4458-4462.
(38) Collins ND, D'Souza C, Albrecht B et al. Proliferation response to interleukin-2 and Jak/Stat activation of T cells immortalized by human T-cell lymphotropic virus type 1 is independent of open reading frame I expression. J Virol. 1999;73:9642-9649.
(39) Rodrigues MS, Echevarria-Lima J, Fernandes dos SN, Rumjanek VM. CD69 expression induced by thapsigargin, phorbol ester and ouabain on thymocytes is dependent on external Ca2+ entry. Life Sci. 2003;73:1037-1051.

163
(40) Owen SM, Rudolph DL, Dezzutti CS et al. Transcriptional activation of the intercellular adhesion molecule 1 (CD54) gene by human T lymphotropic virus types I and II Tax is mediated through a palindromic response element. AIDS Res Hum Retroviruses. 1997;13:1429-1437.
(41) Kimata JT, Wong F, Wang J, Ratner L. Construction and characterization of infectious human T-cell leukemia virus type 1 molecular clones. Virology. 1994;204:656-664.
(42) Lantz RC, Lemus R, Lange RW, Karol MH. Rapid reduction of intracellular glutathione in human bronchial epithelial cells exposed to occupational levels of toluene diisocyanate. Toxicol Sci. 2001;60:348-355.
(43) Yamano Y, Nagai M, Brennan M et al. Correlation of human T-cell lymphotropic virus type 1 (HTLV-1) mRNA with proviral DNA load, virus-specific CD8(+) T cells, and disease severity in HTLV-1-associated myelopathy (HAM/TSP). Blood. 2002;99:88-94.
(44) Alfahim A, Cabre P, Kastrukoff L, Dorovinizis K, Oger J. Blood mononuclear cells in patients with HTLV-I-associated myelopathy: Lymphocytes are highly activated and adhesion to endothelial cells is increased. Cell Immunol. 1999;198:1-10.
(45) Sancho D, Gomez M, Sanchez-Madrid F. CD69 is an immunoregulatory molecule induced following activation. Trends Immunol. 2005;26:136-140.
(46) Sancho D, Gomez M, Viedma F et al. CD69 downregulates autoimmune reactivity through active transforming growth factor-beta production in collagen-induced arthritis. J Clin Invest. 2003;112:872-882.
(47) Kulkarni AB, Huh CG, Becker D et al. Transforming growth factor beta 1 null mutation in mice causes excessive inflammatory response and early death. Proc Natl Acad Sci U S A. 1993;90:770-774.
(48) Yamamoto A, Hara H, Kobayashi T. Induction of the expression of gag protein in HTLV-I infected lymphocytes by anti-ICAM 1 antibody in vitro. J Neurol Sci. 1997;151:121-126.
(49) Guyot DJ, Newbound GC, Lairmore MD. Signaling via the CD2 receptor enhances HTLV-1 replication in T lymphocytes. Virology. 1997;234:123-129.
(50) Fukudome K, Furuse M, Fukuhara N et al. Strong induction of ICAM-1 in human T cells transformed by human T-cell-leukemia virus type 1 and depression of ICAM-1 or LFA-1 in adult T-cell-leukemia-derived cell lines. Int J Cancer. 1992;52:418-427.

164
(51) Imai T, Tanaka Y, Fukudome K et al. Enhanced expression of LFA-3 on human T-cell lines and leukemic. International Journal of Cancer. 1993;55(5):811-816.
(52) Benovic S, Kok T, Stephenson A et al. De novo reverse transcription of HTLV-1 following cell-to- cell transmission of infection. Virology. 1998;244:294-301.
(53) Bondzio A, Abraham-Podgornik A, Blankenstein P, Risse S. Involvement of intracellular Ca2+ in the regulation of bovine leukemia virus expression. Biol Chem. 2001;382:407-416.
(54) Kashanchi F, Brady JN. Transcriptional and post-transcriptional gene regulation of HTLV-1. Oncogene. 2005;24:5938-5951.
(55) Zhou J, Aiken C. Nef enhances human immunodeficiency virus type 1 infectivity resulting from intervirion fusion: evidence supporting a role for Nef at the virion envelope. J Virol. 2001;75:5851-5859.
(56) Wu Y, Marsh JW. Selective transcription and modulation of resting T cell activity by preintegrated HIV DNA. Science. 2001;293:1503-1506.

165
Figure 4.1.Cell surface expression of early activation markers and adhesion
molecules on wild type HTLV-1 infected cells and T cells lacking pX ORF I
expression. Flow cytometric analysis specific for activation markers (A) CD25 and
CD69, adhesion molecules (B) CD58 and CD2, and (C) CD11a and CD54 was
performed on ACH cell line (ACH.2 and ACH.2p) and ACH.p12 cell line (ACH.p12.4
and ACH.p12.4p) (106 cells/sample). The results were representative of at least 2
independent experiments.

166

167
Figure 4.2. Deletion of pX ORF I expression in HTLV-1 infected cells did not affect
bystander proliferation of uninfected target PBMC.
ACH cell line (ACH.2 and ACH.2p) and ACH.p12 cell line (ACH.p12.4 and
ACH.p12.4p) were fixed with 1% PFA and cocultured target PBMC in the presence or
absence of human IL-2 (20 U/ml) and PHA (2 µg/ml) stimulation. Fixed effecter cells
were cocultured with target PBMC at 1:2 ratios. Target PBMC were cultured without
effecter cells for control. The MTS colorimetric assay was performed at 0 days and at 3
days after coculture by measuring optical density (OD) in triplicate samples. Graph
depicts the fold proliferation of target cell OD at day 3 over day 0. Values are mean ±
SEM of triplicate samples of 3 independent experiments. Statistical significance was
analyzed by Student’s t test.
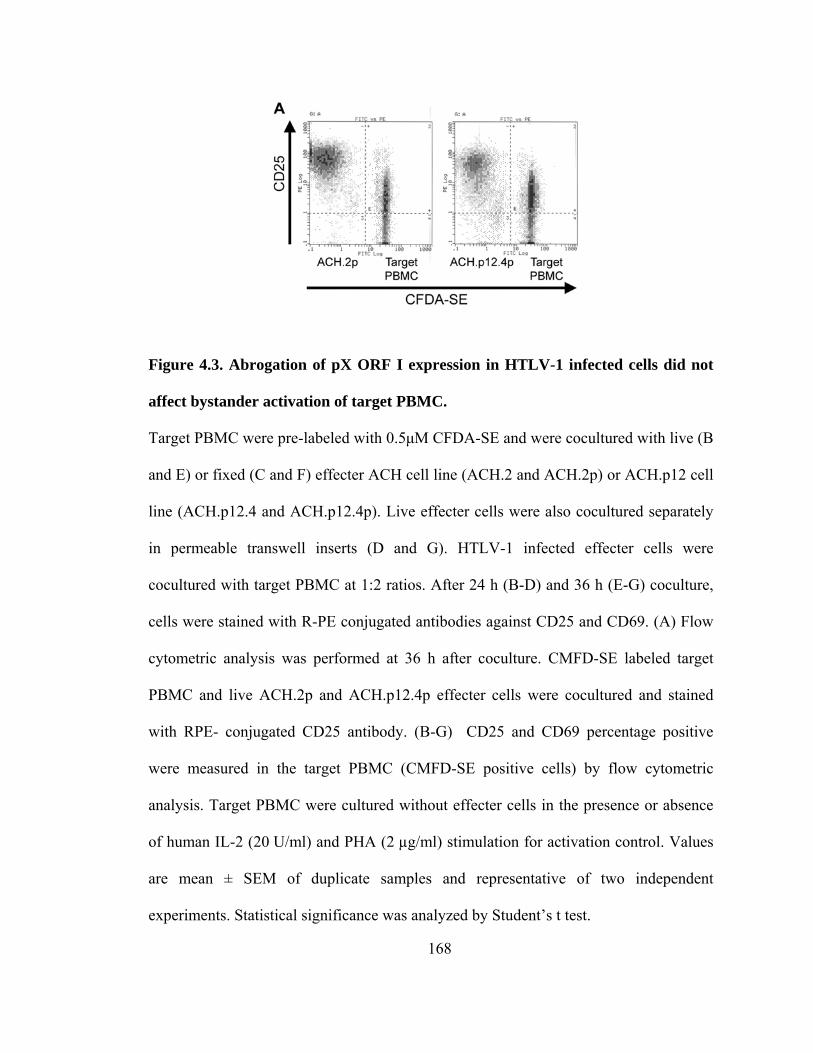
168
Figure 4.3. Abrogation of pX ORF I expression in HTLV-1 infected cells did not
affect bystander activation of target PBMC.
Target PBMC were pre-labeled with 0.5µM CFDA-SE and were cocultured with live (B
and E) or fixed (C and F) effecter ACH cell line (ACH.2 and ACH.2p) or ACH.p12 cell
line (ACH.p12.4 and ACH.p12.4p). Live effecter cells were also cocultured separately
in permeable transwell inserts (D and G). HTLV-1 infected effecter cells were
cocultured with target PBMC at 1:2 ratios. After 24 h (B-D) and 36 h (E-G) coculture,
cells were stained with R-PE conjugated antibodies against CD25 and CD69. (A) Flow
cytometric analysis was performed at 36 h after coculture. CMFD-SE labeled target
PBMC and live ACH.2p and ACH.p12.4p effecter cells were cocultured and stained
with RPE- conjugated CD25 antibody. (B-G) CD25 and CD69 percentage positive
were measured in the target PBMC (CMFD-SE positive cells) by flow cytometric
analysis. Target PBMC were cultured without effecter cells in the presence or absence
of human IL-2 (20 U/ml) and PHA (2 µg/ml) stimulation for activation control. Values
are mean ± SEM of duplicate samples and representative of two independent
experiments. Statistical significance was analyzed by Student’s t test.

169

170

171
Figure 4.4. HTLV-1 pX ORF I and pX ORF III/IV mRNAs were differentially
expressed during the early HTLV-1 transmission.
Tax/Rex, p27I, and p12I mRNA copy numbers were determined as described under
Material and Methods and normalized relative to 1.0 x 106 copies of human GAPDH
mRNA. (A) Relative mRNA copy number of Tax/Rex, p27I, and p12I messages in
HTLV-1 infected cells (MT-2, ACH.2p, and ACH.p12.4p), as well as in uninfected
PBMC. Values are mean ± SEM of duplicate samples and representative at least 3
independent experiments. Relative copy number of (B) Tax/Rex, (C) p27I and (D)12I
mRNA were measured from 3 hours to 24 hours after coculture in unsorted samples.
HTLV-1 infected cells (ACH.2p or ACH.p12.4p) were cocultured with unstimulated
quiescent or pre- activated target PBMC at 1:2 ratios. Values are mean ± SEM of
duplicate samples and representative at least 2 independent experiments.

172

173
Figure 4.5. Target PBMC were selectively sorted from HTLV-1 infected cells.
HTLV-1 infected cells (ACH.2p or ACH.p12.4p) were pre-labeled with CMFDA and
cocultured with unstimulated quiescent or pre-activated target PBMC at 1:2 ratios. (A)
CMFDA negative unstimulated target PBMC and CMFDA positive ACH.2p cells were
analyzed at 3 day after coculture by flow cytometry. (B) Collected target PBMC were
99 % CMFDA negative and (C) labeled ACH.2p cells were 95 % CMFDA positive
after cell sorting.
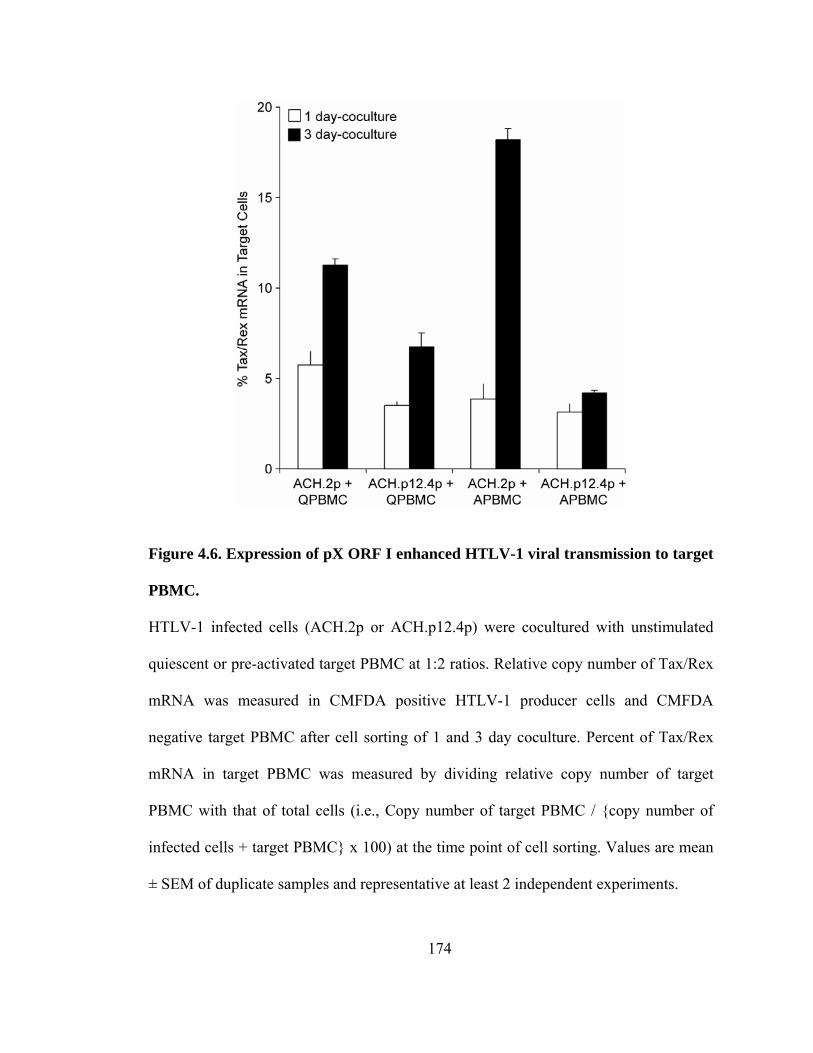
174
Figure 4.6. Expression of pX ORF I enhanced HTLV-1 viral transmission to target
PBMC.
HTLV-1 infected cells (ACH.2p or ACH.p12.4p) were cocultured with unstimulated
quiescent or pre-activated target PBMC at 1:2 ratios. Relative copy number of Tax/Rex
mRNA was measured in CMFDA positive HTLV-1 producer cells and CMFDA
negative target PBMC after cell sorting of 1 and 3 day coculture. Percent of Tax/Rex
mRNA in target PBMC was measured by dividing relative copy number of target
PBMC with that of total cells (i.e., Copy number of target PBMC / {copy number of
infected cells + target PBMC} x 100) at the time point of cell sorting. Values are mean
± SEM of duplicate samples and representative at least 2 independent experiments.

175
Figure 4.7. Expression of HTLV-1 pX ORF I and pX ORF III/IV mRNAs in de
novo HTLV-1 infected target PBMC.
(A, B) HTLV-1 infected cells (ACH.2p or ACH.p12.4p) were cocultured with
unstimulated quiescent or pre-activated target PBMC at 1:2 ratios. (A) Relative copy
number of Tax/Rex mRNA was measured in CMFDA negative target PBMC after cell
sorting of 3 day coculture. (B) Fold increase of Tax/Rex mRNA in sorted PBMC from
day 3 to day5 and from day 5 to day 7 after coculture was measured. Values are mean ±
SEM of duplicate samples and representative 2 independent experiments.

176
(C, D) HTLV-1 infected ACH.2p cells were cocultured with pre-activated target PBMC
at 1:1 ratios. (C) Relative copy number of p27I and p12 I mRNA measured in CMFDA
negative target PBMC after cell sorting of 3 day coculture. (D) p27I and p12 I mRNA
ratio to Tax/Rex mRNA at each day point was measured Values are mean ± SEM of
duplicate samples.

177
CHAPTER 5
SYNOPSIS AND FUTURE DIRECTIONS
Since the initial discovery of HTLV-1, extensive studies have been performed to
understand mechanisms of HTLV-1-mediated lymphoproliferative and
neurodegenerative diseases. Most of these studies have been focused on the role of the
transcriptional activator Tax and its role in immortalization and transformation of T
cells. However, the early events of viral infection, such as cell-to-cell transmission,
regulation of reverse transcription, integration and regulation of viral and cellular gene
transcription are incompletely understood. Recent studies have produced an
accumulating body of data that a critical role for pX ORF I and II encoded proteins in
viral infectivity, maintenance of viral loads, host cell activation, and regulation of gene
transcription 1-5. Particularly, the role of an HTLV-1 p12I encoded by pX ORF I was
discovered to be required for productive infection in a rabbit model and viral infectivity

178
in non-stimulated PBMC 4,5. Moreover, our laboratories data demonstrated the role of
p12I in calcium-dependent signaling, activation of NFAT 1, and enhanced production of
IL-2 6 and p300 expression 7. Data presented in this thesis provided further evidence
that the highly conserved viral accessory protein, p12I is required for early HTLV-1
infection. I demonstrated in this thesis that HTLV-1 p12I modulates NFAT activation
by interacting with calcineurin (Chapter 2), enhances cell-to-cell adhesion by inducing
LFA-1 clustering on the surface of T cells (Chapter 3) and facilitates early viral
infectivity during cell-to-cell transmission (Chapter 4).
5.1 Studies to test the role of p12I binding to calcineurin on HTLV-1-mediated T
cell activation and in vivo and in vitro viral infectivity
We identified a PSLP(I/L)T motif in p12I, which is highly homologous with the
PxIxIT calcineurin-binding motif of NFAT. Through this motif, p12I modulates NFAT
nuclear translocation and transcription activity 8. However it is unclear why p12I has
two regulatory functions for NFAT transcriptional activity; positive modulation by
increasing cytosolic calcium concentration from ER stores 2 and negative modulation by
calcineurin binding. Therefore, further analysis using an infectious HTLV-1 proviral
clone such as ACH that contains mutations in PSLP(I/L)T motif, would be required to
understand the role of p12I binding to calcineurin in context to HTLV-1- mediated cell
activation and during in vivo and in vitro viral infectivity. An alanine substitution in this
motif within an infectious HTLV-1 clone will be helpful to define the role of p12I in
binding to calcineurin in infected cells. The p12I mutant (p12I AxAxAA) exhibited a

179
greatly reduced binding affinity for calcineurin, but retained the ability to enhance
NFAT nuclear translocation and transcriptional activity 8. These properties of p12I,
however, were not tested in context to an HTLV-1 provirus and would be an important
extension of the findings in the thesis. By measuring IL-2 production 6, or detecting
activation markers (e.g., CD25 and CD69), the activation status of HTLV-1 infected
cells expressing an HTLV-1 pX ORF I virus containing AxAxAA mutations could be
evaluated and compared to wild type HTLV-1 infected cells (ACH). Furthermore, in
vitro and in vivo viral infectivity of HTLV-1 infected cells containing this mutant
provirus can be directly compared to wild type HTLV-1 infected cells (ACH) using in
vitro p19 detection 4 or approaches used in Chapter 4 of this thesis such as coculture and
real time RT-PCR methods followed by cell sorting to differentiate target and producer
cells, as well as rabbit inoculation to direct test infectivity in vivo 5.
5.2 Further investigation of the role of p12I in MTOC polarization and virological
synapse formation
In Chapter 3 of this thesis, our data indicated that HTLV-1 p12I promotes cell-
to-cell spread by inducing LFA-1 clustering on T-cells. Furthermore, a recent cross-
linking study indicated that LFA-1 and ICAM-1 interaction is important in HTLV-1
cell-to-cell transmission 9. Antibodies against LFA-1 and ICAM-1 triggered
microtubule-organizing center (MTOC) polarization, which is characteristic of HTLV-
1-induced virological synapses. Taken together my data presented in Chapter 3
suggests that p12I may contribute to MTOC polarization and enhance formation of the
virological synapse by increasing cell adhesion through clustered LFA-1. However, to

180
date, there is no direct published reports that p12I can modulate MTOC polarization and
virological synapse formation. Moreover, p12I may enhance MTOC re-polarization
directly through calcium mediated signaling without upstream activation of signaling
pathway, because calcium-mediated events are required for TCR-mediated MTOC
polarization 10. The role of p12I in MTOC polarization and formation of virological
synapse can be tested by comparing wild type HTLV-1 infected cells (ACH cell line)
and T cells lacking p12I expression (ACH.p12 cell line). The frequency of MTOC
polarization can be measured under the influence of variety of stimulation protocols,
such as engagement of TCR, LFA-1, ICAM-1, CD2 and LFA-3, which are important in
HTLV-1 cell-to-cell mediated bystander target cell activation 11,12. Because LFA-1
clustering was enhanced in wild type ACH cells 13, engagement of LFA-1 may result in
higher MTOC frequency in wild type when compared to ACH.p12 cell lines. By
measuring conjugation rates between HTLV-1 infected cells and target cells, the role of
p12I in formation of the virological synapse could be also tested. Furthermore, studies
designed with exogenously expressed p12I in Jurkat T cell could be expanded to
understand the role of p12I in MTOC polarization and virological synapse formation.
5.3 Studies to explore the mechanisms of LFA-1 affinity regulation influenced by
p12I expression
Our data in Chapter 3 demonstrated that abrogation of ORF I expression in
HTLV-1 infected cells resulted in decreased LFA-1 affinity. However, expression of
p12I in Jurkat T cells did not affect LFA-1 affinity, indicating that the role of p12I is not
directly related to the regulation of LFA-1 affinity control mechanisms. Also these data

181
suggested that in context to HTLV-1 infection the balance of HTLV-1 regulatory and
accessory protein expression is required for LFA-1 affinity control and efficinet cell-to-
cell HTLV-1 transmission. Due to incompletely defined mechanisms of LFA-1 affinity
control in the literature compared to reports of regulation of LFA-1 clustering 14,
investigating mechanism of LFA-1 affinity reduction in p12I expression abrogated cells
(ACH.p12 cell line) may be challenging. In this regard, studies to test the role of other
regulatory or accessory protein in LFA-1 affinity control would be useful to further
understanding the role of this p12I in context to HTLV-1 infection. Interestingly, the
expression of the Lck gene, a critical plasma membrane associated kinase in CD4+ T
cells to maintain the high affinity status of integrin α1β4 15, was down-regulated by the
HTLV-1 accessory protein p30II 16.
5.4 Studies required to test the role of p12I in HTLV-1 envelope glycoprotein
folding and cell surface expression
HTLV-1 p12I was reported to be required for efficient in vivo and in vitro rabbit
viral infectivity 4,5. Because interactions between the HTLV-1 envelope protein and its
receptor are critical for cell-to-cell transmission of the virus, the requirement of p12I on
efficient viral transmission may be due to viral protein’s role or pX ORF I mRNA in
regulating envelope expression. Our previous data demonstrated that p12I interacts with
both calreticulin and calnexin 2, which are molecular chaperones to newly synthesized
and unfolded glycoproteins 17. Furthermore, the folding of most viral envelope proteins
including HIV-1 envelope are associated with calreticulin and calnexin in the lumen of
the endoplasmic reticulum of infected cells 18. Thus the binding of p12I to both

182
calreticulin and calnexin may help the proper folding of HTLV-1 envelope
glycoproteins or increase their cell surface expression to enhance receptor interactions.
5.5 Possible role of p12I in regulating CD69 expression during HTLV-1 infection
The expression of the early activation marker CD69 was consistently higher in
wild type HTLV-1 infected cells compared to ACH.p12 cells. The expression of CD69
is dependent on calcium mediated signaling 19. Thus, higher CD69 expression in wild
type ACH cells may have resulted from activation of p12I-mediated calcium signaling.
The role of CD69 in immune function is still largely unknown, due to the lack of
known ligands and a suitable in vivo model 20. However, recent findings suggested that
CD69 plays a immunoregulatory function by increasing the production of transforming
growth factor-β (TGF-β) 20,21. Interestingly, TGF-β down-regulates antigen-presenting
cell (APC) function 22, suggesting a potential role of CD69 expression in escaping
immune surveillance during HTLV-1 pathogenesis. Further studies will be required to
understand the role of CD69 in HTLV-1 pathogenesis, particularly in escaping immune
surveillance as well as cell-to-cell transmission.
5.6 The potential role of p12I in regulation of viral gene expression during HTLV-1
cell-to-cell transmission
In Chapter 4, my data demonstrated that expression of HTLV-1 pX ORF
messages including Tax/Rex, p27I and p12I mRNA were differentially regulated during
early HTLV-1 transmission. Furthermore, Tax/Rex expression was enhanced by pX
ORF I expression and was modulated by the activation status of target cells. These

183
results suggest that Tax/Rex mRNA expression and subsequent Tax-mediated viral
production may be influenced by pX ORF I mRNA or encoded proteins during cell-to-
cell contact required for efficient HTLV-1 transmission. Therefore, studies designed to
identify stimuli and their corresponding signaling pathways modulating viral gene
expression during early HTLV-1 cell-to-cell transmission will be required to more
completely understand molecular mechanisms of HTLV-1 early cell-to-cell
transmission. Moreover, a number of reports have been identified a role of Tax in
transcriptional regulation of expression of viral and cellular genes 23, however, the
effect of other viral ORF proteins in Tax expression has not been reported.
HTLV-1 pX ORF I encoded p12I is a modulator of calcium signal transduction
1,3-5, suggesting a role of viral gene regulation during early viral transmission.
Interestingly, calcium-mediated signaling induced activation of LTR-mediated
transcription and viral structural protein synthesis in cells infected with bovine
leukemia virus (BLV), another deltaretrovirus 24. Thus further studies are required to
directly test a role of calcium-mediated signaling in the regulation of HTLV-1 gene
expression in particularly during HTLV-1 cell-to-cell transmission. Moreover, other
signaling pathways, particularly, signaling via CD2 and ICAM-125,26, which enhanced
viral replication, may directly enhance Tax/Rex expression during early viral
transmission. Furthermore, a more complete understanding of cell signaling pathways
that modulate viral gene expression during viral transmission would be useful for
development of drugs that are designed to block early viral transmission events.

184
5.7 Studies to test the influence of pX OFR I or p12I in early expression of HTLV-1
regulatory proteins
Our data in Chapter 4 indicated that de novo Tax/Rex mRNA expression was
enhanced in cells expressing pX ORF I, suggesting a role of pX ORF I mRNA or its
encoded proteins in establishing HTLV-1 infection in target cells. Furthermore, our data
is the first to indicate that pX ORF I is expressed early (within 12 h) of cell-to-cell
contact. Therefore, pX ORF I encoded p12I may influence the earliest events of the viral
life cycle directly or indirectly through its known cell activation properties and calcium
signaling properties. In this regard the early events of the viral life cycle such as virus
entry, uncoating, reverse transcription and provirus integration may be modulated by
p12I. However, if p12I influences these early events, the protein would need to be
present in viral pre-integration complexes or expressed early following initial
integration events. Similarly, the HIV accessory protein, Nef, is found as a protein in
HIV virions 27,28 and is selectively expressed before viral integration 29 . Because, some
functions of Nef are similar to that of p12I in T cell activation, it would be interesting to
test if p12I is present in HTLV-1 viral particles or is transferred during the virological
synapse formation. Moreover, tests to determine if p12I is expressed selectively before
viral integration or expressed early following initial integration, would be required for
understanding the role of p12I in early establishment of HTLV-1 infection. Further
studies using single-round infection assays, reverse transcriptase inhibitor or integrase
deficient proviral clones, would be helpful in identifying the function of p12I in the
early viral life cycle. Finally, direct tests to determine if calcium-mediated signaling
could affect HTLV-1 reverse transcription, integration and early viral gene expression

185
would be helpful to indirectly implicate p12I in de novo HTLV-1 infection in target
cells.
Overall, the data from this thesis have contributed to our understanding of the
role of pX ORF I and its encoded p12I in HTLV-1 infection. It appears that pX ORF I
mRNA or proteins are indeed not “accessory”, but essential in the control of the early
events of HTLV-1 infection in T cells.
5.8 REFERENCES
Reference List
(1) Albrecht B, D'Souza CD, Ding W et al. Activation of nuclear factor of activated T cells by human T- lymphotropic virus type 1 accessory protein p12(I). J Virol. 2002;76:3493-3501.
(2) Ding W, Albrecht B, Luo R et al. Endoplasmic Reticulum and cis-Golgi Localization of Human T- Lymphotropic Virus Type 1 p12(I): Association with Calreticulin and Calnexin. J Virol. 2001;75:7672-7682.
(3) Ding W, Albrecht B., Kelley RE et al. Human T lymphotropic virus type 1 p12(I) expression increases cytoplasmic calcium to enhance the activation of nuclear factor of activated T cells. J Virol. 2002;76:10374-10382.
(4) Albrecht B, Collins ND, Burniston MT et al. Human T-lymphotropic virus type 1 open reading frame I p12(I) is required for efficient viral infectivity in primary lymphocytes. J Virol. 2000;74:9828-9835.
(5) Collins ND, Newbound GC, Albrecht B et al. Selective ablation of human T-cell lymphotropic virus type 1 p12I reduces viral infectivity in vivo. Blood. 1998;91:4701-4707.
(6) Ding W, Kim SJ, Nair AM et al. Human T-Cell Lymphotropic Virus Type 1 p12(I) Enhances Interleukin-2 Production during T-Cell Activation. J Virol. 2003;77:11027-11039.

186
(7) Nair A, Michael B, Hiraragi H et al. Human T lymphotropic virus type 1 accessory protein p12I modulates calcium-mediated cellular gene expression and enhances p300 expression in T lymphocytes. AIDS Res Hum Retroviruses. 2005;21:273-284.
(8) Kim SJ, Ding W, Albrecht B, Green PL, Lairmore MD. A conserved calcineurin-binding motif in human T lymphotropic virus type 1 p12I functions to modulate nuclear factor of activated T cell activation. J Biol Chem. 2003;278:15550-15557.
(9) Barnard AL, Igakura T, Tanaka Y, Taylor GP, Bangham CR. Engagement of specific T cell surface molecules regulates cytoskeletal polarization in HTLV-1-infected lymphocytes. Blood. 2005.
(10) Kuhne MR, Lin J, Yablonski D et al. Linker for activation of T cells, zeta-associated protein-70, and Src homology 2 domain-containing leukocyte protein-76 are required for TCR-induced microtubule-organizing center polarization. J Immunol. 2003;171:860-866.
(11) Kimata JT, Palker TJ, Ratner L. The Mitogenic Activity of Human T-Cell Leukemia Virus Type- I Is T-Cell Associated and Requires the CD2/LFA-3 Activation Pathway. J Virol. 1993;67:3134-3141.
(12) Wucherpfennig KW, Hollsberg P, Richardson JH, Benjamin D, Hafler DA. T-cell activation by autologous human T-cell leukemia virus type I- infected T-cell clones. Proc Natl Acad Sci U S A. 1992;89:2110-2114.
(13) Kim SJ, Nair AM, Fernandez S, Mathes L, Lairmore MD. Enhancement of
LFA-1-mediated T cell adhesion by human T lymphotropic virus type 1 p12I1. J Immunol. 2006;176:5463-5470.
(14) Hogg N, Laschinger M, Giles K, Mcdowall A. T-cell integrins: more than just sticking points. J Cell Sci. 2003;116:4695-4705.
(15) Feigelson SW, Grabovsky V, Winter E et al. The Src kinase p56(lck) up-regulates VLA-4 integrin affinity. Implications for rapid spontaneous and chemokine-triggered T cell adhesion to VCAM-1 and fibronectin. J Biol Chem. 2001;276:13891-13901.
(16) Michael B, Nair AM, Hiraragi H et al. Human T lymphotropic virus type-1 p30II alters cellular gene expression to selectively enhance signaling pathways that activate T lymphocytes. Retrovirology. 2004;1:39.
(17) Johnson S, Michalak M, Opas M, Eggleton P. The ins and outs of calreticulin: from the ER lumen to the extracellular space. Trends Cell Biol. 2001;11:122-129.

187
(18) Braakman I, van Anken E. Folding of viral envelope glycoproteins in the endoplasmic reticulum. Traffic. 2000;1:533-539.
(19) Rodrigues MS, Echevarria-Lima J, Fernandes dos SN, Rumjanek VM. CD69 expression induced by thapsigargin, phorbol ester and ouabain on thymocytes is dependent on external Ca2+ entry. Life Sci. 2003;73:1037-1051.
(20) Sancho D, Gomez M, Sanchez-Madrid F. CD69 is an immunoregulatory molecule induced following activation. Trends Immunol. 2005;26:136-140.
(21) Sancho D, Gomez M, Viedma F et al. CD69 downregulates autoimmune reactivity through active transforming growth factor-beta production in collagen-induced arthritis. J Clin Invest. 2003;112:872-882.
(22) Kulkarni AB, Huh CG, Becker D et al. Transforming growth factor beta 1 null mutation in mice causes excessive inflammatory response and early death. Proc Natl Acad Sci U S A. 1993;90:770-774.
(23) Kashanchi F, Brady JN. Transcriptional and post-transcriptional gene regulation of HTLV-1. Oncogene. 2005;24:5938-5951.
(24) Bondzio A, Abraham-Podgornik A, Blankenstein P, Risse S. Involvement of intracellular Ca2+ in the regulation of bovine leukemia virus expression. Biol Chem. 2001;382:407-416.
(25) Yamamoto A, Hara H, Kobayashi T. Induction of the expression of gag protein in HTLV-I infected lymphocytes by anti-ICAM 1 antibody in vitro. J Neurol Sci. 1997;151:121-126.
(26) Guyot DJ, Newbound GC, Lairmore MD. Signaling via the CD2 receptor enhances HTLV-1 replication in T lymphocytes. Virology. 1997;234:123-129.
(27) Kotov A, Zhou J, Flicker P, Aiken C. Association of Nef with the human immunodeficiency virus type 1 core. J Virol. 1999;73:8824-8830.
(28) Zhou J, Aiken C. Nef enhances human immunodeficiency virus type 1 infectivity resulting from intervirion fusion: evidence supporting a role for Nef at the virion envelope. J Virol. 2001;75:5851-5859.
(29) Wu Y, Marsh JW. Selective transcription and modulation of resting T cell activity by preintegrated HIV DNA. Science. 2001;293:1503-1506.

188
BIBLIOGRAPHY
Adachi Y, Copeland TD, Takahashi C et al. Phosphorylation of the Rex protein of human T-cell leukemia virus type I. Journal of Biological Chemistry. 1992;267:21977-21981.
Adedayo O, Grell G, Bellot P. Hospital admissions for human T-cell lymphotropic virus type-1 (HTLV-1) associated diseases in Dominica. Postgrad Med J. 2003;79:341-344.
Ahmed YF, Gilmartin GM, Hanly SM, Nevins JR, Greene WC. The HTLV-1 Rex Response Element Mediates a Novel Form of mRNA Polyadenylation. Cell. 1991;64:727-737.
Alamy AH, Menezes FB, Leite AC, Nascimento OM, Araujo AQ. Dysautonomia in human T-cell lymphotrophic virus type I-associated myelopathy/tropical spastic paraparesis. Ann Neurol. 2001;50:681-685.
Albrecht B, Collins ND, Burniston MT et al. Human T-lymphotropic virus type 1 open reading frame I p12(I) is required for efficient viral infectivity in primary lymphocytes. J Virol. 2000;74:9828-9835.
Albrecht B, D'Souza CD, Ding W et al. Activation of nuclear factor of activated T cells by human T- lymphotropic virus type 1 accessory protein p12(I). J Virol. 2002;76:3493-3501.
Albrecht B, Lairmore MD. Critical role of human T-lymphotropic virus type 1 accessory proteins in viral replication and pathogenesis. Microbiol Mol Biol Rev. 2002;66:396-406.
Alfahim A, Cabre P, Kastrukoff L, Dorovinizis K, Oger J. Blood mononuclear cells in patients with HTLV-I-associated myelopathy: Lymphocytes are highly activated and adhesion to endothelial cells is increased. Cell Immunol. 1999;198:1-10.

189
Allen WE, Zicha D, Ridley AJ, Jones GE. A role for Cdc42 in macrophage chemotaxis. J Cell Biol. 1998;141:1147-1157.
Anderson DW, Epstein JS, Lee TH et al. Serological confirmation of human T-lymphotropic virus type I infection in healthy blood and plasma donors 5242. Blood. 1989;74:2585-2591.
Aramburu J, Garcia-Cozar F, Raghavan A et al. Selective inhibition of NFAT activation by a peptide spanning the calcineurin targeting site of NFAT. Mol Cell. 1998;1:627-637.
Aramburu J, Yaffe MB, Lopez-Rodriguez C et al. Affinity-driven peptide selection of an NFAT inhibitor more selective than cyclosporin A 5399. Science. 1999;285:2129-2133.
Aramburu J, Yaffe MB, Lopez-Rodriguez C et al. Affinity-driven peptide selection of an NFAT inhibitor more selective than cyclosporin A 5399. Science. 1999;285:2129-2133.
Aramburu J, Yaffe MB, Lopez-Rodriguez C et al. Affinity-driven peptide selection of an NFAT inhibitor more selective than cyclosporin A 5399. Science. 1999;285:2129-2133.
Aramburu J, Yaffe MB, Lopez-Rodriguez C et al. Affinity-driven peptide selection of an NFAT inhibitor more selective than cyclosporin A 5399. Science. 1999;285:2129-2133.
Aramburu J, Yaffe MB, Lopez-Rodriguez C et al. Affinity-driven peptide selection of an NFAT inhibitor more selective than cyclosporin A 5399. Science. 1999;285:2129-2133.
Aramburu J, Yaffe MB, Lopez-Rodriguez C et al. Affinity-driven peptide selection of an NFAT inhibitor more selective than cyclosporin A 5399. Science. 1999;285:2129-2133.
Ardouin L, Bracke M, Mathiot A et al. Vav1 transduces TCR signals required for LFA-1 function and cell polarization at the immunological synapse. Eur J Immunol. 2003;33:790-797.
Arias RA, Munoz LD, Munoz-Fernandez MA. Transmission of HIV-1 infection between trophoblast placental cells and T-cells take place via an LFA-1-mediated cell to cell contact. Virology. 2003;307:266-277.
Arisawa K, Soda M, Endo S et al. Evaluation of adult T-cell leukemia/lymphoma incidence and its impact on non-Hodgkin lymphoma incidence in southwestern Japan. Int J Cancer. 2000;85:319-324.

190
Arnold J, Yamamoto B, Li M et al. Enhancement of infectivity and persistence in vivo by HBZ, a natural antisense coded protein of HTLV-1. Blood. 2006.
Asher DM, Goudsmit J, Pomeroy K et al. Antibodies to HTLV-I in populations of the southwestern Pacific. J Med Virol. 1988;26:339.
Azimi N, Mariner J, Jacobson S, Waldmann TA. How does interleukin 15 contribute to the pathogenesis of HTLV type 1- associated Myelopathy/Tropical spastic paraparesis? AIDS Res Hum Retroviruses. 2000;16:1717-1722.
Baeuerle PA, Baltimore D. I kappa B: a specific inhibitor of the NF-kappa B transcription factor. Science. 1988;242:540-546.
Ballard DW. Molecular mechanisms in lymphocyte activation and growth. Immunol Res. 2001;23:157-166.
Ballaun C, Farrington GK, Dobrovnik M et al. Functional Analysis of Human T-Cell Leukemia Virus Type 1 rex-Response Element: Direct RNA Binding of Rex Protein Correlates with In Vivo Activity. J Virol. 1991;65:4408-4413.
Bangham CR. HTLV-1 infections. J Clin Pathol. 2000;53:581-586.
Bangham CR. Human T-lymphotropic virus type 1 (HTLV-1): persistence and immune control. Int J Hematol. 2003;78:297-303.
Bangham CR. The immune control and cell-to-cell spread of human T-lymphotropic virus type 1. J Gen Virol. 2003;84:3177-3189.
Barnard AL, Igakura T, Tanaka Y, Taylor GP, Bangham CR. Engagement of specific T cell surface molecules regulates cytoskeletal polarization in HTLV-1-infected lymphocytes. Blood. 2005.
Barnhart MK, Connor LM, Marriott SJ. Function of the human T-cell leukemia virus type 1 21-base-pair repeats in basal transcription. J Virol. 1997;71:337-344.
Barshira A, Panet A, Honigman A. An RNA Secondary Structure Juxtaposes Two Remote Genetic Signals for Human T-Cell Leukemia Virus Type 1 RNA 3'-End Processing. J Virol. 1991;65:5165-5173.
Bartoe JT, Albrecht B, Collins ND et al. Functional role of pX open reading frame II of human T- lymphotropic virus type 1 in maintenance of viral loads in vivo. J Virol. 2000;74:1094-1100.
Basbous J, Arpin C, Gaudray G et al. The HBZ factor of human T-cell leukemia virus type I dimerizes with transcription factors JunB and c-Jun and modulates their transcriptional activity. J Biol Chem. 2003;278:43620-43627.

191
Bazarbachi A, Hermine O. Treatment of adult T-cell leukaemia/lymphoma: current strategy and future perspectives. Virus Res. 2001;78:79-92.
Benovic S, Kok T, Stephenson A et al. De novo reverse transcription of HTLV-1 following cell-to- cell transmission of infection. Virology. 1998;244:294-301.
Beraud C, Lombard-Platet G, Michal Y, Jalinot P. Binding of the HTLV-I Tax1 transactivator to the inducible 21 bp enhancer is mediated by the cellular factor HEB1 5256. EMBO J. 1991;10:3795-3803.
Berneman ZN, Gartenhaus RB, Reitz MS et al. Expression of alternatively spliced human T-lymphotropic virus type 1 pX mRNA in infected cell lines and in primary uncultured cells from patients with adult T-cell leukemia/lymphoma and healthy carriers. Proc Natl Acad Sci USA. 1992;89:3005-3009.
Bex F, Gaynor RB. Regulation of gene expression by HTLV-I Tax protein. Methods. 1998;16:83-94.
Bex F, Yin MJ, Burny A, Gaynor RB. Differential transcriptional activation by human T-cell leukemia virus type 1 Tax mutants is mediated by distinct interactions with CREB binding protein and p300. Mol Cell Biol. 1998;18:2392-2405.
Bindhu M, Nair A, Lairmore MD. Role of accessory proteins of HTLV-1 in viral replication, T cell activation, and cellular gene expression. Front Biosci. 2005;10:17-37.
Black AC, Chen ISY, Arrigo S et al. Regulation of HTLV-II gene expretion by rex involves positive and negative cis-Acting elements in the 5 long terminal repeat. Virology. 1991;181:433-444.
Black AC, Ruland C, Yip MT et al. Human T-Cell Leukemia Virus Type II Rex Binding and Activity Require an Intact splice Donor Site and a Specific RNA Secondary Structure. J Virol. 1991;65:6645-6653.
Black AC, Ruland CT, Luo J et al. Binding of Nuclear Proteins to HTLV-II Cis-Acting Repressive Sequence (CRS) RNA Correlates with CRS Function. Virology. 1994;200:29-41.
Blattner W, Kalyanaraman V, Robert-Guroff M et al. The human type-C retrovirus, HTLV, in blacks from the Caribbean region and relationchip to adult T-cell leukemia/lymphoma. Int J Cancer. 1982;30:257.
Blayney DW, Blattner WA, Robert-Guroff M et al. The human T-cell leukemia-lymphoma virus in the southeastern United States 5419. JAMA. 1983;250:1048-1052.

192
Blot V, Perugi F, Gay B et al. Nedd4.1-mediated ubiquitination and subsequent recruitment of Tsg101 ensure HTLV-1 Gag trafficking towards the multivesicular body pathway prior to virus budding. J Cell Sci. 2004;117:2357-2367.
Bogerd HP, Echarri A, Ross TM, Cullen BR. Inhibition of human immunodeficiency virus Rev and human T- cell leukemia virus Rex function, but not Mason-Pfizer monkey virus constitutive transport element activity, by a mutant human nucleoporin targeted to Crm1. J Virol. 1998;72:8627-8635.
Bogerd HP, Huckaby GL, Ahmed YF, Hanly SM, Greene WC. The type I human T-cell leukemia virus (HTLV-I) Rex trans-activator binds directly to the HTLV-I Rex and the type 1 human immunodeficiency virus Rev RNA response elements. Proc Natl Acad Sci U S A. 1991;88:5704-5708.
Bondzio A, Abraham-Podgornik A, Blankenstein P, Risse S. Involvement of intracellular Ca2+ in the regulation of bovine leukemia virus expression. Biol Chem. 2001;382:407-416.
Bosselut R, Duvall JF, Gegonne A et al. The product of the c-ets-1 proto-oncogene and the related Ets2 protein act as transcriptional activators of the long terminal repeat of human T cell leukemia virus HTLV-1 5254. EMBO J. 1990;9:3137-3144.
Bosselut R, Lim F, Romond PC et al. Myb protein binds to multiple sites in the human T cell lymphotropic virus type 1 long terminal repeat and transactivates LTR-mediated expression 5258. Virology. 1992;186:764-769.
Bouchard MJ, Wang LH, Schneider RJ. Calcium signaling by HBx protein in hepatitis B virus DNA replication. Science. 2001;294:2376-2378.
Bounou S, Giguere JF, Cantin R et al. The importance of virus-associated host ICAM-1 in human immunodeficiency virus type 1 dissemination depends on the cellular context. FASEB J. 2004;18:1294-1296.
Boustany LM, Cyert MS. Calcineurin-dependent regulation of Crz1p nuclear export requires Msn5p and a conserved calcineurin docking site. Genes Dev. 2002;16:608-619.
Boustany LM, Cyert MS. Calcineurin-dependent regulation of Crz1p nuclear export requires Msn5p and a conserved calcineurin docking site. Genes Dev. 2002;16:608-619.
Braakman I, van Anken E. Folding of viral envelope glycoproteins in the endoplasmic reticulum. Traffic. 2000;1:533-539.

193
Brady JN, Jeang KT, Duvall J, Khoury G. Identification of p40x-Responsive Regulatory Sequences within the Human T-Cell Leukemia Virus Type 1 Long Terminal Repeat. J Virol. 1987;61:2175-2181.
Brauweiler A, Garrus JE, Reed JC, Nyborg JK. Repression of Bax gene expression by the HTLV-I tax protein: Implications for suppression of apoptosis in virally infected cells. Virology. 1997;231:135-140.
Bukrinsky MI, Sharova N, Dempsey MP et al. Active nuclear import of human immunodeficiency virus type 1 preintegration complexes. Proc Natl Acad Sci U S A. 1992;89:6580-6584.
Burton M, Upadhyaya CD, Maier B, Hope TJ, Semmes OJ. Human T-cell leukemia virus type 1 tax shuttles between functionally discrete subcellular targets. J Virol. 2000;74:2351-2364.
Cann AJ, Chen ISY. Human T-Cell Leulemia Virus Types I and II. 1849-1880. 1996. Philadelphia, PA, Lippincott-Raven Publishers. Ref Type: Generic
Cann AJ, Rosenblatt JD, Wachsman W, Shah NP, Chen IS. Identification of the gene responsible for human T-cell leukaemia virus transcriptional regulation 5264. Nature. 1985;318:571-574.
Carman CV, Springer TA. Integrin avidity regulation: are changes in affinity and conformation underemphasized? Curr Opin Cell Biol. 2003;15:547-556.
Castillo LC, Gracia F, Roman GC et al. Spinocerebellar syndrome in patients infected with human T-lymphotropic virus types I and II (HTLV-I/HTLV-II): report of 3 cases from Panama. Acta Neurol Scand. 2000;101:405-412.
Cavrois M, Leclercq I, Gout O et al. Persistent oligoclonal expansion of human T-cell leukemia virus type 1 infected circulating cells in patients with Tropical spastic paraparesis/HTLV-1 associated myelopathy. Oncogene. 1998;17:77-82.
Cereseto A, Berneman Z, Koralnik I et al. Differential expression of alternatively spliced pX mRNAs in HTLV-I-infected cell lines. Leukemia. 1997;11:866-870.
Cereseto A, Giacca M. Integration site selection by retroviruses. AIDS Rev. 2004;6:13-21.
Cereseto A, Kislyakova T, Washington-Parks R, Franchini G. Differential response to genotoxic stress in immortalized or transformed HTLV-I-infected T-cells. J Gen Virol. 1999;80:1575-1581.

194
Chan JR, Hyduk SJ, Cybulsky MI. Chemoattractants induce a rapid and transient upregulation of monocyte alpha4 integrin affinity for vascular cell adhesion molecule 1 which mediates arrest: an early step in the process of emigration. J Exp Med. 2001;193:1149-1158.
Chen YA, Chen S, Fu C, Chen J, Osame M. Antibody reactivities to tumor-suppressor protein p53 and HTLV-I TOF, REX, and TAX in HTLV-I-infected people with differing clinical status. Int J Cancer. 1997;71:196-202.
Chou KS, Okayama A, Tachibana N, Lee TH, Essex M. Nucleotide sequence analysis of a full-length human T-cell leukemia virus type I from adult T-cell leukemia cells: A prematurely terminated pX open reading frame II. Int J Cancer. 1995;60:701-706.
Chrivia JC, Kwok RP, Lamb N et al. Phosphorylated CREB binds specifically to the nuclear protein CBP. Nature. 1993;365(6449):855-859.
Ciminale V, D'Agostino D, Zotti L et al. Expression and characterization of proteins produced by mRNAs spliced into the X region of the human T-cell leukemia/lymphotropic virus type II. Virology. 1995;209:445-456.
Ciminale V, D'Agostino DM, Zotti L, Chieco-Bianchi L. Coding potential of the X region of human T-cell leukemia/lymphotropic virus type II. J Acquir Immune Defic Syndr Hum Retrovirol. 1996;13 Suppl 1:S220-S227.
Ciminale V, Pavlakis GN, Derse D, Cunningham CP, Felber BK. Complex splicing in the human T-cell leukemia virus (HTLV) family of retroviruses: Novel mRNAs and proteins produced by HTLV type I. J Virol. 1992;66:1737-1745.
Ciminale V, Zotti L, Dagostino DM et al. Mitochondrial targeting of the p13(II) protein coded by the x-II ORF of human T-cell leukemia/lymphotropic virus type I (HTLV-I). Oncogene. 1999;18:4505-4514.
Clark NM, Smith MJ, Hilfinger JM, Markovitz DM. Activation of the human T-cell leukemia virus type I enhancer is mediated by binding sites for Elf-1 and the pets factor. J Virol. 1993;67:5522-5528.
Coffin JM. Structure and Classification of Retroviruses. The Retroviridae. 1992;1:19-49.
Coffin JM. Retroviridae: The Viruses and Their Replication. 1767-1835. 1996. Phildaelphia, PA, Lippinocott-Raven Publishers. Ref Type: Generic
Colgin MA, Nyborg JK. The human T-cell leukemia virus type 1 oncoprotein Tax inhibits the transcriptional activity of c-Myb through competition for the CREB binding protein. J Virol. 1998;72:9396-9399.

195
Collins ND, D'Souza C, Albrecht B et al. Proliferation response to interleukin-2 and Jak/Stat activation of T cells immortalized by human T-cell lymphotropic virus type 1 is independent of open reading frame I expression. J Virol. 1999;73:9642-9649.
Collins ND, Newbound GC, Albrecht B et al. Selective ablation of human T-cell lymphotropic virus type 1 p12(I) reduces viral infectivity in vivo. Blood. 1998;91:4701-4707.
Collins ND, Newbound GC, Albrecht B et al. Selective ablation of human T-cell lymphotropic virus type 1 p12I reduces viral infectivity in vivo. Blood. 1998;91:4701-4707.
Collins ND, Newbound GC, Ratner L, Lairmore MD. In vitro CD4(+) lymphocyte transformation and infection in a rabbit model with a molecular clone of human T-cell lymphotropic virus type 1. J Virol. 1996;70:7241-7246.
Constantin G, Majeed M, Giagulli C et al. Chemokines trigger immediate beta2 integrin affinity and mobility changes: differential regulation and roles in lymphocyte arrest under flow. Immunity. 2000;13:759-769.
Courouce AM, Pillonel J, Lemaire JM, Saura C. HTLV testing in blood transfusion. Vox Sang. 1998;74 Suppl 2:165-169.
Cowan EP, Alexander RK, Daniel S, Kashanchi F, Brady JN. Induction of tumor necrosis factor alpha in human neuronal cells by extracellular human T-cell lymphotropic virus type 1 Tax(1). J Virol. 1997;71:6982-6989.
Craig HM, Reddy TR, Riggs NL, Dao PP, Guatelli JC. Interactions of HIV-1 nef with the mu subunits of adaptor protein complexes 1, 2, and 3: role of the dileucine-based sorting motif. Virology. 2000;271:9-17.
Craven RC, Parent LJ. Dynamic interactions of the gag polyprotein. In: Krausslich HG, ed. Morphogenesis and Maturation of Retroviruses. Springer-Verlag Berl; 1996:65-94.
D'Agostino DM, Ciminale V, Zotti L, Rosato A, Chieco-Bianchi L. The human T-cell lymphotropic virus type 1 Tof protein contains a bipartite nuclear localization signal that is able to functionally replace the amino-terminal domain of Rex. J Virol. 1997;71:75-83.
D'Agostino DM, Ranzato L, Arrigoni G et al. Mitochondrial alterations induced by the p13II protein of human T-cell leukemia virus type 1. Critical role of arginine residues. J Biol Chem. 2002;277:34424-34433.
D'Agostino DM, Silic-Benussi M, Hiraragi H, Lairmore MD, Ciminale V. The human T-cell leukemia virus type 1 p13(II) protein: effects on mitochondrial function and cell growth. Cell Death Differ. 2005.

196
D'Agostino DM, Zotti L, Ferro T et al. Expression and functional properties of proteins encoded in the x-II ORF of HTLV-I. Virus Res. 2001;78:35-43.
D'Agostino DM, Zotti L, Ferro T et al. The p13II protein of HTLV type 1: comparison with mitochondrial proteins coded by other human viruses. AIDS Res Hum Retroviruses. 2000;16:1765-1770.
D'Souza-Schorey C, Boettner B, Van AL. Rac regulates integrin-mediated spreading and increased adhesion of T lymphocytes. Molecular & Cellular Biology. 1998;18:3936-3946.
Daenke S, Mccracken SA, Booth S. Human T-cell leukaemia lymphoma virus type 1 syncytium formation is regulated in a cell-specific manner by ICAM-1, ICAM-3 and VCAM-1 and can be inhibited by antibodies to integrin beta(2) or beta(7). J Gen Virol. 1999;80:1429-1436.
Daenke S, Nightingale S, Cruickshank JK, Bangham CRM. Sequence variants of human T-cell lymphotropic virus type I from patients with tropical spastic paraparesis and adult T-cell leukemia do not distinguish neurological from leukemic isolates. J Virol. 1990;64:1278-1282.
Dekaban GA, Peters AA, Mulloy JC et al. The HTLV-I orfI protein is recognized by serum antibodies from naturally infected humans and experimentally infected rabbits. Virology. 2000;274:86-93.
Delamarre L, Rosenberg AR, Pique C, Pham D, Dokhelar MC. A novel human T-Leukemia virus type 1 cell-to-cell transmission assay permits definition of SU glycoprotein amino acids important for infectivity. J Virol. 1997;71:259-266.
Derse D. Bovine leukemia virus transcription is controlled by a virus-encoded trans-acting factor and by cis-acting response elements. J Virol. 1987;61:2462-2471.
Derse D, Mikovits J, Ruscetti F. X-I and X-II open reading frames of HTLV-I are not required for virus replication or for immortalization of primary T-cells in vitro. Virology. 1997;237:123-128.
Ding W, Albrecht B., Kelley RE et al. Human T lymphotropic virus type 1 p12(I) expression increases cytoplasmic calcium to enhance the activation of nuclear factor of activated T cells. J Virol. 2002;76:10374-10382.
Ding W, Albrecht B, Luo R et al. Endoplasmic Reticulum and cis-Golgi Localization of Human T- Lymphotropic Virus Type 1 p12(I): Association with Calreticulin and Calnexin. J Virol. 2001;75:7672-7682.

197
Ding W, Kim SJ, Nair AM et al. Human T-Cell Lymphotropic Virus Type 1 p12(I) Enhances Interleukin-2 Production during T-Cell Activation. J Virol. 2003;77:11027-11039.
Dustin M. Viral spread through protoplasmic kiss. Nat Cell Biol. 2003;5:271-272.
Dustin ML, Bivona TG, Philips MR. Membranes as messengers in T cell adhesion signaling. Nat Immunol. 2004;5:363-372.
Duyao MP, Kessler DJ, Spicer DB et al. Transactivation of the c-myc Promoter by Human T Cell Leukemia virus Type 1 tax is Mediated by NFkB. J Biol Chem. 1992;267:16288-16291.
Eckner R, Yao TP, Oldread E, Livingston DM. Interaction and functional collaboration of p300/CBP and bHLH proteins in muscle and B-cell differentiation. Genes Dev. 1996;10:2478-2490.
Edlich RF, Arnette JA, Williams FM. Global epidemic of human T-cell lymphotropic virus type-I (HTLV-I). J Emerg Med. 2000;18:109-119.
Evangelista A, Maroushek S, Minnigan H et al. Nucleotide sequence analysis of a provirus derived from an individual with tropical spastic paraparesis. Microb Pathog. 1990;8:259-278.
Fagerholm S, Morrice N, Gahmberg CG, Cohen P. Phosphorylation of the cytoplasmic domain of the integrin CD18 chain by protein kinase C isoforms in leukocytes. J Biol Chem. 2002;277:1728-1738.
Fan N, Gavalchin J, Paul B et al. Infection of Peripheral Blood Mononuclear Cells and Cell Lines by Cell-Free Human T-Cell Lymphoma/Leukemia Virus Type 1. J Clin Microbiol. 1992;30:905-910.
Feigelson SW, Grabovsky V, Winter E et al. The Src kinase p56(lck) up-regulates VLA-4 integrin affinity. Implications for rapid spontaneous and chemokine-triggered T cell adhesion to VCAM-1 and fibronectin. J Biol Chem. 2001;276:13891-13901.
Felber BK, Paskalis H, Kleinmanewing C. The pX Protein of HTLV-1 is a Transcriptional Activator of its Long Terminal Repeats. Science. 1985;229:675-679.
Feng P, Park J, Lee BS et al. Kaposi's sarcoma-associated herpesvirus mitochondrial K7 protein targets a cellular calcium-modulating cyclophilin ligand to modulate intracellular calcium concentration and inhibit apoptosis. J Virol. 2002;76:11491-11504.
Ferreira OC, Planelles V, Rosenblatt JD. Human T-cell leukemia viruses: Epidemiology, biology, and pathogenesis. Blood Rev. 1997;11:91-104.

198
Fortin JF, Barbeau B, Hedman H, Lundgren E, Tremblay MJ. Role of the leukocyte function antigen-1 conformational state in the process of human immunodeficiency virus type 1-mediated syncytium formation and virus infection. Virology. 1999;257:228-238.
Fortin JF, Cantin R, Tremblay MJ. T cells expressing activated LFA-1 are more susceptible to infection with human immunodeficiency virus type 1 particles bearing host-encoded ICAM-1. J Virol. 1998;72:2105-2112.
Foyouzi-Youssefi R, Arnaudeau S, Borner C et al. Bcl-2 decreases the free Ca2+ concentration within the endoplasmic reticulum. Proc Natl Acad Sci U S A. 2000;97:5723-5728.
Foyouzi-Youssefi R, Arnaudeau S, Borner C et al. Bcl-2 decreases the free Ca2+ concentration within the endoplasmic reticulum. Proc Natl Acad Sci U S A. 2000;97:5723-5728.
Foyouzi-Youssefi R, Arnaudeau S, Borner C et al. Bcl-2 decreases the free Ca2+ concentration within the endoplasmic reticulum. Proc Natl Acad Sci U S A. 2000;97:5723-5728.
Foyouzi-Youssefi R, Arnaudeau S, Borner C et al. Bcl-2 decreases the free Ca2+ concentration within the endoplasmic reticulum. Proc Natl Acad Sci U S A. 2000;97:5723-5728.
Foyouzi-Youssefi R, Arnaudeau S, Borner C et al. Bcl-2 decreases the free Ca2+ concentration within the endoplasmic reticulum. Proc Natl Acad Sci U S A. 2000;97:5723-5728.
Franchini G. Molecular mechanisms of human T-cell leukemia/lymphotropic virus type I infection. Blood. 1995;86:3619-3639.
Franchini G, Fukumoto R, Fullen JR. T-cell control by human T-cell leukemia/lymphoma virus type 1. Int J Hematol. 2003;78:280-296.
Franchini G, Mulloy JC, Koralnik IJ et al. The human T-cell leukemia/lymphotropic virus type I p12I protein cooperates with the E5 oncoprotein of bovine papillomavirus in cell transformation and binds the 16-kilodalton subunit of the vacuolar H+ ATPase. Journal of Virology. 1993;67:7701-7704.
Franchini G, Nicot C, Johnson JM. Seizing of T cells by human T-cell leukemia/lymphoma virus type 1. Adv Cancer Res. 2003;89:69-132.
Franklin AA, Kubik MF, Uittenbogaard MN et al. Transactivation by the human T-cell leukemia virus Tax protein is mediated through enhanced binding of activating

199
transcription factor-2 (ATF-2) ATF-2 response and cAMP element-binding protein (CREB). J Biol Chem. 1993;268:21225-21231.
Fujii M, Sassone-Corsi P, Verma I. c-fos promotor trans-activation of cAMP on IL-2 stimulated gene expression. Proc Natl Acad Sci U S A. 1988;8526-8530.
Fujii M, Tsuchiya H, Chuhjo T, Akizawa T, Seiki M. Interaction of HTLV-1 Tax1 with p67SRF causes the aberrant induction of cellular immediate early genes through CArG boxes 5270. Genes Dev. 1992;6:2066-2076.
Fujino T, Fujiyoshi T, Yashiki S et al. HTLV-I transmission from mother to fetus via placenta. Lancet. 1992;340:1157. Fujisawa J, Seiki M, Kiyokawa T, Yoshida M. Functional activation of the long terminal repeat of human T-cell leukemia virus type 1 by a trans-acting factor. Proc Natl Acad Sci USA. 1985;82:2277-2281.
Fukudome K, Furuse M, Fukuhara N et al. Strong induction of ICAM-1 in human T cells transformed by human T-cell-leukemia virus type 1 and depression of ICAM-1 or LFA-1 in adult T-cell-leukemia-derived cell lines. Int J Cancer. 1992;52:418-427.
Furukawa K, Furukawa K, Shiku H. Alternatively spliced mRNA of the pX region of human T lymphotropic virus type I proviral genome. FEBS Lett. 1991;295:141-145.
Furukawa Y, Kubota R, Eiraku N et al. Human T-cell lymphotropic virus type I (HTLV-I)-related clinical and laboratory findings for HTLV-I-infected blood donors. J Acquir Immune Defic Syndr. 2003;32:328-334.
Ganem D. Virology. The X files--one step closer to closure. Science. 2001;294:2299-2300.
Garcia-Cozar FJ, Okamura H, Aramburu JF et al. Two-site interaction of nuclear factor of activated T cells with activated calcineurin. J Biol Chem. 1998;273:23877-23883.
Garnier L, Bowzard JB, Wills JW. Recent advances and remaining problems in HIV assembly. AIDS. 1998;12 Suppl A:S5-16.
Garry RF. Sequence similarities between latent membrane protein LMP- 1 of Epstein-Barr virus, integral membrane protein p12(I) of human T cell leukemia lymphotropic virus type 1, E5 transformation protein of bovine papillomavirus, and the transmembrane proteins of slowly transforming retroviruses. AIDS Res Hum Retroviruses. 1995;11:431-432.
Gatza ML, Watt JC, Marriott SJ. Cellular transformation by the HTLV-I Tax protein, a jack-of-all-trades. Oncogene. 2003;22:5141-5149.

200
Gaudray G, Gachon F, Basbous J et al. The complementary strand of the human T-cell leukemia virus type 1 RNA genome encodes a bZIP transcription factor that down-regulates viral transcription. J Virol. 2002;76:12813-12822.
Gerritsen ME, Williams AJ, Neish AS et al. CREB-binding protein/p300 are transcriptional coactivators of p65. Proc Natl Acad Sci U S A. 1997;94:2927-2932.
Gessain A, Barin F, Vernant J et al. Antibodies to human T lymphotropic virus type 1 in patients with tropical spastic paresis. Lancet. 1985;2:407-410.
Gessain A, Mahieux R. [A virus called HTLV-1. Epidemiological aspects] Presse Med. 2000;29:2233-2239.
Giguere JF, Tremblay MJ. Statin compounds reduce human immunodeficiency virus type 1 replication by preventing the interaction between virion-associated host intercellular adhesion molecule 1 and its natural cell surface ligand LFA-1. J Virol. 2004;78:12062-12065.
Gould KG, Bangham CR. Virus variation, escape from cytotoxic T lymphocytes and human retroviral persistence. Semin Cell Dev Biol. 1998;9:321-328.
Grant C, Barmak K, Alefantis T et al. Human T cell leukemia virus type I and neurologic disease: events in bone marrow, peripheral blood, and central nervous system during normal immune surveillance and neuroinflammation. J Cell Physiol. 2002;190:133-159.
Grassmann R, Berchtold S, Aepinus C et al. In Vitro Binding of Human T-Cell Leukemia Virus rex Proteins to the rex-Response Element of Viral Transcripts. J Virol. 1991;65:3721-3727.
Green PL, Chen ISY. Regulation of human T cell leukemia virus expression. FASEB J. 1990;4:169-175.
Green PL, Chen ISY. Molecular features of the human T-cell leukemia virus. Mechanisms of transformation and leukemogenicity. In: Levy JA, ed. The Retroviridae.3. New York: Plenum Press; 1994:277-311.
Griffith JP, Kim JL, Kim EE et al. X-ray structure of calcineurin inhibited by the immunophilin-immunosuppressant FKBP12-FK506 complex. Cell. 1995;82:507-522.
Griffiths EK, Krawczyk C, Kong YY et al. Positive regulation of T cell activation and integrin adhesion by the adapter Fyb/Slap. Science. 2001;293:2260-2263.
Grone M, Koch C, Grassmann R. The HTLV-1 Rex protein induces nuclear accumulation of unspliced viral RNA by avoiding intron excision and degradation. Virology. 1996;218:316-325.

201
Guyot DJ, Newbound GC, Lairmore MD. Signaling via the CD2 receptor enhances HTLV-1 replication in T lymphocytes. Virology. 1997;234:123-129.
Hai TW, Liu F, Coukos WJ, Green MR. Transcription factor ATF cDNA clones: an extensive family of leucine zipper proteins able to selectively form DNA-binding heterodimers. Genes Dev. 1989;3:2083-2090.
Hakata Y, Umemoto T, Matsushita S, Shida H. Involvement of human CRM1 (exportin 1) in the export and multimerization of the Rex protein of human T-cell leukemia virus type 1. J Virol. 1998;72:6602-6607.
Hall WW, Fujii M. Deregulation of cell-signaling pathways in HTLV-1 infection. Oncogene. 2005;24:5965-5975.
Hanon E, Hall S, Taylor GP et al. Abundant Tax protein expression in CD4+T cells infected with human T-cell lymphotropic virus type I (HTLV-I) is prevented by cytotoxic T lymphocytes. Blood. 2000;95:1386-1392.
Harrod R, Kuo YL, Tang Y et al. p300 and p300/cAMP-responsive element-binding protein associated factor interact with human T-cell lymphotropic virus type-1 Tax in a multi- histone acetyltransferase/activator-enhancer complex. J Biol Chem. 2000;275:11852-11857.
Hattori S, Kiyokawa T, Imagawa K et al. Identification of gag and env gene products of human T-cell leukemia virus (HTLV). Virology. 1984;136:338-347.
He H, Lam M, McCormick TS, Distelhorst CW. Maintenance of calcium homeostasis in the endoplasmic reticulum by Bcl-2. J Cell Biol. 1997;138:1219-1228.
Hildreth JEK, Subramanium A, Hampton RA. Human T-cell lymphotropic virus type 1 (HTLV-1)-induced syncytium formation mediated by vascular cell adhesion molecule-1: Evidence for involvement of cell adhesion molecules in HTLV-1 biology. J Virol. 1997;71:1173-1180.
Hino S. Milk-borne transmission of HTLV-I as a major route in the endemic cycle. Acta Paediatr Jpn. 1989;31:428-435. Hino S, Yamaguchi K, Katamine S et al. Mother-to-child transmission of human T-cell leukemia virus type-I. Jpn J Cancer Res. 1985;76:474-480. Hinuma Y, Komoda H, Chosa T et al. Antibodies to adult T-cell leukemia virus-associated antigen (ATLA) in sera from patients with ATL and controls in Japan: a nation-wide seroepidemiological study. Int J Cancer. 1982;29:631.

202
Hinuma Y, Nagata K, Hanaoka M et al. Adult T Cell Leukemia: Antigens in an ATL cell line and detection of antibodies to antigen in human sera. Proc Natl Acad Sci U S A. 1981;78:6476-6480.
Hirai H, Suzuki T, Fujisawa J, Inoue J, Yoshida M. Tax Protein of Human T-Cell Leukemia Virus Type I Binds to the Ankyrin Motifs of Inhibitory Factor kappa b and Induces Nuclear Translocation of Transcription Factor NF- kappa b Proteins for Transcriptional Activation. Proc Natl Acad Sci USA. 1994;91:3584-3588.
Hiraragi H, Kim SJ, Phipps AJ et al. Human T-Lymphotropic Virus Type 1 Mitochondrion-Localizing Protein p13II Is Required for Viral Infectivity In Vivo. J Virol. 2006;80:3469-3476.
Hiraragi H, Michael B, Nair A et al. Human T-lymphotropic virus type 1 mitochondrion-localizing protein p13II sensitizes Jurkat T cells to Ras-mediated apoptosis. J Virol. 2005;79:9449-9457.
Hivin P, Frederic M, rpin-Andre C et al. Nuclear localization of HTLV-I bZIP factor (HBZ) is mediated by three distinct motifs. J Cell Sci. 2005;118:1355-1362.
Hogg N, Henderson R, Leitinger B et al. Mechanisms contributing to the activity of integrins on leukocytes. Immunol Rev. 2002;186:164-171.
Hogg N, Laschinger M, Giles K, Mcdowall A. T-cell integrins: more than just sticking points. J Cell Sci. 2003;116:4695-4705.
Hogg N, Smith A, Mcdowall A et al. How T cells use LFA-1 to attach and migrate. Immunol Lett. 2004;92:51-54.
Holloway MP, Bram RJ. A hydrophobic domain of Ca2+-modulating cyclophilin ligand modulates calcium influx signaling in T lymphocytes. J Biol Chem. 1996;271:8549-8552.
Holloway MP, Bram RJ. Co-localization of calcium-modulating cyclophilin ligand with intracellular calcium pools. J Biol Chem. 1998;273:16346-16350.
Hollsberg P. Mechanisms of T-cell activation by human T-cell lymphotropic virus type I. Microbiol Mol Biol Rev. 1999;63:308-333.
Hollsberg P, Hafler D. Pathogenesis of diseases induced by human lymphotropic virus type I infection. N Engl J Med. 1993;328:1173-1182.
Hollsberg P, Wucherpfennig KW, Ausubel LJ et al. Characterization of HTLV-1 In Vivo Infected T Cell Clones IL-2-independent Growth of Nontransformed T Cells. J Immunol. 1992;148:3256-3263.

203
Hou X, Foley S, Cueto M, Robinson MA. The Human T-Cell Leukemia Virus Type I (HTLV-I) X Region Encoded Protein p13(II) Interacts with Cellular Proteins. Virology. 2000;277:127-135.
Hummel KB, Bellini WJ, Offermann MK. Strain-specific differences in LFA-1 induction on measles virus-infected monocytes and adhesion and viral transmission to endothelial cells. J Virol. 1998;72:8403-8407.
Igakura T, Stinchcombe JC, Goon PK et al. Spread of HTLV-I between lymphocytes by virus-induced polarization of the cytoskeleton. Science. 2003;299:1713-1716.
Imai T, Tanaka Y, Fukudome K et al. Enhanced expression of LFA-3 on human T-cell lines and leukemic. International Journal of Cancer. 1993;55(5):811-816.
Iniguez AM, Otsuki K, Magalhaes GP, Silva EA, Vicente AC. Genetic markers on the HTLV-1 p12I protein sequences from Brazilian HAM/TSP patients and asymptomatic HTLV-1 carrier isolates. AIDS Res Hum Retroviruses. 2005;21:580-582.
Inoue J, Seiki M, Taniguchi T, Tsuru S, Yoshida M. Induction of interleukin 2 receptor gene expression by p40x encoded by human T-cell leukemia virus type 1. EMBO J. 1986;5:2883-2888.
Inoue J, Seiki M, Yoshida M. The second pX product p27 chi-III of HTLV-1 is required for gag gene expression 5294. FEBS Lett. 1986;209:187-190.
Inoue J, Yoshida M, Seiki M. Transcriptional (p40x) and post-transcriptional (p27x-III) regulators are required for the expression and replication of human t-cell leukemia virus type I genes. Proc Natl Acad Sci USA. 1987;84:3653-3657.
Itoyama Y, Minato S, Kira J et al. Spontaneous proliferation of peripheral blood lymphocytes increased in patients with HTLV-I-associated myelopathy. Neurology. 1988;38:1302-1307.
Jacobson S. Immunopathogenesis of human T cell lymphotropic virus type I-associated neurologic disease. J Infect Dis. 2002;186 Suppl 2:S187-S192.
Jacobson S, Zaninovic V, Mora C et al. Immunological findings in neurological diseases associated with antibodies to HTLV-I: activated lymphocytes in tropical spastic paraparesis. Ann Neurol. 1988;23 Suppl:S196-S200.
Jassal SR, Lairmore MD, Leigh-Brown AJ, Brighty DW. Soluble recombinant HTLV-1 surface glycoprotein competitively inhibits syncytia formation and viral infection of cells. Virus Res. 2001;78:17-34.

204
Jeang KT. Functional activities of the human T-cell leukemia virus type I Tax oncoprotein: cellular signaling through NF-kappa B. Cytokine Growth Factor Rev. 2001;12:207-217.
Jeang KT, Chiu R, Santos E, Kim SJ. Induction of the HTLV-I LTR by Jun occurs through the Tax-responsive 21- bp elements 5255. Virology. 1991;181:218-227.
Jeang KT, Giam CZ, Majone F, Aboud M. Life, death, and tax: role of HTLV-I oncoprotein in genetic instability and cellular transformation. J Biol Chem. 2004;279:31991-31994.
Jiang H, Lu H, Schiltz RL et al. PCAF interacts with tax and stimulates tax transactivation in a histone acetyltransferase-independent manner [published erratum appears in Mol Cell Biol 2000 Mar;20(5):1897]. Mol Cell Biol. 1999;19:8136-8145.
Jin DY, Spencer F, Jeang KT. Human T cell leukemia virus type 1 oncoprotein Tax targets the human mitotic checkpoint protein MAD1. Cell. 1998;93:81-91.
Johnson JM, Franchini G. Retroviral proteins that target the major histocompatibility complex class I. Virus Res. 2002;88:119-127.
Johnson JM, Fullen J, Casareto L, Mulloy JC. p12(I) May Contribute TO HTLV-1 Escape from Immune Detection 5313. Leukemia. 2000;14:558.
Johnson JM, Mulloy JC, Ciminale V et al. The MHC class I heavy chain is a common target of the small proteins encoded by the 3' end of HTLV type 1 and HTLV type 2. AIDS Res Hum Retroviruses. 2000;16:1777-1781.
Johnson JM, Nicot C, Fullen J et al. Free Major Histocompatibility Complex Class I Heavy Chain Is Preferentially Targeted for Degradation by Human T-Cell Leukemia/Lymphotropic Virus Type 1 p12(I) Protein. J Virol. 2001;75:6086-6094.
Johnson S, Michalak M, Opas M, Eggleton P. The ins and outs of calreticulin: from the ER lumen to the extracellular space. Trends Cell Biol. 2001;11:122-129.
Kamb A, Gruis NA, Weaver-Feldhaus J et al. A cell cycle regulator potentially involved in genesis of many tumor types. Science. 1994;264:436-440.
Kamihira S, Sohda H, Atogami S et al. Phenotypic diversity and prognosis of adult T-cell leukemia. Leuk Res. 1992;16:435-441.
Kasahata N, Shiota J, Miyazawa Y, Nakano I, Murayama S. Acute human T-lymphotropic virus type 1-associated myelopathy: a clinicopathologic study. Arch Neurol. 2003;60:873-876.

205
Kashanchi F, Brady JN. Transcriptional and post-transcriptional gene regulation of HTLV-1. Oncogene. 2005;24:5938-5951.
Kashishian A, Howard M, Loh C et al. AKAP79 inhibits calcineurin through a site distinct from the immunophilin-binding region. J Biol Chem. 1998;273:27412-27419.
Katagiri K, Hattori M, Minato N et al. Rap1 is a potent activation signal for leukocyte function-associated antigen 1 distinct from protein kinase C and phosphatidylinositol-3-OH kinase. Mol Cell Biol. 2000;20:1956-1969.
Kawakami K, Miyazato A, Iwakura Y, Saito A. Induction of lymphocytic inflammatory changes in lung interstitium by human T lymphotropic virus type I. AMER J RESPIR CRIT CARE MED. 1999;160:995-1000.
Kawano F, Yamaguchi K, Nishimura H, Tsuda H, Takatsuki K. Variation in the clinical courses of adult T-cell leukemia. Cancer. 1985;55:851-856.
Kehn K, Berro R, de La FC et al. Mechanisms of HTLV-1 transformation. Front Biosci. 2004;9:2347-2372.
Khabbaz RF, Douglas JM, Judson FN et al. Seroprevalence of human T-lymphotropic virus type I or II in sexually transmitted disease clinic patients in the USA. J Infect Dis. 1990;162:241-244.
Khabbaz RF, Onorato IM, Cannon RO et al. Seroprevalence of HTLV-1 and HTLV-2 among intravenous drug users and persons in clinics for sexually transmitted diseases. N Engl J Med. 1992;326:375-380.
Kim JH, Kaufman PA, Hanly SM, Rimsky LT, Greene WC. Rex transregulation of human T-cell leukemia virus type II gene expression. J Virol. 1991;65:405-414.
Kim M, Carman CV, Springer TA. Bidirectional transmembrane signaling by cytoplasmic domain separation in integrins. Science. 2003;301:1720-1725.
Kim SJ, Ding W, Albrecht B, Green PL, Lairmore MD. A conserved calcineurin-binding motif in human T lymphotropic virus type 1 p12I functions to modulate nuclear factor of activated T cell activation. J Biol Chem. 2003;278:15550-15557.
Kim SJ, Nair AM, Fernandez S, Mathes L, Lairmore MD. Enhancement of LFA-1-mediated T cell adhesion by human T lymphotropic virus type 1 p12I1. J Immunol. 2006;176:5463-5470.
Kimata JT, Palker TJ, Ratner L. The Mitogenic Activity of Human T-Cell Leukemia Virus Type- I Is T-Cell Associated and Requires the CD2/LFA-3 Activation Pathway. J Virol. 1993;67:3134-3141.

206
Kimata JT, Wong F, Wang J, Ratner L. Construction and characterization of infectious human T-cell leukemia virus type 1 molecular clones. Virology. 1994;204:656-664.
Kimura I, Tsubota T, Tada S, Sogawa J. Presence of antibodies against adult T cell leukemia antigen in the patients with chronic respiratory diseases 5241. Acta Med Okayama. 1986;40:281-284.
Kimzey AL, Dynan WS. Specific regions of contact between human T-cell leukemia virus type I Tax protein and DNA identified by photocross- linking. J Biol Chem. 1998;273:13768-13775.
King JA, Bridger JM, Lochelt M et al. Nucleocytoplasmic transport of HTLV-1 RNA is regulated by two independent LTR encoded nuclear retention elements. Oncogene. 1998;16:3309-3316.
Kinoshita S, Chen BK, Kaneshima H, Nolan GP. Host control of HIV-1 parasitism in T cells by the nuclear factor of activated T cells. Cell. 1998;95:595-604.
Kitado H, Chen IS, Shah NP et al. U3 sequences from HTLV-I and -II LTRs confer pX protein response to a murine leukemia virus LTR 5250. Science. 1987;235:901-904.
Kitze B, Usuku K. HTLV-1-mediated immunopathological CNS disease. Curr Top Microbiol Immunol. 2002;265:197-211.
Kiwaki T, Umehara F, Arimura Y et al. The clinical and pathological features of peripheral neuropathy accompanied with HTLV-I associated myelopathy. J Neurol Sci. 2003;206:17-21.
Kiyokawa T, Seiki M, Iwashita S et al. p27x-III and p21x-III, proteins encoded by the pX sequence of human T-cell leukemia virus type 1. Proc Natl Acad Sci USA. 1985;82:8359-8363.
Koralnik I, Mulloy JC, Andresson T, Fullen J, Franchini G. Mapping of the intermolecular association of the human T-cell leukemia/lymphotropic virus type 1 p12I and the vacuolar H+ATPase 16 kDa subunit protein. J Gen Virol. 1995;76:1909-1916.
Koralnik IJ, Fullen J, Franchini G. The p12, p13 and p30 proteins encoded by human T-cell leukemia/lymphotropic virus type-1 open reading frames I and II are localized in three different cellular compartments. J Virol. 1993;67:2360-2366.
Koralnik IJ, Fullen J, Franchini G. The p12I, p13II, and p30II proteins encoded by human T-cell leukemia/lymphotropic virus type I open reading frames I and II are localized in three different cellular compartments. J Virol. 1993;67:2360-2366.

207
Koralnik IJ, Gessain A, Klotman ME et al. Protein isoforms encoded by the pX region of human T-cell leukemia/lymphotropic virus type 1. Proc Natl Acad Sci USA. 1992;89:8813-8817.
Koralnik IJ, Gessain A, Klotman ME et al. Protein isoforms encoded by the pX region of human T-cell leukemia/lymphotropic virus type I. Proc Natl Acad Sci U S A. 1992;89:8813-8817.
Koralnik IJ, Mulloy JC, Andresson T, Fullen J, Franchini G. Mapping of the intermolecular association of the human T cell leukaemia/lymphotropic virus type I p12(I) and the vacuolar H+- ATPase 16 kDa subunit protein. J Gen Virol. 1995;76:1909-1916.
Kotov A, Zhou J, Flicker P, Aiken C. Association of Nef with the human immunodeficiency virus type 1 core. J Virol. 1999;73:8824-8830.
Krause KH, Michalak M. Calreticulin. Cell. 1997;88:439-443.
Kubota S, Hatanaka M, Pomerantz RJ. Nucleo-cytoplasmic redistribution of the HTLV-I rex protein: Alterations by coexpression of the HTLV-I p21(x) protein. Virology. 1996;220:502-507.
Kucik DF, Dustin ML, Miller JM, Brown EJ. Adhesion-activating phorbol ester increases the mobility of leukocyte integrin LFA-1 in cultured lymphocytes. J Clin Invest. 1996;97:2139-2144.
Kuhne MR, Lin J, Yablonski D et al. Linker for activation of T cells, zeta-associated protein-70, and Src homology 2 domain-containing leukocyte protein-76 are required for TCR-induced microtubule-organizing center polarization. J Immunol. 2003;171:860-866.
Kulkarni AB, Huh CG, Becker D et al. Transforming growth factor beta 1 null mutation in mice causes excessive inflammatory response and early death. Proc Natl Acad Sci U S A. 1993;90:770-774.
Kuroda Y, Matsui M. Cerebrospinal Fluid Interferon-Gamma Is Increased in HTLV- I- Associated Myelopathy. J Neuroimmunol. 1993;42:223-226.
Kwok RP, Lundblad JR, Chrivia JC et al. Nuclear protein CBP is a coactivator for the transcription factor. Nature. 1994;370(6486):223-226.
Kwok RPS, Laurance ME, Lundblad JR et al. Control of cAMP-regulated enhancers by the viral transactivator Tax through CREB and the co-activator CBP. Nature. 1996;380:642-646.

208
Lagrenade L, Hanchard B, Fletcher V, Cranston B, Blattner W. Infective dermatitis of Jamaican children: a marker for HTLV-I infection 5233. Lancet. 1990;336:1345-1347.
Laherty CD, Perkins ND, Dixit VM. Human T-Cell Leukemia Virus Type-I Tax and Phorbol 12- Myristate 13-Acetate Induce Expression of the A20 Zinc Finger Protein by Distinct Mechanisms Involving Nuclear Factor-kappaB. J Biol Chem. 1993;268:5032-5039.
Lai MM, Burnett PE, Wolosker H, Blackshaw S, Snyder SH. Cain, a novel physiologic protein inhibitor of calcineurin. J Biol Chem. 1998;273:18325-18331.
Lairmore MD, Albrecht B, D'Souza C et al. In vitro and in vivo functional analysis of human T cell lymphotropic virus type 1 pX open reading frames I and II. AIDS Res Hum Retroviruses. 2000;16:1757-1764.
Lairmore MD, Lal R. Other Human Retrovirus Infections: HTLV-I and HTLV-II. In: Schocketman G, George JR, eds. AIDS Testing. Methodology and Management Issues. New York: Springer-Verlag; 1995:168-188.
Lantz RC, Lemus R, Lange RW, Karol MH. Rapid reduction of intracellular glutathione in human bronchial epithelial cells exposed to occupational levels of toluene diisocyanate. Toxicol Sci. 2001;60:348-355.
Larocca D, Chao LA, Seto MH, Brunck TK. Human T-cell leukemia virus minus strand transcription in infected T-cells. Biochem Biophys Res Commun. 1989;163:1006-1013.
Le B, I, Grange MP, Delamarre L et al. HTLV-1 structural proteins. Virus Res. 2001;78:5-16.
Le B, I, Prevost MC, Dokhelar MC, Rosenberg AR. The PPPY motif of human T-cell leukemia virus type 1 Gag protein is required early in the budding process. J Virol. 2002;76:10024-10029.
Lefebvre L, Ciminale V, Vanderplasschen A et al. Subcellular localization of the bovine leukemia virus R3 and G4 accessory proteins. J Virol. 2002;76:7843-7854.
Lefebvre L, Ciminale V, Vanderplasschen A et al. Subcellular localization of the bovine leukemia virus R3 and G4 accessory proteins. J Virol. 2002;76:7843-7854.
Lefebvre L, Vanderplasschen A, Ciminale V et al. Oncoviral Bovine Leukemia Virus G4 and Human T-Cell Leukemia Virus Type 1 p13(II) Accessory Proteins Interact with Farnesyl Pyrophosphate Synthetase. J Virol. 2002;76:1400-1414.

209
Lemasson I, Polakowski NJ, Laybourn PJ, Nyborg JK. Transcription factor binding and histone modifications on the integrated proviral promoter in human T-cell leukemia virus-I-infected T-cells. J Biol Chem. 2002;277:49459-49465.
Lenzmeier BA, Baird EE, Dervan PB, Nyborg JK. The Tax protein-DNA interaction is essential for HTLV-I transactivation in vitro. J Mol Biol. 1999;291:731-744.
Leonmonzon M, Illa I, Dalakas MC. Polymyositis in patients infected with human T-cell leukemia virus type I: The role of the virus in the cause of the disease. Ann Neurol. 1994;36:643-649.
Levin MC, Jacobson S. HTLV-I associated myelopathy/tropical spastic paraparesis (HAM/TSP): A chronic progressive neurologic disease associated with immunologically mediated damage to the central nervous system. J Neurovirology. 1997;3:126-140.
Levin MC, Lee SM, Kalume F et al. Autoimmunity due to molecular mimicry as a cause of neurological disease. Nat Med. 2002;8:509-513.
Levin MC, Lee SM, Morcos Y, Brady J, Stuart J. Cross-reactivity between immunodominant human T lymphotropic virus type I tax and neurons: implications for molecular mimicry. J Infect Dis. 2002;186:1514-1517.
Li CC, Ruscetti FW, Rice NR et al. Differential expression of Rel family members in human T-cell leukemia virus type I-infected cells: transcriptional activation of c-rel by Tax protein. J Virol. 1993;67:4205-4213.
Li XH, Murphy KM, Palka KT, Surabhi RM, Gaynor RB. The human T-cell leukemia virus type-1 Tax protein regulates the activity of the I kappa B kinase complex. J Biol Chem. 1999;274:34417-34424.
Lin X, Sikkink RA, Rusnak F, Barber DL. Inhibition of calcineurin phosphatase activity by a calcineurin B homologous protein. J Biol Chem. 1999;274:36125-36131.
Lindholm PF, Reid RL, Brady JN. Extracellular Tax Protein Stimulates Tumor Necrosis Factor-B and Immunoglobulin Kappa Light Chain Expression in Lymphoid Cells. J Virol. 1992;66:1294-1302.
Loh C, Shaw KT, Carew J et al. Calcineurin binds the transcription factor NFAT1 and reversibly regulates its activity. J Biol Chem. 1996;271:10884-10891.
Lundblad JR, Kwok RP, Laurance ME, Harter ML, Goodman RH. Adenoviral E1A-associated protein p300 as a functional homologue of the transcriptional co-activator CBP. Nature. 1995;374:85-88.

210
Lundblad JR, Kwok RP, Laurance ME et al. The human T-cell leukemia virus-1 transcriptional activator Tax enhances cAMP-responsive element-binding protein (CREB) binding activity through interactions with the DNA minor groove. J Biol Chem. 1998;273:19251-19259.
Maggirwar SB, Harhaj E, Sun SC. Activation of NF-kappa B/Rel by Tax involves degradation of I kappa B alpha and is blocked by a proteasome inhibitor. Oncogene. 1995;11:993-998.
Mahieux R, Gessain A. HTLV-1 and associated adult T-cell leukemia/lymphoma. Rev Clin Exp Hematol. 2003;7:336-361.
Mahieux R, Ibrahim F, Mauclere P et al. Molecular epidemiology of 58 new African human T-cell leukemia virus type 1 (HTLV-1) strains: Identification of a new and distinct HTLV-1 molecular subtype in central Africa and in pygmies. J Virol. 1997;71:1317-1333.
Manel N, Battini JL, Taylor N, Sitbon M. HTLV-1 tropism and envelope receptor. Oncogene. 2005;24:6016-6025.
Manel N, Kim FJ, Kinet S et al. The ubiquitous glucose transporter GLUT-1 is a receptor for HTLV. Cell. 2003;115:449-459.
Manel N, Kinet S, Battini JL et al. The HTLV receptor is an early T-cell activation marker whose expression requires de novo protein synthesis. Blood. 2003;101:1913-1918.
Manninen A, Saksela K. HIV-1 Nef interacts with inositol trisphosphate receptor to activate calcium signaling in T cells. J Exp Med. 2002;195:1023-1032.
Manns A, Hisada M, Lagrenade L. Human T-lymphotropic virus type I infection. Lancet. 1999;353:1951-1958.
Manns A, Wilks RJ, Murphy EL et al. A prospective study of transmission by transfusion of HTLV-I and risk factors associated with seroconversion. Int J Cancer. 1992;51:886-891.
Marriott SJ, Lindholm PF, Brown KM et al. A 36-kilodalton cellular transcription factor mediates an indirect interaction of human T-cell leukemia/lymphoma virus type 1 Tax1 with a responsive element in the viral long terminal repeat. Mol Cell Biol. 1990;10:4192-4201.
Martins ML, Soares BC, Ribas JG et al. Frequency of p12K and p12R Alleles of HTLV Type 1 in HAM/TSP Patients and in Asymptomatic HTLV Type 1 Carriers. AIDS Res Hum Retroviruses. 2002;18:899-902.

211
Maruyama M, Shibuya H, Harada H et al. Evidence for aberrant activation of the interleukin-2 autocrine loop by HTLV-1-encoded p40x and T3/Ti complex triggering 5267. Cell. 1987;48:343-350.
Masuda ES, Imamura R, Amasaki Y, Arai K, Arai N. Signalling into the T-cell nucleus: NFAT regulation. Cell Signal. 1998;10:599-611.
Matsui M, Nagumo F, Tadano J, Kuroda Y. Characterization of humoral and cellular immunity in the central nervous system of HAM/TSP. J Neurol Sci. 1995;130:183-189.
Matsumoto J, Ohshima T, Isono O, Shimotohno K. HTLV-1 HBZ suppresses AP-1 activity by impairing both the DNA-binding ability and the stability of c-Jun protein. Oncogene. 2005;24:1001-1010.
Matsumoto M, Sugimoto M, Nakashima H et al. Spontaneous T cell proliferation and release of soluble interleukin-2 receptors in patients with HTLV-I-associated myelopathy. Am J Trop Med Hyg. 1990;42:365-373.
Merino F, Robert-Guroff M, Clark J et al. Natural antibodies to human T-cell leukemia/lymphoma virus in healthy Venezuelan populations. Int J Cancer. 1984;34:501.
Merl S, Kloster B, Moore J et al. Efficient Transformation of Previously Activated and Dividing T Lymphocytes by Human T Cell Leukemia-Lymphoma Virus. Blood. 1984;64:967-974.
Mesnard JM, Devaux C. Multiple control levels of cell proliferation by human T- cell leukemia virus type 1 tax protein. Virology. 1999;257:277-284.
Michael B, Nair A, Lairmore MD. Role of accessory proteins of HTLV-1 in viral replication, T cell activation, and cellular gene expression. Front Biosci. 2004;9:2556-2576.
Michael B, Nair AM, Hiraragi H et al. Human T lymphotropic virus type-1 p30II alters cellular gene expression to selectively enhance signaling pathways that activate T lymphocytes. Retrovirology. 2004;1:39.
Michalak M, Corbett EF, Mesaeli N, Nakamura K, Opas M. Calreticulin: one protein, one gene, many functions. Biochem J. 1999;344 Pt 2:281-292.
Miskin JE, Abrams CC, Dixon LK. African swine fever virus protein A238L interacts with the cellular phosphatase calcineurin via a binding domain similar to that of NFAT. J Virol. 2000;74:9412-9420.
Miskin JE, Abrams CC, Goatley LC, Dixon LK. A viral mechanism for inhibition of the cellular phosphatase calcineurin. Science. 1998;281:562-565.

212
Miyatake S, Seiki M, Malefijt RD et al. Activation of T cell-derived lymphokine genes in T cells and fibroblasts: effects of human T cell leukemia virus type I p40x protein and bovine papilloma virus encoded E2 protein 5266. Nucleic Acids Res. 1988;16:6547-6566.
Miyoshi I, Kubonishi I, Yoshimoto S et al. Type C virus particles in a cord T-cell line derived by co-cultivating normal human cord leukocytes and human leukaemic T cells. Nature. 1981;294:770-771.
Mochizuki M, Watanabe T, Yamaguchi K et al. HTLV-I Uveitis: A distinct clinical entity caused by HTLV-I. Jpn J Cancer Res. 1992;83:236-239.
Molgaard CA, Eisenman PA, Ryden LA, Golbeck AL. Neuroepidemiology of human T-lymphotrophic virus type-I-associated tropical spastic paraparesis. Neuroepidemiology. 1989;8:109-123.
Mori N, Fujii M, Ikeda S et al. Constitutive activation of NF-kappa B in primary adult T- cell leukemia cells. Blood. 1999;93:2360-2368.
Mori N, Mukaida N, Ballard DW, Matsushima K, Yamamoto N. Human T-cell leukemia virus type I Tax transactivates human interleukin 8 gene through acting concurrently on AP- 1 and nuclear factor-kappa B-like sites. Cancer Res. 1998;58:3993-4000.
Mori N, Murakami S, Oda S, Eto S. Human T-cell leukemia virus type I tax induces intracellular adhesion molecule-1 expression in T cells. Blood. 1994;84:350-351.
Mori N, Prager D. Transactivation of the interleukin-1 alpha promoter by human T- cell leukemia virus type I and type II tax proteins. Blood. 1996;87:3410-3417.
Mortreux F, Kazanji M, Gabet AS, de Thoisy B, Wattel E. Two-step nature of human T-cell leukemia virus type 1 replication in experimentally infected squirrel monkeys (Saimiri sciureus). J Virol. 2001;75:1083-1089.
Mosley AJ, Asquith B, Bangham CR. Cell-mediated immune response to human T-lymphotropic virus type I. Viral Immunol. 2005;18:293-305.
Muchardt C, Seeler JS, Nirula A, Gong S, Gaynor R. Transcription factor AP-2 activates gene expression of HTLV-I. EMBO Journal. 1992;11:2573-2581.
Mueller N.E., Blattner W.A. Retroviruses-Human T-Cell Lymphotropic Virus. Fourth Edition, 785-814. 1997. New York, N.Y., Plenum Publishing Corp. Ref Type: Generic
Mulloy JC, Crowley RW, Fullen J, Leonard WJ, Franchini G. The human T-cell leukemia/lymphotropic virus type 1 p12(I) protein binds the interleukin-2 receptor beta

213
and gamma(c) chains and affects their expression on the cell surface. J Virol. 1996;70:3599-3605.
Murakami T, Hirai H, Suzuki T, Fujisawa JI, Yoshida M. HTLV-1 tax enhances NF-kappa B2 expression and binds to the products p52 and p100, but does not suppress the inhibitory function of p100. Virology. 1995;206:1066-1074.
Muraoka O, Kaisho T, Tanabe M, Hirano T. Transcriptional activation of the interleukin-6 gene by HTLV-1. Immunology Letters. 1993;37(2-3):159-165.
Murphy EL, Busch MP, Tong M, Cornett P, Vyas GN. A prospective study of the risk of transfusion-acquired viral infections. Transfus Med. 1998;8:173-178.
Murphy EL, Glynn SA, Fridey J et al. Increased incidence of infectious diseases during prospective follow-up of human T-lymphotropic virus type II- and I-infected blood donors. ARCH INTERN MED. 1999;159:1485-1491.
Nagai M, Osame M. Human T-cell lymphotropic virus type I and neurological diseases. J Neurovirol. 2003;9:228-235.
Nagai M, Usuku K, Matsumoto W et al. Analysis of HTLV-I proviral load in 202 HAM/TSP patients and 243 asymptomatic HTLV-I carriers: high proviral load strongly predisposes to HAM/TSP. J Neurovirol. 1998;4:586-593.
Nagy K, Clapham P, Cheingsong-Popov R, Weiss R. Human T cell leukemia virus type 1: Induction of syncytia and inhibition by patient's sera. Int J Cancer. 1983;32:321-328.
Nair A, Michael B, Hiraragi H et al. Human T lymphotropic virus type 1 accessory protein p12I modulates calcium-mediated cellular gene expression and enhances p300 expression in T lymphocytes. AIDS Res Hum Retroviruses. 2005;21:273-284.
Nam SH, Kidokoro M, Shida H, Hatanaka M. Processing of gag Precursor Polyprotein of Human T-Cell Leukemia Virus Type 1 by Virus-Encoded Protease. J Virol. 1988;62:3718-3728.
Nath MD, Ruscetti FW, Petrow-Sadowski C, Jones KS. Regulation of the cell-surface expression of an HTLV-I binding protein in human T cells during immune activation. Blood. 2003;101:3085-3092.
Nejmeddine M, Barnard AL, Tanaka Y, Taylor GP, Bangham CR. HTLV-1 tax protein triggers microtubule reorientation in the virological synapse. J Biol Chem. 2005.
Nejmeddine M, Barnard AL, Tanaka Y, Taylor GP, Bangham CR. Human T-lymphotropic Virus, Type 1, Tax Protein Triggers Microtubule Reorientation in the Virological Synapse. J Biol Chem. 2005;280:29653-29660.

214
Ng PW, Iha H, Iwanaga Y et al. Genome-wide expression changes induced by HTLV-1 Tax: evidence for MLK- 3 mixed lineage kinase involvement in Tax-mediated NF-kappaB activation. Oncogene. 2001;20:4484-4496.
Nicot C. Current views in HTLV-I-associated adult T-cell leukemia/lymphoma. Am J Hematol. 2005;78:232-239.
Nicot C, Dundr M, Johnson JM et al. HTLV-1-encoded p30II is a post-transcriptional negative regulator of viral replication. Nat Med. 2004;10:197-201.
Nicot C, Harrod RL, Ciminale V, Franchini G. Human T-cell leukemia/lymphoma virus type 1 nonstructural genes and their functions. Oncogene. 2005;24:6026-6034.
Nicot C, Mahieux R, Takemoto S, Franchini G. Bcl-X(L) is up-regulated by HTLV-I and HTLV-II in vitro and in ex vivo ATLL samples. Blood. 2000;96:275-281.
Nicot C, Mulloy JC, Ferrari MG et al. HTLV-1 p12(I) protein enhances STAT5 activation and decreases the interleukin-2 requirement for proliferation of primary human peripheral blood mononuclear cells. Blood. 2001;98:823-829.
Nishioka K, Maruyama I, Sato K et al. Chronic inflammatory arthropathy associated with HTLV-I [letter]. Lancet. 1989;i:441.
Nitta T, Tanaka M, Sun B, Hanai S, Miwa M. The genetic background as a determinant of human T-cell leukemia virus type 1 proviral load. Biochem Biophys Res Commun. 2003;309:161-165.
Niyogi K, Hildreth JE. Characterization of new syncytium-inhibiting monoclonal antibodies implicates lipid rafts in human t-cell leukemia virus type 1 syncytium formation. J Virol. 2001;75:7351-7361.
Northrop JP, Ullman KS, Crabtree GR. Characterization of the nuclear and cytoplasmic components of the lymphoid-specific nuclear factor of activated T cells (NF-AT) complex. J Biol Chem. 1993;268:2917-2923.
Nosaka T, Siomi H, Adachi Y et al. Nucleolar targeting signal of human T-cell leukemia virus type I rex- encoded protein is essential for cytoplasmic accumulation of unspliced viral mRNA 5295. Proc Natl Acad Sci U S A. 1989;86:9798-9802.
Ohshima K, Kikuchi M, Masuda Y et al. Human T-cell leukemia virus type I associated lymphadenitis 5240. Cancer. 1992;69:239-248.

215
Okochi K, Sato H, Hinuma Y. A retrospective study on transmission of adult T cell leukemia virus by blood transfusion: seroconversion in recipients. Vox Sang. 1984;46:245-253.
Ootsuyama Y, Shimotohno K, Miwa M, Oroszlan S, Sugimura T. Myristylation of gag protein in human T-cell leukemia virus type-I and type-II. Jpn J Cancer Res. 1985;76:1132-1135.
Orita S, Saiga A, Takagi S et al. A novel alternatively spliced viral mRNA transcribed in cells infected with human T cell leukemia virus type 1 is mainly responsible for expressing p21X protein 5300. FEBS Lett. 1991;295:127-134.
Oroszlan S. Chemical Analysis of HTLV-1 Structural proteins 5260. In: Gallo RC, Essex ME, Gross L, eds. Human T-cell Leukemia Lymphoma Viruses. Cold Spring Harbor Laboratory Press; 1984:101-110.
Osame M. Pathological mechanisms of human T-cell lymphotropic virus type I-associated myelopathy (HAM/TSP). J Neurovirol. 2002;8:359-364.
Osame M, Igata A, Usuku K, Rosales RL, Matsumoto M. Mother-to-child transmission in HTLV-I associated myelopathy. Lancet. 1987;1:106.
Osame M, Izumo S, Igata A et al. Blood transfusion and HTLV-I-associated myelopathy. Lancet. 1986;104.
Osame M, Janssen R, Kubota H et al. Nationwide survey of HTLV-I-associated Myelopathy in Japan: Association with blood transfusion. Ann Neurol. 1990;28:50-56.
Osame M, Usuku K, Izumo S et al. HTLV-I associated myelopathy, a new clinical entity. Lancet. 1986;i:1031-1032.
Overbaugh J. HTLV-1 sweet-talks its way into cells. Nat Med. 2004;10:20-21.
Owen SM, Rudolph DL, Dezzutti CS et al. Transcriptional activation of the intercellular adhesion molecule 1 (CD54) gene by human T lymphotropic virus types I and II Tax is mediated through a palindromic response element. AIDS Res Hum Retroviruses. 1997;13:1429-1437.
Palmeri D, Malim MH. The human T-cell leukemia virus type 1 posttranscriptional trans- activator Rex contains a nuclear export signal. J Virol. 1996;70:6442-6445.
Park S, Uesugi M, Verdine GL. A second calcineurin binding site on the NFAT regulatory domain. Proc Natl Acad Sci U S A. 2000;97:7130-7135.
Pawson T. Protein modules and signalling networks 5308. Nature. 1995;373:573-580.

216
Perez OD, Mitchell D, Jager GC et al. Leukocyte functional antigen 1 lowers T cell activation thresholds and signaling through cytohesin-1 and Jun-activating binding protein 1. Nat Immunol. 2003;4:1083-1092.
Perini G, Wagner S, Green MR. Recognition of bZIP proteins by the human T-cell leukaemia virus transactivator Tax. Nature. 1995;376:602-605.
Perkins ND, Felzien LK, Betts JC et al. Regulation of NF-kappaB by cyclin-dependent kinases associated with the p300 coactivator. Science. 1997;275:523-527.
Petit C, Buseyne F, Boccaccio C et al. Nef is required for efficient hiv-1 replication in cocultures of dendritic cells and lymphocytes. Virology. 2001;286:225-236.
Petrini M, Pelosi-Testa E, Sposi NM et al. Constitutive expression and abnormal glycosylation of transferrin receptor in acute T-cell leukemia. Cancer Res. 1989;49:6989-6996.
Pinon JD, Klasse PJ, Jassal SR et al. Human T-cell leukemia virus type 1 envelope glycoprotein gp46 interacts with cell surface heparan sulfate proteoglycans. J Virol. 2003;77:9922-9930.
Pique C, Pham D, Tursz T, Dokhelar MC. Human T-Cell Leukemia Virus Type I Envelope Protein Maturation Process: Requirements for Syncytium Formation. J Virol. 1992;66:906-913.
Pique C, Uretavidal A, Gessain A et al. Evidence for the chronic in vivo production of human T cell leukemia virus type I Rof and Tof proteins from cytotoxic T lymphocytes directed against viral peptides. J Exp Med. 2000;191:567-572.
Pise-Masison CA, Jeong SJ, Brady JN. Human T cell leukemia virus type 1: the role of Tax in leukemogenesis. Arch Immunol Ther Exp (Warsz ). 2005;53:283-296.
Poiesz BJ, Ruscetti FW, Gazdar AF et al. Detection and isolation of type C retrovirus particles from fresh and cultured lymphocytes of a patient with cutaneous T-cell lymphoma. Proc Natl Acad Sci USA. 1980;77:7415-7419.
Popovic M, Sarin PS, Robert-Gurroff M et al. Isolation and transmission of human retrovirus (human t-cell leukemia virus). Science. 1983;219:856-859.
Portis T, Harding JC, Ratner L. The contribution of NF-kappa B activity to spontaneous proliferation and resistance to apoptosis in human T-cell leukemia virus type 1 Tax- induced tumors. Blood. 2001;98:1200-1208.
Princler GL, Julias JG, Hughes SH, Derse D. Roles of viral and cellular proteins in the expression of alternatively spliced HTLV-1 pX mRNAs. Virology. 2003;317:136-145.

217
Proietti FA, Carneiro-Proietti AB, Catalan-Soares BC, Murphy EL. Global epidemiology of HTLV-I infection and associated diseases. Oncogene. 2005;24:6058-6068.
Rao A, Luo C, Hogan PG. Transcription factors of the NFAT family: regulation and function. Annu Rev Immunol. 1997;15:707-747.
Ratner L. Regulation of expression of the c-sis proto-oncogene 5269. Nucleic Acids Res. 1989;17:4101-4115.
Reeves W, Saxinger C, Brenes M et al. Human T-cell lymphotropic virus type I (HTLV-I) and risk factors in metropolitan Panama. Am J Epidemiol. 1988;127:539.
Ressler S, Morris GF, Marriott SJ. Human T-cell leukemia virus type 1 Tax transactivates the human proliferating cell nuclear antigen promoter. J Virol. 1997;71:1181-1190.
Robek MD, Wong FH, Ratner L. Human T-Cell leukemia virus type 1 pX-I and pX-II open reading frames are dispensable for the immortalization of primary lymphocytes. J Virol. 1998;72:4458-4462.
Rodrigues MS, Echevarria-Lima J, Fernandes dos SN, Rumjanek VM. CD69 expression induced by thapsigargin, phorbol ester and ouabain on thymocytes is dependent on external Ca2+ entry. Life Sci. 2003;73:1037-1051.
Rodriguez-Fernandez JL, Gomez M, Luque A et al. The interaction of activated integrin lymphocyte function-associated antigen 1 with ligand intercellular adhesion molecule 1 induces activation and redistribution of focal adhesion kinase and proline-rich tyrosine kinase 2 in T lymphocytes. Mol Biol Cell. 1999;10:1891-1907.
Roithmann S, Pique C, Le Cesne A et al. The open reading frame I (ORF I)/ORF II part of the human T-cell leukemia virus type 1 X region is dispensable for p40tax, p27rex, or envelope expression. J Virol. 1994;68:3448-3451.
Rothermel B, Vega RB, Yang J et al. A protein encoded within the Down syndrome critical region is enriched in striated muscles and inhibits calcineurin signaling. J Biol Chem. 2000;275:8719-8725.
Rusnak F, Mertz P. Calcineurin: form and function. Physiol Rev. 2000;80:1483-1521.
Sagara Y, Inoue Y, Kojima E et al. Phosphatidylglycerol Participates in Syncytium Formation Induced by HTLV Type 1-Bearing Cells. AIDS Res Hum Retroviruses. 2001;17:125-135.

218
Sagara Y, Ishida C, Inoue Y, Shiraki H, Maeda Y. 71-Kilodalton heat shock cognate protein acts as a cellular receptor for syncytium formation induced by human T-cell lymphotropic virus type 1. J.Virol. 72, 535-541. 1998. Ref Type: Generic
Saito M, Furukawa Y, Kubota R et al. Frequent mutation in pX region of HTLV-1 is observed in HAM/TSP patients, but is not specifically associated with the central nervous system lesions. J Neurovirol. 1995;1:286-294.
Saksena NK, Srinivasan A, Ge YC et al. Simian T cell leukemia virus type I from naturally infected feral monkeys from Central and West Africa encodes a 91-amino acid p12 (ORF-I) protein as opposed to a 99-amino acid protein encoded by HTLV type I from humans. AIDS Res Hum Retroviruses. 1997;13:425-432.
Sancho D, Gomez M, Sanchez-Madrid F. CD69 is an immunoregulatory molecule induced following activation. Trends Immunol. 2005;26:136-140.
Sancho D, Gomez M, Viedma F et al. CD69 downregulates autoimmune reactivity through active transforming growth factor-beta production in collagen-induced arthritis. J Clin Invest. 2003;112:872-882.
Saxinger WC, Blattner W, Levine P et al. Human T-cell leukemia virus (HTLV-I) antibodies in Africa. Science. 1984;225:1473.
Schreiber GB, Murphy EL, Horton JA et al. Risk factors for human T-cell lymphotropic virus types I and II (HTLV-I and -II) in blood donors: the Retrovirus Epidemiology Donor Study. NHLBI Retrovirus Epidemiology Donor Study. J Acquir Immune Defic Syndr Hum Retrovirol. 1997;14:263-271.
Seiki M, Hattori S, Hirayama Y, Yoshida M. Human adult T-cell leukemia virus: Complete nucleotide sequence of the provirus genome integrated in leukemia cell DNA. Proc Natl Acad Sci. 1983;80:3618-3622.
Seiki M, Hikikoshi A, Taniguchi T, Yoshida M. Expression of the pX Gene of HTLV-1: General Splicing Mechanism in the HTLV Family. Science. 1985;228:1532-1534.
Seiki M, Hikikoshi A, Yoshida M. The U5 sequence is a cis-acting repressive element for genomic RNA expression of human T cell leukemia virus type I. Virology. 1990;176:81-86.
Seiki M, Inoue J, Hidaka M, Yoshida M. Two cis-acting elements responsible for posttranscriptional trans-regulation of gene expression of human T-cell leukemia virus type I. Proc Natl Acad Sci U S A. 1988;85:7124-7128.

219
Semmes OJ, Jeang KT. Localization of human T-cell leukemia virus type 1 Tax to subnuclear compartments that overlap with interchromatin speckles. J Virol. 1996;70:6347-6357.
Shaw GM, Gonda MA, Flickinger GH et al. Genomes of evolutionarily divergent members of the human T-cell leukemia virus family (HTLV-I and HTLV-II) are highly conserved, especially in pX. Proc Natl Acad Sci USA. 1984;81:4544-4548.
Shibasaki F, Kondo E, Akagi T, Mckeon F. Suppression of signalling through transcription factor NF-AT by interactions between calcineurin and Bcl-2. Nature. 1997;386:728-731.
Shimotohno K, Takano M, Teruuchi T, Miwa M. Requirement of multiple copies of a 21-nucleotide sequence in the U3 regions of human T-cell leukemia virus type I and type II long terminal repeats for trans-acting activation of transcription 5252. Proc Natl Acad Sci U S A. 1986;83:8112-8116.
Shimoyama M. Diagnostic criteria and classification of clinical subtypes of adult T- cell leukaemia-lymphoma. A report from the Lymphoma Study Group (1984- 87) 5222. Br J Haematol. 1991;79:428-437.
Silic-Benussi M, Cavallari I, Zorzan T et al. Suppression of tumor growth and cell proliferation by p13II, a mitochondrial protein of human T cell leukemia virus type 1. Proc Natl Acad Sci U S A. 2004;101:6629-6634.
Silverman LR, Phipps AJ, Montgomery A, Ratner L, Lairmore MD. Human T-cell lymphotropic virus type 1 open reading frame II-encoded p30II is required for in vivo replication: evidence of in vivo reversion. J Virol. 2004;78:3837-3845.
Sims TN, Dustin ML. The immunological synapse: integrins take the stage. Immunol Rev. 2002;186:100-117.
Siomi H, Shida H, Nam SH et al. Sequence requirements for nucleolar localization of human T cell leukemia virus type I pX protein, which regulates viral RNA processing 5296. Cell. 1988;55:197-209.
Smith A, Bracke M, Leitinger B, Porter JC, Hogg N. LFA-1-induced T cell migration on ICAM-1 involves regulation of MLCK-mediated attachment and ROCK-dependent detachment. J Cell Sci. 2003;116:3123-3133.
Sodroski J, Rosen C, Goh WC, Haseltine W. A transcriptional activator protein encoded by the x-lor region of the human T-cell leukemia virus 5017. Science. 1985;228:1430-1434.

220
Sodroski JG, Rosen CA, Haseltine WA. Trans-Acting Transcriptional Activation of the Long Terminal Repeat of Human T Lymphotropic Viruses in Infected Cells. Science. 1984;223:381-385.
Srivastava RK, Sasaki CY, Hardwick JM, Longo DL. Bcl-2-mediated drug resistance: inhibition of apoptosis by blocking nuclear factor of activated T lymphocytes (NFAT)-induced Fas ligand transcription. J Exp Med. 1999;190:253-265.
Srivastava RK, Sasaki CY, Hardwick JM, Longo DL. Bcl-2-mediated drug resistance: inhibition of apoptosis by blocking nuclear factor of activated T lymphocytes (NFAT)-induced Fas ligand transcription. J Exp Med. 1999;190:253-265.
Srivastava RK, Sasaki CY, Hardwick JM, Longo DL. Bcl-2-mediated drug resistance: inhibition of apoptosis by blocking nuclear factor of activated T lymphocytes (NFAT)-induced Fas ligand transcription. J Exp Med. 1999;190:253-265.
Stewart M, Hogg N. Regulation of leukocyte integrin function: affinity vs. avidity. J Cell Biochem. 1996;61:554-561.
Stewart MP, Cabanas C, Hogg N. T cell adhesion to intercellular adhesion molecule-1 (ICAM-1) is controlled by cell spreading and the activation of integrin LFA-1. J Immunol. 1996;156:1810-1817.
Stewart MP, Mcdowall A, Hogg N. LFA-1-mediated adhesion is regulated by cytoskeletal restraint and by a Ca2+-dependent protease, calpain. J Cell Biol. 1998;140:699-707.
Stuver SO, Tachibana N, Okayama A et al. Heterosexual transmission of human T cell leukemia/lymphoma virus type I among married couples in southwestern Japan: an initial report from the Miyazaki Cohort Study. J Infect Dis. 1993;167:57-65.
Sun SC, Ballard DW. Persistent activation of NF-kappa B by the Tax transforming protein of HTLV-1: hijacking cellular I kappa B kinases. Oncogene. 1999;18:6948-6958.
Suzuki T, Fujisawa J, Toita M, Yoshida M. The trans-activator Tax of human T-cell leukemia virus type 1 (HTLV-1) interacts with cAMP-responsive element (CRE) binding and CRE modulator proteins that bind to the 21-base-pair enhancer of HTLV-1. Proc Natl Acad Sci U S A. 1993;90:610-614.
Suzuki T, Hirai H, Fujisawa J, Fujita T, Yoshida M. A trans-activator Tax of human T-cell leukemia virus type 1 binds to NF- kappa B p50 and serum response factor (SRF) and associates with enhancer DNAs of the NF-kappa B site and CArG box 5014. Oncogene. 1993;8:2391-2397.

221
Suzuki T, Hirai H, Murakami T, Yoshida M. Tax protein of HTLV-1 destabilizes the complexes of NF- kappa B and I kappa B-alpha and induces nuclear translocation of NF-kappa B for transcriptional activation. Oncogene. 1995;10:1199-1207.
Suzuki T, Hirai H, Yoshida M. Tax protein of HTLV-1 interacts with the Rel homology domain of NF-kappa B p65 and c-Rel proteins bound to the NF-kappa B binding site and activates transcription. Oncogene. 1994;9:3099-3105.
Suzuki T, Kitao S, Matsushime H, Yoshida M. HTLV-1 Tax protein interacts with cyclin-dependent kinase inhibitor p16(INK4A) and counteracts its inhibitory activity towards CDK4. EMBO J. 1996;15:1607-1614.
Suzuki T, Narita T, Uchidatoita M, Yoshida M. Down-regulation of the INK4 family of cyclin-dependent kinase inhibitors by tax protein of HTLV-1 through two distinct mechanisms. Virology. 1999;259:384-391.
Suzuki T, Uchidatoita M, Yoshida M. Tax protein of HTLV-1 inhibits CBP/p300-mediated transcription by interfering with recruitment of CBP/p300 onto DNA element of E-box or p53 binding site. Oncogene. 1999;18:4137-4143.
Takatsuki K, Uchiyama T, Sagawa K, Yodoi J. Adult T-cell leukemia in Japan. In: Seno S, Takaku F, Irino S, eds. Topics in Hematology. Amsterdam: Excerpta Medica; 1977:73-77.
Tanimura A, Teshima H, Fujisawa JI, Yoshida M. A New Regulatory Element that Augments the Tax-Dependent Enhancer of Human T-Cell Leukemia Virus Type 1 and Cloning of cDNAs Encoding Its Binding Proteins. J Virol. 1993;67:5375-5382.
Tendler CL, Greenberg SJ, Blattner WA et al. Transactivation of interleukin 2 and its receptor induces immune activation in human T-cell lymphotropic virus type I-associated myelopathy: pathogenic implications and a rationale for immunotherapy. Proceedings of the National Academy of Sciences of the United States of America. 1990;87:5218-5222.
Terada K, Katamine S, Eguchi K et al. Prevalence of serum and salivary antibodies to HTLV-1 in Sjogren's syndrome. Lancet. 1994;344:1116-1119.
Terashima A, Gotuzzo E, Alvarez H et al. [STRONGYLOIDES STERCORALIS: CLINICAL SEVERE FORMS ASSOCIATED TO HTLV-1 INFECTION]. Rev Gastroenterol Peru. 1999;19:35-40.
Teshigawara K, Maeda M, Nishino K et al. Adult T leukemia cells produce a lymphokine that augments interleukin 2 receptor expression. J Mol Cell Immunol. 1985;2:17-26.

222
Thebault S, Basbous J, Hivin P, Devaux C, Mesnard JM. HBZ interacts with JunD and stimulates its transcriptional activity. FEBS Lett. 2004;562:165-170.
Tian P, Estes MK, Hu Y et al. The rotavirus nonstructural glycoprotein NSP4 mobilizes Ca2+ from the endoplasmic reticulum. J Virol. 1995;69:5763-5772.
Tillmann M, Wessner R, Wigdahl B. Identification of human T-cell lymphotropic virus type I 21-base- pair repeat-specific and glial cell-specific DNA- protein complexes. J Virol. 1994;68:4597-4608.
Toedter GP, Pearlman S, Hofheinz D et al. Development of a monoclonal antibody-based p24 capsid antigen detection assay for HTLV-I, HTLV-II, and STLV-I infection. AIDS Res Hum Retroviruses. 1992;8:527-532.
Torgeman A, Mor-Vaknin N, Aboud M. Sp1 is involved in a protein kinase C-independent activation of human T cell leukemia virus type I long terminal repeat by 12-O-tetradecanoylphorbol-13-acetate. Virology. 1999;254:279-287.
Trovato R, Mulloy JC, Johnson JM et al. A lysine-to-arginine change found in natural alleles of the human T-cell lymphotropic/leukemia virus type 1 p12(I) protein greatly influences its stability. J Virol. 1999;73:6460-6467.
Trovato R, Mulloy JC, Johnson JM et al. A lysine-to-arginine change found in natural alleles of the human T-cell lymphotropic leukemia virus type 1 p12 protein greatly influences its stability. J Virol. 1999;73:6460-6467.
Trovato R, Mulloy JC, Johnson JM et al. A lysine-to-arginine change found in natural alleles of the human T-cell lymphotropic leukemia virus type 1 p12 protein greatly influences its stability. J Virol. 1999;73:6460-6467.
Tsukahara T, Kannagi M, Ohashi T et al. Induction of Bcl-x(L) expression by human T-cell leukemia virus type 1 tax through NF-kappa B in apoptosis-resistant T-cell transfectants with tax. J Virol. 1999;73:7981-7987.
Tsukahara T, Ratner L. Substitution of HIV Type 1 Nef with HTLV-1 p12. AIDS Res Hum Retroviruses. 2004;20:938-943.
Uchiyama T. Human T cell leukemia virus type I (HTLV-I) and human diseases. [Review] [134 refs]. Annual Review of Immunology. 1997;15:15-37:15-37.
Uchiyama T, Yodoi J, Sagawa K, Takatsuki K, Uchino H. Adult T-cell leukemia: clinical and hematologic features of 16 cases 5209. Blood. 1977;50:481-492.

223
Usuku K, Sonoda S, Osame M et al. HLA haplotype-linked high immune responsiveness against HTLV-I in HTLV-I-associated myelopathy: comparison with adult T-cell leukemia/lymphoma. Ann Neurol. 1988;23 Suppl:S143-50.:S143-S150.
van KY, Figdor CG. Activation and inactivation of adhesion molecules. Curr Top Microbiol Immunol. 1993;184:235-256.
Volkov Y, Long A, Kelleher D. Inside the crawling T cell: leukocyte function-associated antigen-1 cross-linking is associated with microtubule-directed translocation of protein kinase C isoenzymes beta(I) and delta. J Immunol. 1998;161:6487-6495.
von Andrian UH, Mackay CR. T-cell function and migration. Two sides of the same coin. N Engl J Med. 2000;343:1020-1034.
von Bulow GU, Bram RJ. NF-AT activation induced by a CAML-interacting member of the tumor necrosis factor receptor superfamily. Science. 1997;278:138-141.
Wang H, Moon EY, Azouz A et al. SKAP-55 regulates integrin adhesion and formation of T cell-APC conjugates. Nat Immunol. 2003;4:366-374.
Wattel E, Vartanian JP, Pannetier C, Wainhobson S. Clonal expansion of human T-Cell leukemia virus type I- infected cells in asymptomatic and symptomatic carriers without malignancy. J Virol. 1995;69:2863-2868.
Weiss RA. Milk-borne transmission of HTLV-1. [Review]. Japanese Journal of Cancer Research. 1993;84(3):inside front cover.
Wessner R, Tillmann-Bogush M, Wigdahl B. Characterization of a glial cell-specific DNA-protein complex formed with the human T cell lymphotropic virus type I (HTLV-I) enhancer. J Neurovirol. 1995;1:62-77.
Wessner R, Wigdahl B. AP-1 derived from mature monocytes and astrocytes preferentially interacts with the HTLV-I promoter central 21 bp repeat. Leukemia. 1997;11 Suppl 3:21-4.:21-24.
Wiktor SZ, Pate EJ, Murphy EL et al. Mother-to-child transmission of human T-cell lymphotropic virus type I (HTLV-I) in Jamaica: association with antibodies to envelope glycoprotein (gp46) epitopes. J Acquir Immune Defic Syndr. 1993;6:1162-1167.
Wu K, Bottazzi ME, de La FC et al. Protein profile of tax-associated complexes. J Biol Chem. 2004;279:495-508.
Wu Y, Marsh JW. Selective transcription and modulation of resting T cell activity by preintegrated HIV DNA. Science. 2001;293:1503-1506.
Wucherpfennig KW, Hollsberg P, Richardson JH, Benjamin D, Hafler DA. T-cell activation by autologous human T-cell leukemia virus type I- infected T-cell clones

224
5375. Proc Natl Acad Sci U S A. 1992;89:2110-2114.
Xu RZ, Gao QK, Wang SJ et al. Human acute myeloid leukemias may be etiologically associated with new human retroviral infection. Leuk Res. 1996;20:449-455.
Yajima A, Kawada A, Aragane Y, Tezuka T. Detection of HTLV-I proviral DNA in sarcoidosis. Dermatology. 2001;203:53-56.
Yamamoto A, Hara H, Kobayashi T. Induction of the expression of gag protein in HTLV-I infected lymphocytes by anti-ICAM 1 antibody in vitro. J Neurol Sci. 1997;151:121-126.
Yamamoto N, Okada M, Koyanagi Y, Kannagi M, Hinuma Y. Transformation of human Leukocytes by cocultivation with an adult T cell leukemia virus producer cell line. Science. 1982;217:737-739.
Yamano Y, Nagai M, Brennan M et al. Correlation of human T-cell lymphotropic virus type 1 (HTLV-1) mRNA with proviral DNA load, virus-specific CD8(+) T cells, and disease severity in HTLV-1-associated myelopathy (HAM/TSP). Blood. 2002;99:88-94.
Yao J, Wigdahl B. Human T cell lymphotropic virus type I genomic expression and impact on intracellular signaling pathways during neurodegenerative disease and leukemia. Front Biosci. 2000;5:D138-D168.
Yoshida M. HTLV-1 Tax: regulation of gene expression and disease. [Review]. Trends in Microbiology. 1993;1(4):131-135.
Yoshida M. Multiple viral strategies of htlv-1 for dysregulation of cell growth control. Annu Rev Immunol. 2001;19:475-496.
Yoshida M, Miyoshi I, Hinuma Y. Isolation and characterization of retrovirus from cell lines of human T-cell leukemia and its implication in the disease. Proc Natl Acad Sci USA. 1982;79:2031-2035.
Yoshida M, Seiki M, Yamaguchi K, Takatsuki K. Monoclonal integration of human T-cell leukemia provirus in all primary tumors of adult T-cell leukemia suggests causative role of human T-cell leukemia virus in the disease. Proc Natl Acad Sci U S A. 1984;81:2534-2537.
Younis I, Boris-Lawrie K, Green PL. Human T-cell leukemia virus open reading frame II encodes a posttranscriptional repressor that is recruited at the level of transcription. J Virol. 2006;80:181-191.
Younis I, Khair L, Dundr M et al. Repression of Human T-Cell Leukemia Virus Type 1 and Type 2 Replication by a Viral mRNA-Encoded Posttranscriptional Regulator. J Virol. 2004;78:11077-11083.

225
Zhang W, Nisbet JW, Albrecht B et al. Human T-lymphotropic virus type 1 p30(II) regulates gene transcription by binding CREB binding protein/p300. J Virol. 2001;75:9885-9895.
Zhang W, Nisbet JW, Bartoe JT, Ding W, Lairmore MD. Human T-lymphotropic virus type 1 p30(II) functions as a transcription factor and differentially modulates CREB-responsive promoters. J Virol. 2000;74:11270-11277.
Zhao LJ, Giam CZ. Human T-cell lymphotropic virus type I (HTLV-I) transcriptional activator, Tax, enhances CREB binding to HTLV-I 21-base-pair repeats by protein-protein interaction. Proceedings of the National Academy of Sciences of the United States of America. 1992;89:7070-7074.
Zhong H, Voll RE, Ghosh S. Phosphorylation of NF-kappa B p65 by PKA stimulates transcriptional activity by promoting a novel bivalent interaction with the coactivator CBP/p300. Mol Cell. 1998;1:661-671.
Zhou J, Aiken C. Nef enhances human immunodeficiency virus type 1 infectivity resulting from intervirion fusion: evidence supporting a role for Nef at the virion envelope. J Virol. 2001;75:5851-5859.
Recommended

















