Embed Size (px)
Citation preview

Aquaculture, 67 (1987) 301-311
Elsevier Science Publishers B.V., Amsterdam - Printed in The Netherlands 301
A Comparison of Survival and Growth of Two Strains of Chinook Salmon (Oncorhynchus tshatoytscha) and Their Crosses Reared in Confinement
KIMBERLY M. CHENG, IAN M. McCALLUM’, ROBERT I. MCKAY and B.E. MARCH’
Department of Animal Science, University o/ British Columbia, Vancouver, B.C. V6T W2 (Canada)
‘Present address: Sea Star Resources, 1066 W. Hasting, Vancouver, B.C. (Canada) ‘To whom correspondence should be addressed.
(Accepted 22 June 1987)
ABSTRACT
Cheng. K.M.. McCallum. I.M.. McKay, R-1. and March, B.E.. 1987. A comparison of survival and growth of two strains of chinook salmon (Oncorhynchus tshawytscha) and their crosses reared in confinement. Aquaculture. 67: 301-311.
The growth rate and survival of the Capilano ( hatchery) and Harrison (wild 1 strains of chi- nook salmon (0. tshawytscha) of British Columbia and their hybrids reared in confinement were compared both during the fresh-water phase and the sea-water phase. Eggs stripped from the Capilano females had better fertility, fingerlings of the Capilano strain were heuvier at 25 weeks of age and had a higher average specific growth rate. The Capilano fish were also heavier both in live weight and gutted weight after 1.5 years in sea-pens. No heterosis was detected for any of the traits measured. Cross-means for body weight and specific growth rate tended to be similar to the
strain-of-sire mean.
INTRODUCTION
Large-scale farming of Pacific salmon is in the developmental stage and few investigations have been carried out on genetic variations of economically im- portant traits. The chinook is the preferred species among fish farmers in Brit- ish Columbia, and research has just begun on the domestication and management of the fish (Kennedy, 1978; Mahnken and Waknitz, 1979; Hunter et al., 1983; Pereira and Adelman, 1985; McCallum et al., 1987).
Fish suitable for aquaculture have to grow rapidly, mature slowly, and sur- vive well under pen conditions. It has been demonstrated in other salmonid species that body weight has a high heritability while survival has a low to
0044-m86/87/%03.50 0 1987 Elsevier Science Publishers B.V.

302
moderate heritability ( Gjedrem, 1983; Gjerde and Gjedrem, 1984; Robison and Luempert, 1984). Donaldson (1970) selected chinook salmon for fast growth and other qualities under sea-ranching conditions and obtained a response in increased body weight of returning fish in two generations. The purpose of the present study was to compare the survival and growth rate of two strains of chinook salmon and their hybrids under pen-rearing conditions.
MATERIALS AND METHODS
Breeding design
This study was conducted concurrently with another study on the genetics of flesh pigmentation ( McCallum et al., 1987) and utilized the same breeding framework. Eggs and milt were obtained from fish caught from the Harrison River, a tributary of the Fraser River system, and from the Capilano River, B.C., Canada. Eggs from each of three Capilano (C) and three Harrison (H) females were divided into six lots. Each lot of eggs from a female was fertilized with sperm from one of six adult males, three Capilano and three Harrison. The 6 x 6 factorial cross resulted in 36 full-sib families with half-sib relations.
Rearing procedure
Following fertilization, the eggs were water-hardened in 10 ppm erythro- mycin for 15 min, rinsed to remove excess milt and ovarian fluid, disinfected in a solution of 200 ppm of iodine, and placed in a Heath stack incubator.
At approximately 5 weeks after hatching (late January), the fish were placed into 36 five-l tanks, keeping the full-sib families separate. Approximately the same number of fish was put into each tank, and a commercial diet (Oregon moist mash) was fed during the next month. At 9 weeks post-hatch, the fish were transferred to 160-l tanks, again keeping the full-sib families separate. Water temperatures increased from 65°C in March to 125°C in the middle of July. Artificial light was provided according to the natural photoperiod. The formula of the pelleted diet fed was given in McCallum et al. (1987).
The fish were freeze-branded at 24 weeks of age to identify full-sib families. Before transfer to sea-pens at Tidal Rush Marine Farms, Nelson Island, sam- ples of the fish were submitted to the Diagnostic Service of the Pacific Biolog- ical Station at Nanaimo to confirm that the fish were free of infectious disease. At 25 weeks of age the fish were vaccinated against Vibrio anguillarum. The smolts were moved to two 7 m x 7 m x 7 m deep net-pens at Tidal Rush Marine Farms at 26 weeks of age (July). Similar numbers of each full-sib family were placed in each pen. The final fish density at harvest time was approximately 3 kg/m3.
The fish were counted and weighed periodically during both the fresh- and

303
sea-water phases. Percent precocious males (jacks) was noted when the fish were 100 weeks old: At approximately 120 weeks of age, fish were harvested, dressed and weighed.
Analyses of data
The 36 full-sib families were classified into four types of crosses: CC, HC, CH, and HH, where the first letter denotes the strain of dam and the second letter denotes the strain of sire. Differences among the progeny of the four types of crosses were analysed ( PROC GLM; SAS, 1985)) using the following models:
Yijk* = u + Ci + Sij + Dik + SD,, + erlkl Model 1
Yij =U+Ci +ei; Model 2
where Y,;,, is the observation on the Ith progeny of thejth sire and the kth dam in the ith cross, and u is the population mean, Ci is the effect of the ith cross, Sij is the effect of thejth sire in the ith cross, Dik is the effect of the kth dam in the ith cross, SD,k is the interaction between the jth sire and the kth dam in the ith cross, and eijkl is the random error. Model 1 was used for traits where individual observations were available and Model 2 was used when only family means were available. When Model 1 was applied for observations in the fresh- water phase, SD,,k was used as the error term because of confounding family with tank effects; for observations obtained during the sea-water phase, eijkl was used as the error term. Orthogonal contrasts were set up to test for differ- ences between pure strains, heterosis effects, strain-of-sire effects, strain-of- dam effects, and reciprocal cross-effects.
Since fish were not identified individually, specific growth rate between any two weighings was calculated using family means as follows:
[In (weight 2) -In (weight 1) ]/age difference in days
All percentages were arcsin transformed prior to analyses ( Bartlett, 1947 1,
and all body weight measurements were log-transformed to decrease hetero- geneity in variances among the crosses.
RESULTS
Fertility and hatchability
There were significant differences ( P-zO.05) in fertility (percent of eggs reaching the eyed stage) among the four crosses. The means for CC, HC, CH, and HH were 97.7%, 65.9%, 90.1%, and 62.1%, respectively. Orthogonal com- parisons indicated that there was no difference between the two types of males
304
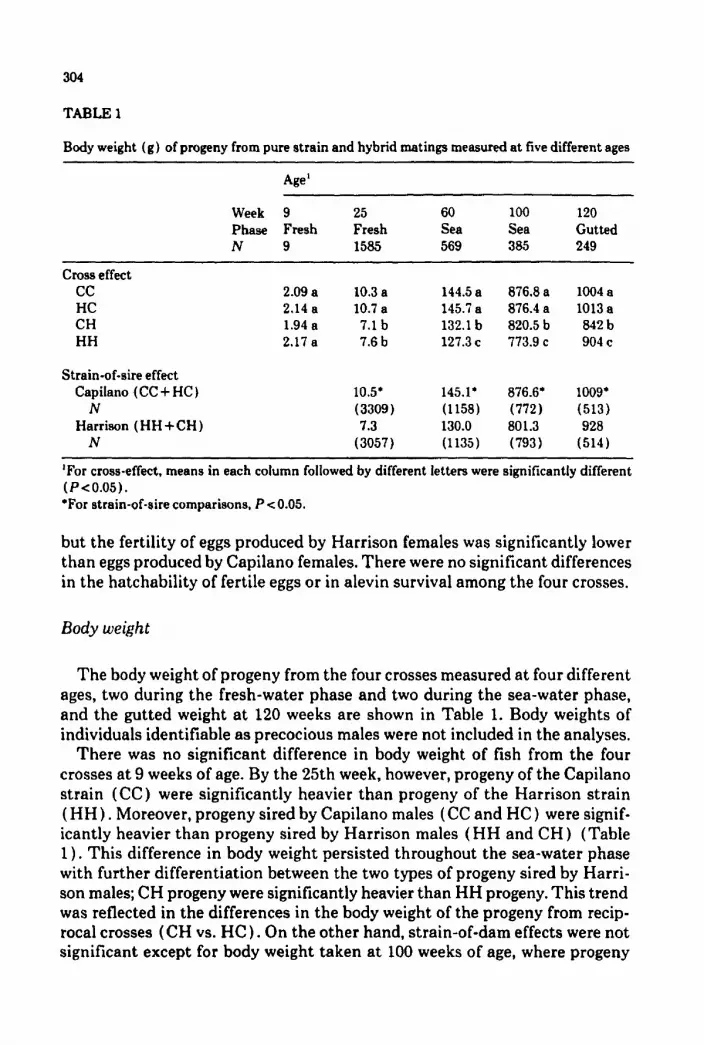
TABLE 1
Body weight (g) of progeny from pure strain and hybrid matings measured at five different ages
Age’
Week 9 25 60
Phase Fresh Fresh Sea
N 9 1585 569
100 120 Sea Gutted 385 249
cross effect cc HC CH HH
Strain-of-sire effect Capilano (CC + HC )
N Harrison (HH+CH)
N
2.09 a 10.3 a 2.14 a 10.7 a
1.94 a 7.1 b 2.17 a 7.6 b
10.5, (3309) 7.3
(3057)
144.5 a 145.7 a 132.1 b 127.3 c
145.1’ (1158) 130.0 (1135)
876.8 a 876.4 a 820.5 b 773.9 c
876.6’
(772) 801.3
(793)
1004 a 1013 a 842b 904 c
1009’
(513) 928
(514)
‘For cross-effect, means in each column followed by different letters were significantly different
(P<O.O5). *For strain-of-sire comparisons, P c 0.05.
but the fertility of eggs produced by Harrison females was significantly lower than eggs produced by Capilano females. There were no significant differences in the hatchability of fertile eggs or in alevin survival among the four crosses.
Body weight
The body weight of progeny from the four crosses measured at four different ages, two during the fresh-water phase and two during the sea-water phase, and the gutted weight at 120 weeks are shown in Table 1. Body weights of individuals identifiable as precocious males were not included in the analyses.
There was no significant difference in body weight of fish from the four crosses at 9 weeks of age. By the 25th week, however, progeny of the Capilano strain (CC) were significantly heavier than progeny of the Harrison strain (HH) . Moreover, progeny sired by Capilano males (CC and HC) were signif- icantly heavier than progeny sired by Harrison males (HH and CH ) (Table 1) . This difference in body weight persisted throughout the sea-water phase with further differentiation between the two types of progeny sired by Harri- son males; CH progeny were significantly heavier than HH progeny. This trend was reflected in the differences in the body weight of the progeny from recip- rocal crosses (CH vs. HC) . On the other hand, strain-of-dam effects were not significant except for body weight taken at 100 weeks of age, where progeny

305
TABLE 2
Specific growth rate ( 56 increase in body weight/day ) of progeny from the four crosses
Periods’
Phase Fresh Seal Sea 2 Sea Overall
Weeks 9-25 26-60 60-100 26-100 9-100 Days’ 112 245 280 525 637
Cross CC HC CH HH
1.79 a 1.10 b 0.67 a 0.88 b 1.01 a
1.76 a 1.10 b 0.66 a 0.87 b 1.00 ab 1.49 b 1.21 a 0.68 a 0.93 a 1.01 a
1.41 b 1.18a 0.67 a 0.91 a 0.96 b
Strain-of-sire Capilano (CC + HC)
Harrison (HH + CH)
1.77’ 1.10. 0.66 0.87. 1.00
1.45 1.20 0.68 0.92 1.00
‘For the crosses, means in each column followed by different letters were significantly different (P<O.OS).
‘Number of days in the period. *For strain-of-sire comparisons, PC 0.05.
from Capilano dams (845.4 g) were heavier than those from Harrison dams (830.4 g). It should be noted that prior to 100 weeks, precocious males were not yet identifiable and their body weights would be included in the analyses, but at 100 weeks they were identified and their measurements were not in- cluded in the analyses.
Gutted weight showed the same pattern of strain differences as body weights measured in the sea-water phase. Orthogonal comparisons indicated that there was no heterosis for body weights or gutted weights.
Within crosses, sire effects were significant for measurements taken at and after 25 weeks of age, whereas dam effects were only significant for measure- ments taken after the fish were in the sea-pens. For all the body weight mea- surements, the mean squares for sires were at least twice those for dams. Sire~dam interactions, measurable only after the fish were in the sea-pens, were significant for all three measurements.
Specific growth rate
Specific growth rates for the three growth periods are shown in Table 2. For the “sea 2” phase, as well as the overall rates, body weights of precocious males were excluded in calculating the family means of body weight from which the specific growth rates were derived. Progeny from Capilano males grew signif- icantly faster than progeny from Harrison males in the fresh-water phase (from

306
TABLE 3
The survival rate ( % ) of progeny from the four crosses (values in parentheses indicate the number of fish at the beginning of the period)
Periods’
Phase Fresh Weeks 9-25 Days’ 112
Sea 1 26-60 245
Sea 2 Overall so-log 9-100 280 637
cc 92.3 b (1788)
33.2 a (1650)
67.7 a
(548)
20.7 a
HC 96.3 ab (1723)
36.8 a (1659)
65.7 a
(610)
23.3 a
CH 92.1 b (1778)
38.4 a (1638)
74.2 a
(629)
26.3 a
HH 97.7 a 35.4 a 65.1 a 22.5 a (1453) (1419) (506)
‘For each column, means followed by different letters were significantly different (P <0.05). ‘Number of days in the period.
9 weeks to 25 weeks) but the reverse was true during the first period when the fish were in sea-water (Table 2). Despite this faster growth rate at the initial sea-water phase, CH and HH progeny were not able to catch up with CC and HC progeny for the final body weight measurements. There was no significant difference in specific growth rate between progeny from the two types of dams during each of the three growth periods, but there was a significant strain-of- dam effect for the overall growth period (9-100 weeks); progeny from Capi- lano dams grew faster than those from Harrison dams. The heterosis effect was not significant.
Survival rate
The survival rate for progeny from the Harrison strain was better than that of the Capilano strain in the fresh-water tanks (Table 3). Orthogonal com- parisons showed that progeny from Harrison females ( HH and HC) had sig- nificantly better survival rates than progeny from Capilano females. Heterosis was not detected. There were no significant differences in survival rate among progeny of the four crosses when the fish were held in the net-pens. The low survival rates observed in these pens were due to predation by sea otters.

307
TABLE 4
Percent of precocious males in progeny of the four crosses sampled at two locations (values in parentheses indicate number of fmh sampled)
Locations
Net-pens Sea-water tanks
cc (371) 9.1 (11) 9.1 HC (401) 9.6 (21) 28.6 CH (467) 13.4 (15) 26.7 HH (327) 19.7 (161 37.5
Precocious males
When the fish were transferred to the net-pens at 26 weeks of age, a small number from each cross were transferred to out-door sea-water tanks at the West Vancouver Laboratory, Fishery and Oceans Canada, to be kept as brood- stocks. The percentage of precocious males from the net-pen population sam- pled at 100 weeks of age, as well as that for fish held in the sea-water tanks, are shown in Table 4. Although the difference in percent jacks in the four crosses was not statistically significant, the trends in both populations were the same; CC had the lowest percent of jacks while HH had the highest, with intermediate value for the hybrids. In all four crosses, the body weight of jacks was not significantly different from that of the rest of the population measured at 100 weeks of age.
DISCUSSION
Pure strain comparison
The present day Capilano stock derives from a transplant from the Big Qual- icum River and has been exposed to a large degree of artificial propagation (Salmon Enhancement Program Annual Reports, 1980-1982). Although there were some stocking efforts in Harrison Lake and the Harrison River in the eighteen-eighties (Are, 1979), the Harrison stock can be considered wild (Withler, 1982). As a result, the Capilano strain may have been selected for adaptation to artificial propagation and performed better both in terms of egg survival and embryonic growth under such conditions (Reisenbichler and McIntyre, 1977).
Progeny of Harrison females survived better in the fresh-water tanks than the progeny of Capilano females. Levy and Northcote (1981) suggested that fry and fingerlings of chinook salmon from stocks further up the Fraser may have better survival than those from stocks closer to the sea, considering the

308
rigours necessary for a longer migration down the turbulent river. Despite ef- forts to start our experiment with the same number of fish in each full-sib family, there were four families with significantly fewer fish because of the poor fertility of eggs from Harrison females. The lower density in tanks containing these families, all involving Harrison dams, may have contributed to the higher survival rate of progeny from Harrison females (Refstie and Kittelsen, 1976). The same density factor also may have resulted in the slightly (but non-sig- nificant ) faster growth in progeny from Harrison dams up to 9 weeks of age.
Although the Capilano strain had a faster growth rate than the Harrison strain in the fresh-water tanks, the reverse was true for their “smolts” during the initial sea-water period. It is unlikely that untimely transfer to sea-water of the Capilano fingerlings would adversely affect their growth rate [e.g., see Clarke and Nagahama (1977) for coho salmon] since chinook fry may safely enter salt water at any time after reaching 4 g (Hoar, 1976; Clarke and Shel- bourn, 1985), and the respective fresh-water and sea-water temperatures in the present study were well within the optimal ranges described by Clarke and Shelbourn (1985). Rather, there may be a heritable difference in the growth curve of the two strains related to fresh/sea-water residence (Refstie et al., 1977; Thorpe and Morgan, 1978). Despite differences in growth pattern, fish of the Capilano strain were heavier at harvest.
In nature, small differences in growth, and hence in juvenile size at the time of entry into the marine environment, are believed to be critical for survival at sea (Parker, 1971; Hoar, 1976). In the net-pens, however, a different set of factors may become important for survival (Mahnken and Waknitz, 1979). We did not find any difference in the survival of the two strains and their hybrids in the net-pens.
The lower incidence of precocious males in the Capilano strain kept under two different conditions (net-pens and sea-water tanks) may have resulted from the fact that in the artificial propagation of the Capilano strain, only adult males and females were used as a source of milt and eggs (E. Stone, personal communication, 1985). This practice has been postulated to lower the percent of precocious males in a population (Gardner, 1976; Naevdal et al., 1978; Gjerde and Gjedrem, 1984; Gjerde, 1984; Hard et al., 1985),
Hybrid performance
Although Gall (1975)) Kincaid (1976) and Gjerde et al. (1983) found evi- dence that non-additive genetic variance is important in affecting body weight of rainbow trout, and Gall (1975) and Ayles and Baker (1983) reported sig- nificant heterosis for body weight among crosses of rainbow trout strains, Gjerde and Refstie (1984) did not find significant heterosis for either growth rate or survival in crosses of five strains of Atlantic salmon. In our experiment with chinook salmon, the dam effect and the dam~sire interaction within cross

309
were significant for body weight measurements taken after the fish were in sea-water, but heterosis was not detected. Furthermore, for body weight and specific growth rate, the strain cross means were similar to the strain-of-sire means. This is consistent with the findings of Bailey and Saunders (1984) for body size in crosses of Atlantic salmon strains. Thorpe and Morgan (1978) found that variation between families in smolting rate and mortality rate in Atlantic salmon is influenced by both parents, but more by the sire than by the dam. Dunham et al. (1982) have reported that blue-channel catfish hy- brids likewise possess traits that are more like the male parent, and referred to the phenomenon as “paternal predominance”. The strong sire effect in these crosses is an interesting phenomenon, and the underlying genetic mechanism deserves further investigation.
ACKNOWLEDGEMENTS
This study was supported by a grant from the Science Council of British Columbia to BEM. E.M. Donaldson generously permitted the use of incuba- tion facilities at the West Vancouver Laboratory, Fisheries and Oceans Can- ada. We also acknowledge the co-operation of Tidal Rush Marine Farms in providing net-pens, and their staff, M. Rudd, J. Powell, and J. Francis for taking care of the fish during the sea-water phase of the study. E Stone, T. Perry, and L. Kahl of the Salmonid Enhancement Program, I. Baker of the West Vancouver Laboratory, and K. Wilcox provided assistance during various stages of the study. We thank C.A.E. Call and C.B. Schom for helpful sugges- tions, and W.S. Hoar and G. Thorgaard for reviewing an earlier draft of this manuscript.
REFERENCES
Are, K.V., 1979. Transfers of eggs and young of Pacific salmon within British Columbia. Tech. Rep. 861, Fish. Mar. Serv. Can., 147 pp.
Ayles, G.B. and Baker, RF., 1983. Genetic differences in growth and survival between strainsand hybrids of rainbow trout (S&no gairdneri) stocked in aquaculture lakes in the Canadian prairies. Aquaculture, 33: 269-280.
Bailey. J.K. and Saunders, R.L.. 1984. Returns of three year-classes of sea-ranched Atlantic salmon of various river strains and strain crosses. Aquaculture. 41: 259-270.
Bartlett. M.S.. 1947. The use of transformations. Biometrics, 3: 39-52. Clarke, WC. and Nagahama, Y.. 1977. Effect of premature transfer to seawater on growth and
morphology of the pituitary, thyroid, pancreas, and interrenal in juvenile coho salmon (On- corhynchus k&t&). Can. J. Zool., 55: 1620-1630.
Clarke, W.C. and Shelbourn. J.E., 1985. Growth and development of seawater adaptability by juvenile fall chinook salmon (Oncorhynchus tshowytscha) in relation to temperature. Aqua- culture, 45: 21-31.

310
Donaldson, LR., 1970. Selective breeding of salmonid ftshea. In: W.J. McNeil (Editor), Marine Aquiculture. Oregon State University Press, Corvallis, OR, pp. 65-74.
Dunham, R-A.. Smitherman. R.O., Brooks, M.J., Ben&&an, M. and Chappell, J.A., 1982. Pater- nal predominance in channel-blue hybrid catfish. Aquaculture. 29: 389-396.
Gall, G.A.E., 1975. Genetics of reproduction in domestic rainbow trout. J. Anim. Sci., 40: 19-28. Gardner, M.L.G., 1976. A review of factors which may influence the sea-age and maturation of
Atlantic salmon Salmo salar, L. J. Fish Biol., 9: 289-327. Gjedrem, T., 1983. Genetic variation in quantitative traits and selective breeding in fish and shell-
fish. Aquaculture, 33: 51-72. Gjerde. B.. 1984. Response to individual selection for age at sexual maturity in Atlantic salmon.
Aquaculture, 38: 229-240. Gjerde. B. and Gjedrem, T., 1984. Estimates of phenotypic and genetic parameters for carcass
traits in Atlantic salmon and rainbow trout. Aquaculture, 36: 97-110. Gjerde, B. and Refstie, T., 1984. Complete diallel cross between five strains of Atlantic salmon.
Livest. Prod. Sci., 11: 207-226. Gjerde, B., Gunnes, K. and Gjedrem, T., 1983. Effect of inbreeding on survival and growth in
rainbow trout. Aquaculture. 34: 327-332. Hard, J.J., Wertheimer, A.C., Heard, W.R. and Martin, R.M., 1985. Early male maturity in two
stocks of chinook salmon (Oncorhynchus tshawytscha) transplanted to an experimental hatchery in southeastern Alaska. Aquaculture, 48: 351-359.
Hoar, W.S., 1976. Smelt transformation: evolution, behavior, and physiology. J. Fish. Res. Board Can., 33: 12234-1252.
Hunter, G.A., Donaldson, E.M., Stoss, J. and Baka, I., 1983. Production of monosex female groups of chinook salmon (Oncorhynchus tshawytscha) by the fertilization of normal ova with sperm from sex reversed females. Aquaculture. 33: 355-364.
Kincaid. H.L.. 1976. Inbreeding in rainbow trorut (Safmo gairdneri) , J. Fish. Res. Board Can., 33: 2420-2426.
Kennedy, W.A., 1978. A Handbook on Rearing Pan-Size Pacific Salmon Using Floating Sea Pens. Ind. Rep. No. 107, Fish. Mar. Serv. Can., 117 pp.
Levy, D.A. and Northcote. T.G., 1981. The distribution and abundance of juvenile salmon in marsh habitats of the Fraser River estuary. Westwater Res. Cent. Tech . Rep. 25. University of British Columbia, Vancouver, B.C., 68 pp.
Mahnken, C.V. and Waknitz, F.W., 1979. Factors affecting growth and survival of coho salmon (Oncorhynchm kisutch) and chinook salmon (0. tshawytscha) in salt-water net-pens in Puget Sound. Proc. World Maricult. Sot., 10: 280-305.
McCallum, I.M., Cheng, K.M. and March, B.E.. 1987. Carotenoid pigmentation in two strains of chinook salmon (Oncorhynchus tshawytscha) and their crosses. Aquaculture. 67: 291-300.
Naevdal, G., Holm, M., Ingebrigtsen, 0. and Moller. D.. 1978. Variation in age at first spawning in Atlantic salmon (Salmo salar). J. Fish. Res. Board Can., 35: 145-147.
Parker, R.R., 1971. Size selective predation among juvenile salmonid fishes in a British Columbia inlet. J. Fish. Res. Board Can., 28: 1503-1510.
Pereira, D.L. and Adelman, I.R., 1985. Interactions of temperature, size and photoperiod on growth and smoltification of chinook salmon (Oncorhynchw tshawytscha), Aquaculture, 46: 185-192.
Refstie, T. and Kittelsen, A., 1976. Effect of density on growth and suntival of artificially reared Atlantic salmon. Aquaculture, 8: 319-326.
Refstie, T., Steine, T.A. and Gjedrem, T., 1977. Selection experiments with salmon. II. Proportion of Atlantic salmon smoltifying at one year of age. Aquaculture, 10: 231-242.
Reisenbichler, R.R. and McIntyre, J.D., 1977. Genetic differences in growth and survival of ju- venile hatchery and wild steelhead trout: Salmogairdneri. J. Fish. Res. Board Can., 34: 123-128.
Robison, O.W. and Luempert, L.C.. III., 1984. Genetic variation in weight and survival of brook trout (Soloelinw fontinalis). Aquaculture. 38: 155-170.

311
Salmonid Enhancement Program Annual Reports, 1980-1982. Fisheries and Oceans Canada, British Columbia, Canada.
SAS Users’ Manual, 1985. SAS Institute, P.O. Box 8000, Gary. NC 27511, U.S.A., 584 pp. Thorpe, J.E. and Morgan, R.I.G.. 1978. Parental influence on growth rate, smoking rate and
survival in hatchery reared Atlantic salmon, Salmo safar. J. Fish Biol., 13: 549-556. Withler. F.C., 1982. Transplanting Pacific salmon. Fish. and Aquatic Sci. Can. Tech. Rep. 1079.
27 pp.





























