Embed Size (px)
Citation preview

A RT I C L E S
968 NATURE CELL BIOLOGY VOLUME 6 | NUMBER 10 | OCTOBER 2004
A new effector pathway links ATM kinase with theDNA damage responseConstantinos Demonacos1,2, Marija Krstic-Demonacos1,2, Linda Smith1, Danmei Xu1, Darran P. O’Connor1,Martin Jansson1 and Nicholas B. La Thangue1,4
The related kinases ATM (ataxia-telangiectasia mutated) and ATR (ataxia-telangiectasia and Rad3-related) phosphorylate alimited number of downstream protein targets in response to DNA damage. Here we report a new pathway in which ATM kinasesignals the DNA damage response by targeting the transcriptional cofactor Strap. ATM phosphorylates Strap at a serine residue,stabilizing nuclear Strap and facilitating formation of a stress-responsive co-activator complex. Strap activity enhances p53acetylation, and augments the response to DNA damage. Strap remains localized in the cytoplasm in cells derived from ataxiatelangiectasia individuals with defective ATM, as well as in cells expressing a Strap mutant that cannot be phosphorylated byATM. Targeting Strap to the nucleus reinstates protein stabilization and activates the DNA damage response. These resultsindicate that the nuclear accumulation of Strap is a critical regulator in the damage response, and argue that this function canbe assigned to ATM through the DNA damage-dependent phosphorylation of Strap.
The DNA damage signalling pathway is a highly conserved response togenotoxic stress1. In mammalian cells, the pathway functions to protectcells from agents that induce cellular death or transformation, where itparticipates in DNA repair and checkpoint control leading to survivalor apoptosis1. The related phosphatidylinositol-3-OH-kinase-likekinases (PI(3)KK) ATM and ATR, which become activated in responseto DNA damage, transduce signals to downstream targets, includingp53 and the checkpoint kinases Chk1 and Chk2 (refs 2–9). In turn,checkpoint kinases phosphorylate key substrates, such as p53, E2F-1,cyclin dependent kinase 25A (Cdc25A) and Cdc25C10–14, to facilitatethe DNA damage response.
The tumour suppressor protein p53 has an important function in thecellular response to DNA damage15, and consequently, TP53 is one of themost frequently altered genes in human cancer16. Under normal condi-tions, p53 is held in a latent inactive state but undergoes a significantincrease in protein stability after DNA damage15. p53 stability control isbelieved to involve both families of DNA damage signalling kinases thatactivate p53 function: the ATM/ATR and Chk1/Chk2 families (refs 6, 7,17, 18). In cells derived from ataxia telangiectasia individuals, in whichATM is missing or inactivated19, the DNA damage pathway and p53response is abnormal2,4. Similarly, in some Li-Fraumeni patients with adefective Chk2, the p53 response is also compromised20.
p53 functions as a stress-responsive transcription factor, inducinggenes that facilitate cell-cycle arrest and apoptosis. The phosphoryla-tion of p53 seems to be tightly coordinated with p53 acetylation, medi-ated by the histone acetyltransferase (HAT) in co-activators such asp300/CBP and PCAF (p300/CBP-associated factor)21–23. During theDNA damage response, two cofactors JMY and Strap bind to p300
(refs 24, 25), resulting in a co-activator complex which enhances p53transcriptional activity. Interestingly, Strap contains a tandem series ofTPR repeats (tetratricopeptide repeats), consistent with its role in pro-tein assembly25. Thus, different domains of Strap interact with p300and JMY, which augments interaction between p300 and JMY.
Here, we have explored the possibility that Strap is regulated by theDNA damage signalling pathway, and report a new pathway throughwhich ATM kinase signals to Strap and activates the DNA damageresponse. ATM phosphorylates Strap at a single serine residue, andphosphorylated Strap accumulates in the nucleus, thereby allowingStrap to assemble into a co-activator complex with p300. Strap activ-ity enhances p53 acetylation and augments the DNA damageresponse. In ataxia telangiectasia cells, Strap remains localized in thecytoplasm under DNA damage conditions, and a mutant derivative ofStrap that cannot be phosphosphorylated by ATM phosphorylationlocalizes to the cytoplasm in cells with normal ATM activity.Moreover, targeting Strap to the nucleus is sufficient to stabilize Strapprotein and activate the DNA damage response. Our results define inmechanistic detail a new checkpoint effector pathway that links ATMkinase with the DNA damage response through the phosphorylationand altered intracellular distribution of Strap.
RESULTSStrap is phosphorylated by ATM kinaseTo investigate the regulation of Strap by the DNA damage signallingpathway we assessed whether Strap was induced by different geno-toxic stresses. In cells treated with etoposide, hydroxyurea or strep-tonigrin, there was an increase in the level of Strap (Fig. 1a; bottom).
1Division of Biochemistry and Molecular Biology, Davidson Building, University of Glasgow, Glasgow G12 8QQ, UK. 2Current address: School of Pharmacy andPharmaceutical Sciences (C.D.), School of Biological Sciences (M.K-D.), University of Manchester, Manchester M13 9PT, UK. 4Correspondence should be addressedto N.B.L.T. (e-mail: [email protected]).
Published online: 19 September 2004; DOI:10.1038/ncb1170
print ncb1170 14/9/04 2:58 PM Page 968
© 2004 Nature Publishing Group
© 2004 Nature Publishing Group

A RT I C L E S
NATURE CELL BIOLOGY VOLUME 6 | NUMBER 10 | OCTOBER 2004 969
A substrate-specific antibody that recognizes phospho-serine/threo-nine (S/T) ATM/ATR was used. This antibody recognised Strap inDNA damaged cells, but not in untreated cells (Fig. 1a; top), suggest-ing that Strap is phosphorylated by ATM or ATR kinase during theDNA damage response.
Several consensus phosphorylation sites (SQ or TQ) for ATM andATR kinase are present in Strap. To identify functionally important ser-ine/threonine residues, each of the three theoretical phosphorylationsites was altered (from serine/threonine to alanine) and the phospho-rylation status and stabilization of the mutant derivative assessed incells treated with etoposide. One derivative, with a serine to alaninechange at residue 203 (referred to as StrapS203A) in the third TPR motif
(Fig. 1b) was of particular interest. This was because, in contrast towild-type Strap, StrapS203A was poorly phosphorylated by immunopre-cipitated ATM kinase (Fig. 1c). Moreover, the protein level ofStrapS203A was not increased in cells treated with etoposide but retainedthe constitutive level of expression observed with wild-type Strap inuntreated cells (Fig. 1d); similar effects were not observed with anyother Strap mutant (data not shown). Thus, Ser 203 is phosphorylatedby ATM kinase and is a functionally important residue in regulatingStrap stability in response to DNA damage. It is consistent with theseresults that Strap is an intrinsically unstable protein, the half life ofwhich is increased on etoposide treatment (30 min to 3 h, respectively;see Supplementary Information, Fig. S1).
IP: Strap
IP: ATM kinase
− E Hu S
IB: anti-P-S/T
IB: Strap
P-Strap
Strap
P-Ser 203 Strap
p53
PCNA
Strapp53
Strap
1 2 3 4
1
0 1 3 6 16
1 2 3 4 5
: Bleomycin (h)
2 3 4
1 2 3 4 5 6
1 440l ll lll lV V Vl
N P K I S Q Q A L : WT
A : StrapS203A
a
c
e
d
b
Strap
p53 −−
− + − + − +WT S203A
WT S203AStrap:
Etoposide:
Figure 1 DNA damage induces phosphorylation of Strap. (a) Lysates fromHeLa cells treated with etoposide (E, 10 µM for 12 h; lane 2),hydroxyurea (Hu; 0.5 µM for 12 h; lane 3), streptonigrin (S; 5 ng µl−1 for1 h; lane 4) or untreated (lane 1) were immunoprecipitated with the anti-Strap peptide 510 antibody that recognizes endogenous Strap and thenimmunoblotted with the anti-phospho-S/T ATM/ATR substrate-specificantibody (anti-P-S/T, top). The bottom panel shows the level of inputStrap by immunoblotting the cell extracts with the anti-Strap 510antibody. Equal amounts of protein (5 µg) were loaded. (b) Schematicrepresentation of Strap, showing the six TPR repeats (dark shading). Thesequence surrounding the serine (S) residue (at position 203) in wild-type (WT) Strap is shown, together with the alanine (A) derivative inStrapS203A. (c) An extract from HeLa cells (about 50 µg) was
immunoprecipitated with the anti-ATM Ab3 antibody and kinase reactionsperformed in the presence of purified His-tagged p53, Strap or StrapS203A
(1 µg of each) as indicated, together with a control treatment lackingrecombinant protein (lane 2). (d) U20S cells transfected with expressionvectors encoding WT Strap (10 µg; lanes 3 and 4), StrapS203A (10 µg;lanes 5 and 6) or empty vector (lanes 1 and 2) were treated with etoposide(10 µM for 12 h) and immunoblotted with an anti-HA monoclonalantibody recognizing exogenous Strap. pCMV-β-gal was co-transfected asan internal control, and used to normalize the amount of protein resolvedfor each treatment. (e) U2OS cells were treated with bleomycin (5 µg ml−1) and harvested at the times indicated. Total cell extracts wereimmunoblotted with the anti-phospho Ser 203 peptide antibody (top),anti-p53 antibody (middle) or anti-PCNA antibody (lower).
print ncb1170 14/9/04 2:58 PM Page 969
© 2004 Nature Publishing Group
© 2004 Nature Publishing Group

A RT I C L E S
970 NATURE CELL BIOLOGY VOLUME 6 | NUMBER 10 | OCTOBER 2004
To gain evidence that Ser 203 is phosphorylated in cells by ATM orATR kinase, we monitored the level of phosphorylation of StrapS203A
with the anti-phospho-S/T ATM/ATR antibody in U2OS cells. Althoughwild-type Strap exhibited an increased level of phosphorylation afteretoposide treatment, phosphorylation of StrapS203A was not evident (seeSupplementary Information, Fig. S1), even though similar levels ofStrapS203A and wild-type Strap were expressed (lower panel). Moreover,we used a phospho-specific peptide antibody against phosphorylated Ser203 (see Supplementary Information, Fig. S1) to confirm the phospho-rylation of Ser 203 in cells. Using a tetracycline-inducible stable cell linein which Strap levels could be regulated, Strap was shown to be specifi-cally phosphorylated on Ser 203 (see Supplementary Information,Fig. S2). The anti-phospho Ser 203 antibody recognised endogenousStrap (see Supplementary Information, Fig. S2), and in cells treated withthe DNA damaging agent bleomycin or ionising radiation, Strap under-went increased phosphorylation at Ser 203 (Fig. 1e; also seeSupplementary Information, Fig. S2). Under the conditions ofbleomycin and etoposide treatment, active ATM kinase26 coincided withthe presence of phosphorylated Ser 203 (see SupplementaryInformation, Fig. S2). Thus, Ser 203 is a major site of phosphorylation byATM kinase and, combined with the earlier results (Fig. 1d), implies thatphosphorylation is required for Strap stabilization.
Phosphorylation of Strap at Ser 203 governs intracellular locationWe investigated the mechanism through which the phosphorylation ofSer 203 may influence Strap activity, and to this end considered a rolein regulated intracellular localization. In normal cells, Strap localized tothe nucleus, with an increase in staining intensity occurring afteretoposide treatment (Fig. 2a, c), correlating with the increased level of
Strap after etoposide treatment (Fig. 1a). A similar effect was observedwith exogenous Strap, again exhibiting enhanced nuclear staining incells treated with etoposide (Fig. 2e, g). Remarkably, the StrapS203A
mutant was excluded from nuclei, being localized to the cytoplasm, andtreating cells with etoposide failed to alter its localization (Fig. 2i, k;also see Supplementary Information, Fig. S4). Residue Ser 203 there-fore regulates the intracellular location of Strap.
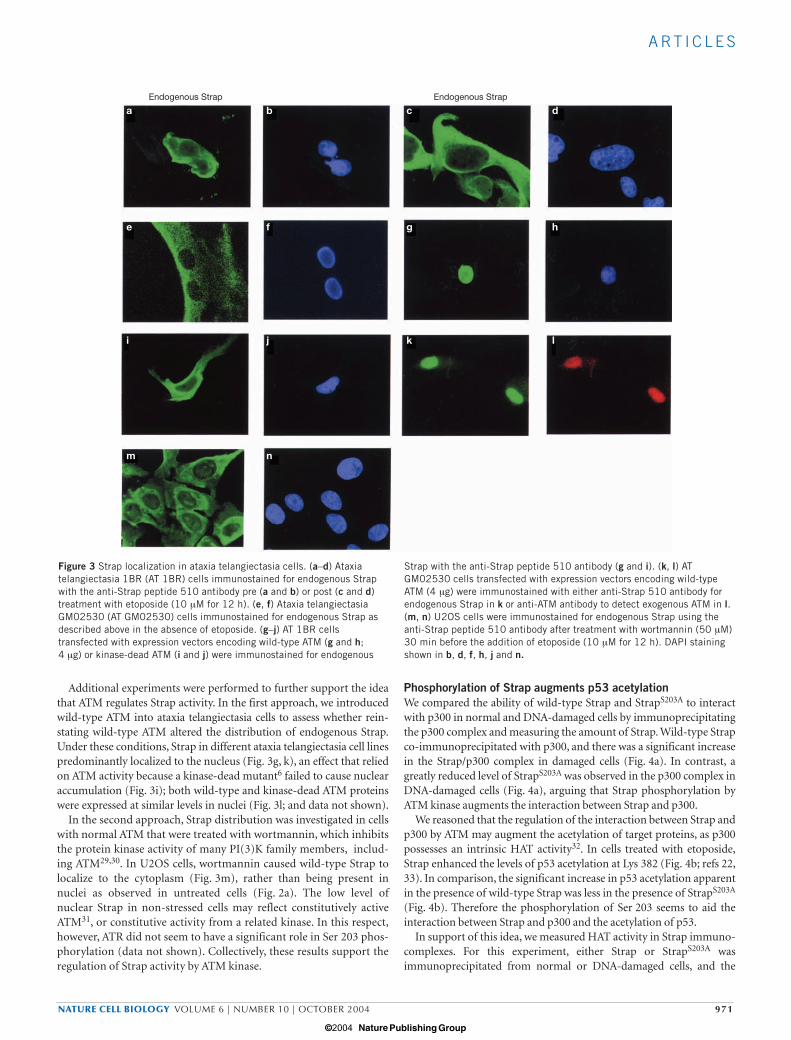
Strap localizes to the cytoplasm in AT cells Ataxia telangiectasia is an autosomal recessive human disease, charac-terized by abnormal chromosome breakage and haematological malig-nancy, that results from mutation in the ATM gene and loss of ATMkinase activity27,28. Given the implied role for ATM kinase in the phos-phorylation and regulation of Strap, we were interested to evaluateStrap in ataxia telangiectasia cells. In two different ataxia telangiectasiacell lines (1BR and GM02530) endogenous Strap localized to the cyto-plasm and was excluded from the nucleus, and etoposide treatmenthad little effect on this distribution (Fig. 3a, c and e). Exogenous wild-type Strap possessed similar properties to endogenous Strap in that itlocalized to the cytoplasm, and similarly was not affected by etoposide(data not shown). Furthermore, Strap did not undergo protein stabi-lization in ataxia telangiectasia cells, and phosphorylated Strap was notapparent in etoposide-treated ataxia telangiectasia cells (seeSupplementary Information, Fig. S1). Thus, the properties of wild-type Strap in ataxia telangiectasia cells recapitulate the properties of theStrapS203A mutant derivative in cells with normal ATM activity (Fig. 2i,k). Because in ataxia telangiectasia cells ATM is missing or inactivated,these results further strengthen the idea that ATM kinase phosphory-lates Ser 203 to regulate the intracellular location of Strap.
a b c d
e f g h
i j k l
Etoposide: − − + +
Endogenous Strap
Exogenous Strap
StrapS203A
Figure 2 Intracellular localization of Strap and StrapS203A. (a–d) U20S cellswere immunostained with the anti-Strap 510 antibody recognizing endogenousStrap pre (a and b) or post (c and d) treatment with etoposide (10 µM for 12 h).(e–l) U20S cells were transfected with expression vectors (5 µg) encoding wild-
type Strap (e–h) or StrapS203A (i–l) and immunostained with the anti-HAmonoclonal antibody pre (e, f, i, j) or post (g, h, k, l) etoposide treatment (10µM for 12 h). DAPI staining shown in b, d, f, h, j, and i. Arrows in k indicate thereduced level of nuclear staining. Original magnification ×630.
print ncb1170 14/9/04 2:58 PM Page 970
© 2004 Nature Publishing Group
© 2004 Nature Publishing Group
A RT I C L E S
NATURE CELL BIOLOGY VOLUME 6 | NUMBER 10 | OCTOBER 2004 971
Additional experiments were performed to further support the ideathat ATM regulates Strap activity. In the first approach, we introducedwild-type ATM into ataxia telangiectasia cells to assess whether rein-stating wild-type ATM altered the distribution of endogenous Strap.Under these conditions, Strap in different ataxia telangiectasia cell linespredominantly localized to the nucleus (Fig. 3g, k), an effect that reliedon ATM activity because a kinase-dead mutant6 failed to cause nuclearaccumulation (Fig. 3i); both wild-type and kinase-dead ATM proteinswere expressed at similar levels in nuclei (Fig. 3l; and data not shown).
In the second approach, Strap distribution was investigated in cellswith normal ATM that were treated with wortmannin, which inhibitsthe protein kinase activity of many PI(3)K family members, includ-ing ATM29,30. In U2OS cells, wortmannin caused wild-type Strap tolocalize to the cytoplasm (Fig. 3m), rather than being present innuclei as observed in untreated cells (Fig. 2a). The low level ofnuclear Strap in non-stressed cells may reflect constitutively activeATM31, or constitutive activity from a related kinase. In this respect,however, ATR did not seem to have a significant role in Ser 203 phos-phorylation (data not shown). Collectively, these results support theregulation of Strap activity by ATM kinase.
Phosphorylation of Strap augments p53 acetylation We compared the ability of wild-type Strap and StrapS203A to interactwith p300 in normal and DNA-damaged cells by immunoprecipitatingthe p300 complex and measuring the amount of Strap. Wild-type Strapco-immunoprecipitated with p300, and there was a significant increasein the Strap/p300 complex in damaged cells (Fig. 4a). In contrast, agreatly reduced level of StrapS203A was observed in the p300 complex inDNA-damaged cells (Fig. 4a), arguing that Strap phosphorylation byATM kinase augments the interaction between Strap and p300.
We reasoned that the regulation of the interaction between Strap andp300 by ATM may augment the acetylation of target proteins, as p300possesses an intrinsic HAT activity32. In cells treated with etoposide,Strap enhanced the levels of p53 acetylation at Lys 382 (Fig. 4b; refs 22,33). In comparison, the significant increase in p53 acetylation apparentin the presence of wild-type Strap was less in the presence of StrapS203A
(Fig. 4b). Therefore the phosphorylation of Ser 203 seems to aid theinteraction between Strap and p300 and the acetylation of p53.
In support of this idea, we measured HAT activity in Strap immuno-complexes. For this experiment, either Strap or StrapS203A wasimmunoprecipitated from normal or DNA-damaged cells, and the
a b c d
e f g h
i j
m n
k l
Endogenous Strap Endogenous Strap
Figure 3 Strap localization in ataxia telangiectasia cells. (a–d) Ataxiatelangiectasia 1BR (AT 1BR) cells immunostained for endogenous Strapwith the anti-Strap peptide 510 antibody pre (a and b) or post (c and d)treatment with etoposide (10 µM for 12 h). (e, f) Ataxia telangiectasiaGM02530 (AT GM02530) cells immunostained for endogenous Strap asdescribed above in the absence of etoposide. (g–j) AT 1BR cellstransfected with expression vectors encoding wild-type ATM (g and h;4 µg) or kinase-dead ATM (i and j) were immunostained for endogenous
Strap with the anti-Strap peptide 510 antibody (g and i). (k, l) ATGM02530 cells transfected with expression vectors encoding wild-typeATM (4 µg) were immunostained with either anti-Strap 510 antibody forendogenous Strap in k or anti-ATM antibody to detect exogenous ATM in l.(m, n) U2OS cells were immunostained for endogenous Strap using theanti-Strap peptide 510 antibody after treatment with wortmannin (50 µM)30 min before the addition of etoposide (10 µM for 12 h). DAPI stainingshown in b, d, f, h, j and n.
print ncb1170 14/9/04 2:58 PM Page 971
© 2004 Nature Publishing Group
© 2004 Nature Publishing Group

A RT I C L E S
972 NATURE CELL BIOLOGY VOLUME 6 | NUMBER 10 | OCTOBER 2004
acetylation of recombinant p53 at Lys 382 was assessed. The acetylaseactivity in the Strap immunocomplex was significantly greater in DNA-damaged cells than normal cells (Fig. 4c). In contrast, the StrapS203A
immunocomplex lacked p53 acetylase activity (Fig. 4d).
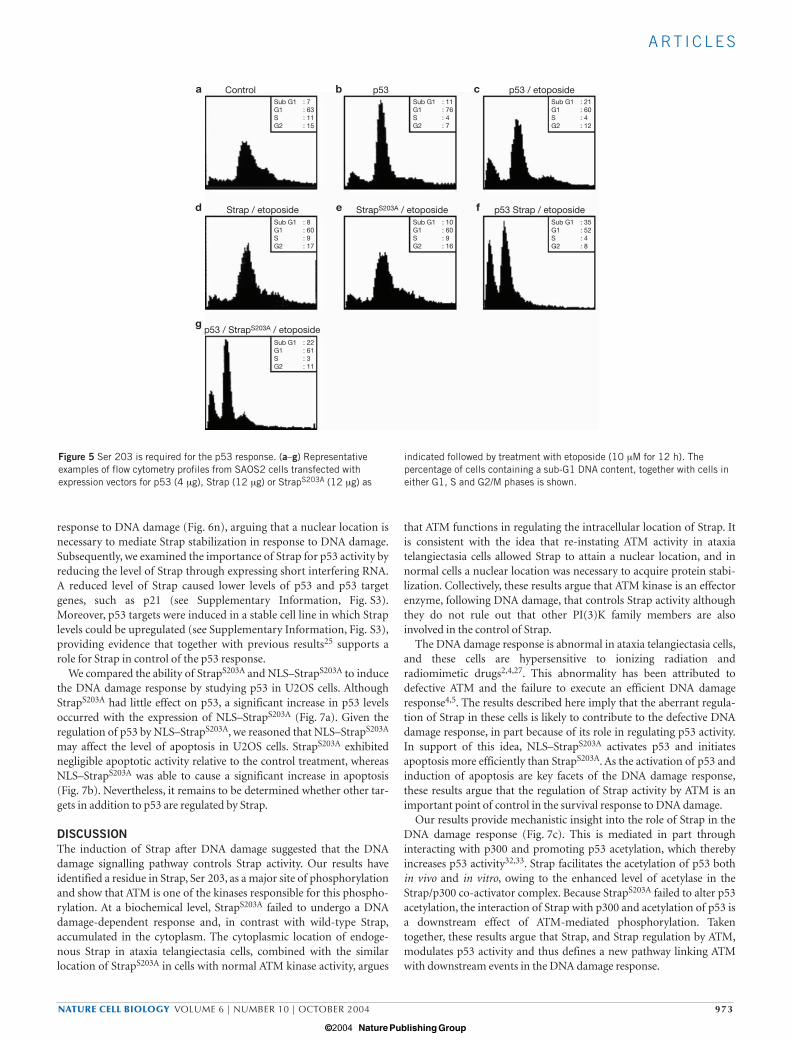
Phosphorylation of Ser 203 is required for apoptosis in responseto DNA damageIt was important to assess the function of Ser 203 in regulating theDNA damage response. Thus, we compared the ability of wild-typeStrap and StrapS203A to enhance p53-dependent apoptosis inresponse to DNA damage. In SAOS2 cells (which lack p53 activity),the introduction of exogenous p53 followed by treatment with etopo-side caused significant cell-cycle delay, increasing the G1 populationand the level of cells undergoing apoptosis (sub-G1) (Fig. 5c). Thepresence of wild-type Strap significantly increased the level of apop-totic cells (Fig. 5f; 21% compared with 35%), whereas in the absence
of p53 Strap had minimal effect (Fig. 5d). In sharp contrast,StrapS203A failed to augment apoptosis (Fig. 5e, g). These results sup-port the idea that Ser 203 phosphorylation by ATM kinase is a func-tionally important event in the DNA damage response.
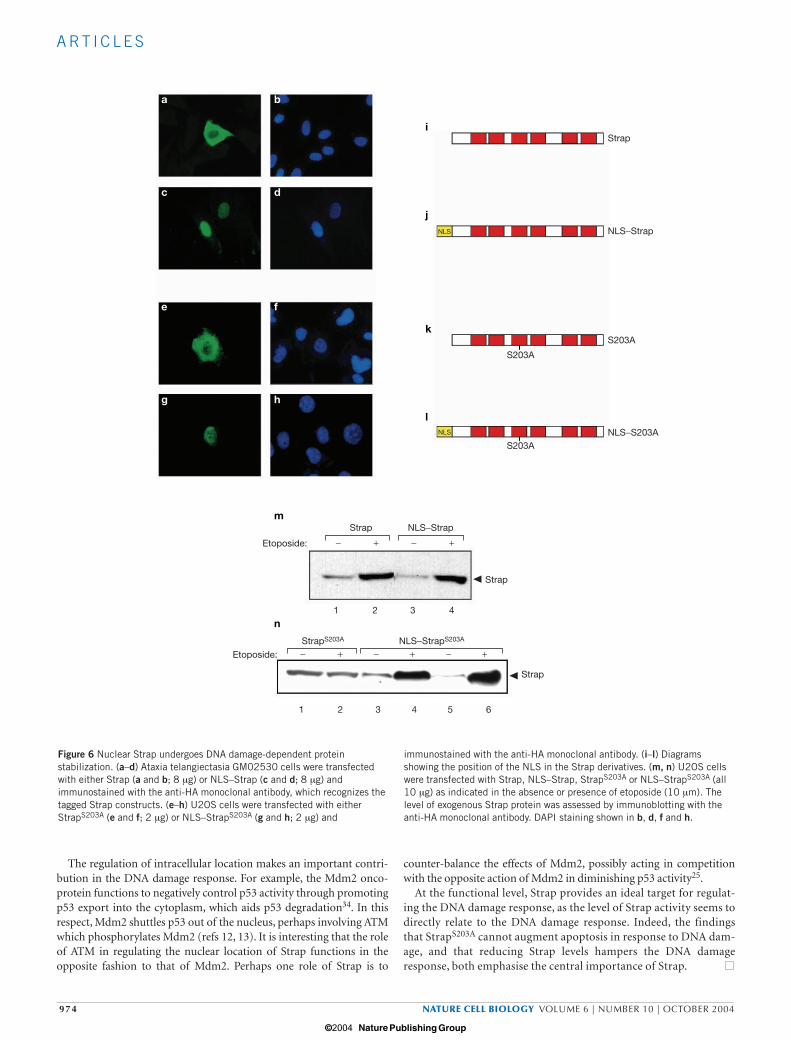
Targeting Strap to the nucleus activates the DNA damage responseTo determine whether the nuclear accumulation of Strap is sufficient toinduce the DNA damage response, we targeted Strap to nuclei by attach-ing a nuclear localization signal (NLS) to both wild-type Strap andStrapS203A (Fig. 6i–l). In ataxia telangiectasia cells, NLS–Strap accumu-lated in nuclei in contrast to the behaviour of wild-type Strap (Fig. 6a–d).Also, in U2OS cells NLS–StrapS203A localized to the nucleus, in contrastwith StrapS203A which remained in the cytoplasm (Fig. 6e–h).
As expected from the earlier results, StrapS203A failed to stabilizeunder DNA damage conditions (Fig. 6m, n). However, targetingStrapS203A to nuclei with NLS–StrapS203A re-instated its stabilization in
a
c
b
+ −− WT S203A − WT S203A− WT S203A
+ −
+ − + − + −
+ − + − + − + − +− +− +−Strap: Strap:
Etoposide: Etoposide:
Input IP: anti-p300 / IB: anti-Strap (HA)
1 2 3 4 5 6
1 2 3 4 5 6
1 2 3 4 5 6
7 8 9 10 11 12
p300
Strap
IP: anti-p53IB: anti-Ac Lys 382
IB: anti-Ac Lys 382
IB: anti-p53
IB: anti-p300
Actin
p53 BSA p53
anti-Strap anti-GAL4
WT Strap
Ac Lys 382-p53
StrapS203A
d
Figure 4 Strap Ser 203 regulates the interaction between Strap and p300and the acetylation of p53. (a) Lysates prepared from U2OS cellstransfected with expression vectors encoding wild-type Strap (15 µg; lanes3, 4, 9 and 10) or StrapS203A (15 µg; lanes 5, 6, 11 and 12) and p300 (10µg; lanes 1–12) and treated with or without etoposide (10 µM for 12 h)were immunoprecipitated with anti-p300 monoclonal antibody, and eitherimmunoblotted with the same antibody (top) or anti-HA polyclonal Y-11antibody for exogenous Strap (bottom). Lanes 1 to 6 show the input level ofp300 (top) and Strap (bottom), and Lanes 7 to 12 show the level of theimmunoprecipitated p300–Strap complex. (b) Lysates prepared from U2OScells transfected with expression vectors encoding wild-type Strap (10 µg;lanes 3 and 4), StrapS203A (10 µg; lanes 5 and 6) or empty vector (10 µg;lanes 1 and 2) were treated with etoposide as above, immunoprecipitated
with anti-p53 DO-1 antibody, and then immunoblotted with anti-acetylatedp53 Lys 382 (top). Input extracts immunoblotted with anti-Ac Lys 382,DO-1, anti-p300 monoclonal antibody or actin are shown. (c, d) Lysatesprepared from SAOS2 (p53−/−) cells transfected with expression vectorsencoding wild-type Strap (10 µg, c) or StrapS203A (10 µg, d) either pre orpost etoposide treatment (10 µM for 12 h) were immunoprecipitated withthe anti-HA monoclonal antibody (lanes 1, 2, 3 and 4) or a control anti-Gal4 antibody (lanes 5 and 6). The level of HAT activity in theimmunocomplex was measured in vitro using His-tagged wild-type p53 (1µg) as the substrate. The acetylation of p53 was assessed byimmunoblotting with an antibody recognizing acetylated Lys 382, a site onp53 which is known to be acetylated in damaged cells by p300 (ref. 22).BSA (1 µg) served as a control substrate.
print ncb1170 14/9/04 2:58 PM Page 972
© 2004 Nature Publishing Group
© 2004 Nature Publishing Group
A RT I C L E S
NATURE CELL BIOLOGY VOLUME 6 | NUMBER 10 | OCTOBER 2004 973
response to DNA damage (Fig. 6n), arguing that a nuclear location isnecessary to mediate Strap stabilization in response to DNA damage.Subsequently, we examined the importance of Strap for p53 activity byreducing the level of Strap through expressing short interfering RNA.A reduced level of Strap caused lower levels of p53 and p53 targetgenes, such as p21 (see Supplementary Information, Fig. S3).Moreover, p53 targets were induced in a stable cell line in which Straplevels could be upregulated (see Supplementary Information, Fig. S3),providing evidence that together with previous results25 supports arole for Strap in control of the p53 response.
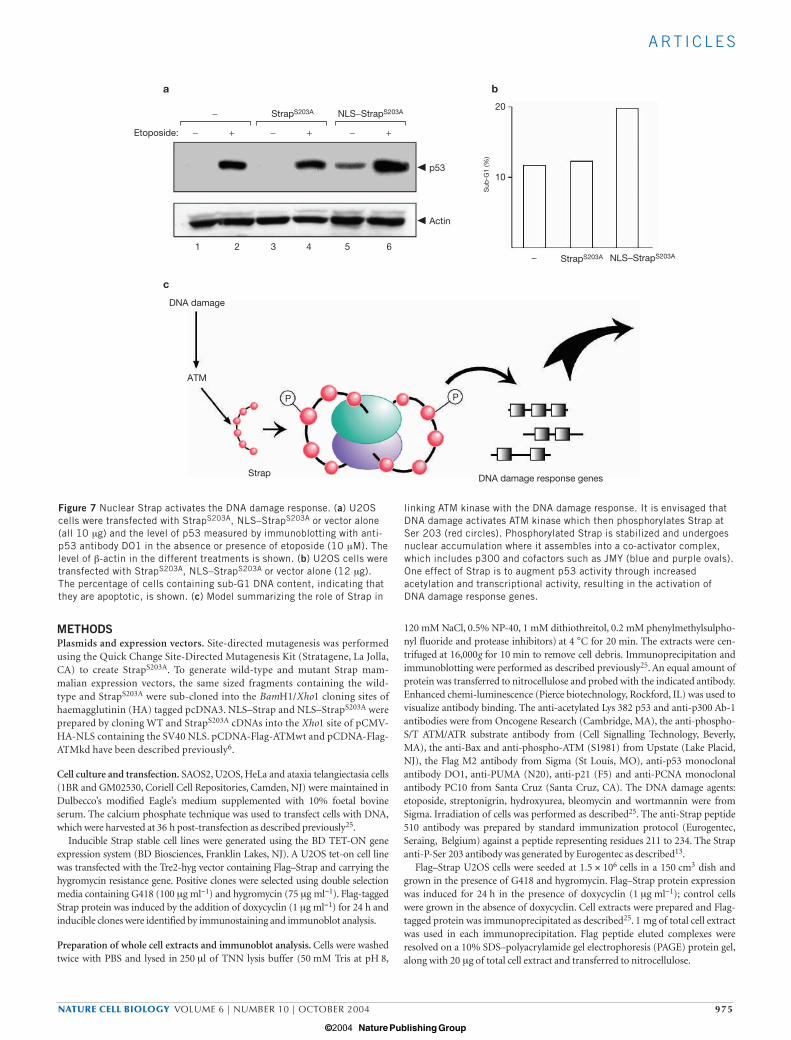
We compared the ability of StrapS203A and NLS–StrapS203A to inducethe DNA damage response by studying p53 in U2OS cells. AlthoughStrapS203A had little effect on p53, a significant increase in p53 levelsoccurred with the expression of NLS–StrapS203A (Fig. 7a). Given theregulation of p53 by NLS–StrapS203A, we reasoned that NLS–StrapS203A
may affect the level of apoptosis in U2OS cells. StrapS203A exhibitednegligible apoptotic activity relative to the control treatment, whereasNLS–StrapS203A was able to cause a significant increase in apoptosis(Fig. 7b). Nevertheless, it remains to be determined whether other tar-gets in addition to p53 are regulated by Strap.
DISCUSSIONThe induction of Strap after DNA damage suggested that the DNAdamage signalling pathway controls Strap activity. Our results haveidentified a residue in Strap, Ser 203, as a major site of phosphorylationand show that ATM is one of the kinases responsible for this phospho-rylation. At a biochemical level, StrapS203A failed to undergo a DNAdamage-dependent response and, in contrast with wild-type Strap,accumulated in the cytoplasm. The cytoplasmic location of endoge-nous Strap in ataxia telangiectasia cells, combined with the similarlocation of StrapS203A in cells with normal ATM kinase activity, argues
that ATM functions in regulating the intracellular location of Strap. Itis consistent with the idea that re-instating ATM activity in ataxiatelangiectasia cells allowed Strap to attain a nuclear location, and innormal cells a nuclear location was necessary to acquire protein stabi-lization. Collectively, these results argue that ATM kinase is an effectorenzyme, following DNA damage, that controls Strap activity althoughthey do not rule out that other PI(3)K family members are alsoinvolved in the control of Strap.
The DNA damage response is abnormal in ataxia telangiectasia cells,and these cells are hypersensitive to ionizing radiation andradiomimetic drugs2,4,27. This abnormality has been attributed todefective ATM and the failure to execute an efficient DNA damageresponse4,5. The results described here imply that the aberrant regula-tion of Strap in these cells is likely to contribute to the defective DNAdamage response, in part because of its role in regulating p53 activity.In support of this idea, NLS–StrapS203A activates p53 and initiatesapoptosis more efficiently than StrapS203A. As the activation of p53 andinduction of apoptosis are key facets of the DNA damage response,these results argue that the regulation of Strap activity by ATM is animportant point of control in the survival response to DNA damage.
Our results provide mechanistic insight into the role of Strap in theDNA damage response (Fig. 7c). This is mediated in part throughinteracting with p300 and promoting p53 acetylation, which therebyincreases p53 activity32,33. Strap facilitates the acetylation of p53 bothin vivo and in vitro, owing to the enhanced level of acetylase in theStrap/p300 co-activator complex. Because StrapS203A failed to alter p53acetylation, the interaction of Strap with p300 and acetylation of p53 isa downstream effect of ATM-mediated phosphorylation. Takentogether, these results argue that Strap, and Strap regulation by ATM,modulates p53 activity and thus defines a new pathway linking ATMwith downstream events in the DNA damage response.
a b c
d
g
e f
Control p53 p53 / etoposide
Strap / etoposide
p53 / StrapS203A / etoposide
StrapS203A / etoposide p53 Strap / etoposide
Sub G1 : 7G1 : 63S : 11G2 : 15
Sub G1 : 11G1 : 76S : 4G2 : 7
Sub G1 : 21G1 : 60S : 4G2 : 12
Sub G1 : 8G1 : 60S : 9G2 : 17
Sub G1 : 22G1 : 61S : 3G2 : 11
Sub G1 : 10G1 : 60S : 9G2 : 16
Sub G1 : 35G1 : 52S : 4G2 : 8
Figure 5 Ser 203 is required for the p53 response. (a–g) Representativeexamples of flow cytometry profiles from SAOS2 cells transfected withexpression vectors for p53 (4 µg), Strap (12 µg) or StrapS203A (12 µg) as
indicated followed by treatment with etoposide (10 µM for 12 h). Thepercentage of cells containing a sub-G1 DNA content, together with cells ineither G1, S and G2/M phases is shown.
print ncb1170 14/9/04 2:58 PM Page 973
© 2004 Nature Publishing Group
© 2004 Nature Publishing Group
A RT I C L E S
974 NATURE CELL BIOLOGY VOLUME 6 | NUMBER 10 | OCTOBER 2004
The regulation of intracellular location makes an important contri-bution in the DNA damage response. For example, the Mdm2 onco-protein functions to negatively control p53 activity through promotingp53 export into the cytoplasm, which aids p53 degradation34. In thisrespect, Mdm2 shuttles p53 out of the nucleus, perhaps involving ATMwhich phosphorylates Mdm2 (refs 12, 13). It is interesting that the roleof ATM in regulating the nuclear location of Strap functions in theopposite fashion to that of Mdm2. Perhaps one role of Strap is to
counter-balance the effects of Mdm2, possibly acting in competitionwith the opposite action of Mdm2 in diminishing p53 activity25.
At the functional level, Strap provides an ideal target for regulat-ing the DNA damage response, as the level of Strap activity seems todirectly relate to the DNA damage response. Indeed, the findingsthat StrapS203A cannot augment apoptosis in response to DNA dam-age, and that reducing Strap levels hampers the DNA damageresponse, both emphasise the central importance of Strap.
a b
i
j
k
l
c d
e f
g
m
h
Strap
NLS−Strap
NLS−S203A
S203A
S203A
S203A
− + − +Strap
− + − + − +StrapS203A NLS−StrapS203A
NLS−Strap
Etoposide:
Etoposide:
Strap
Strap
1 2 3 4
1 2 3 4 5 6
NLS
NLS
n
Figure 6 Nuclear Strap undergoes DNA damage-dependent proteinstabilization. (a–d) Ataxia telangiectasia GM02530 cells were transfectedwith either Strap (a and b; 8 µg) or NLS–Strap (c and d; 8 µg) andimmunostained with the anti-HA monoclonal antibody, which recognizes thetagged Strap constructs. (e–h) U2OS cells were transfected with eitherStrapS203A (e and f; 2 µg) or NLS–StrapS203A (g and h; 2 µg) and
immunostained with the anti-HA monoclonal antibody. (i–l) Diagramsshowing the position of the NLS in the Strap derivatives. (m, n) U2OS cellswere transfected with Strap, NLS–Strap, StrapS203A or NLS–StrapS203A (all10 µg) as indicated in the absence or presence of etoposide (10 µm). Thelevel of exogenous Strap protein was assessed by immunoblotting with theanti-HA monoclonal antibody. DAPI staining shown in b, d, f and h.
print ncb1170 14/9/04 2:58 PM Page 974
© 2004 Nature Publishing Group
© 2004 Nature Publishing Group
A RT I C L E S
NATURE CELL BIOLOGY VOLUME 6 | NUMBER 10 | OCTOBER 2004 975
METHODSPlasmids and expression vectors. Site-directed mutagenesis was performedusing the Quick Change Site-Directed Mutagenesis Kit (Stratagene, La Jolla,CA) to create StrapS203A. To generate wild-type and mutant Strap mam-malian expression vectors, the same sized fragments containing the wild-type and StrapS203A were sub-cloned into the BamH1/Xho1 cloning sites ofhaemagglutinin (HA) tagged pcDNA3. NLS–Strap and NLS–StrapS203A wereprepared by cloning WT and StrapS203A cDNAs into the Xho1 site of pCMV-HA-NLS containing the SV40 NLS. pCDNA-Flag-ATMwt and pCDNA-Flag-ATMkd have been described previously6.
Cell culture and transfection. SAOS2, U2OS, HeLa and ataxia telangiectasia cells(1BR and GM02530, Coriell Cell Repositories, Camden, NJ) were maintained inDulbecco’s modified Eagle’s medium supplemented with 10% foetal bovineserum. The calcium phosphate technique was used to transfect cells with DNA,which were harvested at 36 h post-transfection as described previously25.
Inducible Strap stable cell lines were generated using the BD TET-ON geneexpression system (BD Biosciences, Franklin Lakes, NJ). A U2OS tet-on cell linewas transfected with the Tre2-hyg vector containing Flag–Strap and carrying thehygromycin resistance gene. Positive clones were selected using double selectionmedia containing G418 (100 µg ml−1) and hygromycin (75 µg ml−1). Flag-taggedStrap protein was induced by the addition of doxycyclin (1 µg ml−1) for 24 h andinducible clones were identified by immunostaining and immunoblot analysis.
Preparation of whole cell extracts and immunoblot analysis. Cells were washedtwice with PBS and lysed in 250 µl of TNN lysis buffer (50 mM Tris at pH 8,
120 mM NaCl, 0.5% NP-40, 1 mM dithiothreitol, 0.2 mM phenylmethylsulpho-nyl fluoride and protease inhibitors) at 4 °C for 20 min. The extracts were cen-trifuged at 16,000g for 10 min to remove cell debris. Immunoprecipitation andimmunoblotting were performed as described previously25. An equal amount ofprotein was transferred to nitrocellulose and probed with the indicated antibody.Enhanced chemi-luminescence (Pierce biotechnology, Rockford, IL) was used tovisualize antibody binding. The anti-acetylated Lys 382 p53 and anti-p300 Ab-1antibodies were from Oncogene Research (Cambridge, MA), the anti-phospho-S/T ATM/ATR substrate antibody from (Cell Signalling Technology, Beverly,MA), the anti-Bax and anti-phospho-ATM (S1981) from Upstate (Lake Placid,NJ), the Flag M2 antibody from Sigma (St Louis, MO), anti-p53 monoclonalantibody DO1, anti-PUMA (N20), anti-p21 (F5) and anti-PCNA monoclonalantibody PC10 from Santa Cruz (Santa Cruz, CA). The DNA damage agents:etoposide, streptonigrin, hydroxyurea, bleomycin and wortmannin were fromSigma. Irradiation of cells was performed as described25. The anti-Strap peptide510 antibody was prepared by standard immunization protocol (Eurogentec,Seraing, Belgium) against a peptide representing residues 211 to 234. The Strapanti-P-Ser 203 antibody was generated by Eurogentec as described13.
Flag–Strap U2OS cells were seeded at 1.5 × 106 cells in a 150 cm3 dish andgrown in the presence of G418 and hygromycin. Flag–Strap protein expressionwas induced for 24 h in the presence of doxycyclin (1 µg ml−1); control cellswere grown in the absence of doxycyclin. Cell extracts were prepared and Flag-tagged protein was immunoprecipitated as described25. 1 mg of total cell extractwas used in each immunoprecipitation. Flag peptide eluted complexes wereresolved on a 10% SDS–polyacrylamide gel electrophoresis (PAGE) protein gel,along with 20 µg of total cell extract and transferred to nitrocellulose.
−
−
−
StrapS203A
StrapS203A
NLS−StrapS203A
NLS−StrapS203A
+ − + − +Etoposide:
p53
Actin
1 2 3 4 5 6
a
c
b
20
10
DNA damage
ATM
Strap
P P
DNA damage response genesS
ub-G
1 (%
)
Figure 7 Nuclear Strap activates the DNA damage response. (a) U2OScells were transfected with StrapS203A, NLS–StrapS203A or vector alone(all 10 µg) and the level of p53 measured by immunoblotting with anti-p53 antibody DO1 in the absence or presence of etoposide (10 µM). Thelevel of β-actin in the different treatments is shown. (b) U2OS cells weretransfected with StrapS203A, NLS–StrapS203A or vector alone (12 µg).The percentage of cells containing sub-G1 DNA content, indicating thatthey are apoptotic, is shown. (c) Model summarizing the role of Strap in
linking ATM kinase with the DNA damage response. It is envisaged thatDNA damage activates ATM kinase which then phosphorylates Strap atSer 203 (red circles). Phosphorylated Strap is stabilized and undergoesnuclear accumulation where it assembles into a co-activator complex,which includes p300 and cofactors such as JMY (blue and purple ovals).One effect of Strap is to augment p53 activity through increasedacetylation and transcriptional activity, resulting in the activation ofDNA damage response genes.
print ncb1170 14/9/04 2:58 PM Page 975
© 2004 Nature Publishing Group
© 2004 Nature Publishing Group

A RT I C L E S
976 NATURE CELL BIOLOGY VOLUME 6 | NUMBER 10 | OCTOBER 2004
For the ATM kinase immunoprecipitation, HeLa cells were lysed in TNNbuffer and ATM immunoprecipitated with protein A agarose beads (25 µl) andanti-ATM Ab-3 antibody (Oncogene Research Products, Cambridge, MA).Kinase reactions were performed using precipitated ATM on beads (~15 µl)with either His-tagged Strap, the His-tagged StapS203A mutant or His-taggedp53 (ref. 35) purified from bacteria (~1 µg protein) in a kinase buffer (50 mMHepes at pH 7.4, 150 mM NaCl, 6 mM MgCl2, 4 mM MnCl2, 10% glycerol, 1mM dithiothreitol, 0.1 mM NaOV and 15 µCi of [γ-32P] ATP] and incubated at30 °C for 30 min. Proteins were separated by SDS–PAGE, transferred to nitro-cellulose membranes and visualized by autoradiography.
Acetyl transferase assays. HA-tagged Strap was transfected into SAOS2 cells andimmunoprecipitated using anti-HA monoclonal antibody (Babco, Berkeley, CA).Strap immunocomplexes were dissolved in HAT assay buffer (50 mM Tris at pH8.0, 10% glycerol, 50 mM KCl, 0.1 mM EDTA, 10 mM butyric acid, 1 mM dithio-threitol and 10 µM acetyl-CoA) and incubated with bacterially expressed recombi-nant His-tagged p53 protein. The reaction mixtures were resolved on a SDS–PAGEgel. Proteins were transferred onto nitrocellulose, which was then subjected toimmunoblot analysis and probed with the anti-acetylated Lys 382 p53 antibody.
Immunofluorescence microscopy. Cells were plated on to glass coverslips andtreated as described. Cells were fixed in 4% paraformaldehyde for 30 min, per-meabilized with 0.1% Triton X-100 for 10 min, washed with PBS, blocked with10% foetal calf serum in PBS for 10 min and then washed again with PBS. Theprimary antibodies used were the anti-Strap peptide 510 antibody (detectingendogenous protein, diluted 1 : 200) and the monoclonal anti-HA antibody (forexogenous Strap and StrapS203A, diluted 1 : 1000). After washing with PBS, thesamples were incubated with FITC conjugated goat anti-mouse IgG (diluted 1 :200) and stained with DAPI (0.1 µg m−1) for 30 min. The samples were exam-ined using a fluorescent microscope (Olympus, Melville, NY).
siRNA expression. To generate the pSUPER-Strap knock down mammalianexpression vectors, four different short oligonucleotide fragments wererespectively subcloned into the HindIII/BglII cloning sites of pSUPER36.pSUPER-Strap1 and 4 were found to be functional in reducing Strap proteinexpression. The forward sequence of pSUPER-Strap1 was 5′-GATCCCCAA-G AT G C A G G A C G G A A G C AT T C A A G AT G C T T C C G T C C T G -CATCTTTTTTTGGAAA-3′ and the reverse 5′-AGCTTTTCCAAAAAAAGATGCAGGACGGAAGCATCTCTTGAATGCTTCCGTCCTG-CATCTTGGG-3′. The forward sequence of pSUPER-Strap4 was 5 ′ - G A T C C C C A A G A G A T G G A G A A G A C C C T T T C A A G A -GAAGGGTCTTCTCCATCTCTTTTTTTGGAAA-3′ and the reverse 5 ′ - A G C T T T T C C A A A A A A A G A G A T G G A G A A G A CCCTTCTCTTGAAAGGGTCTTCTCCATCTCTTGGG-3′.
Flow cytometry. U2OS cells were transfected with the indicated expression vec-tors, together with pCMV CD20 (4 µg) to monitor transfection efficiency as pre-viously described25. The profiles of the transfected population of cells are shown.
Note: Supplementary Information is available on the Nature Cell Biology website.
ACKNOWLEDGEMENTSWe thank M. Caldwell for assistance in preparing the manuscript. This work wassupported by the Medical Research Council, the Leukaemia Research Fund, CancerResearch UK and the European Union. We thank M. B. Kastan for the wild-typeand kinase-dead ATM expression vectors.
COMPETING FINANCIAL INTERESTSThe authors declare that they have no competing financial interests.
Received 18 February 2004; accepted 3 August 2004Published online at http://www.nature.com/naturecellbiology.
1. Zhou, B. B. & Elledge, S. J. The DNA damage response: putting checkpoints in per-spective. Nature 408, 433–439 (2000).
2. Abraham, R. T. Cell cycle checkpoint signalling through the ATM and ATR kinases.Genes Dev. 15, 2177–2196 (2001).
3. Khanna, K. K., Lavin, M. F., Jackson, S. P. & Mulhern, T. D. ATM, a central controllerof cellular responses to DNA damage. Cell Death Differ. 8, 1052–1065 (2001).
4. Shiloh, Y. ATM and ATR: networking cellular responses to DNA damage. Curr. Opin.Genet. Dev. 11, 71–77 (2001).
5. Walworth, N. C. Cell-cycle checkpoint kinases: checking in on the cell cycle. Curr.Opin. Cell Biol. 12, 697–704 (2000).
6. Canman, C. E. et al. Activation of the ATM kinase by ionizing radiation and phosphory-lation of p53. Science 281, 1677–1679 (1998).
7. Khosravi, R. et al. Rapid ATM-dependent phosphorylation of MDM2 precedes p53accumulation in response to DNA damage. Proc. Natl Acad. Sci. USA 96,14973–14977 (1999).
8. Maya, R. et al. ATM-dependent phosphorylation of Mdm2 on serine 395: role in p53activation by DNA damage. Genes Dev. 15, 1067–1077 (2001).
9. Cortez, D., Wang, Y., Qin, J. & Elledge, S. J. Requirement of ATM-dependent phospho-rylation of BRCA1 in the DNA damage response to double-strand breaks. Science286, 1162–1166 (1999).
10. Hirao, A. et al. DNA damage-induced activation of p53 by the checkpoint kinaseChk2. Science 287, 1824–1827 (2000).
11. Shieh, S. Y., Ahn, J., Tamai, K., Taya, Y. & Prives, C. The human homologues of check-point kinases Chk1 and Cds1 (Chk2) phosphorylate p53 at multiple DNA damage-inducible sites. Genes Dev. 14, 289–300 (2000).
12. Falck, J., Mailand, N., Syljuasen, R. G., Bartek, J. & Lukas, J. TheATM–Chk2–Cdc25A checkpoint pathway guards against radioresistant DNA synthesis.Nature 410, 842–847 (2001).
13. Stevens, C., Smith, L. & La Thangue, N. B. Checkpoint kinase activates E2F-1 inresponse to DNA damage. Nature Cell Biol. 5, 401–409 (2003).
14. Peng, C. Y. et al. Mitotic and G2 checkpoint control: Regulation of 14-3-3 protein bind-ing by phosphorylation of Cdc25C on serine-216. Science 277, 1501–1505 (1997).
15. Lakin, N. D. & Jackson, S. P. Regulation of p53 in response to DNA damage.Oncogene 18, 7644–7655 (1999).
16. Hollstein, M., Sidransky, D., Vogelstein, B. & Harris, C. C. p53 mutations in humancancers. Science 253, 49–53 (1991).
17. Chehab, N. H., Malikzay, A., Appel, M. & Halazonetis, T. D. Chk2/hCds1 functions asa DNA damage checkpoint in G1 by stabilizing p53. Genes Dev. 14, 278–288 (2000).
18. Giaccia, A. J. & Kastan, M. B. The complexity of p53 modulation: emerging patternsfrom divergent signals. Genes Dev. 12, 2973–2983 (1998).
19. Meyn, M. S. Ataxia-telangiectasia, cancer and the pathobiology of the ATM gene. Clin.Genet. 55, 289–304 (1999).
20. Bell, D. W. et al. Heterozygous germ line hCHK2 mutations in Li-Fraumeni syndrome.Science 286, 2528–2531 (1999).
21. Sakaguchi, K. et al. DNA damage activates p53 through a phosphorylation–acetylationcascade. Genes Dev. 12, 2831–2841 (1998).
22. Liu, L. et al. p53 sites acetylated in vitro by PCAF and p300 are acetylated in vivo inresponse to DNA damage. Mol. Cell. Biol. 19, 1202–1209 (1999).
23. Avantaggiati, M. L. et al. Recruitment of p300/CBP in p53-dependent signal path-ways. Cell 89, 1175–1184 (1997).
24. Shikama, N. et al. A novel cofactor for p300 that regulates the p53 response. Mol.Cell 4, 365–376 (1999).
25. Demonacos, C., Krstic-Demonacos, M. & La Thangue, N. B. A TPR motif cofactor con-tributes to p300 activity in the p53 response. Mol. Cell 8, 71–84 (2001).
26. Bakkenist, C. J. & Kastan, M. B. DNA damage activates ATM through intramolecularautophosphorylation and dimer dissociation. Nature 421, 499–506 (2003).
27. Barlow, C. et al. Loss of the ataxia-telangiectasia gene product causes oxidative dam-age in target organs. Proc. Natl. Acad. Sci. USA 96, 9915–9919 (1999).
28. Beamish, H. & Lavin, M. F. Radiosensitivity in ataxia-telangiectasia: anomalies in radi-ation-induced cell cycle delay. Int. J. Radiat. Biol. 65, 175–184 (1994).
29. Sarkaria, J. N. et al. Inhibition of phosphoinositide 3-kinase related kinases by theradiosensitizing agent wortmannin. Cancer Res. 58, 4375–4382 (1998).
30. Stein, R. C. & Waterfield, M. D. PI3-kinase inhibition: a target for drug development?Mol. Med. Today 6, 347–357 (2000).
31. Di Tullio, R. A. et al. 53BP1 functions in an ATM-dependent checkpoint pathway thatis constitutively activated in human cancer. Nature Cell Biol. 4, 998–1002 (2002).
32. Chan, H- M. & La Thangue, N. B. p300/CBP proteins: HATs for transcriptional bridgesand scaffolds. J. Cell Sci. 114, 2363–2373 (2001).
33. Gu, W. & Roeder, R. G. Activation of p53 sequence specific DNA binding by acetyla-tion of the p53 C-terminal domain. Cell 90, 595–606 (1997).
34. Freedman, D. A. & Levine, A. J. Nuclear export is required for degradation of endogenousp53 by MDM2 and human papillomavirus E6. Mol. Cell. Biol. 18, 7288–7293 (1998).
35. Crighton, D. et al. p53 represses RNA polymerase III transcription by targeting TBPand inhibiting promoter occupancy by TFIIIB. EMBO J. 22, 2810–2820 (2003).
36. Brummelkamp, T. R., Bernards, R. & Agami, R. A system for stable expression of shortinterfering RNAs in mammalian cells. Science 296, 550–553 (2002).
print ncb1170 14/9/04 2:58 PM Page 976
© 2004 Nature Publishing Group
© 2004 Nature Publishing Group

S U P P L E M E N TA RY I N F O R M AT I O N
WWW.NATURE.COM/NATURECELLBIOLOGY 1
Figure S1 a) Strap is an intrinsically unstable protein: U2OS cells were transfected with the expression vector encoding wild-type Strap (10µg) and after 24h extracts harvested from either untreated or treated (cyclohexamide; 10mg/ml) cells at the indicated time points and immunoblotted with anti-HA monoclonal antibody. b) Phosphorylation of Strap and S203A in U2OS and AT cells: Lysates from U20S cells (tracks 1 to 6) transfected and treated as described in (Fig. 1d) were immunoprecipitated with an anti-HA monoclonal antibody followed by immunoblotting with anti-phospho S/T ATM/ATR substrate-specific antibody (top) or the anti-Strap 510 antibody (bottom).
AT 1BR cells (tracks 7 to 10) transfected with the wild-type (WT) Strap (10µg; track 3 and 4) or empty vector (10µg; track 1 and 2) were treated with etoposide (+; 10µM for 12 h) and immunoprecipitated with an anti-HA monoclonal antibody followed by immunoblotting with the anti-ATM/ATR phospho S/T substrate specific antibody (top) or anti-Strap 510 antibody (bottom). c) Strategy for preparing the Strap anti-P-S203 anti-peptide antibody: Strap anti-P-S203 was prepared as described (13). Results from a typical Elisa assay (bottom) representing activity against either the P-S203 or S203 peptide is shown.
© 2004 Nature Publishing Group

S U P P L E M E N TA RY I N F O R M AT I O N
2 WWW.NATURE.COM/NATURECELLBIOLOGY
Figure S2 Phosphorylation of Strap at S203. a) Flag-Strap U2OS cells were either induced (tracks 1, 3, 4, 5 and 6) or not induced (tracks 2, 7, 8, 9 and 10) by doxycyclin and extracts prepared as described. Flag antibody immunoprecipitates were eluted with Flag peptide and immunocomplexes immunoblotted with either anti-Flag (tracks 6 and 10) or the anti-phospho S203 peptide antibody (tracks 3, 4, 5, 7, 8 and 9) in the presence of the phospho S203 peptide ((P); tracks 4 and 8) or non-phosphorylated peptide (+; tracks 3 and 7), or in the absence of peptide (-; tracks 5 and 9). Tracks 1 and 2 represent the input extracts from either induced (+) or un-induced (-) Flag-Strap U2OS cells. b) U2OS cells were treated with etoposide (10µM
for 12h), harvested as described and total cell extracts immunoblotted with anti-Strap 510 antibody (track 1) or anti-phospho S203 peptide antibody (tracks 2, 3 and 4) in the presence of non-phosphorylated peptide (+; track 3) or phospho S203 peptide (P; track 4). c) U2OS cells were treated with ionising radiation (10Gy) and harvested together with control treated cells after 1h, and immunoblotted with anti-phospho S203 peptide antibody (top) or anti-α-tubulin (bottom) as described. d) U2OS cells were treated with etoposide (10µM) or bleomycin (5µg/ml) for the indicated times, harvested and immunoblotted with anti-phospho-ATM (S1981), to detect active ATM (37), or anti-phospho S203.
© 2004 Nature Publishing Group
S U P P L E M E N TA RY I N F O R M AT I O N
WWW.NATURE.COM/NATURECELLBIOLOGY 3
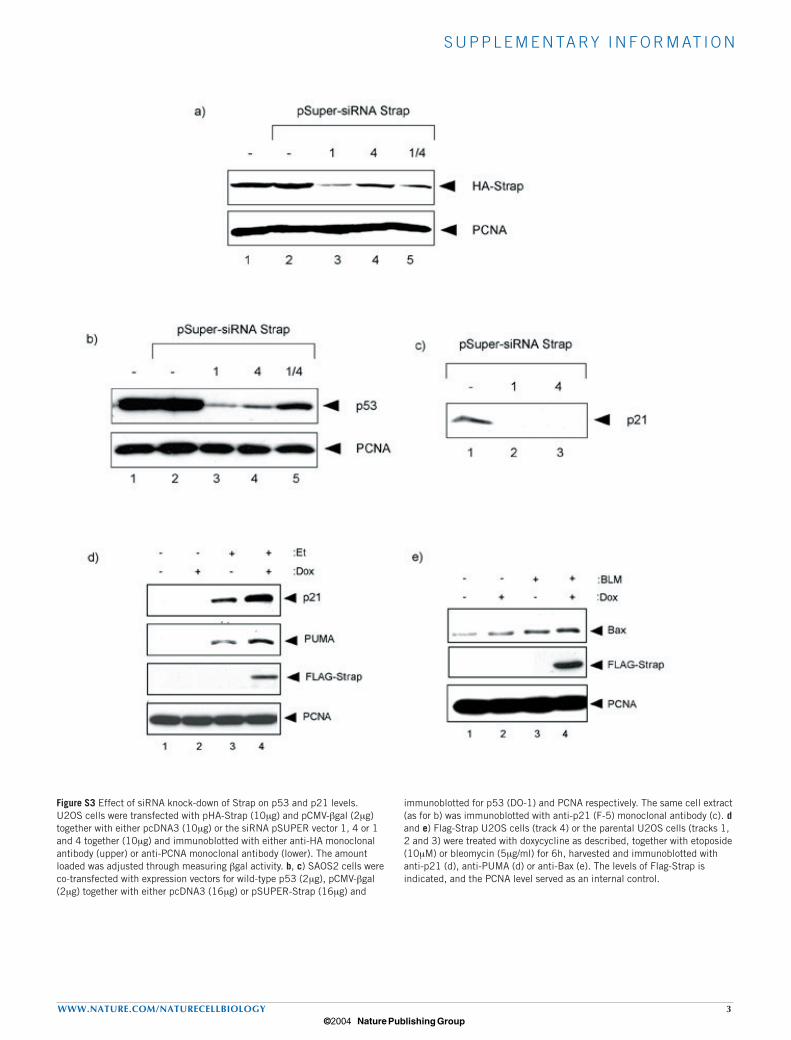
Figure S3 Effect of siRNA knock-down of Strap on p53 and p21 levels. U2OS cells were transfected with pHA-Strap (10µg) and pCMV-βgal (2µg) together with either pcDNA3 (10µg) or the siRNA pSUPER vector 1, 4 or 1 and 4 together (10µg) and immunoblotted with either anti-HA monoclonal antibody (upper) or anti-PCNA monoclonal antibody (lower). The amount loaded was adjusted through measuring βgal activity. b, c) SAOS2 cells were co-transfected with expression vectors for wild-type p53 (2µg), pCMV-βgal (2µg) together with either pcDNA3 (16µg) or pSUPER-Strap (16µg) and
immunoblotted for p53 (DO-1) and PCNA respectively. The same cell extract (as for b) was immunoblotted with anti-p21 (F-5) monoclonal antibody (c). d and e) Flag-Strap U2OS cells (track 4) or the parental U2OS cells (tracks 1, 2 and 3) were treated with doxycycline as described, together with etoposide (10µM) or bleomycin (5µg/ml) for 6h, harvested and immunoblotted with anti-p21 (d), anti-PUMA (d) or anti-Bax (e). The levels of Flag-Strap is indicated, and the PCNA level served as an internal control.
© 2004 Nature Publishing Group
S U P P L E M E N TA RY I N F O R M AT I O N
4 WWW.NATURE.COM/NATURECELLBIOLOGY
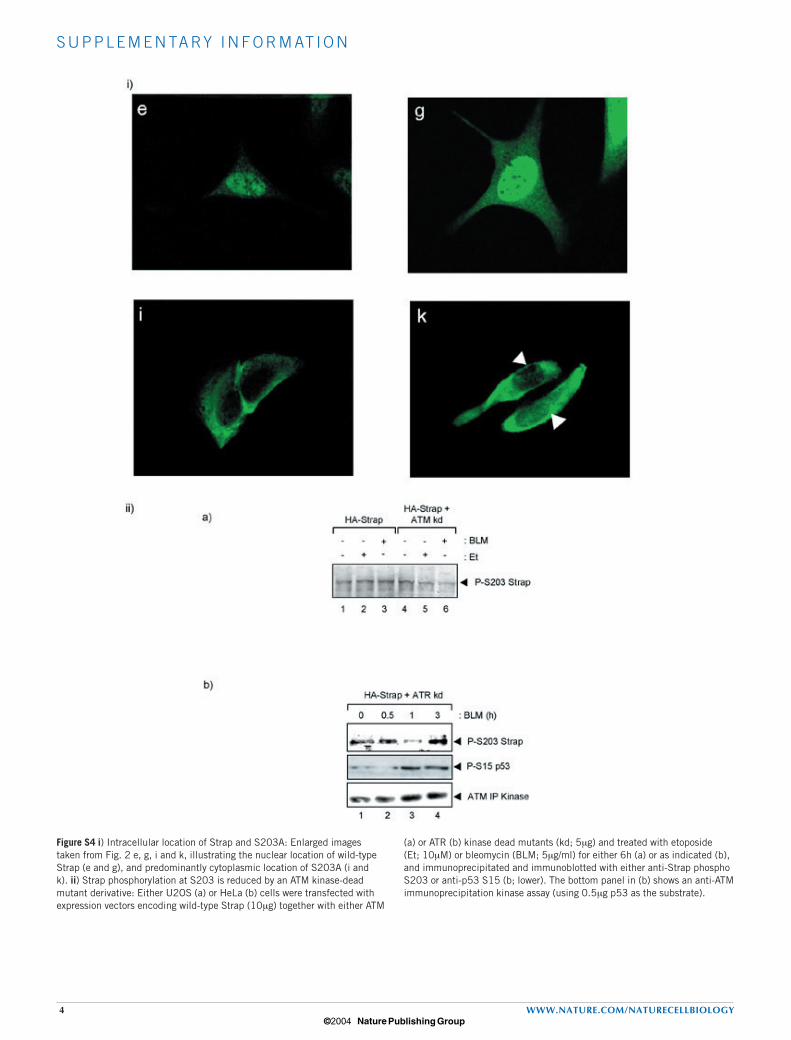
Figure S4 i) Intracellular location of Strap and S203A: Enlarged images taken from Fig. 2 e, g, i and k, illustrating the nuclear location of wild-type Strap (e and g), and predominantly cytoplasmic location of S203A (i and k). ii) Strap phosphorylation at S203 is reduced by an ATM kinase-dead mutant derivative: Either U2OS (a) or HeLa (b) cells were transfected with expression vectors encoding wild-type Strap (10µg) together with either ATM
(a) or ATR (b) kinase dead mutants (kd; 5µg) and treated with etoposide (Et; 10µM) or bleomycin (BLM; 5µg/ml) for either 6h (a) or as indicated (b), and immunoprecipitated and immunoblotted with either anti-Strap phospho S203 or anti-p53 S15 (b; lower). The bottom panel in (b) shows an anti-ATM immunoprecipitation kinase assay (using 0.5µg p53 as the substrate).
© 2004 Nature Publishing Group





























