Embed Size (px)
Citation preview

Somatic Cell Genetics, VoL 8, No. 3, 1982, pp 289-305
a2-Macroglobulin Production by Cultured Human Fibroblasts
Jane E. Brissenden and Diane W. Cox
Research Institute, The Hospital for Sick Children, 555 University Avenue, Toronto, Ontario, Canada, M5G 1X8; and Departments of Medical Genetics, Paediatrics, and Medical Biophysics, University of Toronto
Received 15 September 1981--Final 11 December 1981
Abstraet--Alpha2-macroglobulin (a2M), a high-molecular-weight plasma protease inhibitor has been shown, by both immunological and functional methods, to be produced by cultured adult lung fibroblasts. Cultured skin fibroblasts synthesized approximately one tenth as much a2M as lung fibroblasts. This quantitative difference in a2M production was also demon- strated in fibroblasts of isogenic origin. There was no difference in the amount of a2M produced between adult and fetal fibroblasts o f the same tissue type (i.e., o f lung or of skin origin), a2M was produced in culture during log-phase growth as well as at confluence. Two other plasma protease inhibitors, Cl-esterase inhibitor and a substance immunologically cross- reacting with human inter-a-trypsin inhibitor, were also made by the cultured fibroblasts. Plasma protease inhibitors not detectable in culture supernatants were al-antitrypsin, al-antichymotrypsin, and antithrombin III.
INTRODUCTION
Many aspects of metabolism in mammals have been shown to be controlled by proteases and protease inhibitors (1, 2). These include physio- logical functions mediated by enzyme cascades such as blood coagulation, complement activation and fibrinolysis, and kinin generation (2). Proteases convert zymogens into biologically active products or inactivate biologically active proteins, degrade superfluous or defective protein molecules, and are involved in tissue defence, for example, by inactivating and degrading bacterial proteases (3).
One plasma protease inhibitor, ai-antitrypsin (alAT), is involved in protection of lung elastin integrity by inactivating leukocyte elastase; genetic
289
0098~366/82/0500~289503.00/0 �9 1982 Plenum Publishing Corporation

290 Brissenden and Cox
variants of a~AT leading to lowered serum levels and/or activities of a~AT are detrimental to lung function. Serum levels of other plasma protease inhibitors are not known to be altered in association with lung disease (4), and their overall physiological significance is not understood.
a2M is the second most abundant plasma protease inhibitor on a molar basis (5). a2M has a wide range of substrate specificity and a unique capacity to inhibit endopeptidase activity against high-molecular-weight substrates while retaining such activity against low-molecular-weight substrates (6). This last property could allow controled local regulation of proteolysis. Although the high molecular weight of a2M (725,000 daltons) makes it difficult for the molecule to cross the capillary wall in the absence of previous inflammation (7), thus downplaying its possible significance in primary lung defence, synthesis of a2M in the tissue itself would circumvent this transport problem.
Several recent reports have presented conflicting evidence (8-16) for a2M production by cultured human cells of various origins: monocytes (8), lymphocytes (9, 10), alveolar maerophages (11), and fibroblasts (12-14). Reports that a2M is produced by cultured cells have been questioned because of the cross-reactivity with antiserum to human o~2M of bovine a2M used in the culture growth medium (17) and because of the known endocytosis and recycling of bovine o~2M by fibroblasts in culture (15, 18). Furthermore, one investigator, measuring protease inhibitor production by function, failed to detect a2M or any other known plasma protease inhibitor in supernatants from adult and fetal lung cultures (16). a2M production has been reported to decrease after first passage for cultures of tissues other than of embryonic lung (14).
Some proteins such as fibronectin have been demonstrated to be made throughout culture growth cycle (19), while others such as collagenase are produced only at confluence (20). Such changes in the timing of synthesis might contribute to the variability of the results of studies of a2M production in culture and could as well have functional significance.
Our studies were designed to determine if human a2M is produced by human lung fibroblast cultures of adult origin and to determine if fibroblast cultures of other sources (embryonic lung and adult or embryonic skin) make a2M. We tested for possible effects of culture variables (passage, growth cycle, tissue of origin, and ontological origin) on such production.
We also assayed culture supernatants immunologically for the presence of five other plasma protease inhibitors to determine if they too might be produced by cultured cells.
MATERIALS AND METHODS
Materials. Cell strains used in this study are listed in Table 1. All cultures were tested for Mycoplasma by direct culture and by the UDPase

~2-Maeroglobulin Production 291
Table 1. Cell Strains Used
Figure 2 Strain identification
number number Source Comment
Adult lung L J001 1 T.G.H. a Grossly normal tissue
near carcinoma L J002 2 T.G.H. Grossly normal tissue
near carcinoma L J003 3 T.G.H. Grossly normal tissue
near carcinoma isogenic to S J003
L J004 4 T.G.H. Grossly normal tissue near hamartoma isogenic to S J004
Fetal lung MRC5 1 S. Goldstein, McMaster U. 172 2 H.S.C. b Dept. of Genetics HEL299 3 American Type Culture
Collection LF6 4 D. Hoar, U. of Calgary WI38 5 American Type Culture
Collection Adult skin
160 1 H.S.C. Dept. of Genetics 526 2 H.S.C. Dept. of Genetics 62 3 H.S.C. Dept. of Genetics 527 4 H.S.C. Dept. of Genetics 55 5 H.S.C. Dept. of Genetics S J003 6 T.G.H. S J004 7 T.G.H.
Fetal skin DET551
Isogenic to SF6
Isogenic to L J003 Isogenic to L J004
1 American Type Culture Collection
407 2 H.S.C. Dept. of Genetics 167 3 H.S.C. Dept. of Genetics SF6 4 D. Hoar, U. of Calgary Isogenic to LF6
aToronto General Hospital, Division of Surgical Pathology. bHospital for Sick Children, Toronto.
indirect assay (21). One subcul ture was contaminated and was discarded.
Cul tures were established and propagated f rom minced tissue as described by
Hayfl ick and Moorhead (22) in feeding medium consisting of a l p h a - M E M
( G I B C O ) ( a - M E M ) with 15% fetal cal f serum (FCS) , unless otherwise
stated. Quantitative Methods. Two pre l iminary assays of spent supernatants
(one from embryonic lung fibroblasts and one from adult lung fibroblasts)
were performed by double immunodiffusion (23). a2M concentrat ions in all
remaining supernatants were measured immunological ly by e lec t ro immu-
noassay (EIA) (24), sensitivity l imit 1.4 ~ g / m l , or by radio immunoassay

292 Brissenden and Cox
(RIA) (25), sensitivity limit 5 ng/ml, a2M levels were also measured by function (13), sensitivity limit 10 ~tg/ml.
Supernatants to be assayed by EIA or for functional activity were concentrated 5- to 100-fold by ultrafiltration with a Minicon B15 macrosolute concentrator (Amicon Corporation) or by pressure ultrafiltration with an Amicon ultrafiltration apparatus and a PM 10 membrane. In both cases the membranes had a molecular weight cutoff of 10,000 daltons. Supernatants and cells were stored frozen at - 2 0 ~ until assayed.
To assay intracellular aEM , cells were washed in medium minus FCS then sonicated at a cell density of 2.5 • 10 7 cells/ml in 0.075 M sucrose in 0.02 M sodium phosphate buffer, pH 7.4, using a Fisher Sonic Dismembrator, model 300. OtEM concentrations were measured by RIA.
aRM for RIA was purified by the method of Virca et al. (26) with minor modifications. Whole serum of haptoglobulin 1-1 genotype (typed by starch gel electrophoresis) (27) was applied to a 1.6 x 84-cm column containing sepharose CL6B linked to the group-specific absorbent Cibacron-blue (Phar- macia) and eluted with 0.05 M TrisHC1, pH 8.0, at a flow rate of 3 ml/hr. ohM was eluted in the void volume. Eluted a2M (>95% pure) was stored at - 70~ An impure a2M fraction was further purified by molecular sieving on sepharose CL4B. The resultant aEM was shown to be >95% pure by immunoelectrophoresis and agarose gel electrophoresis (28).
OtEM was iodinated by oxidation with BIORAD Enzymobeads to a specific activity of approximately 1 ~tCi/#g CtEM. Goat antiserum against human ~2M was obtained from Atlantic Antibodies (Westbrook, Maine) and adsorbed against 60-80 mg/ml lyophilized bovine serum until it no longer showed cross-reactivity with bovine teEM by double immunodiffusion. For the RIA, 50 ~tl adsorbed anti-a2M diluted 1/8000 was incubated for 48 h with 100 /~l sample or standard in test medium and 100 ~tl nonimmune goat serum diluted 1/1000. Two days later 10 ng, [125I]o~2M in 25 /~l 0.05 M sodium phosphate buffer, pH 7.2, was added and the mixture incubated for 1 h, after which 200 #1 anti-goat immunoglobulin diluted 1/36 was added. Sixteen to 24 h later, the mixture was centrifuged at 11,000 rpm for 5 min in a Beckman microfuge, the supernatant aspirated, and the ~25I in the precipitate was counted in a Beckman gamma counter. Standard curves from 5-100 ng/ml o~2M were prepared using dilutions of a standard pool of serum from 20 individuals, calibrated from a reference serum obtained from Atlantic Anti- bodies. Dilutions of feeding medium up to 1:3 were tested for cross-reacting material by RIA.
Cultures were grown in medium containing bovine serum. Therefore, other precautions were taken, besides adsorption of cross-reacting material from the first antiserum, to avoid misinterpretation of results due to endocyto- sis and recycling of bovine a2M, which shows some cross-reactivity with

~2-Macroglobulin Production 293
human a2M. Before harvesting of supernatants or cells, all cells in culture were washed twice with medium minus FCS and were fed for two days with medium minus FCS plus bovine serum albumin (test medium). Assay methods were assessed as outlined in Results. Supernatants from lung fibroblast cultures were diluted 10-fold with test medium before analysis.
a2M activity was quantitated functionally by its specific capacity to prevent inhibition by soybean trypsin inhibitor of trypsin, which hydrolyses the small-molecular-weight substrate, toluene-sulfonyl-methyl ester (13, 29). Four hundred microliters of sample mix were added to 300 ~tl TAME for assay. The protected trypsin activity was detected spectrophotometrically by change in light absorbance at 247 nm. The change in light absorbance directly correlated with a2M concentration. A standard curve, from 10 to 100 #g/ml a2M, was prepared using dilutions of serum from a patient genetically deficient (PI null homozygote) for the other major plasma protease inhibitor, alAT , to avoid interference from an interaction in trypsin binding between OtlAT and a2M. The concentrated supernatant samples were centrifuged at 11,000 rpm for 5 min before assay.
When assayed for function, some batches of bovine serum showed detectable functional a2M which did not heat inactivate at 55~ so in each experiment all cultures of both lung and skin origin were fed with the same batch of FCS. This procedure standardized the amount of functional bovine a2M fed to all cultures in each set of experiments.
Other protease inhibitors, a~AT, a~-antichymotrypsin (alX), antithrom- bin III (AT III), inter-a-trypsin inhibitor (laD, and Cl-esterase inactivator (inhibitor) (C1 INA), were quantitated by electroimmunoassay in the culture supernatants concentrated 5- to 25-fold. The sensitivity levels of the assays were 0.4 izg/ml for alAT, 0.9 #g/ml for alX, 1.6 izg/ml for AT III, 1.1 #g/ml for Iod, and 2.5 #g/ml for C1 INA.
a2M Production in Relation to Culture Variables. The effect of passage number on a2M production was examined in explants by collecting spent feeding media at each feeding of the explant and at least two subsequent passages. After vigorous cultures were established, further testing, described in the next section, was done. In these tests cells were grown for the last 48 h in test medium which was then collected and analyzed.
Explants were usually fed at least once before subculturing at time intervals which depended on growth, resulting in an "early" explant superna- tant which was visibly blood-stained in the case of lung explants, and a "late" explant supernatant with no visible blood contamination. The "early" explant supernatant was usually taken before there was an appreciable amount of cellular outgrowth from the explant. Average feeding intervals were 14 days. Alveolar macrophages which may produce aEM (1 l) are removed by trypsin- ization and/or growth for longer than 3 weeks in culture (30).

294 Brissenden and Cox
To investigate possible growth cycle effects on ~2M production, one postnatal skin strain (# 160) and one embryonic lung fibroblast strain (#172) were plated at cell densities of approximately 2 x 105 and 1 x 105 cells/P100 Falcon tissue culture dish, respectively, in feeding medium. Duplicate plates were harvested at 24-h intervals until confluence was reached, o~2M superna- tant concentrations were determined by RIA and cells were counted in an ElectroZone Celloscope (Particle Data Inc.).
e~2M Production by Fibroblasts of Different Origin. Production of o~2M was compared for human lung and skin cells of fetal and of adult origin. Three sets of these fibroblasts were of isogenic origin but of different tissue origin. P100 petri dishes of confluent cells were trypsinized, split 1:8, and fed three days later with feeding medium. Two days later, the medium was removed, cells were washed twice with a-MEM minus FCS, fed with 5 ml of test medium, and harvested two days later. Each fibroblast strain was cultured and analyzed at least three times, with the exception of MRC5 which was only tested once. One representative plate per experiment of each cell line was trypsinized and those cells counted with a Coulter counter.
RESULTS
Assessment of Immunological Methods. Since bovine a2M stored in the cells can be excreted and detected as human ohM, we established that our assay methods (EIA and RIA) did distinguish a2M from the two sources. Figure 1A illustrates the effect of cross-reactivity of bovine ohM in double immunodiffusion. Strains of human lung (#172) and humon skin (#160) fibroblasts both test as positive for a2M by double immunodiffusion. However, EIA detects human o~2M only in the supernatant from the lung fibroblast strain (Fig. 1C). Figure 1B demonstrates that a2M in human and bovine sera could be differentiated by EIA by the marked differences in appearance of their precipitation peaks using antiserum to human a2M. Precipitation peaks from wells containing human serum alone are darkly stained and well defined. Those from wells containing only bovine serum are very faint and diffuse. Double precipitation peaks with the appearance of both human and bovine peaks arise from wells containing mixtures of human and bovine serum. The heights of these peaks were proportional to the relative amounts of human and bovine serum present. The presence of bovine a2M did not interfere with human ~2M quantitation, as verified by studies using known but varied mixtures of human and bovine sera.
For RIA a mixture of 75% feeding medium and 25% test medium, tested with adsorbed antiserum, showed no detectable ~2M (<5 ng/ml). Undiluted feeding medium usually showed no detectable ~zM, but occasionally cross-
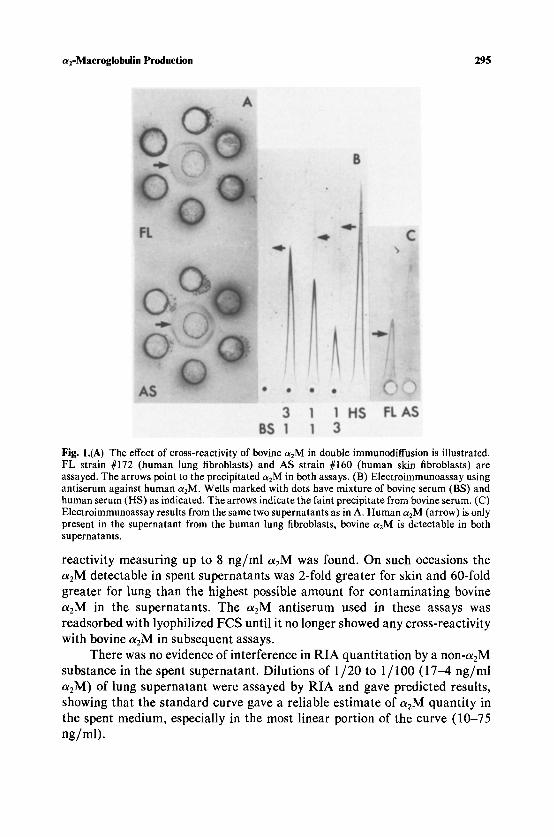
a2-Macroglobulin Production 295
Fig. 1.(A) The effect of cross-reactivity of bovine a2M in double immunodiffusion is illustrated. FL strain #172 (human lung fibroblasts) and AS strain #160 (human skin fibroblasts) are assayed. The arrows point to the precipitated a2M in both assays. (B) Electroimmunoassay using antiserum against human a~M. Wells marked with dots have mixture of bovine serum (BS) and human serum (HS) as indicated. The arrows indicate the faint precipitate from bovine serum. (C) Electroimmunoassay results from the same two supernatants as in A. Human a2M (arrow) is only present in the supernatant from the human lung fibroblasts, bovine c~2M is detectable in both supernatants.
reactivity measuring up to 8 ng /ml teE M was found. On such occasions the ot2M detectable in spent supernatants was 2-fold greater for skin and 60-fold greater for lung than the highest possible amount for contaminat ing bovine a2M in the supernatants. The OtEM antiserum used in these assays was readsorbed with lyophilized FCS until it no longer showed any cross-reactivity with bovine teEM in subsequent assays.
There was no evidence of interference in R I A quanti tat ion by a non-a2M substance in the spent supernatant. Dilutions of 1/20 to 1/100 (17-4 n g / m l a2M) of lung supernatant were assayed by R I A and gave predicted results, showing that the s tandard curve gave a reliable estimate of a2M quanti ty in the spent medium, especially in the most linear portion of the curve (10-75 ng/ml) .

296 Brissenden and Cox
a2M Production and Culture Variables. Initial measurements to deter- mine whether fibroblasts produced ot2M in culture at various times after explant were necessarily done in feeding medium, as we were then establishing growing cultures. All measurements subsequent to significant cell growth were done in test medium. Results are shown in Table 2.
No human a2M was detected in media from the early adult lung explants while they were still blood stained and before appreciable cellular outgrowth had occurred. Human a2M was detectable at least once in each adult lung fibroblast strain tested from late explant onward. The amounts detected varied from about 5 #g/ml to more than 46 #g/ml collected over about two weeks (0.3-2/~g/ml/48 h) and showed no consistent pattern of decline after passage.
LF6, the embryonic lung fibroblast culture established in our laboratory from explant showed similar results (Table 2). These data indicate that a2M is produced by cultured lung fibroblasts and that there is no dramatic decline in human o~2M production between 1 ~ cultures and up to 7 passages of these cultures.
Primary adult and fetal skin cultures also showed oLzM production, but at lower rates than those of lung fibroblast cultures. This production remained at the same level after subculture (Table 2).
T a b l e 2. C o n c e n t r a t i o n o f a 2 - M a c r o g l o b u l i n in S u p e r n a t a n t s of Explan t s a n d F ib rob l a s t Cu l tu re s a f t e r P a s s a g e a
S t a g e of g r o w t h
E a r l y La t e P a s s a g e n u m b e r Cell s t ra in
L u n g L J001 L J 0 0 2
L J003 L J 0 0 4
L F 6 d
Skin ~ S J 0 0 3 S J 0 0 4 S F 6
exp lan t exp lan t 1 2 3 4 5 6 7 8
Id N D b 43 5 9 > 4 6 25 ~ I - - - - - - N D N D 17 12 5 > 4 6 26 ~ - - 2 - -
p . . . . . . . . . . . . . I
N D 7 N D N D [ 1 0.6 - - 0.3 - - < 0 . 0 0 5 N D > 4 6 N D 20 [ 0.2 0.8 0.7 - - - - - -
r . . . . . J
0 .016 0.03 0 .0401 - - 2 - - 1 4 - - - - I
< 0 . 0 0 5 < 0 . 0 0 5 <0 .005~ < 0 . 0 0 5 0 . 0 0 4 - - < 0 . 0 0 5 - - < 0 . 0 0 5 0.03 <0 .0051 - - - - 0.01 - - < 0 . 0 0 5 - - - -
0 .012 < 0 . 0 0 5 0 .0061 - - 0.09 0.01 0 .04 - - - - - - I
a C u l t u r e s to the lef t o f the d a s h e d line were g r o w n in feed ing m e d i u m only for a n ave rage Of two weeks. Those to the r igh t were g r o w n in tes t m e d i u m for the last 48 h in cul ture ; a 2 M c o n c e n t r a t i o n s in t he test m e d i a were m e a s u r e d , c~2M c o n c e n t r a t i o n s a re expressed as ~tg/ml a n d a re the resul t o f one or more de t e rmina t ions .
b N o t de t ec t ab l e CNot tes ted dAssayed b y R I A . All o thers were a s sayed b y E IA .
oq-Maeroglobulin Production 297
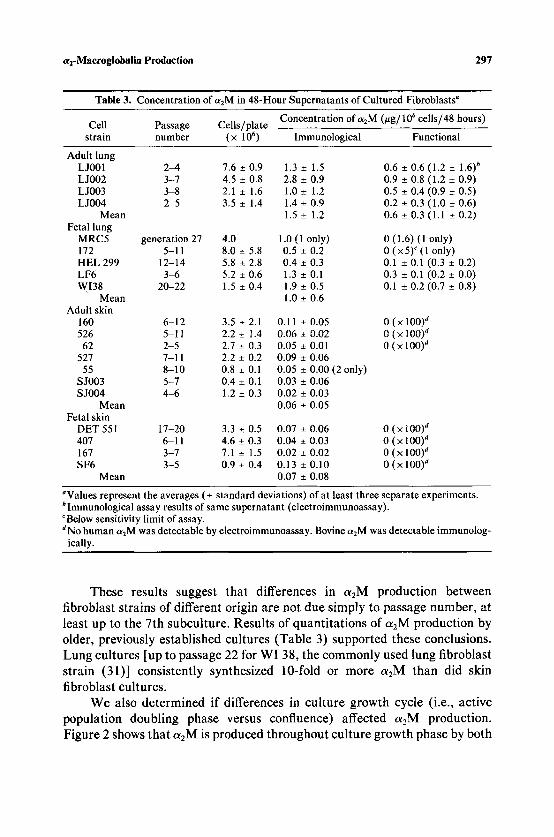
Table 3. Concentration of c~2M in 48-Hour Supernatants of Cultured Fibroblasts a
Concentration of azM (~tg/106 cells/48 hours) Cell Passage Cells/plate
strain number (x 1 0 6 ) Immunological Functional
Adult lung L J001 2~, 7.6 _+ 0.9 1.3 _+ 1.5 L J002 3-7 4.5 +_ 0.8 2.8 _+ 0.9 LJ003 3-8 2.1 _+ 1.6 1.0-+ 1.2 L J004 2-5 3.5 _+ 1.4 1.4 _+ 0.9
Mean 1.5 _+ 1.2 Fetal lung
MRC5 generation 27 4.0 1.0 (1 only) 172 5-11 8.0 _+ 5.8 0.5 _+ 0.2 HEL 299 12-14 5.8 • 2.8 0.4 _+ 0.3 LF6 3q5 5.2 _+ 0.6 1.3 _+ 0.1 WI38 20-22 1.5 _+ 0.4 1.9 _+ 0.5
Mean 1.0 _+ 0.6 Adult skin
160 6-12 3.5 _-!- 2.1 0.11 _+ 0.05 526 5-11 2.2 _+ 1.4 0,06 _+ 0.02
62 2-5 2.7 _+ 0.3 0.05 _+ 0.01 527 7-11 2.2 _+ 0.2 0.09 _+ 0.06
55 8-10 0.8 _+ 0.1 0.05 _+ 0.00 SJ003 5-7 0.4 +_ 0.1 0.03 _+ 0.06 S J004 4 ~ 1.2 _+ 0.3 0.02 +_ 0.03
Mean 0.06 _+ 0.05 Fetal skin
DET 551 17-20 3.3 _+ 0.5 0.07 _+ 0.06 407 6-11 4,6 _+ 0.3 0.04 _+ 0.03 167 3-7 7.1 _+ 1.5 0.02 _+ 0.02 SF6 3-5 0.9 _+ 0.4 0.13 _+ 0.10
Mean 0.07 +_ 0.08
(2 only)
0.6 -+ 0.6 (1.2 +_ 1.6) b 0.9 _+ 0.8 (1.2 _+ 0.9) 0.5 -+ 0.4 (0.9 --+ 0.5) 0.2 _+ 0.3 (1.0 -+ 0.6) 0.6 -+ 0.3 (1.1 _+ 0.2)
0 (1.6) (1 only) 0 (x5) c (1 only) 0.1 _+ 0.1 (0.3 +_ 0.2) 0.3 _+ 0.1 (0.2 + 0.0) 0.1 + 0.2 (0.7 _+ 0.8)
0 (• 100) a 0 (x 100) a 0 (• 100) a
0 (x 100) ~ 0 (x 100) ~ 0 ( x 100) a 0 (x 100) d
~Values represent the averages (_+ standard deviations) of at least three separate experiments. bimmunological assay results of same supernatant (electroimmunoassay). c Below sensitivity limit of assay. aNo human a2M was detectable by electroimmunoassay. Bovine c~2M was detectable immunolog-
ically.
These results suggest that differences in a2M production between fibroblast strains of different origin are not due simply to passage number, at least up to the 7th subculture. Results of quantitations of a2M production by older, previously established cultures (Table 3) supported these conclusions. Lung cultures [up to passage 22 for WI 38, the commonly used lung fibroblast strain (31)] consistently synthesized 10-fold or more a2M than did skin fibroblast cultures.
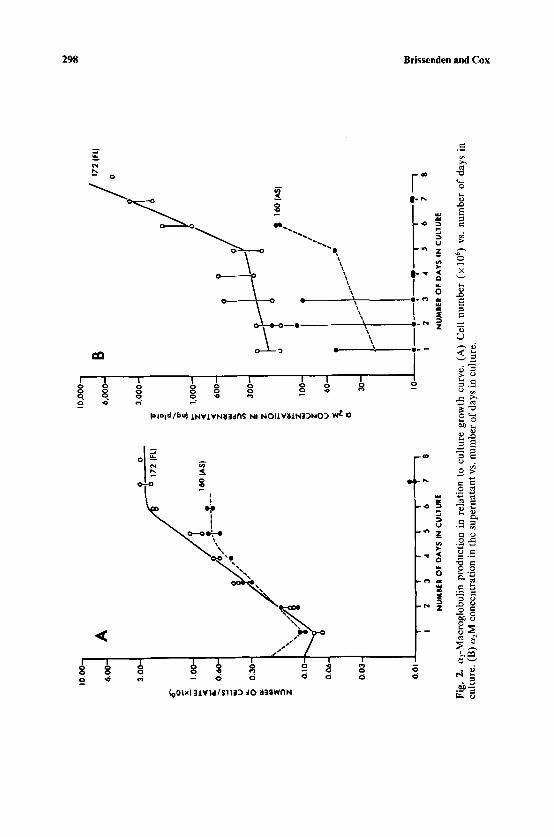
We also determined if differences in culture growth cycle (i.e., active population doubling phase versus confluence) affected a2M production. Figure 2 shows that a2M is produced throughout culture growth phase by both
10.0
0-
6.00
- 3~ %
x ,,
!.
O0-
w
.
8.
0.60
- w
u.m
U
0.3
0-
z 0.
10-
0.0
6-
0,0
3-
A
~ o
;72 c
~
I 0,0
00 -
6,000- B
~/~
/0172 (FL)
3,00
0-
_m
o
1,00
0--
z
60
0-
z_
30
0-
z
�9
/ Z
I00
- �9
/
o /
v 60
- /
* }
" 0.
01
I I
I I
~ t
'~,
I 10
I
: ,~
I
1 'E
3
4 5
6 7
8 1
4 5
6 7
8
NU
MB
ER
OF
DA
YS
iN
CU
LTU
RE
N
UM
BE
R O
F D
AY
S I
N C
ULT
UR
E
Fig.
2.
a2-M
acro
glob
ulin
pro
duct
ion
in r
elat
ion
to c
ultu
re g
row
th c
urve
. (A
) C
ell
num
ber
(x 1
06) v
s. n
umbe
r of
day
s in
cu
lture
. (B
) a2M
con
cent
ratio
n in
the
supe
rnat
ant
vs. n
umbe
r of
day
s in
cultu
re.
ba
OC
g~
r o

a2-Macroglobulin Production 299
lung and skin fibroblasts and that the differences in the amount of a2M produced by lung and skin fibroblast cultures are not dependent on the cultures' stage of growth.
a2M Production by Fibroblasts of Different Origin. We then measured the amount of a2M produced by fibroblasts of different origin to determine if ontological and/or tissue differences in source were associated with differ- ences in a2M production.
Preliminary results with EIA showed that supernatants from lung cultures had to be concentrated 5- to 25-fold (with possible losses of a2M on concentrator membranes) in order to detect human a2M and that human aRM was not detectable by EIA from skin fibroblasts even when concentrated. To increase sensitivity, we developed a RIA capable of measuring 5 ng/ml a 2 M ,
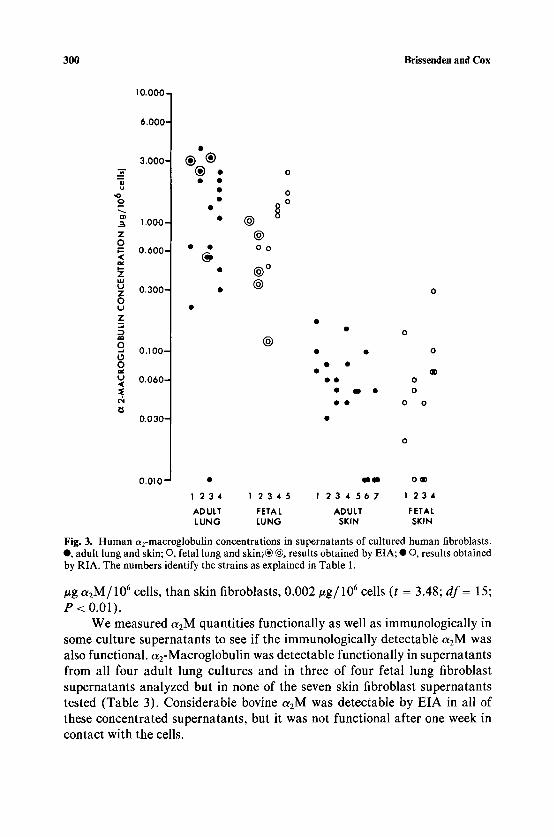
and we were then able to detect aEM in unconcentrated supernatants of both lung and skin fibroblast cultures, a2M quantities in supernatants from the lung and skin fibroblast cultures are shown in Table 3 and in Fig. 3. Each value in Table 3 is the average of quantitations from at least three experiments. Figure 3 shows the values from the individual experiments. EIA and RIA results correspond well, as shown in Fig. 3.
The values are expressed as #g a2M/106 cells/48 h. These measurements are more precise and comparable than are those of Table 2 because they were taken over a defned (48 h) time period from known numbers of cells grown under identical conditions in medium which did not contain FCS.
As seen in Figure 3, all lung fibroblast supernatants, with two exceptions, show immunologically detectable human a2M after 48 h in contact with the confluent fibroblasts. The mean concentration was 1.3 ~tg/106 cells (0.09 _+ 0.05% of total protein). Human a2M detected immunologically in adult and fetal skin fibroblast cultures was significantly lower (t = 7.08; df = 64; P < 0.001) with a mean of 0.06 #g/106 cells. This amount of a2M represented 0.003 _+ 0.002% of the total protein in skin fibroblast supernatants, 1/10 that of lung fibroblast supernatants. Isogenic cultures of different tissue origins retained these differences. There was no significant difference in human a2M levels between supernatants from lung fibroblasts of adult or fetal origin (t = 1.49; df = 31; P < 0.02) or between supernatants of skin fibroblasts of adult or fetal origin (t = 0.48; df = 31; P < 0.9). The consistent differences in a2M quantities detectable have confirmed the differences seen in Table 2 and suggest that there is a tissue-specific difference in the rate of production of human a2M. The results also suggest that there are no ontological differences in production rate of a2M by fibroblasts.
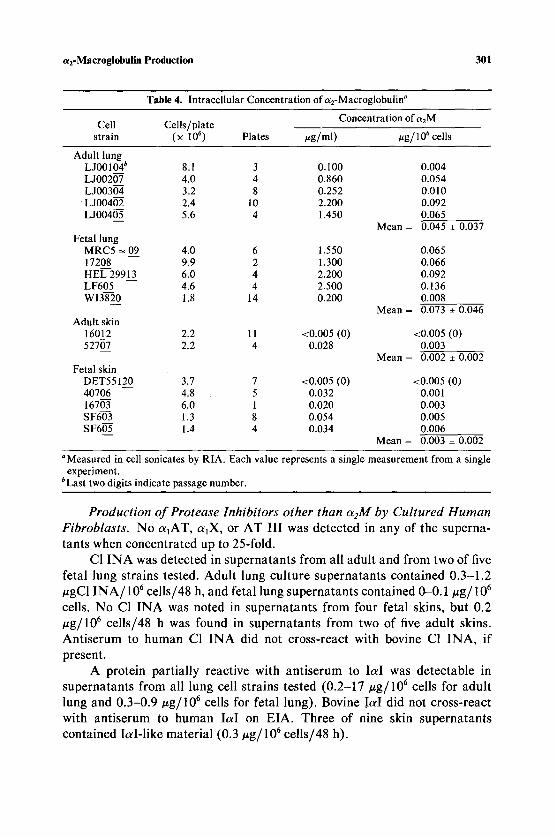
aRM was also quantitated by RIA in cell sonicates (Table 4). a2M appears to be present intracellularly as well as extracellularly in these fibroblast cultures, in contradiction to previously reported results (14). Again lung fibroblasts appear to have higher intracellular aEM concentrations, 0.06
3 0 0 Brissenden and Cox
10.000-
0J u
z 0
uJ
Z 0 u
z D ..a
o 0 0 or U <
r
6.000-
3 .000-
1 . 0 0 0 -
0 . 6 0 0 -
0 . 3 0 0 -
0 . 1 0 0 -
0 . 0 6 0 -
0 . 0 3 0 -
0.010
O O
@ @ O O
@o @
@ �9 O
�9 �9 O
�9 | e e O
�9 �9 O O
1 2 3 4 1 2 3 4 5 1 2 3 4 5 6 7 1 2 3 4
ADULT FETAL ADULT FETAL L UNG LUNG SKiN SKiN
Fig. 3. Human ~2-macroglobulin concentrations in supernatants of cultured human fibroblasts. e , adult lung and skin; O, fetal lung and skin;| @~ results obtained by EIA; �9 O, results obtained by RIA. The numbers identify the strains as explained in Table 1.
#g ct2M / 10 6 cells, than skin fibroblasts, 0.002/~g/10 6 cells (t = 3.48; df = 15; P < 0.01).
We measured a2M quantities functionally as well as immunologically in some culture supernatants to see if the immunologically detectable a2M was also functional, o~2-Macroglobulin was detectable functionally in supernatants from all four adult lung cultures and in three of four fetal lung fibroblast supernatants analyzed but in none of the seven skin fibroblast supernatants tested (Table 3). Considerable bovine o~2M was detectable by EIA in all of these concentrated supernatants, but it was not functional after one week in contact with the cells.
a2-Macroglobulin Production 301
Table 4. Intracellular Concentration of a2-Macroglobulin a
Concentration of a2M Cell Cells/plate
strain (x 10 6) Plates #g/ml) vg/lO 6 cells
Adult lung L J00104 b 8.1 3 0.100 0.004 L J00207 4.0 4 0.860 0.054 L J00304 3.2 8 0.252 0.010 L J00402 2.4 10 2.200 0.092 L J00405 5.6 4 1.450 0.065
Mean = 0.045 _+ 0.037 Fetal lung
MRC5 -~ 09 4.0 6 1.550 0.065 17208 9.9 2 1.300 0.066 HEL 29913 6.0 4 2.200 0.092 LF605 4.6 4 2.500 0.136 WI3820 1.8 14 0.200 0.008
Mean = 0.073 _+ 0.046 Adult skin
16012 2.2 11 <0.005 (0) <0.005 (0) 52707 2.2 4 0.028 0.003
Mean = 0.002 _+ 0.002 Fetal skin
DET55120 3.7 7 <0.005 (0) <0.005 (0) 40706 4.8 5 0.032 0.001 16703 6.0 1 0.020 0.003 SF603 1.3 8 0.054 0.005 SF605 1.4 4 0.034 0.006
Mean = 0.003 _+ 0.002
~ in cell sonicates by RIA. Each value represents a single measurement from a single experiment.
bLast two digits indicate passage number.
Production of Protease Inhibitors other than a:M by Cultured Human Fibroblasts. No a lAT, a~X, or AT II I was detected in any of the superna- tants when concentrated up to 25-fold.
C1 I N A was detected in supernatants from all adult and from two of five fetal lung strains tested. Adul t lung cul ture supernatants contained 0.3-1.2 #gC1 INA/106 cells/48 h, and fetal lung supernatants contained 0-0.1 #g/106 cells. No C1 I N A was noted in supernatants from four fetal skins, but 0.2 #g/106 cells/48 h was found in supernatants from two of five adult skins. Ant i se rum to human C1 I N A did not cross-react with bovine C1 INA, if present.
A protein partially reactive with an t i serum to I a I was detectable in supernatants from all lung cell strains tested (0.2-17 #g/106 cells for adult lung and 0.3-0.9 #g/106 cells for fetal lung). Bovine ! a I did not cross-react with ant iserum to human I a I on EIA. Three of nine skin supernatants contained led-like material (0.3 #g/106 cells/48 h).

302 Brissenden and Cox
These results indicate that C1 INA and an IaI-like material are synthesized by cultured lung and skin fibroblasts. There appear to be differences in the amount of production of these protease inhibitors by fibroblasts of different ontological and/or tissue origin: adult lung fibroblasts appear to produce the highest amounts.
DISCUSSION
In this study we have unequivocally demonstrated that human lung fibroblasts of adult origin produce human azM in culture, a phenomenon not previously reported. We showed this production both immunologically and functionally. We have demonstrated that cultured fetal lung fibroblasts made identical quantities of a2M to cultured adult lung fibroblasts and that skin fibroblasts of both adult and fetal origin made considerably (10 times) less a2M than did lung fibroblasts. These differences were apparent for genetically identical lung and skin fibroblast strains, both adult and fetal.
One earlier report has also convincingly demonstrated a2M production, and some relative a2M quantitative differences in production, by cultured embryonic lung and some other embryonic and non-lung adult fibroblasts (14).
azM was functional in most of the lung fibroblast culture supernatants we assessed. As expected, a2M function was never detectable in supernatants from skin cultures, even when concentrated 100-fold, since immunologically detectable human a2M was below the sensitivity limit of the functional assay in these cases. Usually less functional o~2M than immunologically detectable a2M was present in lung supernatants, suggesting that some a2M in culture supernatants may be nonfunctional, perhaps as a result of binding to a protease, such as collagenase which is known to be produced by cultured fibroblasts (20). A previous assessment of supernatants of human adult and fetal lung cultures for protease inhibitor activity failed to detect functional azM in culture (16).
We have also demonstrated intracellular aRM in both lung and skin fibroblasts by RIA in contrast to previously reported data (14). The intracel- lular a2M concentrations showed the same relative azM levels as the cell supernatants, suggesting that lowered azM levels in skin fibroblast superna- tants do not reflect intracellular trapping of a2M. These findings could be due to differences in either the rate of azM production or degradation between skin and lung fibroblasts.
We have demonstrated the presence of azM in supernatants from explants of human lung and skin after cellular outgrowth but not before. Lack of demonstrable a2M in early lung explants was important since blood cells, lymphocytes and those of the macrophage/monocyte lineage, may produce

~z-Macrogiobulin Production 303
otzM (8-10). Such cells could have been present in the early blood-stained explants and could have contributed to a2M detected early in the life of the culture.
o~2M production by embryonic fibroblasts other than lung and by adult skin and synovium was reported to drop markedly after the first subculture and to remain at the lower level (14). We have observed no similar pronounced drop in o~2M synthesis by any of our 1 ~ cultures after passage. The difference between our results and those reported previously may arise from different techniques used to establish the cultures. Our cultures were established by mincing the tissue and planting small explants under coverslips. Mosher et al. trypsinized their explants and plated them as cell suspensions (14). This latter technique may have allowed more nonfibroblast cells to become established in their cultures than in ours. Many of these nonfibroblast cells might not survive subculture but might make oL2M, which would account for higher c~2M levels being found in their supernatants from 1 ~ cultures than from those from later passages.
Our EIA results indicate that CI INA is produced by cultured lung and skin fibroblasts. This component of the complement pathway has previously been noted to be synthesized in culture by such embryonic tissue slices as lung, liver, skin, and subcutaneous tissue, but not by blood (32). The C1 INA may have been produced by fibroblasts present in the slices. C1 INA was reported previously to be functionally undetectable in supernatants from adult and fetal lung fibroblast cultures (16). This lack of function may occur because of complexing with C1, also noted to be produced by fibroblasts (34) and macrophages (35). The fact that fibroblasts make both these proteins and the complement component C3 (36) may be functionally significant.
A protease inhibitor immunologically related to I~xI, a plasma trypsin inhibitor of molecular weight 160,000 (2), was also found in our culture supernatants. Protease inhibitors said to be immunologically related to Iod have been noted in bronchial mucus and in urine (33). IaI was not detectable by function in a previous study of cultured lung fibroblasts (16). The inhibitor which we have detected may represent partially degraded Iod, which is therefore not fully reactive with Iod antiserum, or some structurally similar protease inhibitor.
If the lung fibroblasts produce o~2M, C1 INA, and IaI in vivo as well as in culture, such local production may be physiologically important in the control of tissue proteolysis, particularly for the large otzM molecule. Such locally produced protease inhibitors could be even more important than those transported by serum in the protection of specialized tissues.
a2M production appears to provide a marker for the identification of lung fibroblasts from fibroblasts of other origins. The difference in quantities of o~zM and possibly other protease inhibitors produced by fibroblasts of different

304 Brissenden and Cox
origin emphasizes the fact that all fibroblasts are not alike and underlines the necessity for care in the choice of the origin of fibroblasts for experimental controls and for cell hybridization experiments where the original biochemical background of the two types of cells is intended to be as similar as possible.
ACKNOWLEDGMENTS
This work was supported by the Canadian Lung Association and by the Medical Research Council of Canada (No. MA-5426). We would like to thank Drs. L. Siminovitch and M. Buchwald for helpful discussion of the manuscript, Drs. D. Hoar and S. Goldstein for their gifts of tissue explants and cultured cells, and Tammy Mansfield for quantitating protease inhibitors, other than a2M, in the supernatants.
LITERATURE CITED
1. Wolf, D.H. (1980). In Advances in Microbiol Physiology, Vol 21, (eds.) Rose, A.H., and Morris, J.G. (Academic Press, London), pp. 267-338.
2. Proteases and Biological Control, Reich, E., Rifkin, D.B., and Shaw, E. (eds.) (1975). Cold Spring Harbor Conferences on Cell Proliferation, Vol. 2. Cold Spring Harbor Laboratory.
3. Werle, E., and Zickgraf-Riidel, G. (1972). Z. Klin. Chem. Klin. Biochem. 10:139-150. 4. Barnett, T.B., Gottovi, D., and Johnson, A.M. (1975). Am. Rev. Resp. Dis. 111:587-593. 5. Laurell, C.B., and Jeppsson, J.O. (1975). In The Plasma Proteins, Vol. 1, 2nd ed., (ed.)
Putnam, F.W. (Academic Press, New York), pp. 229-264. 6. Barrett, A.J., Starkey, P.M., and Munn, E.A. (1974). In Proteinase Inhibitors, (eds.) Fritz,
H., Tschesche, H., Greene, L.J., and Truscheit, E. Bayer Symposium V. (Springer-Verlag, Berlin), pp. 72-77.
7. Wooley, D.E., Roberts, D.R., and Evanson, J.M. (1976). Nature 261:325-327. 8. Hovi, T., Mosher, D., and Vaheri, A. (1977). J. Exp. Med. 145:1580-1589. 9. Tunstall, A.M., and James, K. (1974). Clin. Exp. Immunol. 17:697-701.
10. James, K., Tunstall, A.M., Parker, A.C., and McCormick, J.N. (1975). Clin. Exp. lmmunol. 19:237-249.
11. White, R., Janoff, A., and Godfrey, H.P. (1980). Lung 158:9-14. 12. Harris, J.R., Brown, K.D., and Aiton, J.F. (1976). Biochim. Biophys. Acta 427:727-737. 13. Mosher, D.F., and Wing, D.A. (1976). J. Exp. Med. 143:462-467. 14. Mosher, D.F., Saksela, O., and Vaheri, A. (1977). J. Clin. Invest. 60:1036-1045. 15. Pastan, I., Willingham, M., Anderson, W., and Gallo, M. (1977). Cell 12:609-617. 16. Bernik, M.B. (1977). Thromb. Res. 11:739-746. 17. Shortridge, K.F., Balyavin, G., and Bidwell, D.E. (1976). Comp. Biochem. Physiol.
54A:319-321. 18. Van Leuven, F., Verbruggen, R., Cassiman, J.J., and Van den Berghe, H. (1977). Exp. Cell
Res. 109:468-471. 19. Vlodavsky, I., Johnson, L.K., and Gospodarowicz, D. (1979). Proc. Natl. Acad. Sci. U.S.A.
76:2306-2310. 20. Bauer, E.A. (1977). Exp. Cell Res. 107:269-276. 21. McGarrity, G.J. (1975). Methods Enzymol. 58:21-29. 22. Hayflick, L., and Moorhead, P.S. (1961). Exp. Cell Res. 25:585-621. 23. Ouchterlony, 6 , and Nilsson, L. A. (1978). In Handbook o f Experimental Immunology,
Vol. 1, (ed.) Weir, D.M. (Blackwell Scientific Publications, Oxford), Chapter 19.

~z-Macroglobnlin Production 305
24. Laurell, C.-B. (1972). Scand. J. Clin. Lab. Invest. 29(Suppl. 124):21-37. 25. Hunter, W.M. (1978). In Handbook o f Experimental Immunology, Vol. 1, (ed.) Weir,
D.M. (Blackwetl Scientific Publications, Oxford), Chapter 14. 26. Virca, G.D., Travis, J., Hall, P.K., and Roberts, R.C. (1979). Anal. Biochem. 89:274-278. 27. Smithies, O: (1955). Biochemistry 61:629-641. 28. Johansson, B.G. (1972). Scand. J. Clin. Lab. Invest. 29(Suppl. 124):7-19. 29. Haverbaek, B.J., Dyce, B., Bundy, H.F., Wirtschafter, S.K., and Edmondson, H.A. (1962).
J. Clin. Invest. 41:972-980. 30. Adams, D.O. (1975). Methods Enzymol. 58:494-506. 31. Hayflick, L. (1965). Exp. CellRes. 37:614-636. 32. Gitlin, D., and Biasucci, A. (1969). J. Clin. Invest. 48:1433-1446. 33. Hochstrasser, K., Feuth, H., and Hochgesand, K. (1974). In Proteinase Inhibitors, (eds.)
Fritz, H., Tschesche, H., Greene, L.J., and Truscheit, E. Bayer Symposium V. (Springer- Verlag, Berlin), pp. 111-116.
34. Reid, K.B.M., and Solomon, E. (1977). Biochem. J. 167:647-660. 35. Colter, H.R., Alper, C.A., and Rosen, F.S. (1981). N. Engl. J. Med. 304:653-656. 36. Whitehead, A.S., Sim, R.B., and Bodmer, W.F. (1981). Eur. J. lmmunol. 11:140-146.





























