Embed Size (px)
Citation preview

Aa
Ma
b
a
ARRAA
KNFMRA
p
0d
Journal of Ethnopharmacology 136 (2011) 94–102
Contents lists available at ScienceDirect
Journal of Ethnopharmacology
journa l homepage: www.e lsev ier .com/ locate / je thpharm
brogation of nimesulide induced oxidative stress and mitochondria mediatedpoptosis by Fumaria parviflora Lam. extract
adhulika Tripathia,b, Brijesh Kumar Singha, Sheikh Raisuddinb, Poonam Kakkara,∗
Herbal Research Section, Indian Institute of Toxicology Research (CSIR), P.O. Box-80, M.G. Marg, Lucknow 226001, Uttar Pradesh, IndiaDepartment of Medical Elementology and Toxicology, Hamdard University, New Delhi 110062, India
r t i c l e i n f o
rticle history:eceived 17 August 2010eceived in revised form 3 March 2011ccepted 6 April 2011vailable online 14 April 2011
eywords:imesulideumaria parviflora Lam.itochondria
eactive oxygen speciespoptosis
a b s t r a c t
Ethanopharmacological relevance: Fumaria parviflora Lam. is used for treating aches and pains, diarrhea,fever, influenza and other complications. The herb mixed with honey is taken to prevent vomiting as perAyurvedic text.Aim of the study: In vivo studies were conducted to explore the hepatoprotective potential of Fumariaparviflora Lam. Fp extract against nimesulide induced oxidative stress and regulation of critical events inmitochondria mediated apoptosis.Materials and methods: Group of Wistar rats were fed with nimesulide for 5 days (80 mg/kg/day, po),another group was pre-treated with Fp extract/silymarin (200 mg/kg/day, po) for 5 days followed bynimesulide exposure. Liver serum biomarkers and histopathology were done to assess hepatotoxicitycaused by nimesulide. Antioxidant enzymes (SOD, LPO, GPx, GR) were assessed using biochemical assaysas well as gene expression by RT-PCR. GSH content and ROS generation was also evaluated using flowcytometry. Key apoptotic markers like phosphatidyl serine externalization, Bax, Bcl-2 translocation, mito-chondrial membrane potential, cytochrome c release, caspases (9/3) activation and DNA damage werealso observed in all the groups to confirm involvement of mitochondrial pathway.Results: Pre-treatment with Fp extract for 5 days significantly reduced the impact of nimesulide inducedtoxicity as evident from the serum biomarkers of liver damage and histopathology. It also modulatedantioxidant enzymes mRNA expression as well as activity (SOD, glutathione peroxidase, glutathionereductase) and reduced lipid peroxidation during nimesulide toxicity. Nimesulide exposure decreasedGSH content (92.9%) and increased reactive oxygen species (9.29 fold) which was attenuated in Fp treated
rats. Fp pre-treatment significantly altered key apoptotic events like Bcl2 and Bax translocation, inhibitedmitochondrial depolarization, prevented cytochrome c release, caspase-9/caspase-3 activation and DNAdamage.Conclusion: Our in vivo findings regarding protection accorded by Fp extract against nimesulide toxicitysuggest that Fp not only reduced hepatotoxicity but attenuated critical control points of apoptotic celldeath.Abbreviations: �m, mitochondrial membrane potential; ADR, adverse drug reactiobutylated hydroxyanisole; BHT, butylated hydroxytoluene; CAD, caspases activated Detate; COX-2, cyclo-oxygenase-2; cyt c, cytochrome c; DILI, drug induced liver injurydimethyl sulfoxide; FITC, fluorescein isothiocynate; Fp, Fumaria parviflora Lam.; GSHHEPES, 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid; HPLC, high performance1′ ,1′ ,3′ ,3′-tetraethylbenzamidazolcarbocyanine iodide; LPO, lipid peroxidation; MDpermeability transition; NBT, nitroblue tetrazolium; NSAIDs, non-steroidal anti-inflamreverse phase high performance liquid chromatography; SOD, superoxide dismutase;
∗ Corresponding author. Tel.: +91 0522 2613786/2627586*269; fax: +91 0522 2628227E-mail addresses: madhulika [email protected] (M. Tripathi), singhbrijeshk@hotma
oonam [email protected] (P. Kakkar).
378-8741/$ – see front matter © 2011 Elsevier Ireland Ltd. All rights reserved.oi:10.1016/j.jep.2011.04.014
© 2011 Elsevier Ireland Ltd. All rights reserved.
n; AIF, apoptosis inducing factor; ApoAF-1, apoptosis activating factor-1; BHA,Nase; CMF, chloromethylfluorescein; CMF-DA5′-, chloromethylfluorescein diac-; DCF, dichlorofluorescein; DCFH-DA, 2′ ,7′-dichlorofluorescein diacetate; DMSO,, reduced glutathione; GPX, glutathione peroxidase; GR, glutathione reductase;liquid chromatography; HRP, horseradish peroxidase; JC, -15′ ,5′ ,6′ ,6′-tetrachloro-A, malonyldialdehyde; MFI, mean fluorescence intensity; MPT, mitochondrialmatory drugs; ODS, octa dodcyl selanne; ROS, reactive oxygen species; RP-HPLC,TBARS, thiobarbituric acid reactive substance..il.com (B.K. Singh), [email protected] (S. Raisuddin),

nopha
1
timpeaare
),aecsi(Raammdadr
cleittdoNapb
imaaybtas
ilEtaaPacaat
c
M. Tripathi et al. / Journal of Eth
. Introduction
Hepatotoxicity is the potential complication of most drugherapies prevalent in patients treated with nonsteroidal anti-nflammatory drugs (NSAIDs) (Bjornsson, 2010). NSAIDs are the
ost frequently prescribed therapeutic agents. Due to its over use,ronounced side effects occur, which range from mild, transientlevations in serum transaminases to pronounced hepatocellularnd/or cholestatic injury sometimes leading to fatal fulminant hep-titis. Cases of reported hepatotoxicity is one of the importanteasons resulting in withdrawal of some of the popular NSAIDs (Hant al., 2010).
Nimesulide (N-(4-nitro-2-phenoxyphenyl)methanesulfonamiden NSAID with anti-inflammatory, anti-pyretic and analgesic prop-rties, is one of the most prescribed drug with largest market asompared to other NSAIDs (Kalra et al., 2009). Reactive oxygenpecies (ROS) and resultant mitochondrial dysfunction has beenmplicated as a general mechanism in the toxicity of many NSAIDsHan et al., 2010). Several groups have suggested that intracellularOS generation may also constitute a conserved apoptotic eventnd cite ROS production as a critical determinant of toxicityssociated with exposure to such drugs (Zhang et al., 2010). Asitochondria are the major site of ROS production, hence theyay create adequate oxidative stress which may lead to organelle
ysfunction, ultimately leading to cell death. Mitochondria havelso been found to be frequently involved in the toxicity of manyrugs and other xenobiotics and have been the subject of excellenteviews (Han et al., 2010).
Nimesulide adversely affects mitochondrial function where itauses uncoupling and oxidation of NAD(P)H as studied in isolatediver mitochondria (Singh et al., 2010). However, the sequence ofvents of the mitochondrial injury, as well as its relevance in vivo,s still unclear. In particular, the role of mitochondrial permeabilityransition (MPT), a nonspecific mechanism of mitochondrial injuryhat can lead to apoptosis and/or necrosis has been a matter ofebate and has produced controversial results. Although the causesf cell death are indistinct, the mode can be apoptosis or necrosis.ecrosis is usually a consequence of acute metabolic perturbations occurs in acute drug-induced hepatotoxicity. In contrast, apo-tosis represents the execution of a death program often initiatedy quite specific stimuli.
In chronic health problems where use of NSAIDs becomesmperative, it is logical to search for alternative/complimentary
edicines which can reduce its toxicity. In cases of liver injury,ntioxidants hold promise. Although there are several syntheticntioxidants available in the market such as butylated hydrox-anisole (BHA), butylated hydroxytoluene (BHT) and propyl gallateut some toxicity is reported due to them (Porkey, 2007). Therefore,reating liver diseases with plant-derived compounds which areccessible and do not require laborious pharmaceutical synthesiseems attractive.
Fumaria parviflora Lam. (Fp) belongs to family Fumiraceae ands of great ethanopharmacolgical use as per the Indian Ayurvediciterature. It is distributed throughout India and also found inurope, Africa and many other Asian countries. Based on tradi-ional knowledge, it is widely used as an anti-scabies, anti-scorbite,nti-bronchite, diuretic, expectorant, anti-pyretic, diaphoretic,ppetizer, anti-neoplastic and for management of liver disorders.hytochemical analysis of Fp has indicated the presence of organiccids such as fumaric acid and isoquinoline alkaloids, protropine,ryptopine, sinactine, stylopine, dihydro-fumariline, per-fumidine,nd dihydrosanguirine (Suau et al., 2002) Antinociceptive property,
cetylcholinesterase and butyrylcholinesterase inhibitory activi-ies of Fp have been reported by Orhan et al., 2004.Silymarin, a known hepatoprotectant, was used as a positiveontrol throughout the study. It is known to have clinical appli-
rmacology 136 (2011) 94–102 95
cations in the treatment of toxic hepatitis, fatty liver, cirrhosis,ischemic injury, and viral hepatitis via its anti-oxidative, anti-lipidperoxidative, antifibrotic, anti-inflammatory and liver regeneratingeffects (Girish et al., 2009).
Present study was undertaken to investigate the role of ROSand mitochondrial dysfunction in nimesulide-induced liver injuryin vivo and also to explore the protective efficacy of Fp extractagainst nimesulide-induced liver injury.
2. Materials and methods
Goat polyclonal antibodies against cytochrome c, Bax, rabbitpolyclonal antibodies against Bcl2 and mouse polyclonal antibodyagainst �-actin were procured from Santa Cruz Biotechnology Inc.(CA, USA). Horseradish peroxidase (HRP) conjugated secondaryantibodies were also obtained from the same source. Cytochromec oxidase subunit-IV (COX-IV) antibody was procured from Abcam(Cambridge, UK). Chemiluminescent HRP substrate (ImmobilonTM
Western) was procured from Millipore (Bellarica, USA). CellTracker Green 5′-Chloromethylflouroscein diacetate (CMF-DA)was procured from Molecular Probes (Eugene, OR, USA). 5,5′,6,6′-Tetrachloro-1,1′,3,3′-tetraethylbenzamidazolcarbocyanine iodide(JC-1), 2′,7-dichlorofluorescein diacetate (DCFH-DA), nimesulide(N-(4-nitro-2-phenoxyphenyl)methanesulfonamide), silymarinand all other chemicals were purchased from Sigma ChemicalCo.(St. Louis, MO, USA) unless otherwise mentioned.
2.1. Plant material and extract preparation
Whole plant of Fumaria parviflora Lam. (Fp) was collectedfrom Banthra centre of National Botanical Research Institute, Luc-know, India, authenticated by the taxonomy experts and a voucherspecimen number-93991 was issued. Standardized Fp extract asreported earlier (Tripathi et al., 2010) was used in the presentstudy. In brief, analysis of fumaric acid and protopine in lyophilized50% ethanolic Fp extract was performed using RP-HPLC with pho-todiode array detector on Spherisorb ODS C-18 column. The UVabsorption spectrum of fumaric acid (retention time: 11.948 min)and protopine (5.489 min) showed intense absorption at 226 nmand 280 nm, respectively.
2.2. Animals
Animal handling in all experimental procedures was approvedby the Institutional Animal Ethics Committee (ITRC/IAEC/15/2008).Sprague-Dawley male rats weighing 200 ± 20 g from Indian Insti-tute of Toxicology Research (IITR) animal colony were used forthe experiments. Rats were housed at controlled temperature(25 ± 2 ◦C) humidity (60–70%) and light/dark cycle (12 h). Rats werefed on standard pellet diet (Ashirwad pellet diet, Mumbai, India)and water ad libitum.
2.3. Dosage and drug administration
Rats (36 nos.) were randomly divided into six groups of sixanimals each, and housed in six independent cages. The drug, nime-sulide was dissolved in DMSO at a concentration of 100 mg/mLand filtered through 0.22 �m syringe filters (Millex-GV, Millipore,Ireland). Whole plant extract of Fp was suspended in 10% DMSOand administered po daily (200 mg/kg/day) for 5 days. The sixgroups of animals were as follows: Group I (control) received DMSO(1 �L/g/day, po), Group II received nimesulide (80 mg/kg/day, po),
Groups III and IV received 200 mg/kg body weight/day of Fp extractand silymarin, and Groups V and VI, Fp extract and silymarin(200 mg/kg/day, po) was administered for 5 days followed by nime-sulide (80 mg/kg/day, po) for 5 days as shown below:
9 nopha
(
2
2
taask
2p
fighb(c
iIECs1tf
2
pnpDLP(+4dn
2
oDtDbl3tl
6 M. Tripathi et al. / Journal of Eth
The study was divided in two parts:
(i) To analyze the histopathology, serum biochemistry and molec-ular parameter.
ii) Primary hepatocytes from control and treated rats wereisolated to assess ROS, reduced glutathione (GSH) level, mito-chondrial membrane potential (�m) and apoptosis on flowcytometer.
.4. Hepatotoxicity assessment
.4.1. Blood collection and biochemical parametersBlood samples were collected from the jugular vein and cen-
rifuged at 2000 × g for 15 min at 4 ◦C to separate serum. Levels oflanine aminotransferase (ALT), aspartate aminotransferase (AST)nd serum bilirubin (SB) were determined by an automated analy-is system (Chemwell Awareness Technologies, Inc., Germany) withits purchased from Spinreact (Spain) to assess hepatotoxicity.
.5. Tissue collection and preparation of homogenate andreparation of single cell suspension from liver
Rats were euthanized by chloroform and dissected. Apart fromxed tissue, remaining sections were frozen quickly in liquid nitro-en and stored at −80 ◦C for biochemical/molecular analysis. Tissueomogenate (10%, w/v) was prepared in 10 mM sodium phosphateuffer (pH 7.4) using electric motor with glass/Teflon homogenizerLimaye et al., 2003). All procedures were carried out under coldonditions (0–4 ◦C).
Single cells were isolated from the liver of treated rats accord-ng to the two step collagenase perfusion method (Seglen, 1976).n brief, liver was perfused with HEPES buffer containing 0.1 mMGTA, followed by collagenase and HEPES containing 1.8 mMaCl2·2H2O. Hepatocytes were maintained in RPMI-1640 mediaupplemented with heat-inactivated 10% fetal bovine serum and% of 10,000 units penicillin, 10 mg streptomycin, 25 �g ampho-ericin B, 1 mM sodium pyruvate, 2 mM l-glutamine in the mediaor preparation of single cell suspension.
.6. Annexin V-FITC binding assay
In order to evaluate apoptosis and necrosis, externalization ofhosphotidylserine (PS) during apoptosis and DNA leakage fromecrotic cells was observed by Annexin V/PI dual staining kit (Apo-tosis detection kit-APOAF) procured from Sigma (St. Louis, USA).ifferentiation of cell population was done on flow cytometer (BD-SR) and was assessed as: (a) viable hepatocytes (Annexin −ve;I −ve), (b) early apoptotic hepatocytes (Annexin +ve; PI −ve),c) late apoptotic cells (Annexin +ve; PI +ve), (d) necrotic cells (PIve) (Liu et al., 2003). Samples were analyzed using FL-1 (Ex/Em:90/520 nm)/FL-2 (Ex/Em: 488/617 nm) band pass filters for theetection of Annexin V-FITC and PI simultaneously. Each determi-ation is based on mean fluorescence intensity of 10,000 events.
.7. Measurement of intracellular ROS generation
Intracellular ROS was detected by an oxidation-sensitive flu-rescent probe 2′,7′-dichlorofluorescein diacetate (DCFH-DA).CFH-DA is a membrane permeable dye which de-esterified
o 2′,7′-dichlorodihydrofluorescein (DCFH2) by cellular esterases.CFH2 is further oxidized to fluorescent dichlorofluorescein (DCF)y ROS, mainly by H2O2 and hydroxyl radicals. In brief, cells from
iver of different treatment groups were isolated and incubated for0 min with DCFH-DA (5 �g/mL) at 37 ◦C in dark. Samples werehen analyzed using FL-1 band pass filter at an excitation wave-ength of 480 nm and emission wavelength of 525 nm on flow
rmacology 136 (2011) 94–102
cytometer (BD-LSR). Each determination is based on mean fluo-rescence intensity of 10,000 events (Campbell et al., 2009).
2.8. GSH content
Cell tracker greenTM (5′-chloromethylflouroscein diacetate)(Molecular probes, Oregon, USA) was used to measure the total GSHin the cells, major cellular thiol as per the protocol reported earlier(Singh et al., 2010). The dye passes freely through the cell mem-brane, but once inside the cells is transformed into cell impermeantfluorescent reaction product. The cells were incubated with the dye(5 �g/mL) for 30 min in dark at 37 ◦C. Acquisition was performedwith the treated cells on BD-LSR flow cytometer and analysis wasdone using FL-1 band pass filter.
2.9. Superoxide dismutase (SOD) activity
SOD activity was determined spectrophotometrically by inhi-bition of nicotinamide adenine dinucleotide (reduced)-phenazinemethosulfate-nitroblue tetrazolium reaction system by the methodof Kakkar et al. (1984). 50% inhibition of formazan formation in1 min was taken as one unit activity and represented as per mgprotein.
2.10. MDA determination
Lipid peroxidation was quantified by measuring malondialde-hyde (MDA), a breakdown product formed from polyunsaturatedfatty acids (PUFA) hydroperoxides by the method of Wallin et al.(1993). The concentration of thiobarbituric acid reactive substances(TBARS) was expressed as nmol of MDA formation using a freshlydiluted 1,1,3,3-tetraethoxypropane as standard.
2.11. Glutathione peroxidase (GPx) activity
GPx activity was assayed using the method of Flohe and Gunzler(1984). GPx was measured indirectly by a coupled reaction withglutathione reductase (GR) using t-BHP as substrate. In brief,homogenate was incubated with 1 mL of buffer (50 mM Tris–HCl,pH 7.6, 5 mM EDTA, 1 mM GSH, 0.22 mM NADPH and 0.4 U GR) andthe reaction was initiated by adding t-BHP to a final concentra-tion of 0.22 mM. NADPH oxidation was measured as decrease inabsorbance for 3 min at every 1 min time interval, at 340 nm. 1 �Mof NADPH oxidized per min was considered as one unit activity andrepresented as per mg protein
2.12. Glutathione reductase (GR) activity
GR activity was determined according to Carlberg andMannervik (1985). The GR activity was assayed as NADPH-dependent reduction of oxidized glutathione (GSSG). In brief,homogenate was suspended in 1 mL of reaction buffer (0.125 mMNADPH, 1 mM GSSG, 1 mM EDTA in 100 mM potassium phosphatebuffer, pH 7.0). The reaction was initiated by adding homogenateto the cuvette and the decrease in absorbance at 340 nm was fol-lowed for 3 min at 1 min time interval. 1 �M of NADPH oxidized permin was considered as one unit activity and represented as per mgprotein.
2.13. Reverse transcriptase (RT) PCR
Total RNA was extracted from control and treated liver of rats.
Liver was homogenized using DEPC treated glass/Teflon homog-enizer in TRIzol reagent (Invitrogen, CA, USA) according to themanufacturer’s protocol. Extracted total RNA was further treatedwith DNase I for 15 min at 37 ◦C.
nopharmacology 136 (2011) 94–102 97
upw(idptasa
2
m(lmdwt5aprwr
2
dp11tSR(c4
2
twiaeswmBiaabIlir0o
Table 1Serum biochemical parameters of toxicity.
Group ALT (U/L) AST (U/L) Bilirubin (mg/dL)
Vehicle control 46.7 ± 12.1 46.3 ± 24.2 0.13 ± 0.10Nimesulide 160.7 ± 23.2*** 196.7 ± 31.2*** 0.85 ± 0.04***
Fp alone 58.5 ± 13.3 54.1 ± 13.4 0.17 ± 0.01Sily alone 55.4 ± 12.9 53.5 ± 16.9 0.18 ± .01Fp Pre 71.8 ± 26.2### 77.4 ± 20.9### 0.29 ± 0.02###
Sily Pre 71.3 ± 28.8### 78.2 ± 21.1### 0.42 ± 0.03###
Values are expressed as mean ± SD of 6 animals in each group. Fp: Fumaria parviflora
M. Tripathi et al. / Journal of Eth
First strand cDNA was synthesized from 4 �g of total RNAsing Superscript III (Invitrogen, CA, USA) with 50 �M of oligo dTrimer according to manufacturer’s instruction. Synthesized cDNAas accordingly diluted and used for PCR with suitable primers
Gagliano et al., 2006). Amplification reactions were conductedn a final volume of 20 �L containing 100 pg of cDNA, 10 mMNTP, 50 mM MgCl2, 10 �mol of each primer, and 5 U of Taq DNAolymerase (Invitrogen, CA, USA). The PCR products were elec-rophoresed on 2.0% agarose gels, stained with ethidium bromidend quantified by densitometric analysis (Alfa-Innotech; Cell Bio-ciences, Inc. Santa Clara, CA). In the results �-actin gene was useds internal control.
.14. Measurement of mitochondrial membrane potential (�m)
JC-1 dye is a cationic dye that selectively accumulates withinitochondria. Under normal negative transmembrane potential
polarized), it forms red fluorescent J-aggregates, while in depo-arized conditions it exists as green monomers. This characteristic
akes JC-1 more specific probe for �m measurement than otheryes. Cells were incubated with 2.5 �g/mL JC-1 for 30 min. Afterashing the hepatocytes, �m was assessed by comparing the
wo fluorescence, red (FL-2: 590 nm; emission) and green (FL-1:27 nm; excitation) using flow cytometry (BD-LSR) (Cossarizza etl., 1996) and analysis was performed using Cell Quest software. Dotlot generated by the software is divided in two halves; upper halfepresents polarized mitochondria (fluorescence of J-aggregates)hereas lower half represents mitochondrial depolarization (fluo-
escence of monomeric JC-1).
.15. Preparation of sub-cellular fractions
Cytosolic and mitochondrial fractions were prepared asescribed by Zhang et al. (1999). Briefly, tissue homogenates wererepared in ice-cold RIPA buffer (20 mM HEPES–KOH, pH 7.5,0 mM KCl, 1.5 mM MgCl2, 1.0 mM dithiothreitol, 1.0 mM EDTA,.0 mM EGTA, 1.0 mM phenylmethylsulfonyl fluoride) and cen-rifuged (Sigma 3K18, Germany) at 800 × g for 4 min at 4 ◦C.upernatant was then centrifuged at 22,000 × g for 15 min at 4 ◦C.esulting supernatant was used as cytosolic fraction and pelletmitochondria) was resuspended in ice cold RIPA buffer, and soni-ated for 20 s. The lysate was centrifuged at 15,000 × g for 15 min at◦C. The resultant supernatant was used as mitochondrial fraction.
.16. Immunoblot analysis for translocation of Bax, Bcl2 and cyt c
The protein content corresponding to each treatment was quan-ified using the method of Lowry et al. (1951). Tissue samplesere incubated with Lammeli’s buffer for 5 min at 96 ◦C and
mmediately kept on ice. 30 �g of protein sample from cytosolicnd mitochondrial fraction was separated on 12% SDS-PAGE andlectroblotted on PVDF membrane (HybondTM-P Amersham bio-ciences, UK limited, NA). After blocking with 1× blocking buffer,ashing was done using PBS-T containing 0.1% Tween 20. Theembrane was then incubated for 1 h with primary antibodies.
cl2, cyt c, Bax was used in 1:500, �-actin in 1:1000 and COX-IVn 1:2000 dilution. Incubation was followed by washing with PBSnd incubated with horseradish peroxidase-conjugated secondaryntibodies in 1:1000 dilution for 1 h at room temperature. Mem-rane was rewashed and the immunoblot was visualized using
mmobilonTM Western Chemiluminescent HRP substrate kit. (Mil-ipore, Corporation, MA, USA). �-Actin and COX-IV were used as
nternal loading control for cytosolic and mitochondrial fractions,espectively. PageRulerTM Prestained Protein Ladder (5 �L; SM-671 from Fermentas, EU) was used to determine molecular weightf the protein bands. Densitometry of the bands obtained was doneextract; Sily: Silymarin.*** P < 0.001, as compared to control group.
### P < 0.001, as compared to nimesulide treated group.
using software ImageJ version 1.41o (NIH, USA). Expression levelof cyt c was seen in cytosolic fraction whereas Bcl2 and Bax inmitochondrial fraction.
2.17. Caspases-9 and -3 activity
Enzymatic activities of caspase-9 and -3 were measured inthe liver by the liberation of a colorimeteric substance pNA afterdegradation of peptide substrate specific for caspase-3 (Ac-DEVD-pNA) and caspase-9 (Ac-LEHD-pNA) according to manufacturer’sprotocol. In brief, fresh liver tissue was homogenized in extractionbuffer (25 mM HEPES buffer, pH 7.4, containing 5 mM EDTA,2 mM dithiothreitol, 0.1% CHAPS and 1% protease cocktail). Thehomogenate was centrifuged at 20,000 × g for 25 min. The result-ing supernatants were diluted with assay buffer (50 mM HEPES,10 mM dithiothreitol, 1.0 mM EDTA, 100 mM NaCl, 0.1% CHAPSand 10% glycerol, pH 7.4) and incubated at 37 ◦C with specificsubstrates. Cleavage of the substrates was monitored at 405 nm.The caspase activity was expressed as pmol of pNA produced permilligram of protein.
2.18. Statistical analysis
Data are expressed as mean ± SD. Data were analyzed by one-way ANOVA followed by Tukey test. */#P < 0.05, **/##P < 0.01,***/###P < 0.001 were used as the criterion for significance (SPSSsoftware version 14.0). * Indicates when compared with controland # when compared with the stress.
3. Results
3.1. Liver biomarkers
Administration of nimesulide (80 mg/kg body wt/day for 5 days,po) caused a significant increase in serum biomarkers of hepatotox-icity, namely ALT, AST and serum bilirubin in rats, as compared tocontrol animals (Table 1). The levels of ALT, AST and SB were signif-icantly increased to 3.44, 4.25 and 6.53 fold, respectively (P < 0.001)in the rats treated with nimesulide. In rats pre-administered with Fpextract, these biomarkers of hepatotoxicity were significantly low-ered as compared to nimesulide treated rats and the values werecloser to control levels. Silymarin pre-treatment also preventednimesulide induced increase in ALT, AST and SB levels.
3.2. Apoptosis study (Annexin/PI)
A hallmark of programmed cell death (PCD) is the redistributionof phosphatidyl serine (PS) to the outer side of the plasma mem-brane while on viable cells it is confined to the inner side and is
used as a marker. The relocation of PS to the cell membrane wasdetected by Annexin V/PI dual staining. Treatment with nimesulideincreased the number of apoptotic cells to 43.21% in liver as com-pared to control (2.72%) whereas increase in necrotic cells were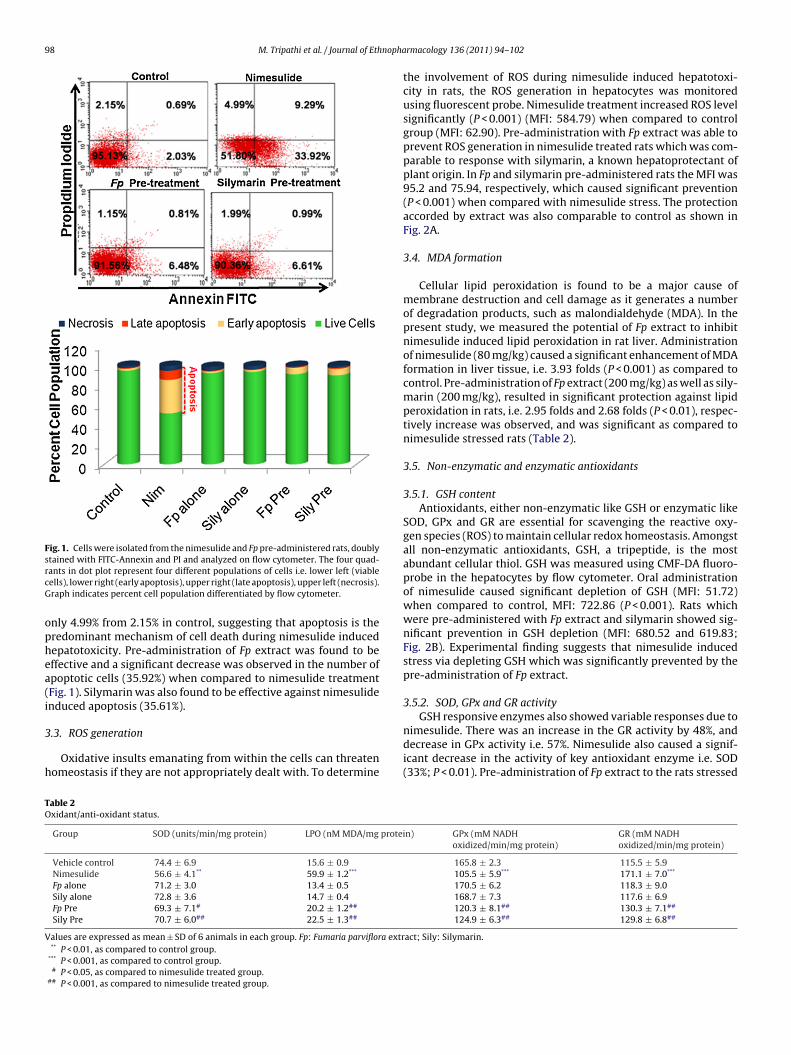
98 M. Tripathi et al. / Journal of Ethnopha
Fig. 1. Cells were isolated from the nimesulide and Fp pre-administered rats, doublystained with FITC-Annexin and PI and analyzed on flow cytometer. The four quad-rcG
ophea(i
3
h
TO
V
ants in dot plot represent four different populations of cells i.e. lower left (viableells), lower right (early apoptosis), upper right (late apoptosis), upper left (necrosis).raph indicates percent cell population differentiated by flow cytometer.
nly 4.99% from 2.15% in control, suggesting that apoptosis is theredominant mechanism of cell death during nimesulide inducedepatotoxicity. Pre-administration of Fp extract was found to beffective and a significant decrease was observed in the number ofpoptotic cells (35.92%) when compared to nimesulide treatmentFig. 1). Silymarin was also found to be effective against nimesulidenduced apoptosis (35.61%).
.3. ROS generation
Oxidative insults emanating from within the cells can threatenomeostasis if they are not appropriately dealt with. To determine
able 2xidant/anti-oxidant status.
Group SOD (units/min/mg protein) LPO (nM MDA/mg protei
Vehicle control 74.4 ± 6.9 15.6 ± 0.9Nimesulide 56.6 ± 4.1** 59.9 ± 1.2***
Fp alone 71.2 ± 3.0 13.4 ± 0.5Sily alone 72.8 ± 3.6 14.7 ± 0.4Fp Pre 69.3 ± 7.1# 20.2 ± 1.2##
Sily Pre 70.7 ± 6.0## 22.5 ± 1.3##
alues are expressed as mean ± SD of 6 animals in each group. Fp: Fumaria parviflora extr** P < 0.01, as compared to control group.
*** P < 0.001, as compared to control group.# P < 0.05, as compared to nimesulide treated group.
## P < 0.001, as compared to nimesulide treated group.
rmacology 136 (2011) 94–102
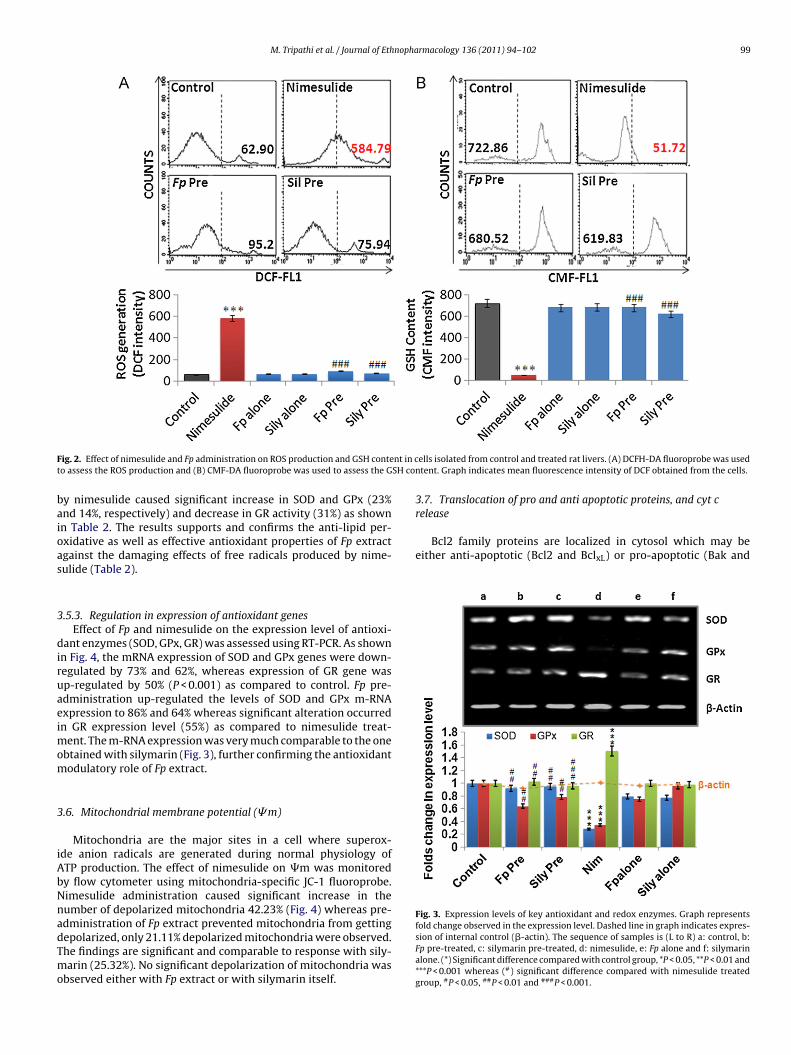
the involvement of ROS during nimesulide induced hepatotoxi-city in rats, the ROS generation in hepatocytes was monitoredusing fluorescent probe. Nimesulide treatment increased ROS levelsignificantly (P < 0.001) (MFI: 584.79) when compared to controlgroup (MFI: 62.90). Pre-administration with Fp extract was able toprevent ROS generation in nimesulide treated rats which was com-parable to response with silymarin, a known hepatoprotectant ofplant origin. In Fp and silymarin pre-administered rats the MFI was95.2 and 75.94, respectively, which caused significant prevention(P < 0.001) when compared with nimesulide stress. The protectionaccorded by extract was also comparable to control as shown inFig. 2A.
3.4. MDA formation
Cellular lipid peroxidation is found to be a major cause ofmembrane destruction and cell damage as it generates a numberof degradation products, such as malondialdehyde (MDA). In thepresent study, we measured the potential of Fp extract to inhibitnimesulide induced lipid peroxidation in rat liver. Administrationof nimesulide (80 mg/kg) caused a significant enhancement of MDAformation in liver tissue, i.e. 3.93 folds (P < 0.001) as compared tocontrol. Pre-administration of Fp extract (200 mg/kg) as well as sily-marin (200 mg/kg), resulted in significant protection against lipidperoxidation in rats, i.e. 2.95 folds and 2.68 folds (P < 0.01), respec-tively increase was observed, and was significant as compared tonimesulide stressed rats (Table 2).
3.5. Non-enzymatic and enzymatic antioxidants
3.5.1. GSH contentAntioxidants, either non-enzymatic like GSH or enzymatic like
SOD, GPx and GR are essential for scavenging the reactive oxy-gen species (ROS) to maintain cellular redox homeostasis. Amongstall non-enzymatic antioxidants, GSH, a tripeptide, is the mostabundant cellular thiol. GSH was measured using CMF-DA fluoro-probe in the hepatocytes by flow cytometer. Oral administrationof nimesulide caused significant depletion of GSH (MFI: 51.72)when compared to control, MFI: 722.86 (P < 0.001). Rats whichwere pre-administered with Fp extract and silymarin showed sig-nificant prevention in GSH depletion (MFI: 680.52 and 619.83;Fig. 2B). Experimental finding suggests that nimesulide inducedstress via depleting GSH which was significantly prevented by thepre-administration of Fp extract.
3.5.2. SOD, GPx and GR activityGSH responsive enzymes also showed variable responses due to
nimesulide. There was an increase in the GR activity by 48%, anddecrease in GPx activity i.e. 57%. Nimesulide also caused a signif-icant decrease in the activity of key antioxidant enzyme i.e. SOD(33%; P < 0.01). Pre-administration of Fp extract to the rats stressed
n) GPx (mM NADHoxidized/min/mg protein)
GR (mM NADHoxidized/min/mg protein)
165.8 ± 2.3 115.5 ± 5.9105.5 ± 5.9*** 171.1 ± 7.0***
170.5 ± 6.2 118.3 ± 9.0168.7 ± 7.3 117.6 ± 6.9120.3 ± 8.1## 130.3 ± 7.1##
124.9 ± 6.3## 129.8 ± 6.8##
act; Sily: Silymarin.
M. Tripathi et al. / Journal of Ethnopharmacology 136 (2011) 94–102 99
F nt in ct SH co
baioas
3
diruaeimom
3
iAbNnadTmo
Bcl2 family proteins are localized in cytosol which may beeither anti-apoptotic (Bcl2 and BclxL) or pro-apoptotic (Bak and
Fig. 3. Expression levels of key antioxidant and redox enzymes. Graph representsfold change observed in the expression level. Dashed line in graph indicates expres-
ig. 2. Effect of nimesulide and Fp administration on ROS production and GSH conteo assess the ROS production and (B) CMF-DA fluoroprobe was used to assess the G
y nimesulide caused significant increase in SOD and GPx (23%nd 14%, respectively) and decrease in GR activity (31%) as shownn Table 2. The results supports and confirms the anti-lipid per-xidative as well as effective antioxidant properties of Fp extractgainst the damaging effects of free radicals produced by nime-ulide (Table 2).
.5.3. Regulation in expression of antioxidant genesEffect of Fp and nimesulide on the expression level of antioxi-
ant enzymes (SOD, GPx, GR) was assessed using RT-PCR. As shownn Fig. 4, the mRNA expression of SOD and GPx genes were down-egulated by 73% and 62%, whereas expression of GR gene wasp-regulated by 50% (P < 0.001) as compared to control. Fp pre-dministration up-regulated the levels of SOD and GPx m-RNAxpression to 86% and 64% whereas significant alteration occurredn GR expression level (55%) as compared to nimesulide treat-
ent. The m-RNA expression was very much comparable to the onebtained with silymarin (Fig. 3), further confirming the antioxidantodulatory role of Fp extract.
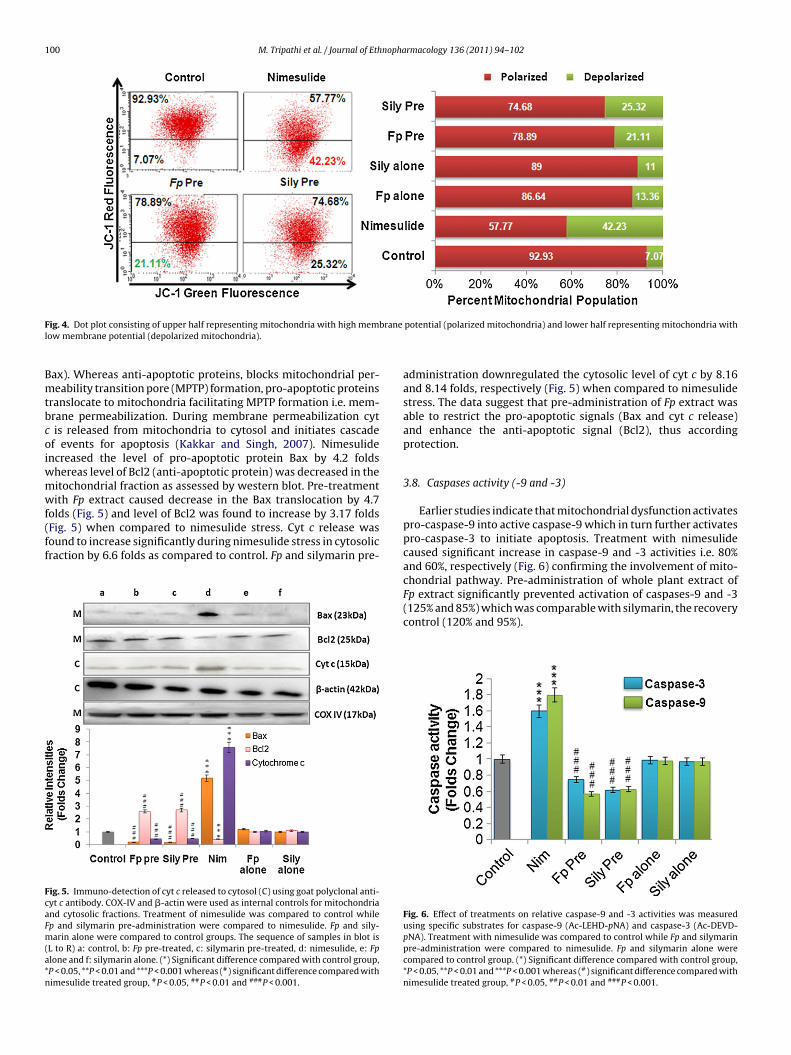
.6. Mitochondrial membrane potential (�m)
Mitochondria are the major sites in a cell where superox-de anion radicals are generated during normal physiology ofTP production. The effect of nimesulide on �m was monitoredy flow cytometer using mitochondria-specific JC-1 fluoroprobe.imesulide administration caused significant increase in theumber of depolarized mitochondria 42.23% (Fig. 4) whereas pre-dministration of Fp extract prevented mitochondria from getting
epolarized, only 21.11% depolarized mitochondria were observed.he findings are significant and comparable to response with sily-arin (25.32%). No significant depolarization of mitochondria wasbserved either with Fp extract or with silymarin itself.
ells isolated from control and treated rat livers. (A) DCFH-DA fluoroprobe was usedntent. Graph indicates mean fluorescence intensity of DCF obtained from the cells.
3.7. Translocation of pro and anti apoptotic proteins, and cyt crelease
sion of internal control (�-actin). The sequence of samples is (L to R) a: control, b:Fp pre-treated, c: silymarin pre-treated, d: nimesulide, e: Fp alone and f: silymarinalone. (*) Significant difference compared with control group, *P < 0.05, **P < 0.01 and***P < 0.001 whereas (#) significant difference compared with nimesulide treatedgroup, #P < 0.05, ##P < 0.01 and ###P < 0.001.
100 M. Tripathi et al. / Journal of Ethnopharmacology 136 (2011) 94–102
F branel
Bmtbcoiwmwf(ff
FcaFm(a*n
ig. 4. Dot plot consisting of upper half representing mitochondria with high memow membrane potential (depolarized mitochondria).
ax). Whereas anti-apoptotic proteins, blocks mitochondrial per-eability transition pore (MPTP) formation, pro-apoptotic proteins
ranslocate to mitochondria facilitating MPTP formation i.e. mem-rane permeabilization. During membrane permeabilization cytis released from mitochondria to cytosol and initiates cascade
f events for apoptosis (Kakkar and Singh, 2007). Nimesulidencreased the level of pro-apoptotic protein Bax by 4.2 folds
hereas level of Bcl2 (anti-apoptotic protein) was decreased in theitochondrial fraction as assessed by western blot. Pre-treatmentith Fp extract caused decrease in the Bax translocation by 4.7
olds (Fig. 5) and level of Bcl2 was found to increase by 3.17 foldsFig. 5) when compared to nimesulide stress. Cyt c release wasound to increase significantly during nimesulide stress in cytosolicraction by 6.6 folds as compared to control. Fp and silymarin pre-
ig. 5. Immuno-detection of cyt c released to cytosol (C) using goat polyclonal anti-yt c antibody. COX-IV and �-actin were used as internal controls for mitochondriand cytosolic fractions. Treatment of nimesulide was compared to control whilep and silymarin pre-administration were compared to nimesulide. Fp and sily-arin alone were compared to control groups. The sequence of samples in blot is
L to R) a: control, b: Fp pre-treated, c: silymarin pre-treated, d: nimesulide, e: Fplone and f: silymarin alone. (*) Significant difference compared with control group,P < 0.05, **P < 0.01 and ***P < 0.001 whereas (#) significant difference compared withimesulide treated group, #P < 0.05, ##P < 0.01 and ###P < 0.001.
potential (polarized mitochondria) and lower half representing mitochondria with
administration downregulated the cytosolic level of cyt c by 8.16and 8.14 folds, respectively (Fig. 5) when compared to nimesulidestress. The data suggest that pre-administration of Fp extract wasable to restrict the pro-apoptotic signals (Bax and cyt c release)and enhance the anti-apoptotic signal (Bcl2), thus accordingprotection.
3.8. Caspases activity (-9 and -3)
Earlier studies indicate that mitochondrial dysfunction activatespro-caspase-9 into active caspase-9 which in turn further activatespro-caspase-3 to initiate apoptosis. Treatment with nimesulidecaused significant increase in caspase-9 and -3 activities i.e. 80%and 60%, respectively (Fig. 6) confirming the involvement of mito-chondrial pathway. Pre-administration of whole plant extract of
Fp extract significantly prevented activation of caspases-9 and -3(125% and 85%) which was comparable with silymarin, the recoverycontrol (120% and 95%).Fig. 6. Effect of treatments on relative caspase-9 and -3 activities was measuredusing specific substrates for caspase-9 (Ac-LEHD-pNA) and caspase-3 (Ac-DEVD-pNA). Treatment with nimesulide was compared to control while Fp and silymarinpre-administration were compared to nimesulide. Fp and silymarin alone werecompared to control group. (*) Significant difference compared with control group,*P < 0.05, **P < 0.01 and ***P < 0.001 whereas (#) significant difference compared withnimesulide treated group, #P < 0.05, ##P < 0.01 and ###P < 0.001.

nopha
4
oriibhmaoiltgcaeawat
tAsei
cvuoaGwafnw
deisiasc
mptauwdsaipmrdt
M. Tripathi et al. / Journal of Eth
. Discussion
Hepatotoxicity is a well known reason for drug withdrawal,r delay in the development of safe therapeutics. A number ofesearches elucidated that NSAIDs are highly liver specific, caus-ng irreversible or reversible toxicity depending on the severity,n humans and animal models (Wallace et al., 2010). It haseen documented by several research groups that drug inducedepatotoxicity is closely associated with intracellular ROS for-ation leading to mitochondrial dysfunction, culminating into
poptosis. Preliminary data from our in vitro study suggests thatnly pre-treatment of Fp extract was effective against nimesulidenduced toxicity (Tripathi et al., 2010). The present study throwsight on the mechanism involved in nimesulide induced hepato-oxicity and the protection accorded by Fp extract in vivo, whereeneration of ROS and alteration in antioxidant status plays a criti-al role. Evidence has been provided that ROS generation is clearlyssociated with nimesulide induced liver damage in vivo. Plantxtracts are rich sources of antioxidant and can minimize the dam-ge caused by oxidative stress. So, the second aspect of the studyas to see if Fp extract pre-administration has the potential to
ttenuate nimesulide induced oxidative stress leading to hepato-oxicity.
Nimesulide caused significant hepatic damage as evident fromhe significant elevation of serum biomarkers of liver injury i.e. ALT,ST and SB. Histopathological studies also confirmed that nime-ulide caused significant toxicity. Oral pre-administration of Fpxtract caused significant prevention of nimesulide induced tox-city, demonstrating a hepatoprotective effect.
Excessive ROS production/loss of antioxidant defenses or bothause an oxidant-antioxidant imbalance making membrane lipidsulnerable to oxidative damage. MDA is a major oxidation prod-ct of polyunsaturated fatty acids, and is an important indicatorf oxidative damage to membrane lipids. In the present study,decrease was found in the SOD, GPx activity and increase inR and MDA content in the liver of rats treated with nimesulide,hich appears to be a result of increased ROS and compromised
ntioxidant defense resulting into oxidative stress. Data obtainedrom m-RNA expression of the antioxidant genes also suggest thatimesulide caused a significant alteration in the antioxidant genes,hich was prevented by Fp extract.
GSH depletion is an early event in the apoptotic cascade,rug exposure (Hammond et al., 2007) and oxidative stress (Hant al., 2008). Our observations revealed that there was significantncrease in ROS generation during nimesulide treatment whereasignificant decrease was found in the GSH content confirming thenvolvement of ROS and GSH. However, pre-administration withntioxidant (Fp) blunted the liver-injury caused by nimesulide anduggested that ROS production lies upstream of the apoptosis cas-ade.
There are evidences that ROS can induce apoptosis via severalechanisms, including mitochondria-dependent or independent
athway in various cell lines and tissues. The present study revealedhat generation of ROS may act on mitochondria, causing loss of �mnd subsequent apoptosis. So, further investigations were takenp to demonstrate how oxidative stress regulates apoptotic path-ay. Detection of �m was carried out as it is assumed that itsisruption is the onset of MPTP formation. Nimesulide caused aignificant decrease in �m as compared to control group, and pre-dministration of Fp extract was able to minimize the decreasen �m. Mitochondria provide the gateway through which apo-tosis can be executed in a rapid and robust manner. Impaired
itochondrial function is largely thought to be a core abnormalityesponsible for hepatic damage (Han et al., 2008). Mitochondria-ependent apoptosis involves outer membrane permeabilizationhat is associated with most pro-apoptotic stimuli which cause per-
rmacology 136 (2011) 94–102 101
turbations in the mitochondrial functions, resulting in dissipationof the trans-membrane potential. This process is controlled by thepro- and anti-apoptotic proteins of the Bcl2 family and leads to thecytosolic release of mitochondrial inter-membrane space proteinsthat can trigger either caspase-dependent or caspase-independentdeath pathways.
In the present study, phosphatidyl serine externalization con-firmed that apoptosis was the primary cause of cell death. Themain apoptotic signals like Bcl2, Bax, cyt c and caspases (-9 and -3)confirmed the involvement of mitochondria in nimesulide inducedapoptosis. Immunoblot analysis revealed significant decrease in thelevel of Bcl2 during nimesulide treatment, increase in the levels ofBax translocated from cytosol to mitochondria, and mitochondrialdepolarization. Data also demonstrate that nimesulide caused sig-nificant increase in cytosolic cyt c, a critical step in the formationof apoptosome, and caspase-9 activation which in turn leads to theactivation of caspase-3 leading to DNA fragmentation, a hallmarkof the apoptotic process. Whole plant extract of Fp conferred sig-nificant protection against these key control points of apoptosis i.e.it inhibited PS externalization, increased Bcl2 level, decreased Baxtranslocation, inhibited mitochondrial depolarization, cyt c release,caspase activities and DNA fragmentation. Thus, confirming theantioxidant efficacy and protective role of Fp extract against nime-sulide induced apoptosis.
5. Conclusion
In conclusion, nimesulide exposure caused significant alter-ations in the antioxidant level leading to oxidative stress.Nimesulide caused rapid generation of ROS, depletion of GSH, alter-ation in antioxidant genes, massive depolarization of mitochondrialmembrane leading to apoptosis. This study concludes that nime-sulide causes hepatotoxicity in rats through the involvement ofmitochondria. The present work suggests a beneficial effect of Fp(50% ethanolic) extract against nimesulide induced oxidative stresswhich also blocks the critical control points of apoptosis.
Conflict of interest statement
The author’s declare that there is no conflict of interest.
Acknowledgments
Authors are grateful to Director, IITR, for his kind support tothis work. The authors are thankful to Dr. Y. Shukla for provid-ing flow cytometer facility and Dr. B.P. Chowdhary for his kindhelp in histopathology work. Financial support received under thesupra-institutional project (SIP-08) is acknowledged. Authors arealso grateful to Indian Council of Medical Research and Councilfor Scientific and Industrial Research, New Delhi for fellowships.Authors are also grateful to IITR publication review committee forallocating the manuscript number, 2829.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.jep.2011.04.014.
References
Bjornsson, E., 2010. Review article: drug-induced liver injury in clinicalpractice. Alimentary Pharmacology and Therapeutics, doi:10.1111/j.1365-
2036.2010.04320.x.Campbell, R.V., Yang, Y., Wang, T., Rachamallu, A., Li, Y., Watowich, S.J., Weinman,S.A., 2009. Effects of hepatitis C core protein on mitochondrial electron trans-port and production of reactive oxygen species. Methods in Enzymology 456,363–380.

1 nopha
C
C
F
G
G
H
H
H
K
K
K
L
02 M. Tripathi et al. / Journal of Eth
arlberg, I., Mannervik, B., 1985. Glutathione reductase. Methods in Enzymology113, 484–490.
ossarizza, A., Ceccarelli, D., Masini, A., 1996. Functional heterogeneity of an isolatedmitochondrial population revealed by cytofluorometric analysis at the singleorganelle level. Experimental Cell Research 22, 84–94.
lohe, L., Gunzler, W.A., 1984. Assays of glutathione peroxidase. Methods in Enzy-mology 105, 114–121.
agliano, N., Donne, I.D., Torri, C., Migliori, M., Grizzi, F., Milzani, A., Filippi, C., Annoni,G., Colombo, P., Costa, F., Ceva-Grimaldi, G., Bertelli, A.A.E., Giovannini, L., Gioia,M., 2006. Early cytotoxic effects of ochratoxin A in rat liver: a morphological,biochemical and molecular study. Toxicology 225, 214–224.
irish, C., Koner, B.C., Jayanthi, S., Ramachandra, R.K., Rajesh, B., Pradhan, S.C., 2009.Hepatoprotective activity of picroliv, curcumin and ellagic acid compared to sily-marin on paracetamol induced liver toxicity in mice. Fundamental and ClinicalPharmacology 23, 735–745.
ammond, C.L., Marchan, R., Krance, S.M., Ballatori, N., 2007. Glutathione exportduring apoptosis requires functional multidrug resistance-associated proteins.Journal of Biological Chemistry 282, 14337–14347.
an, D., Shinohara, M., Ybanez, M.D., Saberi, B., Kaplowitz, N., 2010. Signal trans-duction pathways involved in drug-induced liver injury. In: Uetrecht, J. (Ed.),Adverse Drug Reactions, Handbook of Experimental Pharmacology, vol. 6.Springer-Verlag, Berlin, Heidelberg, pp. 269–310.
an, Y.H., Kim, S.H., Kim, S.Z., Park, W.H., 2008. Intracellular GSH levels ratherthan ROS levels are tightly related to AMA induced HeLa cell death. Chemico-Biological Interactions 171, 67–78.
akkar, P., Das, B., Viswanathan, P.N., 1984. A modified spectrophotometric assayof superoxide dismutase. Indian Journal of Biochemistry and Biophysics 21,130–132.
akkar, P., Singh, B.K., 2007. Mitochondria: a hub of redox activities and cellulardistress control. Molecular and Cellular Biochemistry. 305, 235–253.
alra, B.S., Shalini, Chaturvedi, S., Tayal, V., Gupta, U., 2009. Evaluationof gastro-tolerability, antinociceptive and anti-inflammatory activity of
combination of NSAIDs in rats Indian. Journal of Dental Research 20,418–422.imaye, P.V., Raghuram, N., Sivakam, S., 2003. Oxidative stress and gene expressionof antioxidant enzymes in the renal cortex of streptozotocin induced diabeticrats. Molecular and Cellular Biochemistry 243, 147–152.
rmacology 136 (2011) 94–102
Liu, J., Yagi, T., Sadamori, H., Matsukawa, H., Sun, D.S., Mitsuoka, N., Yamamura,M., Matsuoka, J., Jin, Z., Yamamoto, I., Tanaka, N., 2003. Annexin V assay-provenanti-apoptotic effect of ascorbic acid 2-glucoside after cold ischemia/reperfusioninjury in rat liver transplantation. Acta Medica Okayama. 57, 209–216.
Lowry, O.H., Rosenbourgh, N.J., Farr, A.L., Randall, R.J., 1951. Protein measurementwith folin phenol reagent. Journal of Biological Chemistry 193, 265–275.
Orhan, I., Sener, B., Choudhary, M.I., Khalid, A., 2004. Acetylcholinesterase andbutyrylcholinesterase inhibitory activity of some Turkish medicinal plants. Jour-nal of Ethnopharmacology 91, 57–60.
Porkey, J., 2007. Are natural antioxidants better and safer—than synthetic antioxi-dants? European Journal of Lipid Science and Technology 109, 629–642.
Seglen, P.O., 1976. Preparation of isolated rat liver cells. Methods in Cell Biology 13,29–83.
Singh, B.K., Tripathi, M., Pandey, P.K., Kakkar, P., 2010. Nimesulide aggravates redoximbalance and calcium dependent mitochondrial permeability transition lead-ing to dysfunction in vitro. Toxicology 275, 1–9.
Suau, R., Cabezudo, B., Rico, R., Najera, F., Lopez-Romero, J.M., 2002. Direct determi-nation of alkaloid contents in Fumaria species by GC-MS. Phytochemical Analysis13, 363–367.
Tripathi, M., Singh, B.K., Mishra, C., Raisuddin, S., Kakkar, P., 2010. Involvement ofmitochondria mediated pathways in hepatoprotection conferred by Fumariaparviflora Lam. extract against nimesulide induced apoptosis in vitro. ToxicologyIn Vitro 24, 495–508.
Wallace, K., Cowie, D.E., Konstantinou, D.K., Hill, S., Tjelle, T.E., Axon, A., Koruth,M., White, S.A., Carlsen, H., Mann, D.A., Wright, M.C., 2010. The PXR is a drugtarget for chronic inflammatory liver disease. Journal of Steroid Biochemistryand Molecular Biology 120, 137–148.
Wallin, B., Rosengren, B., Shertzer, H.G., Camejo, G., 1993. Lipoprotein oxidation andmeasurement of thiobarbituric acid reacting substances formation in a singlemicrotiter plate: its use for evaluation of antioxidants. Analytical Biochemistry208, 10–15.
Zhang, Q.H., Sheng, H.P., Brenanner, T.A., Loh, T.T., 1999. Redistribution of
cytochrome c release is not an essential requirement in C2-ceramide inducedapoptosis in HL-60 cells. Life Sciences 65, 1715–1723.Zhang, R., Niu, Y., Zhou, Y., 2010. Increase the cisplatin cytotoxicity and cisplatin-induced DNA damage in HepG2 cells by XRCC1 abrogation related mechanisms.Toxicology Letters 192, 108–114.

























