Embed Size (px)
Citation preview
LS
ELSEVIER
TssueSiCellVolume 48 Issue 2 April 2016 Pages 81-144 ISSN 0040-8166
vj
&&ÿ
nM.
mik s§m
H*
&3& IsV
Xu
k
-1
.
iwjP*
. ...
A %<&,
. ' 231t
.V

Tt
HA
a
ARRAA
KCHSLT
1
rwc2
otd
r(It(c
AS
h0
Tissue and Cell 48 (2016) 81–88
Contents lists available at ScienceDirect
Tissue and Cell
j o ur nal ho mepage: www.elsev ier .com/ locate / t i ce
he role of curcumin in streptozotocin-induced hepatic damage andhe trans-differentiation of hepatic stellate cells
esham N. Mustafa ∗
natomy Department, Faculty of Medicine, King Abdulaziz University, Jeddah, Saudi Arabia
r t i c l e i n f o
rticle history:eceived 5 December 2015eceived in revised form 19 January 2016ccepted 7 February 2016vailable online 9 February 2016
eywords:urcuminepatic stellate cellstreptozotocin
a b s t r a c t
Diabetic patients frequently suffer from non-alcoholic steatohepatitis. The current study aimed to inves-tigate the role of curcumin and the response of hepatic stellate cells in streptozotocin (STZ)-inducedhepatic damage. Sixty male rats were divided into three groups. The normal control injected with acitrate buffer vehicle and the diabetic control group which was injected intraperitoneally (IP) with asingle-dose of streptozotocin (50 mg/kg body weight) and a diabetic group was treated with an oraldose of curcumin at 80 mg/kg body weight daily for 60 days. Curcumin effectively counteracts oxidativestress-mediated hepatic damage and improves biochemical parameters. Alpha-smooth muscle actin (�-SMA) was significantly reduced, and insulin antibodies showed strong positive immunoreactivity withcurcumin administration. These results optimistically demonstrate the potential use of curcumin, which
ipid peroxidationrans-differentiation
is attributed to its antiradical/antioxidant activities and its potential �-cell regenerative properties. Also,it has the capability to encourage the trans-differentiation of hepatic stellate cells into insulin-producingcells for a period of time. In addition, as it is an anti-fibrotic mediator that inhibits hepatic stellatecell activation and the transition to myofibroblast-like cells, this suggests the possibility of considering
utic e
curcumin’s novel therape. Introduction
Diabetic patients frequently suffer from a hepatic impairmentecognized as non-alcoholic steatohepatitis which is associatedith severe complications such as deposition of glycogen, steatosis,
irrhosis, fibrosis and occasionally hepatic cancer (Bugianesi et al.,007).
Streptozotocin (STZ) is an N-nitroso-N-methylurea derivativef 2-deoxy-d-glucose which is a diabetogenic agent that damageshe islet �-cells in the pancreas selectively to produce insulin-ependent diabetes mellitus (IDDM) (Yang et al., 2010).
Alpha-smooth muscle actin (�-SMA) is an indicator for theecognition of myofibroblast-like cells plus hepatic stellate cellsHSCs) which are also known as Ito cells (Clement et al., 2010).
n diabetes, the glucose accessibility is increased and this leadso an accelerated formation of advanced glycation end productsAGEs). AGEs interact with the receptor for AGEs (RAGEs) andonsequently increase oxidative stress and cellular growth. This∗ Corresponding author at: King Abdulaziz University, Faculty of Medicine,natomy Department, Ground Floor Building No. 8, P.O. Box: 80205, Jeddah 21589,audi Arabia. Tel.: +966 566 764 762.
E-mail address: [email protected]
ttp://dx.doi.org/10.1016/j.tice.2016.02.003040-8166/© 2016 Elsevier Ltd. All rights reserved.
ffects in reducing hepatic dysfunction in diabetic patients.© 2016 Elsevier Ltd. All rights reserved.
leads to an increased proliferation of HSCs, which is noticed duringhepatic fibrogenesis that is accompanied by the up-regulation ofRAGEs.
Oxidative stress plays a crucial role in the chronic complicationsof the diabetic liver, where it is associated with an overproduc-tion of oxygen free radicals and lipid peroxidation (Saravanan andPonmurugan, 2011).
The use of herbal medicine in defending against STZ-inducedliver damage looks promising. Curcumin has been gaining atten-tion because of its health benefits, such as its anti-inflammatory,antioxidant, and immune modulatory effects. Curcumin is the chiefcurcuminoid of Curcuma Longa, which is a member of the Zingib-eraceae family (Kumar et al., 2015).
The polyphenol curcumin improves diabetes-induced dysfunc-tion by decreasing the level of glucose, inhibiting protein-kinaseC, and lowering superoxide production (Rungseesantivanon et al.,2010). It reverses insulin resistance, hyperglycemia, and hyper-lipidemia by inhibiting the pro-inflammatory transcription factors,signal transducers and stimulating anti-inflammatory signalingpathways (Tang and Chen, 2010).
This study aimed to investigate the probable influence of hepatic
stellate cells in the protective capability of curcumin in STZ-inducedliver damage. It was also intended to provide an important supportin understanding the mechanism of curcumin treatment for hepaticdysfunction.
8 and C
2
2
MA
2
StpLan
2
wh4sTta
2
occtotB
2
1G(Sibtryp
2
rL
2
iC
2 H.N. Mustafa / Tissue
. Materials and methods
.1. Ethical approval
This study was conducted after receiving the approval of theedical Research Ethics Committee, Faculty of Medicine, King
bdulaziz University, Jeddah, Saudi Arabia.
.2. Chemicals and Reagents
Streptozotocin (STZ) and curcumin were purchased fromigma–Aldrich Chemicals (St. Louis, MO, USA). Alanine amino-ransferase (ALT), aspartate aminotransferase (AST), and alkalinehosphatase (ALP) kits were purchased from Randox Laboratoriestd. (Crumlin, County Antrim, UK). Serum albumin, total protein,nd total bilirubin colorimetric kits were supplied by the Bio Diag-ostic Company (Cairo, Egypt).
.3. Animals
Sixty male adult albino Wistar rats, weighing 190 ± 20 g, thatere used were obtained from the animal house. Animals wereoused in a (24 ◦C ± 3 ◦C) temperature-controlled room with0–70% humidity and 12/12 h light/dark cycle. Rats were fed atandard diet and tap water ad libitum throughout the experiment.he experimental procedures were performed in accordance withhe international guidelines for the care and the use of animals in
laboratory.
.4. Induction of diabetes
Fasted rats (12 h) received a single intraperitoneal (IP) injectionf freshly prepared STZ (50 mg/kg body weight) dissolved in 0.1 Mitrate buffer (pH 4.5). STZ-injected animals were given a 5% glu-ose solution for 24 h to overcome drug-induced hypoglycemia. Onhe third day after STZ injection, glucose levels were estimated bybtaining blood samples from the cut tip of the tail using Diagnos-ic Accu-Chek test strips (Roche Diagnostics, Mannheim, Germany).lood glucose levels of 250 mg/dl or more were considered diabetic.
.5. Experimental design
The animals were distributed into 3 groups (20/group). Group (normal control) was injected IP with a citrate buffer vehicle.roup 2 (diabetic control) received a single IP injection of STZ
50 mg/kg body weight). Group 3 received a single IP injection ofTZ (50 mg/kg body weight) and, on the third day after the STZnjection, curcumin was given orally with a dosage of 80 mg/kgody weight and continued daily for 60 days (Zhang et al., 2013). Athe end of the experiment, blood samples were collected from theetro-orbital sinus in heparinized capillary tubes for serum anal-sis. Animals from all groups were sacrificed, and the livers wererocessed for histological studies.
.6. Plasma glucose estimation
Plasma glucose was measured using an enzymatic colorimet-ic method with commercially available kits (Randox Laboratories,td., Antrim, UK).
.7. Plasma insulin estimation
Plasma insulin was determined using an insulin enzyme-linkedmmunosorbent assay (ELISA) kit (code no. AKRIN-010T; Shibayagio., Ltd., Gunma, Japan).
ell 48 (2016) 81–88
2.8. Serum parameters
ALT, AST, and ALP, which were increased following hepatocyteinjury, were assessed according to the protocol detailed in themanuals of the diagnostic kits (Randox Laboratories Ltd., Crum-lin, County Antrim, UK). Serum albumin, total protein, and totalbilirubin were determined by spectrophotometer using the cor-responding colorimetric kits supplied by Bio Diagnostic Company(Cairo, Egypt) (Lee et al., 2012).
2.9. Histological examination
Livers were washed with a phosphate buffer solution and thenfixed in 10% neutral buffered formalin. Tissues were dehydratedthrough a graded series of alcohol, cleared in xylene, and embed-ded in paraffin wax. Tissues were then cut into sections of 3–5 �min thickness using a microtome and stained with hematoxylin andeosin (H&E) for histopathological evaluation and with periodicAcid-Schiff (PAS) for observation of glycogen. For each specimen,at least three to five slides were examined using an OlympusBX53 microscope equipped with a DP73 camera (Olympus, Tokyo,Japan).
2.10. Histopathological evaluation
The sections were analyzed for hydropic swelling, parenchy-matous degeneration, microvesicular vacuole, macrovesicularvacuole, focal necrosis, inflammatory infiltrations, fibrosis, andsinusoids hyperemia. At the end of the analyses, the findings werepresented in a table which showed the degree of degeneration(Guven et al., 2006). Score levels of 0, +1, +2, +3 were equiva-lent to no, mild, moderate, and severe, respectively. The scoresrepresented values obtained from the tissue sections of six ani-mals from each group with five fields/section (Mustafa et al.,2015).
2.11. Immunohistochemical examination
The standard peroxidase immunohistochemistry technique wasapplied to slides of paraffin-embedded tissue. Sections werede-waxed in xylene, rehydrated, and pretreated with 3% of hydro-gen peroxide solution to block endogenous peroxidase activity.Microwave-assisted antigen retrieval was performed for 20 min.Slides were then incubated overnight at 4 ◦C with the primaryantibody against �-SMA (a mouse monoclonal antibody [Dako,Carpinteria, California, USA] with a dilution of 1:50; cellular site wascytoplasmic) as a marker of activated HSCs with varying degreesof intensity in smooth muscles and myofibroblasts. They weresimilarly incubated with the primary antibody against insulin (amouse monoclonal antibody [Dako, Carpinteria, California, USA]with a dilution of 1:100; cellular site was cytoplasmic). The sectionswere incubated with biotinylated IgG and then with streptavidin-peroxidase conjugate (Zymed Corp, San Francisco, CA, USA).Sections were then washed with phosphate-buffered saline (PBS)and incubated with 3, 3′-diaminobenzidine tetrachloride (DAB)substrate chromogen solution (1 drop of DAB chromogen/1 mL ofsubstrate buffer) for 5 min to detect immunoreactivity. All sectionswere counter-stained with Mayer’s hematoxylin. Negative controlsections were prepared by omitting the primary antibody. Positivecontrol standard laboratory slides were used for all stains to provethe success of the technique. All slides were examined under light
microscopy, and the presence of labeled cells was documented.Absence of staining was recognized as a negative result (−), whilethe presence of brown staining was recognized as positive result(+) (Mustafa et al., 2015).
H.N. Mustafa / Tissue and Cell 48 (2016) 81–88 83
Table 1Body and liver weight of different groups.
Normal control (n = 20) STZ-diabetic control (n = 20) STZ-diabetes + curcumin (n = 20)
Liver weight (g) 11.14 ± 1.54 9.91 ± 1.04P* < 0.01
10.83 ± 0.85*P > 0.05**P < 0.05
Body weight (g) 187.65 ± 6.11 163.94 ± 4.9*P < 0.001
180.47 ± 3.89*P < 0.001**P < 0.001
Values are means ± SD (n = 20 each group). ANOVA followed by Tukey’s post hoc test.*P: compared to normal control. **P: compared to diabetic control.
ven in
2
cIMnJt2
2
pot2
3
3
st
3
iilcl
cws
Fig. 1. Body and liver weight of different groups. The mean is gi
.12. Quantitative morphometric measurements
Ten non-overlapping fields for each animal were selected indis-riminately and analyzed. The measurements were done by usingmage-Pro Plus v6.0 software (Media Cybernetics, Silver Spring,
D, USA) and the NIH ImageJ (v1.50) program (http://rsb.info.ih.gov/ij/) with an Olympus BX53 microscope (Olympus, Tokyo,
apan). The area percentages of �-SMA immunopositive cells andhe insulin-positive cells number were evaluated (Hussein et al.,015).
.13. Statistical analysis
Quantitative data were expressed as the mean ± SD of differentarameters for the treated groups. The data were analyzed usingne-way analysis of variance (ANOVA) followed by Tukey’s post hocest. The statistical analysis was performed using IBM SPSS version3. The values were considered significant when p < 0.05.
. Results
.1. Effect on liver and body weight
At the end of 60 days, curcumin treated specimens showed aignificant increase in the body and liver weight as compared tohe diabetic control (Table 1 and Fig. 1).
.2. Biochemical parameters
The mean glucose level of the diabetic control group was signif-cantly increased while curcumin treatment showed a significantmprovement in glucose level (Table 2 and Fig. 2). The mean insulinevel of the diabetic control group was significantly decreased whileurcumin treatment showed a significant improvement in insulinevel (Table 2 and Fig. 2).
Plasma ALT, AST, and ALP levels were elevated in the diabeticontrol group in comparison to those of the normal control group,hile with curcumin treatment all of these parameters are reduced
ignificantly. Furthermore, there was a significant decrease in the
columns, and error bars represent the standard deviation (SD).
albumin and total protein in the diabetic control group, whilecurcumin treatment showed a significant increase in these param-eters compared to those of the diabetic control group. Conversely,the total bilirubin was increased in the diabetic control groupand decreased significantly with curcumin treatment (Table 2 andFig. 3).
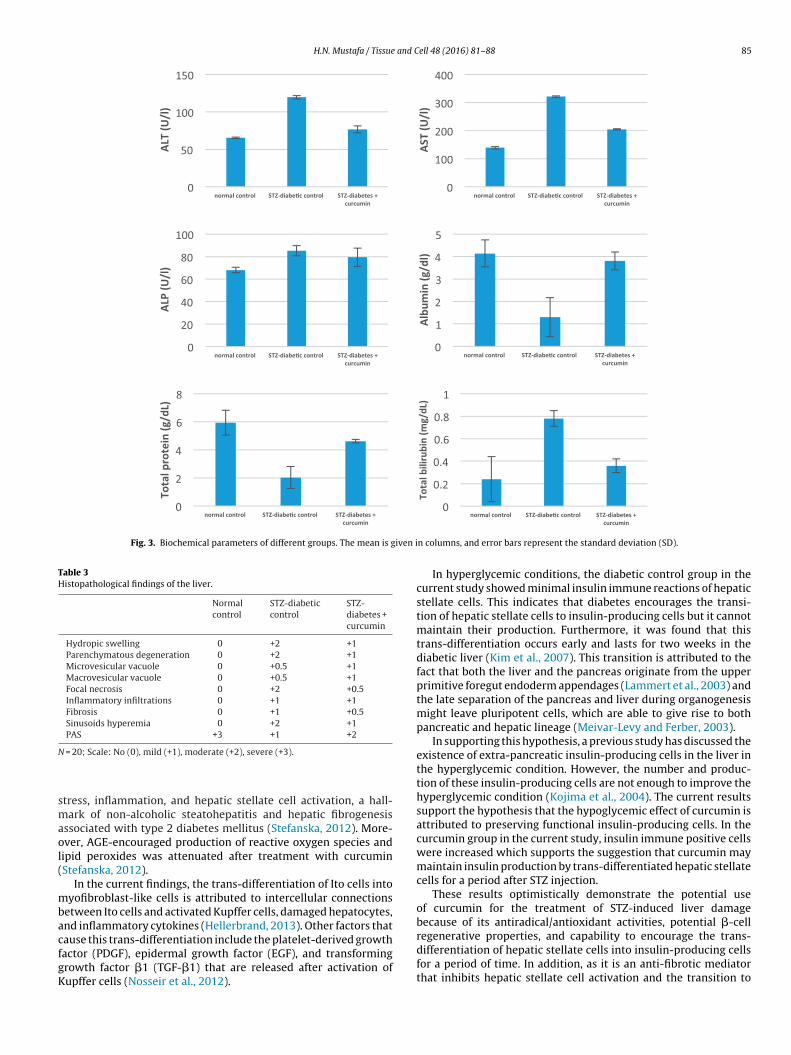
3.3. Histopathological findings
The normal control group showed normal cytoarchitecture. Thediabetic control group showed parenchymatous degeneration andloss of architecture. With curcumin treatment, hepatic changeswere reduced (Table 3).
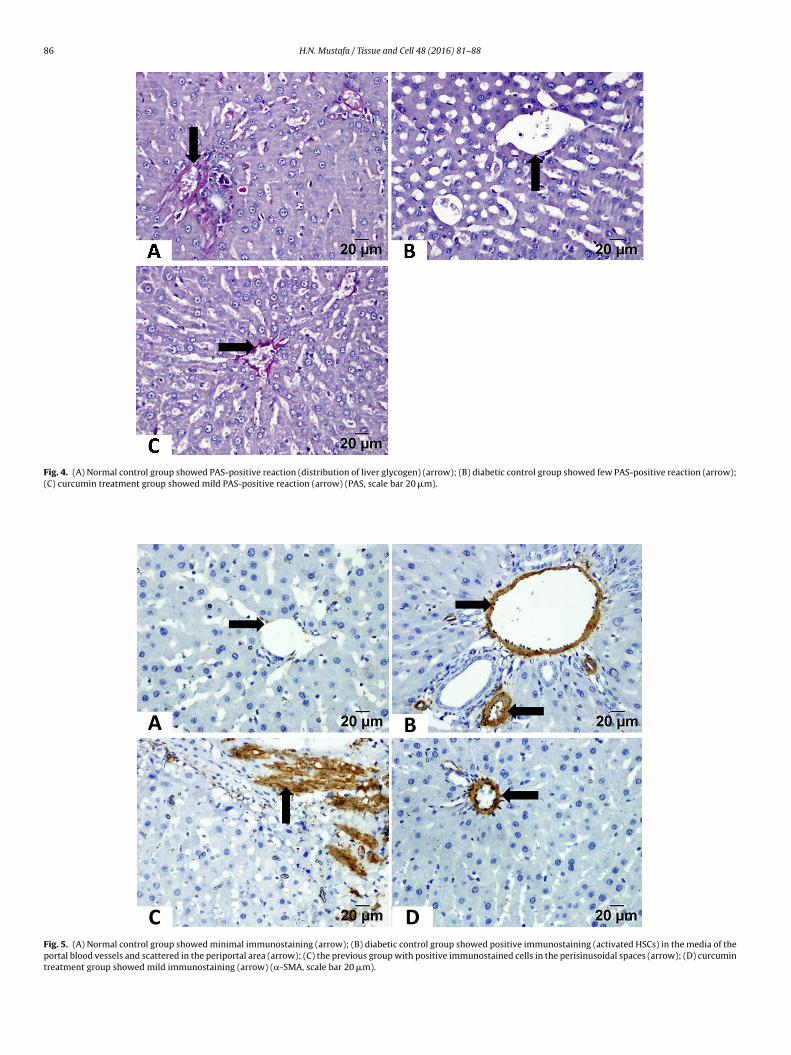
PAS-stained sections of the normal control group showed PAS-positive red granules in the cytoplasm of the hepatocytes andmore granules around the central vein (Fig. 4A), while thoseof the diabetic control group showed few PAS-positive granules(Fig. 4B). With curcumin treatment, mild PAS-positive granuleswere observed (Fig. 4C and Table 3).
3.4. Immunohistochemistry-stained sections
In the normal control group, �-SMA staining was observed onlyin the media of the blood vessels (portal veins, hepatic arterybranches, and central veins) (Fig. 5A). While the diabetic controlgroup showed positive �-SMA immunoreactivity in the media ofthe portal vessels and the periportal area and along the perisinu-soidal spaces (Fig. 5B and C). In the curcumin treatment group,mild immunoreactivity was observed (Fig. 5D). The area percent-age of �-SMA positive cells was significantly higher in the diabeticcontrol group, while in the curcumin treatment group it wasreduced significantly (Table 4 and Fig. 7).
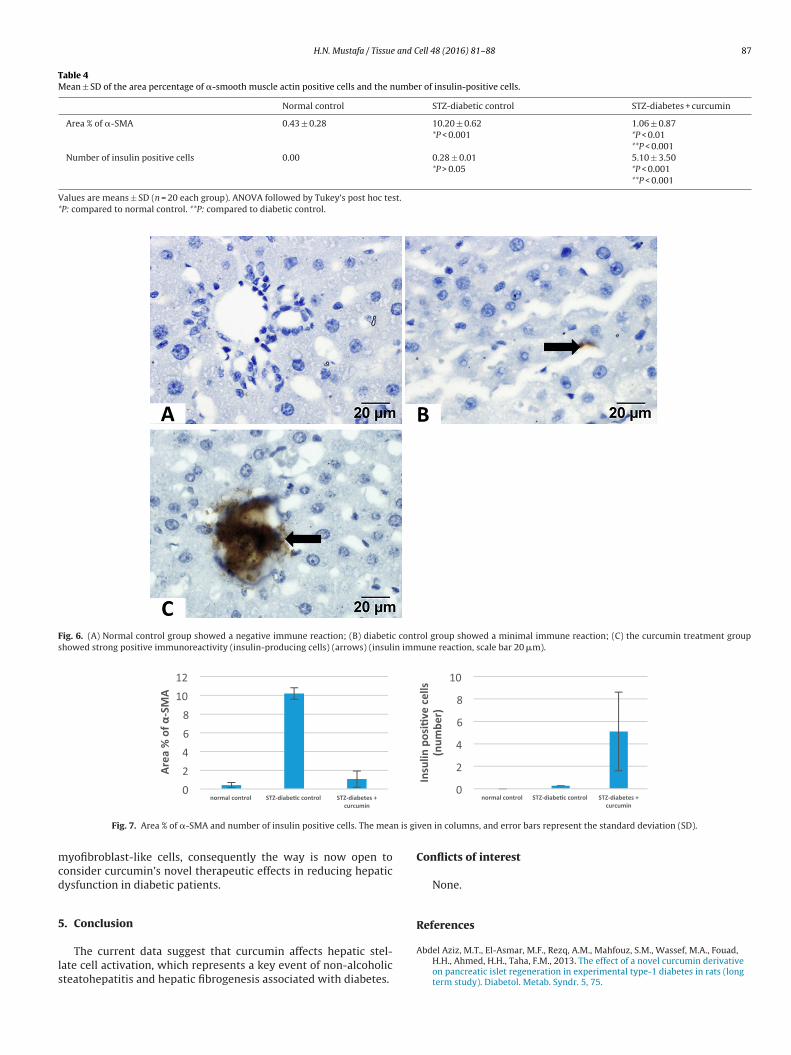
Insulin-immunostained sections for both the normal control(Fig. 6A) and diabetic control (Fig. 6B) groups showed no or mini-mal insulin immune positive cells. The curcumin treatment group
showed many insulin positive immune cells in the portal areaaround the bile ductule (Fig. 6C). The number of insulin positivecells was significantly higher in the curcumin treated group (Table 4and Fig. 7).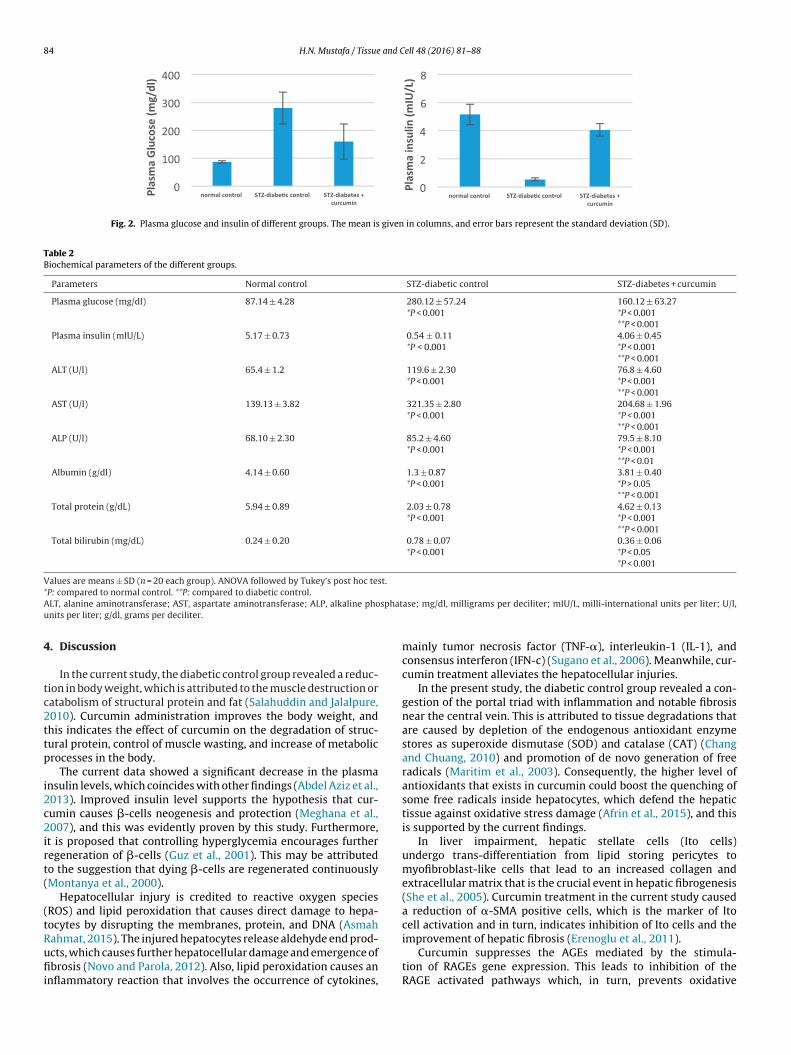
84 H.N. Mustafa / Tissue and Cell 48 (2016) 81–88
Fig. 2. Plasma glucose and insulin of different groups. The mean is given in columns, and error bars represent the standard deviation (SD).
Table 2Biochemical parameters of the different groups.
Parameters Normal control STZ-diabetic control STZ-diabetes + curcumin
Plasma glucose (mg/dl) 87.14 ± 4.28 280.12 ± 57.24*P < 0.001
160.12 ± 63.27*P < 0.001**P < 0.001
Plasma insulin (mIU/L) 5.17 ± 0.73 0.54 ± 0.11*P < 0.001
4.06 ± 0.45*P < 0.001**P < 0.001
ALT (U/l) 65.4 ± 1.2 119.6 ± 2.30*P < 0.001
76.8 ± 4.60*P < 0.001**P < 0.001
AST (U/l) 139.13 ± 3.82 321.35 ± 2.80*P < 0.001
204.68 ± 1.96*P < 0.001**P < 0.001
ALP (U/l) 68.10 ± 2.30 85.2 ± 4.60*P < 0.001
79.5 ± 8.10*P < 0.001**P < 0.01
Albumin (g/dl) 4.14 ± 0.60 1.3 ± 0.87*P < 0.001
3.81 ± 0.40*P > 0.05**P < 0.001
Total protein (g/dL) 5.94 ± 0.89 2.03 ± 0.78*P < 0.001
4.62 ± 0.13*P < 0.001**P < 0.001
Total bilirubin (mg/dL) 0.24 ± 0.20 0.78 ± 0.07*P < 0.001
0.36 ± 0.06*P < 0.05*P < 0.001
Values are means ± SD (n = 20 each group). ANOVA followed by Tukey’s post hoc test.*A sphatu
4
tc2ttp
i2c2irt(
(tRufii
P: compared to normal control. **P: compared to diabetic control.LT, alanine aminotransferase; AST, aspartate aminotransferase; ALP, alkaline phonits per liter; g/dl, grams per deciliter.
. Discussion
In the current study, the diabetic control group revealed a reduc-ion in body weight, which is attributed to the muscle destruction oratabolism of structural protein and fat (Salahuddin and Jalalpure,010). Curcumin administration improves the body weight, andhis indicates the effect of curcumin on the degradation of struc-ural protein, control of muscle wasting, and increase of metabolicrocesses in the body.
The current data showed a significant decrease in the plasmansulin levels, which coincides with other findings (Abdel Aziz et al.,013). Improved insulin level supports the hypothesis that cur-umin causes �-cells neogenesis and protection (Meghana et al.,007), and this was evidently proven by this study. Furthermore,
t is proposed that controlling hyperglycemia encourages furtheregeneration of �-cells (Guz et al., 2001). This may be attributedo the suggestion that dying �-cells are regenerated continuouslyMontanya et al., 2000).
Hepatocellular injury is credited to reactive oxygen speciesROS) and lipid peroxidation that causes direct damage to hepa-ocytes by disrupting the membranes, protein, and DNA (Asmah
ahmat, 2015). The injured hepatocytes release aldehyde end prod-cts, which causes further hepatocellular damage and emergence ofbrosis (Novo and Parola, 2012). Also, lipid peroxidation causes annflammatory reaction that involves the occurrence of cytokines,
ase; mg/dl, milligrams per deciliter; mIU/L, milli-international units per liter; U/l,
mainly tumor necrosis factor (TNF-�), interleukin-1 (IL-1), andconsensus interferon (IFN-c) (Sugano et al., 2006). Meanwhile, cur-cumin treatment alleviates the hepatocellular injuries.
In the present study, the diabetic control group revealed a con-gestion of the portal triad with inflammation and notable fibrosisnear the central vein. This is attributed to tissue degradations thatare caused by depletion of the endogenous antioxidant enzymestores as superoxide dismutase (SOD) and catalase (CAT) (Changand Chuang, 2010) and promotion of de novo generation of freeradicals (Maritim et al., 2003). Consequently, the higher level ofantioxidants that exists in curcumin could boost the quenching ofsome free radicals inside hepatocytes, which defend the hepatictissue against oxidative stress damage (Afrin et al., 2015), and thisis supported by the current findings.
In liver impairment, hepatic stellate cells (Ito cells)undergo trans-differentiation from lipid storing pericytes tomyofibroblast-like cells that lead to an increased collagen andextracellular matrix that is the crucial event in hepatic fibrogenesis(She et al., 2005). Curcumin treatment in the current study causeda reduction of �-SMA positive cells, which is the marker of Itocell activation and in turn, indicates inhibition of Ito cells and the
improvement of hepatic fibrosis (Erenoglu et al., 2011).Curcumin suppresses the AGEs mediated by the stimula-tion of RAGEs gene expression. This leads to inhibition of theRAGE activated pathways which, in turn, prevents oxidative
H.N. Mustafa / Tissue and Cell 48 (2016) 81–88 85
Fig. 3. Biochemical parameters of different groups. The mean is given i
Table 3Histopathological findings of the liver.
Normalcontrol
STZ-diabeticcontrol
STZ-diabetes +curcumin
Hydropic swelling 0 +2 +1Parenchymatous degeneration 0 +2 +1Microvesicular vacuole 0 +0.5 +1Macrovesicular vacuole 0 +0.5 +1Focal necrosis 0 +2 +0.5Inflammatory infiltrations 0 +1 +1Fibrosis 0 +1 +0.5Sinusoids hyperemia 0 +2 +1PAS +3 +1 +2
N
smaol(
mbacfgK
regenerative properties, and capability to encourage the trans-
= 20; Scale: No (0), mild (+1), moderate (+2), severe (+3).
tress, inflammation, and hepatic stellate cell activation, a hall-ark of non-alcoholic steatohepatitis and hepatic fibrogenesis
ssociated with type 2 diabetes mellitus (Stefanska, 2012). More-ver, AGE-encouraged production of reactive oxygen species andipid peroxides was attenuated after treatment with curcuminStefanska, 2012).
In the current findings, the trans-differentiation of Ito cells intoyofibroblast-like cells is attributed to intercellular connections
etween Ito cells and activated Kupffer cells, damaged hepatocytes,nd inflammatory cytokines (Hellerbrand, 2013). Other factors thatause this trans-differentiation include the platelet-derived growth
actor (PDGF), epidermal growth factor (EGF), and transformingrowth factor �1 (TGF-�1) that are released after activation ofupffer cells (Nosseir et al., 2012).n columns, and error bars represent the standard deviation (SD).
In hyperglycemic conditions, the diabetic control group in thecurrent study showed minimal insulin immune reactions of hepaticstellate cells. This indicates that diabetes encourages the transi-tion of hepatic stellate cells to insulin-producing cells but it cannotmaintain their production. Furthermore, it was found that thistrans-differentiation occurs early and lasts for two weeks in thediabetic liver (Kim et al., 2007). This transition is attributed to thefact that both the liver and the pancreas originate from the upperprimitive foregut endoderm appendages (Lammert et al., 2003) andthe late separation of the pancreas and liver during organogenesismight leave pluripotent cells, which are able to give rise to bothpancreatic and hepatic lineage (Meivar-Levy and Ferber, 2003).
In supporting this hypothesis, a previous study has discussed theexistence of extra-pancreatic insulin-producing cells in the liver inthe hyperglycemic condition. However, the number and produc-tion of these insulin-producing cells are not enough to improve thehyperglycemic condition (Kojima et al., 2004). The current resultssupport the hypothesis that the hypoglycemic effect of curcumin isattributed to preserving functional insulin-producing cells. In thecurcumin group in the current study, insulin immune positive cellswere increased which supports the suggestion that curcumin maymaintain insulin production by trans-differentiated hepatic stellatecells for a period after STZ injection.
These results optimistically demonstrate the potential useof curcumin for the treatment of STZ-induced liver damagebecause of its antiradical/antioxidant activities, potential �-cell
differentiation of hepatic stellate cells into insulin-producing cellsfor a period of time. In addition, as it is an anti-fibrotic mediatorthat inhibits hepatic stellate cell activation and the transition to
86 H.N. Mustafa / Tissue and Cell 48 (2016) 81–88
Fig. 4. (A) Normal control group showed PAS-positive reaction (distribution of liver glycogen) (arrow); (B) diabetic control group showed few PAS-positive reaction (arrow);(C) curcumin treatment group showed mild PAS-positive reaction (arrow) (PAS, scale bar 20 �m).
Fig. 5. (A) Normal control group showed minimal immunostaining (arrow); (B) diabetic control group showed positive immunostaining (activated HSCs) in the media of theportal blood vessels and scattered in the periportal area (arrow); (C) the previous group with positive immunostained cells in the perisinusoidal spaces (arrow); (D) curcumintreatment group showed mild immunostaining (arrow) (�-SMA, scale bar 20 �m).
H.N. Mustafa / Tissue and Cell 48 (2016) 81–88 87
Table 4Mean ± SD of the area percentage of �-smooth muscle actin positive cells and the number of insulin-positive cells.
Normal control STZ-diabetic control STZ-diabetes + curcumin
Area % of �-SMA 0.43 ± 0.28 10.20 ± 0.62*P < 0.001
1.06 ± 0.87*P < 0.01**P < 0.001
Number of insulin positive cells 0.00 0.28 ± 0.01*P > 0.05
5.10 ± 3.50*P < 0.001**P < 0.001
Values are means ± SD (n = 20 each group). ANOVA followed by Tukey’s post hoc test.*P: compared to normal control. **P: compared to diabetic control.
Fig. 6. (A) Normal control group showed a negative immune reaction; (B) diabetic control group showed a minimal immune reaction; (C) the curcumin treatment groupshowed strong positive immunoreactivity (insulin-producing cells) (arrows) (insulin immune reaction, scale bar 20 �m).
n is gi
mcd
5
ls
Fig. 7. Area % of �-SMA and number of insulin positive cells. The mea
yofibroblast-like cells, consequently the way is now open toonsider curcumin’s novel therapeutic effects in reducing hepaticysfunction in diabetic patients.
. Conclusion
The current data suggest that curcumin affects hepatic stel-ate cell activation, which represents a key event of non-alcoholicteatohepatitis and hepatic fibrogenesis associated with diabetes.
ven in columns, and error bars represent the standard deviation (SD).
Conflicts of interest
None.
References
Abdel Aziz, M.T., El-Asmar, M.F., Rezq, A.M., Mahfouz, S.M., Wassef, M.A., Fouad,H.H., Ahmed, H.H., Taha, F.M., 2013. The effect of a novel curcumin derivativeon pancreatic islet regeneration in experimental type-1 diabetes in rats (longterm study). Diabetol. Metab. Syndr. 5, 75.

8 and C
A
A
B
C
C
E
G
G
H
H
K
K
K
L
L
M
8 H.N. Mustafa / Tissue
frin, R., Arumugam, S., Soetikno, V., Thandavarayan, R.A., Pitchaimani, V.,Karuppagounder, V., Sreedhar, R., Harima, M., Suzuki, H., Miyashita, S.,Nomoto, M., Suzuki, K., Watanabe, K., 2015. Curcumin amelioratesstreptozotocin-induced liver damage through modulation of endoplasmicreticulum stress-mediated apoptosis in diabetic rats. Free Radic. Res. 49,279–289.
smah Rahmat, A.A., 2015. Effect of pomegranate on histopathology of liver andkidney on generated oxidative stress diabetic induced rats. J. Cytol. Histol. 6, 2.
ugianesi, E., Vanni, E., Marchesini, G., 2007. NASH and the risk of cirrhosis andhepatocellular carcinoma in type 2 diabetes. Curr. Diab. Rep. 7,175–180.
hang, Y.C., Chuang, L.M., 2010. The role of oxidative stress in the pathogenesis oftype 2 diabetes: from molecular mechanism to clinical implication. Am. J.Transl. Res. 2, 316–331.
lement, S., Pascarella, S., Conzelmann, S., Gonelle-Gispert, C., Guilloux, K., Negro,F., 2010. The hepatitis C virus core protein indirectly induces alpha-smoothmuscle actin expression in hepatic stellate cells via interleukin-8. J. Hepatol.52, 635–643.
renoglu, C., Kanter, M., Aksu, B., Sagiroglu, T., Ayvaz, S., Aktas, C., Erboga, M., 2011.Protective effect of curcumin on liver damage induced by biliary obstruction inrats. Balkan Med. J. 28, 352–357.
uven, A., Yavuz, O., Cam, M., Ercan, F., Bukan, N., Comunoglu, C., Gokce, F., 2006.Effects of melatonin on streptozotocin-induced diabetic liver injury in rats.Acta Histochem. 108, 85–93.
uz, Y., Nasir, I., Teitelman, G., 2001. Regeneration of pancreatic beta cells fromintra-islet precursor cells in an experimental model of diabetes. Endocrinology142, 4956–4968.
ellerbrand, C., 2013. Hepatic stellate cells-the pericytes in the liver. Pflügers Arch.465, 775–778.
ussein, A.M., Mustafa, H.N., Badawoud, M.H., 2015. Ameliorative potentials of acombination of fenugreek and alpha-tocopherol on cadmium inducedtesticular toxicity: an ultrastructural study. Folia Morphol. (Warsz) 74,325–334.
im, S., Shin, J.S., Kim, H.J., Fisher, R.C., Lee, M.J., Kim, C.W., 2007.Streptozotocin-induced diabetes can be reversed by hepatic oval cellactivation through hepatic transdifferentiation and pancreatic isletregeneration. Lab. Invest. 87, 702–712.
ojima, H., Fujimiya, M., Matsumura, K., Nakahara, T., Hara, M., Chan, L., 2004.Extrapancreatic insulin-producing cells in multiple organs in diabetes. Proc.Natl. Acad. Sci. USA 101, 2458–2463.
umar, A., Chetia, H., Sharma, S., Kabiraj, D., Talukdar, N.C., Bora, U., 2015.Curcumin Resource Database. Database (Oxford) 2015, bav070.
ammert, E., Cleaver, O., Melton, D., 2003. Role of endothelial cells in early
pancreas and liver development. Mech. Dev. 120, 59–64.ee, T.H., Kim, W.R., Poterucha, J.J., 2012. Evaluation of elevated liver enzymes. Clin.Liver Dis. 16, 183–198.
aritim, A.C., Sanders, R.A., Watkins 3rd, J.B., 2003. Diabetes, oxidativestress, and antioxidants: a review. J. Biochem. Mol. Toxicol. 17, 24–38.
ell 48 (2016) 81–88
Meghana, K., Sanjeev, G., Ramesh, B., 2007. Curcumin preventsstreptozotocin-induced islet damage by scavenging free radicals: aprophylactic and protective role. Eur. J. Pharmacol. 577, 183–191.
Meivar-Levy, I., Ferber, S., 2003. New organs from our own tissues:liver-to-pancreas transdifferentiation. Trends Endocrinol. Metab. 14,460–466.
Montanya, E., Nacher, V., Biarnés, M., Soler, J., 2000. Linear correlation betweenbeta-cell mass and body weight throughout the lifespan in Lewis rats: role ofbeta-cell hyperplasia and hypertrophy. Diabetes 49, 1341–1346.
Mustafa, H., El Awdan, S., Hegazy, G., Abdel Jaleel, G., 2015. Prophylactic role ofcoenzyme Q10 and Cynara scolymus L on doxorubicin-induced toxicityin rats: Biochemical and immunohistochemical study. Indian J. Pharmacol.47, 649.
Nosseir, D.A., Gawish, S.A., Moustafa, A.M., Firgany, A.E.-D.L., 2012. The possibleprotective effect of green tea versus interferon-� on carbontetrachloride-induced hepatic fibrosis in albino rat. Egypt. J. Histol. 35,398–411.
Novo, E., Parola, M., 2012. The role of redox mechanisms in hepatic chronic woundhealing and fibrogenesis. Fibrogenesis Tissue Repair 5, S4.
Rungseesantivanon, S., Thenchaisri, N., Ruangvejvorachai, P., Patumraj, S., 2010.Curcumin supplementation could improve diabetes-induced endothelialdysfunction associated with decreased vascular superoxide production andPKC inhibition. BMC Complement. Altern. Med. 10, 57.
Salahuddin, M., Jalalpure, S.S., 2010. Antidiabetic activity of aqueous fruit extract ofCucumis trigonus Roxb. in streptozotocin-induced-diabetic rats. J.Ethnopharmacol. 127, 565–567.
Saravanan, G., Ponmurugan, P., 2011. Ameliorative potential of S-allyl cysteine onoxidative stress in STZ induced diabetic rats. Chem. Biol. Interact. 189,100–106.
She, H., Xiong, S., Hazra, S., Tsukamoto, H., 2005. Adipogenic transcriptionalregulation of hepatic stellate cells. J. Biol. Chem. 280, 4959–4967.
Stefanska, B., 2012. Curcumin ameliorates hepatic fibrosis in type 2 diabetesmellitus - insights into its mechanisms of action. Br. J. Pharmacol. 166,2209–2211.
Sugano, M., Yamato, H., Hayashi, T., Ochiai, H., Kakuchi, J., Goto, S., Nishijima, F.,Iino, N., Kazama, J.J., Takeuchi, T., Mokuda, O., Ishikawa, T., Okazaki, R., 2006.High-fat diet in low-dose-streptozotocin-treated heminephrectomized ratsinduces all features of human type 2 diabetic nephropathy: a new rat model ofdiabetic nephropathy. Nutr. Metab. Cardiovasc. Dis. 16, 477–484.
Tang, Y., Chen, A., 2010. Curcumin prevents leptin raising glucose levels in hepaticstellate cells by blocking translocation of glucose transporter-4 and increasingglucokinase. Br. J. Pharmacol. 161, 1137–1149.
Yang, Z., Li, K., Yan, X., Dong, F., Zhao, C., 2010. Amelioration of diabetic retinopathy
by engrafted human adipose-derived mesenchymal stem cells instreptozotocin diabetic rats. Graefes Arch. Clin. Exp. Ophthalmol. 248,1415–1422.Zhang, D.W., Fu, M., Gao, S.H., Liu, J.L., 2013. Curcumin and diabetes: a systematicreview. Evid. Based Complement. Alternat. Med. 2013, 636053.