Embed Size (px)
Citation preview
Tetanus Toxin Mechanism
Dariyus Z Kabraji
MSc: 1 Roll No: 18
Department of Biotechnology
Bacteriology Assignment

Tetanus
• Tetanus is one of the most basic fatal diseases in existence.• It is caused by infection of the victim with Clostridium tetani.• Also known as lockjaw, it is characterised by muscular spasms.• In the most common type the spasms begin in the jaw and
then progresses to the rest of the body. These spasms usually last a few minutes each time and occur frequently for three to four weeks.
• Symptoms include bone fractures, fever, headache, trouble swallowing, high blood pressure, and a fast heart rate.
• The incubation period is approximately eight days. In theory, the farther the injury site is from the central nervous system, the longer the incubation period, and the less severe are the symptoms experienced.

Statistics
• Clostridium tetani is considered to be the deadliest bacteriological pathogen in existence, second only to Clostridium botulinum.
• The more frequent cases like neonatal tetanus resulted in over 59000 infant mortalities in 2008 alone.
• The United States alone reports over 30 cases of infection each year.
• Lack of a proper diagnosis test makes Tetanus all the more potent a killer.
• Alongside the neurotoxin, C. tetani also produces the exotoxin tetanolysin, a hemolysin, that causes destruction of tissues.

Mechanism of Infection
• C. tetani usually enter the body through an open wound, leading to spore germination under anaerobic conditions.
• Once spore germination has occurred, toxins are released into the bloodstream and lymphatic system. These toxins act at several locations within the central nervous system, interfering with neurotransmitter release and blocking inhibitor impulses. Such disruptions lead to uncontrollable muscle contractions.
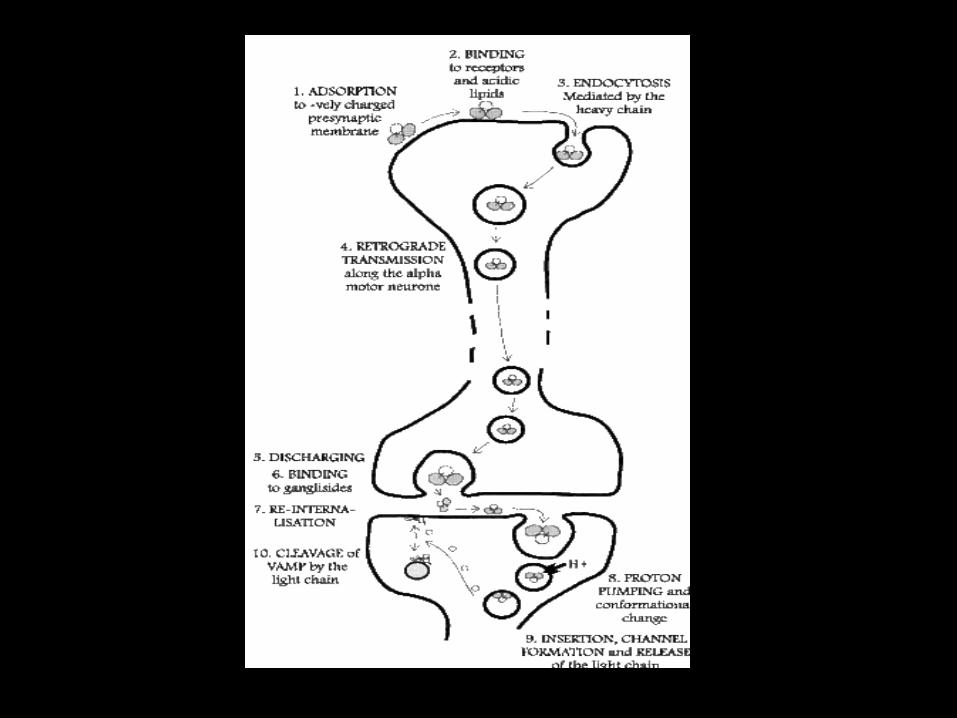

The Neurotoxin
• TeNT is the toxin complex affecting the nervous system of the victim.
• It is also known as Tetanospasmin or as the Spasmogenic toxin.
• It’s course of action involves:
1. Specific binding in the periphery neurons
2. Retrograde axonal transport to the central nervous system (CNS) inhibitory interneurons (Movement toward the cell body is called retrograde transport and movement toward the synapse is called anterograde transport)
3. Transcytosis from the axon into the inhibitory interneurons
4. Temperature and pH mediated translocation of the light chain into the cytosol
5. Reduction of the disulphide bond between the light and heavy chain
6. Cleavage of synaptobrevin / VAMP (Vesicle associated membrane proteins (VAMP) are a family of SNARE proteins with similar structure, and are mostly involved in vesicle fusion.
7. The best studied SNAREs are those that mediate docking of synaptic vesicles with the presynaptic membrane in neurons. These SNAREs are the targets of the bacterial neurotoxins responsible for botulism and tetanus.)

Clathrin Dependant Mechanism
• Neurons have adapted their endocytic pathways to better adjust to their specific requirements. Thus, synaptic vesicle (SV) recycling is the predominant form of neuronal endocytosis at the presynaptic terminal, whereby the fast fusion of neurotransmitter-containing vesicles is coordinated with an efficient mechanism of membrane recovery, which involves clathrin.
• In neurons, clathrin-independent routes have also been documented, although the physiological relevance of endocytosis via caveolae has been questioned in these cells because several of the caveolin isoforms found in other tissues are not detectable.

Course of Action
• Tetanus toxin is composed of a heavy chain and light chain, which are attached by a disulphide bond.
• Tetanus toxin fragment C (TeNT-FC) is a 47-kDa fragment on the heavy chain molecule that contains the ganglioside-binding domain.
• TTFC attaches to gangliosides on the peripheral nerves, and as a result, the toxin is internalized. Through trans-synaptic spread, the toxin can spread to the central nervous system.
• The light chain contains a zinc metalloprotease domain which can cleave proteins that facilitate synaptic vessel fusion with the plasma membrane of the neuron – namely, the integral protein synaptobrevin. As a result, the neurotransmitter g-aminobutyric acid (GABA) is blocked from reaching the synaptic cleft, and the excitation of motor neurons persists. Persistent neuron signalling leads to the motor spasms seen in a typical tetanus patient
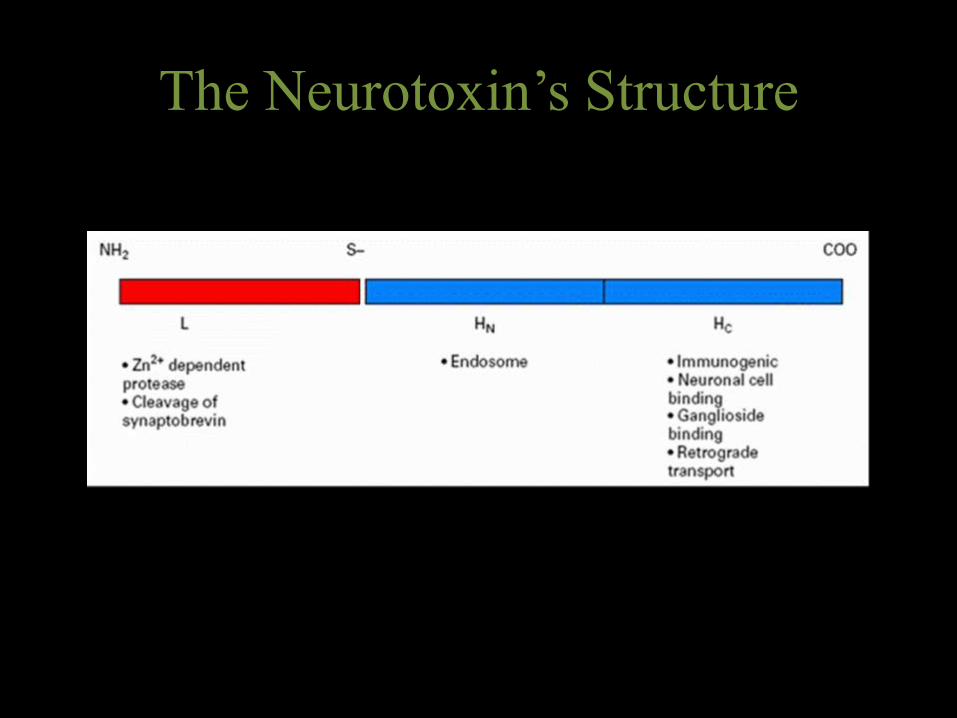
The Neurotoxin’s Structure

Abstract
• Ligand–receptor complexes are internalized by a variety of endocytic mechanisms. Some are initiated within clathrin-coated membranes, whereas others involve lipid microdomains of the plasma membrane. In neurons, where alternative targeting to short- or long-range trafficking routes underpins the differential processing of synaptic vesicle components and neurotrophinreceptors, the mechanism giving access to the axonal retrograde pathway remains unknown. To investigate this sorting process, we examined the internalization of a tetanus neurotoxin fragment (TeNT HC), which shares axonal carriers with neurotrophins and their receptors.
• Previous studies have shown that the TeNT HC receptor, which comprises polysialo-gangliosides, resides in lipid microdomains. We demonstrate that TeNTHC internalization also relies on a specialized clathrin-mediated pathway, which is independent of synaptic vesicle recycling. Moreover, unlike transferrin uptake, this AP-2–dependent process is independent of epsin1. These findings identify a pathway for TeNT, beginning with the binding to a lipid raft component (GD1b) and followed by dissociation from GD1b as the toxin internalizes via a clathrin-mediated mechanism using a specific subset of adaptor proteins.

Introduction
• Endocytosis is essential for a variety of cellular functions, including the internalization of nutrients and communication among cells, or between cells and their environment.
• Internalized molecules must be precisely sorted to their final cellular destinations to fulfil their specific function.
• Distinct endocytic pathways have been described to date, including clathrin- dependent endocytosis and caveolae-mediated uptake, which remain the two best-characterized mechanisms of internalization.
• Transport vesicles bud off as coated vesicles, which have a distinctive cage of proteins covering their cytosolic surface. Before the vesicles fuse with a target membrane, they discard their coat, as is required for the two cytosolic membrane surfaces to interact directly and fuse. The coat performs two main functions. First, it concentrates specific membrane proteins in a specialized patch, which then gives rise to the vesicle membrane. In this way, it selects the appropriate molecules for transport. Second, the coat moulds the forming vesicle. Coat proteins assemble into a curved, basketlike lattice that deforms the membrane patch and thereby shapes the vesicle. This may explain why vesicles with the same type of coat often have a relatively uniform size and shape.
• Clathrin-coated vesicles, for example, mediate transport from the Golgi apparatus and from the plasma membrane, whereas COPI- and COPII-coated vesicles most commonly mediate transport from the ER and from the Golgi cisternae

Materials
• Reagents: Sulfo-NHS-SS-biotin, EZ-link–activated maleimide-HRP (HRP is horseradish peroxidase, an amplifier that’s used by coupling it with TeNT)
• Plasmids: encoding dynamin k44A, epsin 1R63L H73L, and AP180 C-terminal mutants
• Antibodies: 9E10, X22, 12CA5, 69.1, epsin1, IgG3 mouse monoclonal antibody MOG1
• Neurotoxin: TeNT HC • Miscellaneous: Edetic Acid (EDTA), PBS, ConA–
Sepharose, α-methylmannoside, NiNTA-agarose, 20 mM Hepes-NaOH, 150 mM NaCl, and 500 mMimidazole.

Methods
• Protein Labelling
• Microinjection and Internalization assay
• Immunofluorescence
• Confocal microscopy

Observation
• TeNT HC enters clathrin-coated structures in MNs
• MNs were incubated with HRP–TeNT HC for 45 min on ice, and then chased for 45 min at 12 or 18°C

Observation
• A&b =12 degrees
• C= 18 degrees
• TeNT HC internalization in MNs is independent of presynaptic activity
• TeNT HC uptake is dynamin-dependent
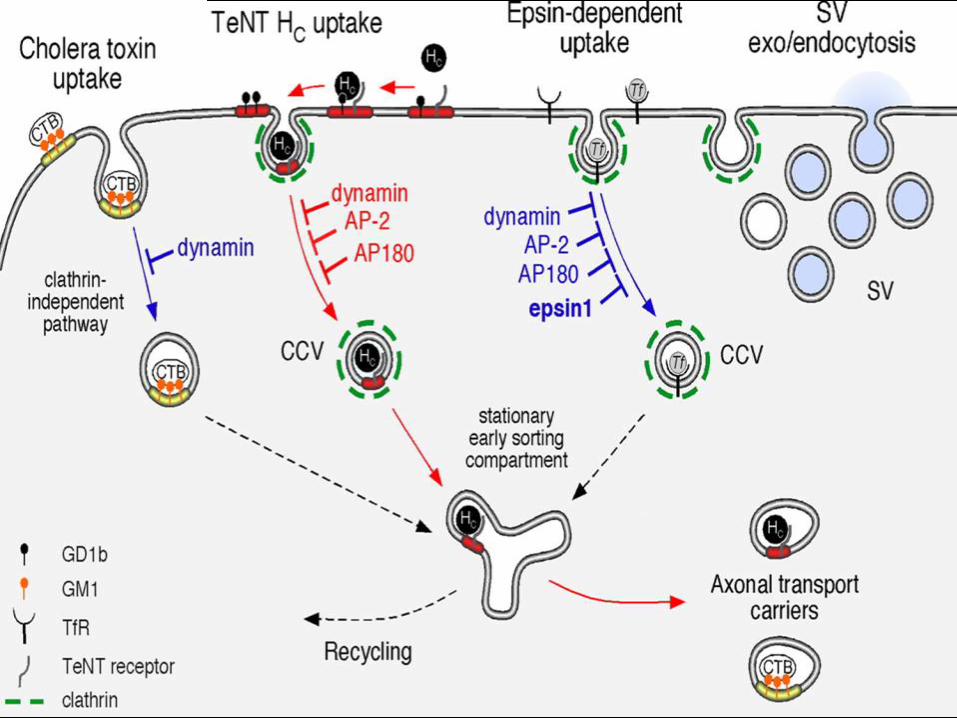

• Transferrin uptake is mediated by a classical clathrin-dependent internalization route occurring in soma and dendrites.
• TeNT HC exploits a pathway requiring lipid rafts and the clathrin machinery, which is distinct from aforementioned routes of internalization.
• At the NMJ, TeNT HC binds to a lipid–protein receptor complex containing the ganglioside GD1b. TeNT HC is then laterally sorted into CCPs and, during this sorting event, GD1b is excluded from the toxin receptor c omplex.
• Internalization of TeNT HC is dependent on dynamin, AP-2, and AP180, but does not require epsin1. Once internalized, TeNT HC is targeted to a stationary early sorting compartment (L akadamyaliet al., 2006), to which other endocytic routes may converge. This early sorting compartment is functionally coupled to the axonal retrograde transport pathway.

References
• Katrin Deinhardt, Otto Berninghausen, Hugh J. Willison, Colin R. Hopkins, and Giampietro Schiavo; Tetanus toxin is internalized by a sequential clathrin-dependent mechanism initiated within lipid microdomains and independent of epsin1; The Journal of Cell Biology, Vol. 174, No. 3, July 31, 2006 459–471
• Farrar JJ, LM Yen, T Cook, N Fairweather, N Binh, J Parry, CM Parry. 2000; Neurological Aspects of Tropical Disease: Tetanus. Journal of Neurology, Neurosurgery, and Psychiatry; 69: 292-301.
• Albert’s Molecular Biology of the Cell, 5th edition, Chapter 13: Intracellular vesicular traffic; 749-812





























