Embed Size (px)
Citation preview

Physical Biomodeling and Foldable, Coarse-Grained
Physical Model of Polypeptide Chain
Promita ChakrabortyFeb 15, 2015
AAAS Annual Meeting 2015, San Jose
Symposium: Emerging Trends in Visualizing Physical Models and Rapid Prototyping for Biological Systems
1

2
A vision: Dynamic physical models of macromolecules that fold and convey
information

BioTableComputer interface and physical models
• BioTable is a computer monitor
• An interactive unit for translation
• Idea was to detect foldable models and molecule-molecule interaction with computers + head-mount cameras
3
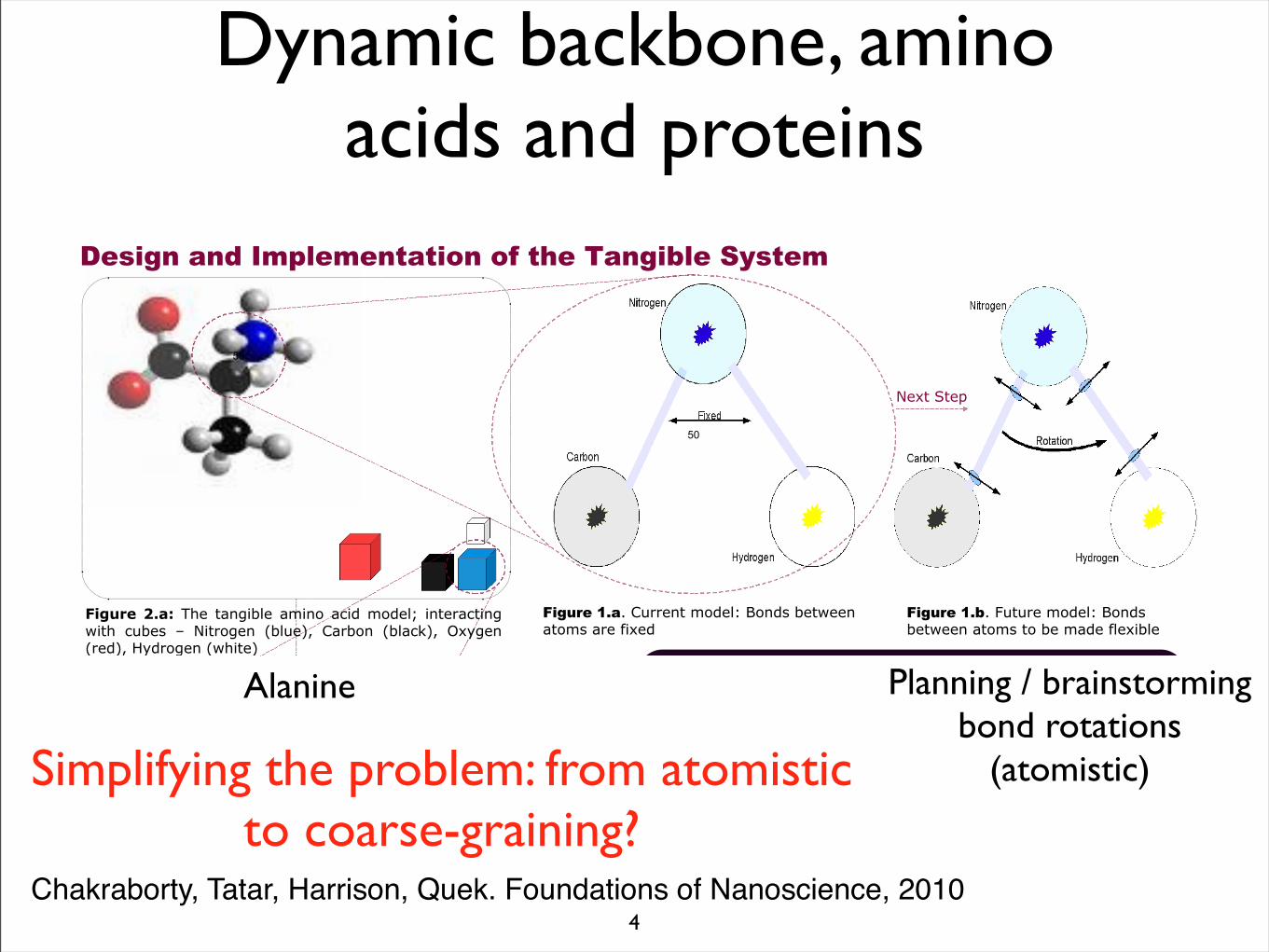
Dynamic backbone, amino acids and proteins
4
! !
"!#$%&'()*!+%,*-$.,'/*!0)$,12-3!,2!45662-,!,7*!
8%9*-:,$%9'%&!21!;<='3*%:'2%$)!+%,*-$.,'2%:!'%!"3'%2!".'9:
!"#$%&'()*'+"',#"&-.(/0,#"'*(1'&'".(2&030(4'""%5#6.(7"'68%5(9:0+!"#$%&'"(&)*+),*'#-&"%)./0"(/"1)20%30(0$)4"/51)67$/89:-%31)20%30(0$1);.<
!"#$%&'()*"$+#,-'./01+0!23
!"#$%&'()*$%+$,")-&"%.$*/+0("12"3'1)&+$"214%+$,5"'*)-&'")-*$"1$4.")-&"2+$*4"214%&%"()*)&
!"613+$,"21'"*"%&&3&'"7$%&'()*$%+$,"12"+$)&'*0)+1$"8&)9&&$"*/+$1"*0+%(
!":1)+;*)+$,"/+%%4&<(0-114"0-+4%'&$")1"4&*'$"27$%*/&$)*4"8+141,."!"#$%&'()*$%+$,"-19")-&.",'*(3"01/34&=")-17,-)(
=*&0>$&0*(
=
!" >" )*$,+84&" +$)&'2*0&" )1" -&43" ()7%&$)(" 4&*'$" )-&"()'70)7'&("12"*/+$1"*0+%("8."+$)&'*0)+$,"9+)-"*"3-.(+0*4"/1%&4"12"*/+$1"*0+%5" 21'"&=*/34&5" 4+,-)+$,"*44"?*'81$"*)1/(5"1'")170-+$,"*$"*)1/")1"4+,-)")-&"81$%("9+)-")-&"$&+,-81'+$,"*)1/(@"
!" >44" &4&0)'1$+0" %&;+0&(" *'&" &$041(&%" +$" *" 04&*'" 34*()+0"8*445"'&3'&(&$)+$,"*$"*)1/@"
!"A)7%&$)(" +$)&'*0)"9+)-")-&"(.()&/"BC+,7'&"D*E"&+)-&'")-'17,-")170-+$,")-&"*)1/("1'"8."(&3*'*)&"078&(")-*)"'&3'&(&$)" *" 3*')+074*'" &4&/&$)" <" F+)'1,&$" B847&E5"?*'81$"B84*0GE5"H=.,&$"B'&%E5"6.%'1,&$"B9-+)&E@
!"I$")-+("3'1)1).3&5")-&"*/+$1"*0+%"/14&074&"+("()*)+0"+$")-&"(&$(&")-*)")-&"81$%"*$,4&("*'&"2+=&%"BC+,7'&"J@*E@"
!"903()$(?)@'#7"'"(&$&0*()*+)&5")4$(30:7").A9&"'
!" >)/&4K(" >LLIFMJN" !?-+3"9+)-+$"&*0-"*)1/
!" ?141'&%" OPQ(" ! ")1"'&3'&(&$)" 81$%(" *$%"*)1/(
!"R-1)1(&$(1'(
B5$&C9)6"0(3);9"?
D03-%")EF$@"?7''&$)"/1%&4S"T1$%("8&)9&&$"*)1/("*'&"2+=&%
D03-%")EF:@"C7)7'&"/1%&4S"T1$%("8&)9&&$"*)1/(")1"8&"/*%&"24&=+84&
L-&" +$21'/*)+1$" 1$" &*0-"078&"'&/+$%("()7%&$)("12")-&"8*(+0" 3-.(+0*4" 3'13&')+&(" 12"&*0-"&4&/&$)"BC+,7'&"D8E@
!" I/34&/&$)+$," 24&=+84&"81$%"BC+,7'&"J@8E
!"L1"4&)")-&"81$%("'1)*)&""8*(&%" 1$" )-&" 21'0&(" )-&"*)1/("2&&4"*'17$%")-&/@"
!" 619" )1" '&3'&(&$)" *$%"+/34&/&$)" )-&" $1)+1$" 12"*))'*0)+1$"*$%"'&374(+1$U
D-&-%")B*%8
!"#$%&'()*)'Q&(+,$"12"&*0-"078&
>?
>?
F&=)"A)&3
!"#$%&'()+,'L-&" )*$,+84&" */+$1" *0+%"/1%&4V" +$)&'*0)+$,"9+)-" 078&(" W" F+)'1,&$" B847&E5" ?*'81$" B84*0GE5" H=.,&$"B'&%E5"6.%'1,&$"B9-+)&E
!"L1"87+4%"34*)21'/(5"+2"31((+84&5")1"+/+)*)&")-&"/14&074*'"(&42"*((&/84."12"3'1)&+$("*("(&&$"!"#$!$%@
@
!"L1"&$-*$0&"*$%"*7,/&$)")-&"7$%&'()*$%+$,"12"8+141,."*)")-&"27$%*/&$)*4"4&;&4"8.")-&"(0-114"0-+4%'&$@
;7&0'$&")G*$79
!" L1" -*;&" &*0-" *)1/" +$047%&" *" 0-+3" 21'"410*4" 01/37)*)+1$"*$%" +$)&'*0)"3-.(+0*44." "9+)-"+)("$&+,-81'(@
!" L1" -*;&" &*0-" 0-+3" ()1'&" +$21'/*)+1$"*817)" )-&" +//&%+*)&" $&+,-81'(" B+@&@"&4&/&$)").3&(5"*$%")-&""81$%").3&(E@
!" L1" -*;&" )-&" (.()&/" 0*4074*)&" )-&" )1)*4"21'0&" 1$" *" 3*')+074*'" *)1/" 8*(&%" 1$" )-&"*81;&"+$21'/*)+1$V")1"/&*(7'&")-&"&22&0)("12"&*0-"*)1/"1$"&*0-"1)-&'@"
H:I"/&0>"9
!" #(&" 078&(" )1" 01$()'70)" )*$,+84&"X7&'+&("*817)")-&"/1%&4@
>$" P=*/34&S" YZ-+0-" ?*'81$" *)1/("21'/"81$%("9+)-"H=.,&$"*)1/(U[
<)4$(30:7")J-"%A)
.A9&"'
&*).-##*%&)K"$%(0(3
L-&" (3*0+*4" 1'+&$)*)+1$" 12" 17'" 3-.(+0*4" /1%&4"/+,-)" -&43" )-&" Y3*'*44&4[" $*)7'&" 12" )-&"01/37)*)+1$"+$;14;&%@
Alanine Planning / brainstorming bond rotations
(atomistic)Simplifying the problem: from atomistic to coarse-graining?
Chakraborty, Tatar, Harrison, Quek. Foundations of Nanoscience, 2010

!"#$%&'()*%")+,-"),.)/%#0"/)&')1&,-,(&$#-).2'$3,'/))2/&'()&'*"+#$34")*#'(&1-")&'*"+.#$"/!
"#$%&'(!)*(+#(,$#'-.!/0,$#(*!1('(#.!2'030!4(##&5$6!
1*0!7&$1(,80!&5!(6!&6'0#9(:0!$6!(!'(,80!5;#9(:0.!(;<%06'0=!>&'*!?*-5&:(8!%(6&?;8(@305A!B0!(#0!&%?80%06@6<!&'!9$#!'0(:*&6<!'*0!!&%?$#'(6:0!$9!%$80:;8(#!5*(?05!$9!,&$8$<&:(8!06@@05!C5;:*!(5!6;:80$@=05.!'DEF!%$80:;805.!0':AG!&6!,&$:*0%&:(8!9;6:@$65A!!
1*0!#050(#:*!&6305@<('05H!!• '*0!?0=(<$<&:(8!3(8;0!$9!?*-5&:(8!30#5;5!IJ=&%065&$6(8!#0?#0506'(@$65!!• '*0!&6'0#(:@$6!,0'>006!IJ=&%065&$6(8!(6=!?*-5&:(8!#0?#0506'(@$65!• =&K0#06:05!&6!5:(80!$9!#0?#0506'(@$65!
L50#5!?*-5&:(88-!&6'0#(:'!>&'*!%(6&?;8(@305!(6=!'*0!'(,80J'$?!5;#9(:0!LM!'$!:(##-!$;'!0N?8$#(@$65!$9!$#&06'(@$6.!5&%&8(#&'-.!5*(?0!#0:$<6&@$6.!(6=!%$80:;8(#!,$6=(,&8&'-!>&'*&6!'*0!:$65'#(&6'5!$9!('$%&:!,&$:*0%&:(8!?#$:05505A!!!
1*0!#050(#:*!*-?$'*05&5H!'*0!&6'0<#(@$6!$9!5?(@(8J&%(<0#-.!'*('!&5!60:055(#-!'$!=00?8-!;6=0#5'(6=!,&$:*0%&5'#-.!&5!%$#0!0K0:@308-!(::$%?8&5*0=!'*#$;<*!0%,$=&0=!&6'0#(:@$6A!
5%#*)#+")*%")6&7"+"'$"/8)
5%9)#+")*%"/")0#&+"6)*,("*%"+8)):#')9,2)*"--).+,;)&'*2&3,'8)
<'6"+/*#'6&'()5,11-")=90,*%"/&/)
1*0!?0=(<$<&:(8!?#$,80%!&5!'$!0N'#(:'!5:&06@O:!:;#&$5&'-!>*&80!'0(:*&6<!*$>!6;:80$@=05!,$6=!>&'*!0(:*!$'*0#!'*#$;<*!*-=#$<06J,$6=&6<!('!5?0:&O:!5&'05.!*$>!'*0!(6@:$=$65!&6!'DEF!,&6=!'$!'*0!:$##05?$6=&6<!:$=$65.!*$>!'*0!>$,,80!*-?$'*05&5!>$#+!0':A!!
� �
������������ ����������� ���������� �����
��� ��������������������������� ����������������������
���������������� ���������������������������������������������������������� ������������������������������� ���������������
������������� ����������
�������������� ������������������������������ ��� ������� ����������������
�������������������������������������������������������������
������������������� ����� �������������������������������⇒������������� ��� ���������������� ��� �
��������
�
�� �� ������� �������� �� ���� ������� ����� �������������������������������������� ��� ������������������������� ������������ ��� ������������������������ �������������� � ���������� � ������ ���������� �
�� ���� ���������� �������� ��� ��������� ��� � ����� ��������������������������� �
��!������ �������� � ��������"#������$%��� ��� ���� ���� ���� ������������������������ ���������� � �������� ������� �� &������� "����%��������"���'%��(������"���%�����������"� ��%
��)�� ������������ ������������������������������ �������� � ����������������������"#������* % �
��������� �������������������������!����
�� ����+�� �,,)&-*.� ⇒� ����� ����� ���
�� �������� /01�� ⇒ ������������ ������ ������
��2 ���������
"���#����������
$�� ��%&� �������������3�4������������������������
$�� ��%&� �#����������3�4���������������������������������
, �� ���������� ��� �� ������������������������ ������� � ������ ���������� ����� ��������"#������$�%
�� )����������� ��������������"#������* �%
��,����� ������������������ ��� �� ������� ����������������� �� �
�� ���� �� ��������� ������������ �� ������ ���������������������5
$ � ��"��
���������1����������� �����
��
��
&���!��
�����������, �� ������� ����� ���������6� ����������� � ������ 7� &������� "����%�� ������ "���'%�� (������"���%�����������"� ��%
��,������������������������������������� �������������������������������������������������
��,���� ������������� �������������������������� �������������������� ���� ����� ������
��������'���
�� ,�� ��� �� � ��� �������� � � ��� ��������� ������������ ������� �������� ��� �������� ����
�� ,�� ��� �� � � ��� ����� �������������� �� �������� ���� ����� "� � ����������������� ������������%
�� ,�� ��� �� ������ ������� �� ��������� ��� � �������� ��� ����� ��� ����������������6���������� �������������� ��������� �� �� �
(�)�������
�� ���� ������ �� �������� �������8����������� �������
��� 0�����3� 9: �� � ������ ����������������� �(���������5;
���������* ��!
�!����
�� ����+�������
, �� ������ ��������� ��� ���� � ������ ��������� � ���� �� 9������;� ����� ��� �������������������
A vision: Dynamic physical models of proteins and DNA that fold
Foundations of Nanoscience, 2010
AAAS Annual Meeting, 2011
Chakraborty, Tatar, Harrison, Quek. 2009-2011
! !
"!#$%&'()*!+%,*-$.,'/*!0)$,12-3!,2!45662-,!,7*!
8%9*-:,$%9'%&!21!;<='3*%:'2%$)!+%,*-$.,'2%:!'%!"3'%2!".'9:
!"#$%&'()*'+"',#"&-.(/0,#"'*(1'&'".(2&030(4'""%5#6.(7"'68%5(9:0+!"#$%&'"(&)*+),*'#-&"%)./0"(/"1)20%30(0$)4"/51)67$/89:-%31)20%30(0$1);.<
!"#$%&'()*"$+#,-'./01+0!23
!"#$%&'()*$%+$,")-&"%.$*/+0("12"3'1)&+$"214%+$,5"'*)-&'")-*$"1$4.")-&"2+$*4"214%&%"()*)&
!"613+$,"21'"*"%&&3&'"7$%&'()*$%+$,"12"+$)&'*0)+1$"8&)9&&$"*/+$1"*0+%(
!":1)+;*)+$,"/+%%4&<(0-114"0-+4%'&$")1"4&*'$"27$%*/&$)*4"8+141,."!"#$%&'()*$%+$,"-19")-&.",'*(3"01/34&=")-17,-)(
=*&0>$&0*(
=
!" >" )*$,+84&" +$)&'2*0&" )1" -&43" ()7%&$)(" 4&*'$" )-&"()'70)7'&("12"*/+$1"*0+%("8."+$)&'*0)+$,"9+)-"*"3-.(+0*4"/1%&4"12"*/+$1"*0+%5" 21'"&=*/34&5" 4+,-)+$,"*44"?*'81$"*)1/(5"1'")170-+$,"*$"*)1/")1"4+,-)")-&"81$%("9+)-")-&"$&+,-81'+$,"*)1/(@"
!" >44" &4&0)'1$+0" %&;+0&(" *'&" &$041(&%" +$" *" 04&*'" 34*()+0"8*445"'&3'&(&$)+$,"*$"*)1/@"
!"A)7%&$)(" +$)&'*0)"9+)-")-&"(.()&/"BC+,7'&"D*E"&+)-&'")-'17,-")170-+$,")-&"*)1/("1'"8."(&3*'*)&"078&(")-*)"'&3'&(&$)" *" 3*')+074*'" &4&/&$)" <" F+)'1,&$" B847&E5"?*'81$"B84*0GE5"H=.,&$"B'&%E5"6.%'1,&$"B9-+)&E@
!"I$")-+("3'1)1).3&5")-&"*/+$1"*0+%"/14&074&"+("()*)+0"+$")-&"(&$(&")-*)")-&"81$%"*$,4&("*'&"2+=&%"BC+,7'&"J@*E@"
!"903()$(?)@'#7"'"(&$&0*()*+)&5")4$(30:7").A9&"'
!" >)/&4K(" >LLIFMJN" !?-+3"9+)-+$"&*0-"*)1/
!" ?141'&%" OPQ(" ! ")1"'&3'&(&$)" 81$%(" *$%"*)1/(
!"R-1)1(&$(1'(
B5$&C9)6"0(3);9"?
D03-%")EF$@"?7''&$)"/1%&4S"T1$%("8&)9&&$"*)1/("*'&"2+=&%
D03-%")EF:@"C7)7'&"/1%&4S"T1$%("8&)9&&$"*)1/(")1"8&"/*%&"24&=+84&
L-&" +$21'/*)+1$" 1$" &*0-"078&"'&/+$%("()7%&$)("12")-&"8*(+0" 3-.(+0*4" 3'13&')+&(" 12"&*0-"&4&/&$)"BC+,7'&"D8E@
!" I/34&/&$)+$," 24&=+84&"81$%"BC+,7'&"J@8E
!"L1"4&)")-&"81$%("'1)*)&""8*(&%" 1$" )-&" 21'0&(" )-&"*)1/("2&&4"*'17$%")-&/@"
!" 619" )1" '&3'&(&$)" *$%"+/34&/&$)" )-&" $1)+1$" 12"*))'*0)+1$"*$%"'&374(+1$U
D-&-%")B*%8
!"#$%&'()*)'Q&(+,$"12"&*0-"078&
>?
>?
F&=)"A)&3
!"#$%&'()+,'L-&" )*$,+84&" */+$1" *0+%"/1%&4V" +$)&'*0)+$,"9+)-" 078&(" W" F+)'1,&$" B847&E5" ?*'81$" B84*0GE5" H=.,&$"B'&%E5"6.%'1,&$"B9-+)&E
!"L1"87+4%"34*)21'/(5"+2"31((+84&5")1"+/+)*)&")-&"/14&074*'"(&42"*((&/84."12"3'1)&+$("*("(&&$"!"#$!$%@
@
!"L1"&$-*$0&"*$%"*7,/&$)")-&"7$%&'()*$%+$,"12"8+141,."*)")-&"27$%*/&$)*4"4&;&4"8.")-&"(0-114"0-+4%'&$@
;7&0'$&")G*$79
!" L1" -*;&" &*0-" *)1/" +$047%&" *" 0-+3" 21'"410*4" 01/37)*)+1$"*$%" +$)&'*0)"3-.(+0*44." "9+)-"+)("$&+,-81'(@
!" L1" -*;&" &*0-" 0-+3" ()1'&" +$21'/*)+1$"*817)" )-&" +//&%+*)&" $&+,-81'(" B+@&@"&4&/&$)").3&(5"*$%")-&""81$%").3&(E@
!" L1" -*;&" )-&" (.()&/" 0*4074*)&" )-&" )1)*4"21'0&" 1$" *" 3*')+074*'" *)1/" 8*(&%" 1$" )-&"*81;&"+$21'/*)+1$V")1"/&*(7'&")-&"&22&0)("12"&*0-"*)1/"1$"&*0-"1)-&'@"
H:I"/&0>"9
!" #(&" 078&(" )1" 01$()'70)" )*$,+84&"X7&'+&("*817)")-&"/1%&4@
>$" P=*/34&S" YZ-+0-" ?*'81$" *)1/("21'/"81$%("9+)-"H=.,&$"*)1/(U[
<)4$(30:7")J-"%A)
.A9&"'
&*).-##*%&)K"$%(0(3
L-&" (3*0+*4" 1'+&$)*)+1$" 12" 17'" 3-.(+0*4" /1%&4"/+,-)" -&43" )-&" Y3*'*44&4[" $*)7'&" 12" )-&"01/37)*)+1$"+$;14;&%@! !
"!#$%&'()*!+%,*-$.,'/*!0)$,12-3!,2!45662-,!,7*!
8%9*-:,$%9'%&!21!;<='3*%:'2%$)!+%,*-$.,'2%:!'%!"3'%2!".'9:
!"#$%&'()*'+"',#"&-.(/0,#"'*(1'&'".(2&030(4'""%5#6.(7"'68%5(9:0+!"#$%&'"(&)*+),*'#-&"%)./0"(/"1)20%30(0$)4"/51)67$/89:-%31)20%30(0$1);.<
!"#$%&'()*"$+#,-'./01+0!23
!"#$%&'()*$%+$,")-&"%.$*/+0("12"3'1)&+$"214%+$,5"'*)-&'")-*$"1$4.")-&"2+$*4"214%&%"()*)&
!"613+$,"21'"*"%&&3&'"7$%&'()*$%+$,"12"+$)&'*0)+1$"8&)9&&$"*/+$1"*0+%(
!":1)+;*)+$,"/+%%4&<(0-114"0-+4%'&$")1"4&*'$"27$%*/&$)*4"8+141,."!"#$%&'()*$%+$,"-19")-&.",'*(3"01/34&=")-17,-)(
=*&0>$&0*(
=
!" >" )*$,+84&" +$)&'2*0&" )1" -&43" ()7%&$)(" 4&*'$" )-&"()'70)7'&("12"*/+$1"*0+%("8."+$)&'*0)+$,"9+)-"*"3-.(+0*4"/1%&4"12"*/+$1"*0+%5" 21'"&=*/34&5" 4+,-)+$,"*44"?*'81$"*)1/(5"1'")170-+$,"*$"*)1/")1"4+,-)")-&"81$%("9+)-")-&"$&+,-81'+$,"*)1/(@"
!" >44" &4&0)'1$+0" %&;+0&(" *'&" &$041(&%" +$" *" 04&*'" 34*()+0"8*445"'&3'&(&$)+$,"*$"*)1/@"
!"A)7%&$)(" +$)&'*0)"9+)-")-&"(.()&/"BC+,7'&"D*E"&+)-&'")-'17,-")170-+$,")-&"*)1/("1'"8."(&3*'*)&"078&(")-*)"'&3'&(&$)" *" 3*')+074*'" &4&/&$)" <" F+)'1,&$" B847&E5"?*'81$"B84*0GE5"H=.,&$"B'&%E5"6.%'1,&$"B9-+)&E@
!"I$")-+("3'1)1).3&5")-&"*/+$1"*0+%"/14&074&"+("()*)+0"+$")-&"(&$(&")-*)")-&"81$%"*$,4&("*'&"2+=&%"BC+,7'&"J@*E@"
!"903()$(?)@'#7"'"(&$&0*()*+)&5")4$(30:7").A9&"'
!" >)/&4K(" >LLIFMJN" !?-+3"9+)-+$"&*0-"*)1/
!" ?141'&%" OPQ(" ! ")1"'&3'&(&$)" 81$%(" *$%"*)1/(
!"R-1)1(&$(1'(
B5$&C9)6"0(3);9"?
D03-%")EF$@"?7''&$)"/1%&4S"T1$%("8&)9&&$"*)1/("*'&"2+=&%
D03-%")EF:@"C7)7'&"/1%&4S"T1$%("8&)9&&$"*)1/(")1"8&"/*%&"24&=+84&
L-&" +$21'/*)+1$" 1$" &*0-"078&"'&/+$%("()7%&$)("12")-&"8*(+0" 3-.(+0*4" 3'13&')+&(" 12"&*0-"&4&/&$)"BC+,7'&"D8E@
!" I/34&/&$)+$," 24&=+84&"81$%"BC+,7'&"J@8E
!"L1"4&)")-&"81$%("'1)*)&""8*(&%" 1$" )-&" 21'0&(" )-&"*)1/("2&&4"*'17$%")-&/@"
!" 619" )1" '&3'&(&$)" *$%"+/34&/&$)" )-&" $1)+1$" 12"*))'*0)+1$"*$%"'&374(+1$U
D-&-%")B*%8
!"#$%&'()*)'Q&(+,$"12"&*0-"078&
>?
>?
F&=)"A)&3
!"#$%&'()+,'L-&" )*$,+84&" */+$1" *0+%"/1%&4V" +$)&'*0)+$,"9+)-" 078&(" W" F+)'1,&$" B847&E5" ?*'81$" B84*0GE5" H=.,&$"B'&%E5"6.%'1,&$"B9-+)&E
!"L1"87+4%"34*)21'/(5"+2"31((+84&5")1"+/+)*)&")-&"/14&074*'"(&42"*((&/84."12"3'1)&+$("*("(&&$"!"#$!$%@
@
!"L1"&$-*$0&"*$%"*7,/&$)")-&"7$%&'()*$%+$,"12"8+141,."*)")-&"27$%*/&$)*4"4&;&4"8.")-&"(0-114"0-+4%'&$@
;7&0'$&")G*$79
!" L1" -*;&" &*0-" *)1/" +$047%&" *" 0-+3" 21'"410*4" 01/37)*)+1$"*$%" +$)&'*0)"3-.(+0*44." "9+)-"+)("$&+,-81'(@
!" L1" -*;&" &*0-" 0-+3" ()1'&" +$21'/*)+1$"*817)" )-&" +//&%+*)&" $&+,-81'(" B+@&@"&4&/&$)").3&(5"*$%")-&""81$%").3&(E@
!" L1" -*;&" )-&" (.()&/" 0*4074*)&" )-&" )1)*4"21'0&" 1$" *" 3*')+074*'" *)1/" 8*(&%" 1$" )-&"*81;&"+$21'/*)+1$V")1"/&*(7'&")-&"&22&0)("12"&*0-"*)1/"1$"&*0-"1)-&'@"
H:I"/&0>"9
!" #(&" 078&(" )1" 01$()'70)" )*$,+84&"X7&'+&("*817)")-&"/1%&4@
>$" P=*/34&S" YZ-+0-" ?*'81$" *)1/("21'/"81$%("9+)-"H=.,&$"*)1/(U[
<)4$(30:7")J-"%A)
.A9&"'
&*).-##*%&)K"$%(0(3
L-&" (3*0+*4" 1'+&$)*)+1$" 12" 17'" 3-.(+0*4" /1%&4"/+,-)" -&43" )-&" Y3*'*44&4[" $*)7'&" 12" )-&"01/37)*)+1$"+$;14;&%@

Lessons learned for these studies
• Foldable macromolecules: Can they be built at all and folded with accuracy?
• How can models interact with computer without cumbersome designs, but retaining accuracy?
• There exists no computational/CAD platform for physical models to biocomputation platforms
6 Color-coded nucleotides
Coarse-graining

Folding with accuracy (alpha-helix and beta-sheet)
7Peppytides
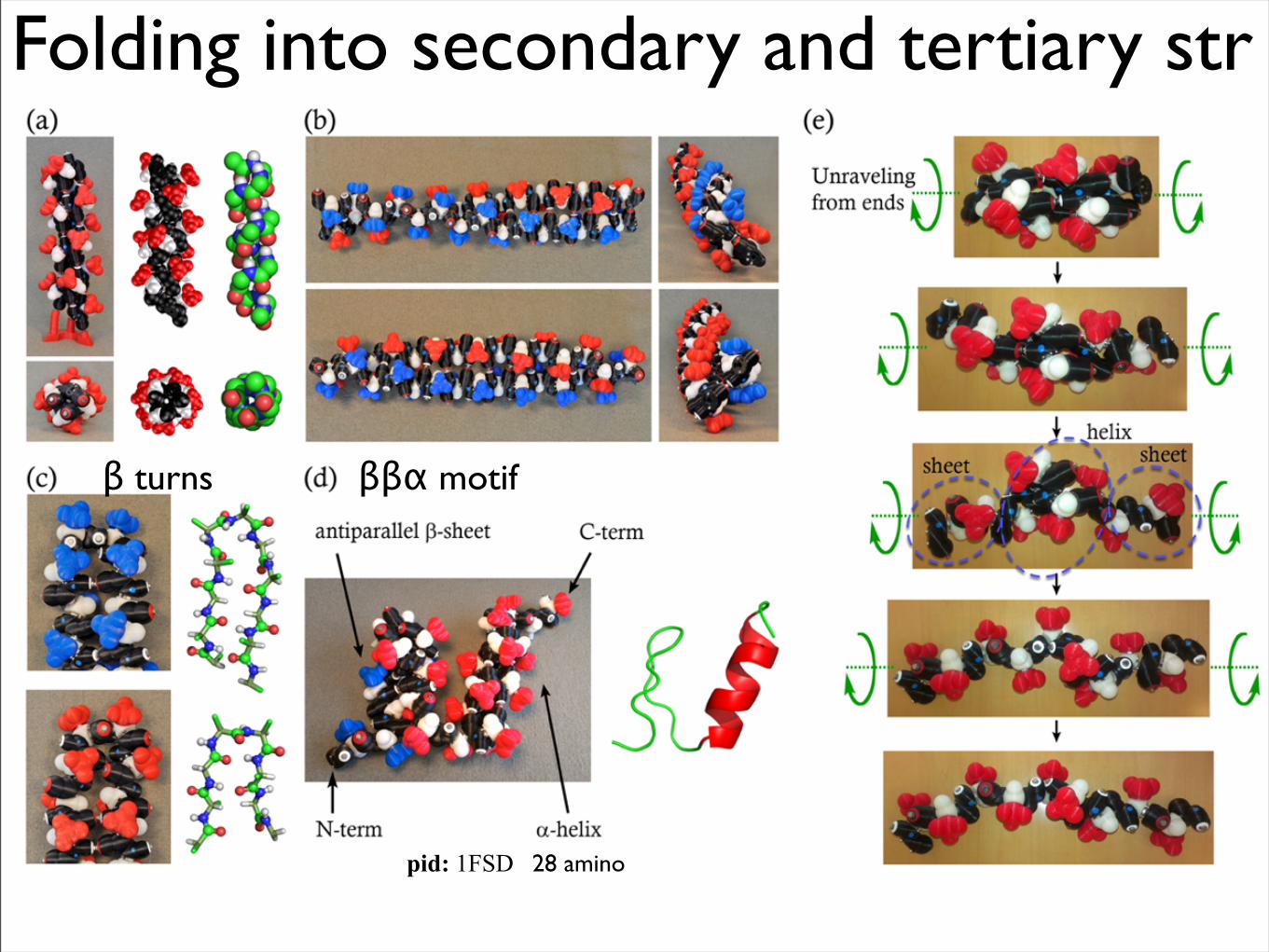
Folding into secondary and tertiary str
pid: 1FSD 28 amino
ββα motifβ turns

Beta turns
9
Type I
Type I'
Type II
Type II'
ααα - Fish Osteocalcin
pid: 1VZM, chain A 45 amino acids
N-term
C-term
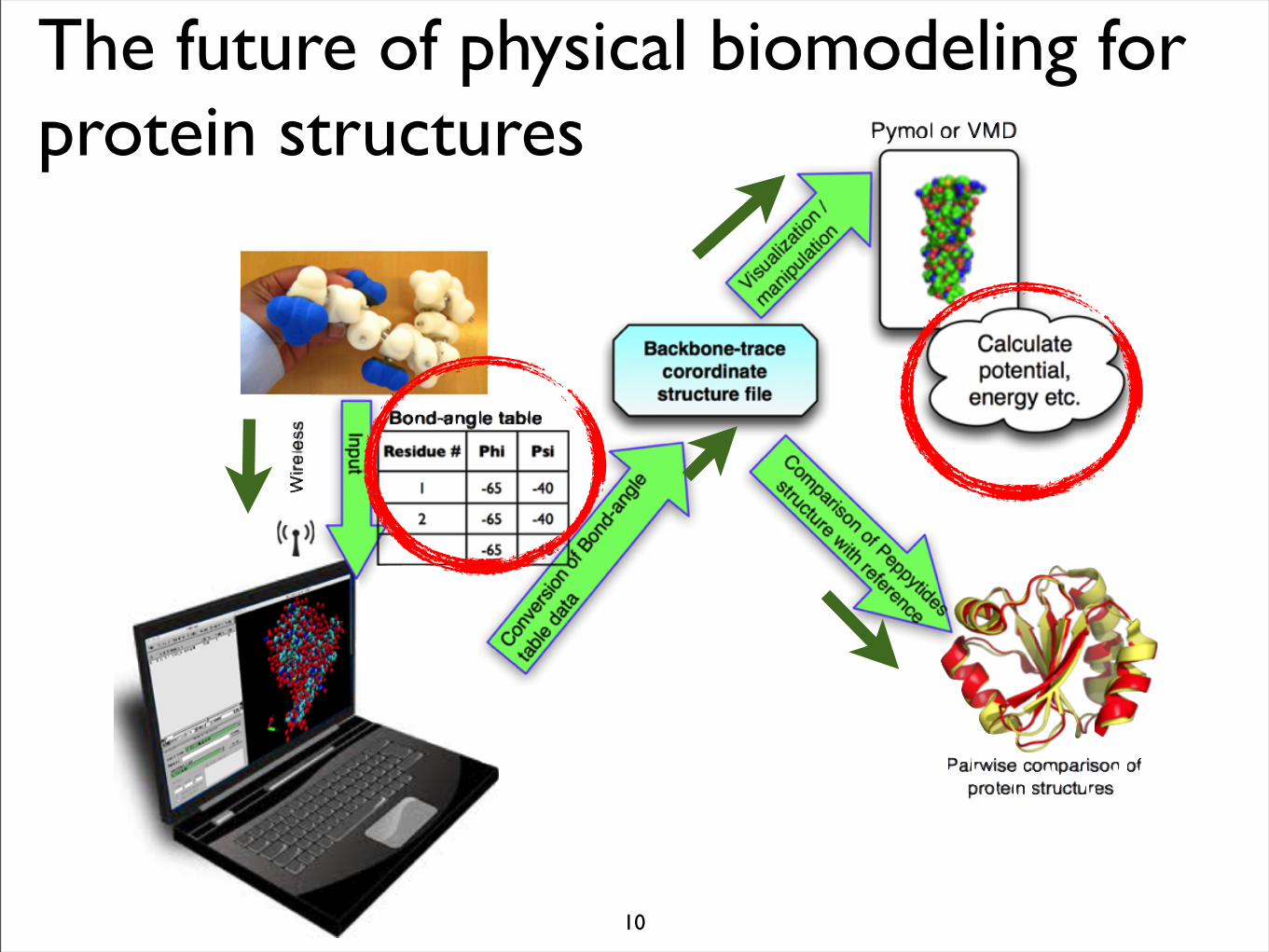
The future of physical biomodeling for protein structures
10
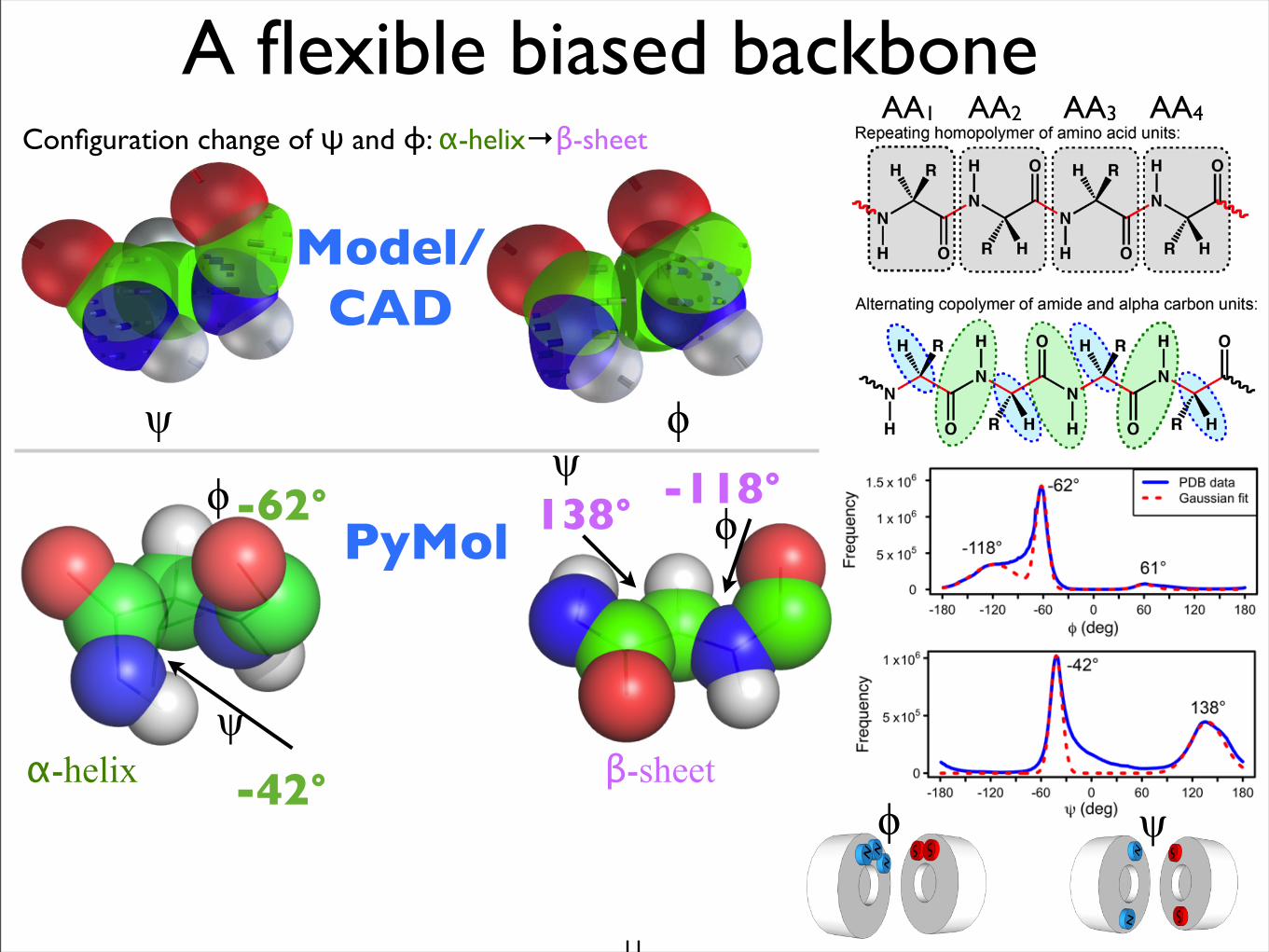
-62°
ψ
ϕ
-42°
-118°ψ
ϕ138°PyMol
Configuration change of ψ and ϕ: α-helix→β-sheet
α-helix β-sheet
ψ ϕ
11
A flexible biased backbone
Model/CAD
AA1 AA2 AA3 AA4
ϕ ψ
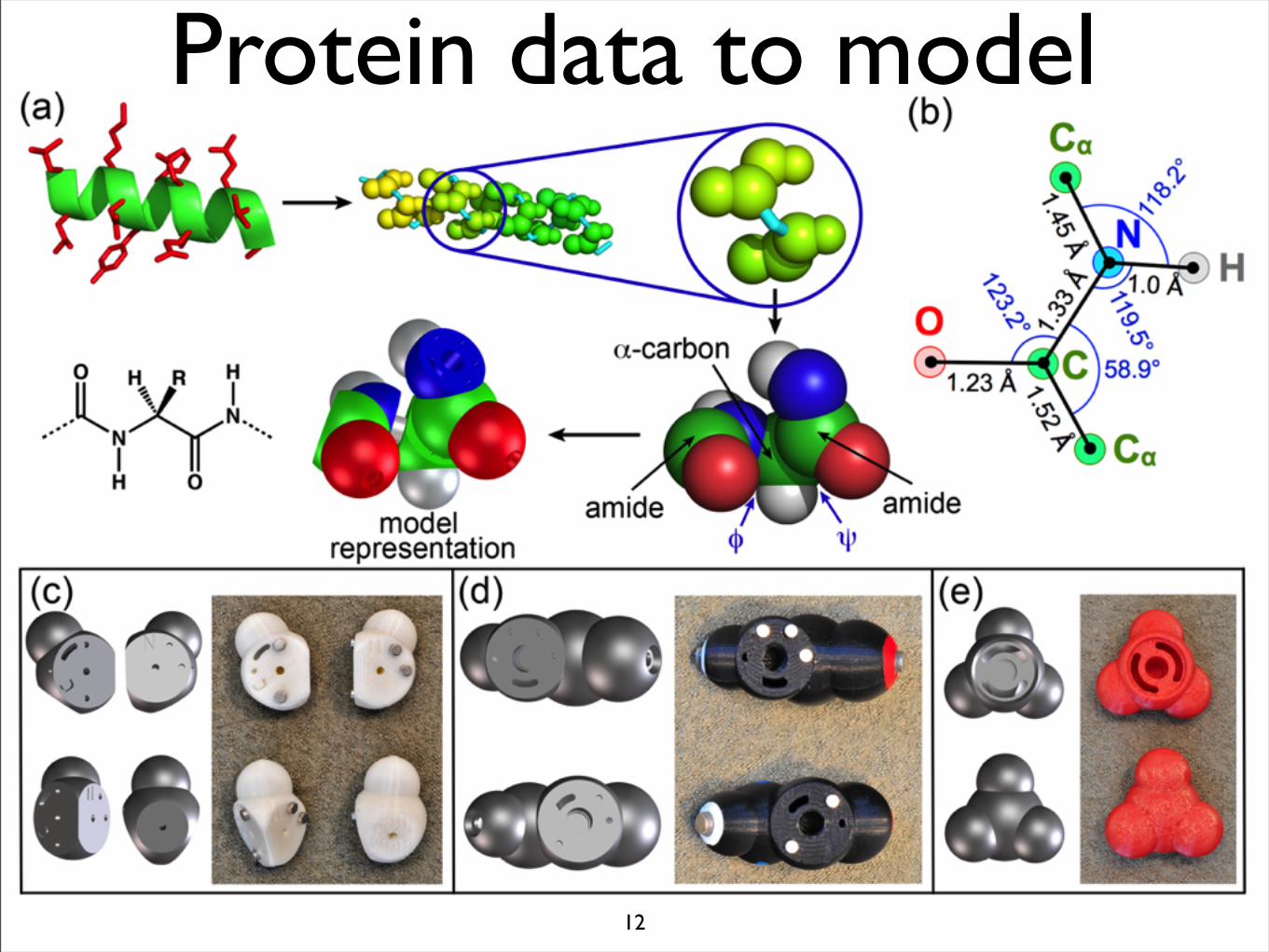
12
Protein data to model
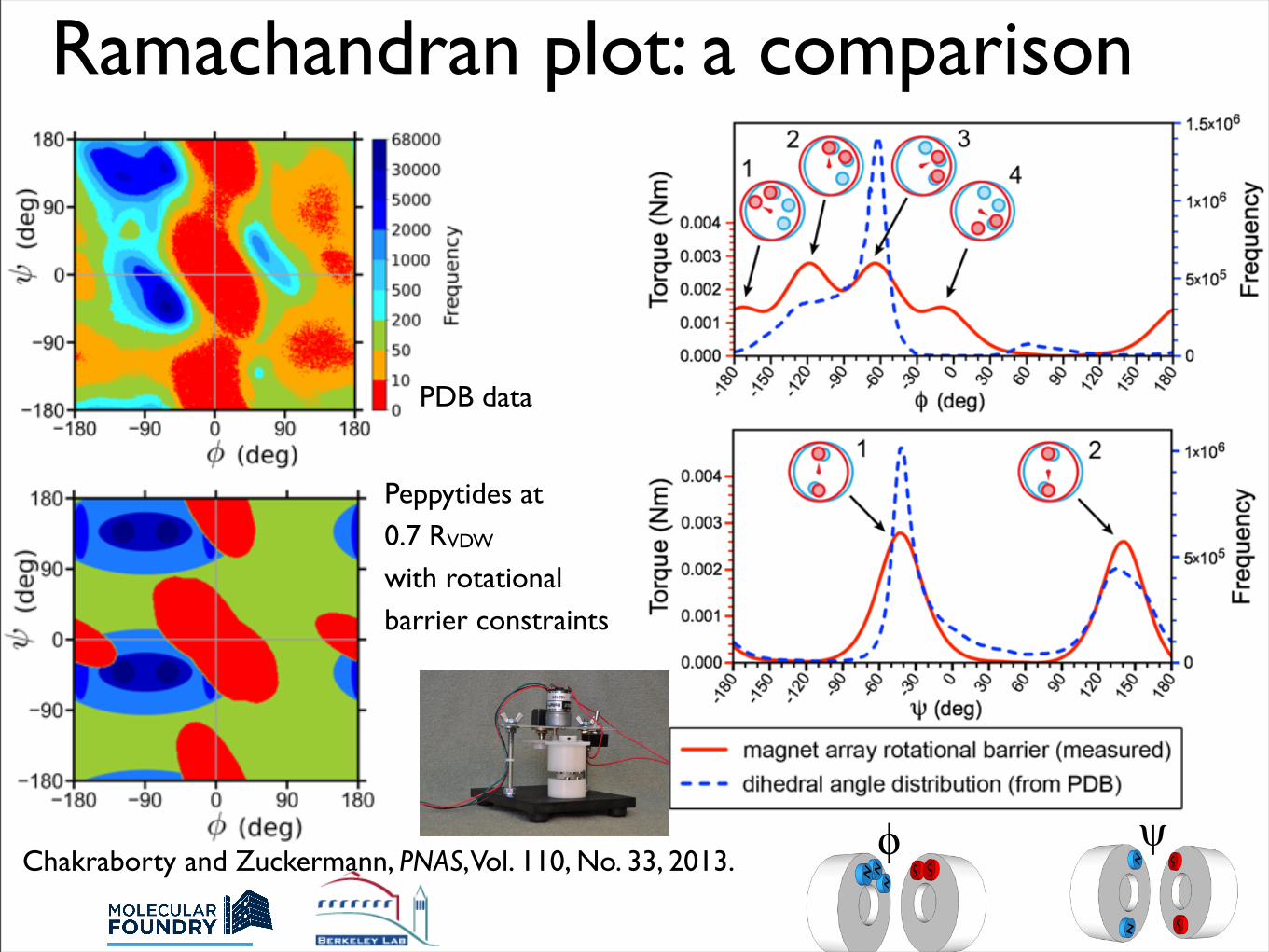
Ramachandran plot: a comparison
PDB data
Chakraborty and Zuckermann, PNAS, Vol. 110, No. 33, 2013.
Peppytides at0.7 RVDW with rotational barrier constraints
ϕ ψ
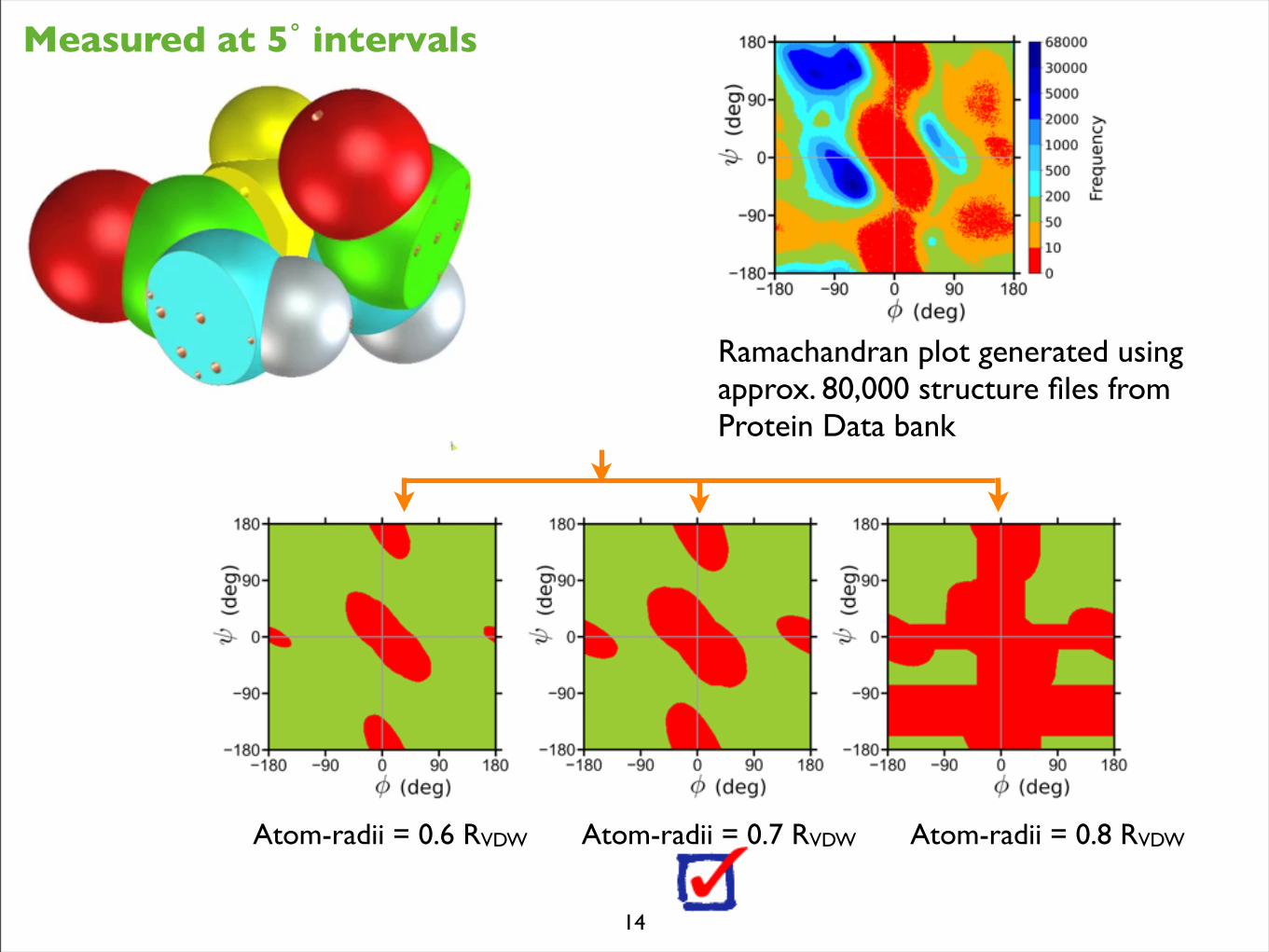
Atom-radii = 0.6 RVDW Atom-radii = 0.7 RVDW Atom-radii = 0.8 RVDW
Measured at 5˚ intervals
Ramachandran plot generated using approx. 80,000 structure files from Protein Data bank
14

O
HN
15
A scaled model: Peppytide
At Lawrence Hall of Science, UC Berkeley
93,000,000 times magnified
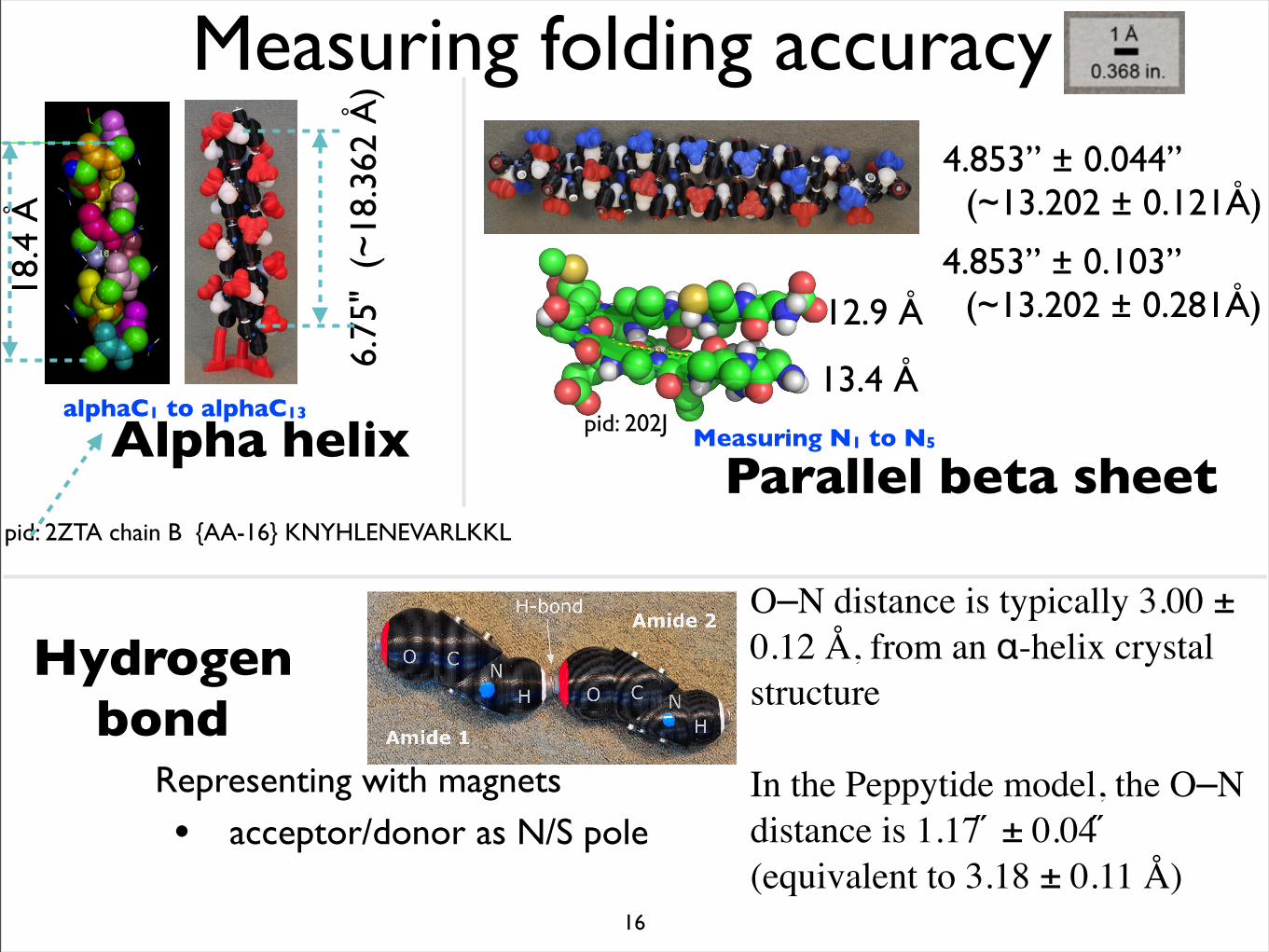
18.4
ÅMeasuring folding accuracy
6.75
" (
~18
.362
Å)
alphaC1 to alphaC13
pid: 2ZTA chain B {AA-16} KNYHLENEVARLKKL
16
Hydrogen bond
Measuring N1 to N5
13.4 Å
12.9 Å
4.853” ± 0.044” (~13.202 ± 0.121Å)
4.853” ± 0.103” (~13.202 ± 0.281Å)
pid: 202J
Representing with magnets
• acceptor/donor as N/S pole
O–N distance is typically 3.00 ± 0.12 Å, from an α-helix crystal structure
In the Peppytide model, the O–N distance is 1.17 ̋ ± 0.04 ̋ (equivalent to 3.18 ± 0.11 Å)
Alpha helixParallel beta sheet
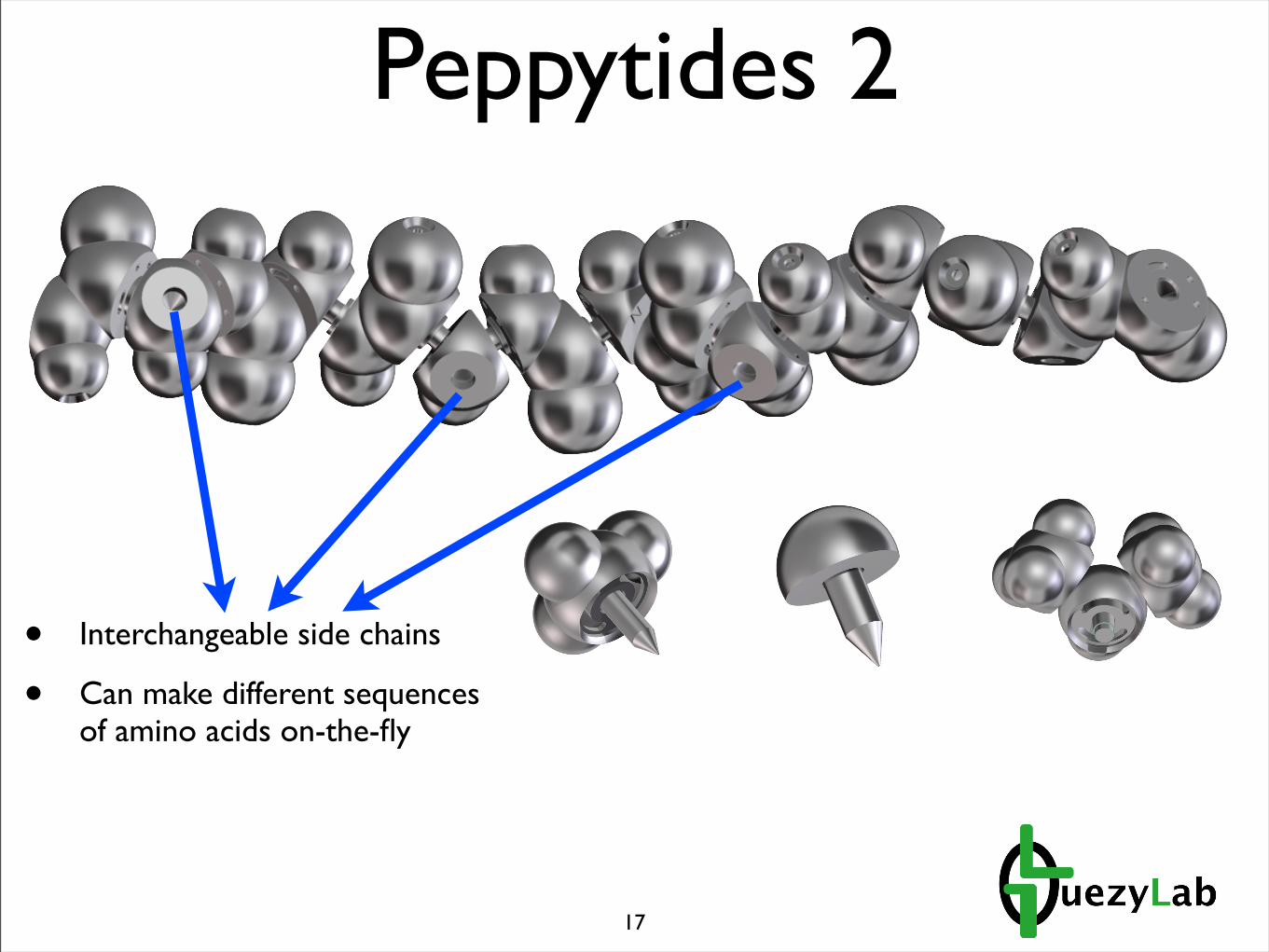
Peppytides 2
17
• Interchangeable side chains
• Can make different sequences of amino acids on-the-fly

PeppyChains
18
PeppyChains (with Ala-Val-Gly sequence)
• 3D-printed as a single unit
• No assembly required
• Without bias magnets
• next step!
• Interchangeable side chains
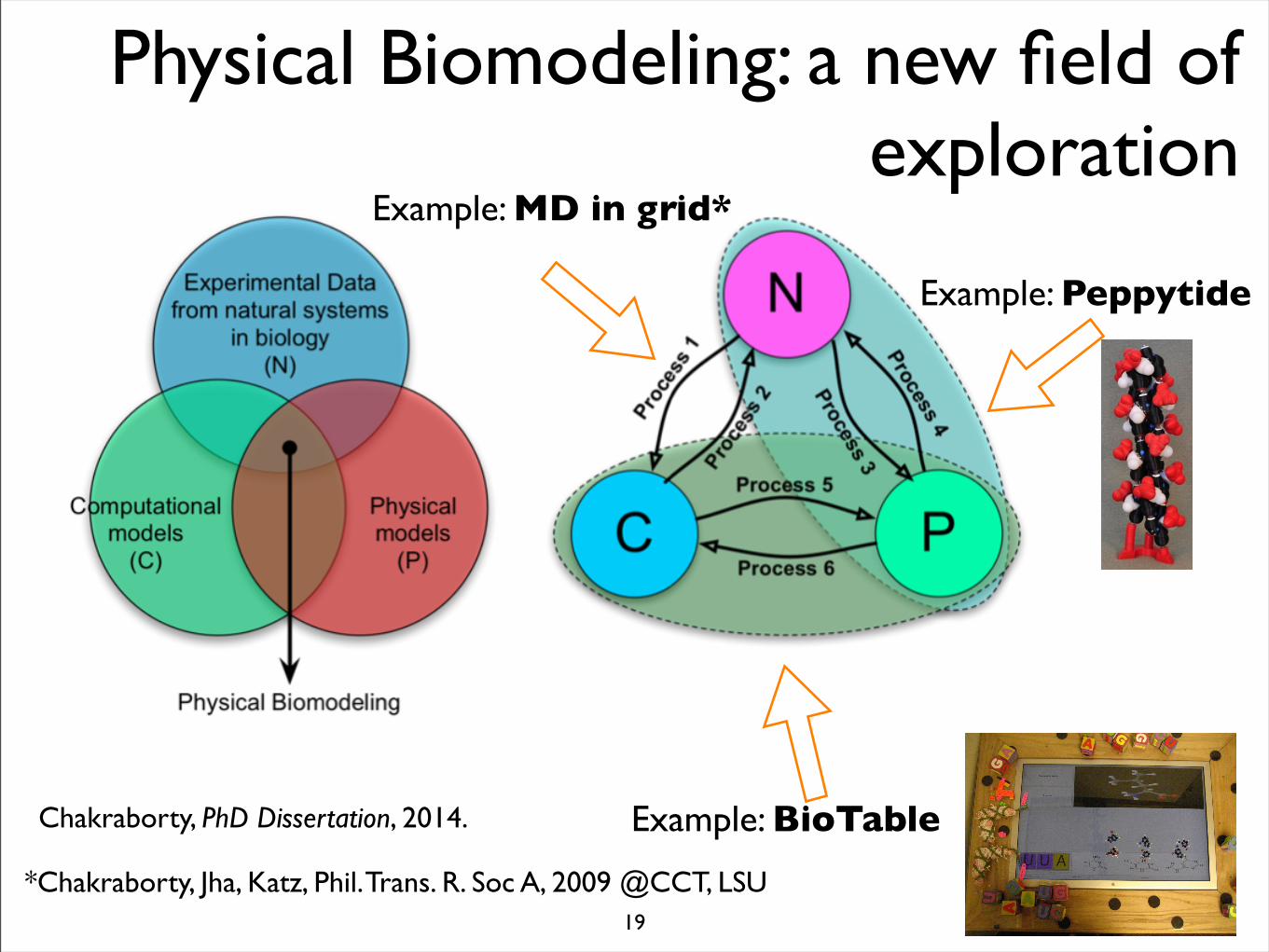
Physical Biomodeling: a new field of exploration
19
Chakraborty, PhD Dissertation, 2014. Example: BioTable
Example: Peppytide
Example: MD in grid*
*Chakraborty, Jha, Katz, Phil. Trans. R. Soc A, 2009 @CCT, LSU
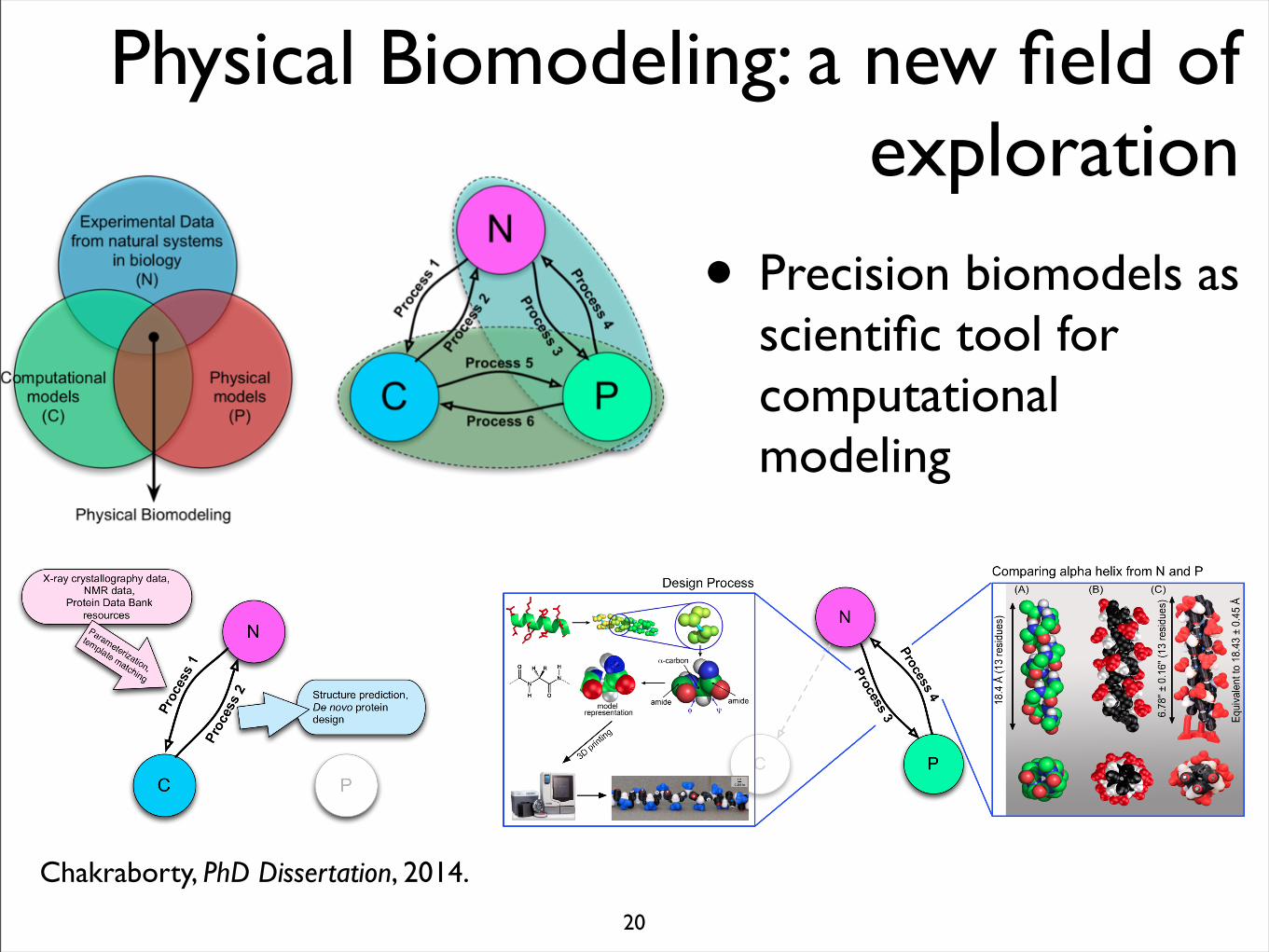
Physical Biomodeling: a new field of exploration
• Precision biomodels as scientific tool for computational modeling
20
Chakraborty, PhD Dissertation, 2014.
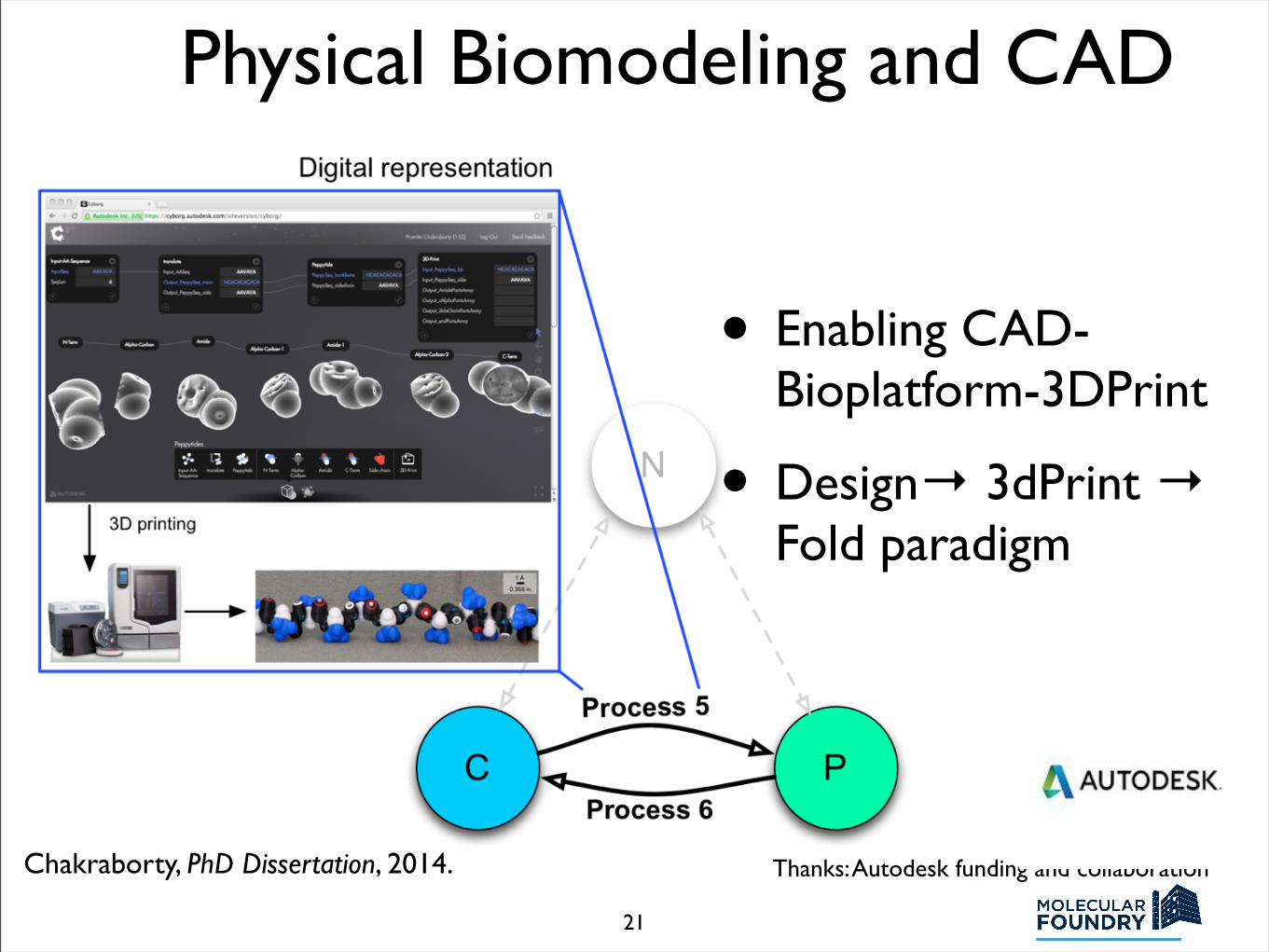
Physical Biomodeling and CAD
21
• Enabling CAD-Bioplatform-3DPrint
• Design→ 3dPrint → Fold paradigm
Thanks: Autodesk funding and collaborationChakraborty, PhD Dissertation, 2014.
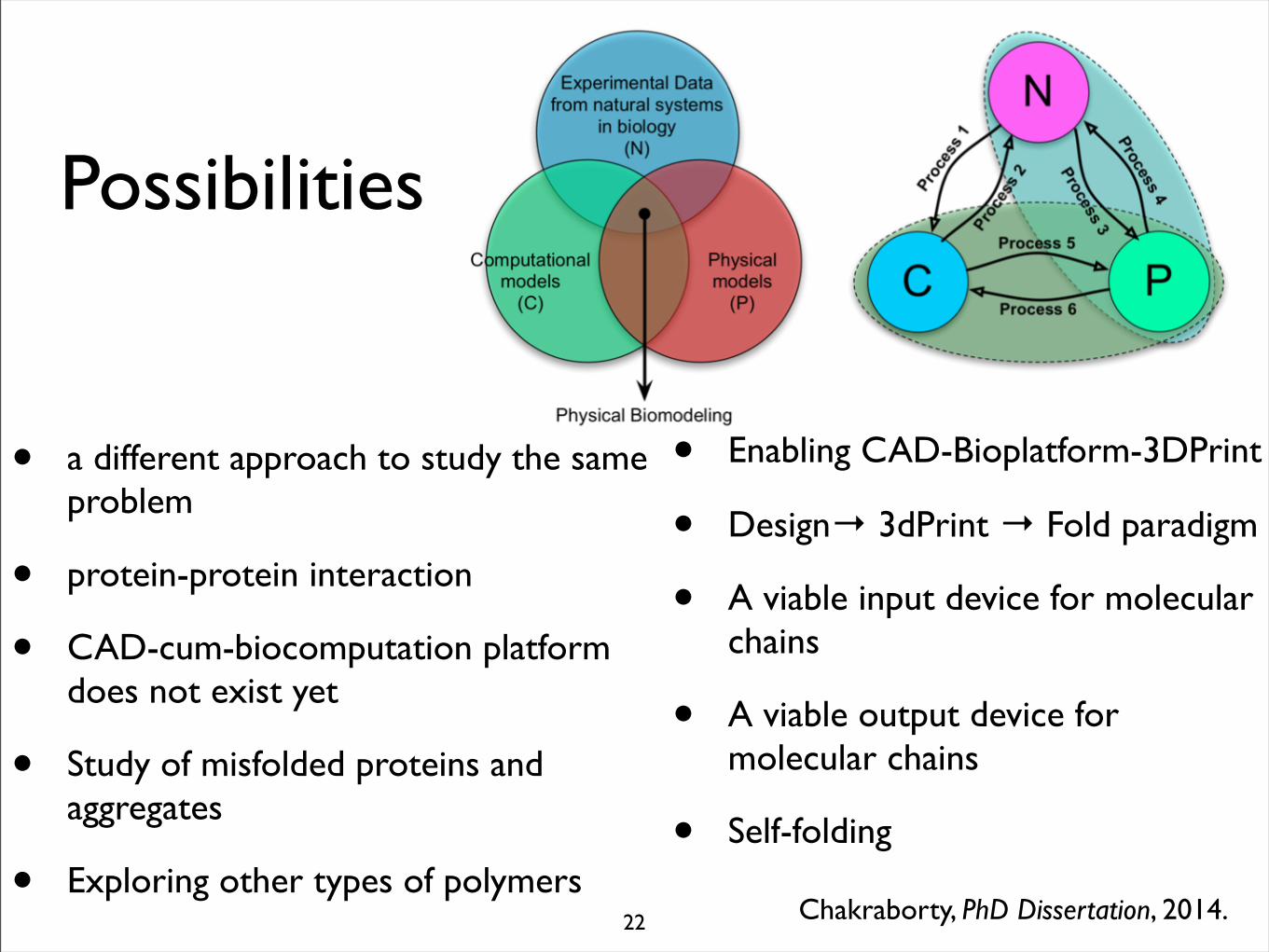
Possibilities
22
• a different approach to study the same problem
• protein-protein interaction
• CAD-cum-biocomputation platform does not exist yet
• Study of misfolded proteins and aggregates
• Exploring other types of polymersChakraborty, PhD Dissertation, 2014.
• Enabling CAD-Bioplatform-3DPrint
• Design→ 3dPrint → Fold paradigm
• A viable input device for molecular chains
• A viable output device for molecular chains
• Self-folding

Envisioning the future of dynamic macromoleculesImagine a world!
23

40!
How to make them?"Open source!
Make Magazine --> Projects!
Peppytides

Acknowledgments and Contacts• QuezyLab
• Collaboration with UCSF Science & Health Education Partnership
• Collaboration with Foothill College, Los Altos
25
• Shantenu Jha, Daniel Katz (CCT, LSU, now at Rutgers U. and Argonne/U. Chicago respectively)
• Deborah Tatar, Steve Harrison, Francis Quek (VT)
• Ronald Zuckermann (LBNL), DoE (Office of Basic Energy Sciences), Defense Threat Reduction Agency (DTRA)
• Alexey Onufriev (VT), Joseph DeRisi (UCSF)
• Molecular Foundry, Lawrence Berkeley National Lab
• Virginia Tech, Dept. of Computer Sc.
• Lawrence Hall of Science Museum
• Industry Collaboration and support by Autodesk Inc.
www.quezylab.com

Peppytide videoshoot Joe DeRisi’s lab, UCSF
Berkeley Lab booth, Berkeley Solano Fest, 2013
Bay Area 2014
Prof. Robert Stroud testing for beta-turns,
UCSF, Dec 2013
Congress Offices, Capitol Hill, Washington DCJun 2014
LBL team with Congressman Jerry McNerney
Nanobio Summer Camp 2014, Foothill College

Extras
27

Existing models
28
CPK models
Dreiding stereomodel
Ball-and-stick
Center for BioMolecular Modeling, Milwaukee School of Engineering http://www.3dmoleculardesigns.com/
Scripps Physical Model Service, Scripps Research Institutehttp://models.scripps.edu/
beta sheet alpha helixDNA double helix
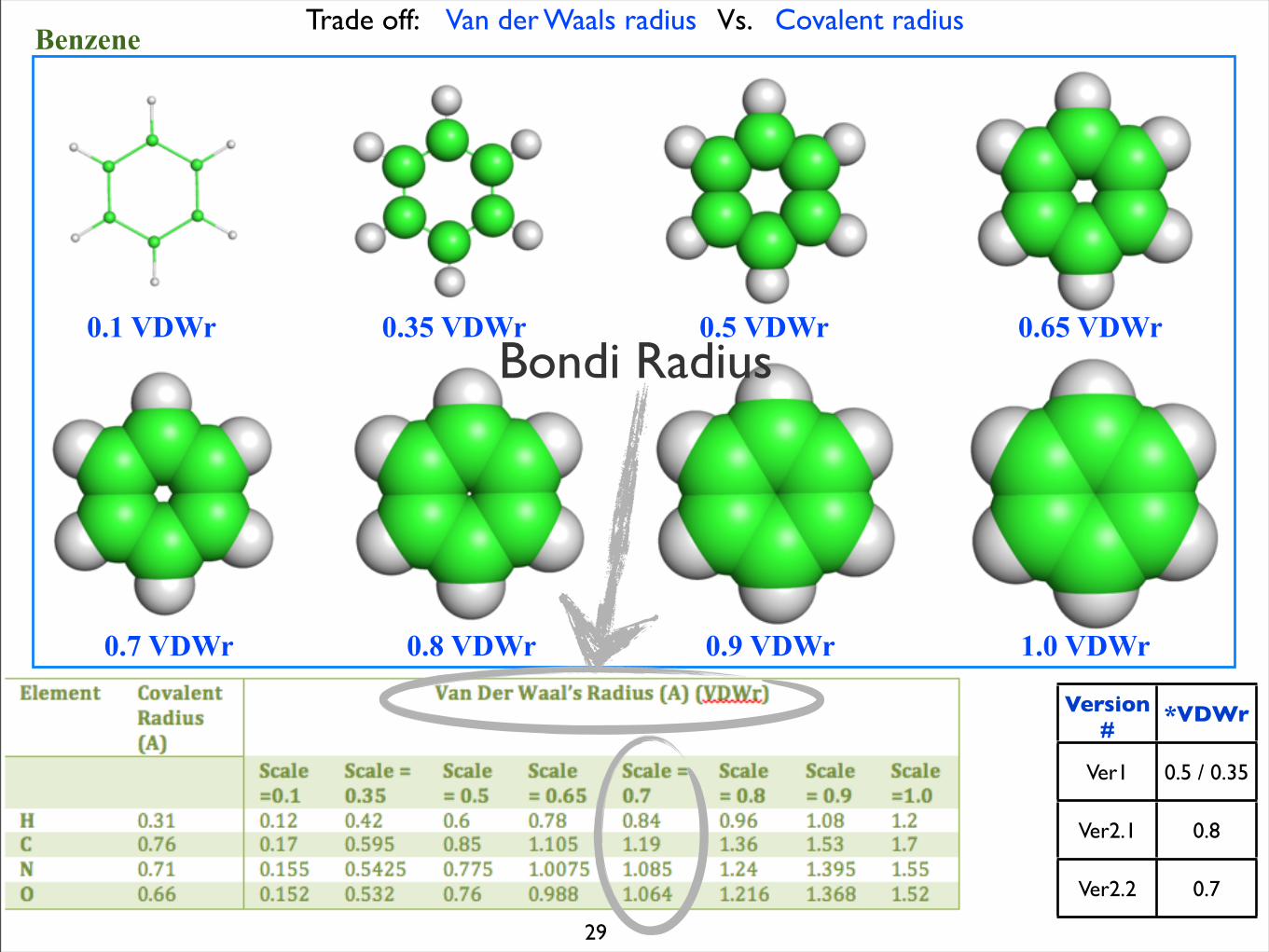
Trade off: Van der Waals radius Vs. Covalent radiusBenzene
0.1 VDWr 0.35 VDWr 0.5 VDWr 0.65 VDWr
0.7 VDWr 0.8 VDWr 0.9 VDWr 1.0 VDWr
Version#
*VDWr
Ver1 0.5 / 0.35
Ver2.1 0.8
Ver2.2 0.7
Bondi Radius
29

30
3D printing
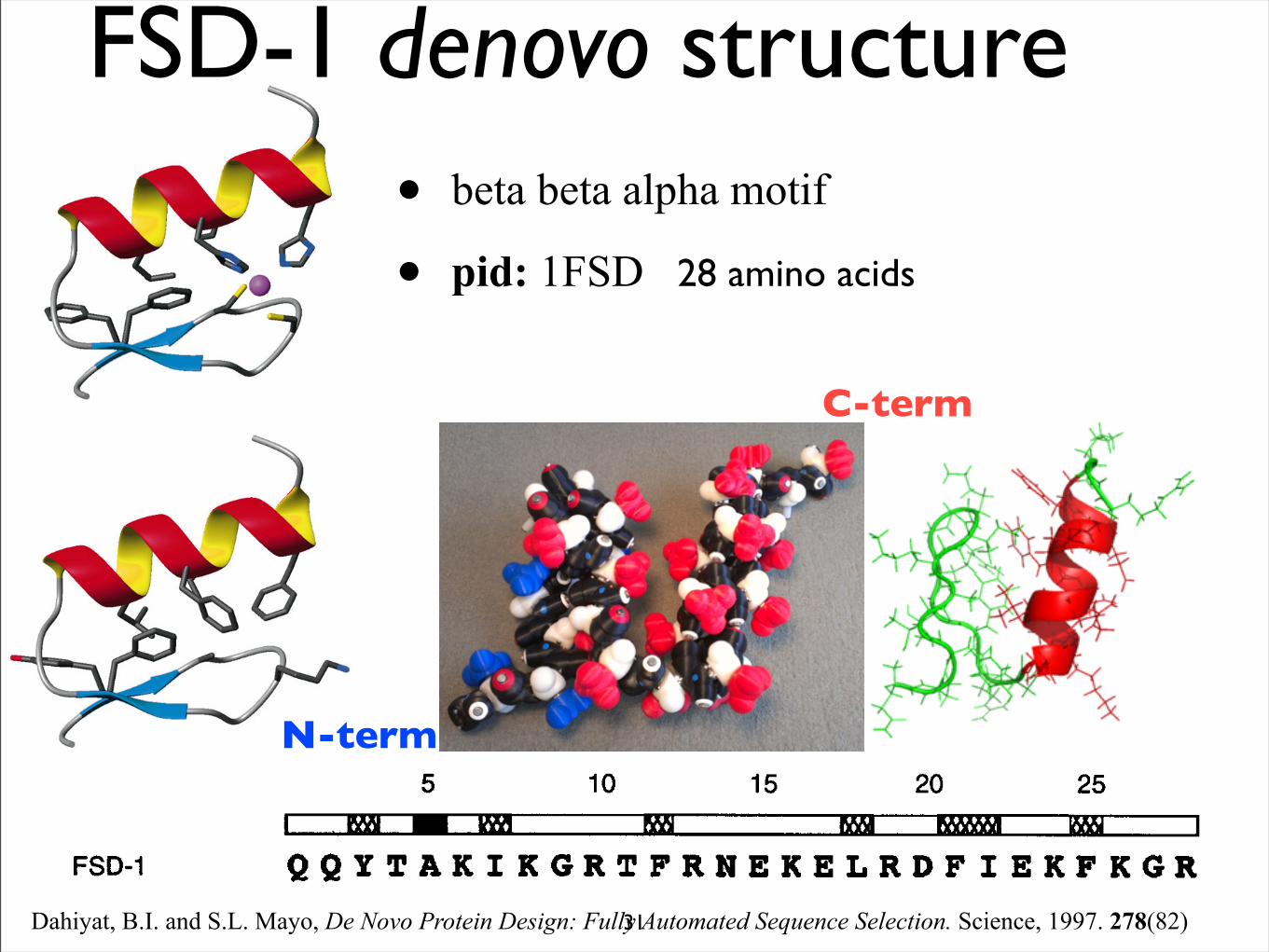
• beta beta alpha motif
• pid: 1FSD 28 amino acids
ther, the BLAST search found only low
identity matches of weak statistical signifi-cance to fragments of various unrelated pro-teins. The highest identity matches were 10
residues (36 percent) with P values ranging
from 0.63 to 1.0, where P is the probability of
a match being a chance occurrence. Random
28-residue sequences that consist of amino
acids allowed in the ��� position classifica-tion described above produced similar
BLAST search results, with 10- or 11-residue
identities (36 to 39 percent) and P values
ranging from 0.35 to 1.0, further suggesting
that the matches for FSD-1 are statistically
insignificant. The low identity with any
known protein sequence demonstrates the
novelty of the FSD-1 sequence and under-scores that no sequence information from
any protein motif was used in our sequence
scoring function.
In order to examine the robustness of the
computed sequence, we used the sequence of
FSD-1 as the starting point of a Monte Carlo
simulated annealing run. The Monte Carlo
search revealed high scoring, suboptimal se-quences in the neighborhood of the optimal
solution (4). The energy spread from the
ground-state solution to the 1000th most
stable sequence is about 5 kcal/mol, an indi-cation that the density of states is high. The
amino acids comprising the core of the mol-ecule, with the exception of position 7, are
essentially invariant (Fig. 1). Almost all of
the sequence variation occurs at surface po-sitions, and typically involves conservative
changes. Asn14, which is predicted to form a
stabilizing hydrogen bond to the helix back-
A
B
Fig. 2. Comparison of Zif268 (9) and computed FSD-1 structures. (A) Stereoview of the second zincfinger module of Zif268 showing its buried residues and zinc binding site. (B) Stereoview of thecomputed orientations of buried side chains in FSD-1. For clarity, only side chains from residues 3, 5, 8,12, 18, 21, 22, and 25 are shown. Color figures were created with MOLMOL (38).
Table 1. NMR structure determination: distance restraints, structural statistics, and atomic root-mean-square (rms) deviations. �SA� are the 41 simulated annealing structures, SA is the average structurebefore energy minimization, (SA )r is the restrained energy minimized average structure, and SD is thestandard deviation.
Distance restraints
Intraresidue 97Sequential 83Short range (�i – j� � 2 to 5 residues) 59Long range (�i – j� � 5 residues) 35Hydrogen bond 10Total 284
Structural statisticsrms deviations �SA� � SD (SA)r
Distance restraints (Å) 0.043 � 0.003 0.038Idealized geometry
Bonds (Å) 0.0041 � 0.0002 0.0037Angles (degrees) 0.67 � 0.02 0.65Impropers (degrees) 0.53 � 0.05 0.51
Atomic rms deviations (Å)*�SA� versus SA � SD �SA� versus (SA)r � SD
Backbone 0.54 � 0.15 0.69 � 0.16Backbone � nonpolar side chains† 0.99 � 0.17 1.16 � 0.18Heavy atoms 1.43 � 0.20 1.90 � 0.29
*Atomic rms deviations are for residues 3 to 26, inclusive. Residues 1, 2, 27, and 28 were disordered [�, �, angularorder parameters (34) � 0.78] and had only sequential and �i – j� � 2 NOEs. †Nonpolar side chains are fromresidues Tyr3, Ala5, Ile7, Phe12, Leu18, Phe21, Ile22, and Phe25, which constitute the core of the protein.
Fig. 3. Circular dichroism (CD) measurements ofFSD-1. (A) Far-UV CD spectrum of FSD-1 at 1°C.The minima at 220 and 207 nm indicate a foldedstructure. (B) Thermal unfolding of FSD-1 moni-tored by CD. The melting curve has an inflectionpoint at 39°C. To illustrate the cooperativity of thethermal transition, the melting curve was fit to atwo-state model [(39) and the derivative of the fit isshown (inset)]. The melting temperature deter-mined from this fit is 42°C.
SCIENCE � VOL. 278 � 3 OCTOBER 1997 � www.sciencemag.org84
on
Augu
st 1
5, 2
012
ww
w.s
cien
cem
ag.o
rgD
ownl
oade
d fro
m
(position 5) the number of residues that canbe assigned unambiguously to the core, where-as seven residues (positions 3, 7, 12, 18, 21,22, and 25) were classified as boundary andthe remaining 20 residues were assigned to thesurface. Whereas three of the zinc bindingpositions of Zif268 are in the boundary orcore, one residue, position 8, has a C�-C�vector directed away from the geometric cen-ter of the protein and is classified as a surfaceposition. As in our previous studies, the aminoacids considered at the core positions duringsequence selection were Ala, Val, Leu, Ile,Phe, Tyr, and Trp; the amino acids consideredat the surface positions were Ala, Ser, Thr,His, Asp, Asn, Glu, Gln, Lys, and Arg; andthe combined core and surface amino acid sets(16 amino acids) were considered at theboundary positions. Two of the residue posi-tions (9 and 27) have � angles greater than 0°and are set to Gly by the sequence selectionalgorithm to minimize backbone strain.
The total number of amino acid sequenc-es that must be considered by the designalgorithm is the product of the number ofpossible amino acids at each residue posi-tion. The ��� motif residue classificationdescribed above results in a virtual combi-natorial library of 1.9 � 1027 possible aminoacid sequences (16). This library size is 15orders of magnitude larger than that acces-sible by experimental random library ap-proaches. A corresponding peptide libraryconsisting of only a single molecule for each28-residue sequence would have a mass of11.6 metric tons (17). In order to accuratelymodel the geometric specificity of side-chain placement, we explicitly consider thetorsional flexibility of amino acid sidechains in our sequence scoring by represent-ing each amino acid with a discrete set ofallowed conformations, called rotamers(18). As a result, the design algorithm mustconsider all rotamers for each possible aminoacid at each residue position. The total sizeof the search space for the ��� motif istherefore 1.1 � 1062 possible rotamer se-quences. We use a search algorithm basedon an extension of the DEE theorem tosolve the rotamer sequence optimizationproblem (4, 8). Efficient implementation ofthe DEE theorem has made complete pro-tein sequence design tractable for about 50residues on current parallel computers in asingle calculation. The rotamer optimizationproblem for the ��� motif required 90 CPUhours to find the optimal sequence (19, 20).
The optimal sequence (Fig. 1) is calledfull sequence design (FSD-1). Even thoughall of the hydrophilic amino acids were con-sidered at each of the boundary positions,the algorithm selected only nonpolar aminoacids. The eight core and boundary positionsare predicted to form a well-packed buriedcluster. The Phe side chains selected by the
algorithm at positions 21 and 25, the zinc-binding His positions of Zif268, are morethan 80 percent buried, and the Ala atposition 5 is 100 percent buried but the Lysat position 8 is more than 60 percent ex-posed to solvent (Fig. 2). The other bound-ary positions demonstrate the steric con-straints on buried residues by packing similarside chains in an arrangement similar to thatof Zif268 (Fig. 2). The calculated optimalconfiguration for core and boundary residuesburies �1150 Å2 of nonpolar surface area.On the helix surface, the algorithm placesAsn14 with a hydrogen bond between itsside-chain carbonyl oxygen and the back-bone amide proton of residue 16. The eightcharged residues on the helix form threepairs of hydrogen bonds, although in ourcoiled-coil designs, helical surface hydrogen
bonds appeared to be less important thanthe overall helix propensity of the sequence(5). Positions 4 and 11 on the exposed sheetsurface were selected by the program to beThr, one of the best �-sheet forming resi-dues (21).
Alignment of the sequences for FSD-1and Zif268 (Fig. 1) indicates that only 6 ofthe 28 residues (21 percent) are identicaland only 11 (39 percent) are similar. Four ofthe identities are in the buried cluster, whichis consistent with the expectation that bur-ied residues are more conserved than sol-vent-exposed residues for a given motif (22).A BLAST (23) search of the FSD-1 se-quence against the nonredundant proteinsequence database of the National Centerfor Biotechnology Information did not re-veal any zinc finger protein sequences. Fur-
Fig. 1. Sequence of FSD-1 aligned with the second zinc finger of Zif268. The bar at the top of the figureshows the residue position classifications: the solid bar indicates the single core position, the hatchedbars indicate the seven boundary positions and the open bars indicate the 20 surface positions. Thealignment matches positions of FSD-1 to the corresponding backbone template positions of Zif268. Ofthe six identical positions (21 percent) between FSD-1 and Zif268, four are buried (Ile7, Phe12, Leu18, andIle22). The zinc binding residues of Zif268 are boxed. Representative nonoptimal sequence solutionsdetermined by means of a Monte Carlo simulated annealing protocol are shown with their rank. Verticallines indicate identity with FSD-1. The symbols at the bottom of the figure show the degree of sequenceconservation for each residue position computed across the top 1000 sequences: filled circles indicatemore than 99 percent conservation, half-filled circles indicate conservation between 90 and 99 percent,open circles indicate conservation between 50 and 90 percent, and the absence of a symbol indicatesless than 50% conservation. The consensus sequence determined by choosing the amino acid with thehighest occurrence at each position is identical to the sequence of FSD-1. Single-letter abbreviations foramino acid residues as follows: A, Ala; C, Cys; D, Asp; E, Glu; F, Phe; G, Gly; H, His; I, Ile; K, Lys; L, Leu;M, Met; N, Asn; P, Pro; Q, Gln; R, Arg; S, Ser; T, Thr; V, Val; W, Trp; and Y, Tyr.
www.sciencemag.org � SCIENCE � VOL. 278 � 3 OCTOBER 1997 83
on
Augu
st 1
5, 2
012
ww
w.s
cien
cem
ag.o
rgD
ownl
oade
d fro
m
N-term
C-term
Dahiyat, B.I. and S.L. Mayo, De Novo Protein Design: Fully Automated Sequence Selection. Science, 1997. 278(82)31
FSD-1 denovo structure
![BIOSTATISTICS AND BIOMODELING - VTUvtu.ac.in/pdf/cbcs/4sem/bio4syll.pdf · BIOSTATISTICS AND BIOMODELING [As per Choice Based Credit System (CBCS) scheme] SEMESTER – IV ... BASIC](https://img.pdfslide.net/doc/110x75/5adce9b97f8b9a4a268cbe8a/biostatistics-and-biomodeling-and-biomodeling-as-per-choice-based-credit-system.jpg)




























