Embed Size (px)
Citation preview

Experimental Neurology 177, 461–474 (2002)doi:10.1006/exnr.2002.8006
Aberrant Sprouting and Downregulation of Tyrosine Hydroxylase inLesioned Nigrostriatal Dopamine Neurons Induced by Long-Lasting
Overexpression of Glial Cell Line Derived Neurotrophic Factorin the Striatum by Lentiviral Gene Transfer
Biljana Georgievska,1 Deniz Kirik, and Anders BjorklundWallenberg Neuroscience Center, Department of Physiological Sciences, Lund University, BMC A11, 221 84 Lund, Sweden
Received March 19, 2002; accepted June 19, 2002
The effects of sustained (up to 9 months) striataloverexpression of glial cell line derived neurotrophicfactor (GDNF) on lesioned nigrostriatal dopamine(DA) neurons was studied using a recombinant lenti-viral (rLV) vector to deliver GDNF into the striatum 4weeks prior to the creation of an intrastriatal 6-hy-droxydopamine lesion. The results of the amphet-amine-induced rotation suggested an initial partialprotection followed by a complete recovery, whereasthe spontaneous motor behaviors remained impaired.There was a clear protection of the nigral tyrosinehydroxylase (TH)-positive neurons in the rLV-GDNFgroup compared to rats injected with the control vec-tor encoding green fluorescent protein (GFP) (70 and20% of the intact side, respectively). However, the stri-atal TH� fiber density was equally reduced (to 20% ofthe intact side) in both groups. Further morphologicalanalyses indicated that the nigrostriatal projections ofthe DA neurons were indeed preserved in the GDNFgroup. The axonal projections were visualized usingtwo independent methods: First, retrograde labelingof the nigral cell bodies by intrastriatal Fluoro-Goldinjections showed that the majority of rescued cells inthe GDNF group had preserved axonal projections tostriatum. Second, injections of a recombinant adeno-associated viral vector expressing GFP into the nigrawas used to anterogradely fill the DA neurons andtheir projections with GFP protein. GFP immuno-staining clearly demonstrated that the fibers of thenigral DA cells were preserved along the nigrostriatalpathway and innervated large parts of the striatum,but did not express TH at detectable levels. In addi-tion, fiber sprouting was observed in the globus palli-dus, entopeduncular nucleus, and substantia nigra,corresponding to areas where GDNF protein was re-leased. The lack of functional recovery in the sponta-neous motor behaviors may, at least in part, be ex-
1 To whom correspondence should be addressed. Fax: �46 46 22205 61. E-mail: [email protected].
461
plained by this extensive aberrant fiber sprouting inthe downstream striatal target nuclei and/or de-creased synthesis of dopamine in the striatum.© 2002 Elsevier Science (USA)
Key Words: behavior; Parkinson’s disease; sprouting;stereology.
INTRODUCTION
Recombinant viral vectors offer powerful tools forlong-term local delivery of glial cell line derived neuro-trophic factor (GDNF) in the brain. In rodent and pri-mate models of Parkinson’s disease (PD) overexpres-sion of GDNF in the striatum and/or substantia nigra(SN) by transfer of the GDNF gene by recombinantlentiviral vector (rLV), recombinant adeno-associatedvirus (rAAV), or adenovirus (Ad) can block the toxin-induced degeneration of the nigral dopamine (DA) neu-rons (1, 2, 7, 10, 18, 21, 33). Injections of the vector inthe striatum, in particular, are effective not only inrescuing the DA cell bodies in the SN, but also inpreserving their axons projecting to the striatum (12,18). In a particularly interesting study, Kordower et al.(21) have shown that rLV-GDNF delivery in the stria-tum and SN can prevent nigrostriatal degenerationand reverse motor deficits in monkeys subjected toMPTP treatment 1 week prior to vector injection.Based on these promising results intracerebral GDNFdelivery by recombinant viral vectors is now beingseriously considered for clinical application in PD pa-tients (5).
However, GDNF delivery by viral vectors is stillhighly experimental. In particular, we need to learnmore about the functional consequences of intracere-bral GDNF delivery and define the conditions (dose,timing, site of transduction) for optimal neuroprotec-tion and functional recovery. Previous studies usingintracerebral administration of recombinant GDNFprotein have shown that GDNF can exert at least three
0014-4886/02 $35.00© 2002 Elsevier Science (USA)
All rights reserved.

distinct effects on degenerating nigral DA neurons: (i)protection of the DA neurons against a toxic or me-chanical insult; (ii) stimulation of axonal sprouting andregrowth from lesioned nigral DA neurons; and (iii)stimulation of DA turnover and function in neuronsspared by the lesion (3, 11, 17, 19, 23, 34). Availabledata suggest that these effects may be, at least in part,independent of each other and may be induced by dif-ferent mechanisms, at different sites, at different timepoints after injury, and at different dose levels ofGDNF. In the 6-hydroxydopamine (6-OHDA) lesionmodel near-complete protection of nigral DA neuronsmay be obtained in the absence of any functional re-covery (17, 34, 40), and GDNF-induced functional re-covery may be obtained in the absence of any directneuroprotective effects (see, e.g., (19, 31)). Similarly, inMPTP-treated monkeys, Kordower et al. (21) observeda highly variable degree of functional recovery afterrLV-GDNF treatment, despite the fact that all treatedanimals exhibited complete protection of the cell bodiesof the nigral DA neurons.
The purpose of the present study was to investigatethe long-term functional consequences of rLV-GDNFmediated gene transfer to the striatum in rats sub-jected to an intrastriatal 6-OHDA lesion. This type oflesion induces a partial and progressive degenerationof the nigrostriatal DA system and marked impair-ments of motor function that are stable over manymonths (16, 20). The results show that the nigral DAneurons and their projections to the striatum were wellpreserved in the rLV-GDNF treated animals, that thestriatal innervation was restored, and that amphet-amine-induced rotation was reversed. Unexpectedly,however, we observed a marked downregulation of thetyrosine hydroxylase (TH) enzyme in large areas of theGDNF transduced striatum and an extensive sprout-ing of TH-positive fibers in downstream striatal tar-gets, most prominently in the globus pallidus, entope-duncular nucleus, and SN, and no functional recoveryin the spontaneous motor tests. These results raise thepossibility that aberrant sprouting induced by long-term GDNF delivery may be functionally detrimentalto the rescued nigrostriatal neurons.
MATERIALS AND METHODS
Subjects
A total of 100 adult female Sprague–Dawley rats(B&K Universal, Stockholm, Sweden) were used in thisstudy. The animals were housed four to five per cageunder a 12-h light–dark cycle with free access to ratchow and water, except when the staircase test wasperformed during which the food consumption was lim-ited to 10–12 g/rat/day. The housing and treatment ofthe animals was conducted according to the rules set
by the Ethical Committee for Use of Laboratory Ani-mals at Lund University.
Experimental Design
All animals received injections of the rLV vectorencoding either GDNF (n � 59) or green fluorescentprotein (GFP) (n � 41) into the striatum. Four weeksafter vector injection, 63 animals received injections of6-OHDA into the striatum. The lesioned animals wererepeatedly tested on a battery of drug-induced andspontaneous motor behaviors as indicated by the timeline given in Fig. 1. Twenty-four rats were killed at 6weeks postlesion for morphological evaluation. The re-maining animals were followed for an additional 26weeks with behavioral tests before being divided intotwo subgroups for anatomical tracing of the nigrostri-atal DA system. In one subgroup (n � 21), Fluoro-Gold(FG) was injected into the striatum in order to retro-gradely label the nigral cell bodies that were still con-nected to the striatum. The animals were killed 7 dayslater (i.e., at 33 weeks postlesion). In a second sub-group (n � 18), rAAV-GFP was injected into the SNfor the purpose of anterogradely filling the nigral DAneurons and their projections to the striatum with GFPprotein. These animals survived 4 weeks after trans-duction (36 weeks postlesion) to allow the GFP proteinto fill the entire processes in the transduced cells. Theintact vector-injected animals were killed 6 (n � 19)and 32 weeks (n � 18) after vector injection for thedetermination of tissue levels of GDNF protein.
Recombinant Lentiviral Vector Production
The rLV-CMV-GFP-W and rLV-CMV-GDNF-W vec-tor constructs were generated as described previously(33). In the present study, the packaging construct,pCMV�R8.91, and the vesicular stomatitis virus Gprotein envelope plasmid, pMD.G (42), were used forlentiviral vector production essentially according to theprocedures described by Zufferey et al. (42). Briefly, thetransfer construct and the helper plasmids were co-transfected into 293T cells and virions released intothe medium were collected at 48 and 72 h posttrans-fection. To concentrate the viral particles, the mediumwas ultracentrifuged for 1.5 h at 26,000 rpm and thepellet dissolved in DMEM, 10% FBS, 1% glutamine.The number of transducing units (TU) of the concen-trated rLV-GFP vector stock was determined on 293Tcells. The cells were plated at a density of 1 � 105 cellsper well in six-well tissue culture dishes. Serial dilu-tions of the vector stock were added and the number ofGFP-expressing cells was analyzed 48 h later. An RNAslot blot technique (38) was subsequently used to de-termine viral particle titer for both the GFP and theGDNF vector stocks. The number of TU of the GDNFexpressing vector was estimated based on the ratiobetween viral particle titer and TU as determined for
462 GEORGIEVSKA, KIRIK, AND BJORKLUND

the rLV-GFP vector. The final titer of the rLV-GFPvector stock was 6.8 � 108 TU/ml and the titer of therLV-GDNF vector stock was estimated at 3.2 � 108
TU/ml. Both vector stocks were diluted fivefold in sa-line before injection.
Surgical Procedures
For all surgical procedures animals received halo-thane anesthesia. All injections were made using a10-�l Hamilton syringe fitted with a glass micropipette(outer diameter � 60–80 �m) using a Kopf stereotaxicframe (Kopf Instruments, Tujunga, CA). The antero-posterior and mediolateral coordinates were calculatedfrom bregma and the dorsoventral coordinates fromthe dural surface.
rLV vector injections. The animals received injec-tions of a total of 3 �l rLV-GDNF (n � 59) or rLV-GFP(n � 41) into the right striatum. The injections weremade as two 0.5-�l deposits, along each of the threeneedle tracts at the following coordinates: (1) AP ��1.4, ML � �2.6, DV � �5.0,�4.0; (2) AP � �0.4,ML � �3.4, DV � �5.0, �4.0; and (3) AP � �0.8,ML � �4.4, DV � �5.0, �4.0. The tooth bar was set at0.0. The injection rate was 1 �l/min and after theinfusion the glass pipette was slowly retracted 1 mmand left in place for an additional 3 min before beingwithdrawn.
6-OHDA lesions. Four weeks after vector injection,63 animals received unilateral injections of a total of 21�g 6-OHDA (calculated as free base; Sigma, St. Louis,MO) dissolved in ascorbate–saline (0.05%), divided intothree 7-�g deposits in the right striatum, according toKirik et al. (16). The injections were made at the fol-lowing coordinates: (1) AP � �1.0, ML � �3.0, DV ��5.0; (2) AP � �0.1, ML � �3.7, DV � �5.0; and (3)AP � �1.2, ML � �4.5, DV � �5.0 and the tooth barwas set at 0.0. The injection rate was 1 �l/min and theglass pipette was left in place for an additional 3 minbefore being withdrawn.
FG injections. Retrograde labeling of the nigrostri-atal neurons was performed by injection of FG into thestriatum at 32 weeks after the 6-OHDA lesion. Twen-ty-one of the 6-OHDA lesioned animals received bilat-eral intrastriatal injections of 0.2 �l of a 2% FG solu-tion (Fluorochrome, Inc., Englewood, CO) dissolved inphysiological saline. The injection coordinates wereAP � �1.0, ML � �3.0, DV � �4.5, with the tooth barset to 0.0. The injection rate was 0.1 �l/min and theglass pipette was left in place for an additional 10 minbefore being withdrawn.
rAAV-GFP injections. The rAAV-CBA-GFP vector(kind gift of Dr. Ronald J. Mandel, Univ. of Florida)was injected unilaterally (n � 12) or bilaterally (n �6) over the substantia nigra at 32 weeks after the6-OHDA lesion. The rAAV-CBA-GFP vector (1.4 � 1011
infectious units/ml) was prepared and titered as de-
scribed elsewhere (9, 27, 41). The animals received asingle 2-�l injection of the vector at coordinates AP ��5.3, ML � �2.0, DV � �7.2, with the tooth bar set to�2.3. The injection rate was 0.5 �l/min and the glasspipette was left in place for an additional 5 min beforebeing withdrawn.
Behavioral Tests
Drug-induced rotation. Drug-induced rotation wasassessed in automated rotometer bowls (37) using D-amphetamine sulfate (2.5 mg/kg, ip; Apoteksbolaget,Lund, Sweden) or apomorphine–HCl (0.25 mg/kg, sc;Sigma–RBI, St. Louis, MO) and monitored for 90 and40 min, respectively. Rotational asymmetry scores areexpressed as net 360° turns/min and ipsilateral rota-tions (i.e., toward the injected side) were assigned apositive value.
The cylinder test. The cylinder test (35) was per-formed at the beginning of the rat’s active (dark) periodas described in Kirik et al. (18). In brief, forelimb place-ment and use during explorative activity was analyzedby videotaping the animal in a clear glass cylinderwhere it was allowed to move freely. Mirrors wereplaced behind the cylinder at an angle to permit re-cording of forelimb movements whenever the animalwas turned away from the camera. The animal was leftin the cylinder until it had performed at least 20touches on the cylinder wall. Forelimb touches werescored by an observer, blinded to the animals’ identi-ties, using a video recorder with slow motion andframe-by-frame capabilities to score the movements.The number of total contacts was counted to a mini-mum of 20 and the percentage of left touches of totalcontacts was calculated.
Stepping (forelimb akinesia) test. The animals weretested for movement initiations using the stepping test(as described (29)). Briefly, the animal was held by theexperimenter fixing the hindlimbs with one hand andholding the forelimb not to be tested with the otherhand, while the animal supported its weight on theunrestrained forelimb. The animal was allowed to ini-tiate stepping movements while being moved sidewaysalong the table surface (90 cm in 4–5 s) in the forehanddirection. This procedure was repeated twice for eachforelimb and the animals were tested on 4 consecutivedays. The mean of the data from the last 3 days con-stituted the final value.
Staircase test. A modified version of the staircasetest described by Montoya et al. (28) was used. Theanimals were food deprived for 2 days before the startof testing. The animals were placed in Plexiglas testboxes and allowed to retrieve pellets for 15 min on 8consecutive days. Forty pellets were placed on eachside of the staircase (10 pellets on each of the fourdifferent levels). After each test the number of pelletstaken and missed was counted on each side. The dif-
463GDNF INDUCES DOWNREGULATION OF TYROSINE HYDROXYLASE

ference between the two counts was considered pelletseaten.
GDNF Determination by ELISA
GDNF protein levels were determined by ELISA onstriatal tissue samples from the intact animals at 6weeks (rLV-GDNF, n � 9; rLV-GFP, n � 10) and 32weeks after the vector injections (rLV-GDNF, n � 9;rLV-GFP, n � 9). The brains were rapidly removedand the striatum was dissected bilaterally, dorsal tothe anterior commissure, and freed from the cortex andseptum. In addition, a 2-mm-diameter punch centeredon the SN was taken from a 3-mm-thick coronal slice.
The tissues from the striatum and SN were soni-cated in a homogenization buffer (50 mM Tris, pH 7.4,150 mM NaCl, 1% Triton X-100, 1.7 �g/ml phenylmeth-ylsulfonyl fluoride, 1.0 �g/ml leupeptin, 10 �g/ml apro-tinin, and 1.0 �g/ml pepstatin) at a tissue concentra-tion of 30 mg/ml. Tissue levels of GDNF were deter-mined on homogenates using ELISA according to thesupplier’s recommendations (G3240, Promega, Madi-son, WI).
Histological Analysis
The animals were sacrificed at 6 weeks (rLV-GDNF,n � 15; and rLV-GFP, n � 9), 33 weeks (FG-injectedrats; rLV-GDNF, n � 14; and rLV-GFP, n � 7), and 36weeks (AAV-GFP-injected rats; rLV-GDNF, n � 12;and rLV-GFP, n � 6) after the 6-OHDA injections. Theanimals were deeply anesthetized with sodium pento-barbital and transcardially perfused with 50 ml of iso-tonic saline followed by 250 ml ice-cold 4% PFA in 0.1M phosphate buffer (PB), pH 7.4. The brains weredissected and postfixed in the same fixative for 2 h andtransferred into 25% sucrose in 0.1 M PB. A freezingmicrotome was used to obtain 40-�m-thick sections,divided into eight series at the level of the striatum andfive series for the midbrain. Immunohistochemistrywas performed as described previously (18) using an-tibodies raised against GDNF (goat IgG, 1:2000; R&DSystems, Minneapolis, MN), GFP (chicken IgG, 1:5000;Chemicon, Temecula, CA), tyrosine hydroxylase (mouseIgG, 1:2000; Chemicon), and vesicular monoamine trans-porter-2 (VMAT, rabbit IgG, 1:2000; Chemicon). The sec-tions were first quenched for 10 min in 3% H2O2, 10%methanol in potassium–phosphate buffer (KPBS), pre-incubated in 5% serum, 0.25% Triton, and incubatedovernight at room temperature with the primary anti-body. This was followed by incubation with 1:200 dilu-tion of biotinylated secondary horse anti-goat (BA9500,Vector Laboratories, Burlingame, CA), rabbit-anti-chicken (G2891, Promega), horse anti-mouse (BA2001,Vector Laboratories), and goat anti-rabbit (BA1000,Vector Laboratories), respectively. The reaction prod-uct was visualized with avidin–biotin–peroxidase com-plex (ABC Elite, Vector Laboratories) using 3�,3�-dia-
minobenzidine as a chromogen. The sections weremounted on chrom–alum-coated glass slides, dehy-drated in ascending alcohol solutions, cleared in xy-lene, and coverslipped with Depex mounting medium.
Immunofluorescence staining was performed usingthe primary antibody against TH as above. The sec-tions were incubated overnight in the primary anti-sera, 5% serum, 0.25% Triton in KPBS. This was fol-lowed by incubation with a biotinylated horse anti-mouse antibody (see above) and Cy3-conjugatedstreptavidin (1:200, Jackson Immunoresearch Labora-tories, Inc., West Grove, PA). The sections weremounted and coverslipped with PVA-DABCO. TH ex-pression was visualized using the green light and na-tive fluorescence of GFP was seen in the blue light.
From the FG-injected animals, one series of sectionswas directly mounted on glass slides and coverslippedwith PVA-DABCO.
Morphometric Analysis
Stereological estimation of the total numbers of TH-positive cells in the SN. Stereological evaluation wasperformed using the optical fractionator (39) as de-scribed previously (16). The sections used for counting(9–11 sections from 1/5 series) covered the entire SN,from the rostral tip of the pars compacta back to thecaudal end of the pars reticulata. The medial border ofthe SN was defined by a vertical line passing throughthe medial tip of the cerebral peduncle and by themedial terminal nucleus of the accessory nucleus of theoptic tract (MTN), when present in the sections,thereby excluding the TH-positive cells in the ventraltegmental area. The ventral border followed the dorsalborder of the cerebral peduncle, including the TH-pos-itive cells in the pars reticulata, and the area extendedlaterally to include the pars lateralis in addition to thepars compacta. Sampling was performed using theOlympus C.A.S.T.-Grid system (Olympus DenmarkA/S, Albertslund, Denmark). A counting frame (4357�m2) was placed randomly over the first counting areaand systematically moved through all counting areas.Quantification of TH-positive perikarya was performedusing a 100X oil objective and the total number ofTH-positive cells was estimated according to the opti-cal fractionator formula (39).
FG-positive cells. The number of FG-positive cellsin the SN pars compacta was assessed in three sectionscentered around the level of the MTN. All FG-positivecells lateral to the MTN were counted at 20X objectivemagnification.
Striatal fiber density measurements. These wereassessed by measuring the optical density (OD) of theTH-positive fibers in the striatum using the NIH 1.62Image program on a Macintosh 9500 computer con-nected to a digital ProgRes camera (Kontron Elek-tronik, Germany) and a constant illumination table.
464 GEORGIEVSKA, KIRIK, AND BJORKLUND

For each animal the OD was measured from the entirecross-section of the striatum at three rostrocaudal lev-els corresponding to AP � �1.0, AP � �0.26, and AP ��0.92, relative to bregma, according to the atlas ofPaxinos and Watson (30). The readings were correctedfor nonspecific background by measuring the corpuscallosum in each section and expressed as a percentageof control side value.
Statistical Analysis
Differences between treatments were assessed usinganalysis of variance and were followed, when signifi-cant, by Student–Newman–Keuls post hoc analysis forgroup comparisons. Significance was accepted at the95% probability level.
RESULTS
GDNF Provides Recovery of Lesion-Induced MotorAsymmetry but Not Spontaneous Motor Behavior
The effect of striatal GDNF overexpression on motorasymmetry in intact animals was assessed at 3 weeksafter the rLV vector injections, just prior to the6-OHDA lesion. The rLV-GDNF treated animals dis-played an enhanced amphetamine-induced contralat-eral turning bias (5.3 � 0.4 turns/min), which did notoccur in the rLV-GFP injected control group (0.1 � 0.5),indicating an upregulation of the DA function on theGDNF-transduced side.
Beginning 4 weeks after the 6-OHDA lesion, theanimals were tested on a battery of drug-induced andspontaneous motor tests over several months (Fig. 1).In the rLV-GFP control group, the 6-OHDA lesion in-duced severe behavioral impairments. In the drug-in-duced rotation tests the control animals displayed asubstantial rotational asymmetry in response to am-phetamine (10.6–13.3 ipsilateral turns/min) and apo-morphine (5.3–7.0 contralateral turns/min) at all timepoints tested (Figs. 2A and 2B). In the tests of sponta-neous motor behavior (cylinder, stepping, and stair-case tests) the control animals showed significant im-pairments on the left side, i.e., the side contralateral tothe lesion (P 0.05 for all three tests). In the cylindertest (Fig. 2C), the left paw use was reduced to 15–25%of total touches. In the staircase test (Fig. 2D), thecontrol animals were able to retrieve on average 17pellets with the left paw compared to 28 pellets withthe right paw. In the stepping test (Fig. 2E), the controlanimals performed on average 1–2 steps with the leftforelimb compared to 10–12 steps with the right fore-limb.
The rLV-GDNF animals showed reduced amphet-amine-induced rotation already at 4 weeks postlesion(6.0 � 1.7 turns/min), suggesting an initial protectionagainst the 6-OHDA lesion. Rotational behavior wasfurther reduced at 10 and 19 weeks, when the asym-metry was completely abolished (1.0 � 0.4 turns/min at19 weeks; Fig. 2A). In the apomorphine rotation test,an average of about 5 turns/min was observed in the
FIG. 1. The animals received injections of the rLV vectors 4 weeks before the 6-OHDA lesion. The behavioral effects of GDNF expressionin the intact dopaminergic system were evaluated using amphetamine-induced motor asymmetry (amph-pre) at 3 weeks after vector injection(�1 week on the time line). 6-OHDA was injected in the striatum at week 0. Postlesion motor impairments were evaluated using repeateddrug-induced rotation tests (amph I–III and apo I–III; shown above the time line) and spontaneous motor tests (cylinder I–III, stepping I–IV,and staircase tests; below the time line). At 6 weeks postlesion a group of animals (n � 24) was killed for morphological evaluation(short-term survival). The motor performance of the remaining animals was monitored until 28 weeks postlesion, as shown on the time line.In order to trace the nigrostriatal DA system, the remaining animals were divided into two groups at 32 weeks after the lesion. One groupwas injected with FG in the striatum, while the second group received injections of rAAV-GFP in the SN. The FG injected and rAAV-GFPinjected animals were killed at weeks 33 and 36 after the lesion, respectively (long-term survival).
465GDNF INDUCES DOWNREGULATION OF TYROSINE HYDROXYLASE

GDNF animals at all time points, which was not sig-nificantly different from the rLV-GFP controls (Fig.2B). In the tests of spontaneous motor behavior, theGDNF animals performed 23–37% left touches in thecylinder test, retrieved on average 22 pellets in thestaircase test, and were able to perform 2–3 steps withthe left forelimb in the stepping test at the differenttesting periods (filled circles and filled bars in Figs.2C–2E). Although these values suggested a trend to-ward better performance in the rLV-GDNF group com-pared to the control group, the differences did not reachstatistical significance (P 0.11 for all contrasts).
Lesioned Nigral DA Neurons Are Rescued by GDNFand Remain Connected to the Striatum
The protection of the nigral DA neurons was evalu-ated by TH immunohistochemistry at 6 and 36 weeksafter the 6-OHDA lesion. In the control rLV-GFPgroup, the intrastriatal 6-OHDA lesion induced a 70–80% reduction in the number of TH-positive cells in theSN pars compacta. rLV-mediated GDNF expression inthe striatum had significant neuroprotective effects onthe TH-positive cells in the SN, amounting to 77% of
intact side at 6 weeks and 65% at 36 weeks (Fig. 3A).By contrast, densitometry measurements showed thatthe striatal TH-positive innervation was reduced by61–76% in the rLV-GDNF group at both time points(Fig. 3B). In fact, at 6 weeks after the 6-OHDA lesion,the striatal TH-positive innervation in the rLV-GDNFgroup (24% of intact side) was significantly lower com-pared with that of the rLV-GFP control group (36% ofintact side). At 36 weeks the density of the striatalTH-positive innervation was similar in both groups(39% vs 32% of the intact side in the rLV-GDNF andrLV-GFP groups, respectively).
At 32 weeks after the lesion one group of animalsreceived bilateral injections of the retrograde tracer FGinto the striatum, and the animals were killed 1 weeklater. The number of FG-positive cells in the SN at 33weeks postlesion was 59% of the intact side in therLV-GDNF group, compared to 23% of the intact sidein the control group (Fig. 4, inset; compare B and C),while in adjacent sections, the numbers of TH-positivecells was 50% in the GDNF group and 22% in thecontrol group (Fig. 4, inset). The close match betweenthe FG and TH-positive cell numbers indicates that the
FIG. 2. Motor performance of the animals in the drug-induced and spontaneous motor tests. (A) The amphetamine-induced rotation wasperformed at 4, 10, and 19 weeks after the 6-OHDA lesion. Already at 4 weeks after the lesion, the rLV-GDNF group displayed reducedrotations compared to the rLV-GFP control group. The rotational asymmetry further declined at 10 and 19 weeks until the side bias wascompletely lost in the rLV-GDNF injected rats. (B) In the apomorphine-induced rotation test, both groups showed a similar numbers of turnsat 4 weeks after the lesion and no significant reduction was observed in the rLV-GDNF group at 10 and 19 weeks postlesion. (C–E) Motorperformance of the forelimbs was assessed using three spontaneous tests: cylinder test (C), staircase test (D), and stepping test (E). Inall three tests, the animals showed an impairment in their left paws and there was no significant difference between the rLV-GFP andthe rLV-GDNF group at any time point (P � 0.30 for the cylinder test; P � 0.11 for the staircase test; P � 0.16 for the steppingtest). *Significant difference from the rLV-GFP group; #Significant difference from earlier time points within the rLV-GDNF group;†Significant difference between the left and right paws.
466 GEORGIEVSKA, KIRIK, AND BJORKLUND

axonal projections to the striatum were to a large ex-tent preserved in the rescued nigral cells in the rLV-GDNF treated animals.
Overexpression of GDNF in the Striatum InducesDownregulation of TH in the Lesioned NigrostriatalSystem
To label the nigral DA neurons and their projectionsto the striatum anterogradely with a marker indepen-dent of TH, an rAAV vector encoding GFP was injectedover the SN in one group of animals at 32 weeks afterthe 6-OHDA lesion. We have previously shown that therAAV vector has a high transduction efficiency in theSN and preferentially transduces the nigral DA neu-rons in the pars compacta (4). The rAAV transducedanimals were killed 4 weeks after the injection to allowsufficient time for the GFP gene to be fully expressed.On the contralateral intact side, transgenic GFP pro-tein (detected by immunohistochemistry) was highlyexpressed in the nigral DA neurons in SN pars com-pacta and their dendrites in SN pars reticulata (Fig. 5,compare A and B) and was transported anterogradelyin the axons, filling out virtually the entire nigrostria-tal pathway and its axonal branches into the entope-duncular nucleus (Fig. 6, compare A and B), globuspallidus (Fig. 7, compare A and B), and the striatum(Fig. 8, compare A and B, for further details, see Bjork-lund et al. (4)). In the rLV-GFP control group, nearly80% of the nigral DA neurons were lost after the6-OHDA lesion, thereby limiting the number of nigral
FIG. 3. (A) The number of TH-positive cells in SN was estimatedusing stereology and expressed as a percentage of the intact side. Inthe rLV-GFP control group there was a 70–80% cell loss at 6 and 36weeks after the 6-OHDA lesion. In the rLV-GDNF group, 77 and 65%of the nigral TH-positive cells were rescued from the lesion at 6 and36 weeks postlesion, respectively. (B) TH-positive striatal fiber den-sity was measured at three rostrocaudal levels and the average datawere expressed as a percentage of the intact side. In the rLV-GFPcontrol group, the TH-positive innervation was reduced to about30–35% at both time points (6 and 36 weeks postlesion). In therLV-GDNF group, the striatal TH-positive innervation was signifi-cantly reduced at 6 weeks (24%) compared to the rLV-GFP group(P 0.05), whereas at 36 weeks postlesion the fiber density was nolonger significantly different from the control group (39%).
FIG. 4. Photomicrographs from the ventral midbrain showing FG-positive cells retrogradely labeled in the SN at 33 weeks after the6-OHDA lesion. The number of FG-positive cells in the rLV-GFP group was greatly reduced to about 20% (B), whereas in the rLV-GDNFtreated animals (C), there was a larger proportion of cells labeled with FG to about 60% compared with the intact side (A). The inset showsthe quantification of the number of FG-labeled cells and the number of TH-positive cells counted in adjacent sections in the SN. Note the closematch between the FG- and TH-positive cell numbers in the rLV-GDNF group, suggesting that the majority of the rescued cells wereconnected to the striatum. The scale bar in A represents 250 �m.
467GDNF INDUCES DOWNREGULATION OF TYROSINE HYDROXYLASE

DA neurons that could be transduced with the rAAV-GFP vector (Fig. 5, compare C and D). The TH-positiveinnervation was restricted to the medial parts of therostral striatum (Fig. 8C), while the lateral (Fig. 8C)and caudal parts (Fig. 7C) were completely denervated.Similarly, GFP-immunoreactive fibers were seen to belimited to the medial parts of rostral striatum (Fig.8D), while the caudal part was completely devoid ofGFP-positive fibers (Fig. 7D). In the rLV-GDNFtreated animals, by contrast, the GFP-positive inner-vation was found to cover large portions of the lateraland medial parts, as well as caudal parts of the stria-tum. This pattern did not match with the TH-positiveinnervation as observed in adjacent sections (Fig. 7,compare E and F; Fig. 8, compare E and F), demon-strating that the fibers were indeed present in thestriatum but did not express TH. The downregulationof TH was particularly apparent in the lateral anddorsal parts of the rostral striatum (Fig. 8E) and in the
caudal striatum (Fig. 7E), i.e., in regions that showed ahigh level of GDNF expression in the GDNF-immuno-stained sections (Fig. 9). The dissociation between theexpression of TH and GFP proteins in the striatal DAterminals was further confirmed using immunohist-ofluorescence for visualization of TH (red) and nativefluorescence of GFP (green) in the same sections. GFP-containing fibers in the medial striatum were found tobe colabeled with the TH antibody (Fig. 8G�–8G��),while the GFP-positive fibers in the lateral part ofstriatum were devoid of TH (Fig. 8H�–8H��). In con-trast to the downregulation of TH, expression of VMATwas maintained in the preserved striatal fibers in theGDNF treated animals. Inspection of adjacent sectionsstained for TH and VMAT showed that the striatalfibers stained for VMAT at a level similar to that on thecontralateral intact side, also in areas that were seem-ingly devoid of TH-positive terminals (Fig. 7, compareG and H).
FIG. 5. Photomicrographs of adjacent sections from substantia nigra (SN) at 36 weeks after the 6-OHDA lesion (rAAV-GFP injectedanimals), immunostained for TH and GFP. Injections of rAAV-GFP in the intact SN efficiently transduced the nigral DA neurons (compareA and B). In the rLV-GFP injected controls, the intrastriatal 6-OHDA lesion induced an extensive loss of nigral TH-positive neurons (C),thereby limiting the number of neurons transduced by the rAAV-GFP vector (D). In the rLV-GDNF treated animals, there was a substantialprotection of the nigral TH-positive neurons (E) and intense axonal sprouting was observed in the SN (arrows in E and F). In the intact SN,the dendrites of the DA neurons extending into the reticulata (SNr) are seen as thick processes (G), while they are absent in the lesioned SNr(H). The sprouting fibers in the rLV-GDNF treated animals appeared disorganized and included an intense network of thin-caliberTH-positive fibers (compare G and I). The scale bar in A represents 250 �m in A–F and the scale bar in G � 50 �m for G–I. SNc, substantianigra pars compacta; SNr, substantia nigra pars reticulata.
468 GEORGIEVSKA, KIRIK, AND BJORKLUND

Anterogradely Transported GDNF Induces Sproutingof TH-Positive Fibers along the NigrostriatalPathway
Striatal injections of rLV-GDNF resulted not only ina widespread distribution of GDNF in the striatum(Fig. 9A), but also the GDNF protein was transportedanterogradely to the globus pallidus (GP; Fig. 9B),entopeduncular nucleus (EP; Fig. 9C), and substantianigra (Fig. 9D). Figure 9 illustrates the distribution ofimmunoreactive GDNF at 10 weeks after vector injec-tion (6 weeks after the 6-OHDA lesion), but the stain-ing pattern remained essentially the same at the long-est survival times as well (36 weeks, data not shown).The levels of GDNF protein, as determined by ELISA,were 2.1 � 0.3 ng/mg tissue in the transduced striatumand 0.21 � 0.07 ng/mg in the ipsilateral nigra at 6weeks after vector injection and 0.9 � 0.2 ng/mg in thestriatum and 0.18 � 0.04 ng/mg in the nigra at 32weeks. The endogenous GDNF expression in the con-tralateral nontransduced striatum was approximately0.01 ng/mg tissue.
In the rLV-GDNF treated animals, intense sprout-ing of TH-positive fibers was observed within the SN
(Fig. 5E), entopeduncular nucleus (Fig. 6E), and globuspallidus (Fig. 7E), i.e., in areas corresponding preciselyto the distribution of anterogradely transported GDNF(Fig. 9). In the animals that had received injections ofrAAV-GFP in the SN to anterogradely label the nigro-striatal projection (see above), the GFP-positivesprouting fibers in the SN and globus pallidus wereclearly more extensive than the TH-positive sproutingfibers, suggesting that TH was downregulated in theseareas as well (Fig. 5, compare E and F for SN, Fig. 7,compare E and F for GP). No apparent sprouting ofTH-positive fibers was observed in the rLV-GFP con-trol animals (Figs. 5C, 6C, and 7C).
DISCUSSION
The results show that overexpression of GDNF in thestriatum can protect nigral DA neurons against thetoxic insult and preserve not only the nigral cell bodiesbut also the axons projecting to the striatum. This is inagreement with previous studies using rAAV (18), Ad(8), or rLV vectors (21) for intrastriatal GDNF deliveryin the rat or monkey PD model. In contrast to these
FIG. 6. Photomicrographs of adjacent sections from entopeduncular nucleus (EP) at 36 weeks after the 6-OHDA lesion (rAAV-GFPinjected animals), immunostained for TH and GFP. The normal nigrostriatal fibers extending into the EP (A and B) were largely lost in therLV-GFP control animals (C and D). In the rLV-GDNF treated animals, there was a substantial protection and intense sprouting responsethat could be visualized by TH (E) and GFP (F) stainings, also shown in high power (compare G and I). The scale bar in A represents 250�m in A–F and the scale bar in G � 50 �m for G–I.
469GDNF INDUCES DOWNREGULATION OF TYROSINE HYDROXYLASE
reports, however, we did not observe any functionalrecovery in tests of spontaneous motor behavior. Nev-ertheless, the rats displayed an almost complete rever-sal in amphetamine-induced rotation (from 10 turns/min in the GFP controls to 1 turn/min in the GDNFtreated animals), which was similar in magnitude tothat reported by Kirik et al. (18) after intrastriatalrAAV-GDNF delivery. In contrast to the Kirik et al.(18) study, however, this was not matched by any sig-nificant recovery in either the cylinder or the paw-reaching tests. This discrepancy may be explained bytwo alternative mechanisms: either by downregulationof TH in part of the striatal TH innervation or byaberrant sprouting in downstream striatal target nu-clei induced by anterograde transport of GDNF.
Downregulation of TH
The recovery seen in amphetamine-induced rotation,which developed progressively over time, is consistentwith a GDNF-induced preservation of at least a portionof the nigrostriatal projection. The retrograde and an-terograde tracing experiment (with FG and GFP, re-spectively) showed that this indeed was the case: the
number of cells in SN pars compacta that could beretrogradely labeled by FG from the striatum wasabout 2.5-fold larger in the rLV-GDNF animals than inthe lesioned controls, and the extent of striatal inner-vation, labeled by GFP from the SN, was considerablylarger in the rLV-GDNF treated rats. While the TH-positive and FG-positive cells in SN matched closely,part of the GFP-positive innervation in the striatumdid not express any detectable level of TH. This mis-match was most pronounced in those parts of the stri-atum (i.e., the dorsolateral and caudal parts) where thetransduced GDNF were most highly expressed. A sim-ilar mismatch between TH-positive fiber staining inthe striatum and rescue of cell bodies in the SN hasbeen reported by Kozlowski et al. (22) after GDNFdelivery to the SN using an Ad vector. Interestingly,the TH-negative fibers maintained their expression ofVMAT, which suggests that not all phenotypic dopa-minergic markers react in the same way to GDNF.This points to the possibility that GDNF delivery af-fects DA synthesis but not DA storage and release. Themechanism underlying GDNF-induced downregula-tion of TH is unclear. In a parallel experiment (Geor-
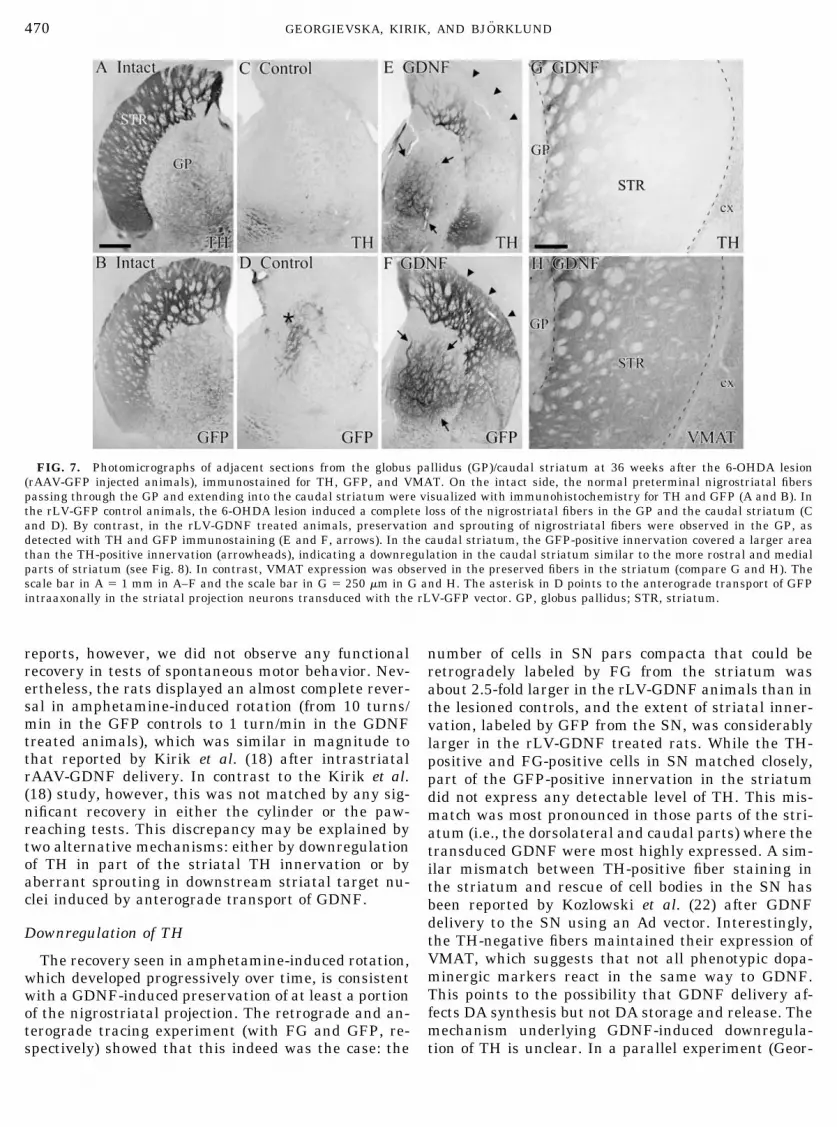
FIG. 7. Photomicrographs of adjacent sections from the globus pallidus (GP)/caudal striatum at 36 weeks after the 6-OHDA lesion(rAAV-GFP injected animals), immunostained for TH, GFP, and VMAT. On the intact side, the normal preterminal nigrostriatal fiberspassing through the GP and extending into the caudal striatum were visualized with immunohistochemistry for TH and GFP (A and B). Inthe rLV-GFP control animals, the 6-OHDA lesion induced a complete loss of the nigrostriatal fibers in the GP and the caudal striatum (Cand D). By contrast, in the rLV-GDNF treated animals, preservation and sprouting of nigrostriatal fibers were observed in the GP, asdetected with TH and GFP immunostaining (E and F, arrows). In the caudal striatum, the GFP-positive innervation covered a larger areathan the TH-positive innervation (arrowheads), indicating a downregulation in the caudal striatum similar to the more rostral and medialparts of striatum (see Fig. 8). In contrast, VMAT expression was observed in the preserved fibers in the striatum (compare G and H). Thescale bar in A � 1 mm in A–F and the scale bar in G � 250 �m in G and H. The asterisk in D points to the anterograde transport of GFPintraaxonally in the striatal projection neurons transduced with the rLV-GFP vector. GP, globus pallidus; STR, striatum.
470 GEORGIEVSKA, KIRIK, AND BJORKLUND

gievska et al., unpublished), we have observed a simi-lar downregulation of the TH protein in the intactstriatum, induced by long-term overexpression ofGDNF by the same rLV-GDNF vector as used here.However, striatal DA synthesis and turnover, mea-sured biochemically, were unaffected or increased,rather than decreased, in these animals, which is con-sistent with several previous studies that have re-
ported a stimulatory effect of GDNF on DA turnover innigrostriatal neurons (13–15, 18, 24–26). These datasuggest that TH downregulation, as observed here,may not be a sign of impaired function, but rather anadaptation of the neurons to a sustained activation ofDA turnover induced by GDNF stimulation. In ourparallel study (Georgievska et al., unpublished), wehave observed that TH downregulation is both time-
FIG. 8. Photomicrographs of adjacent sections from the striatum at 36 weeks after the 6-OHDA lesion (rAAV-GFP injected animals),immunostained for TH and GFP. On the intact side, anterogradely transported transgenic GFP was detected in the striatal terminals (B),similar to TH (A). In the rLV-GFP-injected control animals, a substantial part of the TH-positive innervation was lost (C), mainly in thelateral part of the striatum. Similarly, the GFP-positive fiber innervation in the control animals was only present in the medial parts ofstriatum (D). The asterisk in D marks one of the rLV-GFP injection sites in the striatum. In the rLV-GDNF treated animals, the TH-positivefiber innervation was restricted to the medial parts of striatum, whereas the dorsal and lateral parts were completely denervated (E).However, the GFP-positive striatal fiber innervation in the rLV-GDNF animals was more substantial and covered nearly the entire striatum(F). Visualization of TH and GFP in the same section (G and H) showed a clear colabeling in the medial part (G�–G��), whereas only GFP wasexpressed in the lateral part (H�–H��) of the striatum. The boxes in E and F depict the approximate area from which the high-power picturesare taken. The scale bar in A represents 1 mm in A–F and the scale bar in G � represents 25 �m in G�–G�� and H�–H��.
FIG. 9. Immunohistochemical visualization of the distribution of GDNF at 10 weeks after injection of rLV-GDNF in the striatum (6weeks postlesion). Transduction of the striatal projection neurons resulted in a widespread release of GDNF in the striatum (STR, A) andalong the striatofugal pathway to the globus pallidus (GP, B), entopeduncular nucleus (EP, C), and substantia nigra (SN, D). The scale barin A represents 1 mm. cc, corpus callosum; cx, cerebral cortex; EP, entopeduncular nucleus; GP, globus pallidus; ic, internal capsule; lv,lateral ventricle; SNc, substantia nigra pars compacta; SNr, substantia nigra pars reticulata; STR, striatum.
471GDNF INDUCES DOWNREGULATION OF TYROSINE HYDROXYLASE

and dose-dependent, such that it develops only after 6weeks of continuous GDNF delivery and is most pro-nounced in animals where the level of GDNF in thestriatum exceeds 0.7 ng/mg tissue. This effect is mostlikely due to sustained GDNF delivery and not to therLV vector construct per se. Thus, we have observedthe same magnitude of TH downregulation after 6–8weeks of intrastriatal GDNF delivery using a new hightiter rAAV-GDNF vector; in these animals striatalGDNF levels were 10–16 ng/mg tissue (Kirik et al.,unpublished data).
It should be noted that GDNF delivery over shorterperiods (i.e., up to 4 weeks) by either vector delivery orinfusion of recombinant GDNF protein is associatedwith increased DA turnover and an upregulation(rather than downregulation) of DA synthesis (13–15,18, 24, 26). These data are consistent with a time-dependent compensatory mechanism where continu-ous activation of DA neurons by GDNF is followed by acompensatory downregulation of the TH protein, whichin turn is accompanied by a readjustment of DA neuronactivity to within the normal range (Georgievska et al.,unpublished). While this model is consistent with theobservation made on intrastriatal GDNF delivery inrodents, it does not readily match the data obtained byKordower et al. (21) in intact aged and MPTP lesionedprimates showing that long-term (3 months) rLV-GDNF delivery is accompanied by a sustained increasein striatal DA turnover, without any sign of TH down-regulation (as assessed by immunohistochemistry).The explanation for this discrepancy is unclear, butmay be due to both the level of GDNF expression(which was about 10-fold lower than in the presentstudy) and species differences.
GDNF-Induced Aberrant Sprouting
It is well-known that exogenous GDNF can inducesprouting from lesioned nigrostriatal DA neurons. Thishas been observed in both striatum and globus pallidusfollowing injections of GDNF protein into the striatum(17, 32, 34) and in SN and adjacent parts of the mes-encephalic tegmentum and ventral thalamus after in-tranigral injections of GDNF (6, 17, 36, 40). The pres-ence of aberrant sprouting, i.e., sprouting into areaswith little or no intrinsic DA innervation, is a concernand raises the question of whether such abnormal fiberprojections may be detrimental to functional recoveryin GDNF treated animals. Previous data (18, 21) showthat good GDNF-induced functional sparing and/or re-covery may be obtained also in the presence of a limitedsprouting of TH-positive fibers in parts of the globuspallidus. Signs of overt negative functional effects,however, have been seen in animals with extensivesprouting in, or adjacent to, the SN (17, 18). In thesecases GDNF was delivered (by injection or rAAV vec-tor) directly into the SN. Interestingly, in the study by
Kirik et al. (18), using a rAAV vector to deliver GDNFto the striatum and/or SN, overexpression of GDNF inSN had a negative functional effect, not only when therAAV-GDNF vector was injected into SN alone, butalso when a striatal rAAV-GDNF injection was com-bined with an intranigral injection. In the latter case,the functional recovery seen after striatal GDNF deliv-ery alone was completely blocked by simultaneousoverexpression of GDNF in SN. In these animals,moreover, there was extensive sprouting of TH-posi-tive fibers not only in SN but also in entopeduncularnucleus and the ventral thalamus.
The present data are consistent with these previousobservations and provide further evidence that aber-rant sprouting in downstream striatal target areas, SNand entopeduncular nucleus in particular, may befunctionally detrimental in GDNF treated animals.The ability of intrastriatal rLV-GDNF to inducesprouting in these areas is most probably due to thehigh efficiency by which the transduced GDNF istransported anterogradely along the striatopallidoni-gral projections. In fact, the level of GDNF measuredby ELISA in the SN (0.2 ng/mg tissue) is by itselfsufficient to induce neuroprotective effects in SN (see(4)). In support of this interpretation we observed thatthe distribution of anterogradely transported GDNFcoincided precisely with the localization of the sproutedTH-positive fibers.
Conclusions
The present results provide further evidence thatstriatal GDNF delivery by lentiviral gene transfer ishighly efficient in protecting the nigrostriatal DA neu-rons and inducing significant regenerative sprouting inthe 6-OHDA lesion model. However, GDNF deliveredby the rLV vector (like the AAV vector used in theKirik et al. (18) study) is not only expressed in thetransduced cell bodies of the striatal projection neu-rons, but also efficiently transported anterogradelyalong the striatal efferents. As a result, GDNF is de-livered also to all three principal striatal targets, glo-bus pallidus, entopeduncular nucleus, and SN, at bio-logically active levels (about 0.2 ng/mg tissue, as mea-sured in the SN), sufficient to induce extensivesprouting of the nigrostriatal fibers in these structures.This is a concern. Although we lack direct proof, wethink that there is sufficient circumstantial evidence insupport of the view that this aberrant sprouting—par-ticularly in the entopeduncular nucleus and SN—isfunctionally detrimental and acts to block recovery inspontaneous motor behavior that would otherwisehave been obtained by GDNF-induced regeneration ofthe striatal DA afferents. Since the magnitude of ab-errant sprouting appears to be related to the level ofGDNF expression in these nuclei, it may be possible toavoid, or limit, this negative effect by careful titration
472 GEORGIEVSKA, KIRIK, AND BJORKLUND

of the vector dose (and hence the GDNF expressionlevel). Indeed, in the Kordower et al. (21) and Kirik etal. (18) studies, no or very limited sprouting was ob-served in the entopeduncular nucleus or SN in animalswith a GDNF expression level of about 0.2 ng/mg tissuein the striatum, i.e., about 10 times lower than in thepresent study. In both of these studies significant long-term recovery in spontaneous motor function was ob-served in the GDNF treated animals. Alternatively, itshould be possible to develop constructs that can limitGDNF delivery to the striatum, e.g., by using vectorstargeted to glial cells or by ex vivo gene transfer usingtransplants of cells genetically engineered to secretGDNF.
For any future clinical application of long-term vec-tor-based GDNF delivery it will be essential to deter-mine the optimal therapeutic window and site expres-sion of GDNF. This may require vectors with regulat-able promoters, so that GDNF expression can becontrolled and adjusted to within the therapeuticrange, and/or vector constructs that make it possible tolimit GDNF delivery to the striatal target structure.
ACKNOWLEDGMENTS
The work was supported by grants from the Swedish MedicalResearch Council (National Gene Therapy Program 99-XG-13285)and the Cell Factory Program of the European Commission (QLK3-1999-00702). We thank Ronald J. Mandel and the Gene Vector Coreat University of Florida, Gainesville, for the kind gift of the rAAV-GFP vector; Cecilia Lundberg for help in the production of the rLVvectors; and Birgit Haraldsson, Ulla Jarl, AnnaKarin Olden, andBengt Mattsson for excellent technical support.
REFERENCES1. Bensadoun, J. C., N. Deglon, J. L. Tseng, J. L. Ridet, A. D. Zurn,
and P. Aebischer. 2000. Lentiviral vectors as a gene deliverysystem in the mouse midbrain: Cellular and behavioral im-provements in a 6-OHDA model of Parkinson’s disease usingGDNF. Exp. Neurol 164: 15–24.
2. Bilang-Bleuel, A., F. Revah, P. Colin, I. Locquet, J. J. Robert, J.Mallet, and P. Horellou. 1997. Intrastriatal injection of an ad-enoviral vector expressing glial-cell-line-derived neurotrophicfactor prevents dopaminergic neuron degeneration and behav-ioral impairment in a rat model of Parkinson disease. Proc.Natl. Acad. Sci. USA 94: 8818–8823.
3. Bjorklund, A., C. Rosenblad, C. Winkler, and D. Kirik. 1997.Studies on neuroprotective and regenerative effects of GDNF ina partial lesion model of Parkinson’s disease. Neurobiol. Dis. 4:186–200.
4. Bjorklund, A., D. Kirik, C. Rosenblad, B. Georgievska, C. Lund-berg, and R. J. Mandel. 2000. Towards a neuroprotective genetherapy for Parkinson’s disease: Use of adenovirus, AAV andlentivirus vectors for gene transfer of GDNF to the nigrostriatalsystem in the rat Parkinson model. Brain Res. 886: 82–98.
5. Bjorklund, A., and O. Lindvall. 2000. Parkinson disease genetherapy moves toward the clinic. Nat. Med. 6: 1207–1208.
6. Bowenkamp, K. E., A. F. Hoffman, G. A. Gerhardt, M. A. Henry,P. T. Biddle, B. J. Hoffer, and A. C. Granholm. 1995. Glial cellline-derived neurotrophic factor supports survival of injuredmidbrain dopaminergic neurons. J. Comp. Neurol. 355: 479–489.
7. Choi-Lundberg, D. L., Q. Lin, T. Schallert, D. Crippens, B. L.Davidson, Y. N. Chang, Y. L. Chiang, J. Qian, L. Bardwaj, andM. C. Bohn. 1998. Behavioral and cellular protection of ratdopaminergic neurons by an adenoviral vector encoding glialcell line-derived neurotrophic factor. Exp. Neurol. 154: 261–275.
8. Connor, B., D. A. Kozlowski, T. Schallert, J. L. Tillerson, B. L.Davidson, and M. C. Bohn. 1999. Differential effects of glial cellline-derived neurotrophic factor (GDNF) in the striatum andsubstantia nigra of the aged Parkinsonian rat. Gene Ther. 6:1936–1951.
9. Conway, J. E., S. Zolotukhin, N. Muzyczka, G. S. Hayward, andB. J. Byrne. 1997. Recombinant adeno-associated virus type 2replication and packaging is entirely supported by a herpessimplex virus type 1 amplicon expressing Rep and Cap. J. Virol.71: 8780–8789.
10. Deglon, N., J. L. Tseng, J. C. Bensadoun, A. D. Zurn, Y. Arseni-jevic, L. Pereira de Almeida, R. Zufferey, D. Trono, and P.Aebischer. 2000. Self-inactivating lentiviral vectors with en-hanced transgene expression as potential gene transfer systemin Parkinson’s disease. Hum. Gene Ther. 11: 179–190.
11. Gash, D. M., Z. Zhang, and G. Gerhardt. 1998. Neuroprotectiveand neurorestorative properties of GDNF. Ann. Neurol. 44:S121–S125.
12. Georgievska, B., D. Kirik, C. Rosenblad, C. Lundberg, and A.Bjorklund. 2002. Neuroprotection in the rat Parkinson modelby intrastriatal GDNF gene transfer using a lentiviral vector.Neuroreport 13: 75–82.
13. Hebert, M. A., and G. A. Gerhardt. 1997. Behavioral and neu-rochemical effects of intranigral administration of glial cellline-derived neurotrophic factor on aged Fischer 344 rats.J. Pharmacol. Exp. Ther. 282: 760–768.
14. Horger, B. A., M. C. Nishimura, M. P. Armanini, L. C. Wang,K. T. Poulsen, C. Rosenblad, D. Kirik, B. Moffat, L. Simmons, E.Johnson, Jr., J. Milbrandt, A. Rosenthal, A. Bjorklund, R. A.Vandlen, M. A. Hynes, and H. S. Phillips. 1998. Neurturinexerts potent actions on survival and function of midbraindopaminergic neurons. J. Neurosci. 18: 4929–4937.
15. Hudson, J., A. C. Granholm, G. A. Gerhardt, M. A. Henry, A.Hoffman, P. Biddle, N. S. Leela, L. Mackerlova, J. D. Lile, F.Collins, et al. 1995. Glial cell line-derived neurotrophic factoraugments midbrain dopaminergic circuits in vivo. Brain Res.Bull. 36: 425–432.
16. Kirik, D., C. Rosenblad, and A. Bjorklund. 1998. Characteriza-tion of behavioral and neurodegenerative changes followingpartial lesions of the nigrostriatal dopamine system induced byintrastriatal 6-hydroxydopamine in the rat. Exp. Neurol. 152:259–277.
17. Kirik, D., C. Rosenblad, and A. Bjorklund. 2000. Preservation ofa functional nigrostriatal dopamine pathway by GDNF in theintrastriatal 6-OHDA lesion model depends on the site of ad-ministration of the trophic factor. Eur. J. Neurosci. 12: 3871–3882.
18. Kirik, D., C. Rosenblad, A. Bjorklund, and R. J. Mandel. 2000.Long-term rAAV-mediated gene transfer of GDNF in the ratParkinson’s model: Intrastriatal but not intranigral transduc-tion promotes functional regeneration in the lesioned nigrostri-atal system. J. Neurosci. 20: 4686–4700.
19. Kirik, D., B. Georgievska, C. Rosenblad, and A. Bjorklund.2001. Delayed infusion of GDNF promotes recovery of motorfunction in the partial lesion model of Parkinson’s disease. Eur.J. Neurosci. 13: 1589–1599.
20. Kirik, D., C. Winkler, and A. Bjorklund. 2001. Growth andfunctional efficacy of intrastriatal nigral transplants depend onthe extent of nigrostriatal degeneration. J. Neurosci. 21: 2889–2896.
473GDNF INDUCES DOWNREGULATION OF TYROSINE HYDROXYLASE

21. Kordower, J. H., M. E. Emborg, J. Bloch, S. Y. Ma, Y. Chu, L.Leventhal, J. McBride, E. Y. Chen, S. Palfi, B. Z. Roitberg,W. D. Brown, J. E. Holden, R. Pyzalski, M. D. Taylor, P. Carvey,Z. Ling, D. Trono, P. Hantraye, N. Deglon, and P. Aebischer.2000. Neurodegeneration prevented by lentiviral vector deliv-ery of GDNF in primate models of Parkinson’s disease. Science290: 767–773.
22. Kozlowski, D. A., B. Connor, J. L. Tillerson, T. Schallert, andM. C. Bohn. 2000. Delivery of a GDNF gene into the substantianigra after a progressive 6-OHDA lesion maintains functionalnigrostriatal connections. Exp. Neurol. 166: 1–15.
23. Lapchak, P. A., D. M. Gash, S. Jiao, P. J. Miller, and D. Hilt.1997. Glial cell line-derived neurotrophic factor: A novel ther-apeutic approach to treat motor dysfunction in Parkinson’sdisease. Exp. Neurol. 144: 29–34.
24. Lapchak, P. A., P. J. Miller, and S. Jiao. 1997. Glial cell line-derived neurotrophic factor induces the dopaminergic and cho-linergic phenotype and increases locomotor activity in agedFischer 344 rats. Neuroscience 77: 745–752.
25. Martin, D., G. Miller, T. Cullen, N. Fischer, D. Dix, and D.Russell. 1996. Intranigral or intrastriatal injections of GDNF:Effects on monoamine levels and behavior in rats. Eur. J. Phar-macol. 317: 247–256.
26. Martin, D., G. Miller, N. Fischer, D. Diz, T. Cullen, and D.Russell. 1996. Glial cell line-derived neurotrophic factor: Thelateral cerebral ventricle as a site of administration for stimu-lation of the substantia nigra dopamine system in rats. Eur.J. Neurosci. 8: 1249–1255.
27. McLaughlin, S. K., P. Collis, P. L. Hermonat, and N. Muzyczka.1988. Adeno-associated virus general transduction vectors:Analysis of proviral structures. J. Virol. 62: 1963–1973.
28. Montoya, C. P., L. J. Campbell-Hope, K. D. Pemberton, andS. B. Dunnett. 1991. The “staircase test”: A measure of inde-pendent forelimb reaching and grasping abilities in rats.J. Neurosci. Methods 36: 219–228.
29. Olsson, M., G. Nikkhah, C. Bentlage, and A. Bjoklund. 1995.Forelimb akinesia in the rat Parkinson model: Differential ef-fects of dopamine agonists and nigral transplants as assessedby a new stepping test. J. Neurosci. 15: 3863–3875.
30. Paxinos, G., and C. Watson. 1998. The Rat Brain in StereotaxicCoordinates, 4th ed. Academic Press, San Diego.
31. Rosenblad, C., A. Martinez-Serrano, and A. Bjorklund. 1998.Intrastriatal glial cell line-derived neurotrophic factor pro-motes sprouting of spared nigrostriatal dopaminergic afferentsand induces recovery of function in a rat model of Parkinson’sdisease. Neuroscience 82: 129–137.
32. Rosenblad, C., D. Kirik, B. Devaux, B. Moffat, H. S. Phillips,and A. Bjorklund. 1999. Protection and regeneration of nigraldopaminergic neurons by neurturin or GDNF in a partial lesionmodel of Parkinson’s disease after administration into the stri-atum or the lateral ventricle. Eur. J. Neurosci. 11: 1554–1566.
33. Rosenblad, C., M. Grønborg, C. Hansen, N. Blom, M. Meyer, J.Johansen, L. Dagø, D. Kirik, U. A. Patel, C. Lundberg, D. Trono,A. Bjorklund, and T. E. Johansen. 2000. In vivo protection ofnigral dopamine neurons by lentiviral gene transfer of the novelGDNF-family member neublastin/artemin. Mol. Cell. Neurosci.15: 199–214.
34. Rosenblad, C., D. Kirik, and A. Bjoklund. 2000. Sequentialadministration of GDNF into the substantia nigra and striatumpromotes dopamine neuron survival and axonal sprouting butnot striatal reinnervation or functional recovery in the partial6-OHDA lesion model. Exp. Neurol. 161: 503–516.
35. Schallert, T., D. A. Kozlowski, J. L. Humm, and R. R. Cocke.1997. Use-dependent structural events in recovery of function.Adv. Neurol. 73: 229–238.
36. Tseng, J. L., E. E. Baetge, A. D. Zurn, and P. Aebischer. 1997.GDNF reduces drug-induced rotational behavior after medialforebrain bundle transection by a mechanism not involvingstriatal dopamine. J. Neurosci. 17: 325–333.
37. Ungerstedt, U., and G. W. Arbuthnott. 1970. Quantitative re-cording of rotational behavior in rats after 6-hydroxydopaminelesions of the nigrostriatal dopamine system. Brain Res. 24:485–493.
38. von Schwedler, U., J. Song, C. Aiken, and D. Trono. 1993. Vif iscrucial for human immunodeficiency virus type 1 proviral DNAsynthesis in infected cells. J. Virol. 67: 4945–4955.
39. West, M. J. 1999. Stereological methods for estimating the totalnumber of neurons and synapses: Issues of precision and bias.Trends Neurosci. 22: 51–61.
40. Winkler, C., H. Sauer, C. S. Lee, and A. Bjorklund. 1996.Short-term GDNF treatment provides long-term rescue of le-sioned nigral dopaminergic neurons in a rat model of Parkin-son’s disease. J. Neurosci. 16: 7206–7215.
41. Zolotukhin, S., B. J. Byrne, E. Mason, I. Zolotukhin, M. Potter,K. Chesnut, C. Summerford, R. J. Samulski, and N. Muzyczka.1999. Recombinant adeno-associated virus purification usingnovel methods improves infectious titer and yield. Gene Ther. 6:973–985.
42. Zufferey, R., D. Nagy, R. J. Mandel, L. Naldini, and D. Trono.1997. Multiply attenuated lentiviral vector achieves efficientgene delivery in vivo. Nat. Biotechnol. 15: 871–875.
474 GEORGIEVSKA, KIRIK, AND BJORKLUND






![Chronic lithium administration alters behavioral recovery from nigrostriatal injury: Effects on neostriatal [3H]spiroperidol binding sites](https://img.pdfslide.net/doc/110x75/63556682922cbb7c550c91e1/chronic-lithium-administration-alters-behavioral-recovery-from-nigrostriatal-injury.jpg)






















