Embed Size (px)
Citation preview

Full Terms & Conditions of access and use can be found athttp://www.tandfonline.com/action/journalInformation?journalCode=kmab20
Download by: [T&F Internal Users], [Ms Rita Rosato] Date: 28 October 2015, At: 12:18
mAbs
ISSN: 1942-0862 (Print) 1942-0870 (Online) Journal homepage: http://www.tandfonline.com/loi/kmab20
A comparison of anti-HER2 IgA and IgG1 in vivoefficacy is facilitated by high N-glycan sialylation ofthe IgA
Gerard JA Rouwendal, Miranda M Van Der Lee, Saskia Meyer, Karli R Reiding,Jan Schouten, Guy De Roo, David F Egging, Jeanette HW Leusen, PeterBoross, Manfred Wuhrer, Gijs F Verheijden, Wim H Dokter, Marco Timmers &Ruud Ubink
To cite this article: Gerard JA Rouwendal, Miranda M Van Der Lee, Saskia Meyer, Karli RReiding, Jan Schouten, Guy De Roo, David F Egging, Jeanette HW Leusen, Peter Boross, ManfredWuhrer, Gijs F Verheijden, Wim H Dokter, Marco Timmers & Ruud Ubink (2015): A comparisonof anti-HER2 IgA and IgG1 in vivo efficacy is facilitated by high N-glycan sialylation of the IgA,mAbs, DOI: 10.1080/19420862.2015.1102812
To link to this article: http://dx.doi.org/10.1080/19420862.2015.1102812
View supplementary material Accepted author version posted online: 06Oct 2015.Published online: 06 Oct 2015.
Submit your article to this journal Article views: 32
View related articles View Crossmark data
Citing articles: 1 View citing articles

A comparison of anti-HER2 IgA and IgG1 in vivoefficacy is facilitated by high N-glycan sialylation
of the IgAGerard JA Rouwendal1,*, Miranda M van der Lee1, Saskia Meyer2, Karli R Reiding3, Jan Schouten1, Guy de Roo1,David F Egging1, Jeanette HW Leusen2, Peter Boross2, Manfred Wuhrer3,4, Gijs F Verheijden1, Wim H Dokter1,
Marco Timmers1, and Ruud Ubink1
1Synthon Biopharmaceuticals B.V.; Nijmegen; The Netherlands; 2Laboratory for Translational Immunology; University Medical Center Utrecht; Utrecht; The Netherlands;3Center for Proteomics and Metabolomics; Leiden University Medical Center; Leiden; The Netherlands; 4Division of BioAnalytical Chemistry; VU University
Amsterdam; Amsterdam, The Netherlands
Keywords: cysteine mutation, HER2, monoclonal IgA antibody, N-glycan sialylation, pharmacokinetics, transgenic FcaRImouse model
Abbreviations: ACN, acetonitrile; ADCC, antibody-dependent cell-mediated cytotoxicity; ADCP, antibody-dependent cellularphagocytosis; ASGPR, asialoglycoprotein receptor; AUC, area under the curve; CDC, complement-dependent cytotoxicity; CL, clear-ance rate; Cmax, peak plasma concentration; ER, endoplasmic reticulum; FcaRI, Fc-a receptor I; FcgR, Fcg-receptor; HER2, humanepidermal growth factor receptor 2; HMW, high molecular weight; MALDI-TOF MS, matrix-assisted laser desorption/ionization
time-of-flight mass spectrometry; mAbs, monoclonal antibodies; monocytes, MNC; NK, natural killer; PK, pharmacokinetic; PMN,polymorphonuclear leukocytes; SEC, size exclusion chromatography; SPR, surface plasmon resonance; t1/2, elimination half-life;
UPLC, ultra performance liquid chromatography; Vss, apparent volume of distribution at steady state.
Monomeric IgA has been proposed as an alternative antibody format for cancer therapy. Here, we present ourstudies on the production, purification and functional evaluation of anti-HER2 IgA antibodies as anti-cancer agents incomparison to the anti-HER2 IgG1 trastuzumab. MALDI-TOF MS analysis showed profound differences in glycosylationtraits across the IgA isotypes and cell lines used for production, including sialylation and linkage thereof, fucosylation(both core and antennary) and the abundance of high-mannose type species. Increases in sialylation proved topositively correlate with in vivo plasma half-lives. The polymerization propensity of anti-HER2 IgA2m2 could besuppressed by an 18-aa deletion of the heavy chain tailpiece - coinciding with the loss of high-mannose type N-glycanspecies - as well as by 2 cysteine to serine mutations at positions 320 and 480. The HER2 F(ab’)2-mediated anti-proliferative effect of the IgA2m1 and IgA2m2 subtypes was similar to IgG1, whereas the IgA1 isotype displayedconsiderably lower potency and efficacy. The Fc-mediated induction of antibody-dependent cell-mediated cytotoxicity(ADCC) using human whole blood ADCC assays did not demonstrate such clear differences between the IgA isotypes.However, the potency of the anti-HER2 IgA antibodies in these ADCC assays was found to be significantly lower thanthat of trastuzumab. In vivo anti-tumor activity of the anti-HER2 IgA antibodies was compared to that of trastuzumab ina BT-474 breast cancer xenograft model. Multiple dosing and sialylation of the IgA antibodies compensated for theshort in vivo half-life of native IgA antibodies in mice compared to a single dose of IgG1. In the case of the IgA2m2antibody, the resulting high plasma exposure levels were sufficient to cause clear tumor stasis comparable to thatobserved for trastuzumab at much lower plasma exposure levels.
Introduction
The cancer cell-killing capacity of monoclonal antibodies(mAbs) is determined by Fab- and Fc-mediated effects. Antigen-binding of a mAb to its cancer cell target may trigger a numberof Fab-mediated responses, i.e., internalization, apoptosis andgrowth inhibition. The antibody isotype can influence these Fab-mediated responses as for instance in the case of dimeric IgA or
pentameric IgM isotypes having 4 or 5 Fabs, respectively. Yet,therapeutic efficacy of the different isotypes is deemed to be pre-dominantly determined by the Fc-mediated interactions withmyeloid effector cells and complement, i.e., interactions that caninduce cell killing via antibody-dependent cell-mediated cytotox-icity (ADCC), antibody-dependent cellular phagocytosis(ADCP) and complement-dependent cytotoxicity (CDC).1,2
The most commonly used antibody format comprises an IgG1
*Correspondence to: Gerard Johan Rouwendal; Email: [email protected]: 06/29/2015; Revised: 09/17/2015; Accepted: 09/28/2015http://dx.doi.org/10.1080/19420862.2015.1102812
www.tandfonline.com 1mAbs
mAbs 0:0, 1--13; October 2015; © 2015 Synthon BiopharmaceuticalsREPORT
Dow
nloa
ded
by [
T&
F In
tern
al U
sers
], [
Ms
Rita
Ros
ato]
at 1
2:18
28
Oct
ober
201
5

backbone or engineered versions thereof designed to modulateinteractions with activating and inhibitory Fcg-receptors(FcgRs), and thereby increase cancer cell killing even further.3
The first indications that IgA might be an alternative antibodyformat for cancer therapy came from ADCC experiments thatwere either done with bispecific F(ab’)2 antibodies, in which oneFab was directed at the Fc-a receptor I (FcaRI) and the other atthe target antigen, or with recombinant monomeric IgA1.4,5
One of the anticipated advantages of using IgA was its capacityto also act via the abundant polymorphonuclear leukocytes(PMN), whereas IgG1 is thought to mainly employ natural killer(NK) cells and macrophages as effector cells.6,7 Moreover, IgA-mediated FcaRI activation appears to actively recruit PMNtoward tumor cells.8,9 Since tumor-killing by IgA depends onactivation of the FcaRI and that of IgG1 is assumed to be mainlymediated by FcgRIIIa, it might be superior to the correspondingIgG1 when treating patients carrying the FcgRIIIa F158 allele inwhom IgG1 efficacy is reduced compared to carriers of the V158allele.10,11,12 Very recently, it has been demonstrated not onlythat macrophages can mediate tumor cell killing by IgA2, butmore importantly that this occurs in vivo.13
IgA antibodies have attracted much less attention than IgGantibodies. One of the (practical) reasons might be the fact thatthis isotype utilizes the FcaRI, a receptor that is absent frommice, i.e., the standard animal for pre-clinical testing of cancertherapeutics. However, transgenic mice expressing the humanFcaRI were developed more than a decade ago.14,15 Additionaldiscouragements might have been the significantly lower half-lifeof IgA compared to IgG1, the complexity of IgA N- (and O-)glycosylation profiles complicating control of product consis-tency and the general inexperience with commercial scale IgAproduction in mammalian cell culture and the subsequentpurification.
In humans, secretory IgA is by far the most abundantly pro-duced immunoglobulin, with rates up to about 6 gram per dayin epithelial layers. Serum levels of mainly monomeric and somedimeric IgA are 15-20% of total immunoglobulin levels and con-sist of isotypes IgA1 and IgA2, with the latter occurring as one of2 allotypes, IgA2m1 being predominant in Europe and IgA2m2having the highest frequency in Africa.16 The main differencesbetween IgA1 and IgA2 are located in the heavy chain hingeregion where the IgA1 isotype is 13-amino acid residues longerthan IgA2, and where only this IgA isotype is decorated with upto 5 O-glycans.17 In addition, IgA1 has 2 N-glycosylation sites,whereas the IgA2m1 and IgA2m2 isotypes comprise 4 or 5 N-glycosylation sites, respectively, containing N-glycans with highercomplexity than what is usually found for IgG antibodies.18 TheIgA2m1 allotype is remarkable among human IgA for lacking acovalent disulfide bond between heavy and light chains.19
A meaningful investigation into the potential value of (mono-meric) IgA in oncology can only be performed by direct compari-son with a standard IgG1 having identical variable domains. Inthis study, trastuzumab was compared to different human IgAisotypes containing the trastuzumab variable domains. To thisend, each of the 3 known human IgA isotypes (IgA1, IgA2m1,IgA2m2) and several variants carrying the trastuzumab variable
domains were produced. Production formats include stable PER.C6 pools co-expressing a human a2,3-sialyltransferase or tran-sient expression in HEK293F cells. Pharmacokinetic (PK) studiesdemonstrated a clear correlation between the N-glycan sialylationand half-life in serum. Cysteine to serine mutations in the Ca2domain and the tailpiece of an anti-HER2 IgA2m2 antibodywere shown to reduce undesirable polymerization of (mono-meric) IgA thus reducing a purification bottleneck. ADCC assaysusing human whole blood from different donors as effector cellsshowed that IgA potency tended to be notably lower than that ofIgG1, though maximum cell killing by IgA could sometimes behigher than by IgG1. Isolated human PMN confirmed specificADCC activity mediated by IgAs and lack of activity by IgG1.Finally, a subcutaneous BT-474 xenograft model was used intransgenic severe combined immunodeficient (SCID) miceexpressing the human FcaRI to test in vivo efficacy of the variousanti-HER2 IgA isotypes compared to trastuzumab. These studiesindicate that, in this particular model, the tumor-inhibitorycapacity of the IgG1 antibody was evidently more efficaciousthan the best IgA antibody, despite IgA serum levels that wereequal or even higher than that of the IgG1 antibody.
Results
Production of monomeric IgA from stably transformedPER.C6 or transiently transfected HEK293F
The three anti-HER2 IgA antibodies representing the 3known human IgA isotypes were generated using stable pools ofPER.C6 cells co-transfected with 2 vectors, one encoding thehuman IgA heavy chain and a kappa light chain gene with thecorresponding trastuzumab variable domains and the otherencoding the human a2,3-sialyltransferase ST3GAL4 (Table 1).In addition, a stable pool expressing the human anti-HER2IgA2m1 isotype in the absence of ectopic ST3GAL4 was alsoused for antibody production. The actual production runs wereperformed in 10 liter cultures using 20 liter wave bags andyielded IgA levels between 110–190 mg/L as detected by Alpha-LISA. Size exclusion chromatography (SEC) analysis of KappaSe-lect-purified IgA revealed the presence of substantial amounts of
Table 1. Overview of anti-HER2 IgA antibodies used in this study.
Antibody IgA isotype Host cell Modifications
IgA1-1g5 IgA1 PER.C6 Ectopic a2,3-sialyltransferase
IgA21-2g4 IgA2m1 PER.C6IgA21-2g5 IgA2m1 PER.C6 Ectopic a2,3-
sialyltransferaseIgA22-4g5 IgA2m2 PER.C6 Ectopic a2,3-
sialyltransferaseIgA22-4g2 IgA2m2 HEK293FIgA22D18-5g2 IgA2m2 HEK293F Deletion of tailpieceIgA22-7 IgA2m2 HEK293F C480SIgA22-8 IgA2m2 HEK293F C320SIgA22-9 IgA2m2 HEK293F C320S, C480SIgA22-10 IgA2m2 HEK293F C320S; Deletion of
tailpiece
2 Volume 0 Issue 0mAbs
Dow
nloa
ded
by [
T&
F In
tern
al U
sers
], [
Ms
Rita
Ros
ato]
at 1
2:18
28
Oct
ober
201
5

higher molecular weight (HMW) species in addition to mono-meric IgA and free light chain (Fig. 1). The molecular and bio-logical traits of IgA were determined following a finishing roundof purification by SEC. Using UPLC-SEC, the distribution ofmonomeric, dimeric and multimeric species was quantified.Thus, it was shown that all preparations contained more than90% of the monomeric species (Table S1). The two preparationscontaining wild-type IgA2m2 from PER.C6 and HEK293F con-tained the highest levels of multimers at 3.4 and 3.6%, respec-tively, whereas in contrast none could be detected in the IgA2m2preparation lacking the tailpiece. The final yields of purifiedmonomeric IgA ranged between 10 and 15.7 mg/L. Besides IgAproduced with PER.C6 pools, a number of antibodies were alsoproduced by transient expression in HEK293F. All of these anti-bodies were based on the anti-HER2 IgA2m2 sequence usedabove, but contained one or more mutations, or deletion of thetailpiece. The aim of producing the mutants was to investigatethe nature of the HMW species as well as the putative gel arti-facts that were observed.
Characterization of purified IgAPurified, mainly monomeric IgA variants (Table 1) were
also characterized by non-reducing SDS-PAGE. IgA22-4g5and IgA1-1g5 showed one predominant band at a molecularweight around 160 kDa (Fig. 2), which likely represents thefully assembled molecule with 2 heavy chains and 2 light
chains (H2L2). The same 160 kDa band was also visible inIgA21-2g4 and ¡2g5, albeit at a relatively low level, butinstead these samples displayed major amounts of 115 kDaand 45 kDa proteins, which likely represented heavy (H2) andlight chain dimers (L2), respectively. Several more, but lessconspicuous, bands could be detected, especially in the IgA21-2g4 and ¡2g5 samples that most likely corresponded to speciesconsisting of folded and assembled heavy-heavy-light chains(H2L) and light chain (L). The HMW species above 160 kDaprobably consisted of di- and polymeric forms of IgA thatseemed to be coupled through their tailpieces since, when the18-aa C-terminal tailpiece (containing the penultimate C-ter-minal cysteine) was removed from the anti-HER2 IgA2m2,hardly any HMW species were formed upon transient expres-sion in HEK293F cells (Fig. 1; IgA22D18-5g2). However, theremoval of the tailpiece seemingly caused a deterioration ofantibody stability during electrophoresis as a number of addi-tional bands emerged under non-reducing conditions comparedto intact IgA22-4g2 also produced in HEK293F (see below).HER2 binding of the IgA antibodies using either surface plas-mon resonance (SPR) or cellular binding showed that bindingstrengths of the antigen to the monomeric IgA preparationswere very similar to trastuzumab (Table S2 and Fig. S2).Using SPR to compare binding to the FcaRI receptor, approxi-mate 2-fold differences were found between the monomericIgA preparations in observed KD (KDobs)(Table S3).
Figure 1. Preparative SEC profiles of protein samples from PER.C6 stable pools expressing IgA1-1g5 (A), IgA21-2g4 (B), IgA21-2g5 (C) and IgA22-4g5 (D)and from HEK293F cultures IgA22-4g2 (E) and IgA22D18-5g2 (F) after clarification and KappaSelect purification. HMW D high-molecular weight fraction;LC D light chain.
www.tandfonline.com 3mAbs
Dow
nloa
ded
by [
T&
F In
tern
al U
sers
], [
Ms
Rita
Ros
ato]
at 1
2:18
28
Oct
ober
201
5

Mutation and deletion analysis of anti-HER2 IgA2m2The ostensible instability of IgA22D18-5g2 on non-reducing
SDS-PAGE prompted a further investigation into the role offree thiols in the appearance of the additional bands. To thisend, free thiols were chemically blocked with N-ethylmaleimide(NEM). Non-reducing SDS-PAGE of NEM-treated IgA22D18-5g2 greatly reduced the intensity of these bands, which sug-gested that the penultimate cysteine in its tailpiece-containingIgA22-4g2 counterpart was involved in stabilizing this antibodyduring gel electrophoresis (Fig. S3). A series of mutants weredesigned in which either of the 2 cysteines in the IgA heavychain at positions 320 and 480, which are potentially engagedin SC- and J-chain interactions when present in a secretory IgA,were mutated to serine, or in which the tailpiece was deleted.These mutant antibodies and the wild-type sequences were tran-siently expressed in HEK293F and affinity purified using Kap-paSelect resin only, which yielded intact IgA and light chainmonomers and dimers. Tailpiece deletion (Fig. 3, lane 2)greatly affected the banding pattern compared to the wild-type(Fig. 3, lane 1) and yielded a profile that was very similar toantibody with the C480S mutation (Fig. 3, lane 3). C320Smutation generated a very neat profile lacking some of the lowermolecular weight by-products, but it still showed a significantlevel of HMW product (Fig. 3, lane 4). Strikingly, combining
C320S and C480S into one antibody, or combining C320Swith tailpiece deletion, provided virtually identical profiles con-sisting mainly of (monomeric) IgA and hardly any gel artifactsor HMW species (Fig. 3, lanes 5 and 6).
N-glycosylation of the IgA preparationsMALDI-TOF MS was used to study the N-glycans released
from the various antibodies, with derivatization of the sialic acidsto enhance their stability and allow the discrimination of a2,3-and a2,6-linked variants. Profound N-glycosylation differenceswere revealed between the IgA isotypes produced in PER.C6 sta-ble pools and between antibodies of the same IgA isotype butfrom different expression systems. Structural characterization wasperformed by MALDI-TOF/TOF MS/MS, proving informativein determining antennary compositions and fucose locations(Table S3).
N-glycans from the antibodies IgA1-1g5 and IgA22-4g5 pro-duced in PER.C6 cells with ectopic a2,3-sialyltransferase turnedout to be highly sialylated with those from IgA1-1g5 beingremarkable in having very low levels of core fucose (Fig. 4). Thebulk of the sialic acid residues in these N-glycans were a2,3-linked and a minority of N-glycans was mono-substituted witha2,6-linked residues. In contrast, N-glycans from antibodiesIgA21-2g4 and ¡2g5 mainly consisted of bi-antennary oligosac-charides that were poorly substituted with single a2,3- or a2,6-linked sialic acid residues. At first glance, the 2 antibodies IgA21-2g4 and ¡2g5 produced without and with ectopic a2,3-sialyl-transferase activity, respectively, displayed very similar N-glycanprofiles dominated by a small number of peaks representing bi-antennary N-glycans with both antennae either carrying a fucose(Lewis A or X epitopes) or with one antenna instead carrying asialic acid (visible at m/z 2101.771, 2228.793 and 2274.836).Yet, average sialylation of IgA21-2g5 (41.3%) produced in thepresence of ectopic a2,3-sialyltransferase was considerably higherthan that of IgA22-2g4 (23.6%) (Table S4). In both IgA1-1g5and IgA22-4g5, average sialylation - mostly by a2,3-linked sialicacid residues - is 2-4 fold higher and amounts to 85.3% and94.0%, respectively. In addition, IgA22-4g5, and especiallyIgA1-1g5, contained very few LewisA/X structures, while theseglyco-epitopes were found to be abundant on IgA21-2g4 and¡2g5. Three of the 4 IgA antibodies contained a small, but sig-nificant level of the high-mannose species Man5GlcNAc2 (m/z D1257.423). N-glycan profiles of the HEK293F-produced anti-bodies IgA22-4g2 and IgA22D18-5g2 were found to be quite dif-ferent from those of PER.C6-produced IgA. The strikingdissimilarity between N-glycan profiles from HEK293F- andPER.C6-produced antibodies was due to those from HEK293F-produced antibodies being relatively poorly galactosylated andsialylated, lacking LewisA/X epitopes and displaying significantlevels of bisected N-glycans. Remarkably, the loss of the tailpiecein IgA22D18-5g2 was associated with a decrease of high-man-nose N-glycans beyond Man5GlcNAc2, as seen by the percentageof high-mannose type compositions dropping from 21.0% inIgA22-4g2 to 2.1% in IgA22D18-5g2, and the average highmannose size changing from 7.08 to 5.91 mannose residues(Table S5). As expected, sialic acid residues on N-glycans of
Figure 2. Non-reducing SDS-PAGE of IgA22D18-5g2 (lane 1), IgA22-4g5(lane 2), IgA21-2g4 (lane 3), IgA21-2g5 (lane 4), IgA1-1g5 (lane 5), andIgA22-4g2 (lane 6) obtained by consecutive KappaSelect and SEC purifi-cation steps, and visualized using Criterion TGX Stain-free, precast gelsand Krypton staining. Putative composition of the various bands hasbeen indicated on the right side of the gel.
4 Volume 0 Issue 0mAbs
Dow
nloa
ded
by [
T&
F In
tern
al U
sers
], [
Ms
Rita
Ros
ato]
at 1
2:18
28
Oct
ober
201
5

antibodies coming from thehuman cell line HEK293F werefound to be either a2,3- or a2,6-linked and the sialylation level ofthese IgA antibodies was as low asthat of IgA21-2g4 and ¡2g5.
Cell-based assaysThe ability to inhibit cell pro-
liferation has been studied inHER2-expressing BT-474 breastcarcinoma cells after 6 days oftreatment with anti-HER2 IgAantibodies and compared to theIgG1 antibody trastuzumab. Cellgrowth was inhibited by IgA2antibodies with a similar efficacyand IC50 value as IgG1, whereasIgA1-1g5 was 3-fold less potentand only able to partially inhibitcell growth (Fig. 5). In theADCC assay using human wholeblood as effector cells and theBT-474 human breast carcinomaas target cells, all IgA isotypesshowed similar potency and effi-cacy after 4 h incubation (Fig.S4). Using three donors and theSK-BR-3 human breast carcinomaas target cells, the EC50 values forthe IgA antibodies rangedbetween 30 and 340 ng/mL, andfor IgG1 from 3-7 ng/mL (Fig.S5). The maximal antibody lyticactivity was donor- and isotype-dependent, ranging from 50-80%lysis. Hence, whereas no clear dif-ference in lysis efficacy wasobserved between IgA and theIgG1, the IgG1 variant was con-sistently at least 7-fold morepotent using SK-BR-3 as targetcells. When using whole blood incombination with BT-474 as tar-get cells, IgG1 was 25-fold morepotent than the most active IgAantibodies IgA1-1g5 and IgA22-4g5 (Fig. 6). Following separationof donor cells into different effec-tor cell populations, the ADCCactivity of IgA with PMN wasfound to be much higher thanwith IgG1, whereas IgA and IgG1were equally effective in inducingADCC activity exerted by mono-cytes (Fig. 7). Whole blood
Figure 3. Non-reducing SDS-PAGE of IgA from transiently producing HEK293F following KappaSelect purifi-cation. Lanes: (1) IgA22-4g2, (2) IgA22D18-5g2, (3) IgA22-7, (4) IgA22-8, (5) IgA22-9, (6) IgA22-10. Diagramsdepict cysteines at heavy chain positions 320 and 480 (C), serines at heavy chain positions 320 and 480 (S)and the C-terminal 18 aa tailpiece. Gels were visualized using Criterion TGX Stain-free, precast gels and Kryp-ton staining. Putative composition of the various bands has been indicated on the right side of the gel.
Figure 4. MALDI-TOF MS analysis of N-glycans from IgA1-1g5, IgA21-2g4, IgA21-2g5, IgA22-4g5, IgA22-4g2and IgA22D18-5g2 purified as described in Figure 2 analyzed in positive mode. The diagrams indicate peakswith S/N of >10 in the range of m/z 1200-2400. Blue square indicates N-acetylglucosamine; red triangle,fucose; green circle, mannose; yellow circle, galactose; purple diamond, N-acetylneuraminic acid. Linkagepositions of sialic acid residues are indicated by differing angles.38
www.tandfonline.com 5mAbs
Dow
nloa
ded
by [
T&
F In
tern
al U
sers
], [
Ms
Rita
Ros
ato]
at 1
2:18
28
Oct
ober
201
5

from FcaRI-transgenic mice that mainly contains PMN aseffector cells was likewise observed to mediate ADCC activityin HER2-expressing SK-BR-3 tumor cells (Fig. S6).
Pharmacokinetic (PK) analysisAfter 10 mg/kg intravenous dosing in wild type BALB/c mice,
the PK properties of the different IgA antibodies were highlydependent on the glycosylation status (Fig. 8). The highly sialy-lated IgA1-1g5 and IgA22-4g5 showed similar PK with anAUClast of 1391 and 1827 h*ug/mL, a clearance of 6.0 and4.8 mL/h/kg and a terminal half-life of 32.2 and 27.4 h, respec-tively (Table 2). In contrast, the poorly sialylated IgA21-2g5 and¡2g4 from PER.C6 cells and IgA22-4g2 and IgA22D18-5g2from HEK293F showed a 3-60-fold higher clearance compared
to IgA1-1g5. Intraperitoneal dosing of the highly sialylated IgA1-1g5 and IgA22-4g5 resulted in similar exposure, as expected(Fig. S7). Snapshot PK sampling in xenograft studies, 24 hoursafter the 10th daily IgA dose showed that IgA1-1g5 and IgA22-4g5 reached plasma concentrations that were at least 10-foldhigher at a 25 mg/kg IgA dose than the level of the correspond-ing IgG1 at a single 5 mg/kg dose (Table S6). Multiplying theIgA exposure by 10 for the multiple dose and by 2.5 to compen-sate for the dose difference, results in a theoretical IgA22-4g5AUC of 45675 h*ug/mL, compared to published trastuzumabAUC of 9761 h*ug/mL.20
Xenograft studiesIn the BT-474 xenograft studies in FcaRI-transgenic SCID
mice, single dose treatments with trastuzumab were compared tomultiple dosing with the short-lived IgAs to compensate for largedifferences in exposure levels. The different dose levels of trastu-zumab resulted in a dose-dependent decrease of tumor growth(Fig. 9A). Already at 1 mg/kg, tumor growth was delayed for 10– 20 days, and at 5 mg/kg the IgG1 induced tumor stasis foralmost 40 days, whereas actual tumor regression occurred at25 mg/kg. In contrast, IgA1-1g5 administered on 10 consecutivedays clearly was much less efficacious. Unlike trastuzumab, thelow-dose regimen at 1 mg/kg hardly affected tumor growth. Asimilar delay in tumor growth of about 20 days was seen at both5 and 25 mg/kg (Fig. 9A), which was surprising in view of theclear difference in plasma exposure between them (Table S6). Inthe BT-474 xenograft study shown in Figure. 9B, trastuzumabwas dosed once at the intermediate 5 mg/kg single dose; it dem-onstrated tumor reduction comparable to that in the study shownin Fig. 9A. In contrast to the results obtained with IgA1-4g5, themultiple doses of IgA22-4g5 did achieve a clear dose-dependentreduction of tumor growth at 5 mg/kg, even leading to tumorstasis at 25 mg/kg, i.e., comparable to the efficacy of a single5 mg/kg dose of trastuzumab. Likewise, in a HER2-overexpress-ing A431 intraperitoneal tumor model, but with IgA22-4g5administered directly to the peritoneum, this IgA showed tumorgrowth inhibition in FcaRI-transgenic SCID mice (Fig. S8). In
general, IgA antibodies dosed inthe same tumor model, but usingnon-transgenic SCID mice, dis-played no significant tumorgrowth inhibition (data notshown).13
Discussion
One of the reasons for a pau-city of research into the potentialvalue of anti-cancer IgAs inoncology might be due to thebelief that it would be difficult toproduce high levels of antibodywith consistent quality. Here, weobserve that titers of the anti-
Figure 5. Proliferation inhibition assay of IgA1-1g5, IgA21-2g4, IgA21-2g5, IgA22-4g5 purified as described in Figure. 2, IgG1 trastuzumab andisotype controls. BT-474 cells were incubated with the indicated antibod-ies for 6 days before cell viability was measured using ATP CellTiter-Gloassay. nD 3 replicates; § SEM.
Figure 6. ADCC assays with IgA1-1g5 and IgA22-4g5 purified as described in Figure 2, IgG1 trastuzumab andisotype controls using human whole blood as effector cells and BT-474 (A) or SK-BR-3 (B) as target cells. Celllysis was measured after 24 hours. Representative graphs of data from one donor per target cell are depicted.n D 3 replicates; § SEM.
6 Volume 0 Issue 0mAbs
Dow
nloa
ded
by [
T&
F In
tern
al U
sers
], [
Ms
Rita
Ros
ato]
at 1
2:18
28
Oct
ober
201
5
HER2 IgA antibodies produced in pools of stable, transfectedPER.C6 cells used in this study were similar to that of trastuzu-mab (data not shown). However, the final yield of monomericIgA from the large scale production runs was relatively low dueto the fact that a large amount of IgA is produced in the form ofdimers and polymers instead of monomers, despite the absenceof a J-chain.21 This is in agreement with earlier research showingthat J-chain incorporation is not required for polymerization ofantibodies containing the IgA tailpiece.22 As expression of ananti-HER2 IgA2m2 lacking the penultimate cysteine or even theentire heavy chain tailpiece yielded mostly monomeric species, itcan be concluded that dimers and polymers were primarily dueto tailpiece interactions probably involving disulfide bridge for-mation between the penultimate cysteines.23 The non-reducingSDS-PAGE revealed that especially the IgA2m1 preparationseemed to be unstable as the profile displayed a variety of proteinspecies in addition to the full-length protein, species most likelyrepresenting antibodies lacking one light chain or species consist-ing of homodimers of heavy and light chains.21 However, SECproved conclusively that the anti-HER2 IgA antibodies were
intact. The low level of fully assembled IgA2m1 and the abun-dance of light chain dimers and free light chain was probably dueto inefficient formation of disulfide bonds between heavy andlight chains.24 A similar phenomenon was also observed withIgA2m2 produced in Chinese hamster ovary (CHO) cells, whereit was associated with the presence of free cysteine residues lead-ing to gel electrophoresis artifacts.25
The role of the free thiols in the appearance of additionalbands during non-reducing SDS-PAGE was studied in moredetail. Since blocking the free thiols with NEM before gel elec-trophoresis strongly suppressed the formation of the additionalbands, a series of mutated proteins was made in which the freethiols at positions 320 and 480 were separately or togetherreplaced by serine, or in which the complete tailpiece was deleted.In secretory IgA, one of the 4 heavy chains is covalently bound tothe secretory component through a cysteine corresponding toposition 320 in the IgA2m2 mutant series.26 This position corre-sponds to position 309 in the IgA numbering used by Smith andMorrison.27 Interestingly, these authors observed that engineer-ing a leucine to cysteine mutation at the corresponding positionin an IgG3 triggered apparent polymerization of the otherwisemonomeric IgG3 antibody, which clearly illustrates the potentialreactivity of the cysteine in the Ca2 domain possibly resulting ingel artifacts. The four penultimate cysteine residues, which alsoseem to play an important role in causing gel artifacts, connectthe different monomeric IgAs in secretory IgA by binding to J-chain and probably to each other.28 Clearly, when both typesof free thiols were absent – either through mutation or dele-tion – most of the gel artifacts were gone. More importantly,it is likely that these combined mutations will also greatlyfacilitate commercial manufacture of a consistent, homoge-neous product.
Earlier publications on plasma clearance showed that humanIgA was cleared rapidly in mice with half-lives between 10 to14 hours, whereas human IgG1 half-life in mice is generallyreported to be around 10 days.29,30 Hence, we anticipated thatrepetitive IgA dosing would be essential to maintain adequateand equivalent exposure levels of IgA and IgG1. The fact thatIgA does not bind to FcRn – neither in mice nor in humans –leads to a much shorter half-life for IgA than for IgG1, particu-larly in mice. In humans, the clearance rates of IgG1 and IgA donot differ as strikingly as in mice (IgG1 half-life is »21 days andIgA half-life is »5 days in humans).31,32,33 Our preliminary PKstudies with IgA from HEK293 cells transiently expressing theantibodies confirmed the published data on poor half-life in mice
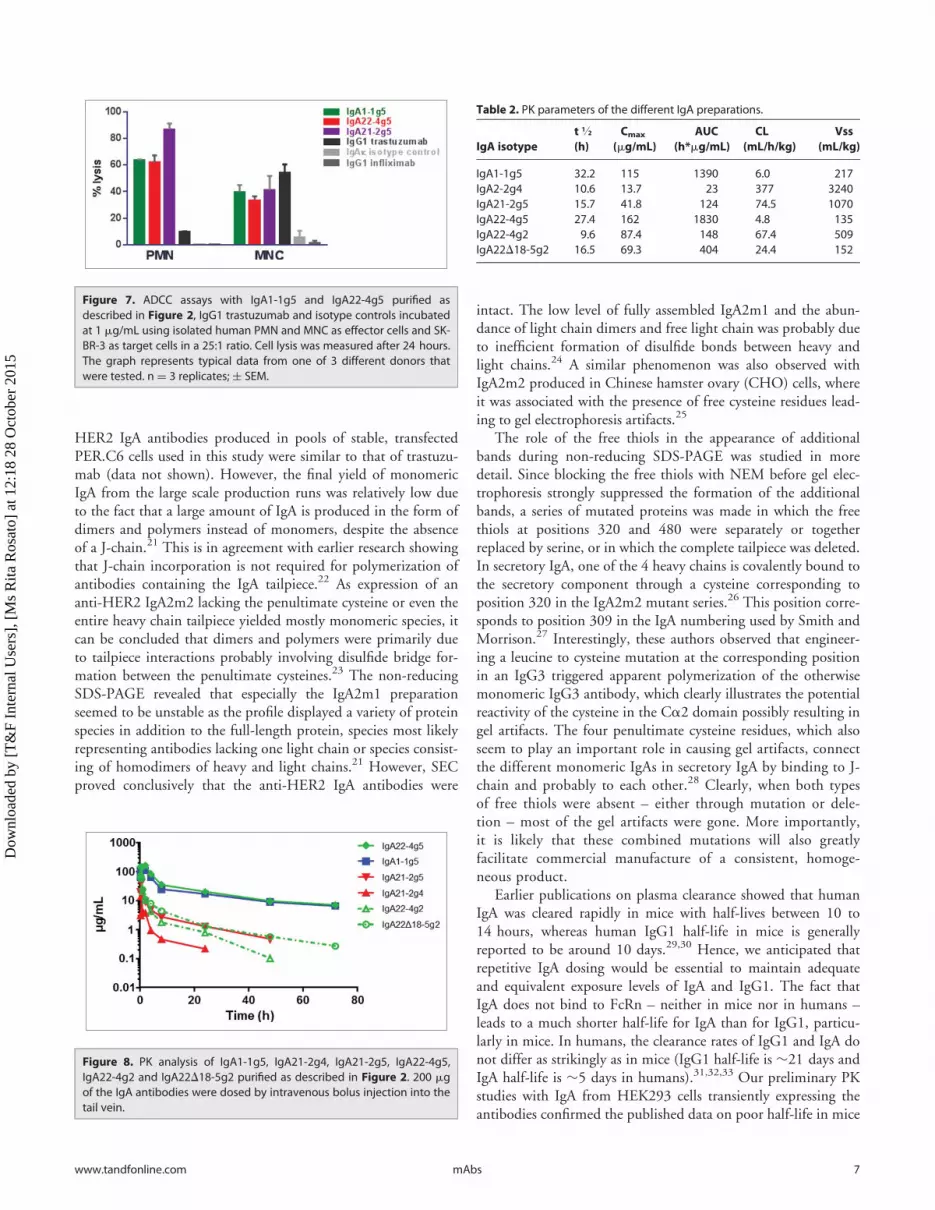
Figure 7. ADCC assays with IgA1-1g5 and IgA22-4g5 purified asdescribed in Figure 2, IgG1 trastuzumab and isotype controls incubatedat 1 mg/mL using isolated human PMN and MNC as effector cells and SK-BR-3 as target cells in a 25:1 ratio. Cell lysis was measured after 24 hours.The graph represents typical data from one of 3 different donors thatwere tested. n D 3 replicates; § SEM.
Figure 8. PK analysis of IgA1-1g5, IgA21-2g4, IgA21-2g5, IgA22-4g5,IgA22-4g2 and IgA22D18-5g2 purified as described in Figure 2. 200 mgof the IgA antibodies were dosed by intravenous bolus injection into thetail vein.
Table 2. PK parameters of the different IgA preparations.
IgA isotypet 1/2
(h)Cmax
(mg/mL)AUC
(h*mg/mL)CL
(mL/h/kg)Vss
(mL/kg)
IgA1-1g5 32.2 115 1390 6.0 217IgA2-2g4 10.6 13.7 23 377 3240IgA21-2g5 15.7 41.8 124 74.5 1070IgA22-4g5 27.4 162 1830 4.8 135IgA22-4g2 9.6 87.4 148 67.4 509IgA22D18-5g2 16.5 69.3 404 24.4 152
www.tandfonline.com 7mAbs
Dow
nloa
ded
by [
T&
F In
tern
al U
sers
], [
Ms
Rita
Ros
ato]
at 1
2:18
28
Oct
ober
201
5

and prompted the step toward glyco-engineering (data notshown).29
The earlier PK studies on human mIgA clearance in mice notonly demonstrated rapid clearance, but they also showed that itwas probably uptake by the liver asialoglycoprotein receptor(ASGPR) that was responsible for the high clearance rate.13,29,34
The involvement of the ASGPR implied that the extent of IgAglycan sialylation would control the antibodies’ fate in serum. Inhuman serum IgA1, 64% of the terminal N-linked galactose resi-dues are sialylated.18 In view of the apparent relevance of highsialylation, it was decided to produce the different mIgA types inPER.C6 cells that co-expressed a sialyltransferase gene to aug-ment capping of the exposed galactose residues on N-glycans.35
Since mouse ASGPR also seems to bind to a2,6-sialylated gly-cans in addition to ß-linked galactose and N-acetylgalactosamine,the human a2,3-sialyltransferase ST3GAL4 - and not anST6GAL1 - was selected for increasing the level of antibody sialy-lation.36,37 In contrast, in human serum IgA1 more than 95% ofsialic acids were linked in the a2,6-configuration.18
The N-glycan profiles of each of the IgA antibodies weredetermined using a novel, robust MALDI-TOF MS method fol-lowing glycan ethyl esterification, which not only avoids the usu-ally occurring loss of sialic acid residues in MALDI-TOFanalysis, but also allows determination of their linkage type.38 N-glycans from the antibodies IgA1-1g5 and IgA22-4g5 werehighly a2,3-sialylated, whereas N-glycans from antibodiesIgA21-2g4 and ¡2g5 exhibited low levels of both a2,3- anda2,6-sialylation and very high levels of Lewis A or X epitope (notdistinguished by MS). These two IgA2m1 antibodies displayedvery similar N-glycan profiles, with IgA21-2g5 displaying a mod-erately higher level of not only a2,3-sialylation, but oddlyenough, also of a2,6-linked sialic acid residues, despite beinggenerated in the presence of ectopically expressed a2,3-sialyl-transferase. Perhaps, extensive sialylation of the exposed LacNActermini by endogenous or co-expressed sialyltransferases was pre-vented by prior a1,3-fucosylation.39 N-glycan profiles of the
antibodies IgA22-4g2 andIgA22D18-5g2 produced in tran-siently transfected HEK293F cellswere remarkably different fromthose produced in stably trans-fected PER.C6 cells, includingIgA22-4g5, i.e., the same IgA2m2sequence as IgA22-4g2, but pro-duced in PER.C6 cells. The twopreparations also differed fromeach other in that IgA22D18-5g2 –lacking the tailpiece – hardly con-tained high-mannose N-glycans incontrast to IgA22-4g2. A analo-gous phenomenon was noticed byBrunke et al. when comparing N-glycans from 2 IgA2m1 antibodiesproduced in CHO-K1, one a wild-type sequence and the other amutated version in which the cyste-
ine in the tailpiece had been deleted.23 Here as well, the IgA witha tailpiece mutation contained significantly lower high-mannoseN-glycans compared to the wild-type IgA. It is quite conceivable,as suggested by these authors, that interfering with intracellulartailpiece interaction by deletion of the penultimate cysteine oreven the complete tailpiece may alter the intracellular fate of theIgA molecule. The reduction of IgA polymerization that weobserved following deletion of the tailpiece in IgA22D18-5g2might facilitate trafficking through the secretory pathway, withthe high-mannose N-glycans in IgA22-4g2 being indicative ofretention in early endoplasmic reticulum (ER) and Golgi com-partments. The sialylation levels of the IgA antibodies producedin HEK293F were as low as those of IgA21-2g4 and ¡2g5.
As anticipated, the plasma exposure levels of the IgA antibod-ies with low sialylation were much lower than of those with highsialylation. Apparently, the high rate of capping of terminalgalactose with sialic acid in IgA1-1g5 and IgA22-4g5 indeed pro-tected these antibodies from rapid clearance via the ASGPR. Thehigh incidence of LewisA/X epitopes on both IgA21-2g4 and¡2g5 may have contributed to their fast clearance through thescavenger receptor C-type lectin, for example.40,41 Likewise,high-mannose N-glycans may enhance antibody clearance.42
All IgA antibodies, except IgA1-1g5, displayed efficacy andpotency similar to trastuzumab in the proliferation inhibition assaywith BT-474 breast carcinoma cells. It is unclear why this IgAshowed only partial inhibition, especially since SPR and cellularbinding analysis revealed that it has HER2 affinity comparable tothe other IgA antibodies. In contrast, performance of all IgA anti-bodies was quite similar in the ADCC assays. These assays con-firmed that IgA could be superior to IgG1 with regard to the levelof cell lysis when using human whole blood or granulocytes aseffector cells, although this result was quite donor dependent incase of human whole blood. On the other hand, the potency oftrastuzumab is better (e.g., > 7-fold lower IC50 in a 24 hoursADCC assay using human whole blood) and would in theoryrequire lower exposure levels compared to the IgA antibodies to be
Figure 9. In vivo activity of IgA1-1g5 (A), IgA22-4g5 (B) purified as described in Figure 2 and IgG1 trastuzu-mab (A, B) in a BT-474 xenograft model in female FcaRI-transgenic SCID mice. Treatment was started at atumor volume of 200-300 mm3 using single-dose trastuzumab (upward arrow) or multiple dosing of theIgA antibodies (small downward arrows). n D 8 animals per group; § SEM.
8 Volume 0 Issue 0mAbs
Dow
nloa
ded
by [
T&
F In
tern
al U
sers
], [
Ms
Rita
Ros
ato]
at 1
2:18
28
Oct
ober
201
5

efficacious. The lower IgA potency might be compensated in vivoby reaching a higher (e.g., tenfold) IgA plasma exposure.
The question as to which mouse model is most likely to pro-vide a fair comparison of the therapeutic value of the differentmIgA isotypes is difficult to answer, especially since it concerns acomparison of human IgG1 and human IgA with identical vari-able domains that utilize different human Fc receptors. Hence,an inevitable methodological flaw arises from the fact that inter-actions of these antibodies with the effector cells in (transgenic)mice will be quite different from those in patients, because thetype, distribution and density of antibody receptors acrosshuman and mouse effector cells are quite different. An addedcomplication comes from the fact that mice lack FcaRI, whichrequires the use of FcaRI-expressing transgenic mice for testingFc-mediated IgA effects. In mice, human IgG1 binds avidly to allmouse FcgRs and is able to induce ADCC/ADCP with mouseNK cells and macrophages.43
In vivo assessment of antibody efficacy is usually performed inxenograft models in SCID mice and is thus restricted to analysesof innate mechanisms of cancer cell killing. Hence, possible anti-body-mediated elicitation or enhancement of anti-tumorhumoral and cellular responses is ignored, even though suchpotentially therapeutic responses have been observed in patientstreated with trastuzumab or cetuximab.44,45,46 As FcaRI-depen-dent internalization of IgA-immune complexes and ensuing pre-sentation has been clearly demonstrated, it is quite conceivablethat IgA could also induce adaptive anti-tumor immunity.47,48
On the other hand, Otten et al. reported that antigen presenta-tion on DC via the FcaRI is low.49 Nevertheless, if a differentialeffect of IgA versus IgG1 with regard to initiation of adaptiveimmunity does exist, our studies will not detect it.
Thus far, in vivo experiments with IgA antibodies were ham-pered by the fast clearance of IgA via the ASGPR. We have beenable to obtain IgA preparations with sufficiently high N-linkedsialylation to allow a direct comparison to an IgG1, albeit withrepetitive dosing for the IgAs vs. a single IgG1 dose to obtaincomparable exposure levels. In a BT-474 xenograft study, IgA1-1g5 displayed only limited efficacy compared to trastuzumab,but, using a similar dosing regimen, IgA22-4g5 showed impres-sive, dose-dependent tumor growth inhibition. The clear in vivoefficacy difference between IgA1-1g5 and IgA22-4g5 concurredwith the results of the in vitro proliferation inhibition assays, butnot with the ADCC data, which showed equivalent efficacy ofthe 2 antibodies with human whole blood and PMN fromFcaRI-expressing transgenic mice as effector cells. Preliminaryimmunohistochemistry analysis indicated that both IgA antibod-ies reached and penetrated the tumors at least as well as trastuzu-mab (data not shown). Thus, with the proliferation inhibitionactivity being the only tangible in vitro efficacy trait differingbetween IgA1-1g5 and IgA22-4g5, we consider that ADCC/ADCP may not be a dominant mode-of-action in this in vivomodel, implying that the superior therapeutic effect of IgA22-4g5 might be purely Fab-mediated. Then again, it has beenshown unequivocally that (relatively high-dose) trastuzumabtreatment in another BT-474 model in athymic nude BALB/cmice requires FcgR functions of effector cells.2 Furthermore, in
the A431 tumor model, efficacy is only observed in FcaRI-expressing transgenic mice. Here as well, the efficacy of a singledose trastuzumab was higher than that of multiple doses ofIgA22-4g5. We conclude that the in vivo tumor cell killingcapacity of IgA antibodies appears to be much less potent thanthat of an equivalent IgG1 in the subcutaneous BT-474 modelused in this study. Consequently, even though the manufactureof high quality IgA antibodies may be perfectly feasible, onewould need to identify tumor types, combination therapies orpatient populations that would merit taking an IgA-basedapproach instead of one using IgG1. As reported recently, hybridIgG1/IgA antibodies might be a promising way to create anti-tumor molecules efficiently employing a broad repertoire ofeffector cells with adequate plasma exposure.50
Materials and Methods
Cell LinesHuman breast carcinoma cell lines SK-BR-3 and BT-474
were obtained from American Type Culture Collection (Rock-ville, MD). SK-BR-3 were cultured in McCoy’s 5A medium(Lonza, BE12-688F ) supplemented with 10% v/w fetal bovineserum (FBS; Invitrogen, 10100-147), Heat-inactivated (HI), andBT-474 cells in RPMI-1640 (Lonza, BE12-702F) containing2 mM L-glutamine (Lonza, BE17-605E) supplemented with10% FBS (Invitrogen, 10100-147), at 37�C in a humidifiedincubator containing 5% CO2.
IgA production and purificationThe heavy and light chain sequences for the antibodies and
the human a2,3-sialyltransferase (ST3GAL4) gene were synthe-sized by GeneArt. Stable PER.C6 pools producing IgA weremade by transfecting 4 mg of pcDNA-based vectors containingthe variable regions of trastuzumab combined with the constantdomains of the human IgA1, IgA2m1 and IgA2m2 heavy chainsand the human kappa light chain. In three out of 4 transfec-tions, PER.C6 cells were co-transfected with 4 mg of apcDNA3.1-Hyg-based vector (Invitrogen, V870-20) containingthe ST3GAL4 gene. Stable PER.C6 pools were cultured inCDM4PERMAb growth medium (Hyclone, SH30871.02) with3 mM L-glutamine (Lonza, BE17-605F), 0.05% Pluronic F68(Sigma-Aldrich, P5556), 62.5 mg/mL G418 (Life Technologies,10131-027), and 6.25 mg/mL hygromycin B (Life Technolo-gies, 10687-010). The four stable pools were expanded to a 10L working volume in CDM4PERMab without selection pres-sure in a 20 L culture bag on an orbital shaker and a batch runof 7 days was performed. Product titers were determined usinga human IgA AlphaLISA kit (PerkinElmer, AL262 C/F) accord-ing to the manufacturer’s instructions. The batch cultures wereharvested and filtered and the antibodies were purified by affin-ity chromatography using KappaSelect chromatography resin(GE Healthcare, 17-5458-01). The bound product was elutedwith 0.1 M glycine (Sigma-Aldrich, 15527) pH 2.5 followed byimmediate neutralization to pH 7 using 1M Tris buffer(Sigma-Aldrich, T1378). As a second purification step, SEC
www.tandfonline.com 9mAbs
Dow
nloa
ded
by [
T&
F In
tern
al U
sers
], [
Ms
Rita
Ros
ato]
at 1
2:18
28
Oct
ober
201
5

was performed using a HiPrepTM 26/60 SephacrylTM S-300HR column (GE Healthcare, 17-1196-01) equilibrated withphosphate-buffered saline (PBS) pH 7.4 (Sigma-Aldrich,P5368). In each run, 10 to 15 mL partially purified IgA (Kap-paSelect eluate, pH adjusted) was loaded to the column usingPBS pH 7.4 as running buffer. Fractions were collected in 96deep-well plates. To prevent cross contamination, the SEC col-umn was cleaned with 2 column volumes of 0.2 M NaOH inbetween each run followed by re-equilibrated with PBS, pH7.4. The preparations were analyzed for aggregates, dimers andmonomers by means of SDS-PAGE and UPLC-SEC analysisusing ACQUITY UPLC BEH200 SEC 1.7 mm (Waters,186005225). Protein concentrations were determined usingA280 absorbance readings obtained with a Nanodrop 1000spectrophotometer. In addition, antibodies were produced usingtransient transfection of HEK293F cells, using similar IgAexpression vectors as described above. Mutations were intro-duced by site-specific mutagenesis. Transient transfections wereconducted using the FreeStyle 293 expression system (LifeTechnologies) according to the manufacturer’s instructions.
SDS-PAGENon-reducing SDS-PAGE was performed by mixing 25 mL
sample with 25 mL Laemmli sample buffer (BioRad, 161-0737)and subsequent loading on 4-20% polyacrylamide gel (CriterionTGX Stain free, Biorad, 567-8094). Protein bands were visual-ized by means of UV using a GelDoc EZ stainfree imager (Bio-Rad) followed by Krypton staining (Thermo Scientific, 46630)according to the manufacturer’s instructions. In some samples,N-ethylmaleimide (Sigma Aldrich, E3876) was added to the mix-ture in a final concentration of 28 mM prior to addition of sam-ple buffer.
N-glycan analysisN-glycans of purified IgAs were analyzed by MALDI-TOF
MS as described previously, with some minor modifications.37
Briefly, 100 mL 2% SDS (Merck, 8170341000) was added to 50mL of IgA1-1g5 (1.12 mg/mL), IgA2m1-2g4 (0.96 mg/mL),IgA2m1-2g5 (1.01 mg/mL), IgA2m2-4g5 (0.98 mg/mL),IgA2m2-4g2 (1.20 mg/mL) and IgA2m2D18-5g2 (1.03 mg/mL), which was then denatured 10 min at 60�C. N-glycanrelease was performed by adding 100 mL release mix (containing2% NP-40 substitute (Sigma-Aldrich, 74385) and 2.5 mUPNGase F (Roche Diagnostics, 11365177001) in 2.5x PBS(Merck)) and incubating 16 h at 37�C. Then, in quadruplicate,the released glycans were ethyl esterified at the a2,6-linked sialicacids, and lactonized at the a2,3-linked sialic acids, by adding 5mL released sample to 35 mL 0.25 M 1-ethyl-3-(3-dimethylami-nopropyl)carbodiimide hydrochloride (Fluorochem, 024810) and0.25 M 1-hydroxybenzotriazole (Sigma-Aldrich, 54802) in etha-nol (Merck, 1009831000), and incubating 1 h at 37�C. Thereaction was stopped by adding 40 mL acetonitrile (ACN)(HPLC Supra gradient, 012035) and incubating 15 min at¡20�C. The derivatized glycans were extracted from the mixtureby hydrophilic interaction liquid chromatography using cotton asstationary phase, and eluted in a 10 mL volume of deionized
water.51 Two mL of the eluted samples was transferred to a MTPAnchorChip 800/384 TF MALDI target (Bruker Daltonics,209514), and mixed on plate with 1 mL 5 mg/mL 2,5-dihydrox-ybenzoic acid (Bruker Daltonics, 201346) 1 mM NaOH(Sigma-Aldrich, 71686) in 50% ACN. After letting the spots dryby air, recrystallization was performed by addition of 0.2 mLethanol.
MALDI-TOF MS measurements were taken using reflectronpositive mode of an UltraFlextreme with Smartbeam-II laser(Bruker Daltonics), controlled by Flexcontrol 3.4 Build 119(Bruker Daltonics). After 140 ns delayed extraction, accelerationwas performed at 25 kV. Laser power was set as high as possibleto still allowing baseline separation of isotopic peaks. Using thesesettings, the instrument was calibrated using a peptide calibrationstandard (Bruker Daltonics, 206195). Sample spectra wereacquired by summing 25000 laser shots at a frequency of2000 Hz, using a window from m/z 1000 to 5000 with suppres-sion up to m/z 950. As shooting pattern, complete sample ran-dom walk was used with 200 shots per raster spot. To confirmthe structures of the most abundant peaks, tandem mass spec-trometry (MALDI-TOF/TOF MS/MS) was performed usinglaser-induced disassociation.
Obtained spectra were processed in flexAnalysis 3.3 build 65(Bruker Daltonics), by first smoothing by Savitzky-Golay filter(width 0.2, cycles 1), followed by a Top-hat filter for baselinesubtraction. Peaks were picked by Snap algorithm with a mini-mum signal-to-noise (S/N) threshold of 6, after which the spec-trum was internally calibrated. Peak lists were exported with S/N,intensity and area, and further analysis was performed in Excel.
Data from 4 independent ethyl esterification procedureswere compared per sample and peaks were selected for furtheranalysis if occurring in all spectra above a S/N threshold of 6.Then, based on the average m/z values, the most likely N-gly-can compositions were assigned. Signals were excluded fromfurther analysis if they could not be assigned to N-glycancompositions within § 0.05 Da. For the remaining glycancompositions, the areas were calculated relative to the sum ofall areas within a spectrum, and these values were also aver-aged for the 4 independent measurements. These average rel-ative areas were used for further calculation.
Percentages of fucosylation, sialylation, a2,3-linked sialyla-tion, a2,6-linked sialylation and high mannose type glycans werecalculated for a sample by summing the relative ratios of compo-sitions meeting the required criteria. Fucosylation, a2,3-linkedsialylation and a2,6-linked sialylation were summed if anyrespective number of fucoses, lactonized or ethyl esterified sialicacids were found, while sialylation was summed if any type ofsialic acid was found. High mannose was summed if the amountof found hexoses was higher than 5, while the amount of N-ace-tylhexosamines did not rise above 2. The average amount of fuco-ses, sialic acids or mannoses in the fucosylated, sialylated andhigh mannose type glycan fractions could be calculated by multi-plying the relative areas by the number of relevant monosacchar-ides found.
Figures were assigned with structures following the CFG nota-tion, made in GlycoWorkbench 2.1 build 146.52
10 Volume 0 Issue 0mAbs
Dow
nloa
ded
by [
T&
F In
tern
al U
sers
], [
Ms
Rita
Ros
ato]
at 1
2:18
28
Oct
ober
201
5

Proliferation inhibitionCells in complete growth medium were plated in 96-well
plates (90 mL/well) and incubated overnight at 37�C, 5% CO2
at the following cell densities of 6,500 SK-BR-3 and 10,000 BT-474 cells. Serial dilutions of each antibody were made in culturemedium. Human IgA1 kappa and IgA2 kappa isotype controlantibodies were obtained from Meridian Bioscience, Inc..(A50555H and A50166H), human IgA kappa isotype controlantibody from SouthernBiotech (0155k-01), and human IgG1Remicade� (infliximab) from Johnson & Johnson. 10 mL ofeach antibody concentration was added to the 96-well plate. Cellviability was assessed after 6 days using the CellTiter-GloTM
(CTG) luminescent assay kit from Promega Corporation(G7572) according to the manufacturer’s instructions. Survivalpercentage was calculated by dividing the measured luminescencefor each mAb concentration with the average mean of the controlwells (no antibody added, only growth medium) multiplied by100.
Antibody-dependent cell-mediated cytotoxicity assayADCC was measured by lactate dehydrogenase (LDH) release
of SK-BR-3 and BT-474 breast carcinoma cells as target cells.Serial dilutions of each antibody were made in complete growthmedium. 50 mL of each antibody solution was incubated for 30minutes at 37�C, 5% CO2 with 50 mL of SK-BR-3 (10,000cells/well) or BT-474 cells (15,000 cells/well) in 96-well round-bottom plates.
Human whole blood from healthy donors was used toisolate monocytes (MNC) and PMN using Density GradientCentrifugation with Histopaque�-1119 (Sigma-Aldrich,11191) and Lymphocyte Separation Medium (Lonza, 17-829F) according to the manufacturer’s instruction. 50 mL of2.5 £ 105 PMN or MNC were added to 104 of the antibody-treated SK-BR-3 or BT-474 cells at an effector-to-target ratioof 25:1. Unseparated human whole blood from healthy donorswas exposed to hypotonic lysis to remove remaining erythro-cytes before 50 mL of human whole blood was added to theantibody-treated target cells. After 4 hours of incubation at37�C, 5% CO2, cell supernatant was transferred to a 96-wellplate to determine the amount of released LDH using a colori-metric assay (Roche, 11644793001). Percent cytotoxicity wascalculated as follows: percent cytotoxicity D (“experimental” –“effector plus target spontaneous”) / (“target maximum” –“target spontaneous”) x 100%, where “experimental” corre-sponds to the signal measured in a treated sample, “effectorplus target spontaneous” corresponds to the signal measuredin the presence of PMN or MNC and tumor cells alone,“target maximum” corresponds to the signal measured in thepresence of detergent lysed tumor cells (Triton X-100, SigmaAldrich, X100), and “target spontaneous” corresponds to thesignal measured in the presence of tumor cells alone.
MiceHuman FcaRI transgenic mice were generated at the UMC
Utrecht, and were backcrossed to the immunodeficient SCIDbackground (CB17/lcr�Prkdcscid/ lcrlcoCrl, Charles River) and
maintained as hemizygotes.14 Transgene-negative littermates,WT BALB/c or C57BL/6 mice (Janvier) were used as controls.All animal studies were approved by local animal ethical commit-tees and performed according to local ethical guidelines of animalexperimentation.
In vivo pharmacokineticsThe IgA antibodies were dosed to 7 weeks old female mice
(CB-17/lcr-Prkdc SCID) via an intravenous bolus injection intothe tail vein. Blood samples were taken via terminal intracardiacpuncture under general anesthesia (isoflurane-O2 mixture) in dif-ferent mice at several time points after dosing and processed toK2-EDTA plasma. Plasma samples were stored at ¡80�C untilbioanalysis. Based on the reported plasma levels, PK parameterswere calculated in WinNonlin version 5.3 using the non-com-partmental analysis for single intravenous bolus injection.
PK ELISAAn ELISA-based method was used for the determination of
trastuzumab and anti-HER2 IgA antibody plasma concentra-tions. Study samples were diluted in appropriate buffers in orderto be able to estimate the antibody concentrations within the ana-lytical range. The solid phase consisted of an anti-idiotypic miniantibody (AbD Serotec, AbD15916) coated on high bindingmicro titer plates (Greiner, 655081). Detection of antibody cap-tured on the solid phase was achieved by a biotinylated anti-idio-type mini antibody, followed by a streptavidin-HRP (R&DSystems, DY998) incubation step and TMB (TeBu-Bio,TMB100) incubation. The color reaction was stopped withH2SO4 and the plate was read at 450 and 630 nm. Each analyti-cal run included appropriate calibrators and quality-control sam-ples. Antibody concentrations of study samples were backcalculated on the calibrator curve.
BT-474 cell-line xenograft studiesBT-474 cell-line xenograft studies were conducted at Oncode-
sign, Dijon, France. Female FcaRI-transgenic SCID mice werehoused in polycarbonate cages (Tecniplast) that are equipped toprovide food and water and contain environmental enrichmentin a specific-pathogen-free (SPF) animal care unit.14 Animalfood (DIETEX) and water are provided ad-libitum. BT-474tumors were induced in mice at the age of 35–46 weeks by a sub-cutaneous injection of 2 £ 107 BT-474 cells in 200 mL of RPMI1640 containing Matrigel (50:50, v:v; BD Biosciences, 356237)into the right flank. Mice were randomized according to theirindividual tumor volume into groups of 8 animals. At start oftreatment, the mean tumor volume (in between 200-300 mm3)of each group did not differ from the others (analysis of variance).Treatment was by a single intravenous bolus injection in the tailvein (trastuzumab) or by intraperitoneal injection 10 times oncedaily (IgA mAbs and vehicle). Individual animals were sacrificedwhen tumor sizes were 2000 mm3 or at the end of the study (typ-ically between day 70-90).
www.tandfonline.com 11mAbs
Dow
nloa
ded
by [
T&
F In
tern
al U
sers
], [
Ms
Rita
Ros
ato]
at 1
2:18
28
Oct
ober
201
5

Disclosure of potential conflicts of interest
GJA Rouwendal, MM van der Lee, J Schouten, G de Roo, DFEgging, GF Verheijden, R Ubink, WH Dokter, and M Timmersare Synthon Biopharmaceuticals B.V. employees. JHW Leusenand MWuhrer received research funding from Synthon Biophar-maceuticals B.V.
Acknowledgments
We gratefully acknowledge technical assistance from Karin deLaat, Jochem Eigenhuijsen, Jos Verhagen, Dirk Glaudemans,
Erin Hendriks-Franssen Wever, Meng Liu, Kim Burgers,Dani€elle Jacobs, Tanja van Achterberg, and Ellen Santegoeds-Lenssen from Synthon Biopharmaceuticals B.V. and MaaikeNederend and Marco Jansen from Laboratory for TranslationalImmunology (UMC Utrecht).
Supplemental material
Supplemental data for this article can be accessed on thepublisher’s website.
References
1. Clynes R, Takechi Y, Moroi Y, Houghton A, RavetchJV. Fc receptors are required in passive and activeimmunity to melanoma. Proc Natl Acad Sci U S A1998; 95:652-6; PMID:9435247; http://dx.doi.org/10.1073/pnas.95.2.652
2. Clynes RA, Towers TL, Presta LG, Ravetch JV. Inhibi-tory Fc receptors modulate in vivo cytotoxicity againsttumor targets. Nat Med 2000; 6:443-6;PMID:10742152; http://dx.doi.org/10.1038/74704
3. Nimmerjahn F, Ravetch JV. Translating basic mecha-nisms of IgG effector activity into next generation can-cer therapies. Cancer Immun 2012; 12:13;PMID:22896758
4. Valerius T, Stockmeyer B, van Spriel AB, Graziano RF,van den Herik-Oudijk IE, Repp R, Deo YM, Lund J,Kalden JR, Gramatzki M, et al. FcaRI (CD89) as anovel trigger molecule for bispecific antibody therapy.Blood 1997; 90:4485-92; PMID:9373259
5. Huls G, Heijnen IAFM, Cuomo E, van der Linden J,Boel E, van de Winkel JGJ, Logtenberg T. Antitumorimmune effector mechanisms recruited by phage dis-play-derived fully human IgG1 and IgA1 monoclonalantibodies. Cancer Res 1999; 59:5778-84;PMID:10582699
6. Stockmeyer B, Dechant M, van Egmond M, Tutt AL,Sundarapandiyan K, Graziano RF, Repp R, Kalden JR,Gramatzki M, Glennie MJ, et al. Triggering Fca-receptor I (CD89) recruits neutrophils as effector cellsfor CD20-directed antibody therapy. J Immunol 2000;16:5954-61; http://dx.doi.org/10.4049/jimmunol.165.10.5954
7. Braster R, O’Toole T, van Egmond M. Myeloid cells aseffector cells for monoclonal antibody therapy of can-cer. Methods 2014; 65:28-37; PMID:23811299;http://dx.doi.org/10.1016/j.ymeth.2013.06.020
8. Otten MA, Bakema JE, Tuk CW, Glennie MJ, TuttAL, Beelen RH, van de Winkel JGJ, van Egmond M.Enhanced FcaRI-mediated neutrophil migrationtowards tumour colonies in the presence of endothelialcells. Eur J Immunol 2012; 42:1815-21;PMID:22535639; http://dx.doi.org/10.1002/eji.201141982
9. Dechant M, Beyer T, Schneider-Merck T, Weisner W,Peipp M, van de Winkel JGJ, Valerius T. Effectormechanisms of recombinant IgA antibodies against epi-dermal growth factor receptor. J Immunol 2007;179:2936-43; PMID:17709508; http://dx.doi.org/10.4049/jimmunol.179.5.2936
10. Cartron G, Dacheux L, Salles G, Solal-Celigny P, Bar-dos P, Colombat P, Watier H. Therapeutic activity ofhumanized anti-CD20 monoclonal antibody and poly-morphism in IgG Fc receptor FcgammaRIIIa gene.Blood 2002; 99:754-8; PMID:11806974; http://dx.doi.org/10.1182/blood.V99.3.754
11. Weng WK, Levy R. Two immunoglobulin G fragmentC receptor polymorphisms independently predictresponse to rituximab in patients with follicular lym-phoma. J Clin Oncol 2003; 21:3940-47;
PMID:12975461; http://dx.doi.org/10.1200/JCO.2003.05.013
12. Musolino A, Naldi N, Bortesi B, Pezzuolo D, Capel-letti M, Missale G, Laccabue D, Zerbini A, Camisa R,Bisagni G, et al. Immunoglobulin G fragment C recep-tor polymorphisms and clinical efficacy of trastuzu-mab-based therapy in patients with HER-2/neu-positive metastatic breast cancer. J Clin Oncol 2008;26:1789-96; PMID:18347005; http://dx.doi.org/10.1200/JCO.2007.14.8957
13. Boross P, Lohse S, Nederend M, Jansen JH, van Teter-ing G, Dechant M, Peipp M, Royle L, Liew LP, BoonL, et al. IgA EGFR antibodies mediate tumour killingin vivo. EMBO Mol Med 2013; 5:1213-26;PMID:23918228; http://dx.doi.org/10.1002/emmm.201201929
14. van Egmond M, van Vuuren AJH, van de Winkel JGJ.The human Fc receptor for IgA (FcaRI, CD89) ontransgenic peritoneal macrophages triggers phagocytosisand tumor cell lysis. Immunol Lett 1999; 68:83-7;PMID:10397160; http://dx.doi.org/10.1016/S0165-2478(99)00034-6
15. Launay P, Grosset̂ete B, Arcos-Fajardo M, Gaudin E,Torres SP, Beaudoin L, Patey-Mariaud de Serre N,Lehuen A, Monteiro RC. Fcalpha receptor (CD89)mediates the development of immunoglobulin A (IgA)nephropathy (Berger’s disease). Evidence for patho-genic soluble receptor-Iga complexes in patients andCD89 transgenic mice. J Exp Med 2000; 191:1999-2009; PMID:10839814; http://dx.doi.org/10.1084/jem.191.11.1999
16. Soua Z, Ghanem N, Ben Salem M, Lefranc G, LefrancMP. Frequencies of the human immunoglobulinIGHA2*M1 and IGHA2*M2 alleles corresponding tothe A2m(1) and A2m(2) allotypes in the French, Leba-nese, Tunisian and black African populations. NucleicAcids Res 1989; 17:3625; PMID:2566982; http://dx.doi.org/10.1093/nar/17.9.3625
17. Tora~no A, Tsuzukida Y, Liu YS, Putnam FW. Locationand structural significance of the oligosaccharides inhuman IgA1 and IgA2 immunoglobulins. Proc NatlAcad Sci U S A 1977; 74:2301-5; http://dx.doi.org/10.1073/pnas.74.6.2301
18. Mattu TS, Pleass RJ, Willis AC, Kilian M, WormaldMR, Lellouch AC, Rudd PM, Woof JM, Dwek RA.The glycosylation and structure of human serum IgA1,Fab, and Fc regions and the role of N-Glycosylation onFca receptor interactions. J Biol Chem 1998;273:2260-72; PMID:9442070; http://dx.doi.org/10.1074/jbc.273.4.2260
19. Tsuzukida Y, Wang CC, Putnam FW. Structure of theA2m(1) allotype of human IgA - a recombinant mole-cule. Proc Natl Acad Sci U S A 1979; 76:1104-8;PMID:286295; http://dx.doi.org/10.1073/pnas.76.3.1104
20. Zhang N, Liu L, Dumitru CD, Cummings NR, CukanM, Jiang Y, Li Y, Li F, Mitchell T, Mallem MR, et al.Glycoengineered Pichia produced anti-HER2 is com-parable to trastuzumab in preclinical study. mAbs
2011; 3:289-98; PMID:21487242; http://dx.doi.org/10.4161/mabs.3.3.15532
21. Beyer T, Lohse S, Berger S, Peipp M, Valerius T,Dechant M. Serum-free production and purification ofchimeric IgA antibodies. J Immunol Methods 2009;346:26-37; PMID:19427867; http://dx.doi.org/10.1016/j.jim.2009.05.002
22. Yoo EM, Coloma MJ, Trinh KR, Nguyen TQ, VuongLU, Morrison SL, Chintalacharuvu KR. Structuralrequirements for polymeric immunoglobulin assemblyand association with J chain. J Biol Chem 1999;274:33771-7; PMID:10559270; http://dx.doi.org/10.1074/jbc.274.47.33771
23. Brunke C, Lohse S, Derer S, Peipp M, Boross P, Kell-ner C, Beyer T, Dechant M, Royle L, Liew LP, et al.Effect of a tail piece cysteine deletion on biochemicaland functional properties of an epidermal growth factorreceptor-directed IgA2 m(1) antibody. MAbs 2013;5:936-45; PMID:24492345; http://dx.doi.org/10.4161/mabs.26396
24. Chintalacharuvu KR, Yu LJ, Bhola N, Kobayashi K,Fernandez CZ, Morrison SL. Cysteine residuesrequired for the attachment of the light chain inhuman IgA2. J Immunol 2002; 169:5072-7;PMID:12391223; http://dx.doi.org/10.4049/jimmunol.169.9.5072
25. Chintalacharuvu KR, Gurbaxani B, Morrison SL.Incomplete assembly of IgA2m(2) in Chinese hamsterovary cells. Mol Immunol 2007; 44:3445-52;PMID:17467056; http://dx.doi.org/10.1016/j.molimm.2006.12.030
26. Fallgren-Gebauer E, Gebauer W, Bastian A, KratzinHD, Eiffert H, Zimmermann B, Karas M, HilschmannN. The covalent linkage of secretory component toIgA. Structure of sIgA. Biol Chem Hoppe Seyler 1993;374:1023-8; http://dx.doi.org/10.1515/bchm3.1993.374.7-12.1023
27. Smith RI, Morrison SL. Recombinant polymeric IgG:an approach to engineering more potent antibodies.Biotechnology (N Y) 1994; 12:683-8;PMID:7764912; http://dx.doi.org/10.1038/nbt0794-683
28. Sørensen V, Rasmussen IB, Sundvold V, MichaelsenTE, Sandlie I. Structural requirements for incorpo-ration of J chain into human IgM and IgA. Int Immu-nol 2000; 12:19-27; PMID:10607746; http://dx.doi.org/10.1093/intimm/12.1.19
29. Rifai A, Fadden K, Morrison SL, Chintalacharuvu KR.The N-glycans determine the differential blood clear-ance and hepatic uptake of human immunoglobulin(Ig)a1 and Iga2 isotypes. J Exp Med 2000; 191:2171-81; PMID:10859341; http://dx.doi.org/10.1084/jem.191.12.2171
30. Petkova SB, Akilesh S, Sproule TJ, Christianson GJ, AlKhabbaz H, Brown AC, Presta LG, Meng YG, Roope-nian DC. Enhanced half-life of genetically engineeredhuman IgG1 antibodies in a humanized FcRn mousemodel: potential application in humorally mediatedautoimmune disease. Int Immunol 2006; 18:1759-69;
12 Volume 0 Issue 0mAbs
Dow
nloa
ded
by [
T&
F In
tern
al U
sers
], [
Ms
Rita
Ros
ato]
at 1
2:18
28
Oct
ober
201
5

PMID:17077181; http://dx.doi.org/10.1093/intimm/dxl110
31. Morell A, Terry WD, Waldmann TA. Metabolic prop-erties of IgG subclasses in man. J Clin Invest 1970;49:673-80; PMID:5443170; http://dx.doi.org/10.1172/JCI106279
32. Blaese RM, Strober W, Levy AL, Waldmann TA.Hypercatabolism of IgG, IgA, IgM, and albumin in theWiskott-Aldrich syndrome. A unique disorder of serumprotein metabolism. J Clin Invest 1971; 50:2331-8;PMID:5096517; http://dx.doi.org/10.1172/JCI106731
33. Delacroix DL, Elkom KB, Geubel AP, Hodgson HF,Dive C, Vaerman JP. Changes in size, subclass, andmetabolic properties of serum immunoglobulin A inliver diseases and in other diseases with high serumimmunoglobulin A. J Clin Invest 1983; 71:358-67;PMID:6401770; http://dx.doi.org/10.1172/JCI110777
34. Moldoveanu Z, Epps JM, Thorpe SR, Mestecky J. Thesites of catabolism of murine monomeric IgA. J Immu-nol 1988; 141:208-13; PMID:2454258
35. Weikert S, Papac D, Briggs J, Cowfer D, Tom S,Gawlitzek M, Lofgren J, Mehta S, Chisholm V,Modi N, et al. Engineering Chinese hamster ovarycells to maximize sialic acid content of recombi-nant glycoproteins. Nat Biotechnol 1999; 17:1116-21; PMID:10545921; http://dx.doi.org/10.1038/15104
36. Park EI, Mi Y, Unverzagt C, Gabius HJ, Baenziger JU.The asialoglycoprotein receptor clears glycoconjugatesterminating with sialic acid a 2,6GalNAc. Proc NatlAcad Sci U S A 2005; 102:17125-9PMID:16286643;http://dx.doi.org/10.1073/pnas.0508537102
37. Steirer LM, Park EI, Townsend RR, Baenziger JU. Theasialoglycoprotein receptor regulates levels of plasmaglycoproteins terminating with sialic acid a2,6-galac-tose. J Biol Chem 2009; 284:3777-83;PMID:19075021; http://dx.doi.org/10.1074/jbc.M808689200
38. Reiding KR, Blank D, Kuijper DM, Deelder AM,Wuhrer M. High-throughput profiling of protein
N-glycosylation by MALDI-TOF-MS employing link-age-specific sialic acid esterification. Anal Chem 2014;86:5784-93; PMID:24831253; http://dx.doi.org/10.1021/ac500335t
39. Kitagawa H, Paulson JC. Cloning of a novel a 2,3-sia-lyltransferase that sialylates glycoprotein and glycolipidcarbohydrate groups. J Biol Chem 1994; 269:1394-1401; PMID:8288606
40. Coombs PJ, Graham SA, Drickamer K, Taylor ME.Selective binding of the scavenger receptor C-type lec-tin to Lewisx trisaccharide and related glycan ligands. JBiol Chem 2005; 280:22993-9; PMID:15845541;http://dx.doi.org/10.1074/jbc.M504197200
41. Graham SA, Antonopoulos A, Hitchen PG, HaslamSM, Dell A, Drickamer K, Taylor ME. Identificationof neutrophil granule glycoproteins as LewisX-contain-ing ligands cleared by the scavenger receptor C-type lec-tin. J Biol Chem 2011; 286:24336-49;PMID:21561871; http://dx.doi.org/10.1074/jbc.M111.244772
42. Goetze AM, Liu YD, Zhang Z, Shah B, Lee E, Bondar-enko PV, Flynn GC. High-mannose glycans on the Fcregion of therapeutic IgG antibodies increase serumclearance in humans. Glycobiology 2011; 21:949-59;PMID:21421994; http://dx.doi.org/10.1093/glycob/cwr027
43. Overdijk MB, Verploegen S, Ortiz Buijsse A, Vink T,Leusen JH, Bleeker WK, Parren PW. Crosstalkbetween human IgG isotypes and murine effector cells.J Immunol 2012; 189:3430-8; PMID:22956577;http://dx.doi.org/10.4049/jimmunol.1200356
44. Disis ML, Calenoff E, McLaughlin G, Murphy AE,Chen W, Groner B, Jeschke M, Lydon N, McGlynn E,Livingston RB, et al. Existent T-cell and antibodyimmunity to HER2/neu protein in patients with breastcancer. Cancer Res 1994; 54:16-20; PMID:7505195
45. Taylor C, Hershman D, Shah N, Suciu-Foca N, Petry-lak DP, Taub R, Vahdat L, Cheng B, Pegram M, Knut-son KL, et al. Augmented HER-2 specific immunityduring treatment with trastuzumab and chemotherapy.Clin Cancer Res 2007; 13:5133-43; PMID:17785568;http://dx.doi.org/10.1158/1078-0432.CCR-07-0507
46. Srivastava RM, Lee SC, Andrade Filho PA, Lord CA,Jie HB, Davidson HC, L�opez-Albaitero A, Gibson SP,Gooding WE, Ferrone S, et al. Cetuximab-activatednatural killer and dendritic cells collaborate to triggertumor antigen-specific T-cell immunity in head andneck cancer patients. Clin Cancer Res 2013; 19:1858-72; PMID:23444227; http://dx.doi.org/10.1158/1078-0432.CCR-12-2426
47. Shen L, van Egmond M, Siemasko K, Gao H, Wade T,Lang ML, Clark M, van De Winkel JGJ, Wade WF.Presentation of ovalbumin internalized via the immu-noglobulin-A Fc receptor is enhanced through Fcreceptor g-chain signaling. Blood 2001; 97:205-13;PMID:11133762; http://dx.doi.org/10.1182/blood.V97.1.205
48. Lang ML, Shen L, Gao H, Cusack WF, Lang GA,Wade WF. Fca receptor cross-linking causes transloca-tion of phosphatidylinositol-dependent protein kinase1 and protein kinase Ba to MHC Class II peptide-loading-like compartments. J Immunol 2001;166:5585-93; PMID:11313398; http://dx.doi.org/10.4049/jimmunol.166.9.5585
49. Otten MA, Groenveld I, van de Winkel JGJ, vanEgmond M. Inefficient antigen presentation via theIgA Fc receptor (FcaRI) on dendritic cells. Immunobi-ology 2006; 211:503-10; PMID:16920489; http://dx.doi.org/10.1016/j.imbio.2006.05.016
50. Borrok MJ, Luheshi NM, Beyaz N, Davies GC, LeggJW, Wu H, Dall’Acqua WF, Tsui P. Enhancement ofantibody-dependent cell-mediated cytotoxicity byendowing IgG with FcaRI (CD89) binding. MAbs2015; 7(4):743-51
51. Selman MHJ, Hemayatkar M, Deelder AM, WuhrerM. Cotton HILIC SPE microtips for microscale purifi-cation and enrichment of glycans and glycopeptides.Anal Chem 2011; 83:2492-9; PMID:21366235;http://dx.doi.org/10.1021/ac1027116
52. Ceroni A, Maass K, Geyer H, Geyer R, Dell A, HaslamSM. GlycoWorkbench: a tool for the computer-assistedannotation of mass spectra of glycans. J Proteome Res2008; 7:1650-9; PMID:18311910; http://dx.doi.org/10.1021/pr7008252
www.tandfonline.com 13mAbs
Dow
nloa
ded
by [
T&
F In
tern
al U
sers
], [
Ms
Rita
Ros
ato]
at 1
2:18
28
Oct
ober
201
5





























