Embed Size (px)
Citation preview

Aquaculture
ELSEVIER Aquaculture 157 (1997) 3 1 I-323
Control of growth and adiposity of juvenile chinook salmon (Oncorhynchus tshawytscha)
Karl D. Shearer a, * , Jeffrey T. Silverstein b, Walton W. Dickhoff ‘yb
’ Northwest Fisheries Science Center, NMFS, NOAA, 2725 Montlake Blvd. E., Seattle, WA 98112, USA
b School of Fisheries, UniL,ersity of Washington, 355100 Seattle, WA 98195, USA
Accepted 24 June 1997
Abstract
Chinook salmon fry (0.7 g, 3.9% whole body lipid) were fed high protein diets containing high lipid (HL) (65% protein, 23% lipid) or low lipid (LL) (85% protein, 3% lipid) at high ration (near satiation = HR) or low ration (one half of high ration = LR) for 247 days to determine if growth and adiposity could be independently controlled. Each treatment was duplicated with each tank containing 400 fish. The amount of feed fed to each tank of fish was adjusted each month so that fish fed at a prescribed ration level (HR or LR) would grow at similar rates. The effects of ration and dietary lipid level on final weight, adiposity, feed efficiency (wet weight gain/dry feed fed) and protein retention efficiency were assessed using regression, two-way ANOVA and ANCOVA (P < O.OS>. Overall feed efficiency (lO9-127%) was lowest in the HR/HL treatment but differences among treatments were not significant. For the different treatments (HR/HL, HR/LL, LR/HL, LR/LL), the amounts of dry feed fed averaged; 17.5, 15.4, 6.7 and 6.1 g/fish, final mean fish weights were, 20.2, 19.5, 9.2 and 8.7 g and final mean whole body lipids were, 11.3, 5.4, 7.1, 3.9%. These results show that ration level had the major influence on growth, whereas dietary lipid level was the major determinant of adiposity. This appears to have occurred because when protein was supplied in excess, it was not spared by lipid. Thus it may be possible for enhancement hatcheries and commercial salmon farms to manipulate growth and adiposity by regulating ration and controlling diet composition. 0 1997 Elsevier Science B.V.
Keywords: Chinook salmon: Growth; Adiposity; Feeding rates; Dietary lipid
* Corresponding author. Tel.: + l-206-860-3393; fax: + I-206-860-3394; e-mail: [email protected].
noaa.gov.
0044.8486/97/$17,00 0 1997 Elsevier Science B.V. All rights reserved PII SOO44-8486(97)00170- I

312 K.D. Shearer et al./Aquaculture 157 (1997) 311-323
1. Introduction
In the Pacific Northwestern United States and Western Canada, hatchery reared chinook salmon return from the ocean to spawn at a lower rate than wild fish (Plotnikoff et al., 1983; McIntyre, 1987; Raymond, 1988; Miller et al., 1990). Possible reasons include increased incidence of disease, behavioral differences and differences in physiol- ogy (Burrows, 1969; NRC, 1996). At the time of release, hatchery reared salmonids are normally larger than wild fish (Kaczynski et al., 1993; Shrimpton et al., 1994; NRC, 1996) and have higher lipid concentrations (Wood et al., 1957, 1960; Ludwig, 1980; Hata et al., 1988). Both size and adiposity (stored lipid) have been shown to affect smoltification and early maturation in salmonids (Bilton, 1984; Ogata and Konno, 1986; Thorpe, 1986; Silverstein et al., 1997).
Several workers (Clarke et al., 1981; Clarke and Shelboum, 1985; Dickhoff et al., 1989; Virtanen et al., 1991) have attempted to determine the effects of size or growth rate on smoltification or return rate of hatchery reared salmonids. Growth has usually been manipulated by varying temperature and/or ration size. Unfortunately, while temperature and ration manipulations modify growth rate, they also influence adiposity (Brett et al., 1969, Reinitz, 1983; Storebakken et al., 1991). It is difficult, therefore, to ascribe differences in smoltification, early maturation or survival to differences in growth per se.
Growth and adiposity can also be affected by diet composition, most notably nutrient balance and nutrient density (Brett and Groves, 1979). Several workers (Clarke et al., 1982; Plotnikoff et al., 1983, 1984; Ogata and Konno, 1986; Higgs et al., 1992) have examined the effects of diet composition on smolt survival or adult return rate. In all of these studies both growth and adiposity were affected, confounding interpretation of observed effects.
Lee and Putnam (1973) demonstrated that varying the dietary protein/energy ratio can affect both growth rate and adiposity in rainbow trout. At low protein concentrations increasing dietary lipid led to increased feed efficiency, growth and adiposity, whereas at high dietary protein concentration (53%) adiposity increased, but growth was not improved by increasing dietary lipid. Takeuchi et al. (1978) reported similar findings when high protein diets (54%) containing 5 to 20% dietary lipid were fed to rainbow
trout. The objective of the current study was to determine if growth and adiposity could be
manipulated independently in juvenile chinook salmon. This was attempted by simulta- neously varying feed supply (ration) and the dietary lipid level in high protein diets. Specifically, our aim was to produce four groups of fish; two groups similar in size to those released from hatcheries, one with lipid levels similar to those of hatchery fish (high lipid) and a second, with lipid levels similar to wild fish (low lipid); and two groups similar in size to wild fish, but containing subgroups with high and low body lipid concentration.
The ability to manipulate growth and adiposity independently would allow examina- tion of the effects of these factors on smoltification, emigration, early maturation and adult return rate. In addition, it might provide insights into the relative effects of ration and dietary lipid level on carcass composition of farmed salmon.

K.D. Shearer et ul./Ayuaculture 157 (1997l311-323 313
2. Materials and methods
2.1. Rearing conditions
Chinook salmon (Oncorhynchus tshawytscha) eggs of mixed parentage, from the stock that returns to Willamette Hatchery, Oregon, were obtained in November 1994 and incubated at Northwest Fisheries Science Center, Seattle, Washington. At first feeding (Jan. 1995) fry were fed a commercial starter feed (manufacturers specifications, 45% protein and 16% lipid) (BioDiet-starter, BIOPRODUCTS, Warrenton, OR). When fry reached a mean weight of 0.7 g, they were counted into lots of 400 and placed into eight 700 1 tanks supplied with dechlorinated municipal water, at 6 l/min. Over the course of the experiment water temperatures ranged from a minimum of 6°C (Jan.> to a maximum of 17°C (Sept.), except for four days in September when temperatures reached 20°C due to a failure of the water-cooling system. The fish were exposed to natural light and photoperiod. The experiment was terminated in Sept. 1995, after 247 days.
2.2. Diets and feeding regime
Diets were prepared monthly, at the appropriate pellet size, and frozen at -20°C until fed. Two diets (Table 1) were prepared by adding different amounts of lipid to a basal mix so that one diet contained (by analysis) 85% protein and 3% lipid (low lipid = LL) and the other contained 65% protein and 23% lipid (high lipid = HL). The calculated gross energy contents were 23.3 and 20.8 kJ/g respectively for the HL and LL diets (Brafield, 1985). The high dietary protein levels were achieved by using dried
Table 1
Experimental diets”
Ingredient Low lipid diet
Dried fish muscleb 662.5
Wheat gluten 58.8
Ascorbic acid 12.9
CaHPO, 29.4
Vitamin mix‘ 17.6
Arginine 5.9
Gelatin 117.6
Herring oil 35.3
Choline Cl 11.8
Trace mineral mixd 1.2
Carboxymethyl cellulose 23.5
Algibind’ 23.5
“Water added (500 ml/kg dry diet).
hSupplied by NMFS, Kodiak, AK: 94% protein, 27~ lipid, 4% ash.
‘USFWS, Abernathy (Hardy, 1989).
‘USFWS, No. 3 (Hardy, 1989).
‘Algea Produkter A/S, Lier, Norway. Values are in g/kg dry weight.
High lipid diet
536.1
47.6
10.5
23.8
14.3
4.8
95.2
219.0
9.5
1 .o
19.0
19.0

314 K.D. Shearer et al./Aquaculture 157 (1997) 311-323
fish muscle (prepared by the National Marine Fisheries Service Laboratory in Kodiak, AK) as the main protein source. Fillets from Pacific cod (Gadus macrocephalus) were minced, washed, air dried (60°C) then ground into meal. This meal contained 94% protein, 2% lipid and 4% ash.
Fish were fed the HL or LL diets at either high ration (HR = near satiation) or low ration (LR = one half of HR). This combination of diet and ration produced four treatment groups: HR/HL, HR/LL, LR/HL, LR/LL, each treatment being run in duplicate tanks.
To insure that the fish fed at a given ration level were similar in weight, an iterative feeding regime was used. On day 1 of the experiment and on the day after each monthly sampling, the fish in the high ration tanks were fed to satiation (until pellets reached the bottom of the tank without being consumed) and the amount of feed eaten was noted. For the remainder of the first month, the high ration groups were fed 90% of the amount of feed consumed by the group that ate the least feed on day 1. Fish were fed slightly below satiation to help insure that all feed was consumed. During the first month low ration groups were fed half this amount. For the remainder of the experiment, the amount (g) to be fed to each tank of fish each day was calculated as follows; in the high ration tanks,
(CX0.9XN) + \ F I d
where: C = the average amount of feed consumed/fish, in the high ration tank that ate the least feed the first day after sampling (g); 0.9 = a factor to help insure that all feed fed was consumed; N = the number of fish in the tank; W,, = the largest mean weight of fish in the tanks fed the high ration (8); W = the mean weight of the fish in the tank (g); F = the feed efficiency in the tank the previous month (weight gain/dry feed fed); d = the number of feeding days before the next sampling; in the low ration tanks,
(Wm-W)XN C x 0.9 x N F
+ 2 d
where: W,, = the largest mean weight of fish in the tanks fed the low ration. Fish were generally fed 5 days per week. Feed was supplied twice each day until fish
ceased to feed or had consumed their allotted ration. If a group of fish failed to consume their feed allotment in five days they were fed on day 6 (Saturday).
2.3. Sampling and analysis
Mortality was recorded daily. The fish in each tank were batch weighed and counted on day one of the experiment and approximately monthly thereafter, to obtain a mean fish weight for each tank. Feed was withheld for one day prior to sampling. After each sampling, 7 fish from each tank were randomly removed for individual lipid analysis

K.D. Shearer et al./Aquaculrure 157 (199713/I-323 315
and three additional fish were removed and pooled for protein analysis. Samples of feed for analysis were taken during the first and last month. Fish were weighed, minced, and dried to constant weight at 105°C. Dried fish were ground with a pestle and mortar and lipid was extracted from either the whole fish or a subsample of 2 g of larger fish. Lipid was extracted using a Soxhlet device using methylene chloride as the solvent. Protein was determined using the Kjeldahl method (N X 6.25). Feed was analyzed in a similar manner.
2.4. Statistical analysis
Treatment effects were compared using linear regression, two-way ANOVA and Fisher’s Protected Least Significant Difference (PLSD) (Statview, 1992) and ANCOVA (SuperANOVA, 1990). U n ess 1 stated otherwise, tank mean values were used as the units of observation. Percentage data were arcsin transformed prior to analysis. Linear regressions were compared using the method of Zar (1974). Differences were accepted as statistically significant if P < 0.05.
3. Results
The different diets and feeding regimes produced four groups of fish: large, high lipid; large, low lipid; small, high lipid; small, low lipid. Within the HR groups, it was one of the tanks of fish in the HR/HL treatment that had the lowest feed consumption and therefore determined the amounts fed to the other tanks of fish. There was a tendency for feed efficiency to be higher in the low lipid (LL) than the high lipid treatments and lower in the high ration than the low ration treatments, but differences among treatments were not significant (Table 2). The average amounts of feed fed per fish differed significantly among treatments (Table 2). Mean final weights of fish fed high rations averaged 20.2 and 19.5 g for the HL and LL groups, respectively (Fig. 1, Table 2). Fish fed low rations averaged 9.2 g and 8.5 g in the HL and LL treatments (Table 2).
Initial whole body lipid was 3.9% and treatment differences were apparent after one month (Fig. 2). Final whole body lipids differed significantly among treatments and ranged from 3.9 to 11.3% (Table 2).
Mortality during the experiment was 7.0%: most fish died either during the first month of feeding or after a failure of the water chilling system (in September), when the temperature reached approximately 20°C for 4 days. Mortality was highest in the HR/HL treatment, but differences in mortality among treatments were not significant (Table 2).
Two-way ANOVA (Table 2) indicated that both ration level and dietary lipid level affected fish growth and adiposity. However, the fact that the HL or LL treatments were fed different amounts of feed complicates interpretation. When the effects of treatment (ration level and dietary lipid) on growth and adiposity were corrected for differences in feed fed and feed efficiency (ANCOVA, using feed efficiency X feed fed as a covariate) the effect of ration on growth was found to be significant, but the effect of lipid on

Tab
le
2
Mea
n (
f S
E);
fin
al w
eigh
t’, m
orta
lity,
fe
ed
fed,
fe
ed
effi
cien
cy
(wet
w
eigh
t ga
in/d
ry
feed
fe
d),
fina
l w
hole
bo
dy
prot
ein
(wet
ba
sis)
pr
oduc
tive
prot
ein
valu
e (P
PV)’
and
fina
l w
hole
bo
dy
lipid
(w
et
basi
s)
of j
uven
ile
chin
ook
salm
on
fed
a hi
gh
or l
ow
ratio
n co
ntai
ning
hi
gh
or l
ow
amou
nts
of l
ipid
(c
alcu
late
d en
ergy
de
nsity
, 23
.3
kJ/g
fo
r th
e hi
gh
lipid
di
et
and
20.8
kJ
/g
for
the
low
lip
id
diet
, B
rafi
eld,
19
85)
Tre
atm
ent
Fina
l w
t. (g
)’
Fina
l w
t. (g
)’
Mor
talit
y (%
) Fe
ed
fed
(dry
) Fe
ed
effi
cien
cy
%
Fina
l pr
otei
n 8
PPV
%
AN
OV
A
AN
CO
VA
* (g
/fish
) Fi
nal
lipid
%
Hig
h ra
tion/
Hig
h lip
id (
HR
/HL
) 20
.2 f
0.
la
22.2
+ 1
.6”
11.8
* 1.
3 17
.5 *
0.5”
10
9+
1.5
16.6
+0.7
28
.9t1
.3”
11.3
+0.2
” H
igh
ratio
n/L
ow
lipid
(H
R/L
L)
19.5
+0.
3b
20.1
f 0
.6a
7.3
+ 4.
5 15
.4 +
0.2
b 12
3kO
.5
17.2
+ 0
.6
25.9
kO.4
” 5.
4+0.
1’
Low
ra
tion/
Hig
h lip
id (
LR
/HL
) 9.
2 i
0.3’
7.
9 +
0.9b
1.
3*0.
9 6.
7 +
0.1’
11
8k8.
5 15
.8 k
O.9
29
.2*
1.2a
7.
1 fO
.lb
Low
ra
tion/
Low
lip
id (
LR
/LL
) 8.
5 +0
.2’
6.9
f 1 .
2b
7.7*
1.
9 6.
1 fO
.ld
127+
1.
5 15
.5+0
.7
22.0
+ 1.
6b
3.9k
O.l”
P (p
roba
bilit
y)
Rat
ion
< 0.
0001
0.
007
0.30
<
0.00
01
0.21
0.
11
0.20
<
0.00
01
Lip
id
0.02
0.
10
0.69
0.
02
0.06
0.
82
0.01
<
0.00
01
Rat
ion
X L
ipid
1 .
o 0.
32
0.26
0.
07
0.60
0.
52
0.15
0.
0001
‘n =
2 f
or
all
anal
yses
, co
lum
n m
eans
w
ith
diff
eren
t su
pers
crip
ts
are
sign
ific
antly
di
ffer
ent
(P
< 0.
05).
*(F
eed
effi
cien
cy
X f
eed
fed)
us
ed
as t
he c
ovar
iate
. T
his
adju
sts
for
diff
eren
ces
in f
eed
fed.
‘Pro
duct
ive
prot
ein
valu
e (P
PV
) =
(inc
reas
e in
tis
sue
prot
ein/
prot
ein
cons
umed
) X
100
. In
itial
w
hole
bo
dy
prot
ein
12.5
%.

K.D. Shearer et al./Aquaculture 157 (1997) 311-323 317
20.0
s =: 15.0 AZ .EF
I
5 10.0
ii
5.0
0.0
0 50 100 150 200 250 300
Day of the year
Fig. 1. Growth of juvenile chinook salmon fed a high (HR = near satiation) or low (LR = one-half satiation)
ration of diets containing high (HL = 23%) or low (LL = 3%) lipid.
growth was not significant (Table 2). Likewise, comparison of fish weight vs. whole body lipid over the course of the study using allometric analysis (Shearer, 1994) indicated that fish fed the same diet but different rations, had similar lipid levels at a given body size (Fig. 3). This indicates that dietary lipid level had a significant effect on whole body lipid but the effect of ration on whole body lipid was not significant.
Productive protein value (PPV) was significantly lower in the (LR/LL) treatment than in the other groups (Table 2). Final whole body protein concentrations were lowest in the LR fish, but differences (ANOVA) were not significant, and regression analysis
12.5 1
10.0 -
T @ 0 .n 7.5 - -
n” B 5.0- s
g 2.5 -
HWHL
LWHL
HWLL
“.O6 300
Day of the year
Fig. 2. Whole body lipid (% wet basis) of juvenile chinook salmon fed a high (near satiation) or low ration (near half satiation) of a high (23%) or low (3%) lipid diet: high ration/high lipid (HR/HL); high ration/low
lipid (HR/LL); low ration/high lipid (LR/HL); or low ration/low lipid (LR/LL). Data points represent the mean of two tanks.
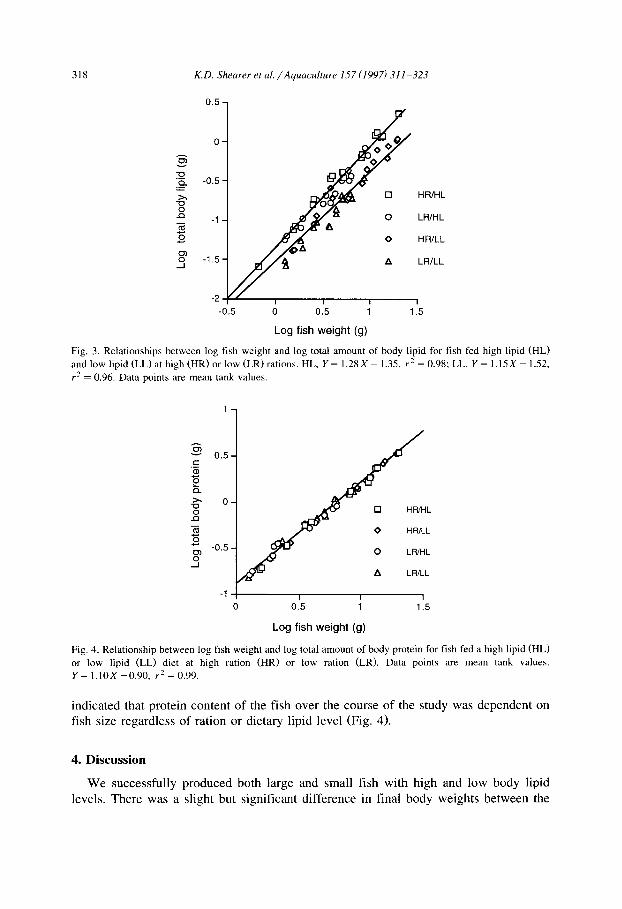
318 K.D. Shearer et al./Aquaculture 157 (1997) 311-323
0
-0.5
-1
-1.5
-2
HRIHL
LWHL
HWLL
LFVLL
-0.5 0 0.5 1
Log fish weight (g)
1.5
Fig. 3. Relationships between log fish weight and log total amount of body lipid for fish fed high lipid (HL)
and low lipid (LL) at high (HR) or low (LR) rations. HL, Y = 1.28 X - 1.35, r’ = 0.98; LL, Y = 1.15X - 1.52,
r* = 0.96. Data points are mean tank values.
0 HWHL
0 HFVLL
.5 0 LWHL
A LR/LL
“V 1.5
Log fish weight (g)
Fig. 4. Relationship between log fish weight and log total amount of body protein for fish fed a high lipid (HL)
or low lipid (LL) diet at high ration (HR) or low ration (LR). Data points are mean tank values.
Y = 1.10x - 0.90, rz = 0.99.
indicated that protein content of the fish over the course of the study was dependent on fish size regardless of ration or dietary lipid level (Fig. 4).
4. Discussion
We successfully produced both large and small fish with high and low body lipid levels. There was a slight but significant difference in final body weights between the

K.D. Shearer et al./Aquaculture 157 (19971311-323 319
high ration treatments due to differences in the amounts fed. Underfeeding of the low lipid groups was due to an overestimation of the feed efficiency projected to occur in the LL groups during the last month of the experiment. Low feed efficiencies may have been due to the high temperatures that occurred during this month, as feed efficiency can be reduced by an increased metabolic requirement at high temperatures (Brett et al., 1969; Wurtsbaugh and Davis, 1977) and the LL diet supplied less energy than the HL diet. The ANCOVA, using feed fed X feed efficiency as the covariate, indicated that if fish had been fed the same amount of feed they would have been the same size regardless of the dietary lipid level. This indicates that dietary lipid level did not influence weight gain (Table 2).
Ration level did not appear to effect adiposity, the controlling factors being dietary lipid level and fish size. Thus, fish fed the same dietary lipid level at high or low ration contained similar amounts of lipid at a given weight (Fig. 2).
Growth rates of the fish fed the HR/HL diet were lower than those that can be achieved under hatchery conditions (BPA, 1994). The fish in our experiment were reared under relatively good rearing conditions, the diets appeared to be palatable and feed efficiencies (wet weight gain/dry feed fed) were 109-127%. However, feed intake was relatively low in the HR/HL group, and it was this group that controlled intake, and consequently the growth rates, of the fish in all treatments. Feed intake averaged about 1.5% of wet body weight/day, on a dry feed basis, while about 2.5% would be fed to hatchery-reared salmonids of similar size at similar temperatures (Halver, 1989, p. 740). Thus the feeding regime used should have resulted in the fish being fed less than they were capable of consuming, but, during the latter part of the experiment, the HR/HL fish frequently failed to consume their allotted feed in 5 days
The control of feed intake is not well understood, but it is known to be multifactorial (Peter, 1979; Vahl, 1979; Fletcher, 1984; Shearer et al., in press). The work of Lee and Putnam (1973) is often cited as evidence that fish eat to meet an energy requirement. Thus, it would be expected that the fish fed the high lipid diet would eat less due to the high energy density of this diet (calculated at, 23.3 kJ/g; Brafield, 1985). The greater appetite, (they always consumed their daily feed allotment in five days) observed in the fish fed the HR/LL diet (20.8 kJ/g), supports this hypothesis.
Work by Metcalfe and Thorpe (19921, Jobling and Miglavs (1993) and a recent experiment in our laboratory (Shearer et al., in press) indicates that high adiposity leads to reduced feed intake. Thus the high adiposity of the HR/HL fish may be responsible for the relatively low feed intake of the fish in this treatment compared to fish reared in hatcheries. The data presented by Lee and Putnam (1973) for rainbow trout and Silver et al. (1993) for juvenile chinook salmon show that feed intake and growth were lower when high protein diets were fed and dietary lipid was high (24 and 25%) than when lower protein diets were fed. Juvenile salmonids generally increase their lipid content as they increase in size (Shearer, 1994) but fish in the HR/HL treatment may have exceeded their capacity to store lipid, which may have led to appetite suppression.
The dietary protein levels in the current experiment (65 and 85%) were much higher than those used by Lee and Putnam (1973) or Takeuchi et al. (1978) and excess protein energy was apparently stored as lipid. Fish fed the low lipid diet at the low ration level (LR/LL) gained more lipid than they were fed. Based on our results and those of Lee

320 K.D. Shearer-et al./Aquaculture 157(1997) 311-323
and Putnam (1973) and Takeuchi et al. (1978) it appears that it should be possible to lower the dietary protein level to slightly over 50% and grow fish at a desired rate and adiposity, by varying the ration and dietary lipid level. Since feed efficiencies were not significantly different among treatments, growth rates should be linearly related to feed intake between the maintenance ration and satiation (Mlkinen, 1993).
The protein level in the HL diet (65%) was well above the 35-45% recommended by Higgs et al. (1995) but the LL diet contained an even higher level of protein (85%) and fish still maintained good appetite (always consumed their daily feed allotment). This indicates that the high protein levels in our diets were not detrimental to fish health or appetite.
The lipid level in the LL diet (3%) was well below that normally fed to juvenile chinook salmon but, no signs of essential fatty acid deficiency were noted. The low lipid diet therefore, appeared to contain sufficient amounts of essential fatty acids and sufficient energy from the high level of protein for maintenance and growth.
The protein/energy ratios of our diets (HL, 28; LL, 41 g/MJ) were well above the protein/energy ratios (23-25 g/MJ) recommended by Higgs et al. (1995) but we observed no apparent health problems in the fish fed the LL diet which had the highest protein/energy ratio.
Higgs et al. (1995) recommend a dietary energy density of 14- 18 MJ/kg. The energy densities of our diets (HL, 23.3 MJ/kg; LL, 20.8 MJ/kg) were considerably higher but were similar to the natural food (19.5-25 MJ/kg dry weight) of chinook salmon (Higgs et al., 1995). The high energy density of the HL diet does not, therefore appear to have been the sole factor responsible for the low feed intake of the HR/HL treatment fish. We believe, based on the high adiposity and low feed intake of the HR/HL fish, that the dietary lipid level (23%) rather than the energy density per se, was too high for juvenile chinook salmon. It also appears that a high energy diet may be appropriate if the energy is supplied by protein, since this is similar to the natural diet of juvenile chinook salmon (Higgs et al., 1995). Our results also confirm the observations of Wathne (1995) who reported that dietary protein to fat ratio has a greater affect on carcass fat in farmed salmon than ration level, and that high growth rates and low adiposity can be achieved by feeding high energy diets where energy is supplied by protein.
In the present study only two levels of ration and two dietary lipid levels were examined. The ration levels were chosen to produce fish with growth rates similar to those of wild or hatchery fish. Similarly, dietary lipid levels were chosen to produce fish with relatively high lipid levels, similar to hatchery fish, and low levels similar to those of wild fish. The range of adiposities produced (3.9 to 11.3%) was higher than those observed by Burrows (1969) (2 and 8%) or Higgs et al. (1995) (1.8 and 6.1%) in wild and hatchery-reared juvenile chinook salmon. This indicates that lower dietary energy density than that used in our study is required to produce a wild type, low adiposity, smolt.
Our study indicates that growth and adiposity can be manipulated independently in juvenile chinook salmon by varying ration and diet composition. This will allow examination of the independent and combined effects of these factors on metabolism, smoltification, early male maturation and immunocompetence in juvenile salmonids.

K.D. Shearer et al. /Aquaculture 157 (1997) 311-323 321
Our results also support the hypothesis that carcass fat in production fish can be reduced by feeding diets high in protein and low in lipid.
Acknowledgements
We wish to thank Cindy Rathbone for the protein determinations and an anonymous reviewer for greatly improving the manuscript. This study was funded in part by USDA grant # 94-37206-1096 to K.D.S., W.W.D. and Dr. Erika M. Plisetskaya, and by projects # DE-A179-93BP55064and # 92-022 from The Division of Fish and Wildlife, Bonneville Power Administration, Department of Energy to Dr. Penny Swanson and W.W.D., respectively.
References
Bilton, H.T., 1984. Returns of chinook salmon in relation to juvenile size at release. Can. Tech. Rept. Fish.
Aquat. Sci., No. 1245. BPA (Bonneville Power Administration), 1994. Policies and Procedures for Columbia Basin Anadronomous
Salmon Hatcheries. US Dept. of Energy, BPA-2432.
Brafield, A.E., 1985. Laboratory studies of energy budgets. In: Tytler, P., Calow, P. (Eds.), Fish Energetics.
New Perspectives. Croom Helm, London, pp. 257-281.
Brett, J.R., Groves, T.D.D., 1979. Physiological energetics. In: Hoar, W.S., Randall, D.J., Brett, J.R. (Eds.),
Fish Physiology. Academic Press, New York, Vol. VIII, pp. 280-352.
Brett, J.R., Shelbourn, J.E., Shoop, C.T., 1969. Growth rate and body composition of fingerling sockeye
salmon (Oncorhynchus nerka) in relation to temperature and ration size. J. Fish. Res. Board Can. 26,
2363-2394.
Burrows, R.E., 1969. The influence of fingerling quality on adult salmon survivals. Trans. Amer. Fish. Sot.
98, 777-784.
Clarke, W.C., Shelbourn, J.E., 1985. Growth and development of seawater adaptability by juvenile fall
chinook salmon (Oncorhynchus tshawytscha) in relation to temperature. Aquaculture 45, 2 l-3 1.
Clarke, W.C., Shelboum, J.E., Brett, J.R., 1981. Effect of artificial photoperiod cycles, temperature, and
salinity on growth and smolting in underyearling coho (Oncorhychus kisutch), chinook (0. tshawytscha),
and sockeye (0. nerka) salmon. Aquaculture 22, 105-116.
Clarke, W.C., Higgs, D.A., Markert, J.R., Shelboum, J.E., Castledine, A.J., 1982. Effects of varying dietary
protein:lipid ratios on growth and body composition of coho salmon fry (Oncorhychus kisutch) reared at
different temperatures. Can. Data Rept. Fish. Aquat. Sci., No. 373.
Dickhoff, W.W., Mahnken, C.V.W., Zaugg, W.S., Waknitz, F.W., Bernard, M.G., Sullivan, C.V., 1989.
Effects of temperature and feeding on smolting and seawater survival of Atlantic salmon (Salmo salar).
Aquaculture 82, 93-102.
Fletcher, D.J., 1984. The physiological control of appetite in fish. Comp. Biochem. Physiol. 78A, 617-628.
Halver, J.E. (Ed.), 1989. Fish Nutrition. Academic Press, San Diego, CA.
Hardy, R.W., 1989. Diet preparation. In: Halver, J.E. (Ed.), Fish Nutrition. Academic Press, San Diego, CA,
pp. 475-548.
Hata. M., Sate, Y., Yamaguchi, T., Ito, M., Kuno, Y., 1988. The chemical and amino acid compositions of cultured and wild coho salmon Oncorhyzchus kisutch. Nippon Suisan Gakkaishi 54, 1365-1370.
Higgs, D.A., Dosanjh, B.S., Plotnikoff, M.D., Markert, J.R., Lawseth, D., McBride, J.R., Buckley, J.T., 1992.
Influence of dietary protein to lipid ratio and lipid composition on the performance and marine survival of hatchery reared chinook salmon (Oncorhyzchus tshaw?;tscha). Bull. Aquacult. Assoc. Can. 92, 46-48.
Higgs, D.A., Macdonald, J.S., Levings, C.D., Dosanjh, B.S., 1995. Nutrition and feeding habits in relation to
life history stage. In: Groat, C., Margolis, L., Clarke, W.C. (Eds.1, Physiological Ecology of Pacific Salmon. UBC Press, Vancouver, Canada, pp. 159-3 16.

322 K.D. Shrurer et al./Aquaculture 157 (19971311-323
Jobling, M., Miglavs, I., 1993. The size of lipid depots-a factor contributing to control of food intake in Arctic charr, Sal~~elinus rrlpinus?. J. Fish Biol. 43, 487-489.
Kacaynski, V.W., Palmisano, J.F., Levin, J.E., 1993. Oregon’s wild salmon and steelhead trout: A review of
the impact of management and environmental factors. Oregon Forest Industries Council, Salem, Oregon.
Lee, D.J., Putnam, G.B., 1973. The response of rainbow trout to varying protein/energy ratios in a test diet. J.
Nutr. 103, 916-922.
Ludwig, B.W., 1980. A morphological and biochemical comparison of artificially and naturally-reared
salmonids. M.Sc. thesis. University of British Columbia, Vancouver, Canada.
Makinen, T., 1993. Effect of temperature. feed ration and other factors on the growth of rainbow trout
(Oncorhvnchus mykiss Walbaum 1792) cultured in Finland. Finnish Fish. Res. 15, 39-64.
McIntyre. J.D., 1987. Smolt quality and the effectiveness of Columbia River hatcheries. Workshop Proceed-
ings: Improving Hatchery Effectiveness as Related to Smoltification, May 1985. US Dept. of Energy, BPA
Project No. 84-52. Portland, OR.
Metcalfe, N.B., Thorpe, J.E., 1992. Anorexia and defended energy levels in over-wintering juvenile salmon. J.
Animal Ecol. 61, 175-181.
Miller, W.H., Coley, T.C. Burge, H.L., Kisanuki, T.T., Steward, C.R., Bjornn, T.C., 1990. Analysis of salmon
and steelhead supplementation. US Dept. of Energy, BPA Project No. 88.100. Portland, OR.
NRC (National Research Council), 1996. Upstream: Salmon and Society in the Pacific Northwest. National
Academy Press, Washington, DC.
Ogata, H., Konno. S., 1986. Growth response and smelt production of 1 year cherry salmon fed diets having
different protein and lipid levels. Bull. Japn. Sot. Sci. Fish. 52, 3 13-3 18.
Peter, R.E., 1979. The brain and feeding behavior. In: Hoar, W.S., Randall, D.J., Brett, J.R. (Eds.), Fish
Physiology. Academic Press, New York, Vol. VIII, pp. 121-159.
Plotnikoff, M.D., Higgs, D.A., Markert, J.R., Dosanjh, S., McBride, J., Buckley, J.T., 1983. Nutrition and
marine survival of chinook salmon (Oncorhynchus tshawytschn): I. Potential role of smolt body composi-
tion (Robertson Creek Hatchery 1980 brood). Can. Tech. Rept. Fish. Aquat. Sci., No. 1206.
Plotnikoff, M.D., Higgs, D.A., Markert, J.R., Dosanjh, S., McBride, J., Buckley, J.T., 1984. Nutrition and
marine survival of chinook salmon (Oncorhyzchus tshawytscha): II. Further investigation of the potential
role of smolt body composition (Robertson Creek Hatchery 1980 brood). Can. Tech. Rept. Fish. Aquat.
Sci., No. 1235.
Raymond, H.L., 1988. Effects of hydroelectric development and fisheries enhancement on spring and summer
chinook salmon and steelhead in the Columbia River Basin, N. Amer. J. Fish. Mgmt. 8, l-24.
Reinitr, G.L., 1983. Relative effect of age, diet, and feeding rate on the body composition of young rainbow
trout (S&no ~nirdneri). Aquaculture 35, 19-27. Shearer, K.D., 1994. Factors affecting the proximate composition of cultured fishes with emphasis on
salmonida. Aquaculture 119, 63-88.
Shearer, K.D., Silverstein, J.T., Plisetskaya, E.M., (in press). The role of adiposity in food intake control of
juvenile chinook salmon (Oncorhwchus tshawytscha). Comp. Physiol. Biochem.
Shrimpton, J.M., Bernier, N., Jiwama, G.K., Randall, D.J., 1994. Differences in measurement of smolt
development between wild and hatchery-reared juvenile coho salmon (Oncorhynchus kisutch) before and
after seawater exposure. Can. J. Fish. Aquat. Sci. 51, 2170-2178.
Silver, G.R., Higgs, D.A., Dosanjh, B.S., Mckeown, B.A., Deacon, G., French, D., 1993. Effect of dietary
protein to lipid ratio on growth and chemical composition of chinook salmon (0ncorhpchu.s tshawytscha)
in sea water. In: Kaushik, S.J., Luquet, P. (Eds.), Fish Nutrition in Practice, IVth International Symposium on Fish Nutrition and Feeding, 24-27 June 1991, Biarritz, France. Institut National de la Recherche
Agronomique, Paris (Les Colloques no. 61). pp. 459-468. Silverstein, J.T.. Shimma, H., Ogata, H., 1997. Early maturation in amago salmon: an association with energy
storage. Can. J. Fish. Aquat. Sci. 54, 444-451. Statview, 1992. Statview 4.02, Abacus Concepts, Berkeley, CA.
Storebakken, T., Hung, S.S.O., Calve& C.C., Plisetsksya, E.M., 1991. Nutrient partitioning in rainbow trout at
different feeding rates. Aquaculture 96, 191-203.
SuperANOVA, 1990. SuperANOVA, Abacus Concepts, Berkeley, CA. Takeuchi, T., Watanabe, T., Ogino, C., 1978. Supplementary effect of lipids in a high protein diet of rainbow
trout. Bull. Japn. Sot. Sci. Fish. 44, 677-681.

K.D. Shearer et al. /Aquaculture 157 (1997) 31 l-323 323
Thorpe, J.E., 1986. Age at first maturity in Atlantic salmon, Salmo s&r, freshwater period influences and
conflicts with smolting. Can. Spec. Pub. Fish. Aquat. Sci. 89. 7-14.
Vahl, 0.. 1979. A hypothesis on the control of food intake in fish. Aquaculture 17, 221-229.
Virtanen, E., Soderholm-Tana, L., Soivio, A., Forsman, L., Muona, M., 1991. Effect of physiological
condition and smoltification status at smelt release on subsequent catches of adult salmon. Aquaculture 97,
23 I-257.
Wathne, E., 1995. Strategies for directing slaughter quality of farmed Atlantic salmon (S&no s&r) with
emphasis on diet composition and fat deposition. Ph.D. thesis. Agricultural University of Norway. As,
Norway.
Wood, E.M., Yasutake, W.T., Woodall, A.N., Halver, J.E., 1957. The nutrition of salmonid fishes: I. Chemical
and histological studies of wild and domestic fish. J. Nutr. 61, 465-478.
Wood, E.M., Yasutake, W.T., Halver, J.E., Woodall, A.N., 1960. Chemical and histological studies of wild
and hatchery salmon in fresh water. Trans. Amer. Fish. Sot. 89, 301-307.
Wurtsbaugh, W.A., Davis, G.E.. 1977. Effects of temperature and ration level on growth and food conversion
efficiency of Salmo gairdneri Richardson. J. Fish Biol. I I, 87-98.
Zar, J.H., 1974. Biostatistical Analysis. Prentice-Hall, Englewood Cliffs, NJ.





























