Embed Size (px)
Citation preview

1 APS406 Research Project Candidate No:100173864
A Comparative Study of the Arthropod
content of an Upper Carboniferous
(Westphalian B) Coal Seam: Clowne
Coal, N.E. Derbyshire
Jamie
Brown
University of
SheffieldDepartment of Animal and Plant
Sciences

2 APS406 Research Project Candidate No:100173864
Abstract
The technique of acid maceration is becoming more and more
frequently used to extract arthropod fragments from rocks of
geological significance that contain trapped organic material.
Thus the classification of arthropods, particularly scorpions,
based on ornamental microstructures has created a new
taxonomic approach to the phylum. The arthropod content of an
Upper Carboniferous coal seam in Clowne Common, N.E.
Derbyshire, has not been analysed before. Rock samples from an
outcrop of the seam located along a disused railway just
outside Clowne were taken for acid maceration analysis
involving the extraction of trapped cuticles within the coal.
The study successfully identified the species Compsoscorpius
buthiformis (Pocock, 1911), an Upper Carboniferous scorpion
previously found in the Westphalian B Kilburn Seam of the
English Midlands. It was identifiable thanks to the extensive
exocuticle analysis conducted by Jeram in his 1994 study of
Upper Carboniferous Orthosterni. The terrestrialisation of
this species reflects its success and abundance leading to the
conclusion that it could be part of the crown group for Recent
scorpions.
Keywords: Compsoscorpius buthiformis, Scorpion, acid maceration,
Light Microscope, SEM, terrestrialisation, British Coal
Measures.

3 APS406 Research Project Candidate No:100173864
1.Introduction
Over the last few decades the described taxa of Carboniferous
arthropods has taken a leap forward due to the extraction of
cuticle fragments from rocks and coals by acid treatment. As
the awareness for the importance of such rock digestion has
grown, lithologies that were thought to be devoid of
arthropods at a macroscopic level have yielded remarkably well
preserved fragments of arthropods at a microscopic level. This
has stimulated a more in-depth taxonomic description of
palaeo-arthropods; based on cuticle structure and
ornamentation.
Coal is an ideal facies to find arthropod remnants using
etching techniques as it is generally considered to have
captured not only aquatic but also terrestrial deposits. As
the records of Palaeozoic terrestrial arthropods are sparse,
finding new forms trapped in their natural habitat would
expand knowledge of palaeoecology and arthropod evolution.
Although plant-arthropod interactions of the Carboniferous are
widely studied (Todd, 1991; Shear & Kukalová-Peck, 1990; Scott
& Rex, 1985; Scott & Taylor, 1983), cuticle analysis has led
to the discovery of the boom in arthropod mouthpart
diversification that occurred around that time allowing
conclusions to be drawn about increased plant defences
(Labanderia, 2000).
In Britain, Yorkshire and Derbyshire have a few arthropod-
wealthy Westphalian B seams, including the Barnsley Seam and
the Lidgett Seam (Bartram et al., 1987), as well as the less

4 APS406 Research Project Candidate No:100173864
documented Clowne Seam (Taylor, 2000; Rippon, 1984). The
former two seams have been analysed for arthropod cuticle
content, but no record can be found for the latter seam. The
purpose of this project is to attempt to identify the
different types of arthropod cuticle present within the Clowne
Seam via analysis using micropalynological techniques, along
with a knowledge of British Carboniferous arthropods (Table
1.) and thus establish what arthropods they came from.
According to a wide variety of studies on cuticle
analysis, it is clear that Eurypterid and Scorpion (two
divergent orders of the true chelicerates; Figure 2)
exoskeletons are the hardiest and best preserved, with
evidence of other arthropods rarely distinguishable due to
decay (Legg et al., 2011; Stankiewicz et al., 1998; Mutvei, 1977).
The arthropod cuticle is one of the most complex natural
tissues which cannot be categorised by reference to only one
of its constituents, for example, the familiar polysaccharide
chitin. The structure of arthropod cuticles fluctuates
significantly depending on the type of arthropod, and on
particular sections of each individual (Richards & Korda,
1948). However, there are fundamental comparisons within the
cuticle composition of all arthropods (Barrington, 1979;
Neville, 1975). It consists of a basal layer of epidermal
cells (the endocuticle), followed by a thick procuticle with a
construction of chitin fibrils, and then a thin epicuticle,
without chitin, comprised chiefly of waterproofing lipids (the
exocuticle) (Bartram et al., 1987). The procuticle is the most
likely to be preserved as it consists of a thick inner zone,

5 APS406 Research Project Candidate No:100173864
whereas preservation of the endocuticle depends on how
calcified it is (as in most Trilobita and Crustacea), and the
thinner exocuticle depends on whether it is sclerotized
(darkened and hardened) and pigmented. The chitin structure of
the procuticle commonly occurs as sheets (lamellae) giving it
a very strong and resistant structure. The chemistry and
natural degradation of arthropod chitin in sediments over time
is exceedingly complex and poorly studied resulting in high
level of ambiguity in the identification and classification of
heavily decayed cuticles.
Nevertheless, due to the unusual hyaline exocuticle (hard
and glassy in appearance) of the scorpions, which is
characterised by the structure and packing of the microfibrils
(Filshie & Hadley, 1979), their unique arrangement has allowed
a vast recovery of cuticles from coals and related shales, but
not of other arthropods. This is the reason they are more
extensively studied than any other chelicerate group
(Dalingwater, 1986) and yield an almost continuous record
ranging from the fully aquatic forms in the Silurian, to both
terrestrial and aquatic faunas in the Carboniferous. It is
apparent from other parallel studies on animal cuticles,
(Stankiewicz et al., 1997; Baas et al., 1995; Briggs et al., 1995)
that the chitin element of arthropod cuticles is rapidly lost
during other forms of diagenesis.
There is a large number of arthropod taxa described in
Britain from the Carboniferous but for this study it is the
orders Eurypterida and Scorpiones that are of particular

6 APS406 Research Project Candidate No:100173864
importance (Table 1, see also Appendix 1 for all non-marine
Carboniferous chelicerates).
Sample location, background and collection.
An array of samples were collected from a single locality along the
Clowne Coal Seam first documented by Taylor in 2000. The outcrop of
the seam is in a disused railway cutting at NGR SK4782675441 ±8
meters identified using a GPS Sat Nav (Map 1). The site was
moderately easy to find but was overgrown; vegetation and debris had
to be cleared and the face cut back to reveal an adequate sampling
surface (Figure 1a). Extensive weathering of the exposure was
apparent from the iron staining and mineralisation in the cleat as
well as the friable nature of the coal.
Figure 1: A) View of sampling site at Clowne Common (SK4782675441 ±8 meters) Scale: Spade is107cm. B) Lithology and sampling intervals. Carbonaceous shale, Dull coalwith some bright bands, Bright coal, * Sampling level and collection range. Samplenames were comprised of the authors initials (JB) and the sample location Clowne (C).
(A (B
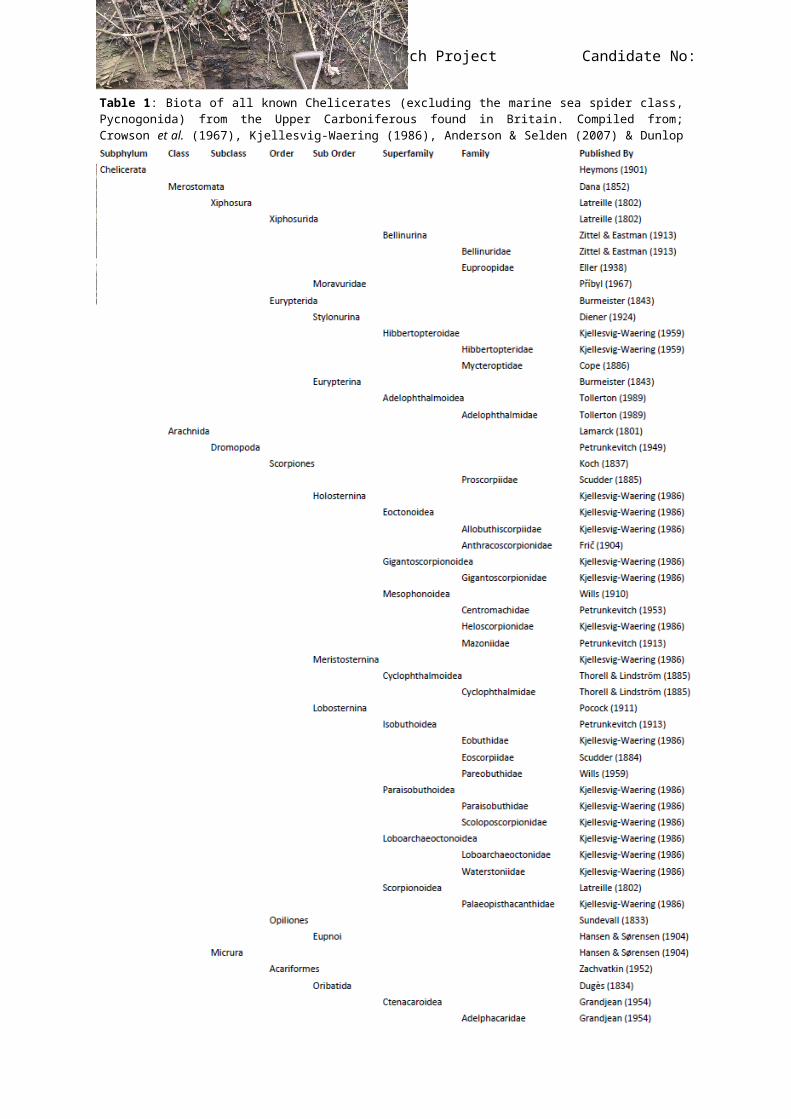
7 APS406 Research Project Candidate No:100173864Table 1: Biota of all known Chelicerates (excluding the marine sea spider class,Pycnogonida) from the Upper Carboniferous found in Britain. Compiled from;Crowson et al. (1967), Kjellesvig-Waering (1986), Anderson & Selden (2007) & Dunlopet al. (2013).

8 APS406 Research Project Candidate No:100173864
Table 1 cont.
Figure 2: Cladogram showing the current thoughts on the evolutionary pathway ofthe Chelicerates (Redrawn from Dunlop & Selden, 1998).

9 APS406 Research Project Candidate No:100173864
A series of ten coal samples (JBC0-JBC8, including JBCC) were
collected. Additionally there was an attempt to collect a sample
from the thin charcoal band described by Taylor (2000) but it proved
hard to find and the sample taken may have been mistaken for a
weathering line or dull coal band. Chunks of coal were clawed away
from the face with a geological hammer into plastic bags, labelled
respectively, and taken back to the palynology lab in Sheffield for
analysis. Not all samples collected were utilised in the
investigation as it became apparent that samples closer to the base
did not produce any identifiable cuticles.
Taylor (2000) included an interpretation of the past environment
in her report based on coal maceral analysis comparing the tissue
preservation index with the gelification index. She concluded that
the Dull Coal band (JBC1-JBC4) was formed by deposits in a clastic
marsh type environment, while the Bright Coal band (JBC5-JBC8) was
formed by deposits in a wet forest swamp environment. This could be
indicative of a move towards a more herbaceous flora. She has also
speculated that the Shale band above at the top of the column may be
a non-marine sediment equivalent to the Black Mudstone Bed, defined
by Rippon (1984), that directly overlies the Clowne Seam in some
parts of North Derbyshire and South Yorkshire. Conversely, the
charcoal band she describes that was hard to trace, indicates a dry
forest swamp/terrestrial environment due to perhaps a temporary drop
in the water table.
2.Methodology
The preservation of arthropod cuticle varies greatly depending on a
large number of factors including what order it’s from and what rock
facies it is trapped in. Often they have been found well preserved,
accidently, in Carboniferous studies on spore analysis and coal

10 APS406 Research Project Candidate No:100173864
balls (Scott, 2002; Scott & Rex, 1985), therefore similar techniques
to these studies are used to acquire arthropod cuticle.
Standard in-house methods were used for the chemical extraction
of the cuticles in the Dainton Palynology lab at Sheffield
University via acid maceration. The morphology of the cuticles and
their relative distribution among the bands were evaluated.
Extraction Procedure
Eight out of the ten sample taken were used in the investigation
(JBC6, JBC7 & JBC9 were not utilised due to a lack of recoverable
organic material after acid maceration). Representative portions of
each sample (~20g) were broken down into roughly 1cm3 chunks small
enough for ease of dissolving yet large enough to retain the
possibility of recovering large cuticle structures without them
being broken/damaged in preparation for acid maceration.
A reagent of saturated potassium chlorate (KClO3) in
concentrated nitric acid (HNO3) [70%] was synthesised in a fume
cupboard, known as Schulze’s reagent. To prevent the possibility of
any ‘energetic’ exothermic reactions generated by the pyrite or
carbonate minerals present in coal, a small amount of concentrated
HNO3 was first added to the coal and left to stand for a few minutes
until any reaction subsided.
Around 200ml of Schulze’s reagent was then added to each beaker
to create an oxidising reaction which also reacts with the humic
acid in the shales/coals, breaking it up. This reaction was left to
occur for 25 ± 10 minutes. When the humic acid was visible on the
surface of the beakers (a light brown froth) water was added to
dilute the acid reagent and slow down the reaction before the
mixture was decanted through an 180μm sieve (to collect matter of a

11 APS406 Research Project Candidate No:100173864
reasonable size) and the contents thoroughly rinsed. The substances
of the sieve were then washed back into the beaker using water.
Next, a 5% potassium hydroxide (KOH) solution was prepared by
dissolving KOH tablets in water, and ~250ml was added to each beaker
and left to stand for 5 minutes. The residue was then rinsed until
the water ran clear. This residue was then washed back into the
beaker again with clean water and swirled. The suspended matter was
transferred into 200ml (labelled screw cap) containers, left to
settle and then some of the excess water drained off.
Examination of Specimens
The sediments of each sample were emptied into glass petri dishes
and methodically examined under a dissecting microscope.
The samples mainly consisted of wood fragments, cutinised plant
material, megaspores and megaspore fragments, and miscellaneous
detritus. The amount of plant cuticle buried is most probably a
significant factor in the reduction of available oxygen in the
sediment which created the anoxic conditions required to preserve
arthropod cuticle (Allison & Briggs, 1993). After giving time to
train the eye it was possible to sift through the debris and pick
out arthropod cuticle with a fine brush and needle. The cuticle
could be either derived from dead animals of moulted exoskeletons.
Evidence for the latter is more frequent in all documented cases
(e.g. Jeram, 1994; Bartram et al., 1987). Aquamount (an aqueous
mounting medium) was used to mount the specimens on slides and hold
coverslips on permanently. Some specimens were able to be unfolded
through careful manipulation in the mounting medium before the cover
slip was positioned. These would then be ready for Light Microscope
(LM) analysis. As many well preserved specimens as possible were
picked from each sample but the amount of usable organic matter

12 APS406 Research Project Candidate No:100173864
decreased the closer the sample height was to the base of the
outcrop column.
The maceration residues contained a variety of material with
different amounts of fragmentation, which on the basis of
carboniferous sclerite sizes must have come from a variety of
different entities. Due to the difference in types of arthropod
sclerite between species, it is reasonable to suggest that some of
the fragmentation occurred before burial.
It became apparent further into the investigation that the three
dimensional fine structures on the cuticles could not be determined.
This meant that it is was necessary to mount some specimens onto
Scanning Electron Microscope (SEM) stubs for examination using
scanning electron microscopy (Plates 7 & 8). Only a few samples were
needed as the fine structures in scrutiny occurred on almost all
specimens.
Systematic Classification of Cuticles
All of the specimens collected and mounted (in total) were extremely
fragmentary which makes classification to the nearest species
impossible. However, the fine ‘hole’ like structures dotted around
the cuticle on the majority of specimens have been determined as
setae; tiny hair like sensory organs occurring on all arachnids.
With appreciation to Jeram’s work (1994) on the family
Palaeopisthacanthidea and other works (namely Legg et al., 2011 and
Wills, 1947), classification to the closest family or genera has
been attempted.
The problems involved in dispersed cuticle taxonomy have been
discussed by Wills (1947). A combination of Wills’ and Jeram’s
methods have been adopted here. It is assumed that only one species
is present at each locality/sample-band unless there are differences

13 APS406 Research Project Candidate No:100173864
between sclerites which can’t be attributed to any of the factors
responsible for variation. These may be any number of large
morphological differences due to ontogeny, intraspecific variation,
sexual dimorphism and taphonomic effects present within a single
species. Therefore, it has been deemed not possible to sub-divide
the sclerite groups reliably due to the limited amount of material
available. Furthermore, to date, from all English Carboniferous
assemblages, no respectable grounds have been found for the sub-
division of the sclerite groups, therefore different species within
this one assemblage can only be determined by comparison of cuticles
with previous work (Legg et al., 2007; Jeram, 1994; Bengtson, 1985;
Wills, 1960; Woodward, 1907).
The described Compsoscorpius elegans (Jeram, 1994) is of particular
interest as there is an extensive description of its external
anatomy, and it’s only documented locality is in the English
Midlands Westphalian B coal seam (Kilburn) (Legg et al., 2011; Fig.
2).

14 APS406 Research Project Candidate No:100173864
3.Results
Results are presented in the form of plates with a description that
attempts to classify the fragments.
Plate 1
A compilation of Light Microscope images of the best cuticlespecimens extracted from the Clowne Common outcrop, horizon JBC0(Slides 3, 4, 5, 6 & 7).
As there is not enough evidence to assign these fragments to oneparticular genus two possible families have been suggested:Palaeopisthacanthidea (Compsoscorpius) and Eoscorpiidae. Determining characteristic: Peg tooth organs.
Description of Figures:
Figure 1 - Hollowed out limb structure with no striking ornaments orsetal follicles apparent. The folding suggests a mild cone-likestructure and the arrow indicates is pointing to the distil endof the limb.
Figure 2 – A remarkably preserved comb-tooth which would have beenattached to a segmented rachi and in turn a pectin. The peg-organs cover the tooth thickly and appear as a minute clear lightcoloured circle (~30µm in diameter) against a background of thedarker skin of the comb-tooth. In the centre of each peg organ isa dark spot which, by analogy with Recent scorpions represents aminute peg (po). The light coloured circle has no rim butotherwise the whole organ has the same general appearance as adenticle. Organs like these have been found on the UpperCarboniferous scorpion Cyclophthalmus (Novojilov & Stormer, 1963),and also resemble the teeth like structure on the basitarus spursfound on Eobuthus by Wills, 1925. The corresponding structures inRecent scorpions are known particularly through the work of O.Schroder (1908), to consist of minute peg-shaped setae projectingfrom papillae and supplied by nerve fibres.
Figure 3 - What looks like a folded ventral plate with atrichobothria follicle clearly present on the proximal end (tf).Exceptional preservation of this cuticle shows tiny holesstructurally dotted about the plate. On close examination theseappear to be wax pores (wp ~ 500nm in diameter) similar to theones found in an SEM study on Calchas birulai (Benli et al., 2009).

15 APS406 Research Project Candidate No:100173864
Figure 4 - Very thin cuticle suggests this cuticle comes from adorsal sternite close to the joint of an appendage (most probablya walking femur). Heavy pigmentation and intricate foldingsuggests a change in cuticle structure around a joint of anappendage. One trichobothrium follicle and one setal follicle arevisible.
Figure 5 – Heavily degraded with only the procuticle layer partiallyleft, folded once in the middle. Its cellular structure can beseen as can traces of setal annulae. Cuticle is too far degradedto speculate on its anatomical position but the large number ofsetae suggest it comes from the lower part of a walkingappendage, such as the tibia or basitarsus.
Figure 6 – Heavily degraded, only procuticle left. A margin oflinear thickening is apparent with outlines of setal folliclesrunning alongside it. Characteristic of a carina on the pedipalpfemur. Thick arrow is pointing toward the distal end. Possiblewax pores penetrating the cuticle have been identified as well(wp).
Figure 7 – Heavily degraded epicuticle displaying the chitinousfibres overlying the procuticle. This means microstructures areimpossible to determine. However, the distinct double fold in thetop left of the figure could be ventral edges along a legappendage.

16 APS406 Research Project Candidate No:100173864
Plate 2
A compilation of Light Microscope images of some specimens of
cuticles extracted from the Clowne Common outcrop, horizon JBC1
(Slides 1, 2, 3, 5, 6, 8 & 9).
Genus: Compsoscorpius (Petrunkevitch, 1949).
Determining characteristic: Trichobothiral distribution on the
ventral side of a sternite.
Description of Figures:
Figure 1 - An almost linear scale like structure is visible here.The fold shows that the patterning is present on both sides andis therefore cellular. Two distinct shaded bands run parallelfollowing the cellular pattern along the cuticle. The width ofthese shaded bands, along with the elongate structure of thecells, suggest the cuticle is from the distal end of the telson.There is one clearly defined setal pit (follicle) (sp) whichjudging by its size most probably housed a trichobothria.
Figure 2 – Comparatively thick piece of cuticle. It is ornamented bya single scale tubercle and the lateral wall of a stigmata (sm).Consequently it is suggested that it comes from the dorsalsurface of the abdomen (due to the thickness of the cuticle andthe positioning of the tubercle and stigmata). It is not possibleto conclude which large sternite this is from.
Figure 3 – Cuticle is slightly degraded and microstructures are hardto place. Cuticle is folded but scale tubercles are apparentalong this fold. The colour of the cuticle, positioning the scaletubercles and distinct lack of setal pits in close proximity tothese mean it is a fragment of the dorsal surface of a metasomalsegment.
Figure 4 – The epicuticle has degraded in places but the smoothsurface of the cuticle can still be seen. Although it has adistinct tooth-like shape the edges are too rough and torn toconclude what is true structure may have look like.
Figure 5 – Exceptionally preserved cuticle showing the curvedstructure of the patella of a walking leg. A single scaletubercle is visible at the proximal end of the cuticle as arethree setal pits. The dorsal (right) and internal (left) sides of

17 APS406 Research Project Candidate No:100173864
the appendage have been preserved. Arrow points to the distal endof the patella.
Figure 6 – Dark brown cuticle implying a similar thickness to thecuticle presented in figure 2 suggesting that it is a fragmentfrom a sternite plate. One setal pit is clearly visible thoughslight degradation of the cuticle has obscured other structures.It is proposed that if the dark pigmented lines in the top rightof the figure is a part of the anterior transverse ridge thenthis specimen could be from the ventral surface of the abdomen.
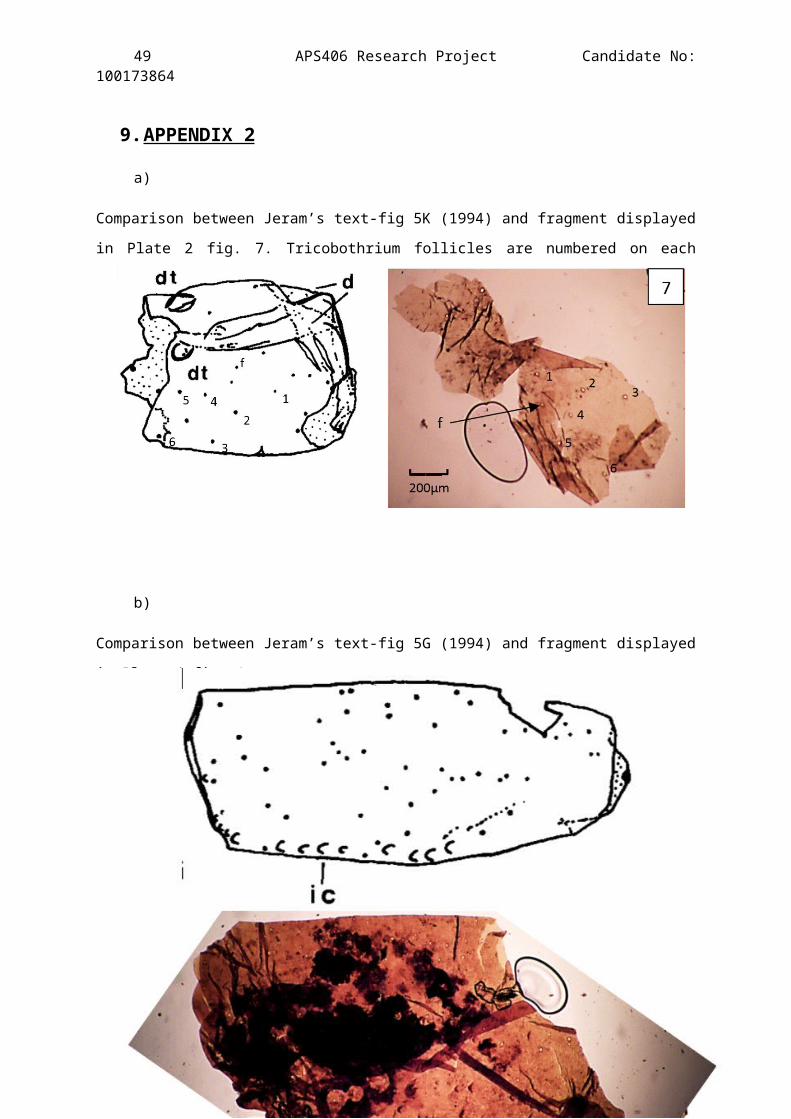
Figure 7 – A very light brown fragment, almost transparent. Thedistinct pattern of setal pits and size of the cuticle fragmentmeans it can be placed on the ventral side of a sternite.Comparable to Jeram’s drawing Text-fig. 3F (1994) (See Appendix2a). On close inspection it was apparent that one of the setalpits occurred on the folded over cuticle misleading its truelocation (f).
Figure 8 – Dark brown in colour, fragment of sternite with a fewscale tubercles and a number of setal pits present. Due to thespacing and rotation of the scale tubercles (st) matched withsimilarities in location and size to Jeram’s Text-fig. 3. E itwould appear to be the dorsal surface of a metasomal segment.

18 APS406 Research Project Candidate No:100173864

19 APS406 Research Project Candidate No:100173864
Plate 3
A compilation of Light Microscope images of the best cuticle
specimens extracted from the Clowne Common outcrop, horizon JBC1
(Slides 10, 13, 15, 16, 20 & 22).
Genus: Compsoscorpius (Petrunkevitch, 1949).
Determining characteristic: Same as Plate 2 (same horizon).
Description of Figures:
Figure 1 – A relatively thick dark brown fragment with a seeminglysporadic distribution of triangular and star shaped holes. Partof sternite. Holes may have been created by minute triangularspinules (not too different from the ones Wills (1959) found onPareobuthus salopiensis, Text-fig 1 (d)) being ripped away from thecuticle.
Figure 2 – Heavily degraded fragment. It is only possible to makeout one setal pit due to the nature of the degradation. Thefrayed edges of this fragment almost looks like it has flakedaway from a larger part during the acid maceration process.
Figure 3 – Exceptionally preserved tibia fragment of a walking legwith the 3-D structures of the annulae of the setal pits clearlyvisible protruding from the cuticle. Distinctive thick banding ispresent almost cutting the fragment in half. The structure is nottoo different from a part of the tibia described for Compsoscorpiuselegans by Jeram (1994) (Plate 3, fig 4).
Figure 4 – A large and extensively folded fragment. On highermagnification granules (g), setal pits (sp) and trichobothriafollicles are present (tf), as is a single still attached setaprobably preserved due to the folding of this structure (s). Thefolding was so intricate that it was impossible to unfold duringthe transition on to the slide. This distorts the visiblestructure and its true shape is undeterminable. The presence ofall these structures on such a large fragment proves that it wasthe ventral side of either a part of a head shield or uppersternite.
Figure 5 – An adaxial side view of part of a walking leg femur.Exceptionally preserved fragment. On increased magnification thetiny hole-like wax pores where the end of the wax canalspenetrate the epicuticle are visible. The distinct difference

20 APS406 Research Project Candidate No:100173864
between follicles for trichobothria (tf) and the touch sensitivehair (ts) here. Trichobothria protrude perpendicular to thecuticle whereas the touch sensitive hairs protrude at an anglegiving the follicle a more elliptical look where one half of theannulus protrudes further than the other half.
Figure 6 – Heavily degraded with only the procuticle layer partiallyleft with one fold to the right. Its cellular structure can beseen although no other microstructures can be confidently placedas they might be mistaken for tears and rips.

21 APS406 Research Project Candidate No:100173864

22 APS406 Research Project Candidate No:100173864
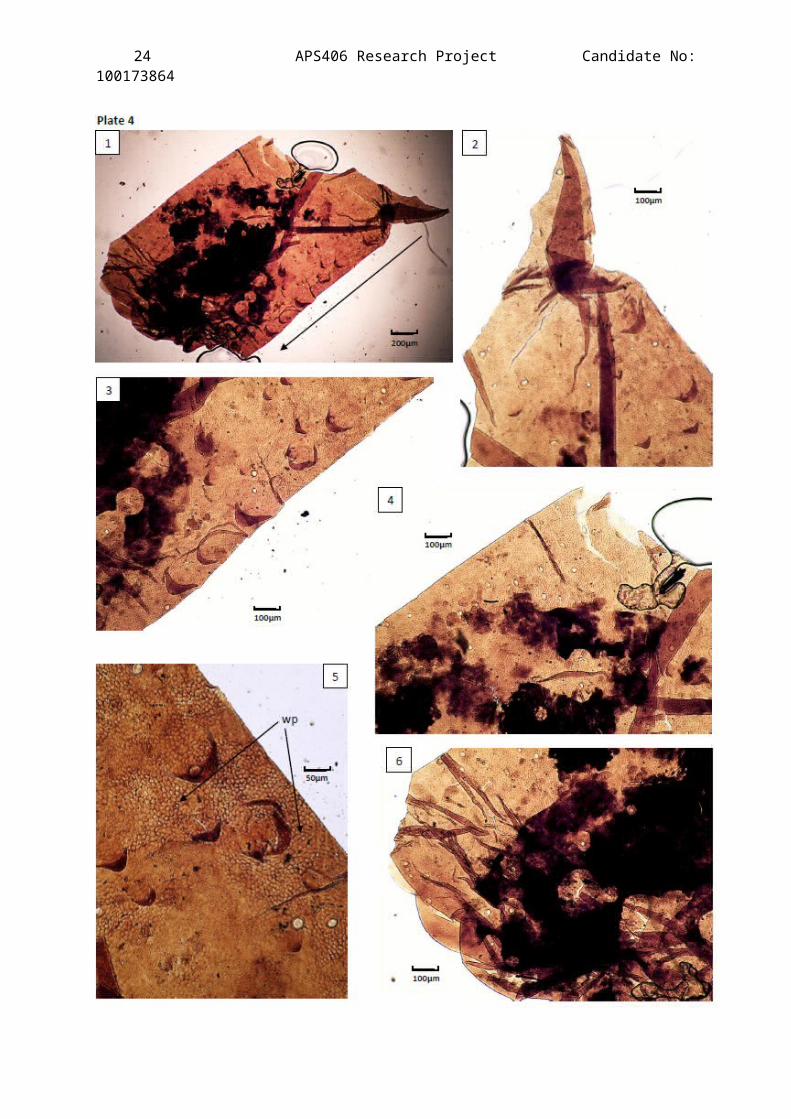
Plate 4
A compilation of Light Microscope images of the best preserved
cuticle fragment recovered from the Clowne Common outcrop. Horizon
JBC1 (Slide 19).
Species: Compsoscorpius buthiformis (Pocock, 1911).
Determining characteristic: ornament for ornament match with thedrawing of the prolateral side of the patella ofwalking leg 3 presented by Jeram (Text-fig. 5G,1994).
Description of Figures:
Figure 1 – A view of the prolateral side of the patella of walkingleg 3. Exceptional preservation of the full specimen showing its2D structure. The fragment is a fully enclosed flattenedcylindrical leg part. The pointed claw like structure in the topright of the figure is clinked fold in the cuticle that would, ifunfolded, run parallel to the scale tubercle ornamented edge. Itis hosting the largest number of microstructures found on any ofthe specimens presented in this investigation. Exceptionalpreservation of trichobothrium follicles, touch sensitive hairfollicles, scale tubercles & wax pores is apparent and thefragment is large enough to place it exactly in comparison withJeram’s detailed drawing of Compsoscorpius elegans’ patella of leg 3(text-fig 5G, 1994)(See Appendix 2b). The arrow is pointing tothe distal end. The dark pigmentation is most probably plantdebris trapped in the cuticle and when it was compressed the moredense follicles pushed it away forming the circles within thedebris.
Figure 2 – 2x magnification of figure 1 focusing on the claw likestructure where the folding can be seen clearly. The size, shapeand structure of the scale tubercles can be clearly seen due tothe immaculate preservation of the scale tubercles. The roughcuticle surface structure is characteristic of appendages of thefamily Palaeopisthacanthidea.
Figure 3 – 2x magnification of figure 1 showing the inferior carina(keel) of the patella of leg 3 of C. buthiformis.
Figure 4 – 2x magnification of figure 1 showing the proximalexternal part showing the distribution of setal pits. Notice the

23 APS406 Research Project Candidate No:100173864
lack of scale tubercles in this entire section – the ornamentalpattern of C. buthiformis on the patella.
Figure 5 – 4x magnification of figure 1 of the inferior carina. Thetransparency of the scale tubercles can be seen and wax pores areapparent (wp).
Figure 6 – 2x magnification of figure 1. Mostly obscured by plant debris and the crushed part of the fragment (distal end). The distribution of follicles can still be seen as well as the distinct lack of scale tubercles.
24 APS406 Research Project Candidate No:100173864

25 APS406 Research Project Candidate No:100173864
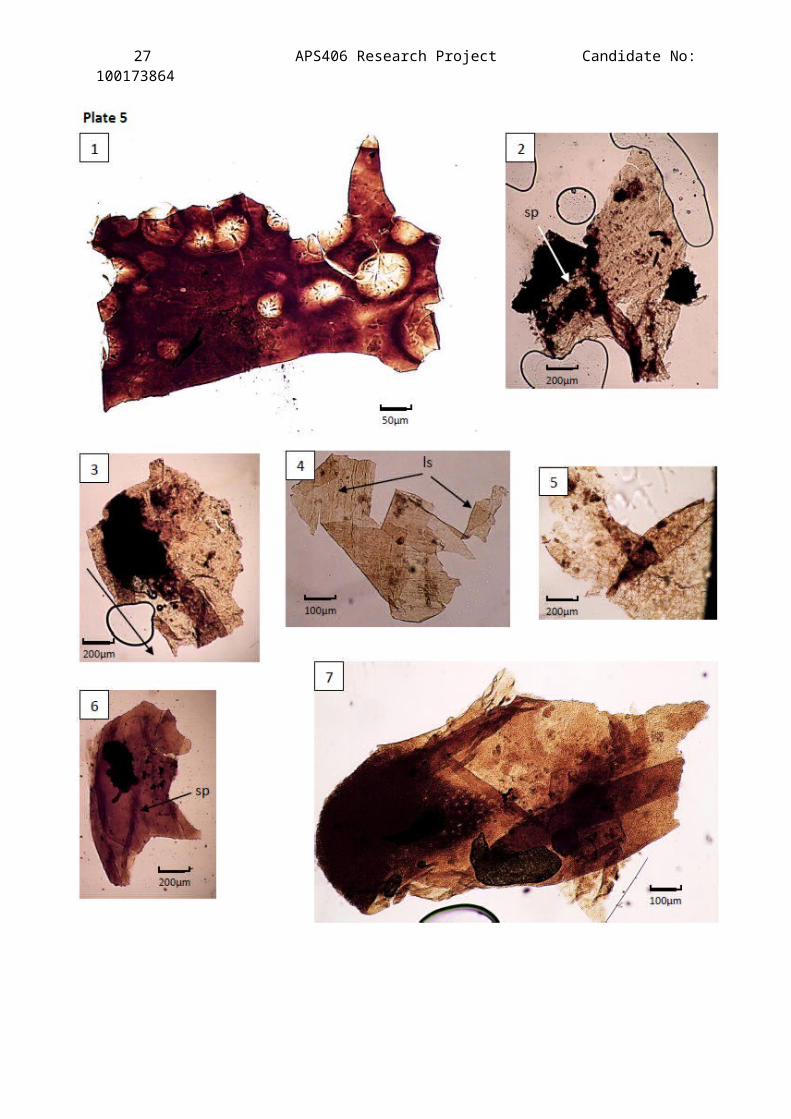
Plate 5
A compilation of Light Microscope images showing the best cuticle
specimens extracted from the Clowne Common outcrop, horizons JBC2
(Slides 1 & 2), JBC3 (Slide 1), JBC4 (Slide 1) and JBC8 (Slides 1-
3).
Not enough evidence to assign these fragments to one particulargenus two possible families have been suggested:Palaeopisthacanthidea (Compsoscorpius) and Eoscorpiidae.Determining characteristics: Ornamentation along the lateral side of
a ventral metasomal segment.
Description of Figures:
Figure 1 – A dark brown fragment of what appears to be arthropodcuticle. The diameters of the holes present are far too large tobe setal which suggests that they must be spinule holes. Withthere being very little documented on the distribution of spineswithin Carboniferous arthropods this cuticle has been dismissedfrom the discussion but left in the plates for the readers owninterpretation.
Figure 2 – Heavily degraded sclerite fragment with onedistinguishable setal pit (sp). The curvature of the cuticle andthe change in pigmentation at the bottom of the figure where theappendage attachment would have been to one of the walking legs.
Figure 3 – Moderately degraded fragment of a femur of one of thewalking legs. Some setal pits are visible but others are hard tomake out meaning their true distribution is hard to plot. Arrowpoints to the distal end of the femur.
Figure 4 - Faint dotted linear patterns can be seen (ls) which,although the folding distorts the true fragment structure, aresimilar to the patterns described by Jeram (1994) for Text-fig.3. (E): the far lateral side of a ventral metasomal segment.
Figure 5 – Heavily decayed cuticle showing the fibrous structure ofthe chitin. Microsctructures are uncertain but it is worth notingthe four setal pits in the top right of the figure which areclose enough together to imply that this fragment is from apedipalp chelicera.
Figure 6 – A sternite fragment viewed from the ventral side.Characteristic dark brown pigmentation. A distinct line of small

26 APS406 Research Project Candidate No:100173864
scale tubercles running down the centre. One setal pit isdistinguishable.
Figure 7 – Cuticle texture implies that this fragment comes from awalking appendage, most probably the femur judging by the size ofthe fragment and the heavy pigmentation to the proximal end.Integrated plant debris means that microstructures are difficultto determine but the clear speckled pattern could be micro-spinules characteristic of the upper ventral section of thefemur.
27 APS406 Research Project Candidate No:100173864

28 APS406 Research Project Candidate No:100173864
Plate 6
A compilation of Light Microscope images of the best cuticle
specimens extracted from the Clowne Common outcrop, horizons JBC8
(Slides 5, 7, 10 & 11) and JBCC (Slides 1, 2 & 8).
Genus: Compsoscorpius (Petrunkevitch, 1949).Determining characteristics: Chelicera showing external row of
teeth, and the pedipalp manus.
Description of Figures:
Figure 1 – A middle ventral sclerite fragment from the metasomal.Note the absence of scale tubercles, visible setal pits anduniform light pigmentation (dark areas are plant debris).
Figure 2 – Part of a pedipal manus with an overlapping fold of thefree finger onto the fixed finger. Comparable in setal pitdistribution and cuticle reflectivity to Jeram’s Gymnoscorpiusmutillidigitus pedipalp manus (Plate 4, Figure 13, 1994). However, thefingers appear to be a distinctly shorter in this fragment.
Figure 3 – Heavily degraded and folded. Deemed unassignable to anyappendage due to the lack of detail and microstructures visible.
Figure 4 – Degraded tibia fragment of a walking leg with closelydistributed setal pits and thick linear banding. The distancesbetween the two thick pigmented bands place this fragment on thetibia. The texture would be similar to figure 3 Plate 3 beforedegradation.
Figure 5 – Chelicera free finger showing the external row of teeth.Comparable to Plate 2; figure 10 in of Compsoscorpius elegans (Jeram,1994). Wax pores are visible and although most of the fragment isfolded the external teeth and setal pits are easy to make out.
Figure 6 – 2x magnification of figure 2. Folding of the free fingeris more visible.
Figure 7 – Fragment from the border of a sternite viewed from theinferior side. Lateral ornamentation along the cuticle indicatesits edge.
Figure 8 – Fragment of femur. Distinctive because of the line of granules (possibly scale tubercles) (g) running along the keel.

29 APS406 Research Project Candidate No:100173864
Heavily degraded but remnants of setal pits (sp) are also discernible. Thick line points towards the distal end.

30 APS406 Research Project Candidate No:100173864

31 APS406 Research Project Candidate No:100173864
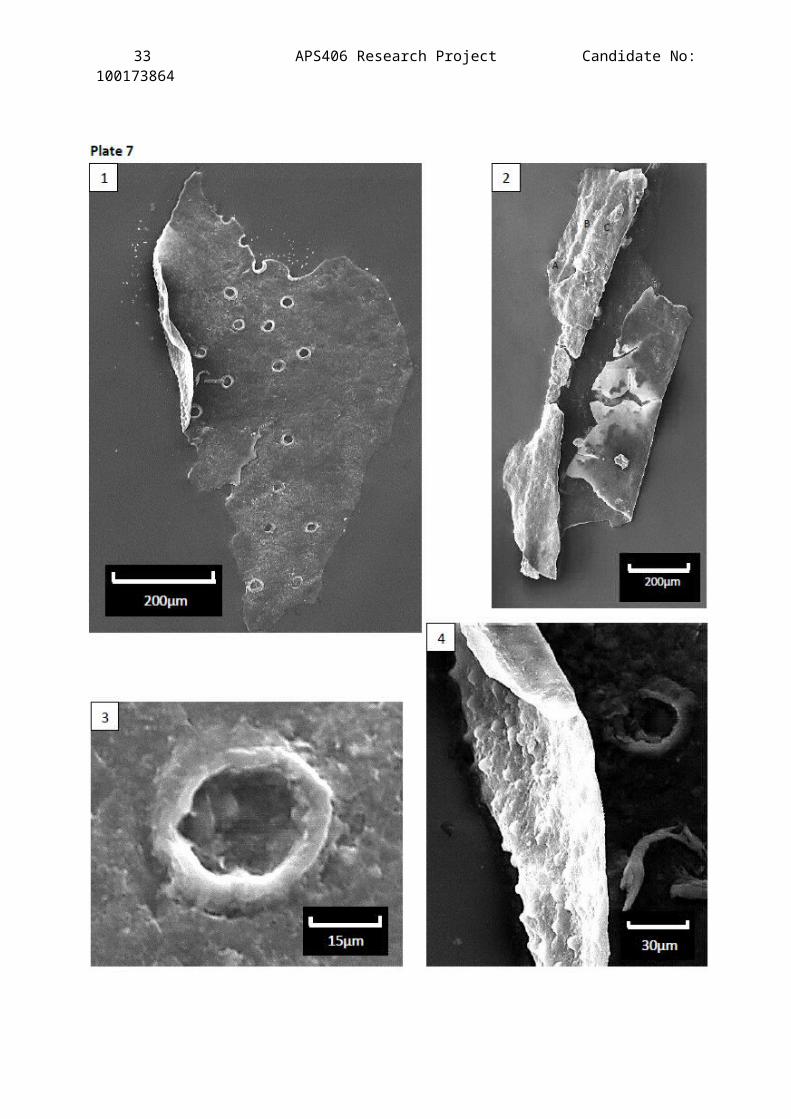
Plate 7
Scanning Electron Microscope images showing the 3-D structures of
two cuticle fragments from the Clowne Common outcrop horizon JBC1
(stub 1).
Description of Figures:
Figure 1 – Annulae of the setal pits are clearly protruding from theepicuticle surface. Setal pits are easier to determine than on LMimages, especially where the fragment is ripped. The distinctpattern of the setal pits is characteristic of a ventral sternitesegment.
Figure 2 – the folding of this cuticle shows the three distinctlayers that a classic scorpion exocuticle comprises of. A – theepicuticle, B – the procuticle and C – the endocuticle. Thelocation of this cuticle in the anatomy of a scorpion ininconclusive as the microstructures cannot be determined due tothe ‘curling’ of the cuticle.
Figure 3 – A highly magnified setal pit (same cuticle as figure 1)showing that the hole does not fully penetrate the cuticle butopens the epicuticle so that the base of the sensory hair organcan access the nervous system of the scorpion.
Figure 4 – Magnification of the cuticle displayed in figure 1showing the interior side of the exocuticle and it’s ‘roughness’in comparison to the smooth exterior side.
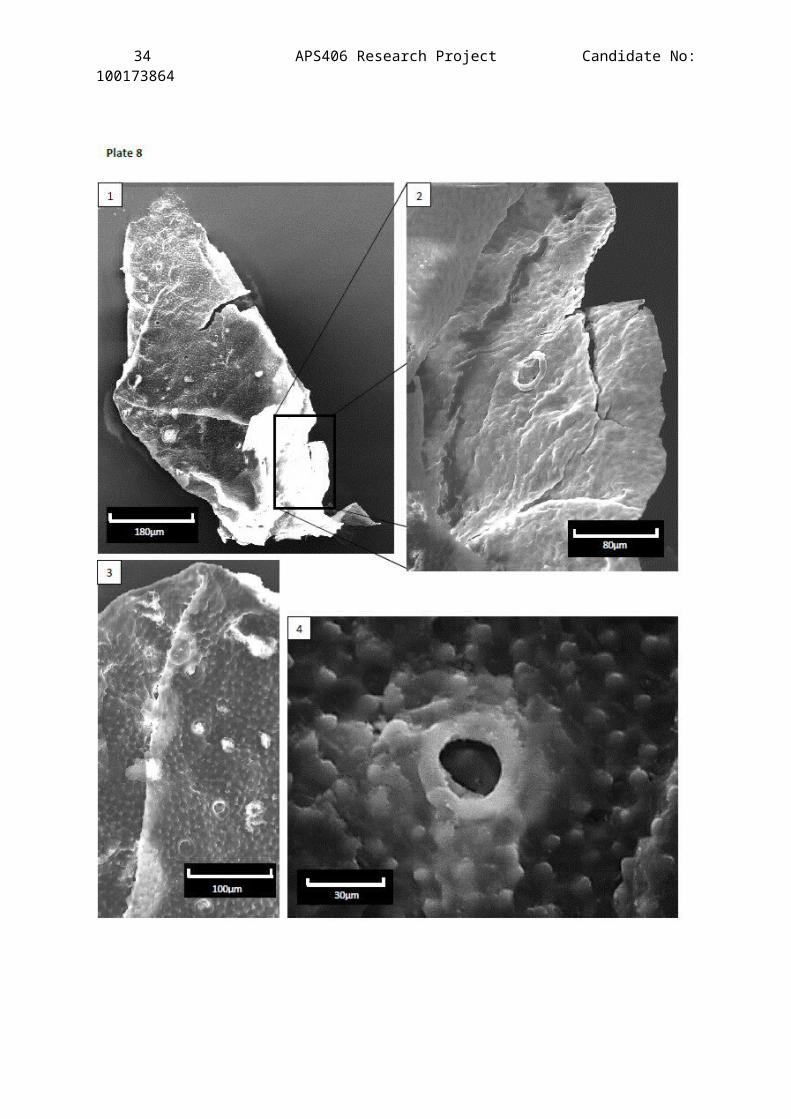
Plate 8
Scanning Electron Microscope images showing the 3-D structures of
one cuticle fragment from the Clowne Common outcrop horizon JBC1
(stub 3).
Description of Figures:
Figure 1 – a fragment of cuticle that has folded in on itselfpresenting the interior side of the exocuticle. The setal pitsare still easy to make out but protrude less prominently than onthe exterior side.
Figure 2 – Magnification of the cuticle where the folding hasuncurled a little to reveal the exterior surface of the

32 APS406 Research Project Candidate No:100173864
exocuticle which is not apparent in figure 1 due to the contrastof the image.
Figure 3 – Magnification of the fold showing setal pit structure onraised and unraised surfaces to help understand some distortionsof them on the LM images.
Figure 4 – A highly magnified setal pit from the interior side ofthe cuticle.
33 APS406 Research Project Candidate No:100173864
34 APS406 Research Project Candidate No:100173864

35 APS406 Research Project Candidate No:100173864
4.Discussion
The results produce conclusive evidence of only one species that was
able to be successfully described from the Clowne Common outcrop –
Compsoscorpius elegans (as described by Jeram, 1994) now known as
Compsoscorpius buthiformis (re-described by Legg et al., 2011). For that
reason the discussion is heavily focused on the mode of life of this
species and the main question being was it a terrestrial or aquatic
organism.
Wax pores
Wax pores have not been studied in any great detail relating to
Carboniferous scorpions, mainly because most species were thought to
be aquatic. The newly found presence of this microstructure during
cuticle analysis provides evidence for terrestrialisation (Plate 1,
fig. 3&6, Plate 4, fig. 5). They appear as tiny circular holes (<5µm
in diameter) were the wax canals penetrate the epicuticle and
secrete a layer of impermeable hydrophobic wax. This feature is key
evidence for the terrestrialisation of arthropods because it
protects the body from losing excessive water to the environment. It
was therefore a fundamental feature for primitive arthropods from
aquatic environments to survive on dry land.
Wax pores are a feature found in all extant species of scorpion
(e.g. Calchas birulai, Benli et al., 2009) and are key survival adaptations
for water retention in arid conditions. This somewhat contradicts
the interpreted palaeoenvironment of the Coal Measures’ non-arid
swampy forests (Taylor, 2000), but evolutionary steps toward
terrestrialisation may have needed this feature to overcome the
sudden change in environment (sub-aqua – to the comparatively dry
environment of the forest floor). The presence of wax pores the C.

36 APS406 Research Project Candidate No:100173864
buthiformis patella fragment (Plate 4, fig. 5) strengthens the argument
for the species’ terrestrialisation.
Peg-organs
The Carboniferous scorpions have always shown large similarities in
their general anatomical organisation, shape and even in some organ
structures to homologous features in Recent forms. The evidence of
the comb organ (tooth from comb organ: Plate 1, fig. 2) is perhaps
the most reliable item of evidence of their close relationship and
has been seen in many specimens (Wills, 1959). This distinct sense
organ is unknown to any other arthropod group, with the exception of
the doubtful claim for its existence in the Eurypterid Glyptoscorpius
(no peg-organs have been found on the teeth).
Despite pectines being sensory structures their true function is
still unclear. In recent scorpions however, they appear to be larger
in males and are believed to play a role in selecting sites for
reproduction (Jacob et al. 2004). However, their overall anatomy has
remained the same since the origin of scorpions so it becomes
conclusive evidence for the presence of scorpions but not the family
or genera.
Cuticular ornamentation
From the directly matched cuticles to the species Compsocorpius
buthiformis (Plate 4, Plate) it is apparent that it is granular.
Ornamentation of a similar kind has been reported from some other
fossil scorpions which were previously placed under the family
Palaeopisthacanthidae, predominantly Palaeopisthacanthus (Petrunkevitch,
1913); Cryptoscorpius (Jeram, 1994); and Corniops (Jeram, 1994). Jeram
(1994) named this granular ornamentation ‘palaeopisthacanthid-type’
cuticle. Granulation has been documented to increase with aridity
(Koch, 1981), which is an expected relationship and increasing the

37 APS406 Research Project Candidate No:100173864
surface texture would hinder air flow over the exoskeleton thus
reducing water loss and evaporation. Its occurrence in Compsoscorpius
may thereby signify a terrestrial adaptation. Nevertheless, this
contradicts the interpretation of the swampy palaeoenvironment and
there are records of other extant species with granular
ornamentation that live in non-arid environments (Prendini, 2001).
Trichobothria
The elongate sense organs, trichobothria, consist of elongate, thin
hairs that project from a cup-like cavity on the epicuticle, called
a bothrium (Reissland & Gorner, 1985). The hairs detect changes in
the air current aiding in predation (prey location). The presence of
them is therefore undisputable evidence for a terrestrial habit as,
due to the intense pressure and density of water, they would not
function aquatically (Selden & Jeram, 1989). Trichobothrial
arrangement and distribution is variable between scorpion families
and is therefore considered to be an important indicator of
systematic relationships (Soleglad & Fet, 2013). Full trichobothria
are seldom preserved during acid maceration and therefore their
presence is rarely recorded (with the exception of Jeram, 1994).
Bothria, however, have been reported in a number of Palaeozoic
scorpions, specifically Palaeopisthacanthus, Cryptoscorpius, Corniops and
Compsoscorpius. Recent XMT scans by Legg et al. have revealed bothria-
like structures on the dorsal surface of the pedipalp femur. Their
distribution is a lot sparser than in Recent scorpions suggesting it
was in the early stages of evolution in the Carboniferous.
Spiracles
The method of respiration in Palaeozoic scorpions is still in
contention as it is tied directly to the issue of
terrestrialisation. The similarities between book-gills in
xiphosurans and book-lungs in arachnids has been recognised for a

38 APS406 Research Project Candidate No:100173864
long time (Van Beneden, 1871) which has led to the belief of a
single terrestrialisation event, preceding the diversification of
the extant arachnids.
The book-gills of xiphosurans can function in air but must be
covered by a thin layer (Reisinger et al., 1991). The extension and
flexion of the abdominal plate muscles, repeatedly, aerates the
gills. In contrary, the scorpion book-lungs are internal. Aeration
is therefore achieved through the use of spiracles, which are slits
in the sternites, sometimes known as stigmata. Very few Palaeozoic
scorpions are known to have these and they are seen best in
Palaeopisthacanthus schucherti (Vogel & Durden, 1966). The side of a
marginal spiracle along the posterior edge of the abdominal plates
is presented in Plate 2 figure 2, which has been assigned to the
genus Compsoscorpius as the most evidence for that genus has been
recovered from the Clowne Common site. Jeram’s assignment of
cuticular macerates bearing spiracles to the genera Compsoscorpius was
an uncertainty due to the missing anatomy of the ventral mesosoma.
However, Legg et al. (2011) have re-examined the cuticle and revealed
structures interpreted as intramarginal spiricles (Fig 5.7-5.8 of
Legg et al., 2011). These features provide more evidence for the
terrestrial habitat of C. buthiformis.
Walking Legs & Coxapophyses
The legs of C. buthiformis bare a strong resemblance to those of extant
scorpions and are, from all records, clearly well-adapted for
terrestrial movement as they are laterally flattened. Legg et al.
conclude that the lateral compression of the appendage adds strength
to the leg allowing the scorpion to support its own weight out of
water (Dalingwater, 1985). Another indicator of the terrestrial
activity of C. buthiformis that was not recovered in this investigation
was their possession of an elongate telotarsus – the tip of the leg

39 APS406 Research Project Candidate No:100173864
which bears the claws – suggestive of plantigrade posture used in
terrestrial locomotion.
Further evidence was found in recent XMT scans conducted by Legg
et al. (2011) which showed the presence of elongate rod-like
structures that form the base of a pre-oral chamber. In extant
scorpions this forms the site of extra-oral digestion which is not
cannot occur effectively in a fluid medium.
Metasoma
Characteristic of the order Scorpiones, C. buthiformis possess a 5-
segmented metasomal with a telson that is modified into a sting. It
is suggested that due to the ratio of the fourth to fifth metasomal
somite C. buthiformis actively used its sting. Legg et al. (2011) also have
discovered that the metasomal is characteristically compressed in
the dorso-ventral plane which, amongst extant species tends to be
found in species that exploit cracks and crevices on the forest
floor. Unfortunately no telson parts or large metasomal fragments
were able to be recovered in this investigation so it is impossible
to speculate on the subject with relation to this study.
5.Conclusions
The evidence presented here indicates that only scorpion cuticle was
preserved well enough in the Clowne Common Coal Seam to undergo acid
maceration and then successful LM analysis. Other arthropod cuticles
may have been present but there was no possible way to determine the
Order of most of the degraded cuticles.
The discovery of an exceptionally preserved pecten tooth-organ,
displaying peg-organs (Plate 1, fig. 2) has led to the educated
assumption that it belonged to the species Compsoscorpius buthiformis.
Furthermore, the recovery of the patella fragment from walking leg 3
(Plate 4) provides conclusive evidence of the distribution of C.

40 APS406 Research Project Candidate No:100173864
buthiformis around the English Midlands, implying that it was a highly
successful and abundant species. Its success as one of the first
terrestrialised scorpions may help to explain the vast similarities
between this species and extant forms of scorpions (crown group
properties).
Another interesting factor was the presence of wax pores visible
on C. buthiformis (Plate 4, fig. 5). This provides further evidence for
the terrestrialisation of the species and strengthens its taxonomic
claim to the crown group of Recent scorpions.
However, with some regret many of the cuticle fragments
displayed in this investigation cannot be plausibly allocated to a
distinct genera due to insufficient collecting and the fragmentary
nature of many of the specimens. For example, although it would
appear that Plate 6, fig. 5 is part of a chelicera free finger from
C. buthiformis, the fragment is not large enough to display unique
ornamentation of the teeth.
6.Recommendations
There is huge variety in degradation between specimens suggesting
some fragmentation occurred before burial where parts were exposed
to harsher aerobic environments. However, it would be beneficial to
carry out a within coal maceral analysis to determine whether there
are different diagenetic rates within horizons or whether it is
consistent within a bed. This then allows assumptions of the within
horizon degradation of samples to be made. With conclusive evidence
it would be possible to speculate on whether degradation was due to
differential diagenesis or different arthropod types.

41 APS406 Research Project Candidate No:100173864
7.Bibliography
Allison, P.A., D.E.G. Briggs. (1993). Exceptional fossil record:Distribution of soft-tissue preservation through the Phanerozoic.Geology. 21, 527-530.
Anderson & Selden. (2007). Opisthosomal fusion and phylogeny ofPalaeozoic Xiphosura. Lethaia. 30, 19-31.
Baas, M., D.E.G. Briggs, J.D.H. van Heemst, A.J. Kear & J.W. Leeuw.(1995). Selective preservation of chitin during the decay ofshrimps. Geochimica et Cosmochimica Acta. 59, 945-951.
Barrington, E.J.W. (1979). Invertebrate Structure and Function. 2nd edition.Nelson, Sunbury on Thames.
Bartram, K.M., A.J. Jeram & P.A. Selden. (1987). Arthropod cuticlesin coal. Journal of the Geological Society, London. 144, 513-517.
Briggs, D.E.G., A.J. Kear, M. Baas, J.W. De Leeuw & S. Rigby.(1995). Decay and composition of the hemichordate Rhabdopleura:implications for the taphonomy of graptolites. Lethaia. 28, 15-23.
Crowson, R.A., W.D.I. Rolfe, J. Smart, C.D. Waterston, E.C. Willey &R.J. Wootton. (1967). Arthropoda: Chelicerata, Pycnogonida,Palaeoisopus, Myriapoda and Insecta. The Fossil Record. 499-534.
Dalingwater, J.E. (1973). The cuticle of a eurypterid. Lethaia. 6,179-186.
Dalingwater, J.E. (1985). Biomechanic approaches to eurypteridcuticle and chelicerate exoskeletons. Transactions of the Royal Society ofEdinburgh. 76, 359-364.
Dunlop, J.A., D. Penney & D. Jekel. (2011). A summary list of fossilspiders and their relatives. The world spider catalog, version, 12. 258.
Dunlop, J.A. & P.A. Selden. (1998). The early history and phylogenyof the chelicerates. In Arthropod Relationships. (pp. 221-235).Springer Netherlands.
Filshie, B.K. & N.F. Hadley. (1979). Fine structure of the cuticleof the Desert Scorpion, Hadrurs arizonensis. Tissue and Cell. 11, 249-262.
Jacob, A., Gantenbein, I., Braunwalder, M. E., Nentwig, W., & Kropf,C. (2004). Morphology and function of male genitalia(spermatophores) in Euscorpius italicus (Euscorpiidae,Scorpiones): Complex spermatophore structures enable safe spermtransfer. Journal of morphology. 260(1), 72-84.

42 APS406 Research Project Candidate No:100173864
Kjellesvig-Waering, E.N. (1986). A restudy of the Fossil Scorpionida of theWorld. Ed. A.S. Caster & K.E. Caster. Palaeontological ResearchInsitution, N.Y. USA.
Koch, L.E. (1981). The scorpions of Aurtalia: aspects of theirecology and zoogeography. P. 875-884. In Keast, A. (ed.), Ecologicalbiogeography of Australia. 41(2).
Labanderia, C.C. & G.J. Eble. (2000). The fossil record of insectdiversity and disparity. In J. Anderson, F. Thackeray, B. Van Wyk,& M. De Wit (Eds.). Gondwana alive: biodiversity and the evolving biosphere.Johannesburg: Witwatersrand University Press.
Legg, D.A., R.J. Garwood, J.A. Dunlop & M.D. Sutton. (2011). Ataxonomic revision of orthosternous scorpions from the EnglishCoal Measres aided by x-ray micro-tomography (XMT). PalaeontologiaElectronica. 15(1), 16.
McCoy, V.E. & D.S. Brandt. (2009). Scorpion taphonomy: criteria fordistinguishing fossil scorpion molts and carcasses. The Journal ofArachnology. 37, 312-320.
Mutvei, H. (1977). SEM Studies on Arthropod Exoskeletons. ZoologicaScripta. 6, 203-213.
Neville, A.C. (1975). Biology of the Arthropod cuticle. Springer Verlad.Berlin.
Novojilov, N., & L. Stormer. (1963). A new scorpion from the UpperCarboniferous of Siberia. Norsk Geologisk Tidsskrift. 43, 83-87.
Petrunkevitch, A. (1953). Palaeozoic and Mesozoic Arachnida ofEurope. Memoirs of the Geological Society of America. 53, 1-112.
Pocock, R.I. (1911). A monograph of the terrestrial CarboniferousArachnida of Great Britain. Monograph of the Palaeontographical Society.34, 1-84.
Prendini, L. (2001). Substratum specialization and speciation insouthern African scorpions: the Effect Hypothesis revisited. P.113-138. In Fet, V. and Selden, P.A. (eds.). Scorpions 2001. InMemoriam Gary A. Polis. British Arachnological Society, Bumham Beeches.
Reisinger, P.W.M., I. Tutter & U. Welsch. (1991). Fine structure ofthe horseshoe crab Limulus Polyphemus and Tachyplus tridentatus and of thebook lung of the spider Eurypelma californicum, 1871. Zoologisches Jahrbuchder Anatomie. 121, 331-357.
Richards, A.G. & F.H. Korda. (1948). Studies on Arthropod Cuticle,II, Electron Microscope Studies of Extracted Cuticle. The BiologicalBilletin. 93(3), 212-235.

43 APS406 Research Project Candidate No:100173864
Rippon, J.H. (1984). The Clowne Seam, Marine Band and overlyingsediments in the Coal Measures (Westphalian B) of NorthDerbyshire. Proceedings for the Yorkshire Geology Society. 45(1&2), 27-43.
Scott, A.C. (2002). Coal petrology and the origin of coal macerals:a way ahead? International Hournal of Coal Geology. 50, 119-134.
Scott, A.C. & G. Rex. (1985). The formation and significance ofcarboniferous coal balls. Philosophical. Transcripts of the. Royal. Society ofLondon. B. 311, 123-137.
Scott, A.C. & T.N. Taylor. (1983). Plant/Animal Interactions duringthe Upper Carboniferous. The Botanical Review. 49(3), 259-307.
Selden, P.A. & A.J. Jeram. (1989). Palaeophysiology ofterrestrialisation in the Chelicerata. Transactions of the Royal Society ofEdinburgh: Earth Sciences. 80:3-4, 303-310.
Shear, W.A. & J. Kukalová-Peck. (1990). The Ecology of Paleozoicterrestrial arthropods: the fossil evidence. Canadian Journal ofZoology. 68(9), 1807-1834.
Soleglad, M.E. & V. Fet. (2013). Evolution of ScorpionOrthobothriotaxy: A Cladistic Approach. Euscorpius, 2001(1), 1-38.
Stankiewicz, B.A., D.E.G. Briggs, R.P. Evershed, M.B. Flannery & M.Wuttke. (1997). Preservation of chitin in 25-million-year-oldfossils. Science. 276, 1541-1543.
Stankiewicz, B.A., A.C. Scott, M.E. Collinson, P. Finch, B. Mosle,D.E.G. Briggs & R.P. Evershed. (1998). Molecular taphonomy ofarthropod and plant cuticles from the Carboniferous of NorthAmerica: implications for the origin of kerogen. Journal of theGeological Society of London. 155, 453-462.
Taylor, K.J. (2000). A comparative study of the petrology andmegaspore content of an Upper Carboniferous (Westphalian B) coalseam: Clowne Coal, N.E. Derbyshire. University of Sheffield PalynologyDepartment Library (Thesis).
Todd. J.A. (1991). A forest-litter animal community from the UpperCarboniferous?: notes on the association of animal body fossilswith plants and lithology in the Westphalian D Coal Measures atWrithlington, Avon. Proceedings of the Geologists’ Association. 102(3), 179-184.
Van Beneden, E. (1871). [No title]. Annals of the Entomological Society ofBelgium. 1871, 9-11.
Vogel, B.R. & C.J. Dursden. (1966). The occurrence of stigmata in aCarboniferous scorpion. Journal of Paleontology. 40, 655-658.

44 APS406 Research Project Candidate No:100173864
Wills, L.J. (1925). The morphology of the Carboniferous ScorpionEobuthus Fritsch. Journal of the Linnean Society of London, Zoology. 36(241),87-97.
Wills, L.J. (1947). The British Triassic scorpions. PalaeontographicalSociety Monograph. 100(437) and 101(441), i-vi & 1-137, plates 1-12.
Wills, L.J. (1959). The external anatomy of some Carboniferous‘scorpions’, part 1. Palaeontology. 1, 261-282.
Wills, L.J. (1960). The external anatomy of some Carboniferous‘scorpions’, part 2. Palaeontology. 3, 276-332.
Woodward, H. (1907). Further notes on the Arthropoda of the BritishCoal-Measures. Geological Magazine (Decade V). 4(12), 539-549.

45 APS406 Research Project Candidate No:100173864
8.Appendix 1 Table of all known genera of Chelicerata (excluding
Pycnogonida) from the Carboniferous as of 2013. Genera in Red havebeen found in Britain.

46 APS406 Research Project Candidate No:100173864

47 APS406 Research Project Candidate No:100173864

48 APS406 Research Project Candidate No:100173864
49 APS406 Research Project Candidate No:100173864
9.APPENDIX 2
a)
Comparison between Jeram’s text-fig 5K (1994) and fragment displayed
in Plate 2 fig. 7. Tricobothrium follicles are numbered on each
picture. Pictures are out of rotation.
b)
Comparison between Jeram’s text-fig 5G (1994) and fragment displayed
in Plate 4 fig. 1





























