Embed Size (px)
Citation preview

Ovulation
IN THE NAME OF GOD
By: M, Hosseinzadaeh

Ovulation Ovulation is the occurrence in themenstrual cycle by which a selectedmature follicle breaks and releases a viableoocyte from the ovary.
Each month, one egg is released inhumans; but occasionally, two or more canerupt during the menstrual cycle.
In women with regular menstrual cycles,the ovulation takes place approximatelytwo weeks after menstruation.
Ovulation is triggered by the pituitary LHsurge, which initiates a series of ovarianevents, generating a cascade ofparacrine/autocrine factors, enzymes andtranscription factors responsible for therupture of the apical follicle wall,remodeling of extracellular matrix (ECM)and cell differentiation.

Ovulation is the result of a well ordered series of events.
Hypothalamus
Pituitary
Ovary
GnRH
LHFSH
Estradiol
The main factors responsible for the
function of this axis are:
•Firstly the hypothalamic hormones,
particularly the gonadotrophin
releasing hormone (GnRH),
•Secondly, the pituitary hormones or
gonadotropins (FSH and LH), and
•Finally, the ovarian steroid hormones
(estradiol and progesterone).
These processes are controlled by
the hypothalamic
-pituitary-ovary axis.
The menstrual cycle is directed
by complex functional
interactions between the ovaries
and the hypothalamus-pituitary
system which control each other
by means of positive or
negative feedback mechanisms.
perogestron

Hypothalamus
GnRH is a decapeptide which is synthesized and released by specific neuronal
endings in the nucleus arcuatus of the hypothalamus.
GnRH is transported through the portal vessels towards the anterior pituitary gland.
If GnRH is released in a constant, non-pulsatile manner, gonadotropin release is
suppressed due to an apparent desensitization of the pituitary GnRH receptors.
The hormone is only detectable in the
portal system, being undetectable in the
systemic circulation.
Small quantities of GnRH are sufficient to
effectuate a release of gonadotropins from
the pituitary gland.
The release pattern of the gonadotropins is
dictated by the frequency of GnRH release.
GnRH must be released in a pulsatile
manner, and its effects depend on the
frequency and amplitude of these pulses.

Effects of GnRH on Gonadotropins
GnRH acts through its receptor on the pituitary gonadotrophcells, stimulating production of phospholipase C.
IP3 pathway causes gonadotropin release, while the DAG/PKCpathway causes gonadotropin synthesis.

Pituitary gland
The last step of the hormone cascade is localized in the ovaries, where steroid
hormones are synthesized caused by the gonadotropins action. These hormones
are estradiol and progesterone .
Ovaries
The next step occurs in the pituitary gland. The varying frequency and amplitude of
GnRH release determines the pattern of release of the gonadotropins (FSH and LH)
during the menstrual cycle.

FSH release pattern during the cycle
In the last phase of the previous cycle, FSH level increases
due to the drop of estradiol, progesterone and inhibin levels.
This is called positive feedback.
The amount and timing of FSH release by the anterior pituitary changes throughout the ovulatory
cycle.
This rise in FSH concentration stimulates the growth of
antral follicles, resulting in an increase of estradiol and
inhibin B concentrations producing a negative feedback,
a reduction of FSH concentrations.
With the formation of the corpus luteum and the outpouring of both oestradiol and progesterone,
the negative feedback mechanism comes into play and continues its suppression of FSH release
until just before the next menstruation.
At mid-cycle, there is a temporary increase in FSH
secretion. It may be due to the GnRH surge and may
have a function in preparing a cohort of small antral
follicles for the next cycle.

Role of FSH
Granulosa cell proliferation and differentiation
Antral follicle development
Estrogen production
Inducing of LH receptor in dominant follicle
Inhibin synthesis
The major role of the FSH surge is to stimulate cumulus expansion and to facilitate the production of the protease
plasmin.

CUMULUS CELLS: EXPANSION
the cumulus granulosa cells undergo a series of structural and functional
changes called mucification.
In response to the preovulatory surge of FSH, the cumulus cells secrete
large quantities of a newly synthesized glycoprotein mucous
substance into the extracellular spaces.
This change results in the dispersal of the cumulus cells and causes the
egg-cumulus complex to expand tremendously.
The process of mucification is physiologically critical for the pickup and
transport of the egg in the fallopian tube.
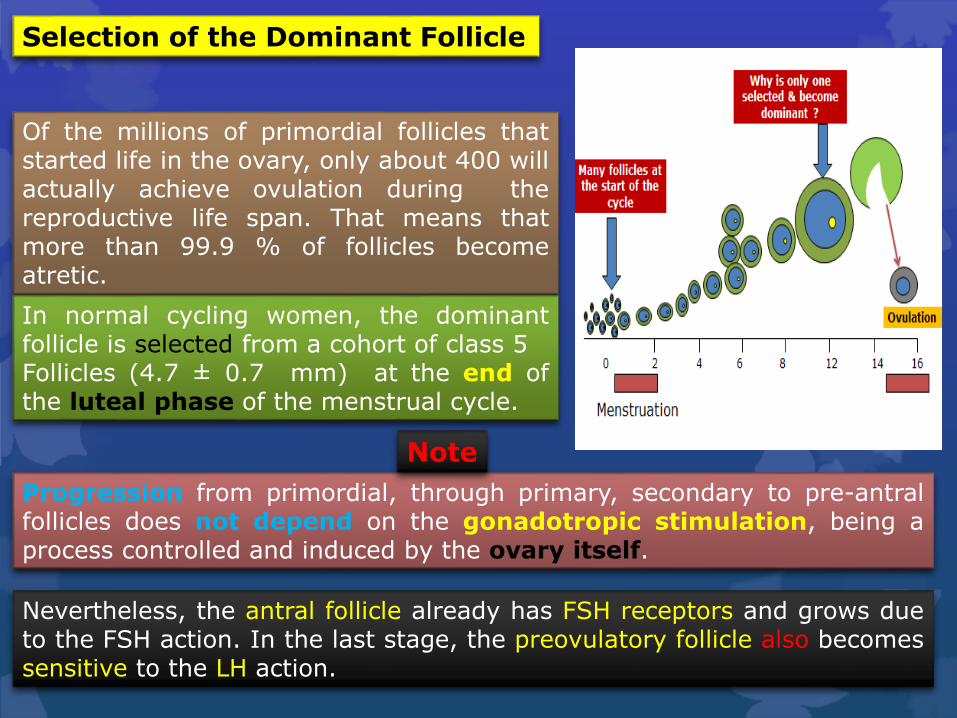
Selection of the Dominant Follicle
Progression from primordial, through primary, secondary to pre-antralfollicles does not depend on the gonadotropic stimulation, being aprocess controlled and induced by the ovary itself.
Of the millions of primordial follicles thatstarted life in the ovary, only about 400 willactually achieve ovulation during thereproductive life span. That means thatmore than 99.9 % of follicles becomeatretic.
Nevertheless, the antral follicle already has FSH receptors and grows dueto the FSH action. In the last stage, the preovulatory follicle also becomessensitive to the LH action.
In normal cycling women, the dominantfollicle is selected from a cohort of class 5Follicles (4.7 ± 0.7 mm) at the end ofthe luteal phase of the menstrual cycle.
Note

At the time of selection, each cohort follicle contains a fully grown oocyte,
about 1 million granulosa cells, a theca interna containing several layers of
TICs, and theca externa composed of smooth muscle cells
A characteristic feature of a dominant follicle is a
high rate of mitosis in the granulosa cells.
As a consequence of increased mitosis, the
dominant follicle continues to grow rapidly
during the follicular phase, reaching 6.9 ± 0.5
mm at days 1 to 5, 13.7 ± 1.2 mm at days 6 to
10, and 18.8 ± 0.5 mm at days 11 to 14.
The key to being chosen as the month’s ovulatory follicle is sensitivity to FSH.
Decreased estradiol production by the corpus luteum is the principal cause for the secondary rise in FSH

As FSH concentrations fall in response torising oestrogen and inhibin B levels andbecome less available, only the mostsensitive follicle to FSH, that with thelowest threshold for a response to FSH,can survive and continue to thrive andproduce the most oestrogen and LHreceptors.
Selection of the dominant follicle in relation to FSH concentrations
The follicles more sensitive to FSHrather than those less mature areselected.The FSH level declines during the midfollicular phase.
The follicles most sensitive to FSH will utilize it to increase aromatase activity and produce oestrogens and inhibin.
The rest, starved of the possibility of FSH stimulation, become atretic.

A. In dominant follicles, FSH in follicular fluid induces P450arom activity that metabolizes androgen substrate to estradiol (E2 ). In such follicles, E2 and androstenedione (A4) accumulate in very high concentrations in the follicular fluid.
B. In nondominant follicles, the low levels of FSH lead to a paucity of granulosa cells (GC) and low concentrations of estradiol, despite the high levels of A4.
In developing healthy (dominant) follicles (class 5 to 8 follicles), the meanconcentration of follicular fluid FSH increases from about 1.3 mIU/ml(about 58 ng/ml) to about 3.2 mIU/ml (about 143 ng/ml) through thefollicular phase.In contrast, the levels of FSH are low or undetectable in themicroenvironment of the nondominant cohort follicles
Dominant follicles have a more vascular theca compared with otherantral follicles, and as a result they display an increased uptake of serumgonadotropins.

LH release pattern during the cycle
During the early and mid-follicular phase, the secretion of LH isrelatively quiet with pulses every 60–90 min and a fairly constant lowconcentration of circulating LH.However, this is the calm before the storm.An enormous climax is reached with the onset of the LH surge in the late follicular phase, the central event of the ovulatory cycle. Concentrations of LH rise to 10–20 times their resting level during the rest of the cycle.

Then, the pituitary gland becomes highly sensitive to GnRH stimulation, due to the
increase of GnRH receptors. Thus, the GnRH surge produces the LH surge.
The causes of the LH surge are:
First, the negative feedback of estradiol at the hypothalamic-pituitary level turns to a
positive feedback when estradiol concentrations reach a critical point.
Following ovulation, increasing concentrations of progesterone slow down the
frequency of LH releasing pulses.
Concentrations of LH once again drop
to baseline levels.

The pre-ovulatory LH surge has a number of key functions:
Triggers of ovulation and follicular rupture about 36 hours after the surge
Disruption of the cumulus oocyte complex
Luteinization of granulosa cells
The major role of the LH surge is to stimulate meiotic maturation
formation of the stigma or site of follicle rupture

The mid-cycle LH surge induces a dismantling of the gap junctionsbetween granulosa cells and the oocyte, thus inhibiting the flow of inhibinfactors to the oocyte and allowing the flow of maturation inducing factorssuch as calcium, maturation promotor factor (MPF) and another growthfactors.
cAMP is an important mediator in the nuclear maturation of the oocyte.cAMP activates a protein-kinase that suppress the activation of MPF ordegrades its subunits. The drop of cAMP levels after LH surge favoursoocyte maturation due to the increasing levels of MPF.
Increasing levels of calcium after the LH surge are also necessary torestore the meiotic arrest.
After oocyte maturation begins andcAMP levels decline, RIIα-PKAmoves to the mitochondria, andmaturation promoting factor (MPF)is activated, leading to germinalvesicle breakdown and maturationto the MII stage.
LH surge is to stimulate meiotic maturation
Meiosis is arrested here and proceeds no further unless the ovulated egg is fertilized

LH
androgens
estradiol
Theca cells
Granulosa cells
aromatase
FSH
“two cell – two gonadotropin hypothesis”
The mid-cycle surge of LH induces the production of androgens by cells of thetheca follicles. The androgens, androstenedione and testosterone, are then ‘passedon’ to the neighbouring granulosa cells, where aromatase (CYP19) converts theminto estrogens, mainly estradiol but also estrone.
These events suggest that the function of theca cells and granulosa cells are controlled by LH and FSH respectively.Moreover, there is also a fine-tuning achieved by other factors such asinhibin, insulin-like growth factors (IGFs) I and II and tumor necrosisfactor-α.
Aromatase activityand therefore,estrogen productionis controlled by FSH.

Oestradiol
Estradiol is the mostimportant estrogen in theovulatory cycle. Duringmenstruation estradiolconcentrations are low, butstart to increase as FSHinduces folliculardevelopment in the mid-follicular phase.
When estradiol levels reach a critical point, they activate a positivefeedback mechanism in the hypothalamus and pituitary resulting in LHand FSH surges.Following ovulation, estradiol concentrations decrease temporarily but arerevived caused by corpus luteum activity. With the demise of the corpusluteum, estradiol concentrations drop rapidly to their lowest levels and bya positive feedback, increase FSH levels immediately precedingmenstruation.

The key functions of estradiol in the ovulatory cycle are:
In the mid-late follicular phase
It suppresses the secretion of FSH due to a negative feedback mechanism leading to the selection of a dominant follicle and preventing multi follicular development.
In mid-cycle
Triggers the LH surge due to a positive feedback mechanism when its concentrations rise to a critical level.
In the follicular phase
As a ‘growth hormone’ for the development of the endometrium.
In the ovulatory period
estradiol stimulates the glands of the cervix to secrete a particular type of mucus which is essential for the sperm to pass through the cervix to reach the ovum.

Progesterone
Progesterone is the main hormone in the luteal phase.
Large quantities are synthesized by luteinized granulosa cells of the corpus
luteum following ovulation.
Progesterone concentrations rise to a peak 7-8 days following ovulation and
fall rapidly with the demise of the corpus luteum.

Together with estradiol,
progesterone suppresses
pituitary gonadotropin
release during the luteal
phase.
The initial rise of progesterone concentrations immediately preceding the
LH surge may play a role In the triggering of this surge.
Progesterone
During this phase, FSH is synthesized and storedready for release when freed from the inhibition imposed by progesteroneand oestradiol when the Corpus luteum fails

Role of progestrone
The main functions of progesterone secreted by the corpus luteum are To
induce a secretory endometrium, capable to enhance embryo
implantation.
Progesterone also plays a role in the expression of genes needed for
implantation at the level of the endometrium.
stimulates mammary growth
Progesterone is responsible for the stimulation of the theca collagenase,
an enzyme which stimulates the dissociation of cells in the area of the
stigma.

Activin is a promoter of many actions ofFSH in that it increases FSH secretion,promotes ovarian follicular development.
Inhibin
Inhibin is secreted by
granulosa cells
Inhibin A
Inhibin B
Inhibin A concentrations are low duringmost of the follicular phase but start torise during its latest stages and peak inthe mid-luteal phase.
inhibin B concentrations start rising earlyin the follicular phase, paralleling butlater than the FSH rise. reduces thesynthesis and secretion of FSH and thenumber of GnRH receptors in the pituitaryand has an inhibitory effect on the growthof antral follicles in the ovary.
Activin and follistatin
Follistatin is an activin-binding proteinthat neutralizes activin bioactivity.
Estrogens and inhibin B are both inhibitory factors for the
secretion of FSH.

Growth factors
insulin-like growth factors (IGFs) I and II which are very active.Insulin, as well as binding to IGF receptors, has its own ovarian receptorsand is known to promote androgen production.The transforming growth factor (TGF) family is also well represented in theovary as is epithelial growth factor (EGF). All play a passive role in theregulation of gonadotropin activity within the follicles.
Anti-Mullerian hormone (AMH)
AMH, a dimeric glycoprotein and member of the transforming growth factor-beta family, is produced by ovarian follicular granulosa cells in late pre-antral and small antral follicles. It seems to have a role in the regulation of folliculogenesis at the two extremes of this process:
(a) by restricting the progression of development of primordial
follicles
(b) by an inhibition of the sensitivity of antral follicles to FSH and
inhibition of aromatase activity during an ovulatory cycle.

Mechanism of follicular rupture
On or about the 15th day of an ideal 28-daycycle, the preovulatory follicle ruptures, andthe egg cumulus complex is released from theovary by a process called ovulation.
The preovulatory surges of LH and FSH play acrucial role in the physiologic mechanism ofovulation.
A stigma in mammalian reproductiveanatomy refers to the area of the ovariansurface where the Graafian follicle will burstthrough during ovulation and release theovum
Stigma formation involves a combination of cell apoptosis, cell migration,and proteolytic digestion of extracellular matrix layers

In response to the LH surge, the preovulatory follicle producesprogesterone and prostaglandin, both of which are obligatory for thestigma to develop.
PGF2a stimulates the rupture of lysosomes in the apex epitheliumfacilitating further stigma formation; it also stimulates ovarian contractionsleading to follicle rupture and; finally stimulates follicle contraction foractual oocyte expulsion
After the surge in luteinizing hormone prior toovulation, vasodilation of capillaries is prominent andthe cross-sectional areas of vasculature luminaincrease. There is evidence of increased vascularpermeability, tissue oedema and ischemia. Capillariesdevelop perforations through which blood cells andplatelets escape when ovulation occurs. Shortlybefore ovulation, blood flow stops in a small area ofthe ovarian surface overlying the bulging follicle.
Prostaglandins production by granulosa cells. Two types of prostaglandins are known to have some role in the process of ovulation.
PGE2, which stimulates the plasminogen activator, an enzyme which dissolves connective tissue in the area of the stigma.

FSH, at very low levels, stimulates the production of plasminogen activator (PA) by the granulosa cells.
1
PA catalyzes the conversion ofplasminogen from thegranulosa cells to plasmin
3
that targets the cells of the follicular wall, weakening their intercellular adhesion.
5
Plasmin is a serine protease derived from plasminogen by enzymaticactivation. Two forms of plasminogen activators have been characterized,urokinase (uPA) and tissue (tPA) types. Both uPA and tPA appear tocontribute to ovary plasmin biosynthesis and ovulation. The follicular fluidcontains relatively high levels of the plasmin precursor, plasminogen. Thepreovulatory surge of FSH appears to stimulate granulosa cells to secreteplasminogen activator, which converts plasminogen to the active proteaseplasmin. Plasmin appears to play a role in the degradation of thegranulosa cells and basal lamina in the presumptive stigma.

FSH also induces the huge expansion of the cumulus oophorus (by mucification) which also includes up regulation of LH receptors on the granulosa cells’ surfaces prior to the LHsurge.
2
The only targets of FSH in the ovary are the granulosa cells
Luteinizing hormone (LH)induces the resumption ofmeiosis (by the mid-cyclesurge)
LH also induces the folliclecorona cells to make andrelease progesterone thatinduces prostaglandins(PG’s) E and F production.
6
7These PG’s trigger the presumptive stigma cells of the ovarian wall to make and release a number of proteases that break down the ovarian matrix and the follicular tunica albuginea
8
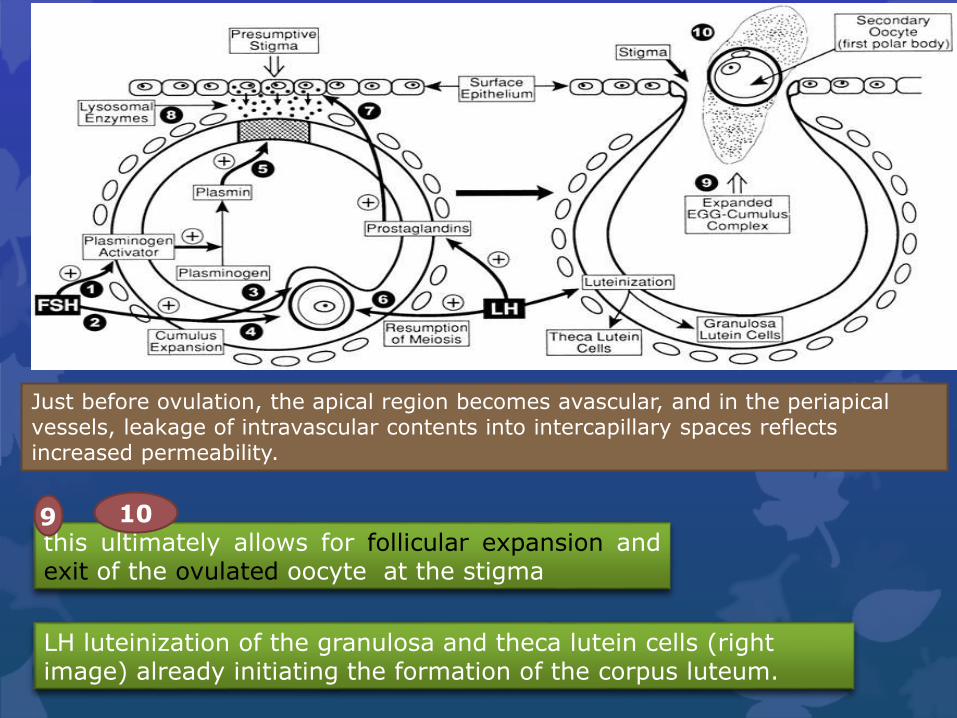
this ultimately allows for follicular expansion andexit of the ovulated oocyte at the stigma
9 10
LH luteinization of the granulosa and theca lutein cells (right image) already initiating the formation of the corpus luteum.
Just before ovulation, the apical region becomes avascular, and in the periapical vessels, leakage of intravascular contents into intercapillary spaces reflects increased permeability.

Scanning electron micrograph of the protruding surface of an ovulatory follicle.

NGF/TrkA interactions lead to a loss of intercellular communication by disrupting gap junctions between theca cells and result in increasedmigratory behavior.
Nerve growth factor (NGF) and one of its receptors, the tyrosine kinase receptor TrkA
NGF and TrkA expression are induced in theca cells in response to the LH surge.
Vascular endothelial growth factor (VEGF) and its downstream signaling pathways are required for follicle angiogenesis during antral follicle growth, and there is evidence in the primate that VEGF is important for follicle rupture.
VEGF
VEGF promotes vascular permeability, allowing more efficient delivery of bloodborne Factors including LH and FSH, and immune
cells to the follicle.
α-Adrenergic agents, such asnorepinephrine, exert astimulatory effect on ovariancontractions, whereas β-adrenergic agents are inhibitory.

Enzymes such as matrix metalloproteinases (MMPs) and ADAMTS
proteases are involved (although not all are essential):
Tissue inhibitors of MMPs (TIMPs) are also involved in the ovulatory
process and the pattern of in vivo expression suggests that decreased
TIMPs levels and increased MMPs are involved in follicle rupture.
The extracellular matrix remodeling is completed by a process similar to
the inflammatory process, with the participation of macrophages,
neutrophils, cytokines produced by leukocytes, platelet activating factor
and free radicals.
Degrading of the basement membrane
Remodeling of the extracellular matrix
Rupturing of the follicular apex.
ovulatory induces with mediators including endothelin-2, interleukin-6and cGMP-dependent protein kinase II.

RelaxinTheca interna constitutes the primary source of follicular relaxin
Oxytocin synthesized by the ovary
Arachidonic acid
play an essential role in gonadotrophin activation of ovarian collagenolysis, needed for follicle rupture at ovulation.
This is supported by the fact that inhibitors of cyclooxygenase and lipoxygenasepathways of arachidonic acid metabolism inhibit follicle rupture at ovulation.
large amounts of sex steroids present in the follicleduring the early preovulatory phase can stimulategrowth of lysosomes
sex steroids
Other substances that appear to influence the process of ovulation at a local ovarian level include various cytokines, oxygen free-radicals, nitric oxide and angiotensin II.


1-5 Minutes before
Follicular Rupture
½-1 Hour before
Follicular Rupture
10 Hours before Follicular
RuptureAnatomy of Ovulation

Ovulation is initiated by the surge of LH (Step 1) that induces the expression of
specific inflammatory-related genes in granulosa cells, which then leads to an
intrafollicular signaling cascade and the expression of similar genes in cumulus cells:
Areg, Ptgs2, Has2 and Tnfaip6. Once activated in COCs, these genes establish an
autocrine regulatory loop whereby PGE regulates Areg expression and AREG
regulates Ptgs2 mRNA; together they induce genes that control the production and
stabilization of HA-rich matrix (Step 2). As expansion proceeds, genes associated
with innate immune cell responses, such as Il6, are induced (Step 3) and continue to
be expressed in ovulated COCs.
hypothesize that
ovulation
involves
coordinated and
sequential
inflammatory
and innate
immune
responses

FSH binds FSH receptors on cumulus cells to activate AC, cAMP production and PKA.
This pathway activates Erk as do Egf receptors in response to Egf-Lamphiregulin (AR), betacellulin (BC) and epiregulin (ER).
Transcription factors regulated by thesekinase cascades include AP-1 factors,Elk-1 and cAMP response element-binding protein (CREB).
In growing follicles, cAMP istranslocated to the oocyte via gapjunctions until these are inactivatedthrough phosphorylation by Erk andcells separate from the oocyte throughcumulus expansion.
Signal transduction in cumulus cells in the mouse periovulatory phase.
Consequently, cAMP levels in the oocyte fall.

In a separate pathway, oocyte-secreted growth and differentiation factor-9 (Gdf-9) and bone morphogenetic protein-15 (BMP-15) activate integralreceptor kinase activity on cumulus cells, which results in phosphorylationof SMAD2/3, which translocate to the nucleus in dimers with SMAD4.
These transcription factorspromote expression of keycumulus genes required forspecification of the cumulus-specific response to theovulatory surge.
PGE2 activates the EP2 receptor to mediate a signal transduction pathway similar to FSH, and RGS-2 may control the activation of G-proteins.
Genes induced via these twosignal transductionmechanisms include HAS-2,TSG-6, PTX-3, COX-2, PGE2receptor (EP2) and regulatorof GPCR-2 (RGS-2).

Transcriptional complexes modify chromatin structure and stabilizetranscriptional machinery to transiently activate high gene expression.Genes expressed in this rapid transient fashion via this signalling cascadeinclude PR, a disintegrin and metalloproteinase with thrombospondinrepeats (ADAMTS-1), Egr-1 cathepsin L and versican.
Signal transduction in mural granulosa cells in the mouse periovulatory phase.
LH interacts with its G-protein (Gas)-coupled receptor activating AC andhence intracellular cAMP productionas well as iP3/Ca Þþ mobilization.
Consequently, PKA and Erk activationresult in phosphorylation (linearrows) of transcription factorsincluding cAMP response element-binding protein (CREB) andstimulatory proteins 1 and 3 (Sp1/3).
Transcriptional complexes assembled on the promoters of periovulatorygenes expressed in mural granulosa cells include Sp1/3 and additional factors including CREB, reproductive homeobox 5 (Rhox5) or other induced transcription factors.

Clinical Signs and Symptoms of Ovulation
Many women refer heightened sense of smell and sexual desire in the several days immediately before ovulation.
Towards the time of ovulation the pH in the vagina becomes less acidic
The cervical mucus becomes more copious and less viscous, all of which favour the progress of motile sperm towards the released oocyte.
Moreover, the action of progesterone increases basal body temperature by one-quarter to one-half degree Celsius (one-half to one
degree Fahrenheit).
Many women experience secondary fertility signs including Mittelschmerz(pain associated with ovulation).

Thanks for your attention





























