Embed Size (px)
Citation preview

1
A capsular polysaccharide impairs microbe-guided chemotactic chasing by
neutrophils
Tamding Wangdi, Cheng-Yuk Lee, Alanna M. Spees, Chenzhou Yu, Dawn D.
Kingsbury, Sebastian E. Winter, Volkmar Heinrich and Andreas J. Bäumler

2
Text
The contribution of capsular polysaccharide to the virulence of
invasive human pathogens is commonly attributed to their anti-phagocytic
properties. Here we show that the virulence-associated (Vi) capsular
polysaccharide of Salmonella enterica serovar Typhi (S. Typhi) also
obstructs neutrophil chemotaxis, a previously overlooked property with
important consequences for host pathogen interaction. In single-cell
experiments, neutrophils extended chemotactic pseudopods toward a non-
capsulated S. Typhi mutant but not toward the capsulated S. Typhi wild
type. Formation of chemotactic pseudopods toward non-capsulated S.
Typhi required complement component 5a receptor (C5aR), suggesting that
neutrophils perceive bacteria by detecting C5a radiating from the microbial
surface. Deletion of capsule biosynthesis genes markedly enhanced uptake
of S. Typhi by neutrophils in vivo through a C5aR-dependent mechanism.
Collectively, these data suggest that the Vi capsular polysaccharide
contributes to virulence by obstructing C5a-guided chemotactic chasing of
bacteria by neutrophils.
Neutrophil chemotaxis toward intruding microbes is an essential host
defense mechanism against bacterial infection. Neutrophil migration into an
infected tissue is initially guided by chemoattractants, such as interleukin (IL)-8, a
chemokine produced by host cells after they detect the presence of bacteria
using pathogen recognition receptors (PRRs). Neutrophils that follow a gradient

3
of IL-8 to enter a site of infection will ultimately reach host cells producing this
chemokine, but this host-guided chemotaxis does not explain how neutrophils
subsequently hunt down bacteria in tissue. Early microscopic observations
suggest that chemotactic chasing of Staphylococcus aureus by neutrophils is
microbe-guided 1. A classic video produced during those studies
(http://www.biochemweb.org/neutrophil.shtml) shows that once neutrophils come
within a certain distance of S. aureus, they move directly toward bacteria by
extending chemotactic pseudopods, pushing aside red blood cells that block a
direct path. Since the identity of the chemoattractant emanating from the microbe
that guides chemotactic chasing by neutrophils remains unknown, no studies
have addressed the question whether bacteria can produce virulence factors that
can overcome this host defense mechanism.
N-formyl peptides represent one chemoattractant derived from bacteria
that might direct microbe-guided neutrophil chemotaxis 2. Alternatively, bacterial
surface carbohydrates might activate complement by the alternative pathway,
thereby giving rise to complement fragment C5a, a potent neutrophil
chemoattractant 3. The latter possibility attracted our attention, since it suggested
that inhibition of complement activation by expression of a capsular
polysaccharide could be a mechanism to evade chemotactic chasing by
neutrophils. Here we investigated this hypothesis and its consequences for host
microbe interaction in vitro and in vivo.
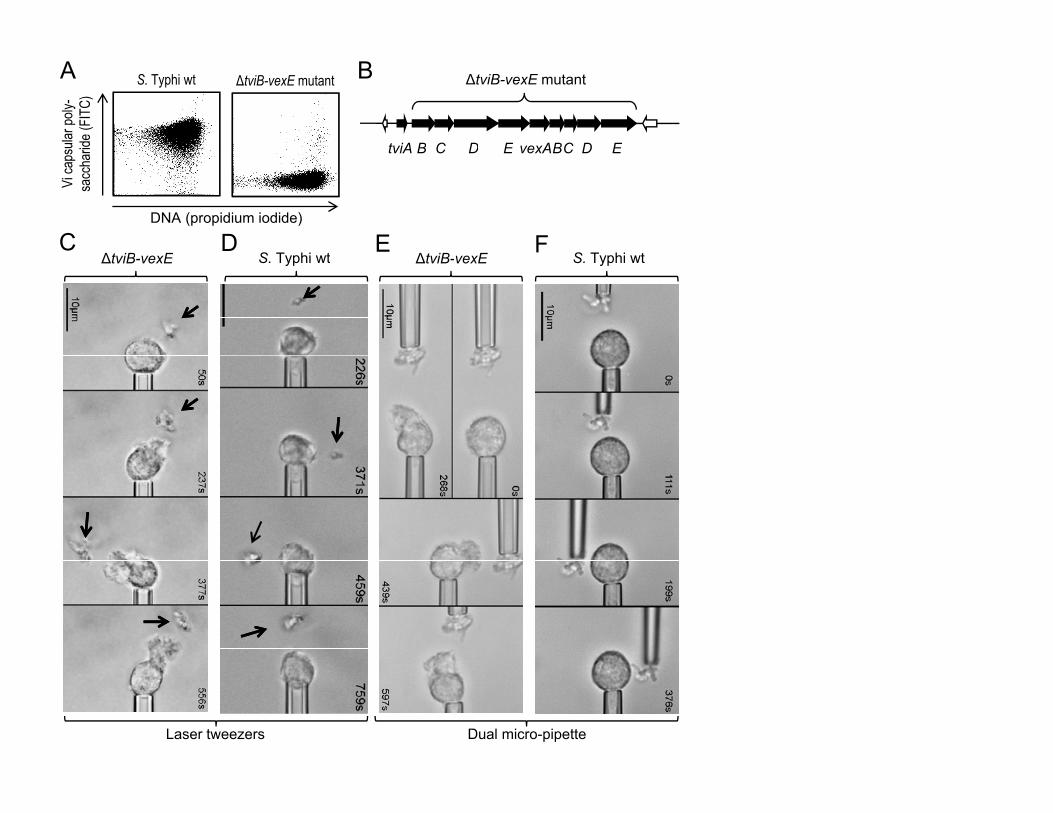
Salmonella enterica serotype Typhi, the causative agent of typhoid fever,
expresses a virulence-associated (Vi) capsular polysaccharide (Fig. 1A) 4. The

4
viaB locus on the S. Typhi chromosome contains genes for the regulation (tviA)
the biosynthesis (tviBCDE) and the export (vexABCDE) of the Vi capsular
polysaccharide (Fig. 1B) 5. To investigate a possible role of the Vi capsular
polysaccharide in evading neutrophil chemotaxis we compared a capsulated S.
Typhi wild-type strain (Ty2) with an isogenic non-capsulated strain lacking the
capsule biosynthesis and export genes (ΔtviB-vexE mutant) (Fig. 1A) 6. The
chemotactic response of neutrophils was first investigated using single-cell
experiments 7, in which an initially quiescent human neutrophil was picked up at
the tip of a micropipette (Fig. S1). Bacteria were immobilized by a laser optical-
trap (laser tweezers) and brought stepwise into close proximity of the neutrophil
in the presence of serum. Bringing the non-capsulated S. Typhi strain (ΔtviB-
vexE mutant) into a certain distance of a neutrophil induced a vigorous
chemotactic response, characterized by formation of a cellular pseudopod, which
protruded toward the bacteria and responded quickly to their relocation (Fig. 1C,
Supplementary video 1). In striking contrast, the capsulated S. Typhi wild-type
strain did not elicit any chemotactic response by neutrophils (Fig. 1D,
Supplementary video 2). Identical results were obtained when agglutinated
bacteria were picked up by a micropipette and brought stepwise into close
proximity of a neutrophil (Fig. 1E and F, Supplementary videos 3 and 4). These
data suggested that expression of the Vi capsular polysaccharide prevents
neutrophil chemotaxis.
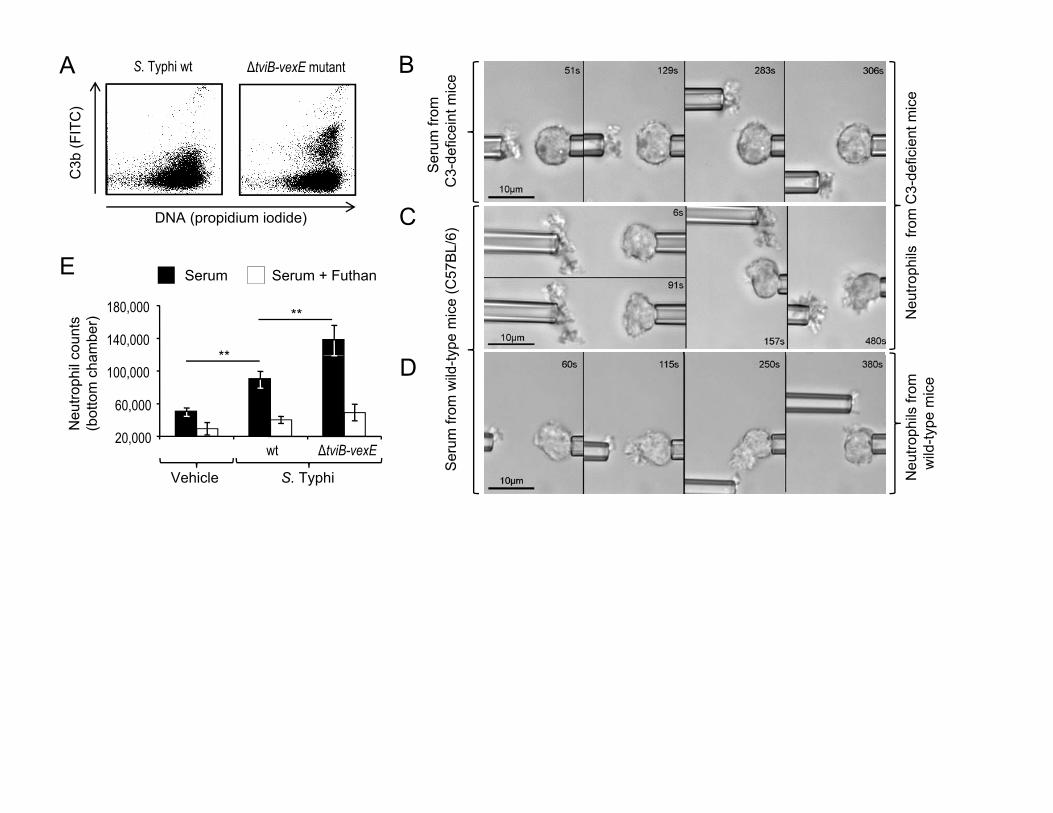
Expression of the Vi capsular polysaccharide reduces complement
activation by the alternative pathway 8,9, as indicated by diminished fixation of

5
complement fragment C3b on the surface of the capsulated S. Typhi wild-type
strain compared to a non-capsulated S. Typhi strain (ΔtviB-vexE mutant) (Fig.
2A). To study the role of complement in microbe-guided chemotaxis, dual-
micropipette manipulation experiments with a non-capsulated S. Typhi strain and
neutrophils from C3-deficient mice were performed in the presence of
homologous or heterologous serum. When the assay was performed in serum
from C3-deficient mice, neutrophils did not exhibit a chemotaxic response toward
the non-capsulated S. Typhi strain (Fig. 2B, suppl. video 5). In contrast, in the
presence of serum from wild type mice (C57BL/6) neutrophils from wild type mice
(C57BL/6) and neutrophils from congenic C3-deficient mice extended cellular
pseudopods toward the bacteria (Fig. 2C and 2D; suppl. video 6 and 7). To
further investigate whether neutrophil chemotaxis elicited by a non-capsulated S.
Typhi strain was complement dependent, we monitored neutrophil migration from
the upper compartment of a Boyden chamber 10 into a bottom reservoir that
contained sterile medium or different bacterial strains. The presence of a non-
capsulated S. Typhi strain in the bottom chamber elicited migration of
significantly (P < 0.05) larger numbers of neutrophils than the presence of the
capsulated S. Typhi wild-type strain. In contrast, no neutrophil migration was
observed when the complement inhibitor Futhan 11 was added to the medium
(Fig. 2E). Collectively, these data suggested that microbe-guided neutrophil
chemotaxis is complement-dependent and can be reduced by the Vi capsular
polysaccharide.

6
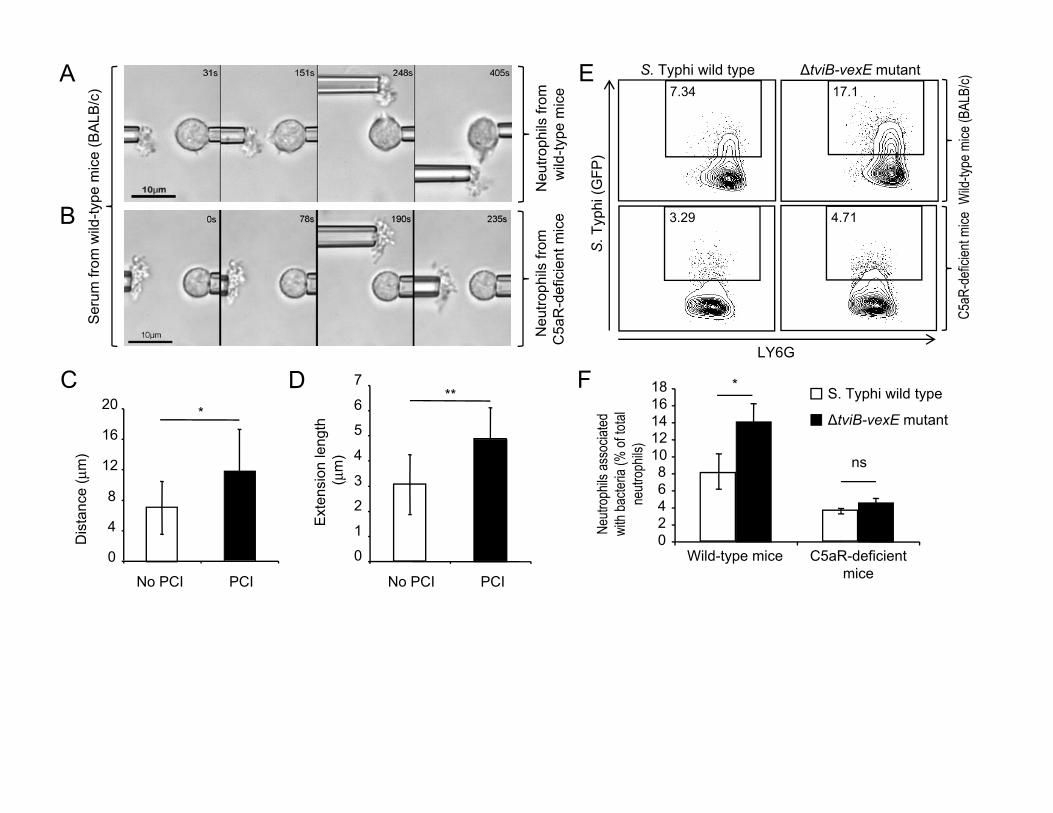
Next, we investigated the mechanism by which complement directs
neutrophil chemotaxis toward bacteria. The alternative pathway of complement
activation is initiated by formation of C3 convertase, C3b fixation on the bacterial
surface and subsequent recruitment of complement components involved in
formation of the membrane attack complex, including C5. Cleavage of C5
generates the neutrophil chemoattractant C5a, which is released from the
bacterial surface 3. To investigate whether C5a emanating from the bacterial
surface was responsible for microbe-guided chemotaxis, we performed dual-
micropipette manipulation experiments with a non-capsulated S. Typhi strain and
neutrophils from C5a receptor (C5aR, also known as CD88)-deficient mice 12.
Remarkably, unlike neutrophils from wild type mice (BALB/c) (Fig. 3A, suppl.
video 8), neutrophils from C5aR-deficient mice did not exhibit any chemotactic
response toward non-capsulated bacteria (Fig. 3B, suppl. video 9). These data
suggested that neutrophils perceive bacteria by detecting a C5a gradient that
radiates from the microbial surface.
Upon cleavage from C5, C5a is quickly metabolized by carboxy-
peptidases, which remove the C-terminal arginine residue to form C5a des Arg, a
molecule with markedly reduced chemotactic activity 13. We reasoned that this
mechanism would result in the presence around a microbe of a chemotactic C5a
corona with a small diameter, which would explain why chemotactic responses
are only elicited when microbes are brought into close proximity to a neutrophil.
This hypothesis would suggest that inhibition of carboxy-peptidase activity would
increase the diameter of the C5a corona, thus increasing the maximum distance

7
between microbe and neutrophil at which a response is still induced. To test this
prediction we used zymosan-particles to model a microbial surface that activates
complement 7. By reducing the distance of a zymosan-particle to a human
neutrophil stepwise using dual-micropipette manipulation experiments, we
observed that supplementation of serum with potato carboxypeptidase inhibitor
(PCI) 14 significantly increased the maximum target perception distance (Fig. 3C
and S2) and the maximum extension of pseudopods (Fig. 3D).
Finally, we wanted to determine the biological significance of our
observations using an animal model. S. Typhi is a human restricted pathogen
and is rapidly cleared from organs of mice 8. However, since chemotactic chasing
and phagocytosis by neutrophils occurs very early during host microbe
interaction and precedes clearance of bacteria, we reasoned that these
processes could be studied at an early time point (1 hour) after injecting S. Typhi
into mice. The capsulated S. Typhi wild-type strain and a the non-capsulated S.
Typhi mutant were transformed with a plasmid encoding green fluorescence
protein (GFP) and injected intraperitoneally into mice (BALB/c). Both strains were
still present in similar numbers one hour after infection and recruited similar
numbers of neutrophils into the peritoneal cavity (Fig. S3 and S4). However, a
significantly larger fraction of neutrophils from mice infected with the non-
capsulated S. Typhi mutant were associated with bacteria compared to mice
infected with the capsulated S. Typhi wild-type strain (Fig. 3E, 3F and S3). To
determine whether these differences were due to a capsule-mediated inhibition
of C5a-guided chasing by neutrophils, congenic C5aR-deficient mice were

8
infected with the capsulated S. Typhi wild-type strain, a non-capsulated S. Typhi
mutant or vehicle control (sterile PBS). Remarkably, the fraction of neutrophils
associated with the non-capsulated S. Typhi mutant was significantly reduced in
C5aR-deficient mice and no differences were observed between capsulated and
non-capsulated bacteria. Collectively, these data suggested that C5a-guided
chemotactic chasing by neutrophils is a host defense mechanism that can be
overcome by expression of the Vi capsular polysaccharide.
Chemotactic chasing by neutrophils was first observed over 50 years ago
1, but the phenomenon remains understudied, in part because most assays
investigating the interaction of microbes with phagocytes do not disentangle this
property from subsequent phagocytic events 15. Furthermore, conventional cell
migration assays, such as the Boyden chamber assay, lack the resolution to
distinguish between host-guided chemotaxis and microbe-guided chemotaxis.
The recent development of single-cell experiments provides now a convenient
assay for studying microbe-guided chemotaxis 15. Using this approach we show
that expression of the Vi capsular polysaccharide inhibits C5a-guided
chemotaxis, suggesting that this virulence factor restrains chemotactic chasing
by neutrophils. Our supplementary videos illustrate that through this mechanism
the Vi capsular polysaccharide can act as a “cloaking device” that makes S.
Typhi practically “invisible” to neutrophils, thereby enabling the pathogen to
evade this arm of the host defense in vivo (Fig. 3E and 3F). This previously
unrecognized property of the Vi capsular polysaccharide is likely relevant to the

9
interaction of our immune system with many other pathogenic microbes that
evade complement activation.
Literature
1. Melly, M.A., Thomison, J.B. & Rogers, D.E. Fate of staphylococci within
human leukocytes. J Exp Med 112, 1121-1130 (1960).
2. Schiffmann, E., Corcoran, B.A. & Wahl, S.M. N-formylmethionyl peptides
as chemoattractants for leucocytes. Proc Natl Acad Sci U S A 72, 1059-
1062 (1975).
3. Manthey, H.D., Woodruff, T.M., Taylor, S.M. & Monk, P.N. Complement
component 5a (C5a). Int J Biochem Cell Biol 41, 2114-2117 (2009).
4. Felix, A. & Pitt, R.M. A new antigen of B. typhosus. Lancet 227, 186-191
(1934).
5. Virlogeux, I., Waxin, H., Ecobichon, C. & Popoff, M.Y. Role of the viaB
locus in synthesis, transport and expression of Salmonella typhi Vi
antigen. Microbiology 141 ( Pt 12), 3039-3047 (1995).
6. Winter, S.E., Raffatellu, M., Wilson, R.P., Russmann, H. & Baumler, A.J.
The Salmonella enterica serotype Typhi regulator TviA reduces
interleukin-8 production in intestinal epithelial cells by repressing flagellin
secretion. Cell Microbiol 10, 247-261 (2008).
7. Mankovich, A.R., Lee, C.Y. & Heinrich, V. Differential effects of serum
heat treatment on chemotaxis and phagocytosis by human neutrophils.
PLoS One 8, e54735 (2013).

10
8. Wilson, R.P., et al. The Vi capsular polysaccharide prevents complement
receptor 3-mediated clearance of Salmonella enterica serotype Typhi.
Infection and immunity 79, 830-837 (2011).
9. Looney, R.J. & Steigbigel, R.T. Role of the Vi antigen of Salmonella typhi
in resistance to host defense in vitro. J Lab Clin Med 108, 506-516 (1986).
10. Boyden, S. The chemotactic effect of mixtures of antibody and antigen on
polymorphonuclear leucocytes. J Exp Med 115, 453-466 (1962).
11. Ikari, N., Sakai, Y., Hitomi, Y. & Fujii, S. New synthetic inhibitor to the
alternative complement pathway. Immunology 49, 685-691 (1983).
12. Hopken, U.E., Lu, B., Gerard, N.P. & Gerard, C. The C5a chemoattractant
receptor mediates mucosal defence to infection. Nature 383, 86-89
(1996).
13. Yancey, K.B., Lawley, T.J., Dersookian, M. & Harvath, L. Analysis of the
interaction of human C5a and C5a des Arg with human monocytes and
neutrophils: flow cytometric and chemotaxis studies. J Invest Dermatol 92,
184-189 (1989).
14. Ryan, C.A., Hass, G.M. & Kuhn, R.W. Purification and properties of a
carboxypeptidase inhibitor from potatoes. J Biol Chem 249, 5495-5499
(1974).
15. Heinrich, V. & Lee, C.Y. Blurred line between chemotactic chase and
phagocytic consumption: an immunophysical single-cell perspective. J
Cell Sci 124, 3041-3051 (2011).

11
16. Cummings, L.A., Wilkerson, W.D., Bergsbaken, T. & Cookson, B.T. In
vivo, fliC expression by Salmonella enterica serovar Typhimurium is
heterogeneous, regulated by ClpX, and anatomically restricted. Mol
Microbiol 61, 795-809 (2006).
17. Raffatellu, M., et al. The capsule encoding the viaB locus reduces
interleukin-17 expression and mucosal innate responses in the bovine
intestinal mucosa during infection with Salmonella enterica serotype
Typhi. Infect Immun 75, 4342-4350 (2007).

12
Supplementary Information accompanies the paper on www.nature.com/nature.
Acknowledgements
This project was supported by Public Health Service grant AI044170 to AJB. We
would also like to acknowledge the NIH Facilities Infrastructure Grant 1-C06-
RR12088-01. The authors do not declare conflicts of interest.
Author Information
Affiliations
Department of Medical Microbiology and Immunology, School of Medicine,
University of California at Davis, One Shields Ave; Davis CA 95616, USA
Tamding Wangdi, Alanna M. Spees, Dawn D. Kingsbury, Sebastian E. Winter,
and Andreas J. Bäumler
Department of Biomedical Engineering, University of California at Davis, One
Shields Ave; Davis CA 95616, USA
Cheng-Yuk Lee, Chenzhou Yu, Volkmar Heinrich
Contributions

13
TW contributed to the experimental design and contributed to Figures 1A, 1C,
1D, 1E, 1F, 2A, 2B, 2C, 2D, 2E, 3A, 3B, 3C, 3D, 3E, 3F, S3, S4A, S4B and S4C.
A.M.S. contributed to Figures 3E, 3F, S3, S4A, S4B and S4C. D.D.K. contributed
to Figures to Figures 3E, 3F, S3, S4A, S4B and S4C. S.E.W. constructed
bacterial plasmids and critically read the manuscript. C.-Y.L. contributed to
Figures 1E, 1F, 3B, 3C, 3D, 3A and 3B. C.Y. contributed to Figures 1C and 1D.
V.H. contributed to Figures S1 and S2. A.J.B. contributed to Figure 1B. A.J.B.
and V.H. provided financial support for the study and contributed to the
experimental design. A.J.B. and V.H. contributed equally to this work.
Competing financial Interests
The authors declare no competing financial interests
Correspondence
Correspondence to: Andreas J. Bäumler ([email protected]).

14
METHODS SUMMARY
The S. Typhi wild-type isolate Ty2 was obtained from the American Type
Culture Collection. A derivative of Ty2 carrying a ΔtviB-vexE deletion has been
described previously 6. To label bacteria with GFP, S. Typhi strains were
transformed with plasmid pDW5 16. All animal experiments were performed
according to Association for Assessment and Accreditation of Laboratory Animal
Care (AAALAC) guidelines and approved by the Institutional Animal Care and
Use Committee at the University of California at (UC) Davis. The UC Davis
Institutional Review Board approved the protocol for obtaining blood draws and
informed consent was obtained from all individuals. Six to eight week old female
mice obtained from The Jackson Laboratory were used for this study. Mice were
injected intraperitoneally with 1 x 107 colony forming units (CFU)/animal
suspended in 0.1 ml phosphate buffered saline (PBS). A neutrophil enrichment
kit (Stemcell Technologies, Vancouver, Canada) was used to collect murine
neutrophils. Human neutrophils were isolated using the Cytoselect Cell Migration
Assay kit (Cellbiolabs Inc., San Diego, CA). Neutrophils were suspended in
chemotaxis medium (1.5x106 cells/ml in Hank's Balanced Salt Solution with 10%
autologous serum). Rabbit anti-Vi serum (for Ty2) or rabbit anti-O9 serum (for
ΔtviB-vexE mutant) were used to generate bacterial micro-agglutinates. To
measure the maximum target perception distance, zymosan (St. Louis, MO)-
particles were prepared as described previously 7. To inhibit cleavage of C5a, 36
μg/ml of potato carboxypeptidase inhibitor (PCI) 14 (Sigma, St. Louis, MO) were
added to the chemotaxis medium. The micropipette setup has been described

15
previously 7. Vi capsule expression and C3b deposition were detected using flow
cytometry as described previously 8,17. Other flow cytometry experiments and the
Boyden chamber assay are described in the supplementary methods. To
determine statistical significance an unpaired Student t-test was used.

16
Figure Legends
Figure 1: The Vi capsular polysaccharide inhibits microbe-guided neutrophil
chemotaxis.
(A) Expression of the Vi capsular polysaccharide (Y-axis) was detected in
cultures of the S. Typhi wild type (wt, left panel) and a non-capsulated S. Typhi
strain (ΔtviB-vexE mutant, right panel) by flow cytometry using rabbit anti-Vi
serum. (B) Schematic drawing of Vi capsular biosynthesis genes (black arrows)
of S. Typhi. (C-F) The indicated bacterial strains were immobilized by laser
tweezers (arrows in C and D) or agglutinated with rabbit serum and picked up
with a micropipette (E and F). Bacteria were brought in close proximity to a
pipette-held human neutrophil and videomicrographs were taken at the indicated
time points. At least four neutrophils were analyzed per blood sample and each
experiment was repeated with neutrophils from different individuals (N = 4). (C
and E) Note chemotactic pseudopods that extend from neutrophils toward
bacteria.
Figure 2: The Vi capsular polysaccharide inhibits complement-dependent
neutrophil chemotaxis.
(A) Fixation of C3b (Y-axis) on the surface of the S. Typhi wild type (wt, left
panel) or a non-capsulated S. Typhi strain (ΔtviB-vexE mutant, right panel) that
had been incubated for 30 minutes in 1% human serum was detected by flow
cytometry using FITC-conjugated goat anti-human C3b antibody. (B-D)

17
Videomicrographs of dual-micropipette manipulation experiments with an
agglutinated non-capsulated S. Typhi strain and murine neutrophils from wild
type (C57BL/6) mice (D) or congenic C3-deficient mice (B and C) performed in
buffer containing serum from wild type mice (C and D) or in serum from C3-
deficient mice (B). For each strain, blood from 4 animals was pooled for isolation
of serum and neutrophils for an experiment. Each experiment was repeated at
least four times. (E) Neutrophil migration into the bottom compartment of a
Boyden chamber containing vehicle control (HBSS) or the indicated bacterial
strains. Bars represent averages ± standard error from 4 donors. **, P < 0.01.
Figure 3: The Vi capsular polysaccharide inhibits C5aR-dependent neutrophil
chemotaxis.
(A and B) Videomicrographs of dual micropipette manipulation experiments with
an agglutinated non-capsulated S. Typhi strain (ΔtviB-vexE mutant) and murine
neutrophils from wild type (BALB/c) mice (A) or congenic C5aR-deficient mice
(B). For each strain, blood from 4 animals was pooled for isolation of serum and
neutrophils for an experiment. Each experiment was repeated at least four times.
(C and D) Maximum target perception distance (C) and the maximum extension
of pseudopods (D) was determined using dual-micropipette manipulation of a
zymosan-particle and a human neutrophil in the presence or absence of potato
carboxypeptidase inhibitor (PCI). Figure represents experiments with blood from
two donors, with at least four neutrophils being tested each per experimental
condition. (E and F) Mice were infected intraperitoneally with the indicated GFP-

18
labeled bacterial strains and cells were collected one hour later by intraperitoneal
lavage. Representative images of bacterial association (Y-axis) with neutrophils
from wild type mice or congenic C5aR-deficient mice (E) and quantitative
analysis of the data (averages ± standard error) from groups of six animals (F)
are shown. (C, D and F) **, P < 0.01; *, P < 0.05; ns, not significantly different.
Figure S1: Single-cell approach to assess neutrophil chemotaxis toward S. Typhi
in buffer containing serum.
S. Typhi (image on the top left) and neutrophils from humans or mice (images on
the top right) were deposited into a microscopy chamber with two open sides
(schematic drawing in the center). Vertically adjustable water reservoirs allowed
accurate pressure application to facing micropipettes inserted into this chamber.
An initially quiescent neutrophil was aspirated at the tip of a micropipette. S.
Typhi was either immobilized with optical tweezers or micro-aggregates of
bacteria were produced and picked up using a second micropipette. Bacteria and
neutrophils were then brought stepwise into close proximity (image at the
bottom).
Figure S2: Measurement of the maximum target perception distance and the
maximum extension of pseudopods.
Using dual-micropipette manipulation, a zymosan-particle was stepwise brought
into close proximity of an initially passive human neutrophil. Relative times are
indicated in the videomicrographs. Images illustrate the determination of the

19
average target distance at which pseudopod formation started within 2 minutes
(first videomicrograph from the left) and the maximum length of chemotactic
pseudopods (fourth videomicrograph from the left).
Figure S3: Gating strategy for detecting an association of S. Typhi with
neutrophils from peritoneal lavage.
Gating strategy for analyzing peritoneal lavage cell suspensions. After doublet
elimination (top left panel) live cells were gated (top middle panel) and CD3+
B220+ NK1.1+ cells eliminated using a dump channel (top right panel). CD3-
B220- NK1.1- cells were then analyzed for expression of CD11B (right panel in
the middle row). Next, CD3- B220- NK1.1- CD11B+ cells were analyzed for
expression of LY6C and LY6G cells (left panel in the middle row). CD3- B220-
NK1.1- CD11B+ LY6C+ LY6G+ cells (neutrophils) were analyzed by microscopy to
confirm neutrophil morphology (bottom right panel). The gate for the detection of
GFP-labeled S. Typhi in neutrophil populations was set using cells isolated from
control mice injected with sterile PBS (bottom left and middle panels).
Figure S4: Quantification of neutrophil infiltrates elicited by infection with S.
Typhi.
Mice (BALB/c) were injected intraperitoneally with sterile PBS or the indicated
GFP-labeled bacterial strains and cells were collected one hour later by
intraperitoneal lavage. (A) Representative images of neutrophil infiltration (LY6C+
LY6G+ cells) in intraperitoneal lavage populations detected by flow cytometry. (B)

20
Quantitative analysis of neutrophil infiltration is shown as geometric means (bars)
± standard error from groups of six animals. (C) Bacterial numbers recovered
from peritoneal lavage. NS, not significantly different.
S. Typhi wt ΔtviB-vexE mutantsu
lar po
ly-rid
e (FI
TC)
A B
EDCBvexAEDCBtviA
ΔtviB-vexE mutantVi
caps
sacc
har
DNA (propidium iodide)
EDC BvexAEDC BtviA
C D E FΔ iB E Δ iB ES T hi t S T hi tΔtviB-vexE ΔtviB-vexES. Typhi wt S. Typhi wt
Laser tweezers Dual micro-pipette
S. Typhi wt ΔtviB-vexE mutant(F
ITC
)A B
rum
from
ef
icei
nt m
ice
cien
t mic
e
C3b
DNA (propidium iodide)
L/6)
Ser
C3-
de
s fr
om C
3-de
fic
C
Serum Serum + Futhan
140,000
180,000
**
**
E
ype
mic
e (C
57B
Neu
troph
il
coun
tsam
ber)
20,000
60,000
100,000
wt ΔtviB-vexE erum
from
wild
-ty
utro
phils
from
ild
-type
mic
eD
Neu
troph
il c
(bot
tom
cha
Vehicle S. Typhi
Se
Neu w
i
A
ophi
ls fr
om
type
mic
e
(BA
LB/c
)E
7.34 17.1
mice
(BAL
B/c)S. Typhi wild type ΔtviB-vexE mutant
B
Neu
trow
ild-
m w
ild-ty
pe m
ice
rom
t m
ice 3.29 4.71
Wild
-type
men
t mice
S. T
yphi
(GFP
)
Ser
um fr
om
Neu
troph
ils fr
C5a
R-d
efic
ien
C5aR
-defi
cie
LY6G
C DLY6G
S. Typhi wild type
ΔtviB-vexE mutant
1012141618 *F
socia
ted%
of to
talils
)16
20
)
*
4
5
6
7
leng
th)
**
02468
10
Wild-type mice C5aR-deficient
ns
Neutr
ophil
s as
with
bacte
ria (%
neutr
oph
0
4
8
12
Dis
tanc
e (μ
m
0
1
2
3
4
Ext
ensi
on
(μm
)
Wild type mice C5aR deficientmice
0 0
PCINo PCI PCINo PCI
















![Monocyte chemotactic protein-1 provokes mast cell aggregation and [3H]5HT release](https://img.pdfslide.net/doc/110x75/634811a4031992cdcf01d95c/monocyte-chemotactic-protein-1-provokes-mast-cell-aggregation-and-3h5ht-release.jpg)












