Embed Size (px)
Citation preview

Rapid Stimulation of L-Arginine Transport by D-GlucoseInvolves p42/44mapk and Nitric Oxide in Human Umbilical
Vein EndotheliumCarlos Flores, Susana Rojas, Claudio Aguayo, Jorge Parodi, Giovanni Mann, Jeremy D. Pearson,
Paola Casanello, Luis Sobrevia
Abstract—D-Glucose infusion and gestational diabetes induce vasodilatation in humans and increase L-arginine transportand nitric oxide (NO) synthesis in human umbilical vein endothelial cells. High D-glucose (25 mmol/L, 2 minutes)induced membrane hyperpolarization and an increase of L-arginine transport (Vmax 6.1�0.7 versus 4.4�0.1 pmol/�gprotein per minute) with no change in transport affinity (Km 105�9 versus 111�16 �mol/L). L-[3H]Citrulline formationand intracellular cGMP, but not intracellular Ca2�, were increased by high D-glucose. The effects of D-glucose weremimicked by levcromakalim (ATP-sensitive K� channel blocker), paralleled by p42/p44mapk and Ser1177–endothelial NOsynthase phosphorylation, inhibited by NG-nitro-L-arginine methyl ester (L-NAME; NO synthesis inhibitor), gliben-clamide (ATP-sensitive K� channel blocker), KT-5823 (protein kinase G inhibitor), PD-98059 (mitogen-activatedprotein kinase kinase 1/2 inhibitor), and wortmannin (phosphatidylinositol 3-kinase inhibitor), but they were unaffectedby calphostin C (protein kinase C inhibitor). Elevated D-glucose did not alter superoxide dismutase activity. Our findingsdemonstrate that the human fetal endothelial L-arginine/NO signaling pathway is rapidly activated by elevated D-glucosevia NO and p42/44mapk. This could be determinant in pathologies in which rapid fluctuations of plasma D-glucose mayoccur and may underlie the reported vasodilatation in early stages of diabetes mellitus. (Circ Res. 2003;92:64-72.)
Key Words: humans � endothelium � glucose � arginine � nitric oxide
The cationic amino acid L-arginine is the substrate fornitric oxide (NO) synthesis via endothelial NO synthase
(eNOS)1 and is taken up primarily by the Na�-independenthigh-affinity (Km �100 to 400 �mol/L) systems y�/CAT-1and y�/CAT-2B (where CAT indicates cationic amino acidtransporter) in human umbilical vein endothelial cells(HUVECs).2,3 L-Arginine transport and NO synthesis (L-arginine/NO pathway) are increased in HUVECs from pa-tients with gestational diabetes.2 Interestingly, long-termincubation (24 hours) of HUVECs from normal pregnancieswith elevated D-glucose mimics the effect of gestationaldiabetes on the L-arginine/NO pathway.4 In addition, elevatedD-glucose for 24 hours4,5 or 5 days6 increases eNOS geneexpression. A recent report shows that D-glucose infusioninduces vasodilatation in humans,7 and in animal models, anelevation of plasma D-glucose results in rapid (seconds tominutes) vasodilatation.8–10 Therefore, rapid fluctuations inthe D-glucose level are crucial in maintaining human fetalendothelial function.2–5,11
D-Glucose activates protein kinase C (PKC), an enzymeinvolved with long-term stimulation of the L-arginine/NOpathway,5,12–14 and (within 1 hour) p42 and p44 mitogen-ac-
tivated protein (MAP) kinases (p42/44mapk).5,14,15 p42/44mapk
activation may itself be dependent on PKC activation and NOsynthesis.5,14 However, the effect of short-term incubationwith elevated D-glucose on the endothelial L-arginine/NOpathway has not been investigated.4,11,16,17
The present study shows that a 2-minute incubation with25 mmol/L D-glucose increases L-arginine transport and NOsynthesis in HUVECs. The underlying cellular mechanismsinvolve phosphorylation of eNOS at Ser1177 via phosphatidyl-inositol 3-kinase (PI3-k) and activation of eNOS and p42/p44mapk by D-glucose.18
Materials and Methods
Cell CultureHuman umbilical vein endothelium was isolated (collagenase diges-tion 0.25 mg/mL) and cultured (37°C, 5% CO2, confluent passage 2)in medium 199 containing 5 mmol/L D-glucose, 10% newborn calfserum, 10% fetal calf serum, 3.2 mmol/L L-glutamine, 100 �mol/LL-arginine, and 100 U/mL penicillin-streptomycin (primary culturemedium).2–4 Before an experiment (24 hours), the incubation me-dium was changed to serum-free medium 199.
Original received July 26, 2002; revision received October 31, 2002; accepted November 11, 2002.From the Cellular and Molecular Physiology Laboratory (C.F., S.R., C.A., J.P., P.C., L.S.), Department of Physiology, Faculty of Biological Sciences,
and the Department of Obstetrics and Gynaecology (P.C.), Faculty of Medicine, University of Concepción, Concepción, Chile, and King’s CollegeLondon (G.M., J.D.P.), Guy’s Campus, London, UK.
Correspondence to Dr L. Sobrevia, Cellular and Molecular Physiology Laboratory (CMPL), Department of Physiology, Faculty of Biological Sciences,University of Concepción, PO Box 160-C, Concepción, Chile. E-mail [email protected]
© 2003 American Heart Association, Inc.
Circulation Research is available at http://www.circresaha.org DOI: 10.1161/01.RES.0000048197.78764.D6
64

L-Arginine TransportL-Arginine transport (1 �Ci/mL, 37°C, 1 minute) was determined incells preincubated (15 seconds to 5 minutes) with Krebs solution(mmol/L: NaCl 131, KCl 5.6, NaHCO3 25, NaH2PO4 1, HEPES 20,CaCl2 2.5, and MgCl2 1 [pH 7.4, 37°C]) containing 5 or 25 mmol/LD-glucose, 25 mmol/L L-glucose, or 5 mmol/L D-glucose plus20 mmol/L D-mannitol (osmotic controls).2–4 L-Arginine transportwas also determined in Krebs solution in which NaCl was replacedby equimolar concentrations of choline chloride2–4 or in cellsincubated (30 minutes) with KCl (5.5 to 131 mmol/L), with NaCldecreased equivalently, or with 131 mmol/L KCl for 2, 4, 10, 20, or30 minutes. In trans-stimulation experiments, cells were preincu-bated (2 hours) with primary culture medium containing 10 mmol/LL-lysine. Cell-associated radioactivity and data analyses were per-formed as described.2–4
L-Arginine transport was assayed in cells preincubated (30 min-utes) with 100 �mol/L NG-nitro-L-arginine methyl ester (L-NAME,eNOS inhibitor),2,3 10 �mol/L PD-98059 (MAP kinase kinase 1/2[MEK1/2] inhibitor),19 100 �mol/L S-nitroso-N-acetyl-L,D-penicillamine (SNAP, NO donor), 1 �mol/L KT-5823 (proteinkinase G [PKG] inhibitor),20 100 nmol/L calphostin C (PKC inhib-itor),21 10 �mol/L glibenclamide (ATP-sensitive K� [K�
ATP] channelblocker),22 1 �mol/L levcromakalim (K�
ATP activator),23 or 30nmol/L wortmannin (PI3-k inhibitor).24
Membrane Potential[3H]Tetraphenylphosphonium ([3H]TPP�) influx (46 nmol/L, 0.5�Ci/mL, 15 to 120 seconds), a membrane potential–sensitiveprobe,2,3,25 was determined in Krebs solution containing 5 or25 mmol/L D-glucose, 100 �mol/L L-arginine, and 5.5 or131 mmol/L KCl.2,3 Resting membrane potential (Em, whole-cellpatch clamp) was recorded using an EPC-7 amplifier (List Medical)as described.2 Em was measured for at least 1 minute in the presenceof 5 or 25 mmol/L D-glucose. Only recordings with �0.1-mVvariations were considered. Em was also determined in cells prein-cubated for 2 hours with 10 mmol/L L-lysine.
PKC ActivityPKC activity (32P incorporation from [�-32P]ATP into a syntheticPKC substrate peptide analogue5) was determined in cells exposed to5 or 25 mmol/L D-glucose (2 minutes) after pretreatment with 100nmol/L phorbol 12-myristate 13-acetate (5 minutes, PKC activator),100 nmol/L 4�-phorbol 12,13-didecanoate (5 minutes, less activephorbol 12-myristate 13-acetate analogue), 100 nmol/L calphostin C(30 minutes), 10 �mol/L PD-98059 (30 minutes), 100 �mol/LSNAP (5 minutes), 100 �mol/L L-NAME (30 minutes), or 30nmol/L wortmannin (30 minutes).
cGMP DeterminationCells preincubated (30 minutes) in Krebs solution (37°C) containingL-arginine (100 �mol/L) and 3-isobutyl-1-methylxanthine(0.5 mmol/L, phosphodiesterase inhibitor),2,3 in the absence orpresence of L-NAME (100 �mol/L), were exposed to 5 or25 mmol/L D-glucose for the last 2 minutes of the 30-minuteincubation period with 3-isobutyl-1-methylxanthine. cGMP wasdetermined in HCl-cell extracts by radioimmunoassay.2,3
L-Citrulline AssayCells were incubated with L-[3H]arginine (100 �mol/L, 4 �Ci/mL,30 minutes, 37°C) in the absence or presence of L-NAME (100�mol/L). Cells were exposed to 5 or 25 mmol/L D-glucose for 30minutes or for the last 2, 4, 10, or 20 minutes of this incubationperiod. Some experiments were performed in cells exposed only toL-[3H]arginine (4 �Ci/mL) and 5 or 25 mmol/L D-glucose. Formicacid–digested cells (200 �L) were passed through a sodium ion formof the cation ion-exchange resin Dowex 50W (50X8-200), andL-[3H]citrulline concentration was determined in the H2O eluate.3
Intracellular Ca2�
Cells on glass coverslips were loaded (30 minutes, 23°C) with theacetoxymethyl derivative of fluo 3 (5 �mol/L). Coverslips weretransferred to an experimental bath with Krebs solution containing 5or 25 mmol/L D-glucose, and Ca2� was imaged using a Zeiss LSM410 confocal microscope.4
Western BlotsAfter pretreatment with 10 �mol/L PD-98059 (30 minutes), 100�mol/L SNAP (2 to 5 minutes), or 30 nmol/L wortmannin (30minutes), the cells were incubated with 5 or 25 mmol/L D-glucose (2minutes). Cell protein extracts were probed with a primary poly-clonal mouse antiphosphorylated (1:1000) or nonphosphorylated(1:1500) p44/p42mapk, rabbit anti-eNOS (1:2500) or anti-phosphory-lated Ser1177–eNOS (1:2500) antibodies, and horseradish peroxidase–conjugated goat secondary antibodies as described.3,5 Primary poly-clonal mouse anti-actin (1:2000) served as the internal control.Proteins were detected by enhanced chemiluminescence and quan-tified by densitometry (Ultroscan XL enhanced laser densitometer,LKB Instruments).3,5
Semiquantitative PCRExtracted mRNA (Dynal) was reversed-transcribed into cDNA usingoligo(dT18) plus random hexamers (10-mer) and M-MLV reversetranscriptase (Promega) for 1 hour at 37°C.3 Polymerase chainreactions (PCRs) were performed in 20-�L samples (2 �L of 10�PCR buffer, 0.8 �L of 50 mmol/L Mg2�, 0.4 �L dNTPs, 13.6 �LRNase-free H2O, 0.2 �L Taq DNA polymerase, and 0.5 �mol/Lsequence-specific oligonucleotide primers for human CAT-1, CAT-2A, or CAT-2B). Samples were incubated (4 minutes, 95°C),followed by 35 cycles of 30 seconds at 95°C, 30 seconds at 57°C, 30seconds at 72°C, and a final extension for 7 minutes at 72°C. �-Actinexpression was used as a reference value. Reverse transcription(RT)-PCR products were sequenced in both directions by Taqdideoxy terminator cycle sequencing (automated DNA sequencer373A, Applied Biosystems).3
Oligonucleotide primers were as follows: hCAT-1 (sense)5�-CCAGTACTTCCGACGAGTTAGA-3�, hCAT-1 (antisense)5�-CATCCACACAGCAAACCGGACC-3�, hCAT-2A (sense) 5�-TATCCCGATTTTTTTGCTGTGTGC-3�, hCAT-2A (antisense)5�-TGCAGTCAACGTGGCAGCAACT-3�, hCAT-2B (sense) 5�-TCCCAATGCCTCGTGTAATCTA-3�, hCAT-2B (antisense)5�-GCATGCTGAAGCCCTGTCTCTGC-3�, �-actin (sense) 5�-AACCGCGAGAAGATGACCCAGATCATCTTT-3�, and �-actin(antisense) 5�-AGCAGCCGTGGCCATCTCTTGCTCGAAGTC-3�.Expected size products were as follows: hCAT-1, 450 bp; hCAT-2A,690 bp; hCAT-2B, 360 bp; and �-actin, 350 bp.
SOD Activity and �-Tocopherol ExperimentsCells were homogenized in buffer containing 50 mmol/L Tris-(hy-droxymethyl)-aminomethane, 100 mmol/L potassium chloride,0.02% Triton X-100, 100 mmol/L sodium pyrophosphate, and100 mmol/L sodium fluoride (pH 7.4), which was supplemented withtrypsin inhibitors (4 mg/mL aprotinin, 1 mg/mL benzamidine, 5�g/mL leupeptin, and 200 �mol/L sodium orthovanadate). Aliquots(1 mg protein/mL) were incubated (25°C, 2 minutes) with potassiumphosphate buffer (50 mmol/L, pH 10.2) containing adrenochrome(200 �mol/L) and epinephrine (10 �mol/L), and absorbance wasmeasured at 480 nm. Superoxide dismutase (SOD) activity wascalculated from the inhibition curve for epinephrine auto-oxidationversus protein concentration. Basal absorbance (100% activity) wasthe reaction in the absence of cell extracts.26 Cells were alsopreincubated (30 minutes) with �-tocopherol (500 �g/mL, 1%ethanol).27
MaterialsSera, agarose, and buffers were from GIBCO Life Technologies.Collagenase type II (Clostridium histolyticum) was from Boehring-er-Mannheim, and Bradford protein reagent was from Bio-RadLaboratories. L-NAME and SNAP were from Calbiochem. Ethidium
Flores et al D-Glucose Acutely Activates L-Arginine/NO Pathway 65
bromide, Dowex (50WX8-400), and all other reagents were fromSigma Chemical Co. L-[2,3-3H]Arginine (36.1 Ci/mmol), D-[1-14C]mannitol (49.3 mCi/mmol), [�-32P]ATP, and [3H]TPP� (37Ci/mmol) were from NEN. 3�,5�-cGMP-TME was from ICN. Anti-bodies were from Cell Signaling, New England Biolabs.
Statistical AnalysisValues are mean�SEM, where n indicates the number of differentcell cultures (4 to 8 replicates per experiment). Statistical analyseswere carried out on raw data using the Peritz F multiple-meanscomparison test.28 A Student t test was applied for unpaired data, anda value of P�0.05 was considered statistically significant.
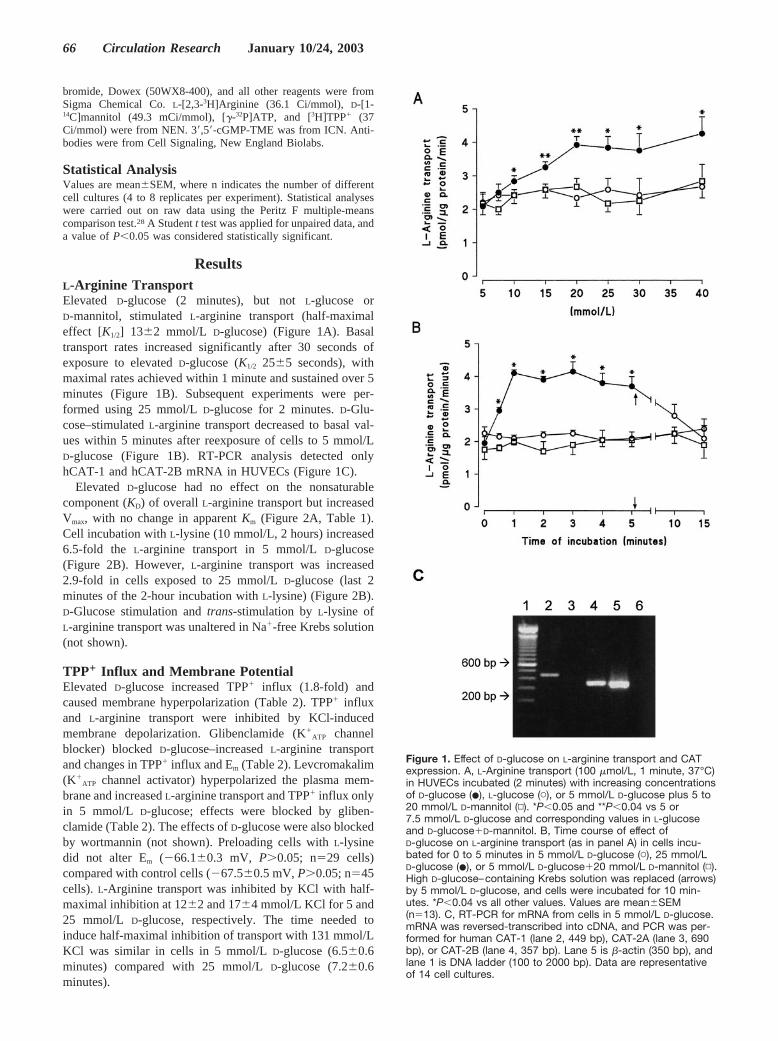
ResultsL-Arginine TransportElevated D-glucose (2 minutes), but not L-glucose orD-mannitol, stimulated L-arginine transport (half-maximaleffect [K1/2] 13�2 mmol/L D-glucose) (Figure 1A). Basaltransport rates increased significantly after 30 seconds ofexposure to elevated D-glucose (K1/2 25�5 seconds), withmaximal rates achieved within 1 minute and sustained over 5minutes (Figure 1B). Subsequent experiments were per-formed using 25 mmol/L D-glucose for 2 minutes. D-Glu-cose–stimulated L-arginine transport decreased to basal val-ues within 5 minutes after reexposure of cells to 5 mmol/LD-glucose (Figure 1B). RT-PCR analysis detected onlyhCAT-1 and hCAT-2B mRNA in HUVECs (Figure 1C).
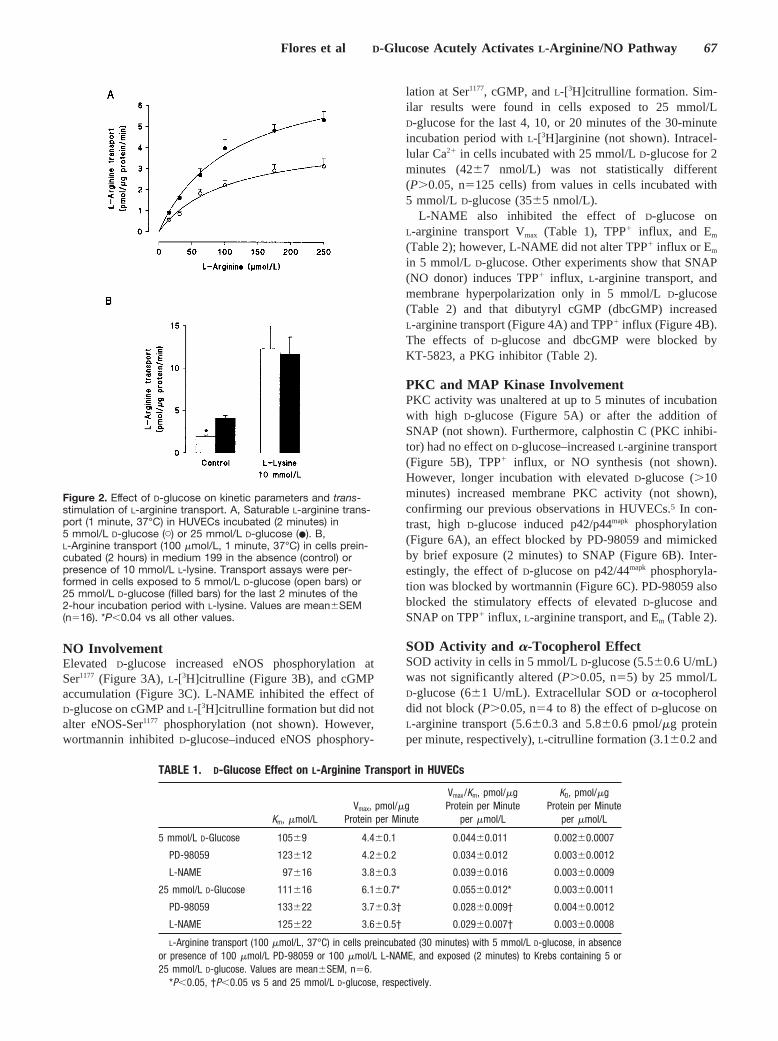
Elevated D-glucose had no effect on the nonsaturablecomponent (KD) of overall L-arginine transport but increasedVmax, with no change in apparent Km (Figure 2A, Table 1).Cell incubation with L-lysine (10 mmol/L, 2 hours) increased6.5-fold the L-arginine transport in 5 mmol/L D-glucose(Figure 2B). However, L-arginine transport was increased2.9-fold in cells exposed to 25 mmol/L D-glucose (last 2minutes of the 2-hour incubation with L-lysine) (Figure 2B).D-Glucose stimulation and trans-stimulation by L-lysine ofL-arginine transport was unaltered in Na�-free Krebs solution(not shown).
TPP� Influx and Membrane PotentialElevated D-glucose increased TPP� influx (1.8-fold) andcaused membrane hyperpolarization (Table 2). TPP� influxand L-arginine transport were inhibited by KCl-inducedmembrane depolarization. Glibenclamide (K�
ATP channelblocker) blocked D-glucose–increased L-arginine transportand changes in TPP� influx and Em (Table 2). Levcromakalim(K�
ATP channel activator) hyperpolarized the plasma mem-brane and increased L-arginine transport and TPP� influx onlyin 5 mmol/L D-glucose; effects were blocked by gliben-clamide (Table 2). The effects of D-glucose were also blockedby wortmannin (not shown). Preloading cells with L-lysinedid not alter Em (�66.1�0.3 mV, P�0.05; n29 cells)compared with control cells (�67.5�0.5 mV, P�0.05; n45cells). L-Arginine transport was inhibited by KCl with half-maximal inhibition at 12�2 and 17�4 mmol/L KCl for 5 and25 mmol/L D-glucose, respectively. The time needed toinduce half-maximal inhibition of transport with 131 mmol/LKCl was similar in cells in 5 mmol/L D-glucose (6.5�0.6minutes) compared with 25 mmol/L D-glucose (7.2�0.6minutes).
Figure 1. Effect of D-glucose on L-arginine transport and CATexpression. A, L-Arginine transport (100 �mol/L, 1 minute, 37°C)in HUVECs incubated (2 minutes) with increasing concentrationsof D-glucose (�), L-glucose (�), or 5 mmol/L D-glucose plus 5 to20 mmol/L D-mannitol (▫). *P�0.05 and **P�0.04 vs 5 or7.5 mmol/L D-glucose and corresponding values in L-glucoseand D-glucose�D-mannitol. B, Time course of effect ofD-glucose on L-arginine transport (as in panel A) in cells incu-bated for 0 to 5 minutes in 5 mmol/L D-glucose (�), 25 mmol/LD-glucose (�), or 5 mmol/L D-glucose�20 mmol/L D-mannitol (▫).High D-glucose–containing Krebs solution was replaced (arrows)by 5 mmol/L D-glucose, and cells were incubated for 10 min-utes. *P�0.04 vs all other values. Values are mean�SEM(n13). C, RT-PCR for mRNA from cells in 5 mmol/L D-glucose.mRNA was reversed-transcribed into cDNA, and PCR was per-formed for human CAT-1 (lane 2, 449 bp), CAT-2A (lane 3, 690bp), or CAT-2B (lane 4, 357 bp). Lane 5 is �-actin (350 bp), andlane 1 is DNA ladder (100 to 2000 bp). Data are representativeof 14 cell cultures.
66 Circulation Research January 10/24, 2003
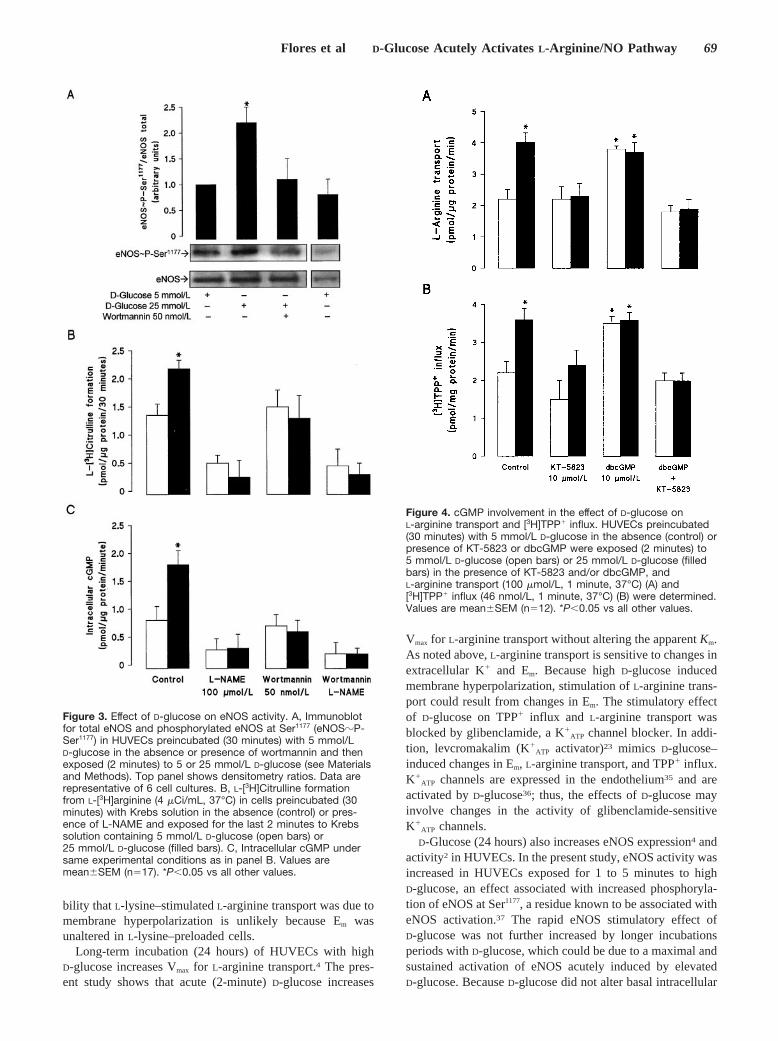
NO InvolvementElevated D-glucose increased eNOS phosphorylation atSer1177 (Figure 3A), L-[3H]citrulline (Figure 3B), and cGMPaccumulation (Figure 3C). L-NAME inhibited the effect ofD-glucose on cGMP and L-[3H]citrulline formation but did notalter eNOS-Ser1177 phosphorylation (not shown). However,wortmannin inhibited D-glucose–induced eNOS phosphory-
lation at Ser1177, cGMP, and L-[3H]citrulline formation. Sim-ilar results were found in cells exposed to 25 mmol/LD-glucose for the last 4, 10, or 20 minutes of the 30-minuteincubation period with L-[3H]arginine (not shown). Intracel-lular Ca2� in cells incubated with 25 mmol/L D-glucose for 2minutes (42�7 nmol/L) was not statistically different(P�0.05, n125 cells) from values in cells incubated with5 mmol/L D-glucose (35�5 nmol/L).
L-NAME also inhibited the effect of D-glucose onL-arginine transport Vmax (Table 1), TPP� influx, and Em
(Table 2); however, L-NAME did not alter TPP� influx or Em
in 5 mmol/L D-glucose. Other experiments show that SNAP(NO donor) induces TPP� influx, L-arginine transport, andmembrane hyperpolarization only in 5 mmol/L D-glucose(Table 2) and that dibutyryl cGMP (dbcGMP) increasedL-arginine transport (Figure 4A) and TPP� influx (Figure 4B).The effects of D-glucose and dbcGMP were blocked byKT-5823, a PKG inhibitor (Table 2).
PKC and MAP Kinase InvolvementPKC activity was unaltered at up to 5 minutes of incubationwith high D-glucose (Figure 5A) or after the addition ofSNAP (not shown). Furthermore, calphostin C (PKC inhibi-tor) had no effect on D-glucose–increased L-arginine transport(Figure 5B), TPP� influx, or NO synthesis (not shown).However, longer incubation with elevated D-glucose (�10minutes) increased membrane PKC activity (not shown),confirming our previous observations in HUVECs.5 In con-trast, high D-glucose induced p42/p44mapk phosphorylation(Figure 6A), an effect blocked by PD-98059 and mimickedby brief exposure (2 minutes) to SNAP (Figure 6B). Inter-estingly, the effect of D-glucose on p42/44mapk phosphoryla-tion was blocked by wortmannin (Figure 6C). PD-98059 alsoblocked the stimulatory effects of elevated D-glucose andSNAP on TPP� influx, L-arginine transport, and Em (Table 2).
SOD Activity and �-Tocopherol EffectSOD activity in cells in 5 mmol/L D-glucose (5.5�0.6 U/mL)was not significantly altered (P�0.05, n5) by 25 mmol/LD-glucose (6�1 U/mL). Extracellular SOD or �-tocopheroldid not block (P�0.05, n4 to 8) the effect of D-glucose onL-arginine transport (5.6�0.3 and 5.8�0.6 pmol/�g proteinper minute, respectively), L-citrulline formation (3.1�0.2 and
Figure 2. Effect of D-glucose on kinetic parameters and trans-stimulation of L-arginine transport. A, Saturable L-arginine trans-port (1 minute, 37°C) in HUVECs incubated (2 minutes) in5 mmol/L D-glucose (�) or 25 mmol/L D-glucose (�). B,L-Arginine transport (100 �mol/L, 1 minute, 37°C) in cells prein-cubated (2 hours) in medium 199 in the absence (control) orpresence of 10 mmol/L L-lysine. Transport assays were per-formed in cells exposed to 5 mmol/L D-glucose (open bars) or25 mmol/L D-glucose (filled bars) for the last 2 minutes of the2-hour incubation period with L-lysine. Values are mean�SEM(n16). *P�0.04 vs all other values.
TABLE 1. D-Glucose Effect on L-Arginine Transport in HUVECs
Km, �mol/LVmax, pmol/�g
Protein per Minute
Vmax/Km, pmol/�gProtein per Minute
per �mol/L
KD, pmol/�gProtein per Minute
per �mol/L
5 mmol/L D-Glucose 105�9 4.4�0.1 0.044�0.011 0.002�0.0007
PD-98059 123�12 4.2�0.2 0.034�0.012 0.003�0.0012
L-NAME 97�16 3.8�0.3 0.039�0.016 0.003�0.0009
25 mmol/L D-Glucose 111�16 6.1�0.7* 0.055�0.012* 0.003�0.0011
PD-98059 133�22 3.7�0.3† 0.028�0.009† 0.004�0.0012
L-NAME 125�22 3.6�0.5† 0.029�0.007† 0.003�0.0008
L-Arginine transport (100 �mol/L, 37°C) in cells preincubated (30 minutes) with 5 mmol/L D-glucose, in absenceor presence of 100 �mol/L PD-98059 or 100 �mol/L L-NAME, and exposed (2 minutes) to Krebs containing 5 or25 mmol/L D-glucose. Values are mean�SEM, n6.
*P�0.05, †P�0.05 vs 5 and 25 mmol/L D-glucose, respectively.
Flores et al D-Glucose Acutely Activates L-Arginine/NO Pathway 67

2.8�0.4 pmol/�g protein per 30 minutes, respectively), TPP�
influx (4.4�0.2 and 4.1�0.3 pmol/mg protein per minute,respectively), or changes in Em (�76�0.3 and �78�0.3 mV,respectively).
DiscussionThe present study establishes that D-glucose induces a rapidconcentration-dependent stimulation of L-arginine transportin HUVECs. This effect requires NO synthesis associatedwith increased phosphorylation of eNOS at Ser1177 and acti-vation of p42/p44mapk and PI3-k, and it is independent of PKCand intracellular Ca2� changes. These findings provide thefirst evidence that short-term hyperglycemia activates theL-arginine/NO signaling pathway in human fetalendothelium.
L-Arginine transport is mediated by systems y�/CATs,2,3,12
y�L,29,30 and b0,�12 in HUVECs, with the first likely predom-inating at the physiological concentration of extracellularL-arginine. The cDNAs for four potential human y� trans-porters (hCAT-1, hCAT-2B, hCAT-2A, and hCAT-4) havebeen sequenced.31 L-Arginine transport in HUVECs occurswith relatively high affinity (Km �80 to 100 �mol/L) and, as
confirmed here, is Na� independent and inhibited by mem-brane depolarization.2,3,32 Because of their similar kineticproperties, CAT-1 and CAT-2B are hard to distinguish at thefunctional level.31 Our results show that both high-affinityhCAT-1 (Km �100 to 200 �mol/L) and hCAT-2B (Km �200to 400 �mol/L), but not the low-affinity hCAT-2A trans-porter (Km �2 to 5 mmol/L), are present in HUVECs,confirming previous reports.3,33 CAT-1 is more sensitive thanCAT-2B to trans-stimulation by cationic amino acids.3,31,34
When we preloaded HUVECs with L-lysine, L-arginine trans-port was increased by �7-fold in 5 mmol/L D-glucose.However, the L-lysine trans-stimulatory effect was less ef-fective (�3-fold) in cells exposed for 2 minutes to 25 mmol/LD-glucose. Because L-arginine transport is trans-stimulatedby 9.8-fold or 1.8-fold in Xenopus oocytes injected withhCAT-1 or hCAT-2B mRNA, respectively,34 trans-stimulation in HUVECs in 5 mmol/L D-glucose may bepreferentially mediated by hCAT-1. The reduced trans-stimulation of transport in high D-glucose may result from astate of maximal activity of L-arginine transporters alreadyinduced by L-lysine; therefore, high D-glucose could notfurther increase L-arginine transport. In addition, the possi-
TABLE 2. D-Glucose Effect on L-Arginine Transport, [3H]TPP� Influx, and Em in HUVECs
L-Arginine Transport, pmol/�gProtein per Minute
[3H]TPP� Influx, pmol/mgProtein per Minute Em, mV
5 mmol/L D-Glucose
Control 1.8�0.3 2.1�0.3 �67.2�0.5
KCl 0.2�0.1* 0.1�0.04* �8.1�0.5*
Glibenclamide 1.3�0.3 1.5�0.3 �63.9�0.6
Levcromakalim 4.6�0.3* 5.6�0.3* �74.2�0.3*
Levcromakalim�glibenclamide 1.6�0.2† 1.4�0.6† �64.5�0.2†
PD-98059 2.1�0.1 1.7�0.2 �64.2�0.5
L-NAME 1.6�0.1 1.5�0.3 �65.1�0.3
SNAP 5.1�0.3* 3.9�0.4* �76.2�1.0*
SNAP�PD-98059 2.1�0.3† 1.9�0.5† �65.3�1.0†
KT-5823 1.9�0.5 1.9�0.3 �66.8�0.9
25 mmol/L D-Glucose
Control 4.0�0.5* 3.7�0.3* �77.1�0.2*
KCl 0.3�0.2*‡ 0.3�0.03*‡ �6.2�0.5*‡
Glibenclamide 2.3�0.4‡ 2.4�0.4‡ �62.7�0.5‡
Levcromakalim 4.3�0.4* 4.7�0.2* �75.1�0.3*
Levcromakalim�glibenclamide 2.2�0.3†‡ 1.8�0.3†‡ �61.9�0.3†‡
PD-98059 0.9�0.3‡ 1.7�0.1‡ �69.9�1.0‡
L-NAME 1.4�0.1‡ 1.8�0.2‡ �65.7�0.7‡
SNAP 4.1�0.4* 3.2�0.2* �78.2�1.0*
SNAP�PD-98059 1.1�0.5†‡ 1.9�0.2†‡ �65.7�1.0†‡
KT-5823 1.9�0.5‡ 1.9�0.3‡ �65.3�0.5‡
L-Arginine transport (100 �mol/L), 3H�TPP� influx (46 nmol/L), and Em (whole-cell patch clamp) in cellspreincubated (30 minutes) with 5 mmol/L D-glucose and 5.5 mmol/L (control) or 131 mmol/L KCl. Cells in 5.5 mmol/LKCl were preincubated with 10 �mol/L glibenclamide (5 minutes), 1 �mol/L levcromakalim (5 minutes), 10 �mol/LPD-98059 (30 minutes), 100 �mol/L L-NAME (30 minutes), 10 �mol/L KT-5823 (30 minutes), or 100 �mol/L SNAP(2 minutes) and exposed (2 minutes) to 5 or 25 mmol/L D-glucose. Values are mean�SEM, n17.
*P�0.05 vs control in 5 mmol/L D-glucose; †P�0.05 vs corresponding values in SNAP or levcromakalim; ‡P�0.05vs control in 25 mmol/L D-glucose.
68 Circulation Research January 10/24, 2003
bility that L-lysine–stimulated L-arginine transport was due tomembrane hyperpolarization is unlikely because Em wasunaltered in L-lysine–preloaded cells.
Long-term incubation (24 hours) of HUVECs with highD-glucose increases Vmax for L-arginine transport.4 The pres-ent study shows that acute (2-minute) D-glucose increases
Vmax for L-arginine transport without altering the apparent Km.As noted above, L-arginine transport is sensitive to changes inextracellular K� and Em. Because high D-glucose inducedmembrane hyperpolarization, stimulation of L-arginine trans-port could result from changes in Em. The stimulatory effectof D-glucose on TPP� influx and L-arginine transport wasblocked by glibenclamide, a K�
ATP channel blocker. In addi-tion, levcromakalim (K�
ATP activator)23 mimics D-glucose–induced changes in Em, L-arginine transport, and TPP� influx.K�
ATP channels are expressed in the endothelium35 and areactivated by D-glucose36; thus, the effects of D-glucose mayinvolve changes in the activity of glibenclamide-sensitiveK�
ATP channels.D-Glucose (24 hours) also increases eNOS expression4 and
activity2 in HUVECs. In the present study, eNOS activity wasincreased in HUVECs exposed for 1 to 5 minutes to highD-glucose, an effect associated with increased phosphoryla-tion of eNOS at Ser1177, a residue known to be associated witheNOS activation.37 The rapid eNOS stimulatory effect ofD-glucose was not further increased by longer incubationsperiods with D-glucose, which could be due to a maximal andsustained activation of eNOS acutely induced by elevatedD-glucose. Because D-glucose did not alter basal intracellular
Figure 3. Effect of D-glucose on eNOS activity. A, Immunoblotfor total eNOS and phosphorylated eNOS at Ser1177 (eNOS�P-Ser1177) in HUVECs preincubated (30 minutes) with 5 mmol/LD-glucose in the absence or presence of wortmannin and thenexposed (2 minutes) to 5 or 25 mmol/L D-glucose (see Materialsand Methods). Top panel shows densitometry ratios. Data arerepresentative of 6 cell cultures. B, L-[3H]Citrulline formationfrom L-[3H]arginine (4 �Ci/mL, 37°C) in cells preincubated (30minutes) with Krebs solution in the absence (control) or pres-ence of L-NAME and exposed for the last 2 minutes to Krebssolution containing 5 mmol/L D-glucose (open bars) or25 mmol/L D-glucose (filled bars). C, Intracellular cGMP undersame experimental conditions as in panel B. Values aremean�SEM (n17). *P�0.05 vs all other values.
Figure 4. cGMP involvement in the effect of D-glucose onL-arginine transport and [3H]TPP� influx. HUVECs preincubated(30 minutes) with 5 mmol/L D-glucose in the absence (control) orpresence of KT-5823 or dbcGMP were exposed (2 minutes) to5 mmol/L D-glucose (open bars) or 25 mmol/L D-glucose (filledbars) in the presence of KT-5823 and/or dbcGMP, andL-arginine transport (100 �mol/L, 1 minute, 37°C) (A) and[3H]TPP� influx (46 nmol/L, 1 minute, 37°C) (B) were determined.Values are mean�SEM (n12). *P�0.05 vs all other values.
Flores et al D-Glucose Acutely Activates L-Arginine/NO Pathway 69

Ca2�, it is likely that rapid eNOS activation by D-glucose isCa2� independent, supporting recent observations of Ca2�-independent eNOS activation in HUVECs.37,38 D-Glucose–induced phosphorylation of eNOS at Ser1177, L-arginine trans-port, and TPP� influx were blocked by wortmannin,suggesting that the PI3-k pathway could be involved in theeffects of D-glucose. L-Arginine transport could be determi-nant for eNOS activity11; however, the possibility that theD-glucose–induced increase of L-citrulline production wasdue to elevated L-arginine transport seems unlikely, inasmuchas D-glucose–induced NO synthesis was unaltered in theabsence of extracellular L-arginine.
L-NAME blocked D-glucose–increased L-arginine trans-port and NO synthesis in HUVECs. This inhibitor does notalter basal L-arginine transport in the endothelium2–4,39; thus,NO most likely mediates changes in L-arginine transport, assuggested in bovine aortic endothelium.40 This result issimilar to that found in HUVECs from patients with gesta-tional diabetes; L-arginine transport in these cells is increasedconcomitantly with membrane hyperpolarization and NOsynthesis, and this increase is inhibited by blocking NO
synthase.2 NO causes membrane hyperpolarization in theendothelium,3,36 and NO (from SNAP) has been shown tocause comparable increases in TPP� influx and L-argininetransport and to cause membrane hyperpolarization to thosecaused by D-glucose, although SNAP treatment did notfurther enhance the effects of high D-glucose, in HUVECs.These findings support the hypothesis that NO acutely mod-ulates L-arginine transport by a mechanism that involvesmembrane hyperpolarization.
NO-altered K� channel activity may occur by both indirectmechanisms via cGMP and direct NO action on channels.36
Figure 5. Lack of effect of calphostin C in the effect ofD-glucose on L-arginine transport. A, PKC activity in cytosolic(open symbols) and membrane (filled symbols) fractions fromHUVECs preincubated (30 minutes) with Krebs solution without(�, �) or with 100 nmol/L calphostin C (▫, �). Cells were incu-bated (30 seconds) in 25 mmol/L D-glucose in the absence orpresence of calphostin C. B, Time-course effect of 25 mmol/LD-glucose on L-arginine transport (100 �mol/L, 1 minute, 37°C)(as in panel A) in Krebs solution without (�) or with 100 nmol/Lcalphostin C (�). Values are mean�SEM (n18). *P�0.05 vs allother values.
Figure 6. NO and PI3-k involvement in effect of D-glucose onp42/44mapk phosphorylation. A, Immunoblot for phosphorylatedp44mapk (p44�P) and p42mapk (p42�P) and nonphosphorylatedp44mapk (p44) or p42mapk (p42) in HUVECs preincubated (30 min-utes) with 5 mmol/L D-glucose in the absence or presence ofPD-98059 and then exposed (2 minutes) to 5 or 25 mmol/LD-glucose. B, Effect of SNAP on p42/p42mapk in HUVECs in5 mmol/L D-glucose. C, Effect of wortmannin on p42/p42mapk inHUVECs (as in panel A). Data are representative of similarresults in 17 cell cultures.
70 Circulation Research January 10/24, 2003

Our results show that the D-glucose increases in L-argininetransport and TPP� influx were mimicked by dbcGMP andblocked by the PKG inhibitor KT-5823. In addition, D-glucose–induced membrane hyperpolarization was alsoblocked by KT-5823. Thus, modulation of ion channelactivity (and hence, L-arginine transport) could be due to theactivation of PKG downstream from NO synthesis.
PKC activity is increased in subjects with diabetes mellitusor in endothelium chronically exposed to highD-glucose.12,16,17 Activation of diacylglycerol/phorbol ester–sensitive PKC isozymes activates eNOS and the NO-dependent increased p42/p44mapk phosphorylation inHUVECs exposed for 24 hours to high D-glucose.5 However,25 mmol/L D-glucose for 1 to 5 minutes did not alter PKCactivity in this cell type, suggesting that the rapid D-glucoseeffect on L-arginine transport was PKC independent. BecauseD-glucose induces a rapid (2-minute) p42/p44mapk phosphor-ylation and because inhibition of the p42/p44mapk phosphory-lation by PD-98059 also inhibits the D-glucose increase inTPP� influx and L-arginine transport, it is likely that p42/p44mapk activation is involved in this pathway. Activation ofp42/44mapk requires PI3-k activity in HUVECs.41 Our resultsshow that D-glucose–induced p42/44mapk phosphorylation isblocked by wortmannin, suggesting that the D-glucose effectrequires PI3-k activity in HUVECs. SNAP-increased p42/p44mapk phosphorylation and L-arginine transport wereblocked by PD-98059, complementing results showing thatNO, via cGMP, causes rapid p42/p44mapk phosphorylation inthe endothelium.4,42 Because D-glucose–induced and NO-induced membrane hyperpolarization are blocked by PD-98059, p42/p44mapk activation could modulate ion channelactivity and L-arginine transport in HUVECs.
Elevated D-glucose leads to overproduction of oxygen-derived free radicals in several cell types.11,16,17 We foundthat SOD activity in HUVECs was unaltered by 25 mmol/LD-glucose and that SOD or �-tocopherol did not block theeffects of D-glucose, suggesting that short-term incubationwith elevated D-glucose would not generate enough oxygen-derived free radicals to induce changes in the L-arginine/NOpathway in HUVECs.
The present study has established that high D-glucoserapidly activates the L-arginine/NO pathway in HUVECs.The effect of D-glucose involves PI3-k–dependent, but PKC-independent and intracellular Ca2�–independent, eNOS andp42/44mapk activation. These results complement previousobservations in animal models in which elevated plasmaD-glucose results in a rapid (seconds to minutes) vasodilata-tion8–10 and observations of D-glucose–induced vasodilatationin humans.7 Local NO synthesis could be one mechanism bywhich rapid alterations in plasma D-glucose result in vasodi-latation and may have important implications in diabeticpatients, in whom plasma D-glucose concentrations maychange rapidly.11,12,16,17
AcknowledgmentsThis study was supported by Fondo Nacional de Ciencia y Tec-nología (FONDECYT 1000354 and 7000354), Universidad de Con-cepción (DIUC 201.084.003-1) (Chile), and The Wellcome Trust(United Kingdom). P. Casanello, J. Parodi, and C. Aguayo hold BecaDocente Universidad de Concepción and CONICYT-PhD (Chile)
fellowships. We thank the midwives of the Hospital Regional-Concepción (Chile) labor ward for the supply of umbilical cords andIsabel Jara for secretarial assistance.
References1. Moncada S, Palmer RMJ, Higgs EA. Nitric oxide: physiology, patho-
physiology and pharmacology. Pharmacol Rev. 1991;43:109–142.2. Sobrevia L, Cesare P, Yudilevich DL, Mann GE. Diabetes-induced acti-
vation of system y� and nitric oxide synthase in human endothelial cellsassociation with membrane hyperpolarization. J Physiol (Lond). 1995;489:183–192.
3. Casanello P, Sobrevia L. Intrauterine growth retardation is associatedwith reduced activity and expression of the cationic amino acid transportsystems y�/hCAT-1 and y�/hCAT-2B, and lower activity of nitric oxidesynthase in human umbilical vein endothelial cells. Circ Res. 2002;91:127–134.
4. Sobrevia L, Nadal A, Yudilevich DL, Mann GE. Activation of L-argininetransport (system y�) and nitric oxide synthase by elevated glucose andinsulin in human endothelial cells. J Physiol (Lond). 1996;490:775–781.
5. Montecinos VP, Aguayo C, Flores C, Wyatt AW, Pearson JD, Mann GE,Sobrevia L. Regulation of adenosine transport by D-glucose in humanfetal endothelial cells: involvement of nitric oxide, protein kinase C andmitogen-activated protein kinase. J Physiol (Lond). 2000;529:777–790.
6. Cosentino F, Hishikawa K, Katusic ZS, Lüscher TF. High glucoseincreases nitric oxide synthase expression and superoxide anion gen-eration in human aortic endothelial cells. Circulation. 1997;96:25–28.
7. Beckman JA, Goldfine AB, Gordon MB, Garrett LA, Creager MA.Inhibition of protein kinase C� prevents impaired endothelium-dependentvasodilation caused by hyperglycemia in humans. Circ Res. 2002;90:107–111.
8. Duckrow RB. Decreased cerebral blood flow during acute hypergly-caemia. Brain Res. 1995;703:145–150.
9. Wascher TC, Bachernegg M, Kickenweiz A, Stark G, Stark U, Toplak H,Graier WF. Involvement of the L-arginine-nitric oxide pathway inhyperglycaemia-induced coronary artery dysfunction of isolated guineapig hearts. Eur J Clin Invest. 1996;26:707–712.
10. Cipolla MJ, Porter JM, Osol G. High glucose concentration dilate cerebralarteries and diminish myogenic tone through an endothelial mechanism.Stroke. 1997;28:405–411.
11. Sobrevia L, Mann GE. Dysfunction of the nitric oxide signalling pathwayin diabetes and hyperglycaemia. Exp Physiol. 1997;82:1–30.
12. Pan M, Wasa M, Lind DS, Gertler J Abbott W, Souba WW. TNF-stimulated arginine transport by human vascular endothelium requiresactivation of protein kinase C. Ann Surg. 1995;221:590–601.
13. Racké K, Hey C, Mössner J, Hammermann R, Stichnote C, Wessler I.Activation of L-arginine transport by protein kinase C in rabbit, rat andmouse alveolar macrophages. J Physiol (Lond). 1998;551:813–825.
14. Haneda M, Araki S, Togawa M, Sugimoto T, Isono M, Kikkawa R.Mitogen-activated protein kinase cascade is activated in glomeruli ofdiabetic rats and glomerular mesangial cells cultured under high glucoseconditions. Diabetes. 1997;46:847–853.
15. Natarajan R, Scott S, Bai W, Yernerni K, Nadler J. Angiotensin IIsignalling in vascular smooth muscle cells under high glucose conditions.Hypertension. 1999;33:378–384.
16. Pieper GM. Review of alterations in endothelial nitric oxide production indiabetes. Hypertension. 1998;31:1047–1060.
17. De Vriese AS, Verbeuren TJ, Van de Voorde J, Lameire NH, VanhouttePM. Endothelial dysfunction in diabetes. Br J Pharmacol. 2000;130:963–974.
18. Flores C, Sobrevia L. Acute effect of D-glucose on L-arginine transportand nitric oxide synthesis in human fetal endothelium. J Physiol (Lond).2000;523:104P. Abstract.
19. Kolch W. Meaningful relationships: the regulation of the Ras/Raf/MEK/ERK pathway by protein interactions. Biochem J. 2000;351:289–305.
20. Grider JR. Interplay of VIP and nitric oxide in regulation of thedescending relaxation phase of peristalsis. Am J Physiol. 1993;264:G334–G340.
21. Kobayashi E, Ando K, Nakano H, Iida T, Ohno H, Morimoto M, TamaokiT. Calphostins (UCN-1028), novel and specific inhibitors of proteinkinase C, I: fermentation, isolation, physico-chemical properties andbiological activities. J Antibiot (Tokyo). 1989;42:1470–1474.
22. Holzmann S, Kukovetz WR, Braida C, Poch G. Pharmacological inter-action experiments differentiate between glibenclamide-sensitive K�
Flores et al D-Glucose Acutely Activates L-Arginine/NO Pathway 71

channels and cyclic GMP as components of vasodilation by nicorandil.Eur J Pharmacol. 1992;29:1–7.
23. Katnik C, Adams DJ. Characterization of ATP-sensitive potassiumchannels in freshly dissociated rabbit aortic endothelial cells. Am JPhysiol. 1997;272:H2507–H2511.
24. Isenovic E, Muniyappa R, Milivojevic N, Rao Y, Sowers JR. Role ofPI3-kinase in isoproterenol and IGF-1 induced ecNOS activity. BiochemBiophys Res Commun. 2001;285:954–958.
25. Schilling WP. Effect of membrane potential on cytosolic calcium ofbovine aortic endothelial cells. Am J Physiol. 1989;257:H778–H784.
26. Misra HP. Adrenochrome assay. In: Greenwald RA, ed. Handbook ofMethods for Oxygen Radical Research. Boca Raton, Fla: CRC Press;1989:237–241.
27. Ganz MB, Seftel A. Glucose-induced changes in protein kinase C andnitric oxide are prevented by vitamin E. Am J Physiol. 2000;278:E146–E152.
28. Harper JF. Peritz’ F test: BASIC program of a robust multiple comparisontest for statistical analysis of all differences among group means. ComputBiol Med. 1984;14:437–445.
29. Arancibia Y, Sobrevia L. Transport of L-arginine mediated by system y�Lin human fetal endothelial cells. J Physiol (Lond). 1999;517:47P.Abstract.
30. Sala R, Rotoli BM, Colla E, Visigalli R, Parolari A, Bussolati O, GazzolaGC, Dall’Asta V. Two-way arginine transport in human endothelial cells:TNF-� stimulation is restricted to system y�. Am J Physiol. 2002;282:C134–C143.
31. Devés R, Boyd CAR. Transporters for cationic amino acids in animalcells: discovery, structure and function. Physiol Rev. 1998;78:487–545.
32. Bussolati O, Sala R, Astorri A, Rotoli BM, Dall’Asta V, Gazzola GC.Characterization of amino acid transport in human endothelial cells. Am JPhysiol. 1993;265:C1006–C1014.
33. Irie K, Tsukahara F, Fujii E, Uchida Y, Yoshioka T, He WR, ShitashigeM, Murota S, Muraki T. Cationic amino acid transporter-2 mRNAinduction by tumor necrosis factor-� in vascular endothelial cells. EurJ Pharmacol. 1997;339:289–293.
34. Closs EI, Graf P, Habermeier A, Föstermann U. Interference of L-arginineanalogues with L-arginine transport mediated by the y� carrier hCAT-2B.Nitric Oxide. 1997;1:65–73.
35. Nilius B, Droogmans G. Ion channels and their functional role in vascularendothelium. Physiol Rev. 2001;81:1415–1459.
36. Sobey CG. Potassium channel function in vascular disease. ArteriosclerThromb Vasc Biol. 2001;21:28–38.
37. Dimmeler S, Fleming I, Fisslthaler B, Hermann C, Busse R, Zeiher AM.Activation of nitric oxide synthase in endothelial cells by Akt-dependentphosphorylation. Nature. 1999;399:601–605.
38. Shaul PW. Regulation of endothelial nitric oxide synthase: location,location, location. Annu Rev Physiol. 2002;64:749–774.
39. Bogle RG, Moncada S, Pearson JD, Mann GE. Identification of inhibitorsof nitric oxide synthase that do not interact with the endothelial cellL-arginine transporter. Br J Pharmacol. 1992;105:768–770.
40. Ogonowski AA, Kaesemeyer WH, Jin L, Ganapathy V, Leibach FH,Caldwell RW. Effects of NO donors and synthase agonists on endothelialcell uptake of L-Arg and superoxide production. Am J Physiol. 2000;278:C136–C146.
41. Lal BK, Varma S, Pappas PJ, Hobson RW II, Duran WN. VEGFincreases permeability of the endothelial cell monolayer by activation ofPKB/akt, endothelial nitric-oxide synthase, and MAP kinase pathways.Microvasc Res. 2001;62:252–262.
42. Parenti A, Morbidelli L, Cui XL, Douglas JG, Hood JD, Granger HJ,Ledda F, Ziche M. Nitric oxide is an upstream signal of vascular endo-thelial growth factor-induced extracellular signal-regulated kinase 1/2activation in postcapillary endothelium. J Biol Chem. 1998;273:4220–4226.
72 Circulation Research January 10/24, 2003





























