
Biochemical Pharmacology 86 (2013) 571–583
The antitumor lignan Nortrachelogenin sensitizes prostate cancer cellsto TRAIL-induced cell death by inhibition of the Akt pathway andgrowth factor signaling
Emilia Peuhu a, Preethy Paul a,b, Mika Remes a, Thomas Holmbom c, Patrik Eklund d,Rainer Sjoholm d, John E. Eriksson a,b,*a Turku Centre for Biotechnology, University of Turku and Abo Akademi University, Biocity, POB 123, FI-20521 Turku, Finlandb Department of Biosciences, Abo Akademi University, Tykistokatu 6, FI-20520 Turku, Finlandc Laboratory of Fibre and Cellulose Technology, Abo Akademi University, Porthaninkatu 3, FI-20500 Turku, Finlandd Laboratory of Organic Chemistry, Abo Akademi University, Piispankatu 8, FI-20500 Turku, Finland
A R T I C L E I N F O
Article history:
Received 4 February 2013
Accepted 16 May 2013
Available online 6 June 2013
Keywords:
Nortrachelogenin
Lignans
TRAIL
Prostate cancer
Receptor tyrosine kinase
Dibenzylbutyrolactone
A B S T R A C T
Prostate cancer cells frequently develop resistance toward androgen-deprivation and chemotherapy. To
identify new approaches to treat androgen-dependent prostate cancer, we have performed a structure–
activity analysis of lignan polyphenols for cancer cell specific sensitization to tumor necrosis factor-
related apoptosis-inducing ligand (TRAIL), a death ligand that has ability to induce tumor-specific cell
death. In this study, we report that the lignan nortrachelogenin (NTG) is the most efficient of the 27
tested lignan compounds in sensitizing prostate cancer cells to TRAIL-induced apoptosis. Importantly,
pretreatment with NTG does not sensitize a non-malignant prostate cell line to TRAIL-induced cell death.
The structural comparison of lignans reveals that the dibenzylbutyrolactone skeleton is required for the
apoptosis-sensitizing activity, while substitutions at the aromatic rings do not seem to play a critical role
in this lignan function. Our study also characterizes the cellular effects and molecular mechanisms
involved in NTG anticancer activity. We previously reported that specific lignans inhibit the Akt survival-
signaling pathway in concert with TRAIL sensitization. While NTG is also shown to be a effective inhibitor
of Akt signaling, in this study we further demonstrate that NTG potently inhibits tyrosine kinase (RTK)
activation in response to growth factors, such as insulin and insulin-like growth factor I (IGF-I). Our
results identify NTG as a novel agent for prostate cancer therapy with ability to inhibit Akt membrane
localization and activity as well as the activation of growth factor receptors (GFRs), thereby efficiently
synergizing with TRAIL exposure.
� 2013 Elsevier Inc. All rights reserved.
Contents lists available at SciVerse ScienceDirect
Biochemical Pharmacology
jo u rn al h om epag e: ww w.els evier .c o m/lo cat e/b io c hem p har m
1. Introduction
Prostrate cancer is the most common type of cancer affectingmen in the western world. Early prostate carcinomas that dependon androgen supply for their growth can metastasize as well asprogress into hormone-refractory prostate cancer (HRPC). Whileandrogen deprivation therapy (ADT) remains as the first-linetreatment of advanced prostate cancer, limited therapeuticapproaches are available and new drug targets are beinginvestigated. Deregulation of the phosphatidylinositol-3-kinase(PI3K)/Akt signaling pathway is common in cancer due tomutations in the key components of the pathway [1,2]. PI3Kactivity is negatively regulated by the tumor suppressor phospha-tase and tensin homolog (PTEN), the function of which is commonly
* Corresponding author.
E-mail address: [email protected] (J.E. Eriksson).
0006-2952/$ – see front matter � 2013 Elsevier Inc. All rights reserved.
http://dx.doi.org/10.1016/j.bcp.2013.05.026
lost in prostate cancer [3,4]. Activation of PI3K/Akt signaling bygrowth factor stimulation via receptor tyrosine kinases (RTKs) mayplay a critical role in allowing prostate tumors to maintaincontinued proliferation in low-androgen environments, therebypromoting development of HRPC [5]. Several recent studies indicatethat inhibition of the RTK/PI3K/Akt pathway with therapeuticagents is an attractive strategy to treat prostate cancers [6].
Lignans are a group of plant-derived polyphenolic compoundsthat have the capability to inhibit Akt signaling in prostatecancer cells [7,8]. These molecules have been reported to act asantioxidants and some lignan types are categorized as phytoestro-gens, naturally occurring estrogen-like chemicals [9]. Lignans areformed primarily from oxidative coupling of two phenylpropanemoieties at their b-carbon atoms. Further oxidations and ringformation reactions result in different classes of lignans with a widestructural diversity. The butyrolactone ring, which is the mainstructural feature of dibenzylbutyrolactone lignans, is one of themost abundant structural moieties in naturally occurring lignans.

E. Peuhu et al. / Biochemical Pharmacology 86 (2013) 571–583572
Substantial health benefits have been implicated with lignan-rich diets, including prevention of cardiovascular disease andreduced risk of hormone-dependent cancers [10]. A meta-analysisof several studies revealed that high lignan exposure might indeedcorrelate with a reduced breast cancer risk in postmenopausalwomen [11]. Lignans may exert the observed anticancer effectsby inhibition of cancer cell proliferation, induction of apoptoticcell death, or reduction of tumor angiogenesis and metastasis[7,12–16]. The molecular mechanisms that have been identifiedfor lignan-mediated cellular events include downregulation ofantiapoptotic proteins [17,18], direct inhibition of kinases [19],and inhibition of cell survival signaling [20–22].
Constitutive Akt signaling has been found to protect androgen-dependent prostate cancer cells from tumor necrosis factor-relatedapoptosis-inducing ligand (TRAIL)-induced apoptosis [23,24]. Aslignans are able to inhibit Akt activity, prostate cancer cells can besensitized to TRAIL-induced apoptosis by lignan treatment [8].Binding of TRAIL to the apoptosis-inducing cell surface receptors,DR4 (TRAIL-R1) and DR5 (TRAIL-R2), initiates caspase activationand cell death selectively in malignant cells [25]. EndogenousTRAIL is widely expressed in hematopoietic cells with a prominentrole in immune surveillance and inhibition of tumor metastasis[26–28]. While several TRAIL-receptor agonists are being evaluat-ed in clinical trials for cancer treatment, resistance has alreadybeen reported, suggesting that combination with other anti-cancertreatments is likely to be required for successful TRAIL-therapy[29]. Thus, new and carefully chosen combinations to TRAIL-therapy are needed.
Our previous observations of lignan anticancer activity led us toexplore, which types of lignans are the most effective insensitization of androgen-dependent prostate cancer cells toTRAIL-induced apoptosis, and what are the critical structuraldeterminants and mechanisms of this activity. Here, we report thatthe nortrachelogenin (NTG) lignan was identified as the mostefficient of the tested lignan compounds in sensitization ofandrogen-dependent LNCaP prostate cancer cells to TRAIL-inducedapoptosis. The mechanism of this reversible action includesinhibition of RTKs in response to growth factor stimulation andinhibition of Akt activity and membrane localization. These resultsprovide interesting possibilities for employment of NTG incombinatorial therapy with TRAIL and other therapeutic agentsagainst prostate cancer.
2. Materials and methods
2.1. Preparation of lignans
In total, 18 lignans (Fig. 1A) and 9 norlignans (Fig. 1B) wereprepared by semisynthetic methods using hydroxymatairesinol(compound 2) as a starting material. A 100 mM stock solution of allcompounds was prepared in EtOH or DMSO at room temperature.All the chemicals used in this study were purchased from Sigma–Aldrich.
Hydroxymatairesinol was isolated from Norway spruce(Picea abies) knotwood material as previously described [30].Matairesinol (MAT) and enterolactone (ENL) were preparedaccording to previously described methods [31]. Dimethylmatair-esinol (MeMR) was prepared by methylation of MAT with MeI andK2CO3 in dry acetone. The corresponding didemethylmatairesinol(demethylMR) was prepared by demethylation of MAT using AlCl3
in pyridine. Oxomatairesinol (OMR) was prepared by oxidationof HMR as previously described [32]. 7-Methoxymatairesinol(7MeOMR) was obtained by treatment of HMR with MeONa inmethanol [33]. The cyclooctadienelignan COD was prepared bycyclisation of MeMR by VOF3 according to the literature [34]. 7-Hydroxysecoisolariciresinol (7-OH SECO), lariciresinol (LARI) and
cyclolariciresinol (CLARI) were prepared according to our previ-ously reported method [35]. Conidendrin was obtained by acidtreatment of HMR and the corresponding dimethylconidendrin(DMCON), by methylation equally as for MAT as described above.Secoisolariciresinol (SECO) was prepared by LiAlH4 reduction ofMAT essentially in the same way as for 7-OH SECO [35]. Pinoresinol(PINO) was isolated from spruce resin as described by Erdtman[36]. Northrachelogenin (NTG) was isolated from pine knotwoodmaterial as previously described [37]. The lactol carissanol (CARS)and the diol carinol (CAR) were prepared by LiAlH4 reduction ofNTG according to the literature [38]. The 9-norlignans X, X-Me andimperanene (IMP) were semisynthetically prepared from HMR aspreviously published [39]. Further acid catalyzed cyclisation ofthese derivatives gave cycloX, cycloXMe, cycloXOH which werefurther transformed to the corresponding arylnaphtalene deriva-tives ArX and ArxMe by DDQ oxidation. The saturated derivativedHX was obtained by hydrogenation with Pd/C and H2 in ethanol.The identity and purity of all compounds was analyzed by NMR-spectroscopy, HRMS and GC–MS.
2.2. Cell culture and treatments
The LNCaP prostate cancer cells (clone FGC from EACC) werecultured in RPMI-1640 medium (Sigma–Aldrich) supplementedwith 10% fetal calf serum (FCS) (BioClear), 2 mM L-glutamin, 100 U/mlpenicillin, 100 mg/ml streptomycin and 1 mM sodium pyruvate(Sigma–Aldrich). LNCaP cells were expanded on delivery, and vialsof low passage cells were preserved in liquid nitrogen. LNCaP cellsused in this study were within 20 passages after resuscitation. Oneday prior to treatment, medium of LNCaP cells was changed tophenol red free RPMI-1640 (Gibco) with or without 1 nM syntheticandrogen (Mibolerone). This medium was supplemented with 10%charcoal/dextran treated fetal bovine serum (Hyclone), L-glutamine,penicillin and streptomycin. PC-3, 22rv1 cells were cultured inRPMI-1640 medium (Sigma–Aldrich) supplemented with 10% fetalcalf serum (BioClear), 2 mM L-glutamin, 100 U/ml penicillin, and100 mg/ml streptomycin (Sigma–Aldrich). SV40 transformed pros-tate epithelial RWPE-1 cells (#CRL-11609, obtained in 2006 fromATCC) were cultured in Keratinocyte medium (KSFM; Gibco) with100 U/ml penicillin, 100 mg/ml streptomycin, 1% fetal calf serum(BioClear), 5 ng/ml human recombinant epidermal growth factor(EGF; Gibco) and 50 mg/ml Bovine Pituitary Extract (Gibco).Authentication of cell lines in ATCC is performed through shorttandem repeat profiling (human cells), karyotyping, and cytochromeC oxidase I testing.
The cells were treated with 0–100 mM lignans or solventcontrol for indicated times. Human recombinant isoleucin-zipperTRAIL (izTRAIL; kindly provided by Professor Henning Walczak)was used at 0–1 mg/ml with an incubation time of 20 h. Prior to10 min treatment with 10 nM IGF-1 (Millipore) or 10 nM insulin(Sigma) the cells were serum-starved (1% FCS) overnight.
2.3. Detection of apoptotic parameters
Flow cytometric analysis of apoptosis with HTS platform wasperformed on cells that were plated on a 96-well plate as triplicatesamples. After 24 h in androgen-deprived conditions, the cellswere pretreated with 40 mM lignans or lignan derivatives for 1 hprior to addition of 10 ng/ml izTRAIL. Twenty hours later the platewas centrifuged with a culture plate rotor (1000 rpm, 3 min). Foranalysis of nuclear fragmentation, propidium iodide (PI) buffer(40 mM Na-citrate, 0.3% Triton X-100, 50 mg/ml PI; Sigma) wasadded to the wells. After 10 min incubation at room temperature,the plate was analyzed with LSRII flow cytometer equipped withHTS platform (PE-A channel). The fraction of sub-G0/G1 events(nuclear fragmentation) was gated as a measure of apoptotic cell

Fig. 1. Molecular formulas of the screened lignans (A) and norlignans (B).
E. Peuhu et al. / Biochemical Pharmacology 86 (2013) 571–583 573
death. For detection of mitochondrial depolarization, the cells weredetached by trypsinization of the cells and incubated in 20 nMtetramethylrhodamine (TMRM; Invitrogen) in warm culturemedium for 10 min before analysis. The plate was analyzed withLSRII flow cytometer equipped with HTS platform (FSC, PE-Achannel). The cells with decreased TMRM incorporation wereconsidered to have undergone mitochondrial membrane depolar-ization. Activated caspase-3 in cells was labeled with phycerythrin(PE)-conjugated antibody according to manufacturer’s protocol
(BD Pharmingen) and analyzed by FacsCalibur flow cytometer(FL-2, FSC).
2.4. Clonogenic assay
The effect of NTG alone and in combination with TRAIL oncolony formation ability of the LNCaP cells was assessed by softagar colony formation assay [40]. Briefly, five hundred LNCaP cellswere suspended in 1 ml of 0.35% soft agar medium (Agarose,

Table 1The screened lignans and norlignans.
Compound
Dibenzylbutyrolactone lignans
1 Matairesinol
2 7-Hydroxymatairesinol
3 7-Oxomatairesinol
4 4,40-Dimethylmatairesinol
5 7-Methoxymatairesinol
6 4,40-Dihydroxyenterolactone/3,30-Didemethylmatairesinol
7 Nortrachelogenin
8 Enterolactone
Aryltetralin lignans
9 Conidendrin
10 Dimethylconidendrin
11 Cyclolariciresinol
12 4,40-Dimethylcyclolariciresinol
Dibenzylbutanediol lignans
13 Secoisolariciresinol
14 Dimethylsecoisolariciresinol
15 7-Hydroxysecoisolariciresinol
Tetrahydrofuro-furan lignans
16 Pinoresinol
Tetrahydrofuran lignans
17 Lariciresinol
Dibenzocyclo-octadiene lignans
18 3a,4,13,13a-Tetrahydro-6,7,10,11-
tetramethoxydibenzo[4,5:6,7]cycloocta[1,2-c]furan-1(3H)-one
9-Norlignans
19 4,40-Dihydroxy-3,30-dimethoxy-90-norlign-70-en-9-oic acid
20 Methyl 4,40-dihydroxy-3,30-dimethoxy-90-norlign-70-en-9-oate
21 Imperanene
22 4,40-Dihydroxy-3,30-dimethoxy-9-norlign-
90-oic acid
23 4,40-Dihydroxy-3,30-dimethoxy-60 ,7-cyclo-9-norlign-90-oic acid
24 Methyl 4,40-dihydroxy-3,30-dimethoxy-60 ,7-cyclo-9-norlign-90-oate
25 4,40-Dihydroxy-3,30-dimethoxy-60 ,7-cyclo-9-norlign-90-ol
26 4,40-Dihydroxy-3,30-dimethoxy-9-nor-60 ,7-cyclolign-7,
70-diene-90-oic acid
27 Methyl 4,40-dihydroxy-3,30-dimethoxy-9-nor-60 ,7-
cyclolign-7,70-diene-90-oate
E. Peuhu et al. / Biochemical Pharmacology 86 (2013) 571–583574
BioNordika, Helsinki, Finland) and plated onto a layer containingsolidified 0.5% soft agar medium in six well plates. 2 mL of RPMI1640 growth media was added onto the top layer and the plateswere maintained at 37 8C. The plates were treated after 24 h withthe indicated compound additions and new media was addedalong with fresh compounds once in every 7 days. Colonyformation was monitored for 21 days. After 21 days, thecolonies were stained with 0.05% crystal violet (Sigma) for 1 hand washed with water. Colonies were photographed under anEVOS digital inverted microscope. Cropped and thresholdedimages were analyzed with ImageJ software for the total area ofcell colonies.
2.5. Confocal microscopy
After treatments, the cells cultured on coverslips were washedwith PBS and fixed with 3% paraformaldehyde for 15 min at roomtemperature. The samples were permeabilized and blocked for 1 hin 3% BSA 0.1% Tween in PBS. Akt was labeled with rabbit anti-Aktantibody (Cell Signaling) and Alexa 546-conjugated anti-rabbitsecondary antibody (Molecular Probes). After washing with PBS,the samples were mounted with DAPI Vectashield (VectorLaboratories) and viewed by Zeiss LSM 510 META laser-scanningconfocal microscope (63� oil objective, 543 nm excitation). Theimages were quantified using ImageJ and presented as histogram(mean � SEM; n � 3; Student’s t-test ***P < 0.001) and line graph.
2.6. Immunoprecipitation
The cells cultured on 10 ml plates were placed on ice and rinsedwith cold PBS. The cells were lysed in 1 ml of cold lysis buffer(50 mM Tris pH 7.4, 150 mM NaCl, 1 mM EDTA, 1.25% CHAPS,1 mM Na3OV4, 2 mM NaF, 10 mM sodium pyrophosphate: Sigma–Aldrich) and complete protease inhibitor cocktail (Roche AppliedScience) for 10 min. The collected lysate was centrifugated at15,000 � g for 10 min. Supernatant was collected and subjected topreclearing with Protein A sepharose beads (Sigma) and Protein Gsepharose beads (Amersham Biosciences) for 1 h at +4 8C. Afterpreclearing, the lysate sample was collected and rest of the samplewas divided for immunoprecipitation with 5 mg anti-insulinreceptor ß or 5 mg anti-IGF-I receptor ß antibody (Santa CruzBiotechnology). After incubation overnight at +4 8C, Protein A andProtein G sepharose beads were added to samples and incubatedfurther for 4 h. The samples were washed 4 times with lysis bufferand the beads were resuspended to 50 ml of Laemmli sample buffer(LSB) and boiled for 10 min.
2.7. Western blotting
Whole cell lysates were prepared by lysing floating andattached cells in Laemmli sample buffer and boiling the samplesfor 10 min. Proteins were separated by SDS-PAGE and transferredon PVDF membrane (Millipore). Western blotting was performedusing antibodies against Bid, caspase 9, Akt and phospho-Akt(ser473) (Cell Signaling Technology), Actin (clone AC-40;Sigma–Aldrich), PI3K p85a (clone 4/PI3-kinase; BD Pharmingen),Insulin receptor ß and IGF-I receptor ß (Santa Cruz Biotechnology),and phospho-tyrosine (clone 4G10; Millipore). HRP-conjugatedsecondary antibodies were from Southern Biotechnology Associ-ates, Promega, and Amersham Biosciences. The results werevisualized using the ECL method (Amersham Biosciences) on x-rayfilm. The presented Western blot results are representative of atleast three independent experiments. Densitometric analysis ofWestern blots was done using scanned films with the MCID M5+software. The values were normalized to the untreated controlsample, which was given the value 1. The histograms in Fig. 6A and
B present a relative decrease in ratio of phosphorylated vs. non-phosphorylated protein compared to control cells.
2.8. Statistical analysis
The graphs were prepared with GraphPad Prism software andthey represent mean values � standard error of mean (SEM). Thenumber of independent experiments and the statistical significance isindicated in the figure legends.
3. Results
3.1. NTG is a novel lignan for sensitization to TRAIL-induced apoptosis
In order to discover new lignan compounds with the capacity tosensitize TRAIL-resistant LNCaP prostate cancer cells to apoptosis-induction by TRAIL, and to further identify the critical structures inthe lignan molecule for this effect, we assayed 27 lignans or lignanderivatives (Table 1, Fig. 1A and B). Androgen-deprived LNCaP cellswere pretreated with the tested compounds or the solvent controlsfor 1 h before addition of human recombinant isoleucine-zipper (iz)TRAIL. After 20 h the cells were analyzed for nuclear fragmentationor mitochondrial depolarization using a LSRII flow cytometer HTS-unit in 96-well format. The formation of propidium iodide labeledDNA fragments (Fig. 2A) was used as the primary indicator ofapoptotic cell death, and a subset of lignans was also assayed forreduction of TMRM labeling in depolarized mitochondria (Fig. 2B).
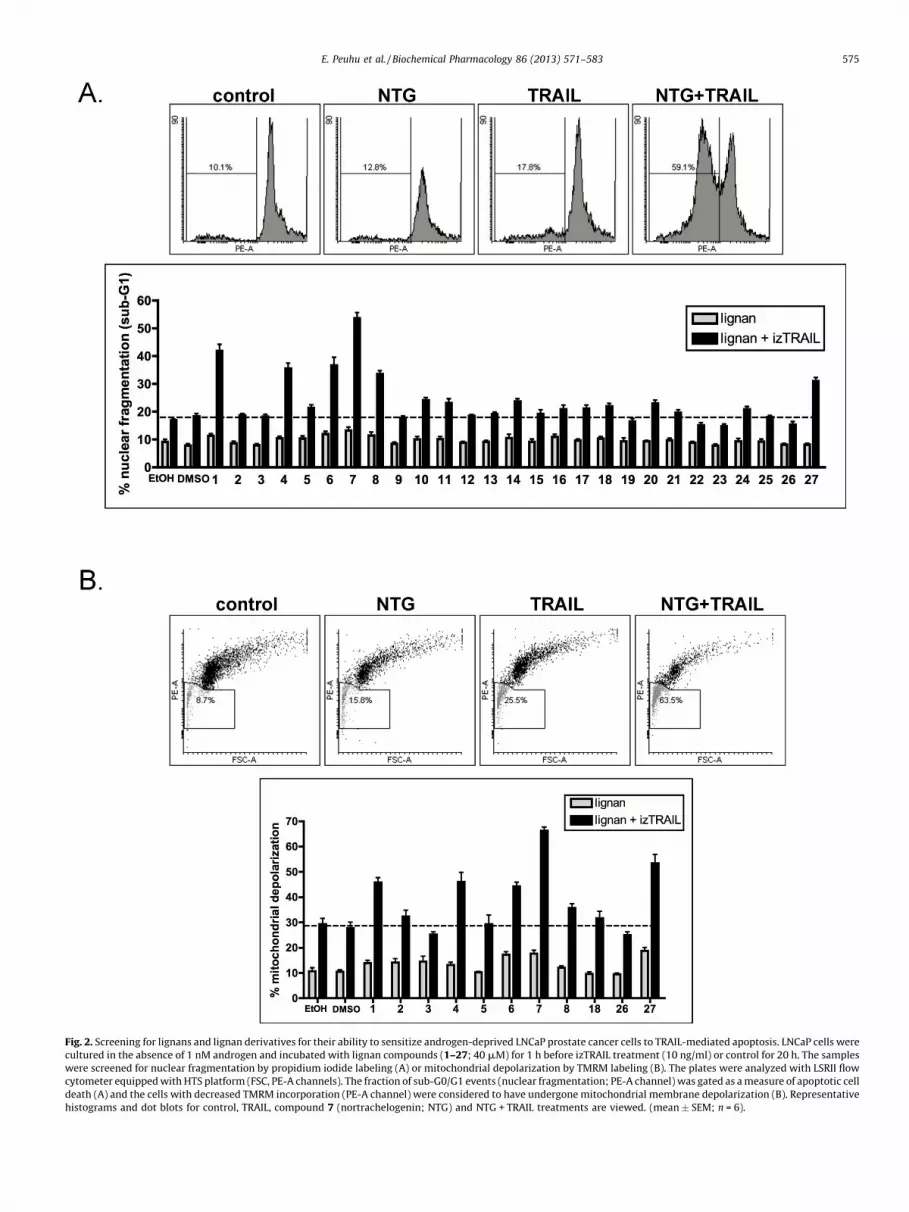
Fig. 2. Screening for lignans and lignan derivatives for their ability to sensitize androgen-deprived LNCaP prostate cancer cells to TRAIL-mediated apoptosis. LNCaP cells were
cultured in the absence of 1 nM androgen and incubated with lignan compounds (1–27; 40 mM) for 1 h before izTRAIL treatment (10 ng/ml) or control for 20 h. The samples
were screened for nuclear fragmentation by propidium iodide labeling (A) or mitochondrial depolarization by TMRM labeling (B). The plates were analyzed with LSRII flow
cytometer equipped with HTS platform (FSC, PE-A channels). The fraction of sub-G0/G1 events (nuclear fragmentation; PE-A channel) was gated as a measure of apoptotic cell
death (A) and the cells with decreased TMRM incorporation (PE-A channel) were considered to have undergone mitochondrial membrane depolarization (B). Representative
histograms and dot blots for control, TRAIL, compound 7 (nortrachelogenin; NTG) and NTG + TRAIL treatments are viewed. (mean � SEM; n = 6).
E. Peuhu et al. / Biochemical Pharmacology 86 (2013) 571–583 575

Fig. 3. NTG amplifies TRAIL-induced caspase-3 activation and inhibits colony formation in prostate cancer cells. (A) LNCaP cells (�1 nM androgen) were incubated with 0–
120 mM NTG for 1 h before addition of izTRAIL (10 ng/ml). After 20 h incubation, the percentage of nuclear fragmentation was detected by flow cytometry (mean � SEM; n � 3;
Student’s t-test *P < 0.05, **P < 0.01, ***P < 0.001). (B) LNCaP cells (�1 nM androgen) were incubated with 40 mM NTG or solvent control for 1 h before addition of izTRAIL in different
concentrations. After 20 h incubation, the percentage of cells with activated caspase-3 was detected by flow cytometry (mean � SEM; n � 3; Student’s t-test **P < 0.01, ***P < 0.001).
E. Peuhu et al. / Biochemical Pharmacology 86 (2013) 571–583576

E. Peuhu et al. / Biochemical Pharmacology 86 (2013) 571–583 577
The role of the lignan skeleton, oxidation degrees, as well as thearomatic substitution pattern was studied by varying these factorsby semisynthetic transformations of the lignan derivatives. Thetested molecules have been listed in Table 1 and their molecularstructures are presented in Fig. 1A and B.
As we previously reported that compound 1 (matairesinol) isable to sensitize the TRAIL-resistant LNCaP cells to apoptosis (8),matairesinol was used as a reference for the activity of the otherlignans. Our results show that none of the tested lignans arecytotoxic to LNCaP cells without combination to TRAIL (Fig. 2A).For further analysis of structural determinants of the biologicalactivity, compounds 1–8 were selected for mitochondrial assay(Fig. 2B). Among these compounds, only dibenzylbutyrolactonelignans without a bond between the benzyl groups (compounds 1,2, 4, 6, and 7) are active in sensitization to TRAIL-induced apoptosis(Fig. 2A and B). Two compounds that have actively studies before,compound 2 (hydroxymatairesinol) and compound 8 (enterolac-tone), were less effective than matairesinol, which is in agreementwith previous observations. Methylation (compound 4) ordemethylation (compound 6) of the aromatic rings did notmarkedly disturb the sensitizing effect of matairesinol, butaddition of methyl (compound 5), hydroxyl (compound 2), oroxo (compound 3) groups at position C-7 completely abolished theactivity (Fig. 2A and B). Also compound 13 (secoisolariciresinol),which differs from matairesinol by the lack of the lactone ring, wascompletely inactive in sensitization to TRAIL, suggesting animportant role of the g-butyrolactone moiety of the molecule(Fig. 2A). Most of the 9-norlignans (compounds 19–27 that lackcarbon 9 from the lignan skeleton) were inactive (Fig. 2A), exceptfor the arylnaphtalene structure, compound 27 (Fig. 2A and B).Interestingly, compound 7 (nortrachelogenin; NTG) was found tobe most efficient of the tested compounds to sensitize theandrogen-deprived LNCaP cells to TRAIL treatment (Fig. 2A and B).
3.2. NTG amplifies TRAIL-induced caspase-3 activation and inhibits
colony formation in prostate cancer cells
As NTG was identified as the most efficient TRAIL-sensitizinglignan, we tested the effective dose range for NTG treatment. LNCaPcells were cultured in androgen-supplied and androgen-deprivedconditions to simulate the effects of androgen-deprivation therapyand pretreated with increasing concentrations of NTG (0–120 mM)before cell death induction with TRAIL. Analysis of nuclearfragmentation shows that both androgen-deprived and androgen-supplied LNCaP cells became sensitized to apoptosis with approxi-mate EC50 values for NTG being 32 mM and 58 mM, respectively(Fig. 3A). We also analyzed the ability of NTG to affect the efficacy ofdifferent TRAIL concentrations in triggering of caspase-3 activation(Fig. 3B). LNCaP cells were treated with increasing concentrations ofTRAIL alone or with the combination of TRAIL and 40 mM NTG(Fig. 3B). The percentage of cells with activated caspase-3 increasedfrom maximum of 15% to 60% for androgen-deprived, and from 13%to 30% for androgen-supplied cells (Fig. 3B). Importantly, evenprolonged incubation (24–72 h) with 0–100 mM NTG alone did notinduce apoptosis in LNCaP cells (Fig. 3C), although dibenzylbutyr-olactone lignans have previously been suggested to facilitate theinitiation of the mitochondrial pathway of apoptosis [7].
To investigate the effects of NTG alone and in combination withTRAIL on colony formation ability, the LNCaP cells were plated at
(C) LNCaP cells (�1 nM androgen) were incubated with 0–100 mM NTG for 24–72 h and t
conjugated antibody according to manufacturer’s protocol (BD Pharmingen) and quant***P < 0.001). (D) LNCaP cells were plated in low confluency in soft agar for 24 h and the
concentrations were used in when these two agents were added individually to the cells. Af
EVOS digital microscope and quantified using ImageJ (mean � SEM; n = 3; Student’s t-test **
before addition of 10 ng/ml izTRAIL for 20 h. Total cell lysates were prepared and analyze
measured as loading control (mean � SEM; n = 3; Student’s t-test *P < 0.05, **P < 0.01, ***P
very low confluency in soft agar, treated and allowed to formcolonies for 21 days. When the colonies were observed and countedafter treatment, 10 ng/ml TRAIL was not able to inhibit LNCaPgrowth (Fig. 3D). However, 40 mM NTG significantly inhibited thegrowth of the plated cells visible as very small colonies, whereas NTGand TRAIL treatment in combination resulted in significant death ofthe cells and virtually no colonies were formed (Fig. 3D). The latterresults are well in agreement with the data showing that NTG incombination with TRAIL is a potent inducer of apoptosis, as reflectedby the total absence of colonies (Fig. 3D). In contrast, in the absenceof TRAIL, NTG did not kill the cells, but inhibited efficiently cellgrowth, as indicated by the presence of miniscule colonies in cellstreated with NTG only. The NTG-mediated effects on cell growthcorrespond well with the effects of NTG on growth factor signaling.NTG increased TRAIL-induced Bid cleavage in androgen-deprivedLNCaP cells and promoted caspase-9 activation in response to TRAILtreatment irrespective the presence of androgen (Fig. 3E), furtherdemonstrating that the mechanisms by which NTG sensitizesprostate cancer cells to TRAIL-induced apoptosis are very similar tothose observed for matairesinol [8].
3.3. Androgen independent PC3 and androgen dependent 22Rv1
prostate cancer cells are not effectively sensitized by NTG to TRAIL-
induced apoptosis
To evaluate how NTG treatment affects TRAIL-induced apopto-sis in other prostate cancer cell lines, we tested androgen-independent PC3 and androgen-dependent 22rv1 cells. PC3 cellsare not resistant to TRAIL and were not further sensitized to TRAIL-induced apoptosis (Fig. 4A). On the other hand, TRAIL-resistant22rv1 cells that have previously been characterized as PTEN-positive [41] and thus exhibit significantly lower levels of Aktactivity [42] could not be sensitized to TRAIL-induced apoptosis byNTG treatment (Fig. 4B). These data suggest that determination ofthe PI3K/Akt signaling status could assist significantly in predictingwhether TRAIL-resistant prostate cancer cells respond to thiscombinatorial treatment.
As both TRAIL and lignans have been demonstrated to displaycancer-specificity [7,8,29], the combinatorial NTG and TRAILtreatment might be well tolerated by non-cancerous tissues. Toexamine this possibility in vitro, we tested how the RWPE-1 non-malignant prostate epithelial cell line responds to the treatment.Interestingly, this cell line did not display any sensitization toTRAIL when pretreated with 40 mM NTG (Fig. 4C), suggestingpotential cancer-specificity of the observed effect.
3.4. The NTG lactone ring has an essential role in sensitization of
prostate cancer cells to TRAIL-induced apoptosis
The structure of the NTG (compound 7) differs from matair-esinol (compound 1) by one hydroxyl group located on the lactonering (Fig. 1). This difference might affect the polarity of themolecule and may also modulate the interaction properties of thelignan. As the lactone ring structure seemed important for MATfunction in sensitization to TRAIL-induced apoptosis, we synthe-sized two derivatives of NTG lacking this functional group. NTGwas reduced to the lactol derivative (carissanol; CARS) and the diol(carinol; CAR) (Fig. 5). When compared to the activity of NTG inpotentiation of TRAIL-induced apoptosis, CARS and CAR were
he percentage of cells with activated caspase-3 was labeled with phycerythrin (PE)-
ified by flow cytometry (mean � SEM; n � 3; Student’s t-test *P < 0.05, **P < 0.01,
n incubated with 40 mM NTG for 1 h before addition of 10 ng/ml izTRAIL. The same
ter 21 days of incubation, cell colonies were stained with crystal violet, imaged using a*P < 0.001). (E) LNCaP cells (�1 nM androgen) were incubated with 40 mM NTG for 1 h
d by Western blotting for cleavage of Bid, caspase-9. Actin (45 kDa) expression was
< 0.001).

Fig. 4. Androgen independent PC3 and androgen dependent 22Rv1 prostate cancer cells are not effectively sensitized by NTG to TRAIL-induced apoptosis. (A) PC3 cells were
treated with 40 mM NTG or solvent control for 1 h prior to addition of izTRAIL 0–20 ng/mL. After 20 h, the cells were stained with propidium iodide and analyzed by flow
cytometry as described in Section 2 (mean � SEM; n = 3). (B) 22Rv1 were cultured in the absence of androgen and treated with 40 mM NTG or solvent control for 1 h prior to
addition of izTRAIL 0–100 ng/mL. After 20 h, the cells were stained with propidium iodide and analyzed by flow cytometry as described in Section 2 (mean � SEM; n = 3). (C) Non-
malignant prostate epithelial RWPE-1 cells were treated with 40 mM NTG or solvent control for 1 h before addition of izTRAIL in different concentrations, and analyzed for caspase-3
activation by flow cytometry (mean � SEM; n = 4).
E. Peuhu et al. / Biochemical Pharmacology 86 (2013) 571–583578
found to be inactive (Fig. 5). CAR completely lacked ability tosensitize androgen-deprived and androgen-supplied LNCaP cells toTRAIL-induced apoptosis, and thereby confirmed the critical roleof an intact hydroxylated g-butyrolactone ring in effectivecombinatorial treatment with TRAIL.
3.5. NTG inhibits Akt and its recruitment to the plasma membrane
As we have previously observed that matairesinol down-regulates Akt activity and subsequently sensitizes LNCaP cells to
Fig. 5. Nortrachelogenin (NTG) lactone ring is critical in sensitization of LNCaP prosta
incubated with 40 mM NTG, CARS, CAR (structures depicted) or solvent control for 1 h be
activated caspase-3 was detected by flow cytometry (mean � SEM; n � 3).
TRAIL-induced apoptosis [8], we wanted to elucidate, if NTG actswith similar mechanisms and determine the kinetics of this effect.Akt activity was determined by Western blotting with phospho-specific antibodies detecting the levels of Akt phosphorylation atserine-473 as compared to the total amount of Akt. As anticipated,Akt phosphorylation was efficiently downregulated by 40–100 mMNTG (Fig. 6A) with the maximum inhibition occurring after 2 hwhen the cells were treated with 100 mM NTG (Fig. 6B). Somerecovery of Akt phosphorylation started to take place around the4 h time point (Fig. 6B). Phosphorylation of a downstream target of
te cancer cells to TRAIL-mediated apoptosis. LNCaP cells (�1 nM androgen) were
fore addition of 10 ng/ml izTRAIL. After 20 h incubation the percentage of cells with

E. Peuhu et al. / Biochemical Pharmacology 86 (2013) 571–583 579
Akt, the GSK-3ß kinase, at serine-9 was also observed in responseto NTG and this inhibition coincided with decreased Aktphosphorylation (data not shown), further corroborating ourresults.
As Akt phosphorylation and activation also involves recruit-ment to the plasma membrane, we determined the subcellularlocalization of Akt in androgen-deprived LNCaP cells before andafter 2 h NTG treatment. Akt phosphorylation was significantlydecreased in response to 40 mM NTG (Fig. 6A and B). Interestingly,when the localization of cellular Akt was assessed by fluorescentmicroscopy, we observed that the fraction of cytoplasmic Akt wasdramatically increased (Fig. 6C), indicating that NTG is likely tohave a synergistic effect influencing both the activity of Akt andsubcellular localization of the kinase. We could not detect anyeffects on phosphorylation of Jun N-terminal kinase (JNK) afterNTG treatment, and interestingly detected very low phosphory-lation of the mitogen-activated kinase ERK in LNCaP cells onlyupon treatment with the phorbol ester PMA (data not shown). Theinhibitory effect of NTG on the Akt pathway was stronger and morepersistent when no androgen was supplied to cells (Fig. 6A and B),demonstrating the counteracting mechanisms induced by andro-gen stimulation in the androgen-dependent prostate cancer cells.
3.6. NTG potently inhibits receptor tyrosine kinases (RTKs) upon
growth factor stimulation
In this study, we wanted to extend the investigations to thepotential effects of lignan action in the upstream regulators of theAkt signaling pathway. Growth factors are powerful activators ofmitogenic signaling that promote the growth and survival ofprostatic tumors [43]. Ligand binding to growth factor receptors(GFRs) leads to autophosphorylation, activation of the involvedRTKs and propagation of a number of key survivalsignalingmodules, including the PI3K/Akt pathway.
Previous results indicate that enterolactone (compound 8)inhibits IGF-I receptor signaling in prostate cancer cells [20]. Inlight of these observations, we determined whether NTG has thecapacity to modulate GFR activity. To this end, we incubated theserum-starved LNCaP cells with 0–100 mM NTG for 2 h prior tostimulation with growth factors and observed the level of receptorautophosphorylation. After LNCaP cells were activated for 10 minwith insulin or IGF-I, insulin receptor (IR) b-subunit and IGF-Ireceptor (IGF-IR) b-subunit were immunoprecipitated. IR becametyrosine phosphorylated in response to both insulin and IGF-I(Fig. 6D), as IGF-I activates IR/IGF-IR hybrid receptors when bothreceptor genes are co-expressed in a cell [44]. The receptor tyrosinekinase activity of IGF-IR was induced effectively by IGF-I (Fig. 6D).The already high Akt phosphorylation level in LNCaP cells was notfurther increased by the growth factors (Fig. 6D). In cells pretreatedwith 40–100 mM NTG, we observed an impressive effect. NTGevoked a drastic decrease in the amount of activation-mediatedreceptor autophosphorylation in response to insulin or IGF-I. Alsothe amount of receptor-associated PI3K p85 regulatory subunitwas clearly reduced upon NTG treatment, also in the absence ofgrowth factor stimulation, which is indicative of reduced PI3Kactivation. Interestingly, our preliminary results suggest that theRTK suppressing activity of NTG is not restricted to insulin and IGF-I signaling, but it extends to inhibition of other RTKs as well (datanot shown).
3.7. NTG has a rapidly reversible effect on Akt activity
and TRAIL-sensitivity
To examine the molecular dynamics of NTG in our experimentalsystem, the androgen and NTG treatments of LNCaP cells werefollowed by medium change and further incubation for 0–2 h.
When recovery of Akt activity was examined by Western blotting,Akt phosphorylation was clearly seen to return 1 h after thewashout (Fig. 7A). This recovery was observed although we used ahigh NTG concentration (100 mM) and 2 h pretreatment for thisassay in order to detect a clear downregulation in Akt phosphor-ylation level. Because similar recovery kinetics applied inandrogen-deprived and androgen-supplied cells, it seems thatandrogen does not affect the persistence of the NTG-induced Aktinhibition (Fig. 7A).
As 1 h pretreatment with 40 mM NTG sensitized to TRAIL-induced apoptosis, we wanted to test how replacement of theculture medium directly before addition of TRAIL would influencethe outcome. Interestingly, this arrangement completely abolishedthe sensitizing effect of NTG with no difference detected betweenandrogen-deprived and androgen-supplied cells (Fig. 7B). Theseresults suggest that the effect of NTG on Akt survival signaling andTRAIL-sensitivity is rapidly reversible and may involve directinhibitory mechanisms at the proximity of the plasma membrane.
4. Discussion
Targeting the growth factor activated PI3K/Akt pathway isconsidered a promising therapeutic approach against prostatecancers with mutations in the PI3K catalytic subunit or deficiencyof the PTEN phosphatase [6]. Our study highlights lignans asinhibitors of this pathway with potential as dietary factors ortherapeutic drugs to be used in treatment of androgen-dependentprostate cancer. Our previous studies showed that the lignanmatairesinol (compound 1) is able to sensitize LNCaP cells toTRAIL-induced apoptosis by inhibition of Akt-signaling [8]. Now,we have screened various lignans and lignan derivatives, identifieda more effective lignan, NTG, for combinatorial therapy with deathreceptor activation by TRAIL, and pointed out the molecularfeatures of NTG that are critical for this application. Interestingly,the combination of TRAIL to NTG treatment did not harm anepithelial prostate cell line, implying that different mechanismsprotect these non-malignant cells from TRAIL.
The screening results clearly showed that the dibenzylbutyr-olactone skeleton is the most active structure. The substitutionpattern at the aromatic rings had only a small effect on the activity(compound 1 vs. compounds 4, 6 and 8) whereas the g-butyrolactone ring was crucial for activity. Reduction of thelactone to the corresponding diol structure led to almost inactivecompounds (compound 1 vs. compound 13), but also modifica-tions such as cyclizations into other types of lignans led to decreasein activity (compound 1 vs. compound 9). Addition of substituentsat the benzylic position (carbon 7) resulted in less activecompounds in all cases. As a result, the butyrolactone as anindependent structure is not responsible for high activity, but theactive structure arises from a dibenzyl substituted g-butyrolactonewithout substituents at the benzylic positions. The introduction ofan a-hydroxyl group in the lactone at C-8 position clearly enhancesthe activity as seen for NTG. The OH-group may influence thepolarity/solubility, as well as the hydrogen-bonding properties.The reduction of NTG to the lactol, resulting in an inactivecompound (carissanol; Fig. 5), indicates that the polarity/solubilitymay not be critical. However, it shows that a carbonyl group isneeded, which indicates that the activity of NTG in modulation ofTRAIL-induced apoptosis requires a hydrogen bond acceptor.Understanding the molecular basis of lignan function as aninhibitor of survival signaling and a sensitizer to TRAIL-inducedapoptosis exposes opportunities to produce lignans with increasedactivity for various forms of cancer therapy.
There are in fact previous indications of favorable bioactivity ofa compound related to NTG. An early report on the NTGenantiomer wikstromol showed antileukemic activity of this

Fig. 6. NTG inhibits receptor tyrosine kinase (RTK) activity and Akt signaling. (A) LNCaP cells (�1 nM androgen) were incubated with 0–100 mM NTG for 1 h or (B) with 100 mM
NTG for 0–8 h after which total cell lysates were prepared. The western blot analysis with Akt and phospho-Akt(ser473) was quantified. The histogram presents a relative decrease in
pAkt/Akt ratio compared to control cells (mean � SEM; n = 3; Student’s t-test *P < 0.05, **P < 0.01, ***P < 0.001). (C) Androgen-deprived LNCaP cells were incubated with 100 mM
NTG for 2 h, fixed and labeled with Akt antibody followed by Alexa 546-conjugated secondary antibody. (D) Serum-starved LNCaP cells were pretreated with 0–100 mM NTG and
E. Peuhu et al. / Biochemical Pharmacology 86 (2013) 571–583580

Fig. 7. The effect of NTG on Akt inhibition and TRAIL-sensitivity is rapidly reversible. (A) LNCaP cells (�1 nM androgen) were treated with 100 mM NTG or solvent control for 2 h
prior to medium change. Total cell lysates were prepared from samples 0, 1 h or 2 h later and analyzed by Western blotting for Akt and phospho-Akt(ser473) expression
(mean � SEM; n = 3; Student’s t-test *P < 0.05, **P < 0.01, ***P < 0.001). (B) LNCaP cells (�1 nM androgen) were treated with 40 mM NTG or solvent control for 1 h, after which the
culture medium was replaced or alternatively left unchanged before addition of 10 ng/ml izTRAIL. After 20 h incubation the percentage of cells with activated caspase-3 was
detected by flow cytometry (mean � SEM; n = 3; Student’s t-test *P < 0.05, **P < 0.01).
E. Peuhu et al. / Biochemical Pharmacology 86 (2013) 571–583 581
compound [45,46]. While these are still early days in terms of the in
vivo effects of lignans in general and more specifically those ofNTG, all accumulated data show that lignans, including NTG, aretolerated well in vivo. NTG is metabolically inert, as it is notmetabolized but is absorbed and excreted in urine as such [47]. Aprevious report by Lee et al. [46] involved NTG doses up to amaximum of 16 mg/kg in mice without signs of toxicity. This dosewould be in the same concentration range as used in our cellularstudies (16 mg/kg of NTG would correspond to a concentration inthe range of 60 mM). It has also been shown by Saarinen et al. [47]that no significant differences in animal body weights or liverweights were observed in long-term studies when 50-day-oldfemale rats were treated with NTG (15 mg/kg body weight, oraladministration) during a 58-day-treatment period. Unpublished in
vivo data in mice from our own laboratory confirm that also mice
stimulated with 10 nM insulin or IGF-I for 10 min. The cell lysates were subjected to immun
(IGF-IR) b-subunit, and the precipitated complexes were analyzed by Western blotting for p
Expression of PI3K p85a, Akt and phospho-Akt(ser473) was probed from the lysates and
**P < 0.01, ***P < 0.001).
tolerate NTG well, as mice can be dosed i.v. (15 mg/kg/day) with noobservable toxicity over period of two weeks. In single NTG dosesto chicken embryos (i.v.) we found no signs of toxicity up to at least40 mg/kg. Taken together, the published reports and our unpub-lished studies demonstrate that lignans, including NTG, are verywell tolerated, without signs of toxicity even at high doses.
The enhanced TRAIL-induced apoptosis now seen with thecombined NTG treatment suggests that the full potential of theantitumorigenic effects of NTG is likely to be revealed only incombinatorial treatments for cancer, perhaps especially thoseaddressing tumor metastasis.
The protein kinase Akt (PKB) is a major regulator of cell survival[48]. Enterolactone and matairesinol can reportedly inhibit Aktphosphorylation in prostate cancer cells [7,8], and our latest resultsindicate that also NTG inhibits Akt phosphorylation in the presence
oprecipitation with antibodies against insulin receptor (IR) b-subunit or IGF-I receptor
resence of PI3K p85a (85 kDa), IRb (95 kDa), IGF-IRb (95 kDa) and phospho-tyrosine.
all the Western blots were quantified (mean � SEM; n = 3; Student’s t-test *P < 0.05,

E. Peuhu et al. / Biochemical Pharmacology 86 (2013) 571–583582
and absence of androgen. Recent advances in lignan researchindicate that many signaling pathways can be inhibited by lignans,including the NF-kB pathway [49,50], the Wnt/b-catenin pathway[21,22], and some signaling cascades that require production ofreactive oxygen species (ROS) for their activation [51]. It iscurrently not known if NTG also effects these cell survival signalingpathways.
As increasing evidence proposes that the malignant behavior ofprostate tumors is sustained by deregulated activation of GFRs, anumber of targeted therapies are currently developed and testedfor treatment of HRPC [52]. The neolignan magnolol has beenreported to inhibit EGFR signaling in prostate cancer cells [53], andenterolactone was seen to inhibit IGF-IR signaling in androgen-independent PC-3 prostate cancer cells [20]. The results of ourstudy now demonstrate that NTG treatment of androgen-depen-dent prostate cancer cells inhibits signaling through GFRs, namelyIR and IGF-IR, but the effect might not be limited to these tworeceptors. Also the colony formation ability of NTG treated LNCaPcells was substantially reduced, indicative of suppressed mitogenicstimulation. Furthermore, we noted that PI3K p85 was associatedwith IR and IGF-IR even in serum-starved LNCaP cells, anobservation that may be related to the reported constitutive basalactivity of particular PI3K p110 subunit isoforms in PTEN-deficientcells [54]. The interaction of PI3K p85 with the RTKs was clearlyreduced upon NTG treatment both in the presence and absence ofgrowth factors. Exactly how NTG inhibits RTK activity and PI3K p85recruitment requires still further investigation.
The effect of NTG treatment on both Akt activity and TRAIL-sensitivity was rapid and reversible, indicating that NTG is likely toact directly on the components regulating these pathways. Also thecyclolignan, picropodophyllin (PPP) specifically inhibits IGF-IRsignaling and increases TRAIL-induced apoptosis in melanomacells [55]. While PPP can also inhibit tumor growth and metastasisof xenografted uveal melanoma cells in vivo [56], it remains to betested if treatment together with TRAIL-receptor agonistsincreases the observed anticancer effects of PPP. The molecularstructure of NTG and our preliminary results on artificial lipidmembrane systems (data not shown) together imply that NTGdoes not spontaneously cross the plasma membrane, but ratherinteracts at the plasma membrane surface. RTKs as cell surfacereceptors fit well in this profile of potential NTG targets, but alsoother cell surface proteins, such as NADPH oxidases, that areinvolved in RTK activation, should be investigated as potentialtargets of NTG activity. The combinatorial treatment of in vivo
prostate tumors with NTG and TRAIL provides an interestingopportunity for development of novel cancer therapy.
In summary, the results presented here demonstrate that thedibenzylbutyrolactone lignan NTG inhibits several components ofthe RTK/PI3K/Akt pathway in LNCaP prostate cancer cells. NTGenhances TRAIL-induced apoptosis of these androgen-dependentprostate cancer cells, whereas a non-malignant prostate cell line isnot sensitized to TRAIL by NTG treatment. Furthermore, theanticancer effects of the TRAIL and lignan combinatorial treatmentcan be enhanced with androgen withdrawal. The lactone structureof NTG has a key function in the TRAIL-sensitizing effect that israpid and reversible in nature. Now, the diversity of cellular andsystemic effects observed for dibenzylbutyrolactone lignans can beexplained to a significant degree by the influences thesecompounds exhibit on a wide range of molecules in the GFRsignaling pathway.
Conflict of interest
Dr Peuhu, Mr Holmbom, Dr Eklund, Prof. Sjoholm and Prof.Eriksson have filed a patent application concerning the findings ofthis study. Mr Holmbom is an owner and employee in Separation
Research Ltd., and Mr Holmbom, Dr Eklund, and Prof. Sjoholm areowners in ArboNova Ltd. Both companies purify and marketnatural polyphenols.
Acknowledgments
We thank Professor Henning Walczak for kindly providing uswith the human izTRAIL and Professor Olli Kallioniemi for sharingthe RWPE-1 cells with us. Dr Anders Bjorkbom is acknowledged forhis help and expertise in evaluating the effects of lignans onartificial membranes. We are also grateful to Dr Annika Meinanderfor valuable comments while writing this manuscript. Weacknowledge Turku Cell Imaging Core for the technical assistancein flow cytometry and confocal microscopy studies. This study issupported by funding from the Finnish Cancer Foundations andfrom Sigrid Juselius Foundation.
References
[1] Engelman JA. Targeting PI3K signalling in cancer: opportunities, challengesand limitations. Nat Rev Cancer 2009;9:550–62.
[2] Wong KK, Engelman JA, Cantley LC. Targeting the PI3K signaling pathway incancer. Curr Opin Genet Dev 2010;20:87–90.
[3] Li J, Yen C, Liaw D, Podsypanina K, Bose S, Wang SI, et al. PTEN, a putativeprotein tyrosine phosphatase gene mutated in human brain, breast, andprostate cancer. Science 1997;275:1943–7.
[4] Vlietstra RJ, van Alewijk DC, Hermans KG, van Steenbrugge GJ, Trapman J.Frequent inactivation of PTEN in prostate cancer cell lines and xenografts.Cancer Res 1998;58:2720–3.
[5] Mulholland DJ, Dedhar S, Wu H, Nelson CC. PTEN and GSK3beta. Key regulatorsof progression to androgen-independent prostate cancer. Oncogene 2006;25:329–37.
[6] Sarker D, Reid AH, Yap TA, de Bono JS. Targeting the PI3K/AKT pathway for thetreatment of prostate cancer. Clin Cancer Res 2009;15:4799–805.
[7] Chen LH, Fang J, Li H, Demark-Wahnefried W, Lin X. Enterolactone inducesapoptosis in human prostate carcinoma LNCaP cells via a mitochondrial-mediated, caspase-dependent pathway. Mol Cancer Ther 2007;6:2581–90.
[8] Peuhu E, Rivero-Muller A, Stykki H, Torvaldson E, Holmbom T, Eklund P, et al.Inhibition of Akt signaling by the lignan matairesinol sensitizes prostatecancer cells to TRAIL-induced apoptosis. Oncogene 2010;29:898–908.
[9] Dixon RA. Phytoestrogens. Annu Rev Plant Biol 2004;55:225–61.[10] Adlercreutz H. Lignans and human health. Crit Rev Clin Lab Sci 2007;44:483–525.[11] Buck K, Zaineddin AK, Vrieling A, Linseisen J, Chang-Claude J. Meta-analyses of
lignans and enterolignans in relation to breast cancer risk. Am J Clin Nutr2010;92:141–53.
[12] Li D, Yee JA, Thompson LU, Yan L. Dietary supplementation with secoisolar-iciresinol diglycoside (SDG) reduces experimental metastasis of melanomacells in mice. Cancer Lett 1999;142:91–6.
[13] Bylund A, Saarinen N, Zhang JX, Bergh A, Widmark A, Johansson A, et al.Anticancer effects of a plant lignan 7-hydroxymatairesinol on a prostatecancer model in vivo. Exp Biol Med (Maywood) 2005;230:217–23.
[14] McCann MJ, Gill CI, Linton T, Berrar D, McGlynn H, Rowland IR. Enterolactonerestricts the proliferation of the LNCaP human prostate cancer cell line in vitro.Mol Nutr Food Res 2008;52:567–80.
[15] Saarinen NM, Warri A, Dings RP, Airio M, Smeds AI, Makela S. Dietarylariciresinol attenuates mammary tumor growth and reduces blood vesseldensity in human MCF-7 breast cancer xenografts and carcinogen-inducedmammary tumors in rats. Int J Cancer 2008;123:1196–204.
[16] Lindahl G, Saarinen N, Abrahamsson A, Dabrosin C. Tamoxifen, flaxseed, andthe lignan enterolactone increase stroma- and cancer cell-derived IL-1Ra anddecrease tumor angiogenesis in estrogen-dependent breast cancer. Cancer Res2011;71:51–60.
[17] Raja SM, Chen S, Yue P, Acker TM, Lefkove B, Arbiser JL, et al. The naturalproduct honokiol preferentially inhibits cellular FLICE-inhibitory proteinand augments death receptor-induced apoptosis. Mol Cancer Ther 2008;7:2212–23.
[18] Hausott B, Greger H, Marian B. Naturally occurring lignans efficiently induceapoptosis in colorectal tumor cells. J Cancer Res Clin Oncol 2003;129:569–76.
[19] Yokoyama T, Okano M, Noshita T, Funayama S, Ohtsuki K. Characterization of(�)-matairesinol as a potent inhibitor of casein kinase I in vitro. Biol PharmBull 2003;26:371–4.
[20] Chen LH, Fang J, Sun Z, Li H, Wu Y, Demark-Wahnefried W, et al. Enterolactoneinhibits insulin-like growth factor-1 receptor signaling in human prostaticcarcinoma PC-3 cells. J Nutr 2009;139:653–9.
[21] Sharma PR, Shanmugavel M, Saxena AK, Qazi GN. Induction of apoptosis by asynergistic lignan composition from cedrus deodara in human cancer cells.Phytother Res 2008;22:1587–94.
[22] Yoo JH, Lee HJ, Kang K, Jho EH, Kim CY, Baturen D, et al. Lignans inhibit cellgrowth via regulation of Wnt/beta-catenin signaling. Food Chem Toxicol2010;48:2247–52.

E. Peuhu et al. / Biochemical Pharmacology 86 (2013) 571–583 583
[23] Nesterov A, Lu X, Johnson M, Miller GJ, Ivashchenko Y, Kraft AS. Elevated AKTactivity protects the prostate cancer cell line LNCaP from TRAIL-inducedapoptosis. J Biol Chem 2001;276:10767–74.
[24] Chen X, Thakkar H, Tyan F, Gim S, Robinson H, Lee C, et al. Constitutively activeAkt is an important regulator of TRAIL sensitivity in prostate cancer. Oncogene2001;20:6073–83.
[25] Gonzalvez F, Ashkenazi A. New insights into apoptosis signaling by Apo2L/TRAIL. Oncogene 2010;29:4752–65.
[26] Finnberg N, Klein-Szanto AJ, El-Deiry WS. TRAIL-R deficiency in mice promotessusceptibility to chronic inflammation and tumorigenesis. J Clin Invest2008;118:111–23.
[27] Grosse-Wilde A, Voloshanenko O, Bailey SL, Longton GM, Schaefer U, CsernokAI, et al. TRAIL-R deficiency in mice enhances lymph node metastasis withoutaffecting primary tumor development. J Clin Invest 2008;118:100–10.
[28] Falschlehner C, Schaefer U, Walczak H. Following TRAIL’s path in the immunesystem. Immunology 2009;127:145–54.
[29] Wiezorek J, Holland P, Graves J. Death receptor agonists as a targeted therapyfor cancer. Clin Cancer Res 2010;16:1701–8.
[30] Willfor S, Hemming J, Reunanen M, Eckerman C, Holmbom B. Lignans andlipophilic extractives in Norway spruce knots and stemwood. Holzforchung2003;57:359–72.
[31] Eklund P, Lindholm A, Mikkola JP, Smeds A, Lehtila R, Sjoholm R. Synthesis of(�) matairesinol, (�)-enterolactone and (�)-enterodiol from the naturallignan hydroxymatairesinol. Org Lett 2003;5:491–3.
[32] Eklund P, Sjoholm R. Oxidative transformation of the natural lignan hydro-xymatairesinol with 2,3-Dichloro-5,6-dicyano-1,4-benzoquinone. Tetrahe-dron 2003;59:4515–23.
[33] Eklund P, Sundell F, Smeds A, Sjoholm R. Reactions of the natural lignanhydroxy-matairesinol in basic and acidic nucleophilic media – formation andreactivity of a quinone methide intermediate. Org Biomol Chem 2004;2:1–8.
[34] Damon RE, Schlessinger RH, Blount JF. A short synthesis (�)-isostegane. J OrgChem 1979;41:3772–3.
[35] Eklund P, Sillanpaa R, Sjoholm R. Synthetic transformation of hydroxymatair-esinol from Norway spruce (Picea abies) to 7-hydroxysecoisolariciresinol, (+)-lariciresinol and (+)-cyclolariciresinol. J Chem Soc Perkin Trans1 2002;16:1906–10.
[36] Erdtman H. Konstitution der Harzphenole und deren biogenetische Zusam-menhange I. Uber Pinoresinol. Svensk Kemisk Tidsskrift 1934;46:229–33.
[37] Ekman R, Willfor S, Sjoholm R, Reunanen M, Maki J, Lehtila R, et al. Identifica-tion of the lignan northrachelogenin in knot and branch heartwood of Scotspine (Pinus sylvestris L). Holzforchung 2002;56:253–6.
[38] Khamlach K, Dhal R, Brown E. Lignans. 13. Total syntheses of (�)-carinol (�)-carissanol and (�)-hydroxythujaplicatin methyl ether. Heterocycles 1990;31:2195–9.
[39] Eklund P, Riska A, Sjoholm R. Synthesis of (�)-R-imperanene from the naturallignan hydroxymatairesinol. J Org Chem 2002;67:7544–6.
[40] Ummanni R, Jost E, Braig M, Lohmann F, Mundt F, Barett C, et al. Ubiquitincarboxyl-terminal hydrolase 1 (UCHL1) is a potential tumour suppressor inprostate cancer and is frequently silenced by promoter methylation. MolCancer 2011;10:129.
[41] Gan L, Chen S, Wang Y, Watahiki A, Bohrer L, et al. Inhibition of the androgenreceptor as a novel mechanism of taxol chemotherapy in prostate cancer.Cancer Res 2009;69:8386–94.
[42] Skjoth IH, Issinger OG. Profiling of signaling molecules in four different humanprostate carcinoma cell lines before and after induction of apoptosis. Int JOncol 2006;28:217–29.
[43] Hellawell GO, Brewster SF. Growth factors and their receptors in prostatecancer. BJU Int 2002;89:230–40.
[44] Belfiore A. The role of insulin receptor isoforms and hybrid insulin/IGF-Ireceptors in human cancer. Curr Pharm Des 2007;13:671–86.
[45] Torrance SJ, Hoffmann JJ, Cole JR. Wikstromol, antitumor lignan from wik-stroemia foetida var oahuensis gray and wikstroemia uva-ursi gray (thyme-laeaceae). J Pharm Sci 1979;68:664–5.
[46] Lee KH, Tagahara K, Suzuki H, Wu RY, Haruna M, Hall IH, et al. Antitumoragents 49 tricin, kaempferol-3-O-beta-D-glucopyranoside and (+)-nortrache-logenin, antileukemic principles from wikstroemia indica. J Nat Prod 1981;44:530–5.
[47] Saarinen NM, Penttinen PE, Smeds AI, Hurmerinta TT, Makela SI. Structuraldeterminants of plant lignans for growth of mammary tumors and hormonalresponses in vivo. J Steroid Biochem Mol Biol 2005;93:209–19.
[48] Datta SR, Brunet A, Greenberg ME. Cellular survival: a play in three Akts. GenesDev 1999;13:2905–27.
[49] Harikumar KB, Sung B, Tharakan ST, Pandey MK, Joy B, Guha S, et al. Sesaminmanifests chemopreventive effects through the suppression of NF-kappaB-regulated cell survival, proliferation, invasion, and angiogenic gene products.Mol Cancer Res 2010;8:751–61.
[50] Corsini E, Dell’Agli M, Facchi A, De Fabiani E, Lucchi L, Boraso MS, et al.Enterodiol and enterolactone modulate the immune response by acting onnuclear factor-kappaB (NF-kappaB) signaling. J Agric Food Chem 2010;58:6678–84.
[51] Cosentino M, Bombelli R, Carcano E, Luini A, Marino F, Crema F, et al.Immunomodulatory properties of achyrocline satureioides (lam.) D.C. infu-sion: a study on human leukocytes. J Ethnopharmacol 2008;116:501–7.
[52] Fizazi K, Sternberg CN, Fitzpatrick JM, Watson RW, Tabesh M. Role of targetedtherapy in the treatment of advanced prostate cancer. BJU Int 2010;105:748–67.
[53] Lee DH, Szczepanski MJ, Lee YJ. Magnolol induces apoptosis via inhibitingthe EGFR/PI3K/Akt signaling pathway in human prostate cancer cells. J CellBiochem 2009;106:1113–22.
[54] Jiang X, Chen S, Asara JM, Balk SP. Phosphoinositide 3-kinase pathwayactivation in phosphate and tensin homolog (PTEN)-deficient prostate can-cer cells is independent of receptor tyrosine kinases and mediated by thep110beta and p110delta catalytic subunits. J Biol Chem 2010;285:14980–89.
[55] Karasic TB, Hei TK, Ivanov VN. Disruption of IGF-1R signaling increases TRAIL-induced apoptosis: a new potential therapy for the treatment of melanoma.Exp Cell Res 2010;316:1994–7.
[56] Girnita A, All-Ericsson C, Economou MA, Astrom K, Axelson M, Seregard S, et al.The insulin-like growth factor-I receptor inhibitor picropodophyllin causestumor regression and attenuates mechanisms involved in invasion of uvealmelanoma cells. Clin Cancer Res 2006;12:1383–91.
Recommended

















